Submitted:
15 January 2026
Posted:
16 January 2026
You are already at the latest version
Abstract
The centrosome, long recognized as the primary microtubule-organizing center (MTOC) of animal cells, is re-examined through the lens of information theory and systems biology. This preprint proposes a unifying hypothesis: the mother centriole within the centrosome acts as a non-genetic cellular ledger, a stable structural repository that accumulates molecular records of a cell’s replicative history and environmental exposures. These records—comprising specific post-translational modification (PTM) signatures, retained proteins, and structural alterations—are subsequently “read” by the cell to inform critical decisions regarding proliferation, differentiation, senescence, and apoptosis. We synthesize evidence from cell biology, gerontology, and evolutionary biology to construct the “Centrosomal Ledger Model.” This model positions the centriole not as a passive cytoskeletal component but as an active, heritable information-processing node that integrates temporal data across scales—from circadian rhythms to organismal aging. We detail the molecular mechanisms of information encoding (e.g., tubulin polyglutamylation, oxidative marks) and decoding (via ciliary signaling, proteostatic feedback, and mechanical transduction). The model’s implications challenge genetic determinism by highlighting structural inheritance, provides a material basis for cellular age, and offers novel, falsifiable avenues for experimental interrogation in aging and cancer research. Crucially, it suggests that modulating the “read-write” cycle of the centrosomal ledger could represent a new frontier in regenerative medicine.
Keywords:
centrosome
; centriole
; cellular memory
; structural inheritance
; post-translational modifications
; aging
; stem cell fate
1. Introduction: Beyond the Microtubule Organizer
The canonical view of the centrosome frames it as an orchestrator of spatial order: it nucleates and anchors microtubules, establishes cell polarity, and forms the poles of the mitotic spindle (Bornens, 2012). However, several persistent observations defy a purely structural explanation. The mother centriole’s remarkable longevity, its asymmetric inheritance during stem cell divisions, and its role as a platform for signaling complexes suggest a deeper, information-centric function (Venkei & Yamashita, 2018). We hypothesize that the mother centriole has evolved to function as a cellular ledger—a durable, non-genetic data storage device that logs biographical data and computes cellular fate.
This preprint integrates disparate lines of evidence into a coherent model. We first define the molecular language of the ledger (Section 2), then describe the cellular machinery for reading it (Section 3). We place this mechanism within the hierarchy of biological timing systems (Section 4), examine its variation across cell types (Section 5), and trace its evolutionary origins (Section 6). We propose critical experimental tests (Section 7), explore broader implications (Section 8), and present a formal integrative model (Section 9).
2. The Molecular Syntax of the Ledger: Encoding Time and Experience
The centriole’s cylindrical, nine-fold symmetric structure of stable microtubule triplets provides an ideal, low-turnover substrate for information encoding. Data is written via two primary classes of molecular notations.
2.1. Post-Translational Modifications as Chronological and Damage Marks
The tubulin backbone undergoes progressive PTMs that serve distinct purposes.
- Neutral PTMs (Replicative Tally): Enzymatic modifications like polyglutamylation accumulate predictably with each cell cycle on the mother centriole. They act as a maturity index, marking functional readiness for roles like basal body formation (Bré et al., 2021). The level of polyglutamylation (PolyG) can be conceptualized as a function of division number (n):where ΔG represents the addition per cycle.PolyG(n) ≈ PolyG(n-1) + ΔG
- Pathological PTMs (Stress Log): Stochastic, damage-associated modifications create a damage index. This includes oxidation of scaffold proteins (e.g., SAS-6), glycation of long-lived proteins (e.g., CEP135), and ubiquitination marking misfolded clients (Wang et al., 2021). These correlate with functional decline.

Table 1.
Molecular “Writes” to the Centrosomal Ledger.
| Modification Type | Example | Proposed Information Encoded | Functional Consequence |
|---|---|---|---|
| Neutral PTM | Polyglutamylation | Number of successful divisions (Maturity) | Stabilization; recruitment of specific effectors (e.g., ciliary assembly proteins) |
| Neutral PTM | Acetylation (K40) | Age of microtubule structure | Reduced dynamics, increased mechanical resistance |
| Pathological PTM | Cysteine Oxidation | History of oxidative stress | Weakened structural integrity, errors in duplication |
| Pathological PTM | Advanced Glycation End-products (AGEs) | Cumulative metabolic stress | Loss of protein function, aggregation propensity |
| Protein Retention | CAFDs (e.g., YAP/TAZ) | Past signaling context (e.g., Hippo pathway activity) | Altered transcriptional preparedness upon release |
2.2. Proteomic Composition as a Snapshot of State
Beyond PTMs, the ledger’s proteome is dynamic. The mother centriole selectively retains Centriole-Associated Fate Determinants (CAFDs)—proteins like transcription regulators and kinases—whose presence reflects the cell’s recent signaling history (Jakobsen et al., 2011). The composition of distal appendage proteins and recruited chaperones (e.g., HSP90) further provides a real-time snapshot of centriolar age and stress.
3. Decoding the Ledger: Cellular Read-Out Mechanisms
The stored information is biologically inert until interpreted. Cells employ parallel pathways to read the ledger’s state.
3.1. Mechanical Read-Out
The centrosome’s core output is microtubule nucleation. A ledger with high damage/maturity indices nucleates a less dynamic, more rigid microtubule array. This altered mechanical output is sensed by +TIP proteins (e.g., EB1) and transmitted via the LINC complex to the nucleus, influencing chromatin organization and gene expression (Lomakin et al., 2015).
3.2. Signaling Read-Out
As the basal body, the mother centriole dictates primary cilium integrity. A damaged ledger leads to a defective cilium, misinterpreting morphogen signals like Sonic Hedgehog (Shh), thereby altering transcriptional programs for fate and proliferation (Breslow & Holland, 2019).
3.3. Proteostatic Read-Out
Accumulated pathological PTMs chronically recruit protein quality control systems (chaperones, ubiquitin ligases). Sustained engagement can spill over to activate global stress responses like the Unfolded Protein Response (UPR), a known driver of senescence (Conroy et al., 2022).
Figure 1: The Information Cycle of the Centrosomal Ledger. (A schematic showing: 1. Inputs of Replication & Stress, 2. Molecular Writing (PTMs, CAFDs), 3. The Stable Ledger (Mother Centriole), 4. Parallel Read-Out Pathways (Mechanical, Signaling, Proteostatic), 5. Fate Decisions (Renewal vs. Aging Programs)).
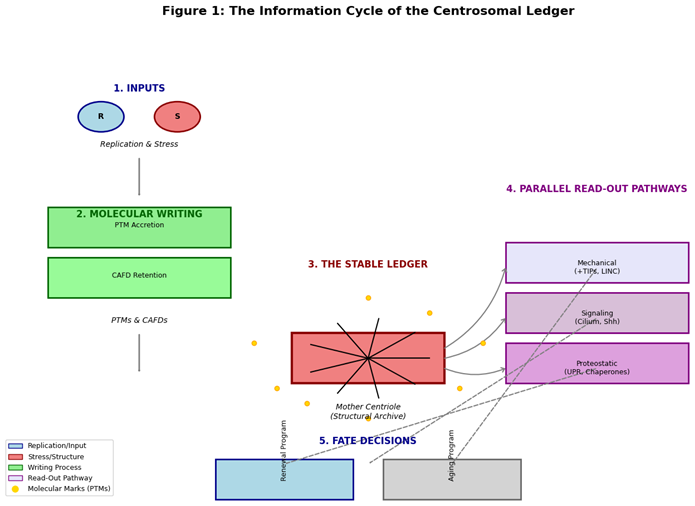
4. The Ledger in the Hierarchy of Biological Clocks
The centrosomal ledger is not an isolated timer but a key node in a network.
- Level 1: Fast Oscillators. Circadian clock proteins (e.g., PER2) localize to centrosomes, suggesting the ledger integrates daily metabolic rhythms with long-term age data (Wils et al., 2021).
- Level 2: Replicative Clocks. The ledger interacts with telomeric and epigenetic clocks. Centriole dysfunction induces genomic instability, accelerating telomere attrition. Conversely, epigenetic changes may lock in a “senescent” interpretation of ledger data.
- Level 3: Systemic Aging. In stem cells, asymmetric inheritance of a “young” vs. “old” ledger determines self-renewal vs. differentiation balance. Age-related decline in this process depletes regenerative niches, driving tissue aging (Geiger et al., 2013).
5. Comparative Cell Biology: Ledger Adaptations
The ledger’s function is tailored to cellular destiny.
- Germline: Employs strict asymmetric inheritance and stringent quality control to reset the ledger, minimizing intergenerational damage transfer (Fishman et al., 2018).
- Somatic Stem Cells: Exhibits “leaky” asymmetry, where imperfect segregation gradually increases damage load in the stem pool, contributing to niche aging.
- Differentiated Cells: The ledger is “paused”; the centriole becomes a static basal body, with age-related damage manifesting as ciliopathies.
- Cancer Cells: The ledger is “hijacked”—amplification and structural distortion disrupt normal timekeeping, enabling unchecked proliferation at the cost of genomic instability (Marteil et al., 2018).
Table 2.
The Centrosomal Ledger Across Cell Types.
| Cell Type | Ledger State | Inheritance Mode | Primary Time-Related Output |
|---|---|---|---|
| Germ Cell | Reset, Immortal | Strictly Asymmetric | Fertility, generational continuity |
| Somatic Stem Cell | Aging, Leaky | Asymmetric (with error) | Tissue homeostasis, niche depletion |
| Differentiated Cell | Paused, Static | Symmetric / Non-dividing | Sensory function, age-related ciliopathies |
| Cancer Cell | Corrupted, Amplified | Aberrant, Symmetric | Uncontrolled proliferation, genomic instability |
6. Evolutionary Origins: Why the Centriole?
The centriole was pre-adapted for this role. Its evolution from a motile basal body in unicellular organisms to a spindle organizer provided structural stability and obligatory inheritance. In metazoans, its inherent asymmetry was co-opted for asymmetric cell division. Finally, in long-lived animals, its durability made it ideal for recording time, transforming it from a structural organelle into an informational ledger (Carvalho-Santos et al., 2011). Plants, lacking centrioles, must utilize alternative systems (e.g., nuclear envelope microtubule nucleation), highlighting the ledger as an animal-specific solution to managing cellular time in complex, motile organisms.
7. Experimental Framework and Falsifiability
The model generates testable predictions:
- Prediction: Targeted reversal of centriolar PTMs (e.g., deacetylation, redox repair) in aged stem cells will rejuvenate function.
- Prediction: Microinjection of in vitro “aged” (glycated/oxidized) centrioles into young cells will induce senescence markers.
- Prediction: Comparative biology will show that centriole-less organisms (plants) rely on fundamentally different primary drivers for stem cell aging.
A critical falsification would be if interventions that successfully reset other aging clocks (telomeres, epigenetics) fully rejuvenate cell function independent of the centriolar state.
8. Philosophical and Practical Implications
This model challenges genetic reductionism, arguing for structural inheritance as a crucial layer of biological information. It materializes time, defining cellular age as the physical state of the ledger. Practically, it identifies the centrosome as a novel therapeutic target. Strategies could include: 1) Ledger “Cleansing” (enzymatic repair of PTMs), 2) Ledger “Stabilization” (chaperone enhancement), and 3) Ledger “Reset” (promoting asymmetric inheritance of young templates in stem cells).
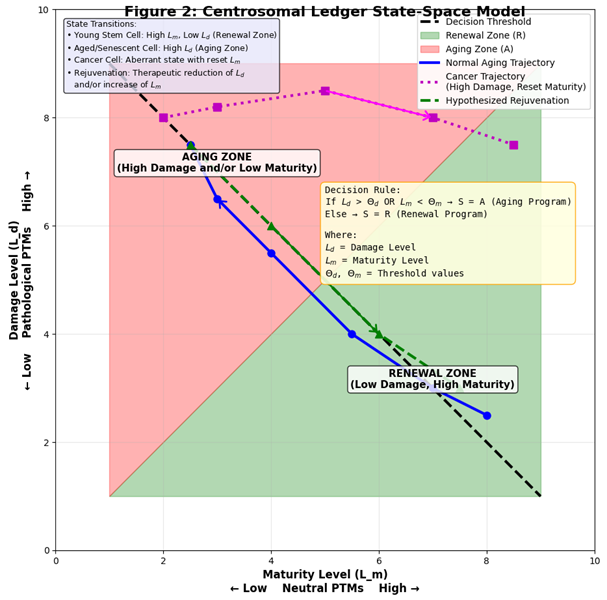
9. The Integrative Centrosomal Ledger Model: A Formal Summary
We propose the cell’s fate (F) is a function of the state (S) of its centrosomal ledger, integrated with other systems (genomic G, epigenetic E, metabolic M).
where:
F(t) = Φ( S(L_m, L_d), G, E, M )
- L_m = Maturity Level (from neutral PTMs)
- L_d = Damage Level (from pathological PTMs)
The ledger state S transitions between zones that promote Renewal (R) or Aging (A) based on threshold crossings:
- If L_d > Θ_d or L_m < Θ_m --> S = A (Engage Senescence/Apoptosis)
- Else --> S = R (Promote Proliferation/Differentiation)
Figure 2: The Centrosomal Ledger State-Space Model. (A graph with Maturity (L_m) on the X-axis and Damage (L_d) on the Y-axis. A diagonal threshold line separates the “Renewal Zone” (low damage, high maturity) from the “Aging Zone” (high damage and/or low maturity). Arrows show trajectories for normal aging, cancer (high damage, reset maturity), and hypothesized rejuvenation.)
Conclusions
The Centrosomal Ledger Model provides a unifying, mechanistic framework that elevates the centrosome from a cytoskeletal organizer to a central processor of cellular time and fate. By reconciling disparate observations in aging, development, and cancer, it opens new avenues for research and suggests that the path to modulating longevity may lie in understanding and maintaining the molecular memory of our cells’ architecture.
References
- Bornens, M. The centrosome in cells and organisms. Science 2012, 335(6067), 422–426. [Google Scholar] [CrossRef] [PubMed]
- Bré, M. H.; Redeker, V.; Levilliers, N. Post-translational modifications of tubulin and microtubule stability in cilia and flagella. In The Cytoskeleton: Diverse Roles in Structure and Mechanism; Springer: Cham, 2021; pp. 47–70. [Google Scholar] [CrossRef]
- Breslow, D. K.; Holland, A. J. Mechanism and regulation of centriole and cilium biogenesis. Annual Review of Biochemistry 2019, 88, 691–724. [Google Scholar] [CrossRef] [PubMed]
- Conroy, P. C.; Saladino, C.; Dantas, T. J.; Lalor, P.; Dockery, P.; Morrison, C. G. Centrosome adaptation to DNA damage by 53BP1 recruitment facilitates error-free mitosis. Nature Communications 2022, 13(1), 620. [Google Scholar] [CrossRef]
- Carvalho-Santos, Z.; Azimzadeh, J.; Pereira-Leal, J. B.; Bettencourt-Dias, M. Evolution: Tracing the origins of centrioles, cilia, and flagella. The Journal of Cell Biology 2011, 194(2), 165–175. [Google Scholar] [CrossRef] [PubMed]
- Fishman, E. L.; Jo, K.; Nguyen, Q. P. H.; Kong, D.; Royfman, R.; Cekic, A. R.; …; Avidor-Reiss, T. A novel atypical sperm centriole is functional during human fertilization. Nature Communications 2018, 9(1), 2210. [Google Scholar] [CrossRef] [PubMed]
- Geiger, H.; de Haan, G.; Florian, M. C. The ageing haematopoietic stem cell compartment. Nature Reviews Immunology 2013, 13(5), 376–389. [Google Scholar] [CrossRef] [PubMed]
- Jaba, T. Dasatinib and quercetin: short-term simultaneous administration yields senolytic effect in humans. Issues and Developments in Medicine and Medical Research 2022, Vol. 2, 22–31. [Google Scholar]
- Jakobsen, L.; Vanselow, K.; Skogs, M.; Toyoda, Y.; Lundberg, E.; Poser, I.; …; Andersen, J. S. Novel asymmetrically localizing components of human centrosomes identified by complementary proteomics methods. The EMBO Journal 2011, 30(8), 1520–1535. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, A. J.; Cattin, C. J.; Cuvelier, D.; Alraies, Z.; Molina, M.; Nader, G. P. F.; …; Piel, M. The nucleus acts as a ruler tailoring cell responses to spatial constraints. Science 2015, 348(6232), 560–563. [Google Scholar] [CrossRef] [PubMed]
- Marteil, G.; Guerrero, A.; Vieira, A. F.; de Almeida, B. P.; Machado, P.; Mendonça, S.; …; Bettencourt-Dias, M. Over-elongation of centrioles in cancer promotes centriole amplification and mitotic defects. The EMBO Journal 2018, 37(22), e100303. [Google Scholar] [CrossRef]
- Venkei, Z. G.; Yamashita, Y. M. Emerging mechanisms of asymmetric stem cell division. The Journal of Cell Biology 2018, 217(11), 3785–3795. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. J.; Acehan, D.; Kao, C. H.; Jane, W. N.; Uryu, K.; Tsou, M. F. B. The deubiquitinase USP33 regulates centrosome biogenesis via the SAS-6 protein. Journal of Cell Science 2021, 134(2), jcs247155. [Google Scholar] [CrossRef]
- Wils, L.; Rombauts, F.; van der Horst, G. T. J. The circadian clock and centrioles: Insights into the regulation of cell division. Seminars in Cell & Developmental Biology 2021, 126, 78–84. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



