
Submitted:
09 January 2026
Posted:
11 January 2026
You are already at the latest version
Abstract
Cardiovascular disease remains one of the leading causes of morbidity and mortality worldwide. Despite its of-ten-asymptomatic progression and therapeutic complexity, CVD is largely preventable through early intervention and lifestyle modification. A persistent challenge in its management, however, is the limited availability of effective pharmacological strategies aimed at reducing disease burden or mitigating associated risk factors. Polyphenols are a diverse class of bioactive compounds that have multiple phenol and hydroxyl groups, making them highly reactive and conferring them an antioxidant capacity, which help to modulate both cardiovascular and oncogenic processes. Polyphenols are naturally found in fruits and vegetables. Regular dietary intake of polyphenols has been linked to protective effects against various chronic conditions. This chapter provides a comprehensive overview of recent ad-vances in the development of polyphenol-based nutraceuticals, highlighting their therapeutic potential for the pre-vention and management of CVD. Experimental evidence, particularly from in vitro studies, suggests that polyphenols can influence cell viability and regulate key signaling pathways involved in the pathogenesis of cancer and CVD. Nonetheless, the biological response to polyphenol exposure varies across individuals, influenced by genetic, metabolic, and environmental factors. Over the past decade, epidemiological studies have consistently demonstrated that pol-yphenols may contribute to the normalization of clinical parameters such as blood pressure, glycemic control, lipid profiles, and body weight-factors intricately linked to cardiovascular risk.
Keywords:
cardiovascular disease
; polyphenols
; reactive oxygen species
; PI3K/Akt signaling
1. Introduction
Cardiovascular diseases (CVDs), encompassing conditions such as coronary artery disease, hypertension, and stroke, remain the foremost cause of mortality worldwide [1,2,3,4,5]. According to the World Health Organization, in 2022, 19.8 million people died because of CVD, which represents 32.2% of the total of deaths worldwide, of whom 85% were due to myocardial and brain-vascular accidents. The development of CVD is strongly influenced by modifiable lifestyle factors, including tobacco use, poor dietary habits, physical inactivity, obesity, dyslipidemia, hypertension, and excessive alcohol consumption [6,7,8]. Encouragingly, epidemiological evidence suggests that up to 80% of CVD-related deaths could be prevented by having a healthy lifestyle, namely, a balanced diet, regular physical activity, smoking cessation, and rigorous control of cardiovascular risk factors [4,6,9]. Among dietary components with therapeutic potential, polyphenols have garnered considerable attention due to their antioxidant, immunomodulatory, and vasodilatory properties, as well as their capacity to influence gene expression [6,10,11,12]. Polyphenols constitute a structurally diverse group of phytochemicals widely distributed in fruits, vegetables, and other plant-based foods. These compounds have been shown to regulate the expression of at least 54 genes and modulate the activity of numerous enzymes, thereby impacting a variety of cellular processes relevant to cardiovascular health.
Polyphenols exhibit potent antioxidant activity, participating in cellular redox reactions and mitigating oxidative stress by reducing the generation of reactive oxygen species (ROS). Specifically, they inhibit the formation of superoxide anions, hydrogen peroxide, and hypochlorous acid, thereby preserving cellular integrity and vascular function [2,4,13]. Among these compounds, flavonoids derived from olive oil have demonstrated the ability to enhance high-density lipoprotein cholesterol (HDL-c) functionality, facilitate reverse cholesterol transport, and protect vascular endothelium from atherogenic damage [1,7,12,14,15]. In addition to their antioxidant effects, polyphenols modulate hemostatic balance by inhibiting platelet aggregation and downregulating the expression of endothelial adhesion molecules. This action contributes to the protection of low-density lipoproteins (LDL) from oxidative modification, a key event in the initiation of atherosclerosis [11,16,17]. Polyphenols also exert anti-inflammatory effects during the progression of atherosclerosis. They suppress the expression of pro-inflammatory cytokines such as interleukins IL-1, IL-6, IL-8, tumor necrosis factor-alpha (TNF-α), and C-reactive protein [11,18]. In vitro studies have further demonstrated that polyphenols downregulate inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), TNF-α, and IL-6, primarily through inhibition of nuclear factor kappa B (NF-κB) signaling [19]. This leads to reduced expression of endothelial dysfunction markers and inflammatory mediators, including IL-1β, IL-2, IL-4, IL-6, E-selectin, and vascular cell adhesion molecule-1 (VCAM-1) [13,18,20]. Moreover, NF-κB interacts with key intracellular pathways such as mitogen-activated protein kinases (MAPKs) and the arachidonic acid cascade. Polyphenolic compounds have been shown to inhibit phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt), IκB kinase/c-Jun N-terminal kinase (IKK/JNK), and the mammalian target of rapamycin complex 1 (mTORC1), a central regulator of protein synthesis. They also modulate the Janus kinase/signal transducer and activator of transcription (JAK/STAT) pathway and suppress toll-like receptor (TLR) signaling, contributing to the downregulation of pro-inflammatory gene expression [21,22,23,24,25]. Scientific interest has been in comprehending the mechanisms of action and therapeutic applications of polyphenols due to their exceptional physiological potential in promoting health and avoiding disease. The importance of polyphenols in extending life and reducing the consequences of aging has been consistent in the literature. This review examines the molecular mechanisms by which polyphenols regulate gene expression and signaling pathways associated with CVD, highlighting their potential as nutraceutical agents in CVD prevention and therapy.
2. Overview of Cardiovascular Diseases
CVD encompasses a broad spectrum of disorders affecting the heart and vascular system, including hypertension, coronary artery disease (CAD), cerebrovascular disease, valvular and rheumatic heart conditions, cardiomyopathies, peripheral arterial disease, congenital heart defects, deep vein thrombosis, and pulmonary embolism [21,26]. Among these, atherosclerosis is recognized as the most prevalent underlying cause. Historically, atherosclerosis has been conceptualized as a form of chronic inflammation. In the mid-19th century, Austrian pathologist Carl von Rokitansky proposed that inflammation was a secondary response to other pathological insults. In contrast, Rudolf Virchow later advanced the notion that atherosclerosis itself constitutes a primary inflammatory disease [27,28]. Another theory postulates that the alteration of the endothelium and intima is due to mechanical injury, toxins, and oxygen free radicals, which lead to endothelial dysfunction [29]. These foundational perspectives have shaped contemporary understanding of vascular pathology. An alternative hypothesis suggests that endothelial and intimal injury, induced by mechanical stress, exposure to toxins, and oxidative damage from reactive oxygen species, initiates a cascade of events leading to endothelial dysfunction and subsequent plaque formation. This multifactorial process underscores the complexity of CVD pathogenesis and highlights the interplay between environmental insults and intrinsic vascular responses.
3. Epidemiology of Cardiovascular Diseases
CVDs remain the leading cause of mortality worldwide, accounting for approximately one in every three deaths globally [30,31]. Among these, ischemic heart disease, primarily driven by atherosclerotic coronary artery pathology, and stroke represent the most prevalent and fatal manifestations. In the United States, heart disease ranks as the foremost cause of death, while stroke occupies the fifth position (6, 25, 29). Notably, more than 75% of CVD-related deaths occur in low- and middle-income countries, where access to timely, equitable, and specialized healthcare services remains limited [6]. In such settings, delayed diagnosis and suboptimal management often result in disease detection at advanced stages, further exacerbating the burden of cardiovascular morbidity and mortality [31].
4. Vascular Endothelium and Its Role
The vascular endothelium is a dynamic monolayer of cells lining the lumen of blood vessels, serving as a critical interface between circulating blood and the vascular wall. Beyond its structural function, the endothelium acts as a highly selective barrier and metabolically active organ, playing a central role in maintaining vascular homeostasis. It regulates the delicate equilibrium between vasodilation and vasoconstriction, modulates the proliferation and migration of vascular smooth muscle cells, and orchestrates processes such as thrombogenesis and fibrinolysis [32]. Vasodilatory responses are primarily mediated by nitric oxide (NO), endothelium-derived hyperpolarizing factors, and prostacyclin. In contrast, vasoconstriction is driven by molecules such as endothelin-1, angiotensin II, thromboxane A2, and prostaglandin H2 [33], among these, NO is considered the most potent endogenous vasodilator. It contributes to vascular wall integrity by inhibiting platelet aggregation, suppressing inflammatory responses and oxidative stress, and preventing the migration and proliferation of smooth muscle cells and leukocyte adhesion [34,35]. These protective mechanisms are relevant across all stages of atherosclerotic development. However, chronic exposure to cardiovascular risk factors, including diabetes, dyslipidemia, hypertension, smoking, obesity, and oxidative stress, can overwhelm endothelial defense systems. This leads to a progressive loss of endothelial integrity, characterized by endothelial dysfunction, increased smooth muscle cell activity, and enhanced leukocyte adhesion and transmigration [36]. Both genetic predisposition and environmental influences may modulate the impact of these risk factors on endothelial function (36) (Figure 1). The preservation of endothelial integrity is essential for vascular health, and its disruption is increasingly recognized as a key contributor to the pathogenesis and clinical manifestation of cardiovascular diseases [35].
5. Nitric Oxide Production and Functions
NO is synthesized in endothelial cells from its precursor L-arginine through the enzymatic activity of endothelial nitric oxide synthase (eNOS), which is localized within caveolae. Under resting conditions, caveolin-1 binds to calmodulin, thereby inhibiting the activity of eNOS. Upon calcium influx, calmodulin binds calcium ions and displaces caveolin-1, resulting in eNOS activation and subsequent NO production. This process requires several cofactors, including tetrahydrobiopterin and nicotinamide adenine dinucleotide phosphate (NADPH), which are essential for optimal enzymatic function [37]. The expression and activity of eNOS are modulated by various physiological and pathological stimuli. For instance, laminar shear stress enhances eNOS expression, whereas asymmetric dimethylarginine (ADMA), an endogenous competitive inhibitor, impairs its activity. Elevated ADMA levels are commonly observed in individuals with cardiovascular risk factors, endothelial dysfunction, or established atherosclerosis [38].
NO plays a pivotal role in vascular homeostasis by mediating endothelium-dependent vasodilation. It inhibits platelet adhesion, leukocyte infiltration, and the proliferation of vascular smooth muscle cells. Additionally, NO prevents the oxidative modification of low-density lipoprotein cholesterol (LDL-c), a key event in the initiation of atherosclerotic plaque formation [39,40]. Conversely, reduced NO bioavailability or impaired NO signaling contributes to pro-atherogenic processes, including vasoconstriction, platelet aggregation, smooth muscle cell migration, leukocyte adhesion, and oxidative stress [40]. LDL-c itself can exacerbate endothelial dysfunction by upregulating caveolin-1 expression, which in turn suppresses NO synthesis via eNOS inhibition [21]. Moreover, oxidative stress interferes with NO signaling by promoting the rapid inactivation of NO through the formation of superoxide radicals and by degrading tetrahydrobiopterin, a critical cofactor for eNOS activity [41]. All these factors that reduce NO bioavailability are involved in the initiation, progression, and complications of atherosclerosis (Figure 2).
6. Endothelial Dysfunction and Atherosclerosis
To understand the potential of polyphenols in mitigating cardiovascular risk, it is essential to examine the pathophysiological mechanisms underlying atherosclerosis and endothelial dysfunction. Endothelial dysfunction is implicated in a range of pathological processes, including the loss of anticoagulant and anti-inflammatory properties, impaired regulation of vascular growth, and disruption of vascular remodeling. It is not only a triggering event but also a critical contributor to the progression of atherosclerotic cardiovascular disease. Importantly, endothelial dysfunction is recognized as an early biomarker of atherosclerosis, often preceding the detection of vascular lesions by angiography or ultrasound. Atherosclerosis itself is a chronic inflammatory condition that typically develops in regions of arterial vulnerability, particularly in medium-sized vessels. Lesions may remain clinically silent for years or even decades before manifesting as acute cardiovascular events. The transition from subclinical to symptomatic disease is often enhanced by the rupture or erosion of vulnerable atherosclerotic plaques. This exposes thrombogenic material to the bloodstream, resulting in the rapid formation of platelet-rich mural thrombi. These thrombi can partially or completely occlude the arterial lumen, leading to ischemic events such as acute myocardial infarction, unstable angina, or sudden cardiac death.
7. Shear Stress
Although the entire vascular system is susceptible to endothelial dysfunction, atherosclerotic lesions tend to develop preferentially in anatomically distinct regions such as arterial bifurcations, branching points, and the inner curvature of coronary segments. These sites are characterized by complex hemodynamic patterns, particularly disturbed shear forces generated by pulsatile blood flow, which act as critical modulators of the atherogenic process. Such localized variations in shear stress contribute to the regional and clinical heterogeneity observed in atherosclerosis. Specifically, areas of low endothelial shear stress (ESS) initiate a cascade of vascular responses that promote the transition toward an unstable atherogenic phenotype. Through mechanosensory and signal transduction mechanisms, low ESS alters the expression of endothelial genes, fostering a pro-inflammatory and pro-thrombotic environment conducive to early plaque formation. This hemodynamic disturbance enhances the production of endothelin-1 while suppressing the synthesis of NO and prostacyclin, key mediators of vessel-protection. Additionally, low ESS impairs lipid uptake and catabolism and promotes oxidative stress and inflammatory activation within endothelial cells, further accelerating the progression of atherosclerotic lesions.
8. Polyphenols and Their Cardiovascular Protective Mechanism
Epidemiological evidence suggests that regular dietary intake of polyphenols is associated with a reduced incidence of CVD, which are often driven by excessive production of ROS. Polyphenols represent the most abundant class of dietary antioxidants and are widely distributed in plant-based foods, including fruits, vegetables, cereals, olives, legumes, chocolate, tea, coffee, and wine [12]. Chemically, polyphenols are characterized by the presence of one or more aromatic rings bearing hydroxyl groups. Over 8,000 distinct polyphenolic structures have been identified, reflecting substantial diversity in their biological activity and molecular complexity [42,43]. Based on their chemical architecture, polyphenols are broadly classified into phenolic acids, non-flavonoids (e.g., stilbenes, lignans), and flavonoids [44,45] (Figure 3). Polyphenols exert antioxidant effects by neutralizing free radicals and protecting cellular components from oxidative damage. They also modulate the activity of various enzymes involved in redox balance and inflammatory signaling. Interestingly, under certain conditions and concentrations, polyphenols may exhibit pro-oxidant properties, which can influence cellular responses in a context-dependent manner [42,43,46]. Numerous preclinical studies have explored the therapeutic potential of polyphenols in the prevention and management of chronic diseases, including cancer, neurodegenerative disorders, diabetes, and CVD [42,43]. Their protective effects are largely attributed to their chemical reactivity and ability to prevent intracellular oxidation. Moreover, polyphenols activate nuclear factor erythroid 2–related factor 2 (Nrf2), a transcription factor that regulates the expression of detoxification and antioxidant enzymes. For instance, in animal models, polyphenolic compounds have been shown to induce the heme oxygenase-1 (HO-1) pathway, contributing to anti-inflammatory and cytoprotective responses relevant to cardiovascular health [47]. Beyond their antioxidant capacity, polyphenols interact with intracellular signaling cascades, modulate gene expression, and influence transcriptional regulators and microRNAs, thereby exerting broad regulatory effects on cellular homeostasis and vascular integrity [12].
9. Polyphenols in Cardiovascular Diseases
A growing body of evidence supports the role of natural products, particularly food-derived compounds and nutraceuticals, in modulating risk factors associated with atherosclerotic CVD (Table 1) [48,49,50]. Nutraceuticals, a term derived from “nutrition” and “pharmaceuticals,” refer to foods or food components that exert health-promoting effects with pharmacological properties. This category includes polyphenols, carotenoids, polyunsaturated fatty acids, and bioactive peptides [51]. Nutraceuticals have received recognition due to their nutritional benefits along with therapeutic effects and safety in a wide range of therapeutic areas. Some of the nutraceuticals and dietary supplements that are used for the treatment and prevention of CVDs are allicin and alliin, omega-3fatty acids, soy isoflavones, antioxidant vitamins, phytosterols, proteins, peptides and amino acids [50].
Atherosclerosis (ATS) is a chronic fibroproliferative condition characterized by low-grade inflammation and endothelial dysfunction, leading to the formation of cholesterol-rich plaques (atheroma) encased in fibrous caps. These lesions are driven by oxidative stress and the upregulation of inflammatory adhesion molecules produced by the endothelium [45]. Specific polyphenolic compounds, including epigallocatechin-3-gallate (EGCG), epigallocatechin (EGC), epicatechin (EC), acai-derived polyphenols (AC), and red muscadine grape phenolics (GP) have been shown to modulate nuclear factor kappa B (NF-κB), a key transcription factor activated during oxidative stress, genotoxic, and DNA damage. These compounds play a central role in regulating inflammatory cascades [52]. Additionally, curcumin, a polyphenol derived from turmeric, has demonstrated lipid-modifying properties, significantly improving parameters such as low-density lipoprotein cholesterol (LDL-C), non–high density lipoprotein cholesterol (non-HDL-C), and total lipoprotein levels [53]. These findings underscore the therapeutic potential of polyphenols in the prevention and management of CVD through multifaceted molecular mechanisms.
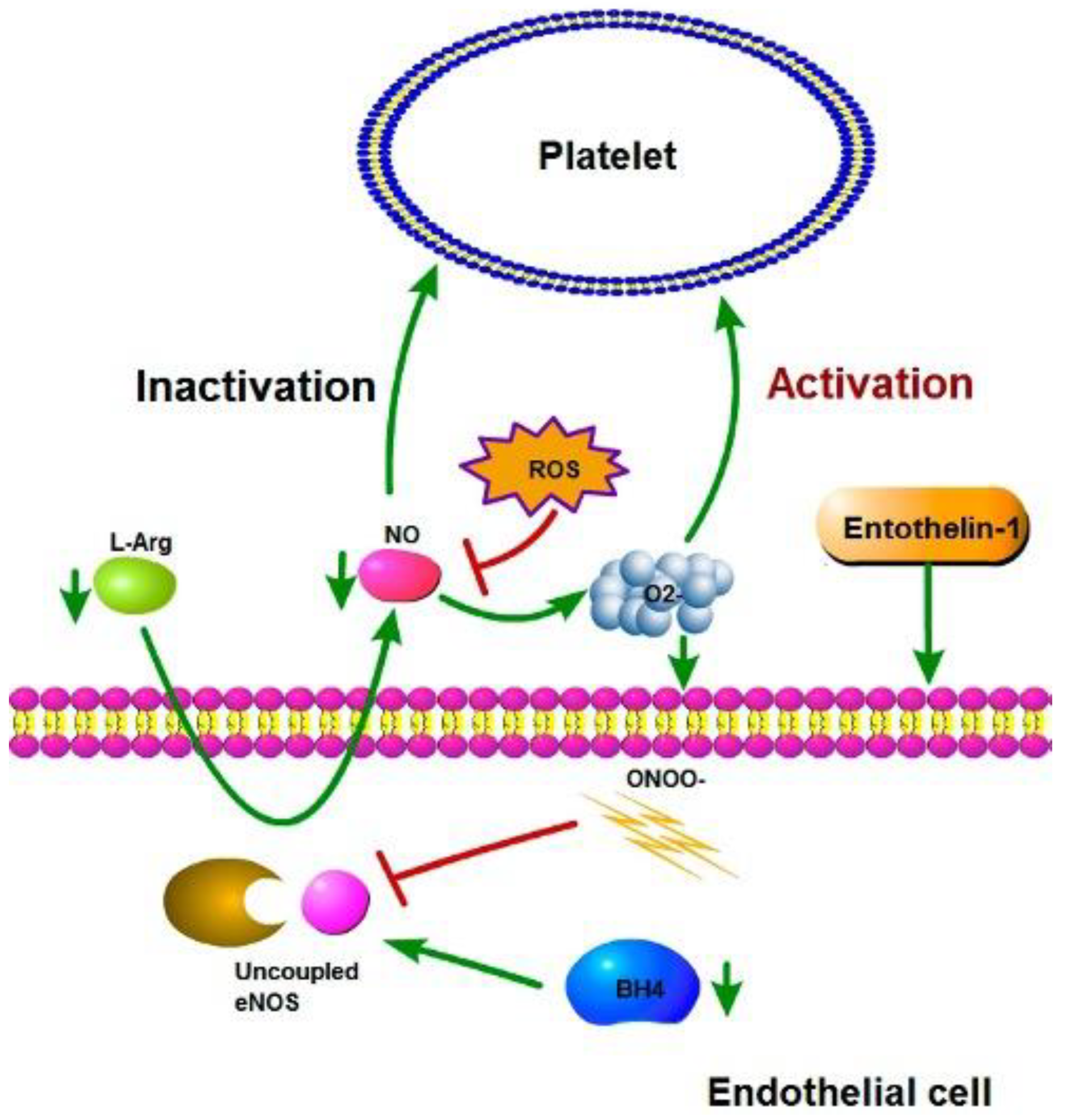
Figure 4.
Activation and inactivation of platelets. Under healthy conditions, eNOS remains coupled and produces NO; the NO production inactivates the platelets. ROS production can inhibit NO production and promote the activation of platelets.
Figure 4.
Activation and inactivation of platelets. Under healthy conditions, eNOS remains coupled and produces NO; the NO production inactivates the platelets. ROS production can inhibit NO production and promote the activation of platelets.
Endothelial dysfunction (ED) is widely recognized as an early and pivotal event in the development of macrovascular complications, particularly coronary artery disease (CAD), for which atherosclerosis remains the primary underlying cause [54]. Dietary flavonoids have been shown to possess vascular protective properties and are linked to a reduced risk of cardiovascular events, including myocardial infarction and stroke. Polyphenols derived from green tea, cocoa, red wine, and citrus fruits have shown notable efficacy in preserving endothelial integrity. For instance, cocoa consumption has been linked to enhance NO bioavailability, improved endothelial NO metabolism, and overall vascular protection. These effects contribute to the attenuation of cardiovascular risk factors such as insulin resistance [55]. Cocoa extracts have been shown to promote endothelial vasorelaxation, reduce circulating cholesterol levels, activate glucose transporter GLUT-2, and enhance insulin receptor phosphorylation thereby strengthening insulin signaling pathways. Epicatechin, the predominant flavonoid in dark chocolate, has been specifically implicated in improving insulin sensitivity and reducing insulin resistance [56]. Clinical studies have demonstrated that daily intake of flavanol-rich cocoa for one month in patients with CAD significantly improves endothelial function. This improvement is associated with the mobilization of functional circulating angiogenic cells, which play a critical role in vascular repair and the maintenance of endothelial homeostasis [57]. Furthermore, in hypertensive patients, consumption of 100 g/day of flavonoid-rich chocolate over 15 days resulted in marked reductions in insulin resistance and improvements in insulin sensitivity and function [58,59]. Additional studies involving healthy, overweight, and obese individuals have shown that daily intake of 500 mg of dark chocolate for 28 days leads to a significant decline in fasting glucose levels and homeostatic model assessment of insulin resistance (HOMA-IR), reinforcing the metabolic benefits of polyphenol-rich interventions [60].
The consumption of white and red wine has been associated with improved endothelial function in patients with CAD, an effect largely attributed to the vascular protective properties of wine-derived flavonoids [61]. In vitro studies have demonstrated that polyphenol-rich extracts from Vitis vinifera seeds and Aronia melanocarpa berries prolong clotting time and reduce the maximal velocity of fibrin polymerization in human plasma, suggesting a potential role in thrombosis prevention [16]. Grape seed proanthocyanidin extract has shown efficacy in reducing thrombus size and weight by downregulating key thrombogenic mediators, including P-selectin, von Willebrand factor, and cellular adhesion molecules (CAMs). These findings support its potential therapeutic application in conditions such as deep vein thrombosis [62]. A wide array of in vitro and in vivo studies has reported beneficial effects of polyphenols derived from natural sources, including plants and dietary components, on various cardiovascular pathologies. These include atherosclerosis, myocardial infarction, hypertension, dyslipidemia, and other vascular disorders [18]. The etiology of these conditions is multifactorial, often linked to lifestyle-related risk factors such as high dietary fat intake, tobacco use, excessive alcohol consumption, physical inactivity, aging, obesity, and metabolic disturbances, including hypertension and hyperglycemia [63]. Polyphenol intake typically occurs through the consumption of antioxidant-rich foods such as fruits, vegetables, green tea, and coffee, as well as through nutritional supplements and nutraceutical formulations [64]. For example, a polyphenolic extract from the Annurca apple has shown clinical benefits in patients with peripheral arterial disease (PAD), improving walking autonomy, hemodynamic parameters, and vascular abnormalities in the lower limbs. These findings suggest its potential utility in alleviating PAD-related symptoms and enhancing vascular function [65].
Polyphenolic compounds are absorbed primarily in the gastrointestinal tract and undergo extensive metabolism in intestinal and hepatic cells. Their bioavailability varies significantly depending on the biological system, metabolic pathways, and chemical structure of the compound [66]. The gut microbial metabolism is deeply involved in the biotransformation of dietary polyphenols into bioactive compounds [67,68]. Healthy gut microbiota can improve the bioavailability of phenolic compounds and contribute to enhancing the protective actions for skeletal and cardiac muscles [69,70,71]. Among their diverse mechanisms of action, the antioxidant capacity of polyphenols is particularly well-documented. These molecules inhibit oxidative DNA damage induced by peroxyl and other reactive radicals, suppress the expression of endothelial adhesion molecules, and reduce platelet aggregation, thereby protecting LDL from oxidative modification [64,72]. In addition to their effects on LDL, polyphenols also enhance the atheroprotective functions of HDL. Flavonoids derived from olive oil have been shown to improve cholesterol transport and promote vascular integrity, thereby reducing the risk of atheroma formation [73].
Furthermore, polyphenols exert anti-inflammatory effects by downregulating key pro-inflammatory cytokines, including interleukins IL-1, IL-6, and IL-8, as well as tumor necrosis factor-alpha (TNF-α) and C-reactive protein biomarkers that play a central role in the pathogenesis of cardiovascular diseases, such as atherosclerosis [18,73]. Polyphenols extracted from grapes have demonstrated significant vascular benefits, particularly in enhancing endothelial function and arterial resilience through NO-dependent mechanisms (Figure 2). These compounds stimulate endothelial NO synthesis, contributing to improved vasodilation and overall cardiovascular protection (Figure 3). Their biological activity encompasses anti-inflammatory, antihypertensive, and antithrombotic effects, largely attributed to their potent vasodilatory and antiplatelet properties [74,75]. Evidence also supports the role of polyphenol-rich dietary patterns, such as the Mediterranean diet, in reducing vascular inflammation. This effect is mediated by the neutralization of free radicals and the downregulation of proatherogenic gene expression, including low-density lipoprotein receptor-related protein 1 (LRP1), cyclooxygenase-2 (COX-2), and monocyte chemoattractant protein-1 (MCP-1). These molecular changes contribute to plaque stabilization and reduce endothelial rupture, thereby lowering the risk of atherothrombotic events [76,77]. Polyphenols further exert immunomodulatory effects by suppressing the expression of inflammatory biomarkers associated with plaque vulnerability. In addition, grape-derived polyphenols influence hepatic cholesterol metabolism and lipoprotein dynamics, reducing intestinal cholesterol absorption and altering its hepatic transport. These compounds modulate apolipoproteins A and B, both of which are implicated in cardiovascular risk and enhance lipoprotein lipase activity, leading to decreased plasma triglyceride levels and reduced circulating concentrations of very low-density lipoproteins (VLDL) and LDL [78]. Resveratrol, a prominent antioxidant found in grape skins, has shown antihypertensive effects through multiple mechanisms. These include activation of endothelial NO synthase (eNOS), modulation of the PI3K/Akt/eNOS signaling pathway, and reversal of endothelial dysfunction via compounds such as epigallocatechin gallate (EGCG), hesperetin, α-linolenic acid, ferulic acid, and catechin hydrate [79,80]. In animal models, resveratrol has also been shown to stimulate NO production, inhibit vascular inflammation, and reduce oxidative damage by upregulating Sirtuin 1 (SIRT1) expression in endothelial cells. Additional mechanisms include suppression of calcium influx and inhibition of platelet aggregation, further contributing to its vascular protective profile [11,81,82].
Resveratrol is a non-flavonoid polyphenol compound that mainly exists in grapes, berries, peanuts, and red wine, and has been reported to have protective effects on the heart by neutralizing reactive oxygen species, preventing inflammation, dilating blood vessels, increasing neoangiogenesis, suppressing apoptosis and delaying atherosclerosis (Table 1) [70,83]. Resveratrol has demonstrated protective effects in animal models of dyslipidemia and insulin resistance, notably reducing cardiac hypertrophy and improving myocardial contractility [81,82,84]. Its cardiovascular benefits are mediated through a range of molecular and physiological mechanisms, including the upregulation of glucose homeostasis, endothelial nitric oxide synthase (eNOS), adenosine monophosphate-activated protein kinase (AMPK), and peroxisome proliferator-activated receptor gamma (PPARγ). Concurrently, resveratrol downregulates glycated hemoglobin (HbA1c), oxidative stress markers, and interleukin-6 (IL-6), all of which are implicated in vascular dysfunction [51]. Most of resveratrol’s beneficial actions are endothelium-dependent and involve key signaling pathways such as AMPK, SIRT1, and nuclear factor erythroid 2-related factor 2 (Nrf2). These pathways contribute to enhanced nitric oxide bioavailability, reduced vascular smooth muscle tone, and diminished cellular contractility, partly through the inhibition of angiotensin II (Ang II) signaling [81,85]. Resveratrol is among the most extensively studied polyphenols for cardiovascular applications. Multiple reports indicate that it suppresses NADPH oxidase activity and reduces the overexpression of Nox2 and Nox4-isoforms strongly associated with heart failure. Elevated NADPH oxidase activity, often triggered by Ang II, endothelin-1, tumor necrosis factor-alpha (TNF-α), or mechanical stress, contributes to oxidative damage and cardiac remodeling. The presence of Nox2 and Nox4 in cardiomyocytes from failing hearts underscores the relevance of resveratrol’s inhibitory effects in conditions such as hypertension and heart failure [2,24].
Another effect of polyphenols is modifying different genes' expression and activating the endogenous antioxidant enzymatic defence system. For example, reports indicate that in murine models with arterial hypertension, a very common dietary polyphenol such as chlorogenic acid decreases the activity of NADPH oxidase and promotes the activation of enzymes such as superoxide dismutase, glutathione peroxidase, catalase, and glutathione-S-transferase in oxidative stress processes in the myocardium of rats [86]. Another polyphenol, such as quercetin, has also been shown to attenuate myocardial ischemia or reperfusion injuries in murine models by activating the phosphatidylinositol 3-kinase (PI3K)/Akt pathway [87]. Quercetin is a flavonoid present in olive oil, apples, onions, green tea, green leafy vegetables, blueberries, nuts, flowers, rinds, broccoli, red grapes, and dark cherries [88]. In vitro, in vivo, and clinical studies have shown that the cardioprotective functions of quercetin can be attributed to its ability to reduce blood pressure, enhance antioxidant potential, and exhibit other beneficial activities [4]. Quercetin has an anti-inflammatory role through the interleukin and TNF-α pathways and an antioxidant function through the cyclooxygenase/lipoxygenase pathways. Additionally, this bio-compound is an antihypertensive molecule that downregulates the autonomic nervous system, a primary cause of hypertension [25].
In human studies, quercetin has demonstrated potent anti-inflammatory effects, notably reducing circulating levels of C-reactive protein (CRP), a key biomarker implicated in the pathogenesis of atherosclerosis [89]. Additionally, quercetin lowers LDL cholesterol and inhibits its oxidative modification by macrophages, particularly in overweight individuals with elevated cardiovascular risk [90,91]. Quercetin’s antidiabetic mechanisms include the suppression of lipid peroxidation and the upregulation of endogenous antioxidant enzymes such as superoxide dismutase (SOD), glutathione peroxidase (GPX), and catalase (CAT). It also inhibits insulin-dependent phosphatidylinositol 3-kinase (PI3K) activation, contributing to improved insulin sensitivity [92]. Furthermore, quercetin stimulates adenosine monophosphate-activated protein kinase (AMPK), promoting the translocation and expression of glucose transporter type 4 (GLUT4) in skeletal muscle, thereby reducing insulin resistance [93]. Its anti-inflammatory, antiproliferative, and anti-atherosclerotic effects are mediated through activation of Sirtuin 1 (SIRT1), which modulates the AMPK/NADPH oxidase/AKT/eNOS signaling axis and suppresses oxidative stress induced by oxidized LDL (ox-LDL) in endothelial cells [94,95].
Beyond polyphenols, several non-polyphenolic natural compounds have shown promise in counteracting cardiovascular dysfunction. Berberine, derived from Berberis species, and curcumin, extracted from Curcuma longa, exert vascular protective effects primarily through the activation of antioxidant defense systems. Both compounds have demonstrated efficacy across a range of cardiovascular conditions [51]. Curcumin is a promising drug candidate, however, pharmacology and mechanism of action of curcumin have not been studied very deeply [96]. Curcumin protects cardiomyocyte injury after ischemia and hypoxia, alleviate vascular endothelial dysfunction, improve diabetic cardiomyopathy, inhibit myocardial hypertrophy and fibrosis, improve ventricular remodeling, reduce drug-induced myocardial injury, inhibit foam cell formation, and reduce vascular smooth muscle cells proliferation [96,97,98]. Curcumin’s mechanisms include inhibition of NADPH oxidase activity, downregulation of c-Jun N-terminal kinase (JNK), upregulation of cardiac metallothionein expression, and enhancement of nitric oxide bioavailability [99]. Brassica vegetables, such as cabbage, broccoli, cauliflower, and sprouts, exert vascular benefits through inhibition of angiotensin-converting enzyme (ACE) and renin, reduction of insulin resistance, and modulation of lipid metabolism [100]. Spirulina, a cyanobacterium rich in bioactive compounds including carotenoids, peptides, and fatty acids, may indirectly support cardiovascular health by improving glucose homeostasis. Its effects are mediated through activation of insulin receptor substrate (IRS)/PI3K/Akt and SIRT1/LKB1/AMPK signaling pathways [101,102].
Dietary polyphenols have been shown to influence the activity of transcription factors and modulate the biogenesis of small non-coding RNA molecules, particularly microRNAs (miRNAs). These miRNAs are critical regulators of metabolic homeostasis and oxidative stress responses, playing a pivotal role in the onset and progression of various diseases, including cardiovascular disorders [12,101,103]. In vitro studies have demonstrated that compounds such as quercetin, epigallocatechin gallate (EGCG), curcumin, and resveratrol can alter miRNA expression profiles in animal models, suggesting a mechanistic link between polyphenol intake and gene regulation [22,104,105,106]. These bioactive compounds are widely available as dietary supplements and have been classified as “Generally Recognized as Safe” (GRAS) by both the U.S. Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA). Their antioxidant properties include protection of DNA, proteins, and lipids from oxidative damage, further supporting their therapeutic potential [12]. A substantial body of preclinical research, complemented by smaller-scale clinical studies, has documented the protective effects of polyphenols against CVD, hypertension, and metabolic syndrome, including dyslipidemia and type 2 diabetes [107]. Notably, Peng et al. (2019) investigated the cardiovascular effects of huangjiu, a traditional Chinese rice wine rich in polyphenols, particularly catechins [108]. In murine models, huangjiu administration significantly reduced atherosclerotic lesion formation. This effect was associated with the inhibition of matrix metalloproteinases MMP-2 and MMP-9, enzymes implicated in plaque destabilization and vascular remodeling. Furthermore, in vitro studies revealed that huangjiu-derived polyphenols exert anti-inflammatory effects by suppressing the production of NO, tumor necrosis factor-alpha (TNF-α), and interleukins IL-6 and IL-1β, key mediators in the inflammatory cascade of atherosclerosis [108]. These findings underscore the potential of culturally rooted nutraceuticals as adjunctive strategies in cardiovascular prevention.
Kosen-cha, a green tea variant rich in catechins, has shown promising cardiometabolic effects in preliminary human studies. In a pilot trial conducted by Katanasaka et al. (2020), six obese individuals consumed Kosen-cha (5 g/L/day) over a period of twelve weeks. The intervention resulted in significant improvements in serum triglyceride (TG) levels and insulin resistance; two key metabolic parameters associated with obesity and CVD risk (97). Additionally, Kosen-cha intake was linked to reduced vascular endothelial dysfunction and attenuation of cardiac hypertrophy, effects attributed to the anti-inflammatory and antioxidant activity of catechins in cardiomyocytes [109]. Polyphenols are known to stimulate the production of vessel-ptotective mediators, such as NO, which contributes to improved endothelial function and vascular homeostasis [15,107]. Chew et al. (2018) reported that daily consumption of 450 mL of cranberry extract beverage (CEB) over eight weeks increased NO levels in overweight individuals. The study also documented reductions in endothelin-1 and oxidized glutathione, alongside elevations in high-density lipoprotein (HDL) and interferon-γ, collectively contributing to a lower CVD risk profile in patients with hypercholesterolemia and type 2 diabetes mellitus (T2DM) [15,94]. Those participants received 450 mL of CEB daily for eight weeks. Results revealed that CBE reduced CVD risk factors by lowering endothelin-1, oxidizing glutathione, and elevating NO, HDL, and interferon-γ [15]. Population-level data further support the cardioprotective role of polyphenols. Adriouch et al. (2018) conducted a prospective study involving 84,158 healthy volunteers, reporting average polyphenol intakes of 975 ± 478 mg/day in women and 1087 ± 498 mg/day in men (95). Tea and herbal infusions were identified as primary sources of catechins, while strawberries and cherries contributed significantly to anthocyanin intake. High consumption of polyphenols, including catechins, flavanols, anthocyanins, and stilbenes, was associated with a reduced risk of coronary heart disease (CHD) and CVD [107]. Despite these encouraging findings, further investigation is warranted to elucidate the molecular mechanisms underlying polyphenol-mediated cardiovascular protection. Specifically, the identification and validation of biomarkers, such as gene and protein expression profiles, related to CHD and CD will be essential to confirm causality and refine therapeutic strategies.
Platelet activation plays a critical role in the pathogenesis of CVD and cerebrovascular disease (CD), contributing to thrombus formation and vascular occlusion [17]. While conventional antiplatelet and anticoagulant therapies offer protection against endothelial dysfunction and platelet hyperactivity, their use is often limited by adverse effects. In contrast, flavonoids have emerged as promising natural agents capable of modulating platelet function and reducing vascular risk. Studies by Bijak et al. (2011) demonstrated that grape seed extract (GSE) influences human coagulation parameters, while subsequent in vitro research revealed that GSE at a concentration of 15 mg/mL significantly prolonged activated partial thromboplastin time (APTT) and prothrombin time (PT), indicating anticoagulant potential (Figure 5) [16]. The antiplatelet activity of GSE is mechanistically linked to the vessel dilator-stimulated phosphoprotein (VASP) pathway. GSE interacts with P2Y1 and P2Y12 purinergic receptors, leading to inhibition of adenylate cyclase (AC) and protein kinase A (PKA). This suppression maintains VASP in its non-phosphorylated state, thereby preventing platelet activation [16,17]. Additionally, GSE flavonoids inhibit platelet aggregation by interfering with the phosphoinositide 3-kinase/protein kinase B (PI3K/PKB) signaling cascade. Upon binding to P2Y12 receptors, GSE prevents the activation of PI3K/PKB and downstream integrins such as αIIbβ3, which are essential for platelet aggregation. The inhibition of αIIbβ3 integrin activation ultimately reduces thrombus formation and contributes to vascular protection [17].
Recent studies have expanded our understanding of the systemic effects of polyphenols on cardiovascular health. Kapolou et al. (2021) evaluated the impact of polyphenol-rich compounds on lipid profiles, endothelial and thrombotic pathways, glycemic control, gene expression, and metabolic regulation in a cohort of 250 Greek individuals, revealing favorable outcomes across multiple cardiovascular parameters [8].
Even more, Taguchi et al. (2019) investigated the association between total polyphenol intake and cardiovascular mortality, reporting that individuals consuming approximately 1905 mg/day of polyphenols exhibited reduced CVD-related mortality. This intake notably exceeds the recommended daily intake of 900 mg/day [6]. Comparative data from other populations show considerable variability: French individuals consume an average of 1193 ± 510 mg/day, Polish populations 1756.5 ± 695.8 mg/day, Spaniards 820 ± 323 mg/day, and Japanese individuals 759 ± 420 mg/day [6,110]. These intake levels have been linked to reductions in inflammatory markers, cardiovascular risk factors, and body mass index (BMI), further supporting the protective role of polyphenols. Although these findings are promising, the precise correlation between polyphenol intake and long-term health outcomes remains to be fully elucidated. Mechanistic studies offer additional insight. For example, polyphenols extracted from black elderberry (BEE) have demonstrated potent antioxidant activity in HepG2 hepatic cells treated with 50–100 mg/mL. These extracts downregulated key genes involved in cholesterol metabolism, including sterol regulatory element-binding protein 2 (SREBP-2), 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGR), and low-density lipoprotein receptor (LDLR) [1,6]. Furthermore, BEE modulated cholesterol transport by suppressing Niemann-Pick C1 Like 1 (NPC1L1) expression and upregulating ATP-binding cassette (ABC) transporters A1, G5/G8, CYP7A1, and ABCB11-proteins involved in cholesterol efflux and bile acid synthesis (Figure 6) (90). Epigenetically, BEE reduced the expression of histone deacetylases HDAC4, HDAC6, and HDAC9, while enhancing the expression of Sirtuin family members SIRT1, SIRT2, and SIRT3, which are associated with improved metabolic and vascular function [1].
10. Future Trends
Diets rich in fruits, vegetables and other plants-based food with high polyphenol content represent a promising non-pharmacological strategy for cardiovascular disease prevention and management. However, further research is needed to elucidate the pharmacokinetics, bioavailability, end precise mechanisms of action of polyphenols. Future studies should also focus on personalized nutrition approaches, considering genetic, metabolic and microbiota-related factors that influence individual response to polyphenol intake. Additionally, the development of standardized polyphenol-based nutraceuticals and clinical trials will be essential to translate experimental findings into effective therapeutic interventions.
11. Conclusions
Polyphenols represent a diverse and biologically potent class of natural compounds with multifaceted roles in cardiovascular protection. Their antioxidants, anti-inflammatory, antihypertensive, and antithrombotic properties have been extensively documented across in vitro, in vivo, and clinical studies. These effects are mediated through complex molecular mechanisms involving modulation of endothelial function, inhibition of platelet activation, regulation of lipid metabolism, and activation of key signaling pathways such as PI3K/Akt/eNOS, AMPK/SIRT1, and Nrf2. Beyond their biochemical activity, polyphenols influence gene expression, epigenetic regulation, and microRNA biogenesis—highlighting their capacity to reshape cellular responses at the transcriptional and post-transcriptional levels. Compounds such as quercetin, resveratrol, curcumin, and catechins have demonstrated efficacy in improving insulin sensitivity, reducing oxidative stress, and stabilizing atherosclerotic plaques.
Population-based studies further support the cardioprotective potential of polyphenol-rich diets, with evidence linking higher intake levels to reduced incidence and mortality from coronary and cerebrovascular diseases. While the precise dose-response relationship and biomarker validation remain areas for future research, the cumulative evidence positions polyphenols as promising nutraceutical agents in the prevention and management of cardiovascular disease. Considering their safety profile, accessibility through dietary sources, and broad spectrum of biological activity, polyphenols offer a compelling adjunctive strategy to complement conventional therapies. Continued investigation into their molecular targets, clinical efficacy, and long-term outcomes will be essential to fully harness their therapeutic potential in cardiovascular medicine.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, A.M.Ch. and writing—original draft preparation, A.M.Ch; writing—review and editing, A.M.Ch., A.C.C.N., I.V.R., C.B.R., G.E.M; supervision, A.M.Ch. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
NA.
Informed Consent Statement
NA.
Data Availability Statement
NA.
Acknowledgments
The authors thank Raúl Rodríguez-Herrera for his valuable feedback and critical review of the manuscript, which greatly contributed to its improvement.
Conflicts of Interest
There are no conflicts of interest.
References
- Jeon, S. et al., The Effects of Polyphenol-Rich Black Elderberry on Oxidative Stress and Hepatic Cholesterol Metabolism. Applied Sciences 2021, 11(21). [Google Scholar] [CrossRef]
- Pechanova, O.; Dayar, E.; Cebova, M. Therapeutic Potential of Polyphenols-Loaded Polymeric Nanoparticles in Cardiovascular System. Molecules 2020, 25(15). [Google Scholar] [CrossRef]
- Parella, P. The value of epigenetic biomarkers in breast cancer. Biomarkers in Medicine 2018, 12(09), p. 4. [Google Scholar] [CrossRef]
- Mirsafaei, L. et al., Molecular and Biological Functions of Quercetin as a Natural Solution for Cardiovascular Disease Prevention and Treatment. Plant Foods Hum Nutr 2020, 75(3), 307–315. [Google Scholar] [CrossRef]
- Zujko, M.E. et al., Dietary Total Antioxidant Capacity and Dietary Polyphenol Intake and Prevalence of Metabolic Syndrome in Polish Adults: A Nationwide Study. Oxid Med Cell Longev 2018, 7487816. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, C. et al., Dietary intake of total polyphenols and the risk of all-cause and specific-cause mortality in Japanese adults: the Takayama study. Eur J Nutr 2020, 59(3), 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. et al., Gut Microbiota: A Novel Regulator of Cardiovascular Disease and Key Factor in the Therapeutic Effects of Flavonoids. Front Pharmacol 2021, 12, 651926. [Google Scholar] [CrossRef] [PubMed]
- Kapolou, A. et al., Association of Mean Daily Polyphenols Intake with Mediterranean Diet Adherence and Anthropometric Indices in Healthy Greek Adults: A Retrospective Study. Applied Sciences 2021, 11(10). [Google Scholar] [CrossRef]
- Godos, J. et al., Dietary Polyphenol Intake, Blood Pressure, and Hypertension: A Systematic Review and Meta-Analysis of Observational Studies. Antioxidants (Basel) 2019, 8(6). [Google Scholar]
- Wisnuwardani, R.W. et al., Polyphenol intake and metabolic syndrome risk in European adolescents: the HELENA study. Eur J Nutr 2020, 59(2), 801–812. [Google Scholar] [CrossRef]
- Dyck, G.J.B. et al., The Effects of Resveratrol in Patients with Cardiovascular Disease and Heart Failure: A Narrative Review. Int J Mol Sci 2019, 20(4). [Google Scholar] [CrossRef] [PubMed]
- Cione, E. et al., Quercetin, Epigallocatechin Gallate, Curcumin, and Resveratrol: From Dietary Sources to Human MicroRNA Modulation. Molecules 2019, 25(1). [Google Scholar] [CrossRef]
- Margina, D. et al., Analysis of the intricate effects of polyunsaturated fatty acids and polyphenols on inflammatory pathways in health and disease. Food Chem Toxicol 2020, 143, 111558. [Google Scholar] [CrossRef]
- Magrone, T. et al., Recent Advances on the Anti-Inflammatory and Antioxidant Properties of Red Grape Polyphenols: In Vitro and In Vivo Studies. Antioxidants (Basel) 2019, 9(1). [Google Scholar]
- Chew, B. et al., Chronic consumption of a low calorie, high polyphenol cranberry beverage attenuates inflammation and improves glucoregulation and HDL cholesterol in healthy overweight humans: a randomized controlled trial. Eur J Nutr 2019, 58(3), 1223–1235. [Google Scholar] [CrossRef]
- Bijak, M. et al., Anticoagulant effect of polyphenols-rich extracts from black chokeberry and grape seeds. Fitoterapia 2011, 82(6), 811–817. [Google Scholar] [CrossRef]
- Bijak, M. et al., Dual Anticoagulant/Antiplatelet Activity of Polyphenolic Grape Seeds Extract. Nutrients 2019, 11(1). [Google Scholar] [CrossRef] [PubMed]
- Ranneh, Y. et al., Honey and its nutritional and anti-inflammatory value. BMC Complement Med Ther 2021, 21(1), 30. [Google Scholar] [CrossRef]
- Nikawa, T.; Ulla, A.; Sakakibara, I. Polyphenols and Their Effects on Muscle Atrophy and Muscle Health. Molecules 2021, 26(16). [Google Scholar] [CrossRef] [PubMed]
- Razquin, C.; Martinez-Gonzalez, M.A. A Traditional Mediterranean Diet Effectively Reduces Inflammation and Improves Cardiovascular Health. Nutrients 2019, 11(8). [Google Scholar] [CrossRef]
- Mozaffari, H. et al., Dietary Total Antioxidant Capacity and Cardiovascular Disease Risk Factors: A Systematic Review of Observational Studies. J Am Coll Nutr 2018, 37(6), 533–545. [Google Scholar] [CrossRef]
- Rasheed, Z. et al., MicroRNA-125b-5p regulates IL-1beta induced inflammatory genes via targeting TRAF6-mediated MAPKs and NF-kappaB signaling in human osteoarthritic chondrocytes. Sci Rep 2019, 9(1), p. 6882. [Google Scholar] [CrossRef]
- Lacroix, S. et al., A computationally driven analysis of the polyphenol-protein interactome. Sci Rep 2018, 8(1), 2232. [Google Scholar] [CrossRef]
- Almajdoob, S.; Hossain, E.; Anand-Srivastava, M.B. Resveratrol attenuates hyperproliferation of vascular smooth muscle cells from spontaneously hypertensive rats: Role of ROS and ROS-mediated cell signaling. Vascul Pharmacol 2018, 101, 48–56. [Google Scholar] [CrossRef]
- Kleemann, R. et al., Anti-inflammatory, anti-proliferative and anti-atherosclerotic effects of quercetin in human in vitro and in vivo models. Atherosclerosis 2011, 218(1), 44–52. [Google Scholar] [CrossRef]
- Tian, J. et al., Comparative study on the effects of apple peel polyphenols and apple flesh polyphenols on cardiovascular risk factors in mice. Clin Exp Hypertens 2018, 40(1), 65–72. [Google Scholar] [CrossRef] [PubMed]
- Frostegard, J. et al., Atherosclerosis in patients with autoimmune disorders. Arterioscler Thromb Vasc Biol 2005, 25(9), 1776–85. [Google Scholar] [CrossRef] [PubMed]
- Mayerl, C. et al., Atherosclerosis research from past to present--on the track of two pathologists with opposing views, Carl von Rokitansky and Rudolf Virchow. Virchows Arch 2006, 449(1), 96–103. [Google Scholar] [CrossRef]
- Ross, R. The pathogenesis of atherosclerosis a perspective for the 1990s. Nature 1993, 362(1), 6. [Google Scholar] [CrossRef]
- Benjamin, E.J. et al., Heart Disease and Stroke Statistics-2017 Update: A Report From the American Heart Association. Circulation 2017, 135(10), e146–e603. [Google Scholar] [CrossRef] [PubMed]
- Soriano, J.B. et al., Global, regional, and national deaths, prevalence, disability-adjusted life years, and years lived with disability for chronic obstructive pulmonary disease and asthma, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. The Lancet Respiratory Medicine 2017, 5(9), 691–706. [Google Scholar] [CrossRef] [PubMed]
- Lusher, *!!! REPLACE !!!*. T.F.A.B., M Biology of the Endothelium. Clin. Cardiol. 1997, 20, 7. [Google Scholar]
- Bonetti, P.O.; Lerman, L.O.; Lerman, A. Endothelial dysfunction: a marker of atherosclerotic risk. Arterioscler Thromb Vasc Biol 2003, 23(2), 168–75. [Google Scholar] [CrossRef]
- Tousoulis, D. et al., Endothelial dysfunction in conduit arteries and in microcirculation. Novel therapeutic approaches. Pharmacol Ther 2014, 144(3), 253–67. [Google Scholar] [CrossRef]
- Vita, J.A.; Keaney, J.F., Jr. Endothelial function: a barometer for cardiovascular risk? Circulation 2002, 106(6), 640–2. [Google Scholar] [CrossRef]
- Deanfield, J.E.; Halcox, J.P.; Rabelink, T.J. Endothelial function and dysfunction: testing and clinical relevance. Circulation 2007, 115(10), 1285–95. [Google Scholar] [CrossRef]
- Behrendt, D.a.G. P, Endothelial Function: From Vascular Biology to Clinical Applications. Am J Cardiol 2002, 90. [Google Scholar]
- C.J., P. Does ADMA Cause Endothelial Dysfunction? Arterioscler Thromb Vasc Biol 2000, 20(1), p. 5. [Google Scholar]
- H, R. et al., Interactions of Nitric Oxide and Peroxynitrite with Low-Density Lipoprotein. Biol.Chem 2002, 383, 5. [Google Scholar]
- Endres, M. et al., Stroke protection by 3-hydroxy-3-methylglutaryl (HMG)-CoA reductase inhibitors mediated by endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 1998, 95(1), p. 5. [Google Scholar] [CrossRef] [PubMed]
- Kinlay, S. et al., Role of Endothelin-1 in the Active Constriction of Human Atherosclerotic Coronary Arteries. Circulation 2001, 104(1), 4. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2(12), 1231–1246. [Google Scholar] [CrossRef]
- Scalbert, A. et al., Dietary polyphenols and the prevention of diseases. Crit Rev Food Sci Nutr 2005, 45(4), 287–306. [Google Scholar]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Current Opinion in Food Science 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Beconcini, D. Antioxidant and Anti-Inflammatory Properties of Cherry Extract: Nanosystems-Based Strategies to Improve Endothelial Function and Intestinal Absorption. Foods 2020, 9(2), 207. [Google Scholar]
- Abbas, M. et al., Natural polyphenols: An overview. International Journal of Food Properties 2017, 20(8), 1689–1699. [Google Scholar]
- Barbagallo, I. et al., Potential therapeutic effects of natural heme oxygenase-1 inducers in cardiovascular diseases. Antioxid Redox Signal 2013, 18(5), 507–21. [Google Scholar] [CrossRef]
- Caliceti, C. et al., New Trends in Antioxidant Compounds: A Precise Nutraceutical in Cardiometabolic Disorders. Oxid Med Cell Longev 2019, 4794563. [Google Scholar]
- Bahorun, T.; Aruoma, O.I.; Neergheen-Bhujun, V.S. Phytomedicines, nutraceuticals, and functional foods regulatory framework, in Nutraceutical and Functional Food Regulations in the United States and around the World; 2019; pp. 509–521. [Google Scholar]
- Puri, V. et al., A Comprehensive Review on Nutraceuticals: Therapy Support and Formulation Challenges. Nutrients 2022, 14(21). [Google Scholar]
- Carrizzo, A. et al., A Novel Promising Frontier for Human Health: The Beneficial Effects of Nutraceuticals in Cardiovascular Diseases. Int J Mol Sci 2020, 21(22). [Google Scholar] [CrossRef] [PubMed]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food and Chemical Toxicology 2018, 113, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Bahramsoltani, R. et al., Dietary polyphenols for atherosclerosis: A comprehensive review and future perspectives. Crit Rev Food Sci Nutr 2019, 59(1), 114–132. [Google Scholar] [CrossRef] [PubMed]
- Golia, E. et al., Inflammation and cardiovascular disease: from pathogenesis to therapeutic target. Curr Atheroscler Rep 2014, 16(9), 435. [Google Scholar] [CrossRef]
- Rabadan-Chavez, G.M. et al., The prothrombotic state associated with obesity-induced hypertension is reduced by cocoa and its main flavanols. Food Funct 2016, 7(12), 4880–4888. [Google Scholar] [CrossRef]
- Grassi, D. et al., Cocoa, glucose tolerance, and insulin signaling: cardiometabolic protection. J Agric Food Chem 2015, 63(45), 9919–26. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C. et al., Improvement of Endothelial Function With Dietary Flavanols Is Associated With Mobilization of Circulating Angiogenic Cells in Patients With Coronary Artery Disease. Journal of the American College of Cardiology 2010, 56(3), 218–224. [Google Scholar] [CrossRef]
- Grassi, D. et al., Short-term administration of dark chocolate is followed by a significant increase in insulin sensitivity and a decrease in blood pressure in healthy persons. Am J Clin Nutr 2005, 81(1), p. 4. [Google Scholar] [CrossRef]
- Grassi, D. et al., Blood Pressure Is Reduced and Insulin Sensitivity Increased in Glucose-Intolerant, Hypertensive Subjects after 15 Days of.
- Consuming High-Polyphenol Dark Chocolate. The Journal of nutritional biochemistry 2008, 5.
- Almoosawi, S. et al., Differential effect of polyphenol-rich dark chocolate on biomarkers of glucose metabolism and cardiovascular risk factors in healthy, overweight and obese subjects: a randomized clinical trial. Food Funct 2012, 3(10), 1035–43. [Google Scholar] [CrossRef]
- Whelan, A. et al., Effects of white and red wine on endothelial function in subjects with coronary artery disease. 2004, 34(5), 224–228. [Google Scholar]
- Zhang, Y. et al., Antithrombotic effect of grape seed proanthocyanidins extract in a rat model of deep vein thrombosis. Journal of Vascular Surgery 2011, 53(3), 743–753. [Google Scholar] [CrossRef]
- Tressera-Rimbau, A. et al., Dietary Polyphenols in the Prevention of Stroke. Oxid Med Cell Longev 2017, 7467962. [Google Scholar] [CrossRef]
- Parrella, E. et al., From Preclinical Stroke Models to Humans: Polyphenols in the Prevention and Treatment of Stroke. Nutrients 2020, 13(1). [Google Scholar] [CrossRef] [PubMed]
- Tenore, G.C. et al., Effect of Annurca Apple Polyphenols on Intermittent Claudication in Patients With Peripheral Artery Disease. The American Journal of Cardiology 2019, 123(5), 847–853. [Google Scholar] [CrossRef]
- Martinez-Huelamo, M. et al., Bioavailability of tomato polyphenols is enhanced by processing and fat addition: Evidence from a randomized feeding trial. Mol Nutr Food Res 2016, 60(7), 1578–89. [Google Scholar] [CrossRef]
- Luca, S.V. et al., Bioactivity of dietary polyphenols: The role of metabolites. Crit Rev Food Sci Nutr 2020, 60(4), 626–659. [Google Scholar] [CrossRef] [PubMed]
- Keohane, E. et al., Comparing Apples to Apples: Evaluating Foodomics in Precision Nutrition Research Featuring the Influence of Polyphenols on the Gut Microbiome. Nutrition Research 2025. [Google Scholar] [CrossRef]
- Ticinesi, A. et al., Accounting Gut Microbiota as the Mediator of Beneficial Effects of Dietary (Poly)phenols on Skeletal Muscle in Aging. Nutrients 2023, 15(10). [Google Scholar] [CrossRef] [PubMed]
- Zivarpour, P. et al., Resveratrol and cardiac fibrosis prevention and treatment. Current pharmaceutical biotechnology 2022, 23(2), 190–200. [Google Scholar] [CrossRef]
- Ticinesi, A. et al., Disentangling the Complexity of Nutrition, Frailty and Gut Microbial Pathways during Aging: A Focus on Hippuric Acid. Nutrients 2023, 15(5). [Google Scholar]
- de Camargo, A.C. et al., Low molecular weight phenolics of grape juice and winemaking byproducts: antioxidant activities and inhibition of oxidation of human low-density lipoprotein cholesterol and DNA strand breakage. J Agric Food Chem 2014, 62(50), 12159–71. [Google Scholar] [CrossRef]
- Millar, C.L.; Duclos, Q.; Blesso, C.N. Effects of Dietary Flavonoids on Reverse Cholesterol Transport, HDL Metabolism, and HDL Function. Adv Nutr 2017, 8(2), 226–239. [Google Scholar]
- Kattoor, A.J. et al., Oxidative Stress in Atherosclerosis. Curr Atheroscler Rep 2017, 19(11), p. 42. [Google Scholar]
- Lin, H.Y. et al., Epigenetic therapy combination of UNC0638 and CI-994 suppresses breast cancer via epigenetic remodeling of BIRC5 and GADD45A. Biomed Pharmacother 2021, 145, 112431. [Google Scholar] [CrossRef] [PubMed]
- Casas, R. et al., Anti-Inflammatory Effects of the Mediterranean Diet in the Early and Late Stages of Atheroma Plaque Development. Mediators Inflamm 2017, 3674390. [Google Scholar]
- Casas, R. et al., Nutrition and Cardiovascular Health. Int J Mol Sci 2018, 19(12). [Google Scholar] [CrossRef] [PubMed]
- Rasines-Perea, Z.; Teissedre, P.L. Grape Polyphenols' Effects in Human Cardiovascular Diseases and Diabetes. Molecules 2017, 22(1). [Google Scholar] [CrossRef]
- Suganya, N. et al., Reversibility of endothelial dysfunction in diabetes: role of polyphenols. British Journal of Nutrition 2016, 116(2), 223–246. [Google Scholar]
- Delmas, D. E.S.a.N.L., Resveratrol, a Phytochemical Inducer of Multiple Cell Death Pathways: Apoptosis, Autophagy and Mitotic Catastrophe. Current Medicinal Chemistry 2011, 18, 21. [Google Scholar] [CrossRef]
- Zordoky, B.N.; Robertson, I.M.; Dyck, J.R. Preclinical and clinical evidence for the role of resveratrol in the treatment of cardiovascular diseases. Biochim Biophys Acta 2015, 1852(6), 1155–77. [Google Scholar]
- Dolinsky, V.W. et al., Resveratrol prevents hypertension and cardiac hypertrophy in hypertensive rats and mice. Biochim Biophys Acta 2013, 1832(10), 1723–33. [Google Scholar]
- Fan, S. et al., Role of resveratrol in inhibiting pathological cardiac remodeling. Front Pharmacol 2022, 13, 924473. [Google Scholar]
- Chan, V.; Fenning, A.; Iyer, A.; Hoey, A.; Brown, L. Resveratrol improves cardiovascular function in DOCA-salt hypertensive rats. Current Pharmaceutical Biotechnology 2011, 12(3), p. 7. [Google Scholar] [CrossRef]
- Cao, X. et al., Resveratrol prevents AngII-induced hypertension via AMPK activation and RhoA/ROCK suppression in mice. Hypertens Res 2014, 37(9), 803–10. [Google Scholar] [PubMed]
- Akila, P.; Vennila, L. Chlorogenic acid a dietary polyphenol attenuates isoproterenol induced myocardial oxidative stress in rat myocardium: An in vivo study. Biomed Pharmacother 2016, 84, 208–214. [Google Scholar] [PubMed]
- Wei, H. et al., Cardioprotective Effects of Malvidin Against Isoproterenol-Induced Myocardial Infarction in Rats: A Mechanistic Study. Med Sci Monit 2017, 23, 2007–2016. [Google Scholar]
- Bhaskar, S. et al., Quercetin alleviates hypercholesterolemic diet induced inflammation during progression and regression of atherosclerosis in rabbits. Nutrition 2013, 29(1), 219–29. [Google Scholar]
- Schulza, H.U. et al., Investigation of Pharmacokinetic Data of Hypericin, Pseudohypericin, Hyperforin and the Flavonoids Quercetin and Isorhamnetin Revealed from Single and Multiple Oral Dose Studies with a Hypericum Extract Containing Tablet in Healthy Male Volunteers. Drug Res 2005, 55(10), p. 7. [Google Scholar]
- Xue, F. et al., Quercetin Inhibits LPS-Induced Inflammation and ox-LDL-Induced Lipid Deposition. Front Pharmacol 2017, 8, 40. [Google Scholar] [CrossRef]
- Mbikay, M. et al., Quercetin-3-glucoside increases low-density lipoprotein receptor (LDLR) expression, attenuates proprotein convertase subtilisin/kexin 9 (PCSK9) secretion, and stimulates LDL uptake by Huh7 human hepatocytes in culture. FEBS Open Bio 2014, 4, 755–62. [Google Scholar] [CrossRef]
- Stewart, L.K. et al., Failure of dietary quercetin to alter the temporal progression of insulin resistance among tissues of C57BL/6J mice during the development of diet-induced obesity. Diabetologia 2009, 52(3), 514–23. [Google Scholar] [CrossRef]
- Eid, H.M. et al., The molecular basis of the antidiabetic action of quercetin in cultured skeletal muscle cells and hepatocytes. Pharmacogn Mag 2015, 11(41), 74–81. [Google Scholar]
- Hung, C.H. et al., Quercetin is a potent anti-atherosclerotic compound by activation of SIRT1 signaling under oxLDL stimulation. Mol Nutr Food Res 2015, 59(10), 1905–17. [Google Scholar] [CrossRef]
- Kaur, G. M. C, Amelioration of obesity, glucose intolerance, and oxidative stress in high-fat diet and low-dose streptozotocin-induced diabetic rats by combination consisting of "curcumin with piperine and quercetin. ISRN Pharmacol 2012, 2012, 957283. [Google Scholar] [CrossRef]
- Yang, C. et al., Review of the Protective Mechanism of Curcumin on Cardiovascular Disease. Drug Des Devel Ther 2024, 18, 165–192. [Google Scholar] [CrossRef] [PubMed]
- Li, H. et al., Curcumin, the golden spice in treating cardiovascular diseases. Biotechnol Adv 2020, 38, 107343. [Google Scholar] [CrossRef] [PubMed]
- Pourbagher-Shahri, A.M. et al., Curcumin and cardiovascular diseases: Focus on cellular targets and cascades. Biomed Pharmacother 2021, 136, 111214. [Google Scholar] [CrossRef] [PubMed]
- Wongeakin, N.; Bhattarakosol, P.; Patumraj, S. Molecular mechanisms of curcumin on diabetes-induced endothelial dysfunctions: Txnip, ICAM-1, and NOX2 expressions. Biomed Res Int 2014, 161346. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Potential efficacy of broccoli sprouts as a unique supplement for management of type 2 diabetes and its complications. J Med Food 2013, 16(5), 375–82. [Google Scholar] [CrossRef]
- Cannataro, R. et al., Ketogenic Diet Acts on Body Remodeling and MicroRNAs Expression Profile. Microrna 2019, 8(2), 116–126. [Google Scholar] [CrossRef]
- Ren, Z. et al., C-Phycocyanin inhibits hepatic gluconeogenesis and increases glycogen synthesis via activating Akt and AMPK in insulin resistance hepatocytes. Food Funct 2018, 9(5), 2829–2839. [Google Scholar] [CrossRef]
- Cannataro, R. et al., Ketogenic Diet and microRNAs Linked to Antioxidant Biochemical Homeostasis. Antioxidants (Basel) 2019, 8(8). [Google Scholar]
- Boesch-Saadatmandi, C. et al., Effect of quercetin and its metabolites isorhamnetin and quercetin-3-glucuronide on inflammatory gene expression: role of miR-155. J Nutr Biochem 2011, 22(3), 293–9. [Google Scholar] [CrossRef]
- Taganov, K.D. et al., NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A 2006, 103(33), 12481–6. [Google Scholar] [CrossRef]
- Tili, E. J.J. Michaille, Resveratrol, MicroRNAs, Inflammation, and Cancer. J Nucleic Acids 2011, 102431. [Google Scholar]
- Adriouch, S. et al., Prospective Association between Total and Specific Dietary Polyphenol Intakes and Cardiovascular Disease Risk in the Nutrinet-Sante French Cohort. Nutrients 2018, 10(11). [Google Scholar] [CrossRef]
- Peng, L. et al., Polyphenols extracted from huangjiu have anti-inflammatory activity in lipopolysaccharide stimulated RAW264.7 cells. RSC Advances 2019, 9(10), 5295–5301. [Google Scholar] [CrossRef]
- Katanasaka, Y. et al., Kosen-cha, a Polymerized Catechin-Rich Green Tea, as a Potential Functional Beverage for the Reduction of Body Weight and Cardiovascular Risk Factors: A Pilot Study in Obese Patients. Biol. Pharm. Bull 2020, 43(4), p. 6. [Google Scholar] [CrossRef]
- Jadeski, L. et al., IQGAP1 stimulates proliferation and enhances tumorigenesis of human breast epithelial cells. J Biol Chem 2008, 283(2), 1008–17. [Google Scholar] [CrossRef]
Figure 1.
Pathophysiological mechanisms in the progression of atherosclerosis and cardiovascular events. Various risk factors such as diabetes, dyslipidemia, hypertension, obesity, smoking, aging, physical inactivity, and inflammation contribute to oxidative stress and endothelial dysfunction. This reduces the bioavailability of NO, promoting leucocyte adhesion and progression of atherosclerotic lesions, thrombosis, and cardiovascular events.
Figure 1.
Pathophysiological mechanisms in the progression of atherosclerosis and cardiovascular events. Various risk factors such as diabetes, dyslipidemia, hypertension, obesity, smoking, aging, physical inactivity, and inflammation contribute to oxidative stress and endothelial dysfunction. This reduces the bioavailability of NO, promoting leucocyte adhesion and progression of atherosclerotic lesions, thrombosis, and cardiovascular events.
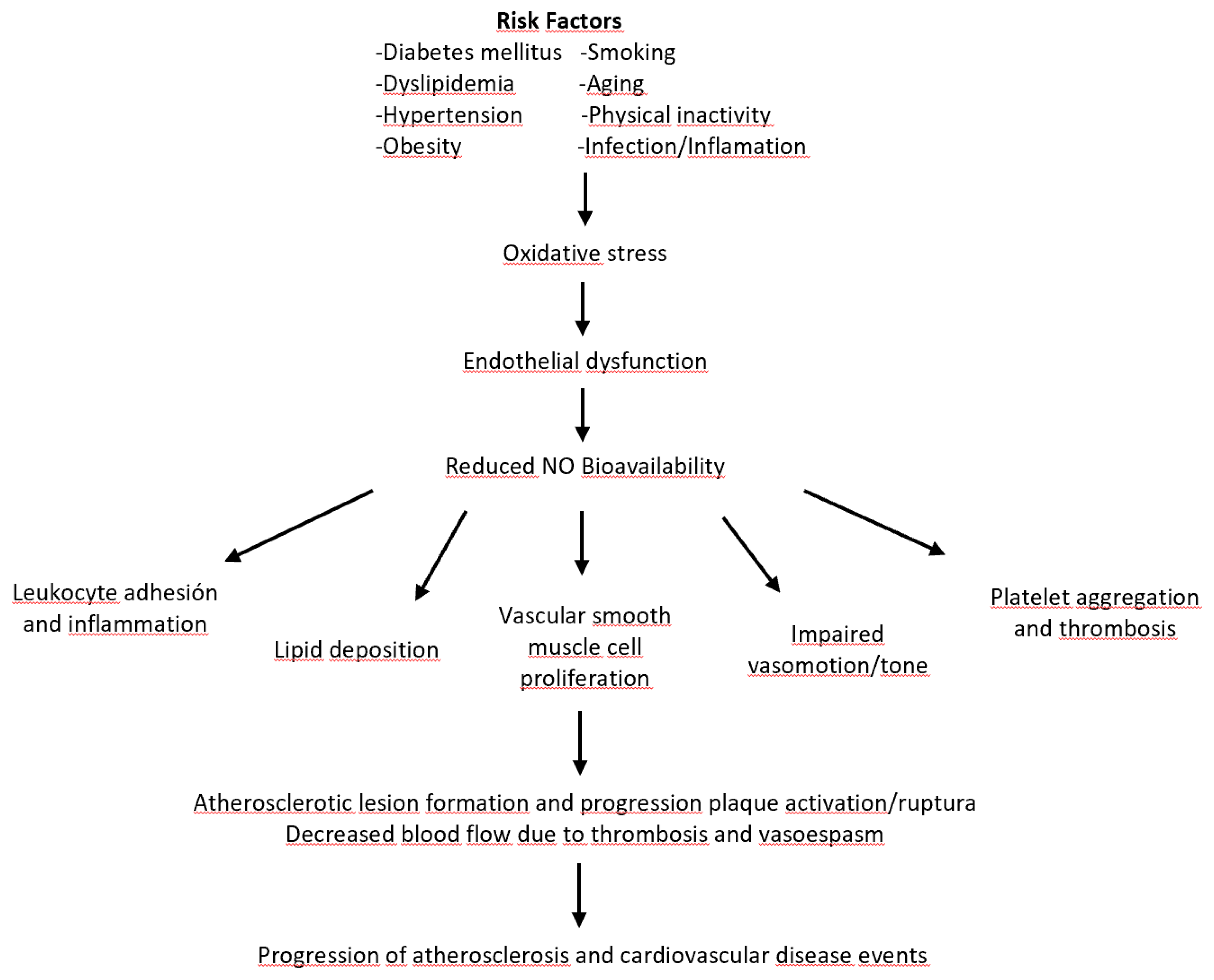
Figure 2.
Typic NO production in endothelial cells. The NO production is activated by increasing Ca2+ levels and the response to endothelin 1. NO subsequently participates in the activation of the SGC to relax or contract the muscles. The green arrows indicate activation or increasing expression levels, and the red lines represent blocking.
Figure 2.
Typic NO production in endothelial cells. The NO production is activated by increasing Ca2+ levels and the response to endothelin 1. NO subsequently participates in the activation of the SGC to relax or contract the muscles. The green arrows indicate activation or increasing expression levels, and the red lines represent blocking.
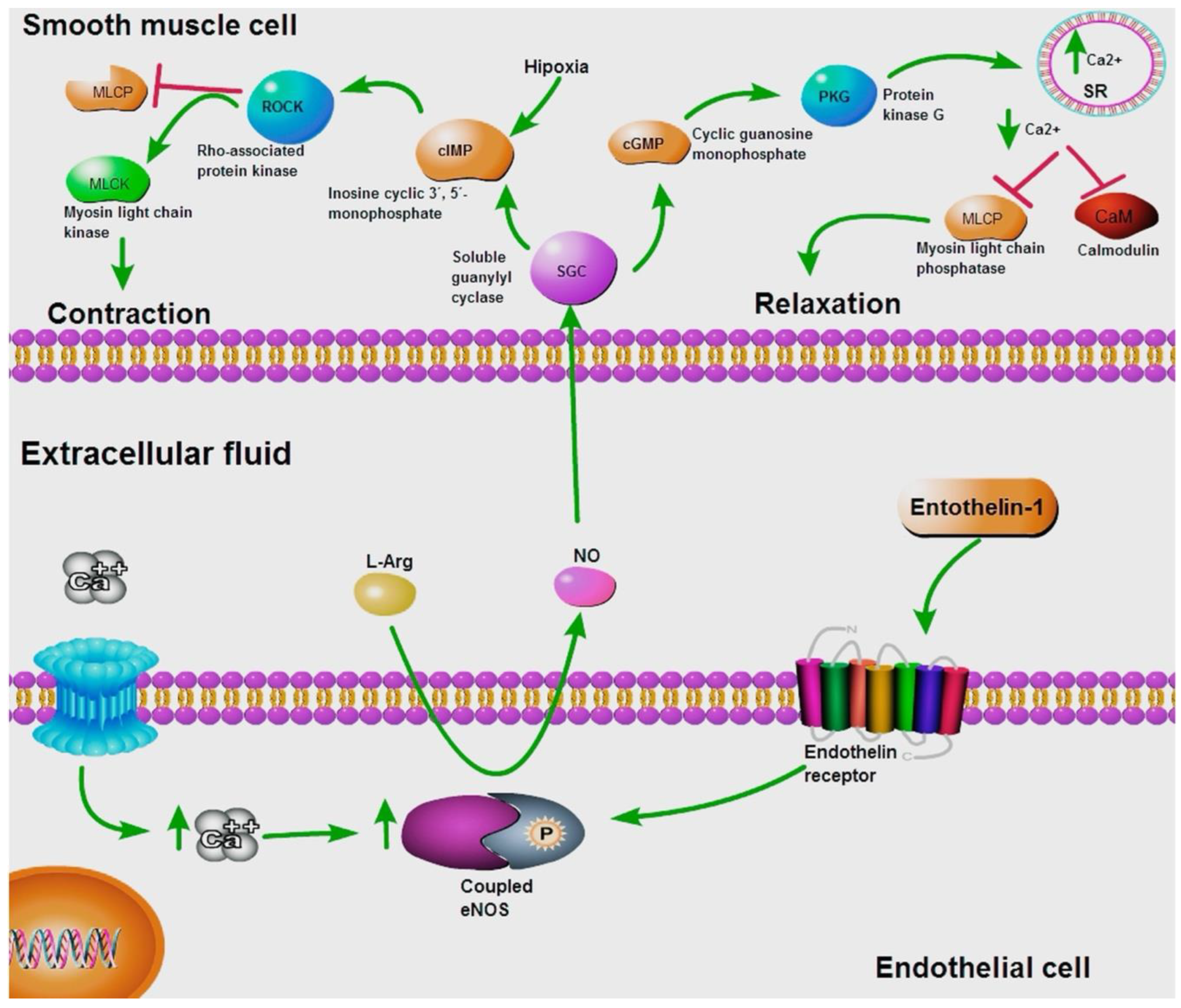
Figure 3.
Polyphenol classification and structure.
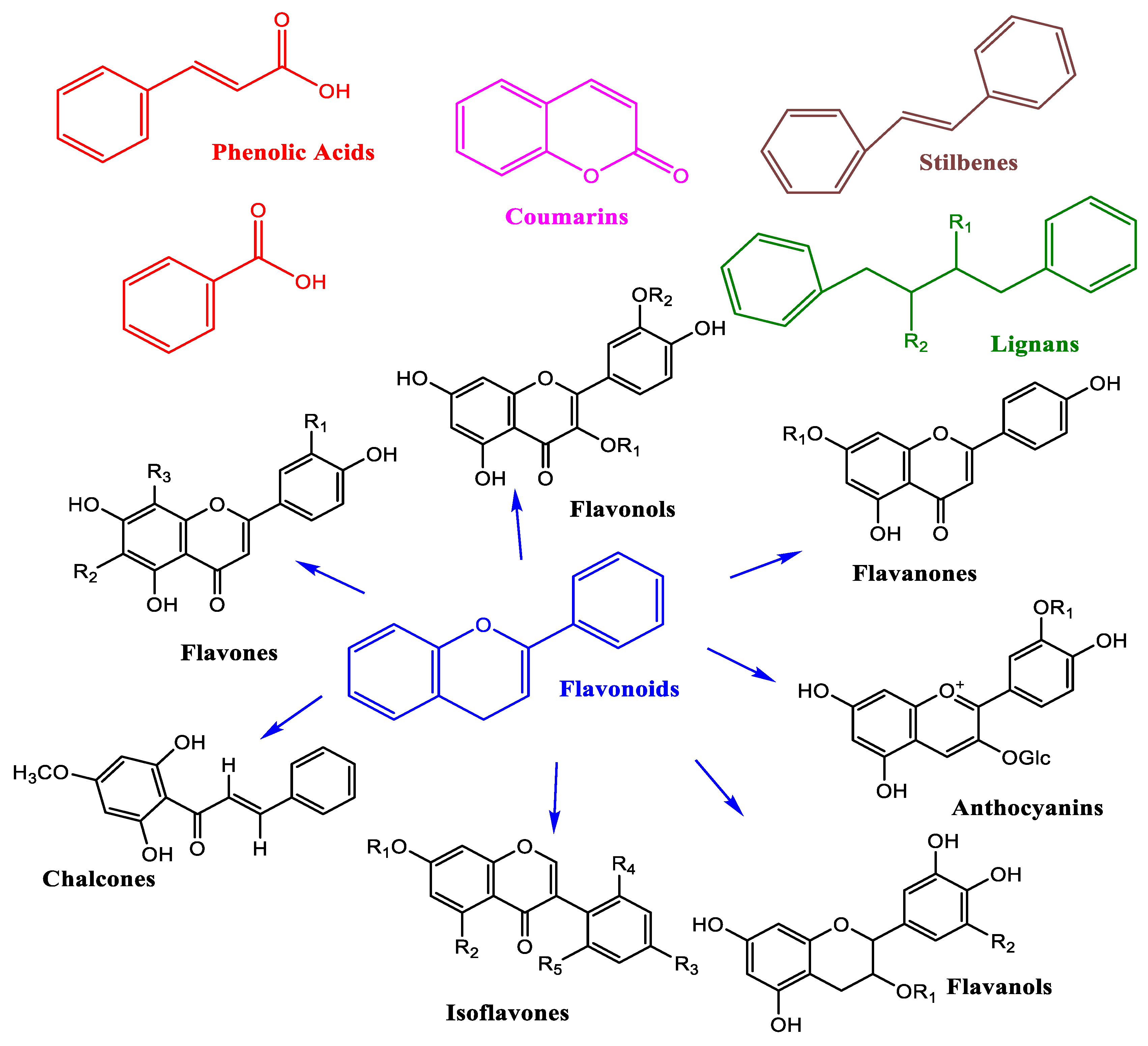
Figure 5.
Activation and Inactivation of platelets. Polyphenols block the receptor P2Y12 and adenylate cyclase cannot convert the ATP onto cAMP. In the absence of cAMP, PKA continues to be inactivated, and VASP keeps dephosphorylated; then, the platelets are inactivated.
Figure 5.
Activation and Inactivation of platelets. Polyphenols block the receptor P2Y12 and adenylate cyclase cannot convert the ATP onto cAMP. In the absence of cAMP, PKA continues to be inactivated, and VASP keeps dephosphorylated; then, the platelets are inactivated.
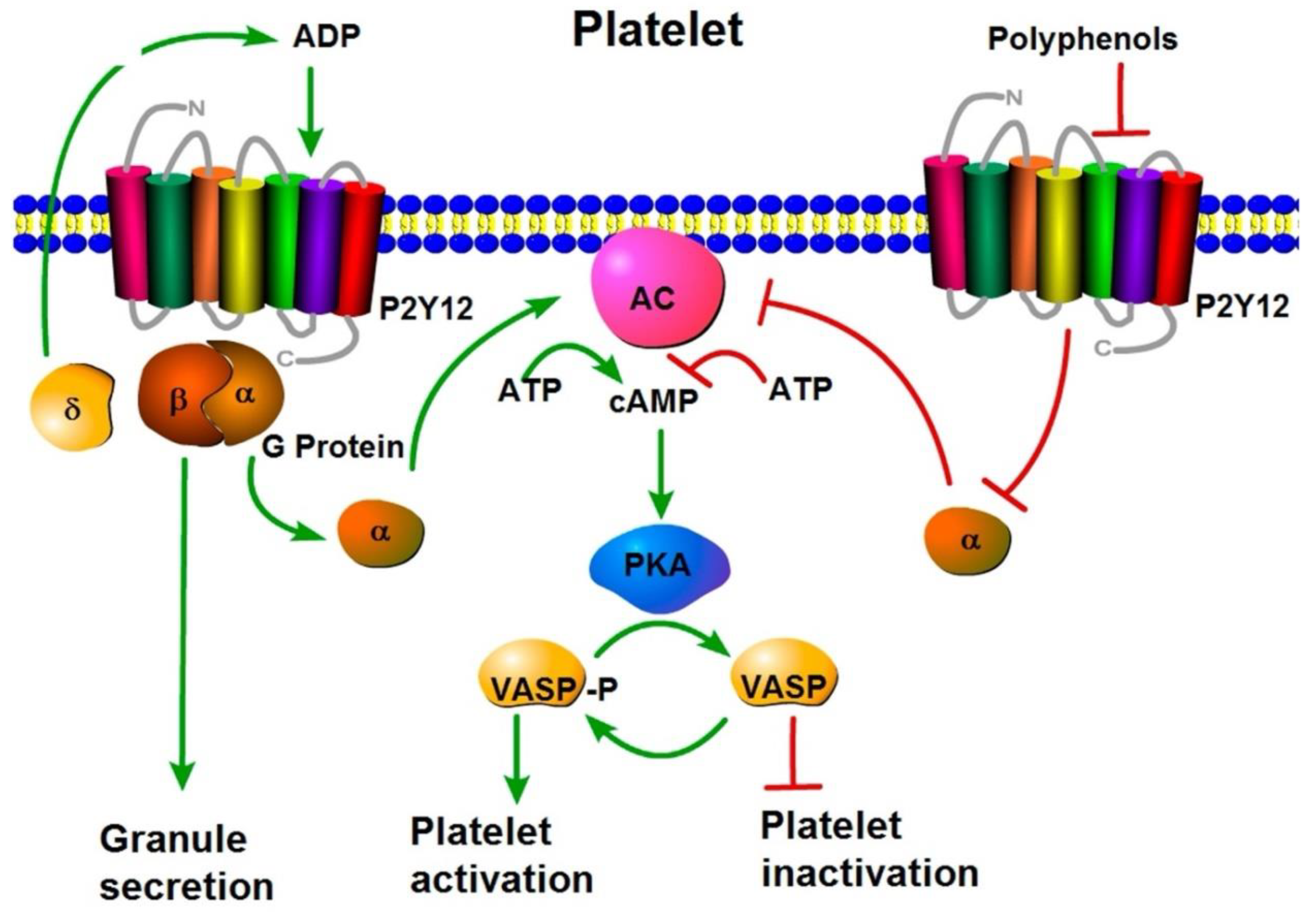
Figure 6.
BEE inhibits the protein SREP and LDLR. The phenolic compounds block the LDLR. Also, in the interior of hepatic cells, BEE down-regulates the SREBP-2 proteins, and endogenous cholesterol is poorly produced. Bee can increase the flow of cholesterol across ABCG5/8 channels.
Figure 6.
BEE inhibits the protein SREP and LDLR. The phenolic compounds block the LDLR. Also, in the interior of hepatic cells, BEE down-regulates the SREBP-2 proteins, and endogenous cholesterol is poorly produced. Bee can increase the flow of cholesterol across ABCG5/8 channels.
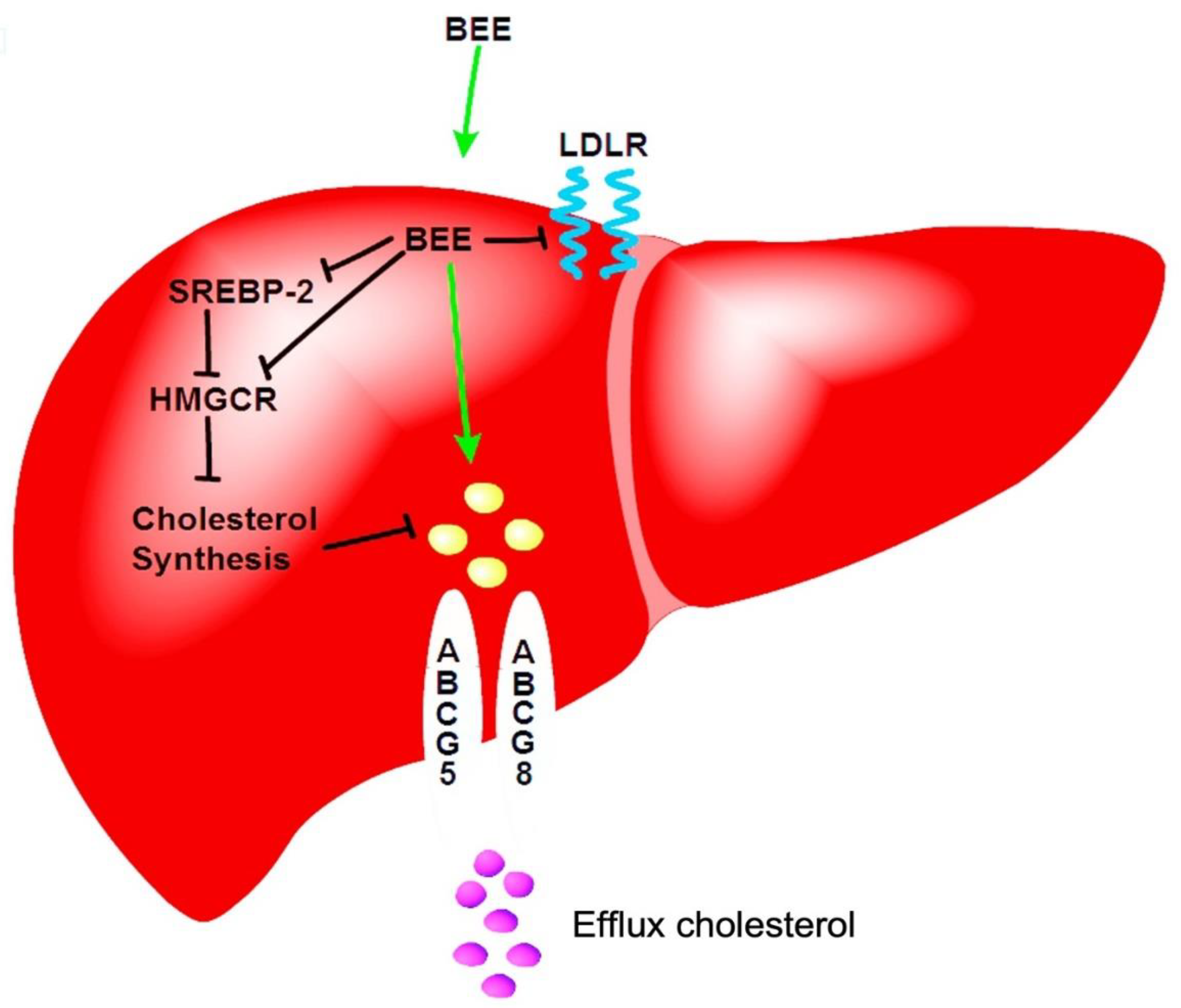

Table 1.
Summary of dietary polyphenols, molecular mechanisms, and cardiovascular effects.
| Polyphenol | Dietary source | Molecular mechanisms | Cardiovascular effects |
| Quercetin | Apples, onions, green tea, grapes, leafy greens | Activates PI3K/Akt, AMPK, SIRT1; inhibits COX/LOX, TNF-α, IL-6 | ↓ Blood pressure ↓ LDL ↑ Insulin sensitivity ↑Anti-inflammatory |
| Resveratrol | Red grapes, red wine, berries | Activates eNOS, SIRT1, Nrf2, AMPK; inhibits NADPH oxidase, Ang II | ↓ Cardiac hypertrophy ↓ Inflammation ↑ Endothelial function |
| Epicatechin |
Cocoa, dark chocolate |
Stimulates NO, GLUT-2, insulin receptor phosphorylation | ↓ Insulin resistance, ↑ Vasodilation ↓ Cholesterol |
| EGCG | Green tea |
Activates PI3K/Akt/eNOS; modulates miRNA expression | ↓ Oxidative stress ↓ Inflammation ↑ Vascular protection |
| Curcumin |
Turmeric root | Inhibits NADPH oxidase, JNK; ↑ NO, ↑ metallothionein expression | ↓ LDL, ↓ Inflammation ↑ Lipid metabolism |
| Anthocyanins |
Strawberries, cherries, blueberries | Modulates pro-atherogenic genes; ↑ antioxidant activity | ↓ P-selectin ↓ Arterial inflammation ↓ CHD/CD risk |
| Catechins | Green tea, Kosen-cha, huangjiu | ↑ NO, ↓ TNF-α, IL-6, IL-1β; inhibits MMP-2/MMP-9 | ↓Atherosclerosis ↓ Hypertension ↑ Endothelial function |
| Proanthocyanidins |
Grape seeds | Inhibits VASP, PI3K/PKB, αIIbβ3 integrins | ↓ Platelet aggregation ↑ Coagulation time |
| Elderberry Polyphenols (BEE) | Black elderberry | ↓ SREBP-2, HMGR, LDLR, NPC1L1; ↑ ABC transporters, SIRT1/2/3 | ↓ Hepatic cholesterol, ↑ Lipid metabolism, ↑ Antioxidant defense |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



