
Submitted:
15 January 2026
Posted:
16 January 2026
You are already at the latest version
Abstract
Infrasound, physically defined as sound at frequencies below 20 Hertz, can travel long distances with minimal attenuation and permeate biological tissues due to its marked particle displacement and deep penetration. Generated by both natural phenomena and human-made systems, infrasound has drawn increasing scientific and public attention regarding its potential physiological and psychological effects. Experimental studies demonstrate that infrasound can modulate mechanosensitive structures at the cellular level, particularly pressure-sensitive ion channels such as PIEZO1 and TRPV4, leading to intracellular calcium influx, oxidative stress, altered intercellular communication, and in some settings, apoptosis. These responses vary according to sound pressure levels, frequencies, exposure duration and tissue type. In the cardiovascular system, higher sound pressures have been associated with mitochondrial injury and fibrosis, whereas low sound pressures may exert context-dependent protective effects. In animal models, prolonged or intense exposure to infrasound has been shown to induce neuroinflammatory responses and memory impairment. Short-term studies in humans at moderate intensities have reported minimal physiological changes, with psychological and contextual factors influencing symptom perception. Occupational environments such as factories and agricultural settings may contain elevated levels of infrasound, underscoring the importance of systematic measurements and exposure assessments. At the same time, controlled infrasound stimulation has shown potential as an adjunct modality in bone repair and tissue regeneration, highlighting its dual capacity as both a biological stressor and a possible therapeutic tool. Overall, existing data indicate that infrasound may be harmful at chronic exposure depending on intensity and frequency, yet beneficial when precisely regulated. Future research should standardize exposure metrics, refine measurement technologies, and clarify dose–response relationships to better define the health risks and therapeutic applications of infrasound.
Keywords:
infrasound
; mechanotransduction
; PIEZO1
; TRPV4
; inflammation
1. Introduction
Sound consists of rhythmic fluctuations in air pressure superimposed on the constant atmospheric background [1]. These oscillations vary in frequency, forming the sound’s spectral composition. Complex non-sinusoidal acoustic patterns predominate in the environment, they lead to several integer multiples of the determined fundamental frequency, the so-called harmonics.
When sufficiently intense, sound can be perceived as noise or felt as vibration [2]. Although sound intensity depends on the amplitude of pressure variations, particle displacement - the movement of air particles - increases as frequency decreases, meaning low-frequency waves induce more pronounced mechanical movement than high-frequency waves of equal pressure [1,2].
Infrasound is sound with frequencies below the range of human hearing. Complex acoustic and electromagnetic patterns can mediate information at the cellular level even at very low intensity. Frequencies below 10 Hz are considered to have a particularly high information content. However, the information content of a spectrum depends on all characteristics, including, of course, the duration or time of occurrence [3,5]. It can be generated from natural sources such as wind, volcanoes, avalanches, and earthquakes. It can also be generated by man-made sources, e.g. wind turbines, diesel engines, and ventilation systems. Among man-made sources, wind turbine-generated infrasound is distinctive for its rhythmic, repetitive pattern, setting it apart from most other environmental low-frequency sources and drawing particular attention in both public discourse and scientific investigation.
To understand the effect of infrasound on biological mechanisms and public health relevance, we review and summarize current knowledge of infrasound’s physiological and psychological impact, discuss associated risks and therapeutic applications, and identify remaining research gaps. This manuscript employs several definitions. Primarily, we discuss frequency. We explain frequency in terms of infrasound, which is defined as sound below 20 Hz in humans (ISO 226:2003). Furthermore, we discuss the term amplitude. This refers to the change in value of a measure, specifically, in this case, the change in pressure from the pressure before the change. Additionally, we discuss sound pressure. Sound pressure is a measurement of the change in pressure of the air around the space where a sound is being produced. Finally, we measure sound intensity. Sound intensity is a measure of the energy per unit volume in a given space caused by sound. Sound intensity is proportional to the pressure of sound in that field.
2. Cellular and Molecular Responses
In biological tissues, infrasound propagates as longitudinal mechanical waves shaped by tissue density and elasticity. Because it undergoes minimal attenuation, infrasound can penetrate deeply and may produce resonance when its frequency aligns with that of specific organs or tissues [4]. In resonance, the absolute strength of the stimulus is less important than the alignment of its frequency with the system’s intrinsic vibrations. Even weak stimuli, when temporally and spatially synchronized, can significantly affect system stability. The human body, as an energetic organism, is comparable to an oscillator with a complex waveform, whereby different organs have different resonance solutions [5].
Resonance can intensify mechanical strain, promoting tissue deformation, mechanotransduction, and, under sustained exposure, structural or functional damage. Modeling studies indicate that tissues with distinct acoustic impedances exhibit different resonance responses, with certain frequencies enhancing energy absorption and localized effects [6,7].
In order to maintain the integrity of cells and tissues in organisms in terms of structure, function and communication, the unimpeded transmission of mechanical forces into biological and biochemical information is one of the most important fundamentals. At the cellular level, mechanical oscillations such as infrasound activate mechanosensitive ion channels, e.g., PIEZO1, TRPV4, and Connexin 43 (Cx43) hemichannels, by inducing membrane tension or substrate deformation [6]. These channels open to cation influx (mainly Ca²⁺), initiating signaling cascades that modulate proliferation, differentiation, and apoptosis [6]. PIEZO1 efficiently transduces low-frequency stimuli within the 0.5–50 Hz range, including infrasound [7]. TRPV4 is robustly activated around 16 Hz, particularly in glial cells [8,9]. Finally, Cx43 hemichannels, downstream of PIEZO1 or TRPV4, mediate ATP and glutamate release, sustaining intercellular communication [10,11]. Crosstalk among these channels forms a coordinated mechanosensitive network through which infrasound can influence tissue physiology and cellular homeostasis.
It is important to remember the dual nature of PIEZO channels in their context. PIEZO channels often participate in pro-inflammatory signals, but depending on the environment, they can also promote anti-inflammatory or homeostatic effects, mainly influenced by the mechanical surroundings. For example, in endothelial cells, activation of PIEZO channels by laminar shear stress helps maintain vascular integrity. Conversely, when oscillatory forces dominate, pro-inflammatory responses tend to prevail, weakening vascular integrity and immune regulation [12].
At the cellular level, the effects of infrasound can influence multiple pathways, including proliferation, apoptosis, and intracellular signalling. For instance, studies on rat bone marrow mesenchymal stem cells (BMSCs) demonstrated that infrasound enhances proliferation and inhibits apoptosis, effects mediated in part through activation of the Notch signaling pathway and upregulation of surviving, a key anti-apoptotic protein [13,14]. In parallel, neuronal studies revealed that infrasound exposure induces oxidative stress and apoptosis in the hippocampus, which could be mitigated by antioxidant compounds that modulate the expression of Bcl-2, Bax, and caspase-3, underscoring the role of reactive oxygen species and apoptotic signaling in infrasound-induced cellular injury [15]. Furthermore, infrasound was found to alter blood-retinal barrier permeability by increasing the activity of calcium-activated potassium channels in retinal microvascular endothelial cells, demonstrating that infrasound can directly affect ion channel dynamics and cellular excitability [16]. These results emphasize that infrasound can actively reshape key regulatory pathways governing cell survival and functional integrity.
Infrasound has also been proven to impact glial and microbial cells. Astrocytes in the rat hippocampus responded to infrasound with increased A1 reactive astrocyte formation and elevated glutamate release via Cx43 hemichannels, effects that were modulated by the FGF2/FGFR1 pathway and microglial activation, highlighting the involvement of neuron–glia interactions in central nervous system injury [14,17]. Similarly, microbial studies demonstrated that mechanical vibration at infrasound frequencies can either stimulate or inhibit bacterial growth depending on the exposure duration and frequency, suggesting that both extracellular medium properties and cellular mechanosensors mediate microbial responses [18]. Finally, investigations in plant systems revealed that infrasound-induced changes in water structure and cell hydration may act as a sensitive signal for intracellular metabolic activity, indicating that physical cues can propagate molecular responses across diverse organisms [19]. Altogether, these findings reveal the multifaceted nature of infrasound, demonstrating its capacity to influence cellular processes through interconnected mechanical, biochemical, and biophysical mechanisms.
3. Systemic Effects on Organs and Tissues
3.1. Cardiovascular System
Results from animal studies indicate that exposure to infrasound at high sound pressure levels (SPLs) can cause a significant impact on components of the cardiovascular system. In contrast, human studies (particularly those conducted under controlled laboratory or epidemiological conditions) provide conflicting and complex results and often do not lead to definitive conclusions.
Research focusing on rodent models has provided considerable insight into the myocardial effects of high SPL infrasound. Pei et al. (2007) exposed rats to 5 Hz infrasound at 130 dB for 2 hours daily over 1, 7, and 14 days, and evaluated hemodynamic, molecular, and ultrastructural indices at the end of each exposure period. The study showed a time-dependent change in the function and structure of the heart. Left ventricular systolic pressure, together with + dP/dt and – dP/dt, showed a progressive increase and a decrease, respectively. Mitochondrial swelling and intracellular calcium levels also significantly and progressively increased during the exposure period. Inhibition of SERCA2 (Sarco/endoplasmic reticulum Ca²⁺-ATPase 2) protein expression showed a unique pattern of predominance: after a one-day increase, there was a significant decrease after prolonged seven- and fourteen-day exposures. The varied muscular and ultrastructural disorganization was then accompanied by disrupted calcium homeostasis and impaired excitation-contraction-sarcomere coupling and relaxation mechanisms, which certainly deserve more attention [20].
Subsequent studies showed that exposure to infrasound induced apoptosis in neonatal rat cardiomyocytes by altering the expression of proteins involved in apoptosis pathways. including upregulation of Bax and caspases, the pro-apoptotic proteins, and downregulation of Bcl-2, the anti-apoptotic protein [21]. The same research group further demonstrated that exposure to infrasound increased oxidative stress by reducing the expression of PPAR-γ and the activity of antioxidant enzymes, resulting in cardiomyocyte damage. Pharmacological stimulation of PPAR-γ significantly reduced these damages, as evidenced by decreased oxidative injury (lower lipid peroxidation), preservation of mitochondrial ultrastructure, and reduced cardiomyocyte apoptosis [22]. Overall, these findings indicate that increased intracellular calcium concentration, impaired mitochondrial function, activation of apoptotic pathways, and oxidative stress are among the most important mechanisms involved in myocardial damage caused by exposure to infrasound.
Other animal studies corroborate these observations. Lousinha et al (2018) demonstrated that rats exposed to infrasound developed coronary perivascular fibrosis, while dexamethasone attenuated the effect, implicating inflammatory pathways in the fibrotic response [23]. Similarly, Antunes et al (2013) reported myocardial fibrosis in rats chronically exposed to low-frequency noise (≤500 Hz, including infrasonic components), within the conceptual framework of ”vibroacoustic disease” [24].
In contrast to the findings described above regarding infrasound-induced cardiomyocyte damage through increased apoptosis and oxidative stress [21,22], Jin and co-workers studied the effects of infrasound on rat cardiac fibroblasts and demonstrated that low-SPL infrasound (80-86 dB) actually suppressed pathological processes in the fibroblasts. Specifically, low SPL exposure inhibited angiotensin II-induced proliferation and collagen synthesis, acting through the microRNA-29a/TGF-β/Smad3 and Nrf2/HO-1 pathways. These findings raise the intriguing possibility that infrasound may exert bidirectional effects depending on dose and cellular context, being harmful at high intensities but potentially protective at lower, sub-threshold SPLs [25].
Experiments in ex vivo human tissue extend these insights. Exposure of atrial appendage muscle samples obtained during cardiac surgery to infrasound >100 dB (Z) for one hour showed a graded reduction in myocardial contractility, averaging a 9% decline in force for each 10 dB increase above 100 dB(Z). These results provide rare direct evidence that human myocardium is functionally sensitive to infrasound at supraphysiological intensities [26].
The translation of these findings to intact humans is complex. Controlled laboratory studies from the 1980s onward have yielded heterogeneous results. Danielsson and his co-workers reported increases in diastolic blood pressure in healthy volunteers during acute exposure to 6-16 Hz infrasound at high SPLs, without concomitant changes in heart rate [27]. Likewise, Qibai et al. found that an hour of exposure to infrasound at 2.1 Hz (110 dB) or 4.1 Hz (120 dB) in healthy volunteers resulted in an increase in systolic and diastolic blood pressure, an increase in heart rate was generally observed at 120 dB, but at 110 dB, the heart rate response was inconsistent (increase or decrease). Discomfort, headache, ear pressure, and fatigue were also reported by participants [28]. In an early study, Moller et al. observed no cardiovascular changes when subjects were exposed to short-term near-threshold levels (<20 dB above auditory threshold). However, annoyance and sensations of ear pressure were common [29]. Birnie reported small, non-significant increases in blood pressure under unspecified infrasonic exposures, again accompanied by subjective symptoms [30]. These early experiments suggested that only high-intensity infrasound (>100d(B)Z) elicits measurable cardiovascular responses in humans, whereas near-threshold exposures are more likely to cause annoyance than physiological changes. Unfortunately, short-term studies are not eligible to draw conclusions about long-term effects on the cardiovascular system.
More recent and methodologically rigorous studies have focused on infrasound and low frequency noise generated by wind turbines. Health Canada’s large cross-sectional survey of over 1200 adults living near turbines found no association between exposure time and blood pressure or heart rate [31]. This study was performed in 2012 and is based on data on dwellings in the vicinity of smaller wind turbines with rated electrical power output ranging from 660 kW to 3MW. In a 72-hour, double-blind, crossover study of noise-sensitive adults exposed to simulated wind turbine infrasound (1.6-20 Hz; 90 dB peak), no cardiovascular effects were detected [32]. However, those studies do not consider the current state of knowledge regarding the impact of infrasound on cellular mechanotransduction, as reflected in blood pressure and heart rate measurements. Exploratory field studies have suggested possible associations between low-frequency components of wind turbine noise (20-200 Hz) and altered heart rate variability, but these findings cannot be attributed to infrasound and remain inconclusive [33].
Nonetheless, there is evidence for acute vascular effects. Deng et al. reported that exposure to short-term low-frequency noise (70-100 Hz ≤85 dB) increased cutaneous blood flow in both humans and mice, mediated in part by nitric oxide. Although these frequencies extend slightly above the infrasonic range, the findings suggest a mechanosensory role for the endothelium and highlight the need to examine vascular regulatory mechanisms at infrasonic frequencies [34,35,36].
Despite decades of research, many gaps remain, particularly regarding the intensity threshold of infrasound exposure-related effects, the differences between animal and human studies, the precise cellular mechanosensory pathways, and the associated markers of vascular health and blood flow at the microcirculatory level. Recent microvascular modeling has established standard reference values for blood velocity in exchange microvessels. This enables distinguishing between healthy and diseased states using biophysical flow indicators [37]. To gain a clearer understanding, future studies should be conducted using precisely calibrated chambers, double-blind designs, and advanced mechanical analyses. In addition, standard cardiovascular and microcirculatory markers should be used to properly elucidate the true impact of infrasound on cardiovascular functions. Research using Laser Doppler Flowmetry during local thermal hyperemia has demonstrated that skin microvascular reactivity is highly sensitive to endothelial dysfunction. This underscores the importance of microcirculatory assessments in detecting early vascular impairment [38]. Normative datasets for sublingual microcirculation obtained through SDF imaging in healthy pediatric populations further demonstrate the robustness and physiological sensitivity of microvascular imaging methods [39]. Complementary findings from adult cohorts using incident dark-field imaging show age- and sex-specific variation in sublingual microcirculatory parameters, broadening the reference framework for assessing vascular function across populations [40].
3.2. Central Nervous System
Infrasound exposure (particularly when delivered at high SPL) has repeatedly been linked in animals and in vitro models to neuronal apoptosis and oxidative stress, while human data remains less clear-cut. Here, we summarize findings from diverse experimental paradigms, comparing rodent studies, cellular assays, and limited human research, evaluating mechanistic pathways, neurofunctional outcomes, and potential protective responses.
Extensive rodent experiments have demonstrated that infrasound exposure induces hippocampal neuronal apoptosis alongside oxidative imbalance. For example, rats exposed to 8 Hz at 140 dB for 2 h/day across three days exhibited increased neuronal apoptosis in hippocampal regions, correlating with impaired spatial learning and memory [41]. In mice, being repetitively exposed to 16 Hz infrasound at 130 dB for 2 hours per day for 8 days caused increased inflammatory cytokine levels in the CNS as well as elevated oxidative stress, leading again to impaired memory and spatial learning due to neuronal damage in the prefrontal cortex and hippocampus [42]. Mechanistically, epigallocatechin gallate (EGCG), an antioxidant, has been shown to suppress infrasound-induced neuronal apoptosis in rat hippocampi, confirming oxidative stress as a causal mediator [43]. Together, these findings support a model in which infrasound triggers mitochondrial dysfunction, reactive oxygen species (ROS) generation, and apoptotic cascades involving p53 upregulation and Bcl-2 suppression.
Evidence on synaptic plasticity indicates that high-intensity acoustic stimulation can disrupt long-term potentiation (LTP), a core mechanism underlying learning and memory. In one study, a single 1-minute exposure to 110 dB audible sound spanning approximately 3–15 kHz inhibited hippocampal Schaffer-CA1 LTP for 24 hours, while spatial learning ability remained unaffected [44]. Although this sound exposure did not involve infrasound, these data suggest that similarly intense acoustic inputs may impair synaptic strengthening, most likely through alterations in postsynaptic signaling mechanisms.
Emerging evidence also points to disturbances in intracellular calcium homeostasis. For example, exposure in the 90-130 dB range was shown to alter free intracellular calcium levels and modify NMDAR1 expression in hippocampal neurons, implicating calcium dysregulation and impaired glutamatergic signaling as key mechanisms [45]. Complementary in vitro work further demonstrated that cultured hippocampal neurons developed axonal degeneration following infrasound exposure, potentially reflecting cytoskeletal compromise or excitotoxic damage [46].
Animal studies largely highlight CNS changes, whereas human data remain limited. In a randomized pilot trial, bedroom-based infrasound exposure (6 Hz, 80-90 dB) over 28 consecutive nights induced detectable changes in resting-state brain networks, though behavioural or cognitive correlates were not clearly documented [47]. Broader epidemiological surveys of low-frequency noise (including infrasound) have associated such exposures with annoyance, subjective ear pressure, and sleep disturbance, but consistent links to cognitive performance remain elusive [48]. An earlier report noted that infrasound near the auditory threshold heightened pressure sensations and annoyance, but impaired performance only in one of nine tasks [30]. Interestingly, some brief exposure studies even suggested transient improvements in working memory, though these effects proved inconsistent across replications [49].
Animal studies have shown further vulnerabilities in the auditory system. In chinchillas, simultaneous exposure to low-frequency sound (30Hz, 100 dB SPL) and high-intensity auditory noise (4 kHz octave band, 108 dB) caused greater damage to outer hair cells throughout the cochlea than exposure to auditory noise alone. These findings suggest that low-frequency components can exacerbate noise-induced cochlear damage and raise concerns about possible synergistic effects on the inner ear [50]. Low-frequency noise from environmental sources such as wind turbines, which often includes infrasound components, has been associated with disrupted sleep and reduced performance on higher-order cognitive tasks (e.g., logical reasoning, mathematical calculations). However, causality remains debated, with some reports emphasizing annoyance, restlessness, or emotional discomfort in the absence of consistent objective deficits [29,51]. The results of the Weichenberger study could provide an explanation, as could the fact that infrasound can cause a temporary increase in alertness. It has been proven that infrasound activates three centres responsible for fear, flight, emotion, and central RR and pulse regulation. Importantly, since some of these centers are involved in attentional control and arousal, it has been suggested that activation of this pathway could, for example, wake people up at night, while leaving them unable to pin down what it actually was that caused them to waken [52].
At the cellular level, rodent studies consistently implicate oxidative stress as a central driver of neuronal injury. ROS accumulation, mitochondrial dysfunction, and downstream apoptotic cascades appear to dominate. The protective effect of antioxidants such as EGCG reinforces this interpretation [43]. Beyond oxidative stress, neuroinflammatory processes have emerged as key contributors: astrocyte activation during infrasound exposure impairs learning and memory, while pharmacological blockage of astroglial connexin-43 hemichannels mitigates these deficits, highlighting the role of glial-driven inflammation in neuronal survival [53]. Similarly, activation of the FGF2/FGFR1 pathway suppresses astrocyte-derived inflammatory cytokines and neuronal loss, whereas antagonism of FGFR1 exacerbates injury through NF-κB activation [54]. Together, these data converge on astroglial NF-κB signalling and the release of proinflammatory mediators (IL-1β, TNF, and IL-6) as critical pathways.
Some studies also highlight adaptive capacity. Environmental enrichment before infrasound exposure provided robust neuroprotection in rats, preserving memory performance following 16 Hz, 130 dB exposure. This effect was accompanied by lower hippocampal oxidative stress, reduced pro-inflammatory cytokines, enhanced antioxidant defences, and decreased apoptosis, with molecular shifts favouring cell survival (increased Bcl-2, reduced p53) [55]. Such findings suggest that preconditioning strategies may buffer the brain against infrasound-induced injury.
Across rodent and cellular models, the evidence is remarkably consistent: high-intensity infrasound produces oxidative stress, neuronal apoptosis, synaptic impairment, and neuroinflammation. By contrast, human studies tend to show weaker or absent effects at moderate or near-threshold levels. This discrepancy may reflect differences in exposure intensity, interspecies sensitivity, adaptive mechanism, or methodological heterogeneity. Notably, brief human exposures have even been linked to transient cognitive enhancement [49]. In rodents, extreme intensities (≥130 dB) reliably generate profound neurotoxicity levels [56].
3.3. Other Systems
The biological effects of infrasound on organ systems beyond the cardiovascular and CNS domains have been investigated in a variety of animal, human, and technical studies. Although the evidence is still fragmented and often derived from high-intensity exposures, several consistent patterns emerge. Below, the findings are discussed by system, highlighting major outcomes, mechanistic insights, and limitations in the current literature.
In the auditory domain, psychophysical studies in humans as early as the 1970s demonstrated that infrasound is perceptible at sufficiently high sound pressure levels, with clear thresholds across the 0.5-16 Hz range [57]. Subsequent integrative work has reinforced that audibility scales steeply with SPL [58]. Vestibular studies add an important dimension: controlled laboratory exposures have shown significant increases in postural sway and the induction of vertical nystagmus around 7 Hz at high SPLs [59]. Reports on non-aural sensations, such as chest or abdominal vibration, support when exposures are strong enough [60]. The data collectively provide strong evidence that sensory structures respond to infrasound; however, they also highlight that the effective levels are often higher than what is typically encountered in everyday environments. This situation is currently changing. Recent studies published by Professor Ken Mattsson and the Colas et al. group indicate that sound pressure levels (SPLs) have been significantly underestimated under various atmospheric conditions and near several wind turbines. Current plans for 7 MW-class wind turbines suggest a further increase in sound pressure levels, reaching up to 120 dB(Z) [61].
Animal studies suggest possible effects of infrasound on physical performance. In rats, endurance capacity decreased after repeated exposure, especially in animals with lower baseline ability, suggesting that infrasound may accelerate fatigue under certain conditions [62]. While this may reflect muscular strain, it is equally plausible that vestibular disturbance or altered proprioceptive feedback contributed to reduced performance. Evidence in humans is essentially absent, and there are no dedicated in vitro studies to clarify whether muscle fibers themselves are mechanically vulnerable.
The respiratory literature is sparse. Early human observations reported minimal changes in breathing patterns under infrasonic exposure, although some participants described discomfort or nonspecific symptoms at higher SPLs [63]. Systematic measurements of lung function or airway physiology remain lacking, and no controlled animal or cell culture models have yet clarified potential mechanistic pathways. There are also changes in the bronchial mucous membranes, as described by Alves Pereira in 2007. The images for this can be found in the first paper. The new PIEZO research now confirms that lung tissue also has a large number of PIEZO channels, i.e., it is sensitive to overstimulation [37,64].
More direct pathological evidence comes from animal work. In rats, repeated infrasound exposure at 130 dB produced slowed gastric emptying, mucosal injury, and increased nitric oxide synthase activity; partial recovery was observed after cessation of exposure [65]. These findings align with the broader pattern of oxidative and inflammatory responses described in other organ systems.
Endocrine effects have been less studied, but intriguing signals exist. For example, chronic exposure in rats increased corticosterone levels and activated brown adipose tissue metabolism without causing overt glucose intolerance [66]. This points toward possible activation of stress axes and thermogenic pathways. Immune effects are mostly inferred indirectly from studies showing oxidative stress and inflammatory markers in gastrointestinal, hepatic, or reproductive tissues. Dedicated cellular and clinical studies are still missing.
Older experiments reported frequency- and duration-dependent hepatocyte changes, with longer or more intense exposures producing irreversible alterations [67]. These findings suggest a vulnerability of hepatic tissue, possibly mediated by oxidative stress and mitochondrial dysfunction. Yet, given the age of these studies and the lack of biochemical or omics-level validation, conclusions must be cautious.
Male reproductive toxicity has been demonstrated in rodents. Repeated exposure reduced sexual behavior, suppressed testosterone, and altered the expression of steroidogenic genes [68], while other work described ultrastructural damage to testes [69]. Oxidative stress and apoptotic pathways are the most plausible mechanisms. No equivalent data exists in humans, and no in vitro germ-cell or Leydig cell studies have been published, leaving a major translational gap. According to the current state of knowledge, the importance of external forces, in particular repeated exposure to low frequencies and vibration during pregnancy, must be classified as significantly more harmful than previously assumed. The high sensitivity is related to various stages, particularly of embryonic development, which are physiologically based on an undisturbed capillary flow. Examples of particular phases of increased sensitivity are, e.g., the vasculogenesis. After differentiation of endothelial progenitor cells [70] and their fusion into a primary capillary plexus [71], the growth direction of the vascular tree is essentially regulated by shear stress of the blood stream and thus by mechanotransduction, Hahn and Schwartz [72]. Together, these data illustrate how infrasound-triggered cellular and molecular mechanisms result in dysfunction across various organ systems, as summarized in Figure 1.
4. Psychological and Behavioral Responses
Infrasound exposure, particularly generated by wind turbines for electricity production, has been associated with a range of psychological and behavioral effects, although the underlying mechanisms remain complex and context-dependent. Experimental studies have demonstrated that expectation and framing can influence symptom perception [73,74]. For instance, participants exposed to wind turbine sound containing sub-audible infrasound reported fewer symptoms and lower annoyance when provided with positive information about infrasound, whereas negative expectations increased symptom reporting, consistent with nocebo effects [74,75]. Surveys of individuals reporting low-frequency noise complaints highlight a similar pattern, with many respondents describing sensations of vibration, sleep disturbance, and secondary effects such as headaches and palpitations, despite measured sound levels often being below hearing thresholds [76]. Laboratory investigations further emphasize that the perception of infrasound varies widely between individuals: small changes in sound pressure at infrasonic frequencies can lead to large differences in perceived annoyance, and sensations beyond hearing (such as ear pressure, vibration, and discomfort) may occur even when tones are inaudible [77,78].
Beyond subjective reports, recent research has explored potential objective correlates of prolonged infrasound exposure in humans. Long-term studies with controlled exposure to inaudible infrasound (6 Hz, 80–90 dB) found no significant effects on global cognitive performance, psychiatric symptoms, or sleep quality, though subtle trends in self-reported physical weakness were noted [47]. Research and case studies have shown that wind turbine noise can diminish concentration and cognitive performance. Correlation factors include sleep disturbances, increased annoyance, and stress. Lab studies that include EEG and fMRI data show that relaxation and concentration become more difficult as infrasound levels increase. Thus, it can be assumed that in proximity to wind turbines, there is a diminished cognitive function [79]
Functional connectivity analyses revealed changes in specific brain networks, including the default mode, sensorimotor, and executive control networks, which were associated with self-reported somatic symptoms and annoyance [80]. These results suggest that while infrasound may not broadly impair psychological function in healthy individuals, it can modulate brain activity and bodily perception in ways that correlate with subjective annoyance. These findings, together with behavioral studies on expectation effects, highlight the complex interplay between perceptual, cognitive, and psychosocial factors in responses to infrasound, including individual sensitivity, contextual influences, prior information.
5. Therapeutic and Biomedical Applications
Despite the plethora of studies highlighting the negative and potentially harmful consequences of infrasound exposure (from sleep disturbances to cardiovascular and neurological effects), there is a lesser-studied area: if this powerful physical phenomenon is used in controlled, dosed, targeted settings, it can become a therapeutic and innovative tool. Just as many natural agents produce beneficial biological effects at specific ranges and under safe conditions, infrasound has the potential, at appropriate levels, to modulate vital physiological pathways and stimulate the body's repair responses. The traditional view of infrasound often sees it as a dangerous "sound pollutant", but emerging evidence suggests that these low-frequency waves can have a beneficial therapeutic effect under controlled conditions. Long et al. demonstrated in an in vivo study using an animal model (mice) that applying 16 Hz sound increased bone density and osteogenesis. Subsequently, he and Fan reported positive effects on proliferation, osteogenic differentiation, and mineralization using the same frequency on bone marrow mesenchymal stem cells in vitro [81]. Additionally, Enomoto et al. showed that not just within the infrasonic range, but beyond, one can exert vibrational stimulation that changes cellular migratory and metabolic activities, indicating that there are greater cellular activities responsive to the mechanobiological effects of the vibrational stimulation [82]. Likewise, Rachlin et al. showed that low-intensity infrasound enhanced cytotoxic effects of cisplatin by merely increasing membrane permeability and did so without causing apoptosis, which, by itself, was a claim [83]. Even so, the observation must not be made in comparison to the systems that have higher-intensity ultrasound, but a direct study of systems that have higher-intensity ultrasound has not been performed. Thus, claims of greater safety and/or selectivity in treatments are made without adequate evidence, and as such, the need for caution is apparent.
Early experimental studies in rats revealed that high-intensity low-frequency sound could reduce postoperative adhesion formation, likely through micromotion of abdominal organs that prevents fibrous bridging between tissues, without detectable adverse effects [84,85]
While results lend support to the potential of infrasound as a treatment for solid tumors, studies in immune cell-derived malignancies (such as B-cell lymphoma and leukemia) have shown no significant alterations in either cell proliferation or apoptosis. However, ultrastructural changes were observed that suggest subtle cellular effects worthy of further investigation. Compared to invasive methods, such as electroporation, infrasound presents a non-invasive technique for enhancing the delivery of chemotherapeutic agents. Long wavelengths penetrate deeply into tissue, making them particularly appropriate for targeting tumors located deep in tissue. Although promising, detailed mechanisms through which infrasound can induce membrane permeability and chemosensitizing effects are not yet well understood. Furthermore, its effect varies across different cell types and is dependent on several parameters, including frequency, sound pressure, and duration of exposure. The combination of infrasound and chemotherapy presents a promising approach in therapeutics, potentially improving the effectiveness of chemotherapeutic drugs while reducing systemic side effects. Infrasound could optimize treatment outcomes by selectively enhancing drug delivery to tumor cells [86]. A study in 2017 first demonstrated that low-intensity infrasound can inhibit the proliferation and collagen synthesis of Ang II-stimulated cardiac fibroblasts. These findings suggest the potential of low-intensity infrasound as a novel therapeutic approach to prevent or reduce cardiac fibrosis [25].
6. Occupational Health and Environmental Aspects
Occupational exposure to infrasound has been extensively documented in industrial and agricultural settings. Measurements across various machinery, including tractors, grinders, conveyors, and combine harvesters, have shown that infrasound levels often exceed recommended limits, particularly during prolonged operation [87,88]. The use of modern cab-type tractors, while reducing audible noise exposure, still exposes operators to significant infrasonic pressure, sometimes surpassing occupational exposure limits within minutes of operation [88]. In addition to auditory considerations, these levels may contribute to non-auditory effects, such as fatigue, sleep disturbances, and stress, underscoring the need for effective technical and organizational interventions to reduce worker exposure [89,90,91]
The accuracy of describing and predicting a phenomenon depends heavily on the measurement methods and instruments used. As Alves-Pereira (1999) and Castelo Branco and Alves-Pereira note, conventional sound-level meters often underestimate total acoustic energy. To assess potential biobehavioral effects, it is therefore essential to measure the full frequency spectrum, capturing not just what is heard but all energy present in the environment [61,92,93].
Sound-level weighting filters are commonly applied to account for the human ear’s varying sensitivity across frequencies. A-weighting emphasizes mid-range frequencies (around 500 Hz–6 kHz) where human hearing is most sensitive, attenuating low- and high-frequency sounds, and is widely used for general noise assessment. C-weighting flattens the response at higher sound levels, giving more weight to low frequencies, and is often applied in evaluating loud environments. G-weighting targets infrasound, emphasizing frequencies below 20 Hz and reflecting pressure fluctuations interacting with the body rather than perceived loudness. Z-weighting applies no frequency correction, providing a flat response across the audible and low-frequency range, essential for capturing the full energy content of environmental sound.
Salt and Kaltenbach demonstrated this underestimation in wind turbine sounds. At 10 Hz, A-weighting recorded no measurable sound, while C- and G-weighted, as well as unweighted measures, indicated pressures of 40–60 dB. In the 1-Hz range, C- and G-weighted measures remained similar, whereas the unweighted measure reached ~90 dB. These levels can stimulate inner hair cells of the basilar membrane and various body cavities, showing that sub-10 Hz turbine sounds are potent biological stimuli. The C-weighted scale, reflecting ear sensitivity at higher sound levels, provides uniform gain from ~50 Hz to ~10 kHz and may capture effects on other organ systems. In contrast, the G-weighted scale, less aligned with perceived loudness, better represents pressure fluctuations interacting with the human body’s intrinsic properties [94].
Recent regulatory proposals and risk assessment frameworks aim to mitigate occupational infrasound exposure [95,96,97]. For example, Polish standards recommend a maximum continuous G-weighted sound level of 102 dB over an 8-hour workday, with unweighted peak levels not exceeding 145 dB [97]. In contrast, Balážiková advocates using Z-weighting in the Comprehensive Method of Acoustic Risk Assessment to avoid underestimating potential health risks [95]. Although there is still no consensus on the optimal measurement approach, these discussions highlight the importance of accurately capturing low-frequency sound to guide effective occupational safety strategies.
Individual sensitivity and context play major roles in occupational risk. Studies investigating highly sensitive populations, such as paramedics, have demonstrated that personal factors, including sensory processing sensitivity, age, and experience, can influence perceived burdensomeness of noise and infrasound exposure [90]. Moreover, phenomena such as motion sickness, simulator sickness, or idiopathic environmental intolerance illustrate that symptoms may emerge even at exposure levels below conventional safety thresholds. These findings highlight the need for occupational health protocols to consider both physical measurements and psychosocial factors when assessing potential risks, particularly in environments combining noise, vibration, confinement, and high cognitive demand [90,91].
7. Concluding Remarks
Exposure to infrasound is an important environmental stressor that has received little attention and can cause significant biological impact to various body systems. Scientific evidence from cellular and animal studies, as well as a limited number of human studies, indicates that high-intensity infrasound can lead to oxidative stress, mitochondrial dysfunction, calcium accumulation, and stimulation of apoptotic pathways, which can ultimately cause tissue damage and functional disorders in the cardiovascular, nervous, and other systems. In vitro rodent models also provide compelling evidence of myocardial fibrosis, neuronal apoptosis, and oxidative imbalance in the hippocampus. Although human studies have yielded conflicting results, they indicate vascular and neurophysiological sensitivities at higher and chronic levels of exposure. Ongoing PIEZO channel research is increasingly revealing the importance of PIEZO channels in individual organs. Overall, evidence suggests that infrasound can act as a mechanical and biological pathway modulator, affecting the body’s homeostatic balance.
Author Contributions
Conceptualization, M.D. and C.L.; literature review and synthesis, M.D., E.D.P.R.; writing and original draft preparation, M.D., E.D.P.R.; writing, review, and editing, U.B.-S., J.Z., and C.L.; supervision, C.L.; project administration, C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Bernhard Scherzinger and Marcel Bremekamp for their helpful technical discussions and insights that contributed to the development of this review.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| IS | Infrasound |
| SPL | Sound pressure level |
| TRPV4 | Transient receptor potential vanilloid 4 |
| Cx43 | Connexin 43 |
| Hz | Hertz |
| ROS | Reactive oxygen species |
| NF-κB | Nuclear factor kappa B |
| TNF | Tumor necrosis factor |
| IL-1β | Interleukin 1 beta |
| IL-6 | Interleukin 6 |
| LTP | Long-term potentiation |
| AD | Alzheimer’s disease |
| PPAR-γ | Peroxisome proliferator-activated receptor gamma |
References
- Ginsberg, J. Acoustics; American Institute of Physics: Woodbury (NY), 1996; Vol. 1, pp. 1–7 p. [Google Scholar]
- Salt, AN; Hullar, TE. Responses of the ear to low frequency sounds, infrasound and wind turbines. Hear Res. 2010, 268(1–2), 12–21. [Google Scholar] [CrossRef] [PubMed]
- Leventhall, G. What is infrasound? Prog Biophys Mol Biol. 2007, 93(1–3), 130–7. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, N; Zinkin, V; Slivina, L. Biomechanical mechanisms of action of low-frequency acoustic vibrations on a person. Russian Journal of Biomechanics 2020, 24(2), 188–200. [Google Scholar] [CrossRef]
- Persinger, MA. Infrasound, human health, and adaptation: An integrative overview of recondite hazards in a complex environment. Natural Hazards 2014, 70(1), 501–25. [Google Scholar]
- Gao, W; Hasan, H; Anderson, DE; Lee, W. The Role of Mechanically-Activated Ion Channels Piezo1, Piezo2, and TRPV4 in Chondrocyte Mechanotransduction and Mechano-Therapeutics for Osteoarthritis. In Frontiers in Cell and Developmental Biology; Frontiers Media S.A., 2022; Vol. 10. [Google Scholar]
- Lewis, AH; Cui, AF; McDonald, MF; Grandl, J. Transduction of Repetitive Mechanical Stimuli by Piezo1 and Piezo2 Ion Channels. Cell Rep. 2017, 19(12), 2572–85. [Google Scholar] [CrossRef] [PubMed]
- Sianati, S; Schroeter, L; Richardson, J; Tay, A; Lamandé, SR; Poole, K. Modulating the Mechanical Activation of TRPV4 at the Cell-Substrate Interface. In Front Bioeng Biotechnol; 18 Jan 2021; p. 8. [Google Scholar]
- Folwaczny, M; Steinecker-Frohnwieser, B; Lohberger, B; Toegel, S; Windhager, R; Glanz, V; et al. Activation of the Mechanosensitive Ion Channels Piezo1 and TRPV4 in Primary Human Healthy and Osteoarthritic Chondrocytes Exhibits Ion Channel Crosstalk and Modulates Gene Expression; 2023. [Google Scholar]
- Shahidullah, M; Delamere, NA. Mechanical Stretch Activates TRPV4 and Hemichannel Responses in the Nonpigmented Ciliary Epithelium. Int J Mol Sci. 2023, 24(2). [Google Scholar]
- Kim, YJ; Hyun, J. Role of Piezo1 in cell death.
- Albarrán-Juárez, J; Iring, A; Wang, SP; Joseph, S; Grimm, M; Strilic, B; et al. Piezo1 and Gq/G11 promote endothelial inflammation depending on flow pattern and integrin activation. Journal of Experimental Medicine 2018, 215(10), 2655–72. [Google Scholar] [CrossRef]
- He, R; Liu, Y; Fan, J. The role of Notch signaling pathway in mechanical stimulation of infrasound altering activities of bone marrow mesenchymal stem cells [Internet]. Int J Clin Exp Pathol 2017, Vol. 10. [Google Scholar]
- He, R; Fan, J. Effects of infrasound on the growth of bone marrow mesenchymal stem cells: A pilot study. Mol Med Rep. 2014, 10(5), 2427–32. [Google Scholar]
- Wang, H; Wang, J; Yang, Q; Zhang, X; Gao, P; Xu, S; et al. Synthesis of a Novel Nitronyl Nitroxide Radical and Determination of its Protective Effects Against Infrasound-Induced Injury. Neurochem Res [Internet] 2015, 40(7), 1526–36. [Google Scholar] [PubMed]
- Qui, P; Li, Y; Gao, W; Guo, Q; Zhang, ZM; Jiang, Y; et al. Effects of infrasound on Ca2+ -activated-K+ channel of the bovine retinal microvascular endothelial cells. Chinese Journal of Clinical Rehabilitation [Internet] 2005, 9(6), 198–9. [Google Scholar]
- Zou, LH; Shi, YJ; He, H; Jiang, SM; Huo, FF; Wang, XM; et al. Effects of FGF2/FGFR1 pathway on expression of A1 astrocytes after infrasound exposure. Front Neurosci. 2019, 13(MAY). [Google Scholar] [CrossRef]
- Martirosyan, V; Markosyan, L; Hovhanesyan, H; Hovnanyan, K; Ayrapetyan, S. The frequency-dependent effect of extremely low-frequency electromagnetic field and mechanical vibration at infrasound frequency on the growth, division and motility of Escherichia coli K-12. Environmentalist 2012, 32(2), 157–65. [Google Scholar] [CrossRef]
- Ayrapetyan, S; De, J. Cell Hydration as a Biomarker for Estimation of Biological Effects of Nonionizing Radiation on Cells and Organisms. Scientific World Journal 2014. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z; Sang, H; Li, R; Xiao, P; He, J; Zhuang, Z; et al. Infrasound-induced hemodynamics, ultrastructure, and molecular changes in the rat myocardium. Environ Toxicol. 2007, 22(2), 169–75. [Google Scholar] [CrossRef]
- Pei, ZH; Chen, BY; Tie, R; Zhang, HF; Zhao, G; Qu, P; et al. Infrasound exposure induces apoptosis of rat cardiac myocytes by regulating the expression of apoptosis-related proteins. Cardiovasc Toxicol 2011, 11(4), 341–6. [Google Scholar] [CrossRef]
- Pei, Z; Meng, R; Zhuang, Z; Zhao, Y; Liu, F; Zhu, MZ; et al. Cardiac peroxisome proliferator-activated receptor-γ expression is modulated by oxidative stress in acutely infrasound-exposed cardiomyocytes. Cardiovasc Toxicol 2013, 13(4), 307–15. [Google Scholar] [CrossRef] [PubMed]
- Lousinha, A; Maria, MJ; Borrecho, G; Brito, J; Oliveira, P; Oliveira de Carvalho, A; et al. Infrasound induces coronary perivascular fibrosis in rats. Cardiovascular Pathology 2018, 37, 39–44. [Google Scholar] [CrossRef]
- Antunes, E; Oliveira, P; Borrecho, G; Oliveira, MJR; Brito, J; Águas, A; et al. Myocardial fibrosis in rats exposed to low frequency noise. Acta Cardiol. 2013, 68(3), 241–5. [Google Scholar] [CrossRef]
- Jin, W; Deng, QQ; Chen, BY; Lu, ZX; Li, Q; Zhao, HK; et al. Inhibitory effects of low decibel infrasound on the cardiac fibroblasts and the involved mechanism. Noise Health 2017, 19(88), 149–53. [Google Scholar] [PubMed]
- Chaban, R; Ghazy, A; Georgiade, E; Stumpf, N; Vahl, CF. Negative effect of high-level infrasound on human myocardial contractility: In-vitro controlled experiment. Noise Health 2021, 23(109), 57–66. [Google Scholar] [CrossRef]
- DANIELSSON, Å; LANDSTRÖM, U. Blood Pressure Changes in Man during Infrasonic Exposure: An Experimental Study. Acta Med Scand. 1985, 217(5), 531–5. [Google Scholar] [CrossRef]
- Yuan, C; Qibai, H; Shi, H. An Investigation on the Physiological and Psychological Effects of Infrasound on Persons; 2004. [Google Scholar]
- Møller, H; M<t>ller, Henrik. Physiological and psychological effects of infrasound on humans. Journal of Low Frequency Noise, Vibration and Active Control. 1984, Vol. 3. Available online: http://lfn.sagepub.com/content/3/1/1.refs.
- Birnie, S; Hall, F; Taylor, S. The effects of infrasound on human health. Canadian Acoustics cited. 1983, 11(4), 46–55. [Google Scholar]
- Health Canada. Wind Turbine Noise and Health Study: Summary of Results, Background and Rationale. 2019. [Google Scholar]
- Marshall, NS; Cho, G; Toelle, BG; Tonin, R; Bartlett, DJ; D’rozario, AL; et al. The Health Effects of 72 Hours of Simulated Wind Turbine Infrasound: A Double-Blind Randomized Crossover Study in Noise-Sensitive, Healthy Adults. Environ Health Perspect 2023, 131(3). [Google Scholar] [CrossRef]
- Chiu, CH; Lung, SCC; Chen, N; Hwang, JS; Tsou, MCM. Effects of low-frequency noise from wind turbines on heart rate variability in healthy individuals. Sci Rep. 2021, 11(1). [Google Scholar] [CrossRef] [PubMed]
- Deng, Y; Ohgami, N; Kagawa, T; Kurniasari, F; Chen, D; Kato, M; et al. Vascular endothelium as a target tissue for short-term exposure to low-frequency noise that increases cutaneous blood flow. Science of the Total Environment, 2022 Dec 10; p. 851. [Google Scholar]
- Bräuner, E V.; Jørgensen, JT; Duun-Henriksen, AK; Backalarz, C; Laursen, JE; Pedersen, TH; et al. Long-term wind turbine noise exposure and the risk of incident atrial fibrillation in the Danish Nurse cohort. Environ Int. 2019, 130. [Google Scholar] [CrossRef] [PubMed]
- Koutsiaris, AG; Riri, K; Boutlas, S; Daniil, Z; Tsironi, EE. A normative blood velocity model in the exchange microvessels for discriminating health from disease: Healthy controls versus COVID-19 cases. Clin Hemorheol Microcirc. 2023, 84(2), 215–26. [Google Scholar] [CrossRef] [PubMed]
- Fang, XZ.; Zhou, T.; Xu, JQ.; et al. Structure, kinetic properties and biological function of mechanosensitive Piezo channels. Cell Biosci 2021, 11, 13. [Google Scholar] [CrossRef]
- Glazkov, AA; Ulbashev, DS; Borshchev, GG; Pulin, AA; Glazkova, PA; Kulikov, DA. Skin microcirculation reactivity to local thermal hyperaemia in patients with COVID-19 - A pilot observational study. Clin Hemorheol Microcirc. 2022, 83(1), 19–29. [Google Scholar] [CrossRef]
- Krausova, V; Neumann, D; Kraus, J; Dostalova, V; Dostal, P. Sublingual microcirculation in healthy pediatric population using the sidestream dark-field imaging method. Clin Hemorheol Microcirc. 2023, 85(2), 163–71. [Google Scholar] [CrossRef]
- Yang, YP; Chiu, CT; Chao, A; Yeh, YC; Sun, WZ; Liu, CM; et al. An observational study of microcirculation among healthy individuals by age and sex. Clin Hemorheol Microcirc. 2024, 86(4), 407–17. [Google Scholar] [CrossRef]
- ZHANG, MY; CHEN, C; XIE, XJ; XU, SL; GUO, GZ; WANG, J. Damage to Hippocampus of Rats after Being Exposed to Infrasound. Biomedical and Environmental Sciences 2016, 29(6), 435–42. [Google Scholar] [PubMed]
- Zhou, X; Yang, Q; Song, F; Bi, L; Yuan, J; Guan, S; et al. Tetrahydroxystilbene Glucoside Ameliorates Infrasound-Induced Central Nervous System (CNS) Injury by Improving Antioxidant and Anti-Inflammatory Capacity. Oxid Med Cell Longev 2020. [Google Scholar] [CrossRef]
- Cai, J; Jing, D; Shi, M; Liu, Y; Lin, T; Xie, Z; et al. Epigallocatechin gallate (EGCG) attenuates infrasound-induced neuronal impairment by inhibiting microglia-mediated inflammation. Journal of Nutritional Biochemistry 2014, 25(7), 716–25. [Google Scholar] [CrossRef]
- De Deus, JL; Cunha, AOS; Terzian, AL; Resstel, LB; Elias, LLK; Antunes-Rodrigues, J; et al. A single episode of high intensity sound inhibits long-term potentiation in the hippocampus of rats. Sci Rep. 2017, 7(1). [Google Scholar] [CrossRef]
- Liu, ZH; Chen, JZ; Ye, L; Liu, J; Qiu, JY; Xu, J; et al. Effects of infrasound at 8 Hz 90 dB/130 dB on NMDAR1 expression and changes in intracellular calcium ion concentration in the hippocampus of rats. Mol Med Rep. 2010, 3(6), 917–21. [Google Scholar]
- Cheng, H; Wang, B; Tang, C; Feng, G; Zhang, C; Li, L; et al. Infrasonic noise induces axonal degeneration of cultured neurons via a Ca 2+ influx pathway. Toxicol Lett. 2012, 212(2), 190–7. [Google Scholar] [CrossRef]
- Ascone, L; Kling, C; Wieczorek, J; Koch, C; Kühn, S. A longitudinal, randomized experimental pilot study to investigate the effects of airborne infrasound on human mental health, cognition, and brain structure. Sci Rep. 2021, 11(1). [Google Scholar] [CrossRef] [PubMed]
- Alves, JA; Paiva, FN; Silva, LT; Remoaldo, P. Low-frequency noise and its main effects on human health-A review of the literature between 2016 and 2019. In Applied Sciences (Switzerland); MDPI AG, 2020; Vol. 10. [Google Scholar]
- Weichenberger, M; Kühler, R; Bauer, M; Hensel, J; Brühl, R; Ihlenfeld, A; et al. Brief bursts of infrasound may improve cognitive function - An fMRI study. Hear Res. 2015, 328, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Harding, GW; Bohne, BA; Lee, SC; Salt, AN. Effect of infrasound on cochlear damage from exposure to a 4 kHz octave band of noise. Hear Res. 2007, 225(1–2), 128–38. [Google Scholar] [CrossRef]
- Liang, P; Li, J; Li, Z; Wei, J; Li, J; Zhang, S; et al. Effect of low-frequency noise exposure on cognitive function: a systematic review and meta-analysis. BMC Public Health 2024, 24(1). [Google Scholar] [CrossRef]
- Weichenberger, M; Bauer, M; Kuhler, R; Hensel, J; Garcia Forlim, C; Ihlenfeld, A; et al. Altered cortical and subcortical connectivity due to infrasound administered near the hearing threshold-Evidence from fMRI. PLoS One 2017, 12(4). [Google Scholar] [CrossRef] [PubMed]
- Zhang, W; Yin, J; Gao, BY; Lu, X; Duan, YJ; Liu, XY; et al. Inhibition of astroglial hemichannels ameliorates infrasonic noise induced short-term learning and memory impairment. Behavioral and Brain Functions 2023, 19(1). [Google Scholar] [CrossRef] [PubMed]
- Shi, YJ; Shi, M; Xiao, LJ; Li, L; Zou, LH; Li, CY; et al. Inhibitive effects of FGF2/FGFR1 pathway on astrocyte-mediated inflammation in vivo and in vitro after infrasound exposure. Front Neurosci 2018, 12(AUG). [Google Scholar] [CrossRef]
- Jiang, S; Wang, YQ; Tang, YF; Lu, X; Guo, D. Pre-Exposure to Environmental Enrichment Protects against Learning and Memory Deficits Caused by Infrasound Exposure. Oxid Med Cell Longev 2022. [Google Scholar] [CrossRef]
- Araújo, Amilcar; Carranca, Joaquim; Alves-Pereira, Mariana; Castelo Branco, Nuno A. A. ECHOCARDIOGRAPHY IN VIBROACOUSTIC DISEASE; 2005. [Google Scholar]
- Yeowart, NS; Evans, MJ. Thresholds of audibility for very low-frequency pure tones. J Acoust Soc Am. 1974, 55(4), 814–8. [Google Scholar] [CrossRef]
- Møller, H; Pedersen, CS. Hearing at Low and Infrasonic Frequencies; 2004. [Google Scholar]
- Hiroshi, Takigawa; Fumiyo, Hayashi; Shizuko, Sugiura; Hiroshi, Sakamoto. Effects of Infrasound on Human Body Sway. Journal of Low Frequency Noise, Vibration and Active Control [Internet] 1988, 7(2), 66–73. [Google Scholar]
- Landström, Ulf; Lundström, Ronnie; Byström, Marianne. Exposure to Infrasound — Perception and Changes in Wakefulness. Journal of Low Frequency Noise, Vibration and Active Control [Internet] 1983, 2(1), 1–11. [Google Scholar]
- Mattsson, K; Eriksson, G; Persson, L; Chilo, J; Tatar, K. Efficient finite difference modeling of infrasound propagation in realistic 3D domains: Validation with wind turbine measurements. Applied Acoustics, 2026 Feb 5; 243. [Google Scholar]
- Yamamura, K; Kishi, R. Effects of infrasound on the rota-rod treadmill performance of rats. Eur J Appl Physiol Occup Physiol. 1980, 45(1), 81–6. [Google Scholar] [CrossRef]
- Okamoto, K; Yoshida, A; Inoue, J; Takyu, H. [The influence of infrasound upon human body]. J UOEH 1986, 8 Suppl, 135–49. [Google Scholar] [PubMed]
- Alves-Pereira, M; Castelo Branco, NAA. Vibroacoustic disease: Biological effects of infrasound and low-frequency noise explained by mechanotransduction cellular signalling; Progress in Biophysics and Molecular Biology, 2007; Vol. 93, pp. 256–79. [Google Scholar]
- Zhao, JH; Wang, JH; Luo, JY; Guo, XY; Wang, Y; Cheng, Y. Effects of Infrasound on Gastric Motility, Gastric Morphology and Expression of Nitric Oxide Synthase in Rat. Biomed Environ Sci 2018, 31(5), 399–402. [Google Scholar]
- Pereira, GM; Santos, M; Pereira, SS; Borrecho, G; Tortosa, F; Brito, J; et al. High-intensity infrasound effects on glucose metabolism in rats. Sci Rep. 2021, 11(1). [Google Scholar]
- Nekhoroshev, AS; Glinchikov, V V. [Morphological research on the liver structures of experimental animals under the action of infrasound]. Aviakosm Ekolog Med. 1992, 26(3), 56–9. [Google Scholar]
- Zhuang, Z; Pei, Z; Chen, J. Infrasound-induced changes on sexual behavior in male rats and some underlying mechanisms. Environ Toxicol Pharmacol 2007, 23(1), 111–4. [Google Scholar] [CrossRef]
- Wei, YN; Liu, J; Shu, Q; Huang, XF; Chen, JZ. [Effects of infrasound on ultrastructure of testis cell in mice]. Zhonghua Nan Ke Xue 2002, 8(5), 323–5, 328. [Google Scholar] [PubMed]
- Bellut-Staeck, U. Die Mikrozirkulation und ihre Bedeutung für alles Leben [Internet]. In (essentials); Springer Berlin Heidelberg: Berlin, Heidelberg, 2022. [Google Scholar]
- Belvitch, P; Htwe, YM; Brown, ME; Dudek, S. Cortical Actin Dynamics in Endothelial Permeability. In Current Topics in Membranes; Academic Press Inc., 2018; pp. 141–95. [Google Scholar]
- Hahn, C; Schwartz, MA. Mechanotransduction in vascular physiology and atherogenesis; Nature Reviews Molecular Cell Biology, 2009; Vol. 10, pp. 53–62. [Google Scholar]
- Crichton, F; Chapman, S; Cundy, T; Petrie, KJ. The link between health complaints and wind turbines: Support for the nocebo expectations hypothesis. Front Public Health [Internet] 2014, 2(NOV). [Google Scholar]
- Crichton, F; Petrie, KJ. Health complaints and wind turbines: The efficacy of explaining the nocebo response to reduce symptom reporting. Environ Res [Internet] 2015, 140, 449–55. [Google Scholar]
- Crichton, F; Dodd, G; Schmid, G; Petrie, KJ. Framing sound: Using expectations to reduce environmental noise annoyance. Environ Res [Internet] 2015, 142, 609–14. [Google Scholar]
- Møller, H; Lydolf, M. A questionnaire survey of complaints of infrasound and low-frequency noise. Journal of Low Frequency Noise Vibration and Active Control [Internet] 2002, 21(2), 53–64. [Google Scholar]
- Andresen, J; Moller, H. Equal annoyance contours for infrasonic frequencies. Journal of Low Frequency Noise and Vibration [Internet] 1984, 3(3), 1–9. [Google Scholar] [CrossRef]
- Rajala, V; Hakala, J; Alakoivu, R; Koskela, V; Hongisto, V. Hearing threshold, loudness, and annoyance of infrasonic versus non-infrasonic frequencies. Applied Acoustics 2022, 198. [Google Scholar] [CrossRef]
- Dumbrille, A; McMurtry, RY; Krogh, CM. Wind turbines and adverse health effects. Environ Dis. 2021, 6(3), 65–87. [Google Scholar] [CrossRef]
- Forlim, CG; Ascone, L; Koch, C; Kühn, S. Resting state network changes induced by experimental inaudible infrasound exposure and associations with self-reported noise sensitivity and annoyance. Sci Rep. 2024, 14(1). [Google Scholar] [CrossRef] [PubMed]
- He, R; Fan, J. Effects of infrasound on the growth of bone marrow mesenchymal stem cells: A pilot study. Mol Med Rep. 2014, 10(5), 2427–32. [Google Scholar] [CrossRef]
- Enomoto, U; Imashiro, C; Takemura, K. Collective cell migration of fibroblasts is affected by horizontal vibration of the cell culture dish. Eng Life Sci. 2020, 20(9–10), 402–11. [Google Scholar] [CrossRef]
- Rachlin, K; Moore, DH; Yount, G. Infrasound sensitizes human glioblastoma cells to cisplatin-induced apoptosis. Integr Cancer Ther. 2013, 12(6), 517–27. [Google Scholar] [CrossRef]
- Colasante, DA; Au, FC; Sell, HW; Tyson, RR. Prophylaxis of adhesions with low frequency sound. Surg Gynecol Obstet. 1981, 153(3), 357–9. [Google Scholar]
- Zhang, H; Qi, P; Si, SY; Ma, WM. Effect of infrasound on the growth of colorectal carcinoma in mouse. Chinese Journal of Cancer Prevention and Treatment 2013, 20, 1145–9. [Google Scholar]
- Vahl, JM; von Witzleben, A; Reiter, R; Theodoraki, MN; Wigand, M; Hoffmann, TK; et al. Infrasound a new weapon in cancer therapy? Explore [Internet] 2022, 18(3), 366–70. [Google Scholar]
- Bilski, B. Exposure to infrasonic noise in agriculture. Annals of Agricultural and Environmental Medicine 2017, 24(1), 86–9. [Google Scholar] [CrossRef] [PubMed]
- Bilski, B. Audible and infrasonic noise levels in the cabins of modern agricultural tractors - Does the risk of adverse, exposure-dependent effects still exist? Int J Occup Med Environ Health 2013, 26(3), 488–93. [Google Scholar] [CrossRef]
- Cybulski, M; Wiecheć, K; Zieliński, M; Bilski, B. How paramedics perceive internal noise in ambulance? sensory processing sensitivity (SPS) and Subjective Noise Assessment. Noise Health 2019, 21(103), 242–7. [Google Scholar] [PubMed]
- Van Den Bergh, O; Bräscher, AK; Witthöft, M; Gutenberg, J. Idiopathic Environmental Intolerance: A Treatment Model. [CrossRef]
- Kalandrik, D; Jacob, R; Saunders, M; Quaiser-Pohl, C; Kap-herr, KH; von; Koenig, P. Investigation of Acoustic Influencing Parameters to Increase the Human Performance Potential in Simultaneous Driving and Working Operation of Armoured Vehicles. International Journal of Vehicle Structures and Systems 2021, 13(3). [Google Scholar] [CrossRef]
- Castelo Branco, NAA; Monteiro, E; Silva, ACE; Dos Santos, JM; Reis Ferreira, JM; Alves-Pereira, M. The lung parenchyma in low frequency noise exposed wistar rats. Rev Port Pneumol [Internet] 2004, 10(1), 77–85. [Google Scholar]
- Colas, J; Emmanuelli, A; Dragna, D; Stevens, RJAM. Modeling wind farm noise emission and propagation: effects of flow and layout; 19 Aug 2025. [Google Scholar]
- Salt, AN; Hullar, TE. Responses of the ear to low frequency sounds, infrasound and wind turbines. Hear Res. 2010, 268(1–2), 12–21. [Google Scholar] [CrossRef]
- Balážiková, M; Pačaiová, H; Tomašková, M. A Proposal for Risk Assessment of Low-Frequency Noise in the Human–Machine–Environment System. Applied Sciences (Switzerland) 2023, 13(24). [Google Scholar] [CrossRef]
- Pawlaczyk-Luszczynska, M. Evaluation of occupational exposure to infrasonic noise in Poland. Int J Occup Med Environ Health 1999, 12(2), 159–76. [Google Scholar]
- Pawlaczyk-ŁUszczýnska, M; Kaczmarska-Kozłowska, A; Augustyńska, D; Kameduła, M. Proposal of new limit values for occupational exposure to infrasonic noise in Poland. Journal of Low Frequency Noise Vibration and Active Control. 2000, 19(4), 183–93. [Google Scholar] [CrossRef]
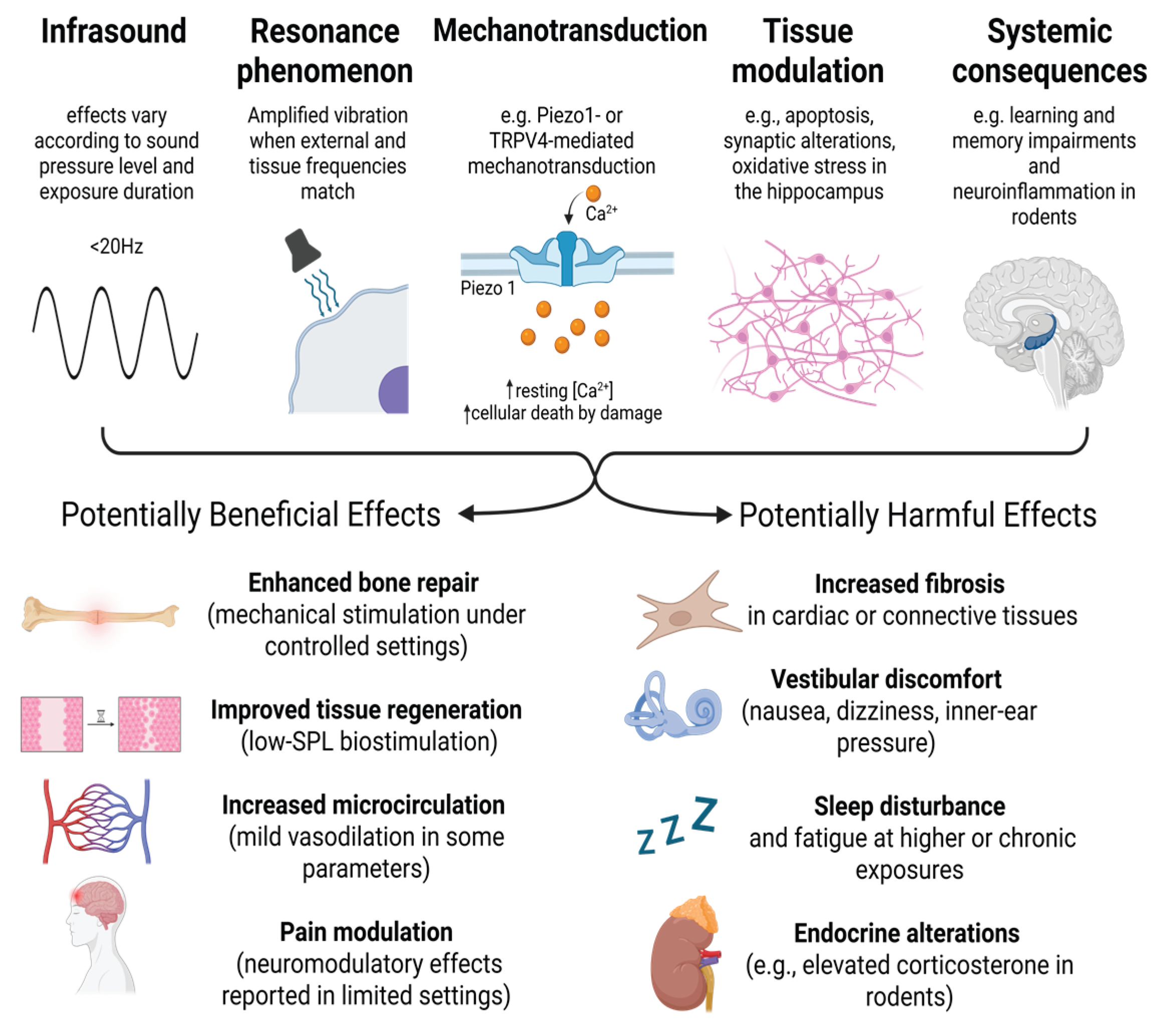
Figure 1.
Overview of the biological mechanisms of infrasound. SPL- Sound pressure level, TRPV4 - transient receptor potential cation channel subfamily V member 4. Created with BioRender.com.
Figure 1.
Overview of the biological mechanisms of infrasound. SPL- Sound pressure level, TRPV4 - transient receptor potential cation channel subfamily V member 4. Created with BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



