
Submitted:
12 January 2026
Posted:
14 January 2026
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and influenza virus are dangerous respiratory pathogens with high pandemic potential. Since 2021, these two viruses have been co-circulating, which implies additional risks of co-infection with both pathogens. According to clinical data, influenza and SARS-CoV-2 cause sim-ilar symptoms, and co-infection can increase disease severity and significantly enhance the risks of pneumonia and acute respiratory distress syndrome progressing with a poor outcome. Therefore, management of such patients requires special consideration. Prophylactic vaccination is widely recognized as the most effective way to prevent COVID-19 and influenza and to reduce the severity of these diseases. A range of influ-enza and COVID-19 vaccines built on different technological platforms is currently available on the market, with proven effectiveness, immunogenicity, and safety. Im-portantly, multiple countries have approved recommendations for simultaneous vac-cination against both viral pathogens. This approach is more convenient for patients and is associated with better response to treatment, while also improving vaccine cov-erage and compliance and offering significant resource savings for healthcare systems. This review analyzes recent data on the simultaneous circulation of influenza and SARS-CoV-2 viruses worldwide. We review epidemiological data and the pathogenetic mechanisms of co-infection with these two viruses. Next, we focus on current ap-proaches to simultaneous and combined vaccination against influenza and COVID-19. We outline the types of vaccines and summarize the available findings on the effec-tiveness and safety of co-vaccination.
Keywords:
SARS-CoV-2
; influenza virus
; co-circulation
; co-infection
; simultaneous vaccination
; next-generation vaccines
1. Introduction
Influenza A virus (IAV) is a widespread and dangerous pathogen that causes seasonal epidemics and periodic global pandemics with serious medical and socio-economic consequences [1]. Seasonal epidemics are associated with the circulation of IAV subtypes H1N1 and H3N2. Each year, influenza causes between 3 to 5 million cases of severe illness and 290,000 to 650,000 deaths from respiratory causes [2]. Of especial WHO concern is the risk of zoonotic transmission to humans of other IAV subtypes, primarily the highly pathogenic H5N1 avian influenza, clade 2.3.4.4 b, due to the high mortality rate during human infection and the significant pandemic potential of this pathogen [3].
In addition to the flu, in recent years the world has faced the emergence of the SARS-CoV-2 virus and the unprecedented COVID-19 pandemic. According to WHO data, from 31.12.2019 to 07.12.2025, 778,994,897 cases of COVID-19 in humans were officially confirmed worldwide, including 7,106,996 COVID-19 deaths (0.9% of the number of cases) [4]. The European region ranks first among the regions of the world in terms of the total number of detected cases. In Russia, from 03.02.2020 to 05.05.2024, 24,184,485 cases of COVID-19 were registered in 85 regions of the country [5]. The COVID-19 pandemic has had a dramatic negative impact on the global economy and health systems around the world.
Though the COVID-19 pandemic officially ended in 2023 [6], people continue to encounter this virus. SARS-CoV-2 is evolving, and new contagious strains are emerging, so the healthcare system must be prepared for different scenarios. Apparently, COVID-19 has become a seasonal epidemic, and both influenza and SARS-CoV-2 viruses will continue to represent a significant threat to public health for the foreseeable future. In terms of epidemiology, COVID-19 seems to exhibit seasonal trends similar to influenza. SARS-CoV-2 and influenza viruses can co-circulate in autumn and winter, leading to overlapping outbreaks [7]. The dynamic circulation of influenza viruses in conjunction with SARS-CoV-2 is registered all over the world, including in Russia. This raises concerns about the potential risk of infection by each of these two pathogens or co-infection by both of them, which significantly increases the risk to public health.
Despite the differences in the entry mechanisms into target cells, similar modes of transmission and infection sites in the respiratory tract are characteristic of both viral pathogens [8,9]. Influenza and COVID-19 exhibit a generally similar array of symptoms, such as fever, sore throats, chills, chest and abdominal pain, vomiting, diarrhea, nasal symptoms, and loss of taste [10,11,12]. The simultaneous or sequential presence of these two pathogens in lung tissue enhanced the risk of serious complications and aggravation of the clinical picture of the disease. Studies conducted in several countries around the world have documented cases of co-infection with SARS-CoV-2 and influenza viruses of types A or B [13]. Co-infections cause reasonable concern among specialists, as they can worsen the course of the disease and introduce additional difficulties into the treatment process. Co-infection with SARS-CoV-2 and influenza A virus leads to a worsening of respiratory ailments such as pneumonia, sinus infections, and bronchitis and increases the danger of acute respiratory failure and cardiovascular complications [14,15]. The mechanisms that cause a more severe course of the disease in co-infection can be multifactorial, including an imbalance in immune regulation, which, in turn, can lead to more significant damage to lung tissue and increased vulnerability to the development of acute respiratory distress syndrome [16,17]. Thus, in the case of co-infection, one of the viruses can enhance the pathogenic effects of the other, which makes it difficult to accurately interpret the clinical manifestations of the disease.
Vaccination remains the most effective strategy to mitigate the burden of both influenza and COVID-19. Simultaneous administration of influenza and COVID-19 vaccines is being actively explored as a way to improve immunization coverage and streamline public health efforts. In October 2021, WHO approved the combined use of vaccines against COVID-19 and seasonal influenza [18]. The Russian Ministry of Health has also authorized simultaneous vaccination and updated the instructions for use of the vaccine "Sputnik V" [19]. Simultaneous vaccination is the administration of several vaccines on the same clinical day in different anatomical areas, while it is possible to use vaccines with different routes of administration (for example, intramuscularly and intranasally). The simultaneous use of influenza and SARS-CoV-2 vaccines provides more benefits than either of them alone. Clinical trials have further confirmed the feasibility of simultaneous vaccination against influenza and SARS-CoV-2 as an effective approach to immunization [20]. However, it is important to consider such factors as vaccine compatibility, as well as differences in storage requirements and administration schedules.
Another approach is to use combination vaccines, which consist of two or more components that are physically combined and injected simultaneously into the same anatomical site. Combined vaccines targeting both SARS-CoV-2 and influenza are an important area in the new vaccines’ development. With that, it should be considered that the combination of several vaccines can affect the immunogenic properties of individual antigens. Simultaneous vaccination or the use of combined vaccines is more economical, as well as more convenient for patients. A number of vaccines under development have confirmed their immunogenicity in preclinical trials, and several are undergoing clinical trials.
Our review analyzes recent data on the co-circulation of influenza and SARS-CoV-2 viruses worldwide. We highlight the pathogenetic mechanisms of co-infection with these two viruses. Next, we explore the rationale, current evidence, and future potential of simultaneous and combination vaccination strategies targeting influenza and SARS-CoV-2 as an effective approach to immunization. We outline the types of vaccines on various platforms and summarize the available findings on the effectiveness and safety of co-vaccination.
2. SARS-CoV-2 and Influenza Virus Co-Circulation and Co-Infection
The emergence of SARS-CoV-2 in 2019 had a significant impact on the global epidemiology of respiratory viruses, including influenza (Figure 1). After a period of practical disappearance from circulation in 2020 and the first half of 2021, influenza viruses resumed circulation at the end of 2021 and, according to epidemiological data, continue to show pronounced genetic variability. As per surveillance data, during the COVID-19 pandemic, the circulation of influenza viruses is reported worldwide (Figure 1A) [21], including in Russia (Figure 1B) [22]. To investigate the impact of SARS-CoV-2 on influenza activity worldwide, WHO global surveillance data [23,24] were analyzed to compare the number of cases of COVID-19 and influenza in 22 different countries. In that work, Takashita et al. claimed that though both viruses co-circulate, there is rather an alternating pattern in the prevalence of SARS-CoV-2 and influenza virus during the reported period [25].
The dynamic circulation raises concerns about the risk of infection with either of these two pathogens as well as the risk of co-infection with two viruses. In the context of co-circulation, it is important to understand the mechanisms of interaction between SARS-CoV-2 and influenza viruses as well as the consequences of such concomitant infections. In general, competitive interference has been described in the case of co-circulation between respiratory viruses, notably influenza [26]. Possible mechanisms underlying these processes can be associated with competition of viruses for host cell resources and the activation of the immune response [27,28,29]. Studies have indicated a significant contribution of the innate immune response to inter-viral interactions, especially when co-infections occur simultaneously or consistently close in time [29,30].
An in vitro approach using the human epithelial airway cells has shown that in the case of sequential infection with SARS-CoV-2 and influenza, respiratory syncytial virus (RSV), or rhinovirus, the type of virus and the order of infections are crucial factors in virus-virus interactions. In this regard, primary infection plays a significant role in shaping the immune response and influencing the outcome of subsequent infections [30]. Pinky et al. created a mathematical model of co-infection with two viruses and showed that SARS-CoV-2 has a lower rate of spread than influenza and can be suppressed if infections start simultaneously. However, if influenza infection occurs later than SARS-CoV-2 infection starts, co-infection may appear [31].
The results of a recent study have shown that the influenza A virus can promote SARS-CoV-2 virus infectivity due to an increase in the expression of angiotensin-converting enzyme 2 (ACE2), allowing the SARS-CoV-2 virus to enter the cells [32]. In addition to the similarities in symptoms between the two infections, SARS-CoV-2 specifically infects type II alveolar cells (AT2 pneumocytes), which are also the site of IAV replication [33,34]. The study by Ziegler et al. analyzed single-cell RNA sequencing data from human lung explants infected with the influenza A virus ex vivo for 24 hours. The data obtained established that ACE2 is expressed within type II pneumocytes. Furthermore, after IAV infection, ACE2 expression was elevated [35]. Such elevation of ACE2 expression and of SARS-CoV-2 infectivity is characteristic only of IAV, but not of other respiratory viruses [32].
In animal models, co-infection with influenza A/H1N1 and SARS-CoV-2 viruses clearly prolongs COVID-19 symptoms, causes more severe and prolonged pneumonia, and increases lung damage and weight loss, leading to high mortality [36,37,38].
Clinically, co-infection of the SARS-CoV-2 and influenza A viruses was first reported in a Chinese patient suffering from pneumonia in 2020 [39]. Later, numerous case reports on the co-infection from medical centers around the world were published. These results were summarized in several informative meta-analytical reviews [13,40,41]. According to the results of a recent comprehensive meta-analysis by Golpour et al. [13], the average prevalence of influenza A/B co-infection in COVID-19-positive patients was 14 percent, which is certainly a significant value that requires the careful attention of medical personnel.
Numerous clinical observations clearly indicate that SARS-CoV-2 and influenza co-infection has been associated with significantly worse outcomes, posing a serious public health challenge [24,42]. Given the concurrent circulation, in terms of diagnostics, it is important to consider the COVID-19 co-infection possibility, even if other respiratory viral pathogens have been identified. Particularly, Ma et al. found that a significant number of critically ill COVID-19 patients were co-infected with influenza, potentially leading to earlier cytokine storms and organ damage [43]. Studies have shown that influenza and SARS-CoV-2 co-infection distinctly contribute to the severe form of the disease and, as a result, increase mortality rates [44,45].
Thus, clinical investigations indicate that co-circulation of influenza and SARS-CoV-2 significantly increases public health risk, whether infections occur individually or concurrently. Simultaneous or combined vaccinations are important strategies for preventing influenza and COVID-19, reducing the risk of infection and co-infection, and minimizing the burden on medical care.
3. Concepts of Simultaneous and Combined Vaccination Against COVID-19 and Influenza
Simultaneous vaccination is the administration of multiple vaccines on the same clinic day at different anatomic sites, aiming to induce immune responses to each vaccine while reducing the burden of multiple visits [46,47]. Simultaneous vaccination offers several benefits, including convenience, improved coverage, enhanced compliance, and cost-effectiveness. It reduces a number of required visits, diminishes missed doses, and improves adherence to recommended immunization schedules [48,49]. Additionally, simultaneous vaccination may increase vaccine uptake without compromising the effectiveness of either vaccine [50].
However, when considering simultaneous vaccination, healthcare professionals should account for various factors, such as vaccine compatibility, the individual’s health status, and differences in storage requirements and administration schedules.
Another strategy is to use combination vaccines, which consist of two or more vaccines physically combined and administered simultaneously at the same anatomic site. Combination vaccines offer additional advantages such as reduced storage and handling requirements, increased vaccine efficacy, and convenience for patients [51,52]. Administration of combination vaccines may be more effective in reducing the overall number of injections required, which can be particularly important for patients with needle phobia or anxiety [48,53,54].
To prevent both COVID-19 and seasonal influenza with a single vaccine, the development of combination vaccines targeting these two respiratory diseases has become increasingly relevant. In summary, combination vaccines against COVID-19 and seasonal influenza employ various biotechnological platforms (Figure 2). Inactivated vaccines use killed viruses to stimulate immunity; vectored vaccines deliver genetic material via harmless viruses; virus-like particle vaccines mimic viral structures to provoke an immune response; recombinant nanoparticle vaccines present antigens using engineered nanoparticles; recombinant protein vaccines introduce recombinant viral proteins; and mRNA vaccines instruct cells to produce viral antigens. Each platform offers distinct approaches to generating protective immunity against both diseases.
However, combining multiple vaccines may interfere with the immunogenic properties of individual antigens and complicate vaccination schedules [55,56,57,58].
Therefore, extensive research is needed on the safety and efficacy of simultaneous and combination vaccination, as well as on the characteristics of the post-vaccination immune response in experimental and clinical studies.
4. Animal Studies of Co-Vaccination Against COVID-19 and Influenza
Co-vaccination against influenza and COVID-19 with vaccines developed on different technological platforms has been repeatedly researched in relevant animal models, such as K18-hACE2 mice and hamsters.
The effectiveness of the co-vaccination against influenza A (H1N1) and SARS-CoV-2 was investigated using the transgenic K18-hACE2 mouse model. This study assessed the efficacy of the PiCoVacc SARS-CoV-2 vaccine, the flu vaccine, and a co-vaccination in K18-hACE2 mice, using various infection and vaccination groups. The results indicated that mice infected with H1N1 experienced significant weight loss and viral loads, while those infected with SARS-CoV-2 did not survive the observation period. Mice with sequential infection with H1N1 followed by SARS-CoV-2 also showed accelerated mortality due to compromised immune responses. In terms of vaccination, mice that received only the PiCoVacc SARS-CoV-2 vaccine exhibited a strong neutralizing antibody response to SARS-CoV-2, while mice immunized with the H1N1 flu vaccine alone showed a positive response to the flu vaccine. The combined vaccination group demonstrated neutralizing antibodies for both viruses, with levels comparable to single vaccines. It can be concluded that simultaneous vaccination against H1N1 and SARS-CoV-2 provides effective protection against both infections [38].
An inactivated virus - based vaccine targeting both COVID-19 and influenza was studied by Singh et al. [59]. This combination vaccine encapsulated inactivated whole viruses of SARS-CoV-2 (Delta and Omicron variants) and Influenza A (H1N1 and H3N2 strains) within biodegradable PLGA polymer microparticles, formulated with the AddaVax™ adjuvant. Administered to mice via the intranasal route, this microparticulate system aims to provide sustained antigen release and target the mucosal immune system at the primary site of infection. The vaccine successfully induced a robust dual immune response, generating significant levels of virus-specific IgG and mucosal IgA antibodies, alongside strong activation of CD4+ and CD8+ T-cells. The immune response elicited was comparable to that achieved by traditional intramuscular vaccination, supporting the potential of this intranasal combination vaccine as a convenient and effective strategy for simultaneous protection against both respiratory pathogens [59].
Chaparian et al. developed a chimeric influenza virus which simultaneously displays influenza HA and the SARS-CoV-2 RBD domain. In prime-boost mode, this vaccine was successfully used in mice in live attenuated (prime vaccination) and inactivated (boost vaccination) form. Vaccination with this combination vaccine elicited neutralizing antibodies and provided protection from lethal challenge with both influenza and SARS-CoV-2 [60].
Wang et al. developed a virus-like particle (VLP) vaccine by conjugating the recombinant RBD of the SARS-CoV-2 spike protein onto an inactivated influenza A virus. The resulting vaccine, Flu-RBD, induced protective immunity against SARS-CoV-2 while retaining functionality as an influenza vaccine. In a hamster model, the vaccine conferred protection against live SARS-CoV-2 infection. It exhibited strong neutralization activity against both the SARS-CoV-2 Delta pseudovirus and the wild-type influenza A (H1N1) inactivated virus in mice [61].
Recently, another VLP-based combination vaccine candidate was created by Sanchez-Martinez et al. [62] VLPs produced in CHO cells contained full-length SARS-CoV-2 S-protein, influenza H1N1 hemagglutinin, and neuraminidase (S+H1+N1) incorporated into the VLPs’ envelope. The authors demonstrated that two doses of trivalent VLPs elicit specific antibodies and cellular immunity in a mouse model.
Shi et al. developed a combination liposome-enclosed vaccine candidate by mixing recombinant RBD-trimer and HA1-trimer, which conferred protection against SARS-CoV-2 and a lethal homologous H1N1 influenza challenge. Indeed, the RBD-trimer elicited significantly higher neutralizing antibody titers compared to the RBD-monomer, RBD-dimer, and spike ectodomain trimer. Additionally, the vaccine induced a balanced T helper cells (Th1/Th2) cellular immune response in mice [63].
Huang et al. designed a Flu-COVID combo vaccination using the AddaVax adjuvant, including the influenza virus hemagglutinin and SARS-CoV-2 spike proteins. This vaccine effectively protected mice from both influenza and SARS-CoV-2 challenges by preventing weight loss and disease progression, eliciting protective immune responses comparable to monovalent influenza or COVID-19 recombinant protein vaccines [64].
Flu-COVID pentavalent recombinant protein-based vaccine was developed by Krasilnikov and co-authors [65]. This vaccine contained SARS-CoV-2 RBD fused with the Fc fragment of the human IgG and HAs of four influenza viruses: A/H1N1- pdm09, A/H3H2, B/Yamagata, and B/Victoria. Betulin was used as an adjuvant to enhance the vaccine’s immunogenicity. In a mouse model, vaccination provided high titers of specific antibodies to all antigens administered in the vaccine, as well as SARS-CoV-2 and influenza virus neutralization.
Several combined mRNA-based candidate vaccines for COVID-19 and influenza were developed. A combined mRNA vaccine (AR-CoV/IAV) for COVID-19 and influenza was developed using a lipid nanoparticle-encapsulated mRNA platform (LNP-mRNA) [66]. This vaccine encodes IAV-HA and SARS-CoV-2-RBD and elicits robust hemagglutination inhibition (HAI) antibodies against IAV, as well as neutralizing antibodies against SARS-CoV-2. It also protects mice from co-infection with IAV and the SARS-CoV-2 Alpha and Delta variants. Moreover, this vaccine t induced Th1 cytokine-secreting CD4+ T cells and interferon gamma (IFN-γ+) or tumor necrosis factor-alpha (TNF-α+) CD8+ T cells, demonstrating enhanced antiviral activity without causing severe disease [66].
FLUCOV-10 is a LNP-mRNA vaccine that encodes full-length hemagglutinin proteins from four seasonal influenza viruses (A/Wisconsin/588/2019 (H1N1) pdm09, A/Darwin/6/2021 (H3N2), B/Austria/1359417/2021, and B/Phuket/3073/2013), two avian influenza viruses posing potential pandemic risks (A/Thailand/NBL1/2006 (H5N1) and A/Anhui/DEWH72-03/2013 (H7N9)), and spike proteins from four SARS-CoV-2 variants (ancestral SARS-CoV-2, BQ.1.1, BA.2.75.2, and XBB.1.5 omicron variants). It has been shown to elicit robust immune responses in mice, including the production of immunoglobulin G (IgG), neutralizing antibodies, and antigen-specific cellular responses against all vaccine-matched influenza and SARS-CoV-2 viruses, as well as complete protection in mouse models against both homologous and heterologous strains of influenza and SARS-CoV-2 [67].
Recently, a study in mice evaluated the co-administration of the seasonal quadrivalent influenza vaccine (QIV) and the Pfizer-BioNTech COVID-19 mRNA vaccine (BNT162b2). The research specifically tested three methods of simultaneous administration: injection into opposite limbs, the same limb, or mixing both vaccines in one syringe. The key finding was that co-administration, particularly when vaccines were given in the same limb or mixed, led to a significantly enhanced antibody response against influenza viruses. Co-administration resulted in a slight reduction in antibody levels against SARS-CoV-2 compared to giving the COVID-19 vaccine alone. Despite this modest reduction, all co-administered regimens provided complete protection, with vaccinated mice showing 100% survival, minimal weight loss, and low viral loads in the lungs after lethal challenge with either virus. This suggests that simultaneous vaccination induces robust protective immunity, though it may differentially modulate the strength of the immune response to each pathogen [68].
A 2025 study developed a novel mRNA-LNP combination vaccine for both influenza and COVID-19. To address the known challenge of low immunogenicity for influenza B strains in mRNA platforms, the researchers used an innovative antigen design. They engineered fusion proteins where hemagglutinin (HA) antigens are linked by a stabilizing bacteriophage T4 foldon domain, creating "dumbbell" or trimeric structures expressed from a single mRNA species. In mice, this combination vaccine elicited hemagglutination inhibition (HAI) antibody titers against seasonal influenza strains that were significantly higher than those induced by a commercial high-dose inactivated vaccine (Fluzone HD). Simultaneously, it generated superior neutralizing antibody responses against the SARS-CoV-2 XBB.1.5 variant compared to a commercial COVID-19 mRNA vaccine (Spikevax). The study demonstrates a promising antigen design strategy to enhance immunogenicity in multivalent mRNA vaccines [69].
Viral vectors to deliver genetic material encoding target antigens demonstrated good potential for creating new vaccines for respiratory infection prevention [70]. Immunization with vector vaccines elicits strong immune responses against both the vector and the embedded vaccine antigen. Moreover, viral vector vaccines can be easily administered non-invasively, via nasal sprays or nebulization, and are associated with fewer adverse reactions and improved acceptance of vaccines. The administration of intranasal vaccines augments protection against respiratory viruses through the stimulation of the immune system at the primary site of viral infections, thereby fostering a balanced and efficacious immune response. Importantly, immunization with vector vaccines also promotes the production of cytokines and chemokines as part of a defensive inflammatory reaction [71].
Implementation of adenoviral vector vaccines during the COVID-19 pandemic underscored their evident efficacy, favorable safety characteristics, and immunogenicity. Cao et al. described a strategy for developing a chimpanzee adenovirus 68 (AdC68)-based vaccine targeting both SARS-CoV-2 and IAV using a fusion immunogen [72]. It focused on an immunogen created by combining the SARS-CoV-2 receptor-binding domain (RBD) with the conserved stalk of H7N9 hemagglutinin (HA). Ferritin was used as a platform to improve the vaccine’s immunogenicity. The AdC68-CoV/Flu vaccine elicited antibody response against both viruses. The spike protein elicited strong neutralizing antibody responses against wild-type SARS-CoV-2 strains but lower responses against variants such as Beta (B.1.351, B.1.627) and Gamma (P.1). Extensive RBD-specific T cell responses of splenocytes were revealed. In addition, AdC68-CoV/Flu vaccine provided effective protection against lethal SARS-CoV-2 challenge in hACE2-C57BL/6 mice [72].
More recently, another new AdC68-HATRBD vaccine was developed using the same AdC68 vector [73]. This vaccine encoded two RBD dimers from various SARS-CoV-2 variants, namely Beta-Alpha chimeric dimer and Omicron-Delta chimeric dimer, as well as numerous T cell epitopes of SARS-CoV-2 and full-length HA of A/California/07/2009 (pH1N1). When administered intranasally, the AdC68-HATRBD vaccine provided comprehensive immune responses, including IgG, mucosal IgA, and memory T cell responses, which protected the mice from BA.5.2 and pandemic H1N1 infections.
The influenza vector platform is based on integrating foreign proteins into attenuated or replication-deficient influenza viruses for developing vaccines targeting various human respiratory pathogens [74,75,76].
Delta-19 is a nasal spray vaccine designed to protect against both COVID-19 and influenza. It is built on Delta NS1 vaccine vector technology, which expresses key immunogenic proteins of both viruses. According to the developer's (Vivaldi Biosciences) information, the vaccine is currently undergoing challenge-protection studies in animal models and is being prepared for an Investigational New Drug (IND) application and clinical trials [77].
A study by Sergeeva et al. investigated the effectiveness of an intranasal vaccine using a modified influenza vector that encodes the nucleoprotein (N protein) of SARS-CoV-2 and has a truncated NS1 gene, designed to stimulate a robust local immune response, particularly targeting essential CD8+ T-cells. The study revealed that intranasal immunization with the influenza vector significantly reduced weight loss and viral load in the lungs of naïve mice after exposure to the SARS-CoV-2 beta variant, indicating the vaccine's effectiveness in providing protection in previously unexposed individuals. Additionally, in seropositive Th2-prone mice that had been primed with alum-adjuvanted inactivated SARS-CoV-2, a single intranasal boost with the vaccine was able to prevent disease enhancement, such as early weight loss and eosinophilia in the lungs during infection. The vaccine successfully modulated the immune response to mitigate these symptoms. Overall, the findings highlight that intranasal immunization with influenza vector-based SARS-CoV-2 vaccine holds significant potential for preventing COVID-19 and associated immunopathology, though the direct anti-influenza protective effect of the vaccine has not been assessed in this work [78].
During development of the Pneucolin dNS1-RBD vector vaccine, the nonstructural-1 (NS1) gene of influenza was replaced with the SARS-CoV-2 RBD region. Studies in hamsters have shown that this nasal vaccine stimulated both systemic and local immune responses and preserved body weight after challenge. Moreover, the vaccine provided cross-protection against H1N1 and H5N1 influenza, as well as protection against various variants of SARS-CoV-2. Attenuating pro-inflammatory cytokine levels post SARS-CoV-2 challenge was registered, thereby reducing excess immune-induced lung tissue injury [79,80].
Loes and co-authors developed an influenza virus vector where the SARS-CoV-2 RBD region was inserted in place of the neuraminidase coding sequence. The authors demonstrated that this candidate vector vaccine elicits high titers of serum neutralizing antibodies against both SARS-CoV-2 and influenza viruses [81].
Licensed seasonal trivalent live attenuated influenza vaccine was used as the basis for the development of a modified 3×LAIV/CoV-2 vaccine, where H1N1 and H3N2 LAIV strains encoded an immunogenic cassette enriched with conserved T-cell epitopes of SARS-CoV-2, whereas a B/Victoria lineage LAIV strain was unmodified. This modified trivalent live attenuated influenza vaccine encoding the T-cell epitopes of SARS-CoV-2 revealed effective protection against challenges from either influenza strain as well as against SARS-CoV-2. It can be considered a promising tool for combined protection against seasonal influenza and COVID-19 [82].
Thus, the results of animal studies demonstrate that simultaneous vaccination and combination vaccines developed using different platforms are effective and immunogenic against SARS-CoV-2 and influenza. These vaccination strategies provide effective protection against both infections.
Animal studies of co-vaccination against SARS-CoV-2 and influenza virus are summarized in Table 1.
5. Clinical Studies of Co-Administration of Influenza and COVID-19 Vaccines
COVID-19 now is becoming an established seasonal epidemic disease, and its severe public health impact has diminished from the pandemic's initial phase. Nevertheless, it continues to present a substantial health burden as the respiratory infectious disease accountable for a number of hospitalizations and intensive care unit admissions, especially in patients from high-risk groups. The possibility of future outbreaks highlights the importance of up-to-date vaccination strategies, which are developing all over the world. In October 2021, the World Health Organization (WHO) approved co-administration of vaccines against COVID-19 and seasonal influenza [18]. Subsequently, the Russian Ministry of Health also authorized simultaneous vaccination and updated the instructions for the use of the Sputnik V vaccine accordingly [19].
During the COVID-19 pandemic, clinical trials have been conducted in several countries, and the results show that simultaneous administration of the vaccines is generally safe and produces comparable reactogenicity and immune responses to separate administration [83,84,85,86,87,88,89,90,91,92,93,94]. Additionally, simultaneous vaccination may increase vaccine uptake without compromising the effectiveness of either vaccine [50]. Figure 3 summarizes the geography of these clinical trials and lists the countries that recommend simultaneous vaccination against COVID-19 and seasonal influenza.
Clinical research on the safety and effectiveness of simultaneous vaccination against COVID-19 and influenza is a task of primary importance for public health.
Toback et al. investigated the safety and immunogenicity profiles of NVX-CoV2373 (Novavax; US) vaccinations administered simultaneously with seasonal influenza vaccines. The incidence and severity of local and systemic reactogenicity events after co-administration were generally comparable to those when each vaccine was administered separately. Furthermore, post-vaccination geometric mean titers and seroconversion rates remained high for each strain, regardless of whether the influenza vaccine was administered with placebo or NVX-CoV2373, despite a generally reduced response to the influenza B strains among all influenza vaccine recipients. However, co-administration of NVX-CoV2373 and an influenza vaccine resulted in a modest decrease in anti-spike protein IgG. The levels of anti-spike protein IgG in those who received the two vaccines remained more than threefold higher than those seen in convalescent serum, implying that these levels may be protective [87].
Hause et al. showed that among individuals who received both the seasonal influenza vaccine and either the Pfizer-BioNTech or Moderna COVID-19 mRNA booster simultaneously, systemic reactions were 8%–11% more common compared to those who received the COVID-19 booster alone [83].
Izikson et al. conducted a study on the safety and immunogenicity of co-administering a high-dose quadrivalent influenza vaccine (QIV-HD) and a mRNA-1273 vaccine booster dose in older adults. The study found no safety concerns or immune interference in older adults who received a third dose of the mRNA-1273 vaccine with QIV-HD up to 21 days after vaccination. Similar hemagglutination inhibition and SARS-CoV-2 binding antibody responses were observed between the co-administration and QIV-HD groups and between the co-administration and mRNA-1273 groups [84].
A study comparing the effectiveness of co-administering the BNT162b2 BA.4/5 bivalent mRNA COVID-19 vaccine and seasonal influenza vaccines (SIV) in a community setting (which included 3,442,996 commercially insured US adults aged 18 years or older) found that outcomes of co-administration of both vaccines were similar when compared to those of each vaccine against COVID-19 and SIV alone. It suggests that co-administration may improve the uptake of both vaccines [85].
Moro et al. compared reports of systemic reactions and injection site reactions in COVID-19 patients who received a booster dose of a mRNA COVID-19 booster vaccine with a quadrivalent inactivated influenza vaccine (QIV) and those who received a booster dose alone. Systemic reactions were slightly more frequent in reports with the QIV vaccine, while injection site reactions and COVID-19 infection were slightly more frequent in reports with only the mRNA COVID-19 vaccine booster dose [86].
The combining influenza and COVID-19 vaccination (ComFluCOV) study was undertaken to determine the side effects, including fever and tiredness, experienced by participants when their second COVID-19 vaccine dose is administered simultaneously with the flu vaccine. In the multicenter, randomized, controlled, phase 4, trial 679 adult participants were enrolled. They receive a single dose of ChAdOx1 or BNT162b2 and concomitant administration of either an age-appropriate seasonal, inactivated vaccines or placebo alongside their second dose of COVID-19 vaccine. 3 weeks later the group who received placebo received the influenza vaccine, and vice versa. It was established that most systemic reactions to vaccination were mild or moderate, rates of adverse local and systemic reactions were similar between the groups, and immune responses were not adversely affected. Concomitant vaccination with ChAdOx1 or BNT162b2 plus an age-appropriate influenza vaccine raises no safety concerns and preserves antibody responses to both vaccines [88].
In the study involving 1231 participants, individuals who had received two doses of the BNT162b2 mRNA vaccine followed by a third dose (either BNT162b2 or mRNA-1273) were assessed for co-administration with the influenza vaccine. Co-administration of the influenza vaccination was an option chosen by part of the participants. Although co-administration led to a modest reduction in anti-SARS-CoV-2 spike IgG levels—particularly for mRNA-1273 and BNT162b2—it did not result in increased health-related absenteeism among healthcare workers and thus does not jeopardize public healthcare capacity [92].
A prospective cohort study involving healthcare workers at a large tertiary medical center in Israel assessed the reactogenicity and immunogenicity of co-administration of the Omicron BA.4/BA.5-adapted bivalent COVID-19 vaccine with SIV. Results showed that those who received SIV alone experienced the least reactogenicity, while COVID-19 vaccination alone elicited similar reactogenicity to the co-administration of the vaccine with SIV. The study also revealed a 16% decrease in anti-spike IgG titers, which did not significantly impact vaccine effectiveness or protection against symptomatic disease, based on previous studies [93].
A similar study found that co-administering the BNT162b2 vaccine alongside the seasonal inactivated influenza vaccine (SIIV) in adults aged 18–64 was safe and well-tolerated and demonstrated robust immune responses that were not inferior to separate administration of BNT162b2 and SIIV [94].
Moderna successfully conducted Phase 1 and 2 trials to evaluate the safety, reactogenicity, and immunogenicity of mRNA-1073 (SARS-CoV-2 and Influenza Vaccine) when compared to co-administered mRNA-1010 (Influenza) and mRNA-1273 (SARS-CoV-2) vaccines and to mRNA- 1010 and mRNA-1273 vaccines in healthy adults 18-75 years [95].
Lee et al. performed a randomized trial that enrolled 56 adults who received the Afluria QIV and the Moderna monovalent SARS-CoV-2 XBB.1.5 mRNA vaccine, either in separate arms or both in the same arm at the same anatomical site. Influenza vaccination's immunogenicity is comparable regardless of whether it's administered in the same or opposite arms as the SARS-CoV-2 vaccine. However, it may be advisable to administer the SARS-CoV-2 vaccine at a distinct site from influenza vaccines [96].
The recent systematic review consolidates available evidence on the prevalence, safety, immunogenicity, and effectiveness of co-administering BNT162b2 and licensed SIVs [97]. Twenty studies (15 observational and 5 clinical trials) conducted in seven countries in Europe and North America were included. Eight observational studies reported prevalence, twelve reported safety/reactogenicity, six reported immunogenicity, and three evaluated effectiveness. Reported co-administration of BNT162b2 vaccines with SIVs increased over time. The percentage of individuals who received BNT162b2 and also reported co-administered SIVs rose significantly, from 2.7% in 2021 to 34.1% in 2023. Effectiveness was not observed to change when BNT162B2 was co-administered with SIVs. The incidence of systemic and local adverse events was comparable between individuals receiving the vaccines separately and those receiving them co-administered.
Table 2 represents key parameters of clinical studies of influenza and COVID-19 vaccines co-administration.
Thus, clinical studies of co-administration of vaccines against COVID-19 and influenza proved safety in evaluated scenarios [97,98]. Most side effects were mild to moderate and temporary. While some studies noted a slight increase in reactogenicity, serious adverse events or safety issues were not inherent. Importantly, nearly all studies confirmed that simultaneous vaccination did not significantly alter the immune response for either vaccine. The advantages conferred by vaccine co-administration demonstrably outweigh any associated risks. This emphasizes the benefit of combined administration, potentially improving vaccine acceptance by simplifying immunization procedures and decreasing healthcare visits. However, despite these findings, the optimal approach to co-vaccination against COVID-19 and influenza remains the subject of continuous discussion and research [99].
6. Clinical Studies of Combined Influenza and SARS-CoV-2 Vaccines
Combination influenza and SARS-CoV-2 vaccines are aimed to provide protection against both infections with a single-dose administration, as a more favorable alternative to simultaneous vaccination with two vaccines. A number of combined vaccines on various biotechnological platforms have proven their immunogenicity and beneficial safety profile in preclinical studies. Several combined vaccines reached clinical trials. A summary of clinical studies on combination vaccines targeting SARS-CoV-2 and influenza is provided in Table 3.
A part of these clinical trials involves the mRNA LNP vaccine platform, which previously became the basis for the globally approved monovalent SARS-CoV-2 mRNA vaccines. mRNA LNP vaccines have a modular technological engineering that allows to rapidly amplify their manufacturing, as well as easily reconfigure the vaccine’s design following the drift in actual viral antigens.
Pfizer and BioNTech launched a Phase 3 trial to evaluate the safety, tolerability, and immunogenicity of a combined modified RNA COVID-19 and influenza vaccine (NCT06178991) [100].
Moderna performed two trials of the safety, reactogenicity, and immunogenicity of SARS-CoV-2 and influenza vaccines, mRNA-1073 (NCT05375838) Phase 1,2 [101] and mRNA-1083 (NCT06097273) Phase 3 [102].
GlaxoSmithKline recently initiated a Phase 1,2 clinical trial (NCT06680375) to assess the reactogenicity, safety, and immune response of the mRNA Flu/COVID-19 vaccine [103].
Novavax performed a clinical study evaluating the safety and immunogenicity of a SARS-CoV-2 rS nanoparticle and quadrivalent hemagglutinin nanoparticle influenza combination vaccine with Matrix-M adjuvant [104]. This vaccine is designed as adjuvant nanoparticles bearing recombinant antigen proteins of SARS-CoV-2 and the influenza virus.
In addition to the mRNA LNP and nanoparticle-based combination vaccines listed above, an attenuated influenza virus with a deleted NS segment (delta NS technology) has proven to be a promising biotechnological platform. Delta NS technology was applied in the development of the dNS1-RBD Pneucolin vaccine with an intranasal route of administration. In general, intranasal vaccines provide additional immune protection via mucosal resident memory B- and T-cells, as well as secretory IgA stimulation. dNS1-RBD Pneucolin’s efficacy and safety have been demonstrated in several clinical trials (ChiCTR2000037782, ChiCTR2000039715, ChiCTR21000483160) [105,106]. dNS1-RBD was the first intranasal vaccine against COVID-19, which obtained emergency use authorization in China in December 2022.
Delta NS technology was also used in the design of another vaccine, Corfluvec. Corfluvec contains two influenza virus-based vectors (H3N2 and H1N1pdm09) that carry a modified NS gene encoding the N protein of SARS-CoV-2. A phase 1,2 clinical trial assessing the safety and immunogenicity of this intranasal vaccine was carried out (NCT05696067) [107].
In clinical trial results, favorable safety profiles of the vaccines listed above were reported. Nevertheless, further extensive large-scale research of their efficacy in human populations is clearly needed for combined vaccines’ implementation into the routine practice of influenza and COVID-19 medical management.
7. Conclusions
The co-circulation of COVID-19 and influenza highlights the potential for simultaneous outbreaks, particularly during peak respiratory illness seasons, which could significantly impact public health. This dynamic circulation raises concerns about the risk of infection with either virus. Co-infection with influenza viruses and SARS-CoV-2 poses a serious threat to global health and underscores the need for a multifaceted vaccination strategy. Co-infection with these viruses exacerbates respiratory disease, prolongs pneumonia, and increases mortality.
Healthcare professionals should remain vigilant, monitoring both infections and promoting simultaneous or combination vaccination against COVID-19 and influenza. The main advantage of simultaneous vaccination is improved efficiency, allowing multiple vaccines to be administered at once. This approach reduces the number of visits, minimizing scheduling challenges and missed doses. In addition, it increases the vaccination rate and adherence to recommended vaccine schedules, contributing to higher community immunity and overall safety. Moreover, it can reduce healthcare costs by minimizing the resources required for multiple appointments, thereby improving the overall efficiency of vaccination programs.
Future studies should focus on elucidating the mechanisms underlying the exacerbation of respiratory disease caused by co-infection, as well as the optimal strategies for co-administering influenza and COVID-19 vaccines. Simultaneous and combination vaccination strategies should be evaluated across diverse populations and settings to inform public health policy and guide future pandemic preparedness. Together, these insights support the development and integration of dual-targeted vaccines as a practical and effective tool for long-term control of seasonal and potentially pandemic respiratory viruses.
Author Contributions
Conceptualization, E.V., M.K., A.G., and M.S.; formal analysis, E.V., M.K. A.G., and M.S.; writing—original draft preparation, E.V., A.G., and M.K.; writing—review and editing, A.G., E.V., and M.S., funding acquisition, M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Ministry of Health of the Russian Federation government contract, grant 125020401358-1.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ACE2 | Angiotensin-Converting Enzyme 2 |
| AdC68 | Chimpanzee adenovirus 68 |
| BAL | Bronchoalveolar lavage |
| HA | Hemagglutinin |
| HAI | Hemagglutination Inhibition Assay |
| HI | Hemagglutination Inhibition |
| IAV | Influenza A virus |
| Ig | Immunoglobulin |
| IM | Intramuscular |
| IN | Intranasal |
| IP | Intraperitoneal |
| IND | Investigational New Drug |
| LNP | Lipid Nanoparticle |
| PLGA | Co-polymer of lactic and glycolic acids |
| qIRV | Quadrivalent influenza modRNA vaccine |
| QIV | Quadrivalent Inactivated Influenza Vaccine |
| RBD | Receptor-Binding Domain |
| SARS-CoV-2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
| SIV | Seasonal Influenza Vaccines |
| SIIV | Seasonal Inactivated Influenza Vaccine |
| TIV | Trivalent Inactivated Influenza vaccine |
| QIV-HD | High-dose quadrivalent influenza vaccine |
| V-safe | Vaccine safety |
| VAERS | Vaccine Adverse Event Reporting System |
| VLP | Virus-Like Particle |
References
- Taubenberger, J.K.; Morens, D.M. Influenza: The Once and Future Pandemic. Public health reports 2010, 125, 15–26. [Google Scholar] [CrossRef]
- Influenza (Seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal).
- Peacock, T.P.; Moncla, L.; Dudas, G.; VanInsberghe, D.; Sukhova, K.; Lloyd-Smith, J.O.; Worobey, M.; Lowen, A.C.; Nelson, M.I. The Global H5N1 Influenza Panzootic in Mammals. Nature 2025, 637, 304–313. [Google Scholar] [CrossRef]
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/2023.
- The Current Epidemic Situation in Russia and the World. Available online: https://www.rospotrebnadzor.ru/region/korono_virus/epid.php.
- World Health Organization Statement on the Fifteenth Meeting of the IHR (2005) Emergency Committee on the COVID-19 Pandemic. Available online: https://www.who.int/news/item/05-05-2023-statement-on-the-fifteenth-meeting-of-the-international-health-regulations-(2005)-emergency-committee-regarding-the-coronavirus-disease-(covid-19)-pandemic.
- Inaida, S.; Paul, R.E.; Matsuno, S. Viral Transmissibility of SARS-CoV-2 Accelerates in the Winter, Similarly to Influenza Epidemics. American Journal of Infection Control 2022, 50, 1070–1076. [Google Scholar] [CrossRef]
- Coleman, K.K.; Tay, D.J.W.; Tan, K.S.; Ong, S.W.X.; Than, T.S.; Koh, M.H.; Chin, Y.Q.; Nasir, H.; Mak, T.M.; Chu, J.J.H. Viral Load of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in Respiratory Aerosols Emitted by Patients with Coronavirus Disease 2019 (COVID-19) While Breathing, Talking, and Singing. Clinical Infectious Diseases 2022, 74, 1722–1728. [Google Scholar] [CrossRef] [PubMed]
- Chow, V.T.; Tay, D.J.W.; Chen, M.I.; Tang, J.W.; Milton, D.K.; Tham, K.W. Influenza A and B Viruses in Fine Aerosols of Exhaled Breath Samples from Patients in Tropical Singapore. Viruses 2023, 15, 2033. [Google Scholar] [CrossRef]
- Riou, J.; Althaus, C.L. Pattern of Early Human-to-Human Transmission of Wuhan 2019 Novel Coronavirus (2019-nCoV), December 2019 to January 2020. Eurosurveillance 2020, 25, 2000058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, L.; Deng, X.; Liang, R.; Su, M.; He, C.; Hu, L.; Su, Y.; Ren, J.; Yu, F. Recent Advances in the Detection of Respiratory Virus Infection in Humans. Journal of Medical Virology 2020, 92, 408–417. [Google Scholar] [CrossRef]
- Khorramdelazad, H.; Kazemi, M.H.; Najafi, A.; Keykhaee, M.; Emameh, R.Z.; Falak, R. Immunopathological Similarities between COVID-19 and Influenza: Investigating the Consequences of Co-Infection. Microbial Pathogenesis 2021, 152, 104554. [Google Scholar] [CrossRef]
- Golpour, M.; Jalali, H.; Alizadeh-Navaei, R.; Talarposhti, M.R.; Mousavi, T.; Ghara, A.A.N. Co-Infection of SARS-CoV-2 and Influenza A/B among Patients with COVID-19: A Systematic Review and Meta-Analysis. BMC Infectious Diseases 2025, 25, 145. [Google Scholar] [CrossRef]
- Alosaimi, B.; Naeem, A.; Hamed, M.E.; Alkadi, H.S.; Alanazi, T.; Al Rehily, S.S.; Almutairi, A.Z.; Zafar, A. Influenza Co-Infection Associated with Severity and Mortality in COVID-19 Patients. Virology Journal 2021, 18, 127. [Google Scholar] [CrossRef]
- Yue, H.; Zhang, M.; Xing, L.; Wang, K.; Rao, X.; Liu, H.; Tian, J.; Zhou, P.; Deng, Y.; Shang, J. The Epidemiology and Clinical Characteristics of Co-infection of SARS-CoV-2 and Influenza Viruses in Patients during COVID-19 Outbreak. Journal of Medical Virology 2020, 92, 2870–2873. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, S.; Kim, H.; Gil, D.; Han, H.-J.; Thimmulappa, R.K.; Choi, J.-H.; Kim, J.-H. Reciprocal Enhancement of SARS-CoV-2 and Influenza Virus Replication in Human Pluripotent Stem Cell-Derived Lung Organoids. Emerging Microbes & Infections 2023, 12, 2211685. [Google Scholar] [CrossRef]
- Ma, S.; Lai, X.; Chen, Z.; Tu, S.; Qin, K. Clinical Characteristics of Critically Ill Patients Co-Infected with SARS-CoV-2 and the Influenza Virus in Wuhan, China. International Journal of Infectious Diseases 2020, 96, 683–687. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Co-administration of Seasonal Inactivated Influenza and COVID-19 Vaccines. Available online: https://www.who.int/publications/i/item/WHO-2019-nCoV-vaccines-SAGE_recommendation-co-administration.
- Russian Ministry of Health The Russian Ministry of Health has authorized simultaneous vaccination against COVID-19 and influenza. Available online: https://minzdrav.gov.ru/news/2021/10/22/17665-minzdrav-rossii-razreshil-odnovremennuyu-vaktsinatsiyu-ot-kovida-i-grippa.
- Xie, Z.; Hamadi, H.Y.; Mainous, A.G.; Hong, Y.-R. Association of Dual COVID-19 and Seasonal Influenza Vaccination with COVID-19 Infection and Disease Severity. Vaccine 2023, 41, 875–878. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Influenza and SARS-CoV-2 Virus Detections Reported to FluNet from Countries, Areas and Territories. Available online: https://app.powerbi.com/view?r=eyJrIjoiNzc4YTIxZjQtM2E1My00YjYxLWIxMDItNzEzMjkyY2E1MzU1IiwidCI6ImY2MTBjMGI3LWJkMjQtNGIzOS04MTBiLTNkYzI4MGFmYjU5MCIsImMiOjh9.
- Karpova, L.S.; Pelikh, M.Y.; Stolyarov, K.A.; Volik, K.M.; Stolyarova, T.P.; Danilenko, D.M. Analysis of Influenza Epidemics during the COVID-19 Pandemic Using an Improved Surveillance System (from 2021 to 2024). Journal of Microbiology, Epidemiology and Immunobiology 2024, 101, 679–691. [Google Scholar] [CrossRef]
- FluCov Dashboard Understanding the Impact of SARS-CoV-2 on Influenza. Available online: https://www.nivel.nl/en/dossier-epidemiology-respiratory-viruses/flucov-dashboard.
- World Health Organization Global Influenza Programme. Available online: https://www.who.int/tools/flunet.
- Takashita, E.; Watanabe, S.; Hasegawa, H.; Kawaoka, Y. Are Twindemics Occurring? Influenza and Other Respiratory Viruses 2023, 17, e13090. [Google Scholar] [CrossRef]
- Opatowski, L.; Baguelin, M.; Eggo, R.M. Influenza Interaction with Cocirculating Pathogens and Its Impact on Surveillance, Pathogenesis, and Epidemic Profile: A Key Role for Mathematical Modelling. PLoS Pathogens 2018, 14, e1006770. [Google Scholar] [CrossRef]
- Shinjoh, M.; Omoe, K.; Saito, N.; Matsuo, N.; Nerome, K. In Vitro Growth Profiles of Respiratory Syncytial Virus in the Presence of Influenza Virus. Acta Virologica 2000, 44, 91–97. [Google Scholar]
- Piret, J.; Boivin, G. Viral Interference between Respiratory Viruses. Emerging Infectious Diseases 2022, 28, 273. [Google Scholar] [CrossRef]
- Pinky, L.; Dobrovolny, H.M. Coinfections of the Respiratory Tract: Viral Competition for Resources. PloS ONE 2016, 11, e0155589. [Google Scholar] [CrossRef]
- Pizzorno, A.; Padey, B.; Dulière, V.; Mouton, W.; Oliva, J.; Laurent, E.; Milesi, C.; Lina, B.; Traversier, A.; Julien, T. Interactions between Severe Acute Respiratory Syndrome Coronavirus 2 Replication and Major Respiratory Viruses in Human Nasal Epithelium. The Journal of Infectious Diseases 2022, 226, 2095–2104. [Google Scholar] [CrossRef] [PubMed]
- Pinky, L.; Dobrovolny, H.M. SARS-CoV-2 Coinfections: Could Influenza and the Common Cold Be Beneficial? Journal of Medical Virology 2020, 92, 2623–2630. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Zhao, Y.; Dong, J.; Liang, S.; Guo, M.; Liu, X.; Wang, X.; Huang, Z.; Sun, X.; Zhang, Z.; et al. Coinfection with Influenza A Virus Enhances SARS-CoV-2 Infectivity. Cell Research 2021, 31, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H.; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef]
- Ibañez, L.; Martinez, V.; Iglesias, A.; Bellomo, C.; Alonso, D.; Coelho, R.; Martinez Peralta, L.; Periolo, N. Decreased Expression of Surfactant Protein-C and CD74 in Alveolar Epithelial Cells during Influenza Virus A(H1N1)Pdm09 and H3N2 Infection. Microbial Pathogenesis 2023, 176, 106017–106017. [Google Scholar] [CrossRef]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]
- Kinoshita, T.; Watanabe, K.; Sakurai, Y.; Nishi, K.; Yoshikawa, R.; Yasuda, J. Co-Infection of SARS-CoV-2 and Influenza Virus Causes More Severe and Prolonged Pneumonia in Hamsters. Scientific Reports 2021, 11, 21259–21259. [Google Scholar] [CrossRef]
- Zhang, A.J.; Lee, A.C.-Y.; Chan, J.F.-W.; Liu, F.; Li, C.; Chen, Y.; Chu, H.; Lau, S.-Y.; Wang, P.; Chan, C.C.-S.; et al. Coinfection by Severe Acute Respiratory Syndrome Coronavirus 2 and Influenza A(H1N1)Pdm09 Virus Enhances the Severity of Pneumonia in Golden Syrian Hamsters. Clinical Infectious Diseases 2021, 72, e978–e992. [Google Scholar] [CrossRef]
- Bao, L.; Deng, W.; Qi, F.; Lv, Q.; Song, Z.; Liu, J.; Gao, H.; Wei, Q.; Yu, P.; Xu, Y.; et al. Sequential Infection with H1N1 and SARS-CoV-2 Aggravated COVID-19 Pathogenesis in a Mammalian Model, and Co-Vaccination as an Effective Method of Prevention of COVID-19 and Influenza. Signal Transduction and Targeted Therapy 2021, 6, 200–200. [Google Scholar] [CrossRef]
- Wu, X.; Cai, Y.; Huang, X.; Yu, X.; Zhao, L.; Wang, F.; Li, Q.; Gu, S.; Xu, T.; Li, Y.; et al. Co-Infection with SARS-CoV-2 and Influenza A Virus in Patient with Pneumonia, China. Emerging Infectious Diseases 2020, 26, 1324–1326. [Google Scholar] [CrossRef]
- Dadashi, M.; Khaleghnejad, S.; Abedi Elkhichi, P.; Goudarzi, M.; Goudarzi, H.; Taghavi, A.; Vaezjalali, M.; Hajikhani, B. COVID-19 and Influenza Co-Infection: A Systematic Review and Meta-Analysis. Frontiers in Medicine 2021, 8, 681469. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Wang, Z.; Ye, L.; He, X.; Wei, X.; Ma, Y.; Li, H.; Chen, L.; Wang, X.; Zhou, Q. Co-Infection of SARS-COV-2 and Influenza A Virus: A Case Series and Fast Review. Current Medical Science 2021, 41, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.S.; Shu, Y. Public Health Control Measures for the Co-Circulation of Influenza and SARS-CoV-2 During Influenza Seasons. China CDC Weekly 2022, 4, 22–26. [Google Scholar] [CrossRef]
- Ma, S.; Lai, X.; Chen, Z.; Tu, S.; Qin, K. Clinical Characteristics of Critically Ill Patients Co-Infected with SARS-CoV-2 and the Influenza Virus in Wuhan, China. International Journal of Infectious Diseases 2020, 96, 683–687. [Google Scholar] [CrossRef]
- Iacobucci, G. Covid-19: Risk of Death More than Doubled in People Who Also Had Flu, English Data Show. BMJ 2020, m3720–m3720. [Google Scholar] [CrossRef] [PubMed]
- Stowe, J.; Tessier, E.; Zhao, H.; Guy, R.; Muller-Pebody, B.; Zambon, M.; Andrews, N.; Ramsay, M.; Bernal, J.L. Interactions between SARS-CoV-2 and Influenza, and the Impact of Coinfection on Disease Severity: A Test-Negative Design. International Journal of Epidemiology 2021, 50, 1124–1133. [Google Scholar] [CrossRef]
- Sørup, S.; Benn, C.S.; Poulsen, A.; Krause, T.G.; Aaby, P.; Ravn, H. Simultaneous Vaccination with MMR and DTaP-IPV-Hib and Rate of Hospital Admissions with Any Infections: A Nationwide Register Based Cohort Study. Vaccine 2016, 34, 6172–6180. [Google Scholar] [CrossRef]
- Sansone, V.; Miraglia del Giudice, G.; Della Polla, G.; Angelillo, I.F. Simultaneous Vaccination against Seasonal Influenza and COVID-19 among the Target Population in Italy. Frontiers in Public Health 2024, 12. [Google Scholar] [CrossRef]
- Dudley, M.Z.; Salmon, D.A.; Halsey, N.A.; Orenstein, W.A.; Limaye, R.J.; O’Leary, S.T.; Omer, S.B. Do Combination Vaccines or Simultaneous Vaccination Increase the Risk of Adverse Events? In The Clinician’s Vaccine Safety Resource Guide; Springer International Publishing: Cham, 2018; pp. 157–165. [Google Scholar]
- Walter, E.B.; Schlaudecker, E.P.; Talaat, K.R.; Rountree, W.; Broder, K.R.; Duffy, J.; Grohskopf, L.A.; Poniewierski, M.S.; Spreng, R.L.; Staat, M.A.; et al. Safety of Simultaneous vs Sequential mRNA COVID-19 and Inactivated Influenza Vaccines: A Randomized Clinical Trial. JAMA Network Open 2024, 7, e2443166–e2443166. [Google Scholar] [CrossRef]
- Bonanni, P.; Steffen, R.; Schelling, J.; Balaisyte-Jazone, L.; Posiuniene, I.; Zatoński, M.; Van Damme, P. Vaccine Co-Administration in Adults: An Effective Way to Improve Vaccination Coverage. Human Vaccines & Immunotherapeutics 2023, 19. [Google Scholar] [CrossRef]
- Decker, M.D. Combination Vaccines. In Clinics in Office Practice; Primary Care, 2001; Volume 28, pp. 739–761. [Google Scholar] [CrossRef]
- Skibinski, D.; Baudner, B.; Singh, M.; O′Hagan, D. Combination Vaccines. Journal of Global Infectious Diseases 2011, 3, 63–63. [Google Scholar] [CrossRef]
- Maman, K.; Zöllner, Y.; Greco, D.; Duru, G.; Sendyona, S.; Remy, V. The Value of Childhood Combination Vaccines: From Beliefs to Evidence. Human Vaccines & Immunotherapeutics 2015, 11, 2132–2141. [Google Scholar] [CrossRef]
- Rubin, R. Combined Vaccines Against COVID-19, Flu, and Other Respiratory Illnesses Could Soon Be Available. JAMA 2024, 331, 1880–1880. [Google Scholar] [CrossRef]
- Guess, H.A. Combination Vaccines: Issues in Evaluation of Effectiveness and Safety. Epidemiologic Reviews 1999, 21, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Elliman, D.; Bedford, H. Safety and Efficacy of Combination Vaccines: Combinations Reduce Distress and Are Efficacious and Safe. Bmj 2003, 326, 995–996. [Google Scholar] [CrossRef] [PubMed]
- Halsey, N.A. Safety of Combination Vaccines: Perception versus Reality. The Pediatric Infectious Disease Journal 2001, 20, S40–S44. [Google Scholar] [CrossRef]
- Tafreshi, S.H. Efficacy, Safety, and Formulation Issues of the Combined Vaccines. Expert Review of Vaccines 2020, 19, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Gulani, M.; Vijayanand, S.; Arte, T.; Adediran, E.; Pasupuleti, D.; Patel, P.; Ferguson, A.; Uddin, M.; Zughaier, S.M. An Intranasal Quadruple Variant Vaccine Approach Using SARS-CoV-2 and Influenza A: Delta, Omicron, H1N1and H3N2. International Journal of Pharmaceutics 2025, 126043. [Google Scholar] [CrossRef]
- Chaparian, R.R.; Harding, A.T.; Hamele, C.E.; Riebe, K.; Karlsson, A.; Sempowski, G.D.; Heaton, N.S.; Heaton, B.E. A Virion-Based Combination Vaccine Protects against Influenza and SARS-CoV-2 Disease in Mice. Journal of Virology 2022, 96, e00689-22. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Z.; Shi, W.; Zhu, D.; Hu, S.; Dinh, P.-U.C.; Cheng, K. A SARS-CoV-2 and Influenza Double Hit Vaccine Based on RBD-Conjugated Inactivated Influenza A Virus. Science Advances 2023, 9. [Google Scholar] [CrossRef]
- Sanchez-Martinez, Z.V.; Alpuche-Lazcano, S.P.; Stuible, M.; Akache, B.; Renner, T.M.; Deschatelets, L.; Dudani, R.; Harrison, B.A.; McCluskie, M.J.; Hrapovic, S. SARS-CoV-2 Spike-Based Virus-like Particles Incorporate Influenza H1/N1 Antigens and Induce Dual Immunity in Mice. Vaccine 2024, 42, 126463. [Google Scholar] [CrossRef]
- Shi, R.; Zeng, J.; Xu, L.; Wang, F.; Duan, X.; Wang, Y.; Wu, Z.; Yu, D.; Huang, Q.; Yao, Y.-G.; et al. A Combination Vaccine against SARS-CoV-2 and H1N1 Influenza Based on Receptor Binding Domain Trimerized by Six-Helix Bundle Fusion Core. eBioMedicine 2022, 85, 104297–104297. [Google Scholar] [CrossRef]
- 64 Huang, Y.; Shi, H.; Forgacs, D.; RossT, M. Flu-COVID Combo Recombinant Protein Vaccines Elicited Protective Immune Responses against Both Influenza and SARS-CoV-2 Viruses Infection. Vaccine 2024, 42, 1184–1192. [Google Scholar] [CrossRef]
- Krasilnikov, I.; Isaev, A.; Djonovic, M.; Ivanov, A.; Romanovskaya-Romanko, E.; Stukova, M.; Zverev, V.; 65. Transformative Vaccination: A Pentavalent Shield against COVID-19 and Influenza with Betulin-Based Adjuvant for Enhanced Immunity. Vaccine 2024, 42, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wu, M.; Zhou, C.; Lu, X.; Huang, B.; Zhang, N.; Zhao, H.; Chi, H.; Zhang, X.; Ling, D.; et al. Rational Development of a Combined mRNA Vaccine against COVID-19 and Influenza. npj Vaccines 2022, 7, 84–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, Q.; Li, M.; Mai, Q.; Ma, L.; Zhang, H.; Zhong, H.; Mai, K.; Cheng, N.; Feng, P.; et al. A Decavalent Composite mRNA Vaccine against Both Influenza and COVID-19. mBio 2024, 15. [Google Scholar] [CrossRef]
- Abbad, A.; Yueh, J.; Yellin, T.; Singh, G.; Carreño, J.M.; Clark, J.J.; Muramatsu, H.; Tiwari, S.; Bhavsar, D.; Alzua, G.P. Co-Administration of Seasonal Quadrivalent Influenza and COVID-19 Vaccines Leads to Enhanced Immune Responses to Influenza Virus and Reduced Immune Responses to SARS-CoV-2 in Naive Mice. Vaccine 2025, 50, 126825. [Google Scholar] [CrossRef]
- Thornhill-Wadolowski, E.; Ruter, D.L.; Yan, F.; Gajera, M.; Kurt, E.; Samanta, L.; Leigh, K.; Zhu, J.; Guo, Z.; Wang, Z. Development of an Influenza/COVID-19 Combination mRNA Vaccine Containing a Novel Multivalent Antigen Design That Enhances Immunogenicity of Influenza Virus B Hemagglutinins. Vaccines 2025, 13, 628. [Google Scholar] [CrossRef]
- McCann, N.; O’Connor, D.; Lambe, T.; Pollard, A.J. Viral Vector Vaccines. Current Opinion in Immunology 2022, 77, 102210–102210. [Google Scholar] [CrossRef] [PubMed]
- Vasilyev, K.; Shurygina, A.-P.; Sergeeva, M.; Stukova, M.; Egorov, A. Intranasal Immunization with the Influenza A Virus Encoding Truncated NS1 Protein Protects Mice from Heterologous Challenge by Restraining the Inflammatory Response in the Lungs. Microorganisms 2021, 9, 690–690. [Google Scholar] [CrossRef]
- Cao, K.; Wang, X.; Peng, H.; Ding, L.; Wang, X.; Hu, Y.; Dong, L.; Yang, T.; Hong, X.; Xing, M.; et al. A Single Vaccine Protects against SARS-CoV-2 and Influenza Virus in Mice. Journal of Virology 2022, 96. [Google Scholar] [CrossRef]
- Xing, M.; Hu, G.; Wang, X.; Wang, Y.; He, F.; Dai, W.; Wang, X.; Niu, Y.; Liu, J.; Liu, H.; et al. An Intranasal Combination Vaccine Induces Systemic and Mucosal Immunity against COVID-19 and Influenza. npj Vaccines 2024, 9, 64. [Google Scholar] [CrossRef]
- Li, J.; Arévalo, M.T.; Zeng, M. Engineering Influenza Viral Vectors. Bioengineered 2013, 4, 9–14. [Google Scholar] [CrossRef]
- García-Sastre, A.; Egorov, A.; Matassov, D.; Brandt, S.; Levy, D.E.; Durbin, J.E.; Palese, P.; Muster, T. Influenza A Virus Lacking the NS1 Gene Replicates in Interferon-Deficient Systems. Virology 1998, 252, 324–330. [Google Scholar] [CrossRef]
- Egorov, A.; Brandt, S.; Sereinig, S.; Romanova, J.; Ferko, B.; Katinger, D.; Grassauer, A.; Alexandrova, G.; Katinger, H.; Muster, T. Transfectant Influenza A Viruses with Long Deletions in the NS1 Protein Grow Efficiently in Vero Cells. Journal of Virology 1998, 72, 6437–6441. [Google Scholar] [CrossRef]
- Biosciences, V. Delta-19, Covid-19 + Universal Influenza Combination Vaccine. Available online: https://vivaldibiosciences.com/delta19.
- Sergeeva, M.V.; Vasilev, K.; Romanovskaya-Romanko, E.; Yolshin, N.; Pulkina, A.; Shamakova, D.; Shurygina, A.-P.; Muzhikyan, A.; Lioznov, D.; Stukova, M. Mucosal Immunization with an Influenza Vector Carrying SARS-CoV-2 N Protein Protects Naïve Mice and Prevents Disease Enhancement in Seropositive Th2-Prone Mice. Vaccines 2024, 13, 15–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, P.; Yuan, L.; Zhang, L.; Zhang, L.; Zhao, H.; Chen, C.; Wang, X.; Han, J.; Chen, Y. A Live Attenuated Virus-Based Intranasal COVID-19 Vaccine Provides Rapid, Prolonged, and Broad Protection against SARS-CoV-2. Science bulletin 2022, 67, 1372–1387. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, Y.; He, J.; Chen, J.; Qi, R.; Yuan, L.; Shao, T.; Zhao, H.; Chen, C.; Chen, Y. Intranasal Influenza-Vectored COVID-19 Vaccine Restrains the SARS-CoV-2 Inflammatory Response in Hamsters. Nature Communications 2023, 14, 4117. [Google Scholar] [CrossRef]
- Loes, A.N.; Gentles, L.E.; Greaney, A.J.; Crawford, K.H.; Bloom, J.D. Attenuated Influenza Virions Expressing the SARS-CoV-2 Receptor-Binding Domain Induce Neutralizing Antibodies in Mice. Viruses 2020, 12, 987. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, E.; Matyushenko, V.; Mezhenskaya, D.; Bazhenova, E.; Kotomina, T.; Rak, A.; Donina, S.; Chistiakova, A.; Kostromitina, A.; Novitskaya, V. Safety, Immunogenicity and Protective Activity of a Modified Trivalent Live Attenuated Influenza Vaccine for Combined Protection Against Seasonal Influenza and COVID-19 in Golden Syrian Hamsters. Vaccines 2024, 12, 1300. [Google Scholar] [CrossRef] [PubMed]
- Hause, A.M.; Zhang, B.; Yue, X.; Marquez, P.; Myers, T.R.; Parker, C.; Gee, J.; Su, J.; Shimabukuro, T.T.; Shay, D.K. Reactogenicity of Simultaneous COVID-19 mRNA Booster and Influenza Vaccination in the US. JAMA Network Open 2022, 5, e2222241–e2222241. [Google Scholar] [CrossRef]
- Izikson, R.; Brune, D.; Bolduc, J.-S.; Bourron, P.; Fournier, M.; Moore, T.M.; Pandey, A.; Perez, L.; Sater, N.; Shrestha, A.; et al. Safety and Immunogenicity of a High-Dose Quadrivalent Influenza Vaccine Administered Concomitantly with a Third Dose of the mRNA-1273 SARS-CoV-2 Vaccine in Adults Aged ≥65 Years: A Phase 2, Randomised, Open-Label Study. The Lancet Respiratory Medicine 2022, 10, 392–402. [Google Scholar] [CrossRef]
- McGrath, L.J.; Malhotra, D.; Miles, A.C.; Welch, V.L.; Di Fusco, M.; Surinach, A.; Barthel, A.; Alfred, T.; Jodar, L.; McLaughlin, J.M. Estimated Effectiveness of Co-administration of the BNT162b2 BA.4/5 COVID-19 Vaccine With Influenza Vaccine. JAMA Network Open 2023, 6, e2342151–e2342151. [Google Scholar] [CrossRef]
- Moro, P.L.; Zhang, B.; Ennulat, C.; Harris, M.; McVey, R.; Woody, G.; Marquez, P.; McNeil, M.M.; Su, J.R. Safety of Co-Administration of mRNA COVID-19 and Seasonal Inactivated Influenza Vaccines in the Vaccine Adverse Event Reporting System (VAERS) during July 1, 2021–June 30, 2022. Vaccine 2023, 41, 1859–1863. [Google Scholar] [CrossRef]
- Toback, S.; Galiza, E.; Cosgrove, C.; Galloway, J.; Goodman, A.L.; Swift, P.A.; Rajaram, S.; Graves-Jones, A.; Edelman, J.; Burns, F.; et al. Safety, Immunogenicity, and Efficacy of a COVID-19 Vaccine (NVX-CoV2373) Co-Administered with Seasonal Influenza Vaccines: An Exploratory Substudy of a Randomised, Observer-Blinded, Placebo-Controlled, Phase 3 Trial. The Lancet Respiratory Medicine 2022, 10, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, R.; Baos, S.; Cappel-Porter, H.; Carson-Stevens, A.; Clout, M.; Culliford, L.; Emmett, S.R.; Garstang, J.; Gbadamoshi, L.; Hallis, B.; et al. Safety and Immunogenicity of Concomitant Administration of COVID-19 Vaccines (ChAdOx1 or BNT162b2) with Seasonal Influenza Vaccines in Adults in the UK (ComFluCOV): A Multicentre, Randomised, Controlled, Phase 4 Trial. The Lancet 2021, 398, 2277–2287. [Google Scholar] [CrossRef]
- Venuto, R.; Giunta, I.; Cortese, R.; Denaro, F.; Pantò, G.; Privitera, A.; D’Amato, S.; Genovese, C.; La Fauci, V.; Fedele, F.; et al. The Importance of COVID-19/Influenza Vaccines Co-Administration: An Essential Public Health Tool. Infectious Disease Reports 2022, 14, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Pascucci, D.; Lontano, A.; Regazzi, L.; Marziali, E.; Nurchis, M.C.; Raponi, M.; Vetrugno, G.; Moscato, U.; Cadeddu, C.; Laurenti, P. Co-Administration of SARS-CoV-2 and Influenza Vaccines in Healthcare Workers: Results of Two Vaccination Campaigns in a Large Teaching Hospital in Rome. Human Vaccines & Immunotherapeutics 2023, 19. [Google Scholar] [CrossRef]
- Costantino, C.; Mazzucco, W.; Conforto, A.; Cimino, L.; Pieri, A.; Rusignolo, S.; Bonaccorso, N.; Bravatà, F.; Pipitone, L.; Sciortino, M.; et al. Real-Life Experience on COVID-19 and Seasonal Influenza Vaccines Co-Administration in the Vaccination Hub of the University Hospital of Palermo, Italy. Human Vaccines & Immunotherapeutics 2024, 20. [Google Scholar] [CrossRef]
- Wagenhäuser, I.; Reusch, J.; Gabel, A.; Höhn, A.; Lâm, T.-T.; Almanzar, G.; Prelog, M.; Krone, L.B.; Frey, A.; Schubert-Unkmeir, A.; et al. Immunogenicity and Safety of Co-administration of COVID-19 and Influenza Vaccination. European Respiratory Journal 2023, 61, 2201390–2201390. [Google Scholar] [CrossRef] [PubMed]
- Gonen, T.; Barda, N.; Asraf, K.; Joseph, G.; Weiss-Ottolenghi, Y.; Doolman, R.; Kreiss, Y.; Lustig, Y.; Regev-Yochay, G. Immunogenicity and Reactogenicity of Co-administration of COVID-19 and Influenza Vaccines. JAMA Network Open 2023, 6, e2332813–e2332813. [Google Scholar] [CrossRef]
- Murdoch, L.; Quan, K.; Baber, J.A.; Ho, A.W.Y.; Zhang, Y.; Xu, X.; Lu, C.; Cooper, D.; Koury, K.; Lockhart, S.P.; et al. Safety and Immunogenicity of the BNT162b2 Vaccine Coadministered with Seasonal Inactivated Influenza Vaccine in Adults. Infectious Diseases and Therapy 2023, 12, 2241–2258. [Google Scholar] [CrossRef]
- ModernaTX; Inc. A Safety, Reactogenicity, and Immunogenicity Study of mRNA-1073 (COVID-19/Influenza) Vaccine in Adults 18 to 75 Years Old. Available online: https://clinicaltrials.gov/study/NCT05375838.
- Lee, W.S.; Selva, K.J.; Audsley, J.; Kent, H. E.; Reynaldi, A.; Schlub, T. E.; Cromer, D.; Khoury, D. S.; Peck, H.; Aban, M.; et al. Randomized trial of same- versus opposite-arm coadministration of inactivated influenza and SARS-CoV-2 mRNA vaccines. JCI Insight 2025, 10, e187075. [Google Scholar] [CrossRef]
- Boikos, C.; Schaible, K.; Nunez-Gonzalez, S.; Welch, V.; Hu, T.; Kyaw, M.H.; Choi, L.E.; Kamar, J.; Goebe, H.; McLaughlin, J. Co-Administration of BNT162b2 COVID-19 and Influenza Vaccines in Adults: A Global Systematic Review. Vaccines 2025, 13, 381. [Google Scholar] [CrossRef]
- Janssen, C.; Mosnier, A.; Gavazzi, G.; Combadière, B.; Crépey, P.; Gaillat, J.; Launay, O.; Botelho-Nevers. Coadministration of seasonal influenza and COVID-19 vaccines: A systematic review of clinical studies. Human Vaccines & Immunotherapeutics 2022, 18, 2131166. [Google Scholar] [CrossRef]
- Al Hashimi, F.; Shuaib, S.E.; Bragazzi, N.L.; Chen, S.; Wu, J. COVID-19 Vaccine Timing and Co-Administration with Influenza Vaccines in Canada: A Systematic Review with Comparative Insights from G7 Countries. Vaccines (Basel) 2025, 13, 670. [Google Scholar] [CrossRef] [PubMed]
- BioNTech, S.E. A Study to Evaluate the Safety, Tolerability, and Immunogenicity of a Combined Modified RNA Vaccine Candidate Against COVID-19 and Influenza. Available online: https://clinicaltrials.gov/study/NCT06178991.
- ModernaTX; Inc. A Safety, Reactogenicity, and Immunogenicity Study of mRNA-1073 (COVID-19/Influenza) Vaccine in Adults 18 to 75 Years Old. Available online: https://clinicaltrials.gov/study/NCT05375838.
- ModernaTX, Inc. A Study of mRNA-1083 (SARS-CoV-2 and Influenza) Vaccine in Healthy Adult Participants, ≥50 Years of Age. Available online: https://clinicaltrials.gov/study/NCT06097273.
- GlaxoSmithKline. A Study of an Investigational Flu Seasonal/SARS-CoV-2 Combination Vaccine in Adults. Available online: https://clinicaltrials.gov/study/NCT06680375.
- A Study to Evaluate the Safety and Immunogenicity of COVID-19 and Influenza Combination Vaccine (COVID-19). Available online: https://clinicaltrials.gov/study/NCT05519839.
- Zhu, F.; Zhuang, C.; Chu, K.; Zhang, L.; Zhao, H.; Huang, S.; et al. Safety and immunogenicity of a live-attenuated influenza virus vector-based intranasal SARSCoV-2 vaccine in adults: randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Respir Med. 2022, 10, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Huang, S.; Liu, X.; Chen, Q.; Zhuang, C.; Zhao, H.; Han, J.; Jaen, A. M.; Do, T. H.; Peter, J. G.; et al. Safety and efficacy of the intranasal spray SARS-CoV-2 vaccine dNS1-RBD: a multicentre, randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir Med. 2023, 11, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Smorodintsev Research Institute of Influenza; Russia. Evalution of Corfluvec Vaccine for the Prevention of COVID-19 in Healthy Volunteers. Available online: https://clinicaltrials.gov/study/NCT05696067.
Figure 1.
Epidemiological situation of COVID-19 and influenza from 2020. (A) Epidemiological situation of COVID-19 and influenza in the world, adapted from World Health Organization (WHO) 2025, Influenza and SARS-CoV-2 virus detections reported to FluNet [21]; (B) Circulation of influenza and SARS-CoV-2 viruses in Russia based on data from the A.A. Smorodintsev Influenza Research Institute. Modified from [22].
Figure 1.
Epidemiological situation of COVID-19 and influenza from 2020. (A) Epidemiological situation of COVID-19 and influenza in the world, adapted from World Health Organization (WHO) 2025, Influenza and SARS-CoV-2 virus detections reported to FluNet [21]; (B) Circulation of influenza and SARS-CoV-2 viruses in Russia based on data from the A.A. Smorodintsev Influenza Research Institute. Modified from [22].
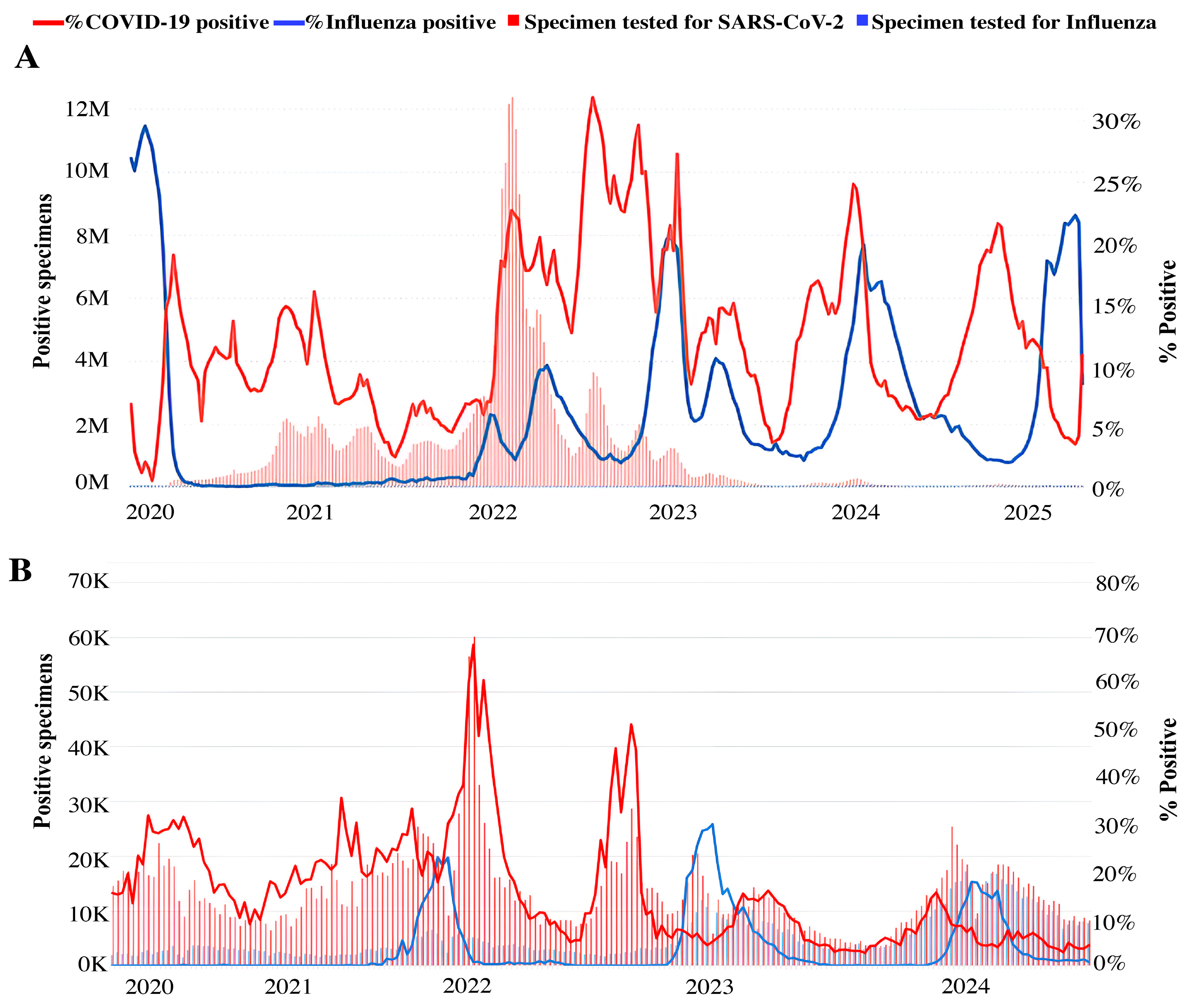
Figure 2.
Main biotechnological platforms used in combination vaccines against COVID-19 and seasonal influenza.
Figure 2.
Main biotechnological platforms used in combination vaccines against COVID-19 and seasonal influenza.
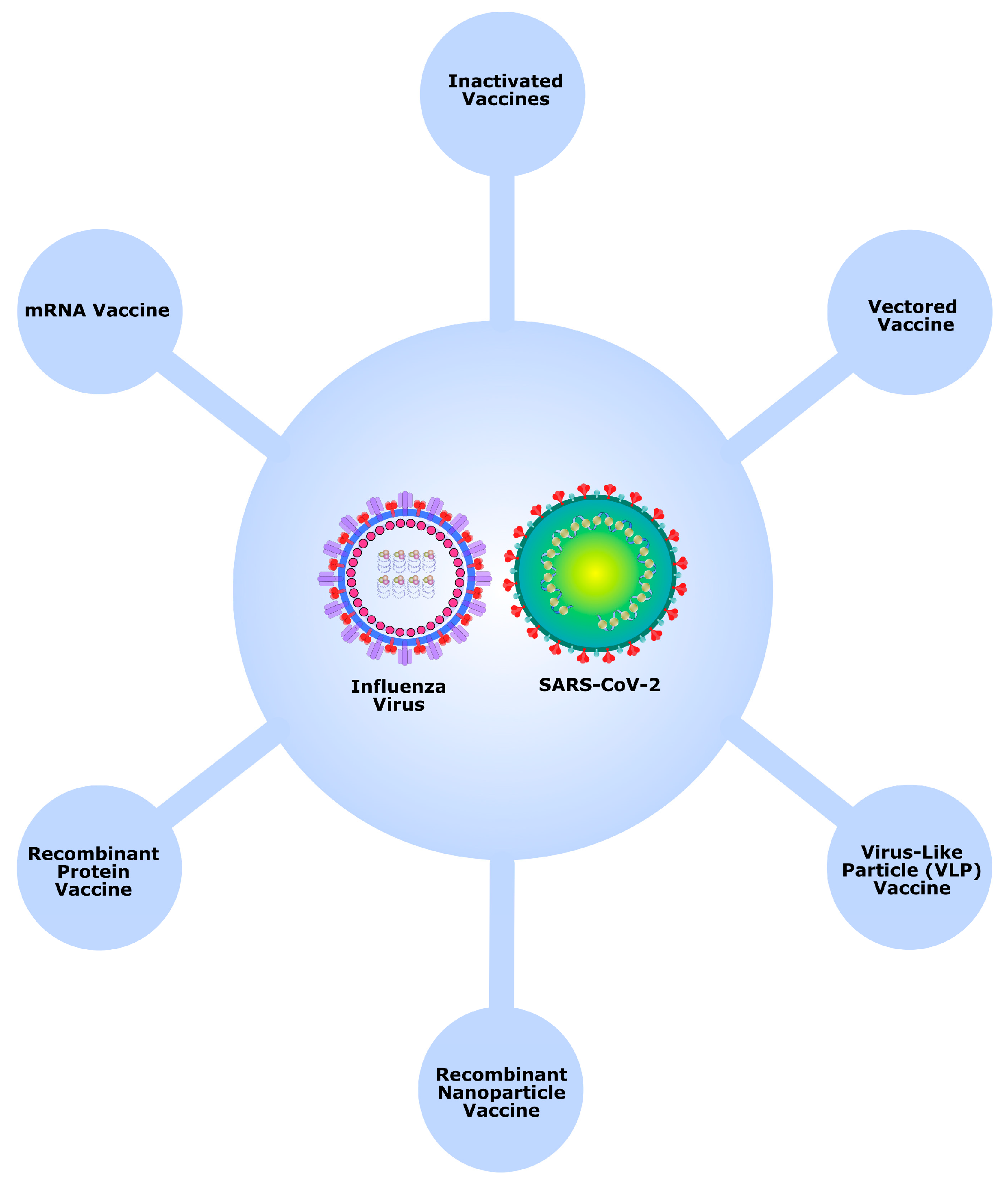
Figure 3.
Global participation in the simultaneous vaccination of COVID-19 and influenza. Green: countries that recommended simultaneous use. The table provides data from relevant clinical trials: A[83] B[84] C[85] D[86] E[87] F[88] G[89] H[90] I91] J[92] K[93] L[94].
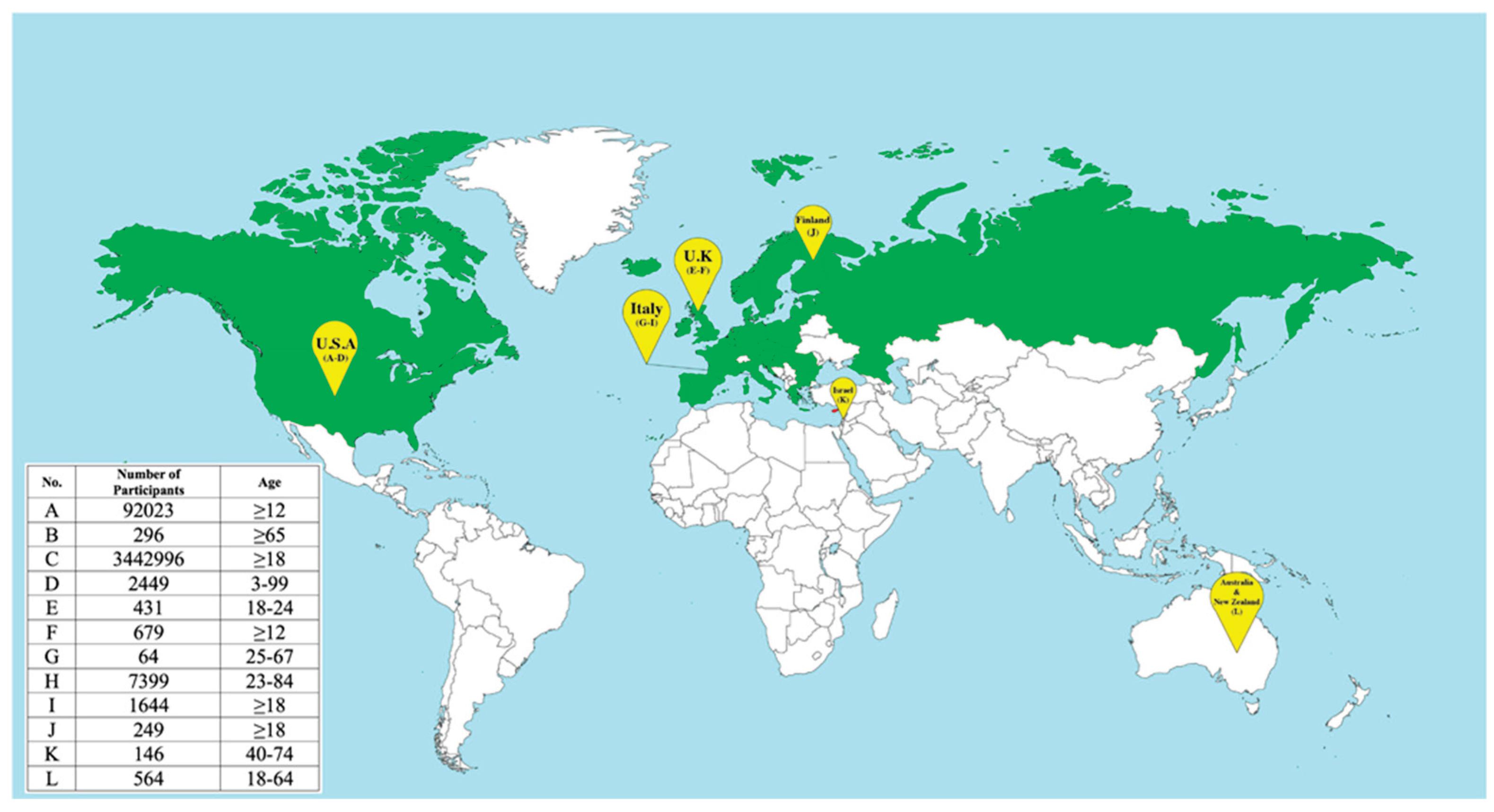

Table 1.
Animal studies of co-vaccination against SARS-CoV-2 and influenza virus.
| Vaccine | Platform | Composition | Administration route | Key results | Ref. |
|---|---|---|---|---|---|
| PiCoVacc/Flu vaccine | Inactivated SARS-CoV-2 Split-virion influenza Vaccine | Inactivated SARS-CoV-2 virus Split-virion influenza virus |
IP | Neutralizing antibodies Protection against SARS-CoV-2 and H1N1 infection |
[38] |
| Quadruple microparticulate vaccine | Inactivated viruses encapsulated into PLGA polymer microparticles | Inactivated SARS-CoV-2 Delta & Omicron variants. Inactivated Influenza A H1N1 & H3N2 variants. AddaVax adjuvant |
IN | Antigen-specific IgG (serum) and mucosal IgA (lung). Activation of cytotoxic (CD8+) and helper (CD4+) T-cells in lymph nodes and spleen. |
[59] |
| Chimeric Influenza virus | Live attenuated or inactivated virus | Chimeric virus in live attenuated or inactivated form displaying influenza HA and SARS-CoV-2 RBD on its envelope | IN, IM | Neutralizing antibodies Protection from lethal challenge with both pathogens in mice. |
[60] |
| Double-hit Flu-RBD vaccine |
VLP Vaccine | SARS-CoV-2 RBD conjugated onto inactivated influenza A virus |
IM | RBD-specific IgG2a and IgG1 Th1/Th2 balanced cellular immune response High protection efficacy against SARS-CoV-2 challenge in hamsters Strong neutralization activity against wild-type influenza A H1N1 inactivated virus in mice |
[61] |
| Trivalent S/H1/N1 enveloped VLP | VLP vaccine | Full length SARS-CoV-2 S-protein, H1N1 hemagglutinin (H1) and neur aminidase (N1) co-incorporated into enveloped VLP SLA Archaeosome adjuvant | IM | Specific IgG to S, H1, N1, and cellular immune responses stimulation | [62] |
| Self-assembling SARS-CoV-2 RBD-trimer and Influenza H1N1 HA1-trimer | Recombinant protein vaccine | SARS-CoV-2-RBD-trimer and HA1-trimer Liposomal saponin-based MA103 adjuvant |
IM | HAI for Influenza RBD-specific IgG Neutralizing antibody for SARS-CoV-2 Th1/Th2 balanced cellular immune response High protection efficacy against lethal SARS-CoV-2 and homogenous H1N1 influenza co-infection |
[63] |
| Flu-COVID combo vaccine | Recombinant protein vaccine | Truncated H1 and H3 hemagglutinin (aa 1–528) and SARS-CoV-2 S protein (aa 1–1213) AddaVax adjuvant |
IM | Neutralizing antibodies against both influenza and SARS-CoV-2 Specific IgGs against HA and S protein Protection from lethal challenge with both viruses in mice. |
[64] |
| Flu-COVID pentavalent vaccine | Recombinant protein vaccine | SARS-CoV-2 RBD fused with the Fc fragment of the human IgG hemagglutinin surface antigens of the viruses A/H1N1- pdm09, A/H3H2, B/Yamagata, B/Victoria Betulin-based adjuvant |
IM | RBD-specific and HA- specific IgG HAI for A/H1N1- pdm09, A/H3H2, B/Yamagata, B/Victoria viruses SARS-CoV-2 neutralization |
[65] |
| AR-CoV/IAV | mRNA-LNP | LNP-encapsulated mRNA encoding HA fromH1N1 and RBD from SARS-CoV-2 S protein | IM | Robust protective antibodies Antigen-specific cellular immune responses against SARS-CoV-2 and IAV Mice protection from coinfection with IAV and the SARS-CoV-2 Alpha and Delta variants |
[66] |
| FLUCOV-10 | mRNA- LNP | LNP-encapsulated mRNA encoding HA from H1N1 pdm09, H3N2, B/Victoria, B/Yamagata, H5N1, H7N9, S protein from four SARS-CoV-2 variants |
IM |
IgG antibodies, neutralizing antibodies, and antigen-specific cellular immune responses against all the vaccine-matched viruses of influenza and SARS-CoV-2
Complete protection in mouse models against both homologous and heterologous strains of influenza and SARS-CoV-2 |
[67] |
| QIV & BNT162b2 | QIV: split-virion vaccine BNT162b2: mRNA-LNP |
QIV: split-virion from 4 strains (A/H1N1, A/H3N2, B/Victoria, B/Yamagata) BNT162b2: LNP-encapsulated mRNA encoding SARS-CoV-2 S protein |
IM | HAI & Binding Antibody Titers (HA, NA) Binding & Neutralizing Antibody Titers Complete protection in mouse models against lethal challenge with either virus |
[68] |
| Influenza/COVID-19 Combination mRNA Vaccine | mRNA-LNP | LNP-encapsulated mRNA encoding: Influenza HA fusion proteins (dumbbell/trimer design with bacteriophage T4 foldon) from 4 strains (A/H1N1, A/H3N2, B/Victoria, B/Yamagata) SARS-CoV-2 RBD fusion protein (bivalent dumbbell). |
IM | Hemagglutination Inhibition (HAI) titers against A/H1N1, A/H3N2, B/Victoria, B/Yamagata. - SARS-CoV-2-specific: Neutralizing antibody titers against XBB.1.5 variant. |
[69] |
| AdC68-CoV/Flu | Vectored vaccine | Chimpanzee adenovirus 68 (AdC68) vector encoding SARS-CoV-2 RBD and H7N9 hemagglutinin | IM | Anti-H7 IgG Neutralizing antibody for SARS-CoV-2 Extensive RBD-specific T cell responses of splenocytes Protection against lethal SARS-CoV-2 challenge |
[72] |
| AdC68-HATRBD | Vectored Vaccine | Chimpanzee adenovirus 68 (AdC68) vector encoding SARS-CoV-2 RBD Beta-Alpha chimeric dimer and Omicron-Delta chimeric dimer, numerous SARS-CoV-2 T cell epitopes, and full-length HA of H1N1 pdm09 | IN | IgG, mucosal IgA, neutralizing antibodies, and memory T cells, protecting the mice from SARS-CoV-2 BA.5.2 and pandemic H1N1 infections. | [73] |
| Delta-19 | Vectored Vaccine | Influenza Delta NS1 vector expressesing key immunogenic proteins of SARS-CoV-2 and influenza viruses | IN | Neutralizing antibodies against SARS CoV-2 and Influenza viruses | [77] |
| FluVec-N | Vectored Vaccine | Influenza A/PR/8/34 (H1N1) vector carrying the SARS-CoV-2 N protein C terminal fragment (aa211–369) fused to the truncated NS1 gene |
IN | Neutralizing antibody for SARS-CoV-2 N-Protein Specific antibodies in BAL Virus-specific effector and resident CD8+ lymphocytes in lungs Reduced weight loss and viral load in the lungs following infection with the SARS-CoV-2 beta variant. |
[78] |
| dNS1-RBD Pneucolin | Vectored Vaccine | Live attenuated H1N1 pdm09 virus with NS1 gene replaced by SARS-CoV-2 RBD | IN | Local RBD-specific T cell response in the lung RBD-specific IgA and IgG response Attenuating pro-inflammatory cytokine levels post SARS-CoV-2 challenge, thereby reducing excess immune-induced tissue injury Cross-protection against H1N1 and H5N1 viruses |
[79,80] |
| ∆NA(RBD)-Flu | Vectored Vaccine | Influenza A/WSN/33 (H1N1) with H3 from A/Aichi/2/1968 and NA gene replaced by SARS-CoV-2 RBD |
IN | Neutralizing antibodies against SARS-CoV-2 and Influenza virus | [81] |
| 3×LAIV/CoV-2 | Vectored Vaccine | Licensed trivalent LAIV with H1N1/H3N2 strains encoding conserved SARS-CoV-2 T-cell epitopes. B/Victoria strain is unmodified | IN | Serum antibodies specific to all three influenza strains Protection against challenge from either influenza strain and SARS-CoV-2 challenge. T-cell response to SARS-CoV-2 epitopes |
[82] |
Abbreviations: IM - intramuscular, IN- intranasal, IP – intraperitoneal, HA – hemagglutinin, HAI - hemagglutination inhibition, NA – neuraminidase, RBD - receptor-binding domain, Ig – Immunoglobulin, BAL - Bronchoalveolar lavage, LNP - Lipid Nanoparticles, PLGA - Co-polymer of lactic and glycolic acids.
Table 2.
Parameters of clinical studies of influenza and COVID-19 vaccines co-administration.
| Vaccine | Target | Manufacturer | Group of Participants (age) |
Type of trial | Safety and reactogenicity profile | Immunogenicity | Ref. |
|---|---|---|---|---|---|---|---|
| NVX-CoV2373_ recombinant spike protein with matrix-M adjuvant |
SARS-CoV-2 |
Novavax |
Adult ≥18 (n = 15187) |
Clinical trial Phase 3 |
Safety- Local / Systemic Reaction–Adverse Events |
HAI for Seasonal Influenza A and B strains Anti-SARS-CoV-2-spike IgG |
[87] |
| Flucelvax Quadrivalent |
Influenza |
Seqirus |
|||||
| Fluad | |||||||
| COVID-19 mRNA booster vaccine | SARS-CoV-2 | Pfizer-BioNTech | ≥12 (n = 61390) |
Study on v-safe platform |
Safety–Local / Systemic Reaction |
N/A |
[83] |
| SIV | Influenza | N/A | |||||
| COVID-19 mRNA booster vaccine | SARS-CoV-2 | Moderna | ≥12 (n = 30633) |
||||
| SIV | Influenza | N/A | |||||
| mRNA-1273 | SARS-CoV-2 | Moderna |
Adult ≥65 (n = 306) |
Clinical trial Phase 2 |
Safety–Adverse Events | HAI for Seasonal Influenza A and B strains Anti-SARS-CoV-2-spike IgG |
[84] |
| Fluzone QIV-HD | Influenza | Sanofi Pasteur | |||||
| BNT162b2 BA.4/5 bivalent_mRNA | SARS-CoV-2 | Pfizer-BioNTech |
Adult ≥18 (n = 3442996) |
Retrospective comparative effectiveness study | Safety–Weighted hazard ratio | N/A |
[85] |
| SIV | Influenza | N/A | |||||
| COVID-19 mRNA booster vaccine | SARS-CoV-2 | N/A |
Median: 48 years (n = 2449) |
Vaccine Adverse Event Reporting System (VAERS) | Safety–Adverse Events | N/A |
[86] |
| QIV | Influenza | ||||||
| ChAdOx1 |
SARS-CoV-2 |
Pfizer-BioNTech |
Adult ≥!8 years (n=679) |
Multicentre, randomised, controlled, phase 4 trial ConFluCov study |
Safety- Local / Systemic Reaction–Adverse Events | HAI for Seasonal Influenza A and B strains Anti-SARS-CoV-2-spike IgG |
[88] |
| BNT162b2 | Pfizer-BioNTech | ||||||
| Adjuvanted TIV (FluAd (MF59) |
Influenza |
Seqirus
|
|||||
| Flucelvax QIV | Seqirus | ||||||
| Flublok Quadrivalent (QIVr) | Sanofi Pasteur | ||||||
| BNT162b2mRNA | SARS-CoV-2 | Pfizer-BioNTech |
Adult (n = 1231) |
Preference-based non-randomised controlled study | Safety |
Anti-SARS-CoV-2-spike IgG |
[92] |
| mRNA-1273 | SARS-CoV-2 | Moderna | |||||
| InfluvacTetra | Influenza | Abbott | |||||
| Omicron BA.4/BA.5–adapted bivalent_mRNA | SARS-CoV-2 | Pfizer-BioNTech |
Adult (n = 588) |
Prospective cohort study | Safety- Local / Systemic Reaction–Adverse Events |
Anti-SARS-CoV-2-spike IgG |
[93] |
| Influvac Tetra | Influenza | Abbott | |||||
| BNT162b2_mRNA | SARS-CoV-2 | Pfizer-BioNTech |
Adult 18–65 (n = 1134) |
Clinical trial Phase 3 |
Safety- Local / Systemic Reaction–Adverse Events | HAI for Seasonal Influenza A and B strains Anti-SARS-CoV-2-spike IgG |
[94] |
| Afluria Quad | Influenza | Seqirus | |||||
| mRNA-1273 | SARS-CoV-2 |
Moderna |
Adult 18-75 |
Clinical trial Phase 1/2 | Safety–Local / Systemic Reaction–Adverse Events | HAI for Seasonal Influenza A and B strains Titer of VAC62 Neutralizing Antibody for SARS-CoV-2 |
[95] |
| mRNA-1010 | Influenza | ||||||
| mRNA-1073 | SARS-CoV-2/influenza | ||||||
| Omicron XBB.1.5–containing COVID-19 mRNA booster vaccine (Spikevax) | SARS-CoV-2 | Moderna |
Adult (n=56) |
Open-label, randomized trial |
Safety- Local / Systemic Reaction–Adverse Events |
Neutralizing antibody, Anti-SARS-CoV-2-spike IgG |
[96] |
| Afluria Quad | Influenza | Seqirus | HAI for Seasonal Influenza H1, H3, and B-Vic |
Abbreviations: HAI - hemagglutination inhibition, N/A - not available, SIV - seasonal influenza vaccine, TIV - trivalent influenza vaccine, QIV - quadrivalent influenza vaccine, QIV-HD - high-dose quadrivalent influenza vaccine, V-safe - vaccine safety. Note: in all cases, the vaccines were administered intramuscularly.
Table 3.
Clinical trials of combined influenza and SARS-CoV-2 vaccines.
| Vaccine | Manufacturer | Platform | Administration route | Type of trial | Ref. |
|---|---|---|---|---|---|
| Combined modified RNA COVID-19 and Influenca vaccine |
Pfizer BioNTech |
mRNA LNP |
IM |
Phase 3 | [100] |
| Combined COVID-19 and Influenca mRNA 1073 |
Moderna |
mRNA LNP |
IM |
Phase 1,2 | [101] |
| Combined COVID-19 and Influenca mRNA 1083 |
Moderna |
mRNA LNP |
IM |
Phase 3 |
[102] |
| mRNA Flu/COVID-19 | GlaxoSmithKline | mRNA LNP | IM | Phase1 | [103] |
| qNIV/CoV2373 |
Novavax |
Nanoparticle vaccine |
IM | Phase 2 | [104] |
| dNS1-RBD, Pneucolin | Beijing Wantai Biological Pharmacy Enterprise | Vector vaccine |
IN |
Phase 1-3 | [105,106] |
| Corfluvec | Smorodintsev Research Institute of Influenza, Russia | Vector vaccine |
IN |
Phase 1,2 | [107] |
Abbreviations: LNP - lipid nanoparticles, IM - intramuscular, IN- intranasal.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



