Submitted:
05 January 2026
Posted:
05 January 2026
You are already at the latest version
Abstract
Glioblastoma (GBM) remains among the most treatment-refractory human malignancies, shaped by profound molecular heterogeneity, extensive genomic instability and an immunosuppressive tumor microenvironment. Radiotherapy represents a cornerstone of current management; however, its therapeutic benefit is frequently limited by adaptive resistance and ineffective antitumor immunity. Emerging evidence indicates that ionizing radiation acts not only as a cytotoxic modality, but also as a potent immunological trigger through the release of damage-associated molecular patterns (DAMPs) and the induction of immunogenic cell death (ICD).
In this review, we synthesize recent advances describing how canonical DAMPs—including HMGB1, ATP, calreticulin exposure, mitochondrial and nuclear DNA fragments—coordinate innate and adaptive immune activation via TLR-, RAGE- and cGAS–STING-dependent pathways. We further discuss the dual nature of DAMP signaling, which can either promote durable antitumor immunity or foster chronic inflammation, myeloid reprogramming and tumor tolerance, depending on radiation dose, fractionation, tumor context and concomitant therapies.
Special emphasis is placed on how different radiation qualities, particularly proton versus photon irradiation, differentially modulate DAMP release, ICD dynamics and microenvironmental remodeling. Finally, we highlight translational opportunities to exploit DAMP-related signatures as liquid-biopsy biomarkers of response, as rational selectors for combination strategies (including immunotherapy and radiosensitizers), and as biological guides for personalized and adaptive radiotherapy in GBM.
Collectively, DAMP-centered radiobiology provides a conceptual framework to integrate immunity into radiation planning and may enable a new generation of biologically informed treatment strategies for glioblastoma.
Keywords:
glioblastoma
; radiotherapy
; proton therapy
; damage-associated molecular patterns (DAMPs)
; Immunogenic cell death
; cGAS–STING pathway
; tumor microenvironment
; radioresistance
; immune modulation
; personalized radiotherapy
1. Introduction
Glioblastoma (GBM) remains one of the most aggressive and treatment-refractory human malignancies, with median survival rarely exceeding 15 months despite maximal safe resection, concurrent chemoradiotherapy and adjuvant temozolomide administration. Standard radiotherapy regimens, including photons, provide modest improvements in local control, but tumor recurrence within the irradiation field is almost universal and strongly associated with intrinsic and acquired resistance mechanisms, including glioma stem cell survival, hypoxia and potent immunosuppression in the tumor microenvironment (TME).
Recent advances in radiation oncology have expanded interest beyond purely physical dose delivery to include biological modulation of treatment response. In particular, there is increasing recognition that ionizing radiation can act as a stressor that not only induces DNA lesions but also activates complex immunological pathways through the release of damage-associated molecular patterns (DAMPs). DAMPs are endogenous danger signals—including high-mobility group box 1 (HMGB1), ATP, heat-shock proteins and mitochondrial DNA—that are released during radiation-induced cell damage and immunogenic cell death (ICD) and can engage pattern recognition receptors (PRRs) on innate and adaptive immune cells. [1,2]
This paradigm reflects an important evolution in radiobiology: whereas early models focused primarily on direct DNA damage and clonogenic cell death, modern frameworks encompass immune reprogramming, inflammation and systemic host responses as critical determinants of therapeutic outcome. ICD and associated DAMP signaling can either promote antitumor immunity by fostering dendritic cell maturation and T-cell priming or conversely contribute to chronic inflammation and tumor tolerance depending on context and radiation quality. [3]
In GBM, an inherently “cold” tumor with profound immunosuppressive networks, the interplay between DAMPs, ICD and the TME is of particular relevance. Strategies that harness DAMP-mediated immune activation may enhance the efficacy of radiotherapy and immunotherapy combinations, overcome resistance, and enable personalized approaches that integrate real-time biological response monitoring. Moreover, differential radiation modalities—including proton beam therapy with its distinct ionization patterns—may variably modulate DAMP release and immune outcomes, offering opportunities for theranostic integration. [1,2,3]
In this review, we critically examine the current understanding of DAMP signaling and ICD in the context of GBM radiotherapy, emphasize immunological consequences and translational opportunities, and outline how DAMP-centered biomarkers and modulators can inform nanotheranostic strategies to improve precision treatment.
2. Radiotherapy-Induced DAMP Signaling and Immunogenic Cell Death
2.1. Canonical DAMPs Released After Irradiation
Among the best-characterized DAMPs induced by radiation are calreticulin (CRT) exposure on the cell surface, extracellular ATP, nuclear proteins such as HMGB1, mitochondrial and nuclear DNA fragments, and heat-shock proteins. Each plays a temporally coordinated role.CRT exposure provides an early “eat-me” signal, promoting phagocytosis of dying tumor cells by dendritic cells. ATP released into the extracellular space activates purinergic receptors (e.g., P2RX7), favoring inflammasome assembly and dendritic cell maturation. HMGB1 subsequently binds pattern-recognition receptors including TLR4 and RAGE, supporting efficient cross-presentation of tumor antigens. Meanwhile, cytosolic leakage of nuclear or mitochondrial DNA activates cGAS–STING signaling, resulting in type I interferon production — a central axis for recruiting and priming cytotoxic T lymphocytes [4,5,6].
2.2. ICD as the Immunological Outcome of Radiation-Induced Damage
ICD represents a form of regulated cell death that is inherently pro-immunogenic. Rather than being defined solely by morphology, ICD is characterized by the coordinated presence of DAMP hallmarks (CRT, ATP, HMGB1) and robust type I interferon signaling. Radiotherapy can drive ICD, converting otherwise “silent” death into a vaccine-like event capable of eliciting durable T-cell responses. Importantly, the immunogenicity of ICD is context-dependent — influenced by dose, fractionation, tumor genotype, hypoxia, and the immunological state of the microenvironment.
In glioblastoma, where T-cell infiltration is limited and suppressive myeloid populations dominate, partial ICD induction may paradoxically reinforce immunosuppression if DAMPs promote chronic, non-productive inflammation. Thus, understanding how radiation shapes the quality and kinetics of DAMP release is crucial for rationally combining radiotherapy with immunotherapies [3,5,7].
2.3. Receptor Pathways Linking DAMPs to Immune Activation
Multiple receptor systems translate DAMPs into immune signals. TLR2/4, RAGE, and NLRP3 coordinate early innate responses; cGAS–STING couples cytosolic DNA sensing to interferon-driven cross-priming; and purinergic signaling regulates inflammasome activation and antigen presentation. These pathways converge on dendritic cells, macrophages, and endothelial cells, ultimately influencing T-cell trafficking and effector function. Intriguingly, glioma cells themselves can sense DAMPs, creating feed-forward loops that shape local inflammation and radioresistance.
2.4. Proton Versus Photon Irradiation: Qualitative Differences in DAMP Dynamics
Beyond physical dose distribution, emerging evidence suggests that radiation quality influences immunological consequences. Proton irradiation, characterized by distinct linear energy transfer patterns, may differentially modulate DAMP exposure, mitochondrial stress, and cGAS–STING activation compared with photons. Preliminary studies indicate altered profiles of extracellular ATP, HMGB1, and DNA-containing vesicles following proton exposure, accompanied by differences in bystander signaling and myeloid polarization. These qualitative distinctions support the concept that protons may be harnessed not only to spare normal tissue, but also to shape immune composition as they spare normal tissue from damaging, presenting a hypothesis requiring rigorous translational validation in GBM. In further section we will discuss the hypothesis [8,9,10,11,12]. Figure 1
3. Immune Modulation in Glioblastoma after Radiotherapy: DAMP Signaling in a Highly Suppressive Tumor Microenvironment
Radiotherapy induces profound changes within the glioblastoma microenvironment that extend beyond direct cytotoxicity. Experimental and clinical studies consistently show that ionizing radiation leads to the exposure and release of canonical damage-associated molecular patterns (DAMPs), including calreticulin, extracellular ATP, HMGB1, and fragments of nuclear or mitochondrial DNA. These signals engage pattern-recognition receptors such as TLR4, RAGE, and the cGAS–STING pathway, thereby linking cellular damage to antigen cross-presentation, dendritic cell maturation, and type I interferon production. In glioblastoma, this sequence of events has been documented as a mechanism by which radiotherapy increases antigen visibility and transiently promotes conditions permissive for T-cell priming and trafficking [12,13,14].
At the same time, glioblastoma is characterized by a strongly immunosuppressive tumor microenvironment dominated by tumor-associated macrophages and microglia, regulatory T cells, and limited infiltration of cytotoxic lymphocytes. Multiple analyses demonstrate that radiation-induced inflammation in this setting is rapidly counterbalanced by compensatory mechanisms, including upregulation of immune checkpoints, accumulation of myeloid-derived suppressor populations, and metabolic features such as hypoxia and adenosine production. As a result, although DAMP signalling initiates immunogenic processes, subsequent remodeling frequently favors restoration of tolerance rather than sustained antitumor immunity. [14,15] This pattern reflects a reproducible biological property of glioblastoma rather than a failure of any single pathway.
DAMP signalling therefore functions as an organizing node that integrates radiation-induced cell injury with immune reprogramming. Early exposure of calreticulin and ATP release has been shown to facilitate phagocytosis and inflammasome activation, while HMGB1 and DNA sensing promote interferon-driven recruitment of effector cells. Parallel studies indicate that persistent activation of these same pathways correlates with myeloid polarization toward tissue-repair phenotypes and functional exhaustion of infiltrating lymphocytes. Importantly, these observations have been reported across multiple preclinical glioma models and validated in analyses of patient material, establishing DAMP-driven immune dynamics as a reproducible feature of radiotherapy response in this disease [16,17,18,19].
These immune consequences are closely linked to the structural and metabolic organization of glioblastoma, including hypoxic niches, disrupted vasculature, and high intracellular antioxidant capacity. Such characteristics influence both the magnitude of DAMP release and the downstream composition of infiltrating immune cells. In parallel, radiation-conditioned extracellular vesicles containing proteins, nucleic acids, and DAMP-related molecules have been documented as additional mediators of communication between tumor cells and the surrounding microenvironment. The findings show a biologically consistent framework in which radiotherapy induces a defined cascade of immune-relevant signals that are subsequently shaped by local microenvironmental constraints [19,20].
Because DAMP levels and associated immune signatures reflect the biological effectiveness of radiotherapy, they are being increasingly investigated as measurable indicators of treatment response and as candidates for integration into precision-oriented treatment strategies. This evidence-based perspective provides a logical entry point for translational research aimed at linking molecular monitoring with therapeutic decision-making, particularly in the context of biologically informed radiotherapy approaches.
4. DAMP-Driven Biomarkers and Pathways Relevant to Precision Radiotherapy
Importantly, the evaluation of DAMPs extends beyond single molecules. Composite signatures integrating HMGB1, CRT exposure, and interferon responses have been shown to align with hallmarks of immunogenic cell death across several tumor types, including glioma. In clinical contexts, such signatures have been associated with immune activation and, in selected settings, with improved treatment outcomes when radiotherapy is combined with immunomodulatory approaches. Although glioblastoma remains challenging, the consistency of these biological relationships underscores their relevance for biomarker development rather than isolated experimental phenomena.
Another translational area informed by DAMP biology is treatment monitoring. Serial sampling studies demonstrate that radiation-induced DAMP release occurs dynamically over the course of therapy (hours, days) and can be detected in liquid biopsy formats, including plasma and cerebrospinal fluid. At the same time, imaging techniques capable of capturing inflammation, perfusion changes, or microenvironmental remodeling offer complementary information on downstream immune effects. Integrating molecular and imaging biomarkers therefore provides a realistic pathway toward biologically guided adaptation of therapy, where biological signals reflect treatment effectiveness more directly than anatomical change alone [12,15,21,22,23].
Nanotechnology contributes further precision by enabling controlled delivery and measurement within this biological framework. Several nanotheranostic platforms have been engineered to co-localize therapeutic effects with imaging components, allowing simultaneous intervention and monitoring. In preclinical glioma models, nanoscale radiosensitizers, immunomodulators, and targeted carriers have been used to enhance local radiation effects while permitting in-vivo visualization of distribution and response. When evaluated alongside DAMP-related markers, such systems provide complementary information: DAMPs characterize biological impact, whereas nanotheranostic probes inform localization and kinetics of therapy [23,25].
Radiation modality is another dimension with direct translational relevance. Comparative investigations indicate that distinct radiation qualities may differentially influence DAMP release, mitochondrial stress responses, and downstream immune activation. Studies of proton irradiation, in particular, have reported differences in DNA damage patterning and associated immune signalling relative to photon therapy. [23] These observations do not imply superiority of one modality over another; rather, they indicate that radiation type contributes to the biological signature that can be captured and potentially used to guide clinical decision-making. In this context, the alignment of dosimetry, molecular biomarkers and imaging represents a coherent strategy to move toward biologically informed radiotherapy planning.
In Table 1 we summarized published evidence comparing photon, proton, carbon-ion and BNCT irradiation with respect to the induction of canonical DAMPs. Across modalities, the same core DAMPs (ecto-calreticulin, HMGB1, ATP and extracellular DNA) are consistently observed, with differences mainly reflecting quantitative and kinetic variation rather than modality-specific DAMP species.
DAMP-related signatures and ICD hallmarks provide quantifiable indicators of radiation-induced immune engagement; imaging-enabled nanoplatforms allow localized therapy and simultaneous monitoring; and modality-specific biological effects create opportunities to match treatment delivery with tumor biology. The cumulative evidence indicates that integrating these components is feasible within prospective clinical research, offering a structured pathway toward precision radiotherapy that is grounded in measurable biological parameters rather than theoretical assumptions.
4.1. Do Different Radiation Modalities Induce Different DAMPs?
Across preclinical and early translational studies, photon, proton and high-LET radiations (carbon ions, BNCT) consistently induce the same canonical DAMPs associated with immunogenic cell death (ICD): surface-exposed calreticulin (ecto-CRT), extracellular ATP, HMGB1 release and the presence of nuclear or mitochondrial DNA in the extracellular space. These DAMPs signal through shared receptor systems – CD91 for calreticulin, P2RX7/P2RY2 for ATP, TLR4 and RAGE for HMGB1, and the cGAS–STING pathway for cytosolic DNA – driving the established ICD cascade. Comprehensive reviews by Vaes et al. and Zhu et al. emphasize that these molecules and pathways are common across different radiation regimens, rather than specific to a particular modality [10].
For photons vs. protons, the most direct mechanistic comparison comes from Gameiro et al. In multiple human tumor cell lines, including radioresistant subsets, they showed that proton and photon irradiation at iso-effective doses induced a “common immunogenic modulation signature”: upregulation of MHC class I, ICAM-1, tumor antigens (CEA, MUC1) and calreticulin exposure on the cell surface, which in turn increased susceptibility to antigen-specific T-cell killing [16]. The authors explicitly concluded that tumor cells surviving proton or photon radiation share the same immunogenic phenotype, with no evidence of modality-specific DAMPs [16,20,21,22,23,24,25,26,27,28,29]
Huang et al. extended this observation by directly comparing ecto-CRT induction after photon, proton and carbon-ion irradiation in several tumor cell lines, including U251 glioma. All three modalities increased ecto-CRT in a time- and dose-dependent manner. At iso-doses, proton and photon beams were essentially equivalent in inducing ecto-CRT, whereas carbon ions produced stronger CRT exposure at 2 and 4 Gy but not at higher doses. This confirms that the DAMP species are the same for low-LET photons and protons; high-LET heavy ions change the intensity, not the identity, of DAMP signalling. [27,28,29]
The same pattern holds for HMGB1. Yoshimoto et al. examined HMGB1 release from multiple human cancer cell lines after X-ray versus carbon-ion irradiation and found that both modalities induced HMGB1 release, and when doses were normalised to iso-survival levels, X-rays and carbon ions were equally effective at inducing HMGB1 [14]. Onishi et al. later showed that increasing LET within carbon-ion beams further enhances HMGB1 release, reinforcing that high-LET radiation can quantitatively boost HMGB1 while still utilising the same molecular mediator [29].
For BNCT, Imamichi et al. demonstrated that both BNCT and γ-irradiation trigger extracellular HMGB1 release in vitro, but BNCT produce a more pronounced and sustained HMGB1 elevation in mouse plasma, peaking around day 3 after treatment. Again, the DAMP is HMGB1 in both conditions; BNCT modifies the kinetics and amplitude, not the DAMP repertoire. [13]. The data support a conclusion: photon, proton, carbon-ion and BNCT all induce the same canonical DAMPs associated with ICD. High-LET modalities can enhance the magnitude and timing of DAMP release, but no modality-specific DAMP species have been consistently identified. Differences are primarily quantitative and kinetic, not qualitative.
4.2. How can DAMPs Influence Radiotherapy Outcomes – and How Does RT Choice Interact with This Biology?
DAMPs influence radiotherapy outcomes by shaping the balance between effective antitumor immunity and tolerogenic or chronically inflamed microenvironments.
Mechanistically, the sequence is well defined. Early after irradiation, calreticulin translocates to the cell surface, acting as an “eat-me” signal that facilitates phagocytosis of dying tumor cells by dendritic cells via CD91. Extracellular ATP is released and engages P2RX7, promoting NLRP3 inflammasome activation and IL-1β secretion, which supports dendritic cell maturation. Subsequently, HMGB1 is released and binds TLR4 and RAGE on antigen-presenting cells, enhancing antigen processing and cross-presentation. In parallel, leakage of nuclear and mitochondrial DNA into the cytosol or extracellular space activates cGAS–STING, leading to type I interferon production and a transcriptional programme that recruits and primes CD8+ T cells [9].
Vaes et al. systematically reviewed clinical data where radiotherapy-induced ICD markers were assessed in patients and found that elevated HMGB1, ecto-CRT and activation of IFN-I–related signatures correlate with features of immune activation and, in selected settings, with improved outcomes in combination with immunotherapies [9]. Zhu et al. summarised similar relationships across radiotherapy trials where ICD hallmarks were linked to enhanced infiltration of effector T cells and better responses to immune checkpoint blockade, particularly in tumors historically considered “cold” [10].
Another clinical example is provided by Clasen et al., who performed weekly plasma sampling in head-and-neck squamous cell carcinoma patients receiving definitive radiochemotherapy with photons. Plasma HMGB1, measured by ELISA, increased dynamically over the course of treatment in most patients. Importantly, specific HMGB1 trajectories correlated with oncologic outcome: patients with a sustained increase in HMGB1 during radiochemotherapy had more favourable local control, whereas decreasing or flat profiles were associated with higher progression risk. HMGB1 dynamics also correlated with tumor volume, CRP and treatment-related toxicity [27]. This study shows that DAMP levels are not merely a byproduct of damage but carry prognostic information relevant to treatment success.
In preclinical models, high-LET modalities (carbon ions, BNCT) amplify these same DAMP-mediated mechanisms. Yoshimoto and Onishi demonstrated that carbon ions induce HMGB1 release at levels comparable or higher than X-rays at iso-survival doses and can more strongly enhance downstream immune responses in combination with immunotherapy [14,29]. Authors showed that BNCT produces an early and sustained HMGB1 plasma peak, which they propose as a biomarker of BNCT effectiveness. High-LET radiations magnify the same DAMP-driven immunological cascade and can, under controlled conditions, translate into superior antitumor immunity – but the underlying DAMP biology remains shared.
For proton versus photon radiotherapy, the key message from Gameiro and Huang is that at the tumor-cell level, DAMP-related immunogenic modulation is very similar: both modalities induce ecto-CRT, upregulate antigen presentation machinery and enhance susceptibility to CTL killing. The clinically relevant difference arises more at the systemic immune level than at the level of DAMP species per se. Multiple clinical and dosimetric studies (in glioblastoma and other tumors) have demonstrated that proton plans substantially reduce low-dose exposure of circulating blood and lymphoid organs compared to photons, resulting in lower rates of severe radiation-induced lymphopenia. Because DAMP-driven ICD requires functional dendritic cells and T-cell populations to translate into effective immunity, preservation of the host immune compartment is likely to enhance the clinical impact of a given DAMP/ICD signal.
Thus, from a translational standpoint:
- Different RT types do not fundamentally change which DAMPs are generated,
- high-LET beams (C-ions, BNCT) can increase how much and how fast they are released,
- protons may improve the conditions in which DAMP-induced immunity operates, by sparing blood and immune organs, even if the local DAMP pattern is similar to photons.
This implies that the choice of RT modality can influence outcome indirectly via DAMP biology: by modulating the intensity of ICD (high-LET) or preserving the immune system’s ability to respond (protons), rather than by switching to entirely different DAMP pathways.
4.3. Can DAMPs be Markers of Radiotherapy Success?
The accumulated evidence strongly supports the idea that DAMPs and ICD signatures are promising biomarkers of radiotherapy effectiveness, although they are not yet fully validated predictive or surrogate markers in glioblastoma.
In the clinical setting, Vaes et al. summarised multiple studies where ICD markers were measured during or after radiotherapy in different tumor types. HMGB1 increases after CRT for oesophageal, rectal and head-and-neck cancers; ecto-CRT in tumor biopsies correlates with dendritic cell infiltration and favourable prognosis in cervical cancer; and type I interferon–related gene signatures have been associated with better response to RT plus immunotherapy in several solid tumors. Clasen’s weekly HMGB1 study is a particularly clear demonstration that serial DAMP measurements during RT carry prognostic information and can be used to stratify patients by risk, fulfilling key criteria for biomarkers of treatment success.
Preclinical data provide mechanistic support: blocking HMGB1, CRT or ATP signalling attenuates the immune contribution to tumor control after RT, while enhancing these pathways (e.g. via STING agonists) augments T-cell responses and abscopal effects. These interventions modulate radiotherapy outcome in a manner tightly coupled to DAMP biology, strengthening the rationale to use DAMP signatures as biomarkers and potential therapeutic targets.
In glioblastoma specifically, systematic liquid-biopsy studies have shown that circulating DNA, exosomes and immune-related proteins can be monitored over time and correlate with disease burden and treatment response, even though DAMP-specific panels (HMGB1, ecto-CRT, mtDNA) are only beginning to be explored [21]. The analytical tools – ELISA, qPCR/digital PCR, EV proteomics and, increasingly, DAMP-sensitive imaging probes – already exist and have been proven feasible in other tumor types.
DAMP-related markers fulfil the criteria of “promising biomarkers” of radiotherapy success: they can be repeatedly measured, show dynamic changes over the course of treatment, and correlate with immune activation and clinical outcome in several tumor entities. However, they have not yet reached the status of fully validated predictive or surrogate endpoints in glioblastoma, and their integration into clinical decision-making will require prospective, biomarker-driven trials.
In this framework, our own experience – that proton radiotherapy appears to produce a qualitatively different immune context than photon RT – fits very well with the available data: the DAMP biology at the tumor level is shared; the systemic environment in which these signals act is improved by proton plans that better preserve the immune system.
4.4. When DAMPs Promote Tolerance Rather than Immunity
DAMP signaling is fundamentally dualistic. Although acute, well-orchestrated release of calreticulin, ATP, HMGB1, and extracellular DNA promotes ICD, persistent or dysregulated DAMP release generates a biological milieu that paradoxically sustains tumor survival and treatment resistance. In the context of chronic tissue injury, repeated fractions of radiotherapy and persistent tumor necrosis, DAMP-driven activation of pattern-recognition receptors (PRRs)—including TLR4, RAGE, and cGAS–STING—can shift from initiating protective immunity toward reinforcing chronic, tumor-supportive inflammation [1,2,3].
This tolerogenic reprogramming proceeds through several, partly overlapping mechanisms. First, chronic HMGB1 exposure enhances NF-κB signaling, angiogenesis, DNA repair capacity and autophagy, ultimately conferring radioresistance at the tumor-cell level [4,5,6,7]. Experimental models demonstrate that blocking extracellular HMGB1 reverses myeloid recruitment and restores radiosensitivity after irradiation, highlighting HMGB1 as a central node of therapy resistance [8,9,10].
Second, ATP released as an early DAMP is rapidly catabolized by CD39 and CD73, generating adenosine, a potent immunosuppressive metabolite. Adenosine signaling via A2A and A2B receptors limits cytotoxic T- and NK-cell function, promotes regulatory T cells, and fosters vascular remodeling—thereby attenuating radiotherapy-induced antitumor immunity and reducing responses to immune checkpoint inhibitors. Therapeutic targeting of the CD39–CD73–A2AR axis restores sensitivity to radiotherapy in multiple preclinical models, illustrating how a DAMP that initially promotes ICD can evolve into a dominant suppressive pathway when chronically present [11,12,13].
Third, persistent DAMP signaling drives the accumulation and functional polarization of myeloid-derived suppressor cells (MDSC) and tumor-associated macrophages (TAM), which inhibit antigen presentation, suppress T-cell infiltration, and upregulate immune checkpoints such as PD-L1. This phenomenon has been extensively documented across tumor types, where DAMP-dependent sterile inflammation correlates with immune escape, metastatic dissemination, and therapeutic failure [14,15,16]. Recent genomic signatures enriched for DAMP-related genes further link these pathways with immunosuppressive tumor microenvironments and inferior clinical outcomes [17].
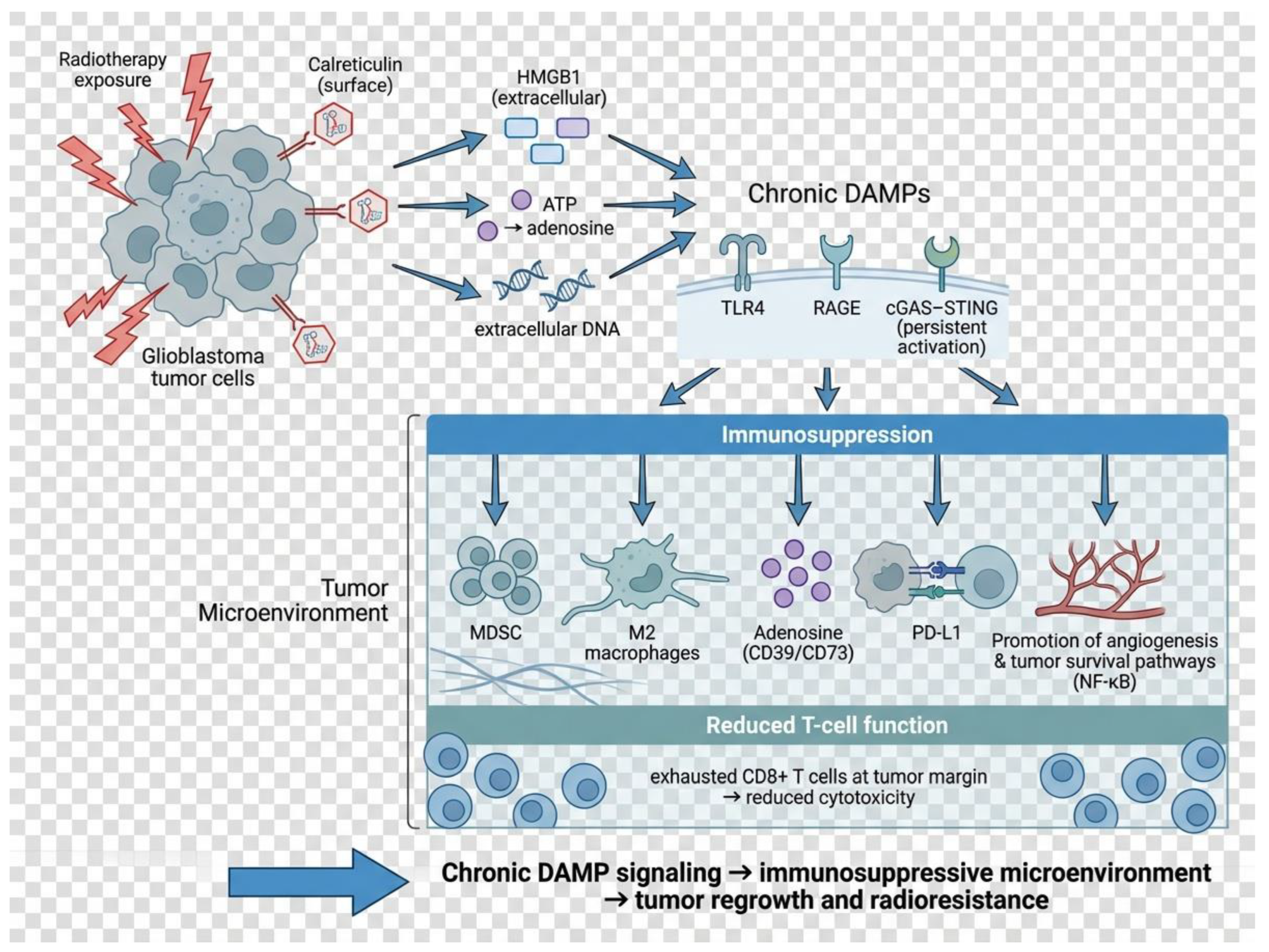
Figure 2.
DAMP-guided nanotheranostics enhance radiation-induced antitumor immunity. Radiotherapy induces immunogenic cell death and release of DAMPs (calreticulin, HMGB1, ATP, extracellular DNA). Nanotheranostic platforms sense radiation-induced stress, deliver radiosensitizers or immunomodulatory payloads, and provide imaging feedback (PET/MRI/fluorescence). DAMP signaling activates dendritic cells and macrophages via TLR4, RAGE and cGAS–STING, leading to T-cell priming and tumor infiltration. Integration of molecular monitoring with imaging enables adaptive, biology-guided radiotherapy.
Figure 2.
DAMP-guided nanotheranostics enhance radiation-induced antitumor immunity. Radiotherapy induces immunogenic cell death and release of DAMPs (calreticulin, HMGB1, ATP, extracellular DNA). Nanotheranostic platforms sense radiation-induced stress, deliver radiosensitizers or immunomodulatory payloads, and provide imaging feedback (PET/MRI/fluorescence). DAMP signaling activates dendritic cells and macrophages via TLR4, RAGE and cGAS–STING, leading to T-cell priming and tumor infiltration. Integration of molecular monitoring with imaging enables adaptive, biology-guided radiotherapy.
Glioblastoma exemplifies this biology. Necrotic tumor cores continuously release DAMPs—including HMGB1, heat-shock proteins, and other TLR ligands—driving polarization of glioma-associated macrophages toward an M2-like phenotype, suppression of MHC expression, and induction of checkpoint molecules. These effects create a TME that is highly resistant to radiotherapy and poorly responsive to immunotherapy, despite the presence of abundant inflammatory signals [18,19,20]. Thus, in GBM, DAMP signaling frequently becomes tolerogenic rather than immunogenic, converting radiation-induced injury into a chronic, self-reinforcing program of immune suppression.
The published data argue that ICD should not be conceptualized as a binary switch but rather as a continuum. At one end, coordinated, transient DAMP release promotes effective dendritic-cell activation and cytotoxic T-cell recruitment; at the other, sustained or excessive DAMP signaling drives myeloid suppression, checkpoint induction, vascular remodeling, and radiotherapy resistance. Understanding this spectrum—and the conditions that shift DAMP signaling from immunity to tolerance—is essential for designing biomarker-guided strategies and rational combinations capable of converting radiation-induced damage into durable antitumor immunity.
5. Measuring DAMPs: From Biological Readouts to Actionable Nanotheranostics
The translational relevance of DAMP biology depends on whether these signals can be reliably quantified, monitored longitudinally, and integrated with therapeutic decision-making. Over the past decade, multiple platforms have matured to the point where DAMP assessment is technically feasible and increasingly suitable for clinical research. Rather than focusing on methodological detail, the key question is how each compartment informs treatment biology and how such information can be linked to image-guided and nanotechnology-enabled interventions.
5.1. Blood-Based Assays
Plasma HMGB1, circulating cfDNA/mtDNA, ecto-calreticulin–related signatures and interferon-stimulated gene panels can be obtained repeatedly during therapy. Serial sampling studies in solid tumors demonstrate that these markers change dynamically over the course of radiotherapy and correlate with tumor burden, inflammation, and, in some settings, outcome. Their minimally invasive nature makes them attractive for biologically guided treatment adaptation, particularly when combined with other systemic biomarkers.
5.2. Cerebrospinal Fluid
For brain tumors, CSF provides a compartment that is biologically closer to the tumor microenvironment than peripheral blood. DAMP-related nucleic acids and proteins detected in CSF show greater specificity for intracranial pathology and may capture microenvironmental remodeling that is not visible on structural imaging. Although still underexplored for DAMP biology specifically, CSF profiling is poised to become a high-value matrix for response monitoring in glioblastoma.
5.3. Extracellular Vesicles (EVs)
EVs package DAMPs, danger-associated peptides, nucleic acids, and immunomodulatory mediators released after irradiation. Their stability and ability to reflect tumor–immune crosstalk make EV-associated DAMPs promising candidates for liquid-biopsy theranostics, potentially linking biological activity with both therapeutic delivery and monitoring strategies.
5.4. Imaging Correlates
Advanced MRI and PET tracers can visualize inflammation, perfusion, hypoxia, and innate immune activation—processes closely intertwined with DAMP signaling. The development of probes targeting ICD-associated pathways (e.g., STING activation or macrophage/microglial dynamics) creates opportunities to co-register molecular DAMP activity with functional imaging, moving toward biologically informed, image-adaptive radiotherapy.
These approaches transform DAMPs from purely mechanistic phenomena into measurable, clinically interpretable signals. Importantly, they provide the informational backbone for nanotheranostic strategies: nanosystems can be engineered to respond to DAMP-associated cues, co-deliver radiosensitizers or immunomodulators, and simultaneously report biological effects through imaging. In this emerging paradigm, radiotherapy induces damage; DAMPs quantify the biological consequence; and nanotheranostic platforms close the loop by enabling tailored intervention and monitoring. This integration offers a realistic pathway to truly personalized, DAMP-guided radiotherapy in glioblastoma and head-and-neck cancers.
6. From Measurement to Nanotheranostic Design
The ability to quantify DAMPs in blood, cerebrospinal fluid, extracellular vesicles, and through molecular imaging provides more than descriptive biological information; it offers actionable guidance for therapeutic development. Patterns of HMGB1 release, calreticulin exposure, cfDNA dynamics, and interferon-related signatures collectively define whether radiotherapy is inducing immunogenic cell death or generating a more tolerogenic inflammatory state. When these circulating and imaging-based readouts are integrated longitudinally, they create a biologically anchored framework capable of informing intervention timing, dose scheduling, and the rational selection of combination strategies.
Within this context, nanotheranostic platforms can be designed not only as carriers of therapeutic payloads but as responsive systems engineered around measurable biological cues. Tumor hypoxia, redox status, and inflammation — all processes linked to DAMP signalling — are already used to trigger the release of radiosensitizers, immunomodulatory compounds, or DNA-damage amplifiers in preclinical glioma models. Incorporating imaging components into these platforms enables simultaneous visualization of nanoparticle distribution and correlation with DAMP-derived biomarkers, thereby linking delivery, biological effect, and outcome within the same system.
Such alignment between measurable DAMP biology and bioresponsive nanosystems establishes a closed diagnostic–therapeutic loop: radiotherapy initiates damage signalling; DAMP assays and molecular imaging quantify the resulting immune and microenvironmental response; and nanotheranostic agents are deployed or adjusted according to these signals. This iterative, data-driven model represents a realistic pathway by which biological monitoring can be translated into precision radiotherapy strategies, particularly in tumors like glioblastoma where conventional imaging alone provides limited insight into treatment biology.
7. Conclusions
DAMPs released during radiotherapy represent a crucial biological interface between tumor cell damage and the immune system. Radiation-induced immunogenic cell death is characterized by the exposure and release of molecules such as calreticulin, HMGB1, ATP, and extracellular DNA, which can activate dendritic cells and trigger adaptive immune responses through pattern-recognition receptors and type I interferon pathways [1,2,3]. These responses underpin the emerging understanding of RT as not merely a cytotoxic modality, but as a therapy that modifies the tumor microenvironment and systemic immunity, thereby potentially enhancing antitumor effects beyond direct DNA damage.
A major translational implication of DAMP biology is their potential role as biomarkers of treatment response. Serial measurement of DAMPs and downstream signals could provide real-time insight into whether RT is eliciting effective ICD within an individual patient. For example, studies have identified HMGB1, calreticulin exposure and interferon signatures as correlates of immunogenic responses in irradiated tumors, suggesting that such markers might predict which patients mount meaningful antitumor immunity following RT [1,4]. Because these signals may change dynamically over the course of treatment, monitoring DAMPs could inform adaptive strategies, such as interplay with immunotherapy or modification of dose/fractionation.
Moreover, DAMP levels might reflect intrinsic tumor radiosensitivity. Tumors that fail to generate robust DAMP signals after irradiation may be less likely to engage immune effector mechanisms, contributing to resistance. While genomic radiosensitivity indices such as RSI and GARD integrate gene expression patterns to predict intrinsic radiosensitivity [5], DAMP profiling offers a complementary, functionally grounded approach that directly measures the immunogenic consequences of irradiation at the cellular level.
Critically, the impact of DAMPs is context dependent. Excessive or persistent DAMP release can drive chronic inflammation and immune suppression, recruiting myeloid-derived suppressor cells and regulatory networks that blunt antitumor immunity and promote tumor regrowth [6]. This dualism underscores the need not only to measure DAMPs, but to interpret their temporal patterns and biological context.
From a clinical personalization standpoint, integrating DAMP biomarkers into RT planning could enable stratification of patients according to their baseline and early treatment immunobiology. For example, a TME with pre-existing immunogenic signaling might benefit from conventional dose schedules combined with immune checkpoint inhibitors, whereas a poorly immunogenic TME might require strategies to enhance ICD, such as high-LET particles or immune modulators. Similarly, DAMP signatures could inform decisions about combining RT with targeted agents or nanomedicines designed to modify the TME.
Our insights suggest that DAMPs constitute a future-oriented complement to existing RT biomarkers, providing both mechanistic depth and temporal resolution. By capturing the biological response to radiation, rather than simply the physical dose delivered, DAMP-based biomarkers could help bridge the gap between radiobiology and immunology, enabling more nuanced and personalized RT regimens tailored to individual patients and tumor phenotypes.
References
- Hernández C, Huebener P, Schwabe RF. Damage-associated molecular patterns in cancer: a double-edged sword. Oncogene. 2016;35:5931–5941. [CrossRef]
- Lin H et al. Damage-associated molecular patterns (DAMPs) in diseases. Signal Transduct Target Ther. 2025 (ahead of print).
- Chen F et al. DAMPs in immunosenescence and cancer. Semin Immunol. 2024;101813. [CrossRef]
- Liao Y et al. HMGB1 in radiotherapy: a two-headed signal regulating tumor radiosensitivity and immunity. Onco Targets Ther. 2020;13:6859–6871. [CrossRef]
- Alhasan BA et al. HMGB1: A central node in cancer therapy resistance. Int J Mol Sci. 2025;26(24):12010. [CrossRef]
- Ayoub M et al. Immune-mediated role of extracellular HMGB1 in a bladder cancer model. Sci Rep. 2019;9:16876. [CrossRef]
- Dong J et al. HMGB1 overexpression promotes malignant phenotype and radioresistance in ESCC. J Cancer. 2022;13:2717–2726. [CrossRef]
- Lv G et al. Multiple functions of HMGB1 in cancer. Front Oncol. 2024;14:1384109. [CrossRef]
- Vaes RDW et al. Biomarkers of radiotherapy-induced immunogenic cell death. Cancers (Basel). 2021;13(8):1781. [CrossRef]
- Zhu H et al. Immunogenic cell death induction by ionizing radiation. Front Immunol. 2021;12:705361. [CrossRef]
- Fucikova J et al. Detection of immunogenic cell death and its relevance for cancer therapy. Cell Death Dis. 2020;11:1013. [CrossRef]
- Han Y et al. Clinical application of immunogenic cell death inducers in cancer immunotherapy. Front Cell Dev Biol. 2024;12:1363121. [CrossRef]
- Imamichi S et al. Extracellular release of HMGB1 as an early biomarker in BNCT. Biology (Basel). 2022;11:420. [CrossRef]
- Yoshimoto Y et al. Carbon-ion beams induce production of HMGB1 comparable with X-rays. J Radiat Res. 2015;56:509–514. [CrossRef]
- Onishi M et al. High LET carbon-ion irradiation increases HMGB1 release. Oncotarget. 2018;9:24532–24543.
- Gameiro SR et al. Shared immunogenic modulation after proton and photon radiation. Int J Radiat Oncol Biol Phys. 2016;95:120–130. [CrossRef]
- Huang Y et al. Photon, proton and carbon-ion effects on ecto-calreticulin exposure. Ann Transl Med. 2019;7(20):542. [CrossRef]
- de Leve S et al. Targeting CD73/adenosine pathway to improve RT. Cancers (Basel). 2019;11:1573. [CrossRef]
- Leone RD, Emens LA. Targeting adenosine for cancer immunotherapy. J Immunother Cancer. 2018;6:57. [CrossRef]
- Xia C et al. CD39/CD73/A2AR pathway and cancer immunotherapy. Mol Cancer. 2023;22:72. [CrossRef]
- Shalapour S, Karin M. Immunity, inflammation, and cancer. J Clin Invest. 2015;125:3347–3355. [CrossRef]
- Li K et al. Myeloid-derived suppressor cells as therapeutic targets. Signal Transduct Target Ther. 2021;6:362. [CrossRef]
- Ostrand-Rosenberg S, Fenselau C. Myeloid-derived suppressor cells in cancer. J Immunol. 2018;200:422–431. [CrossRef]
- Wu Q et al. DAMP prognostic signature predicts TME and outcome. Front Immunol. 2025;16:1516457.
- Lin YJ et al. Myeloid cells in glioblastoma immunosuppression. Front Immunol. 2022;13:887781. [CrossRef]
- Chen J et al. Tumor-associated macrophages in glioblastoma. Cells. 2025;14:1458. [CrossRef]
- Clasen K et al. Dynamics of HMGB1 during chemoradiotherapy in head and neck cancer. Strahlenther Onkol. 2022;198:194–200. [CrossRef]
- De Martino M et al. Exploiting radiation immunostimulatory effects in glioblastoma. Neuro Oncol. 2023;25:433–447.
- Onishi M, Okonogi N, Oike T, et al. High linear energy transfer carbon-ion irradiation increases the release of the immune mediator high mobility group box 1 from human cancer cells. J Radiat Res. 2018;59(4):541–546.
Figure 1.
Chronic DAMP signaling promotes an immunosuppressive tumor microenvironment. Radiotherapy causes glioblastoma cells to release DAMPs (calreticulin, HMGB1, ATP/adenosine, extracellular DNA). When signaling becomes persistent, activation of TLR4, RAGE and cGAS–STING drives recruitment of MDSCs, polarization of M2-like macrophages, accumulation of adenosine (CD39/CD73) and up-regulation of PD-L1. These processes suppress cytotoxic T-cell activity, promote angiogenesis and survival signaling, and ultimately contribute to tumor regrowth and radioresistance.
Figure 1.
Chronic DAMP signaling promotes an immunosuppressive tumor microenvironment. Radiotherapy causes glioblastoma cells to release DAMPs (calreticulin, HMGB1, ATP/adenosine, extracellular DNA). When signaling becomes persistent, activation of TLR4, RAGE and cGAS–STING drives recruitment of MDSCs, polarization of M2-like macrophages, accumulation of adenosine (CD39/CD73) and up-regulation of PD-L1. These processes suppress cytotoxic T-cell activity, promote angiogenesis and survival signaling, and ultimately contribute to tumor regrowth and radioresistance.
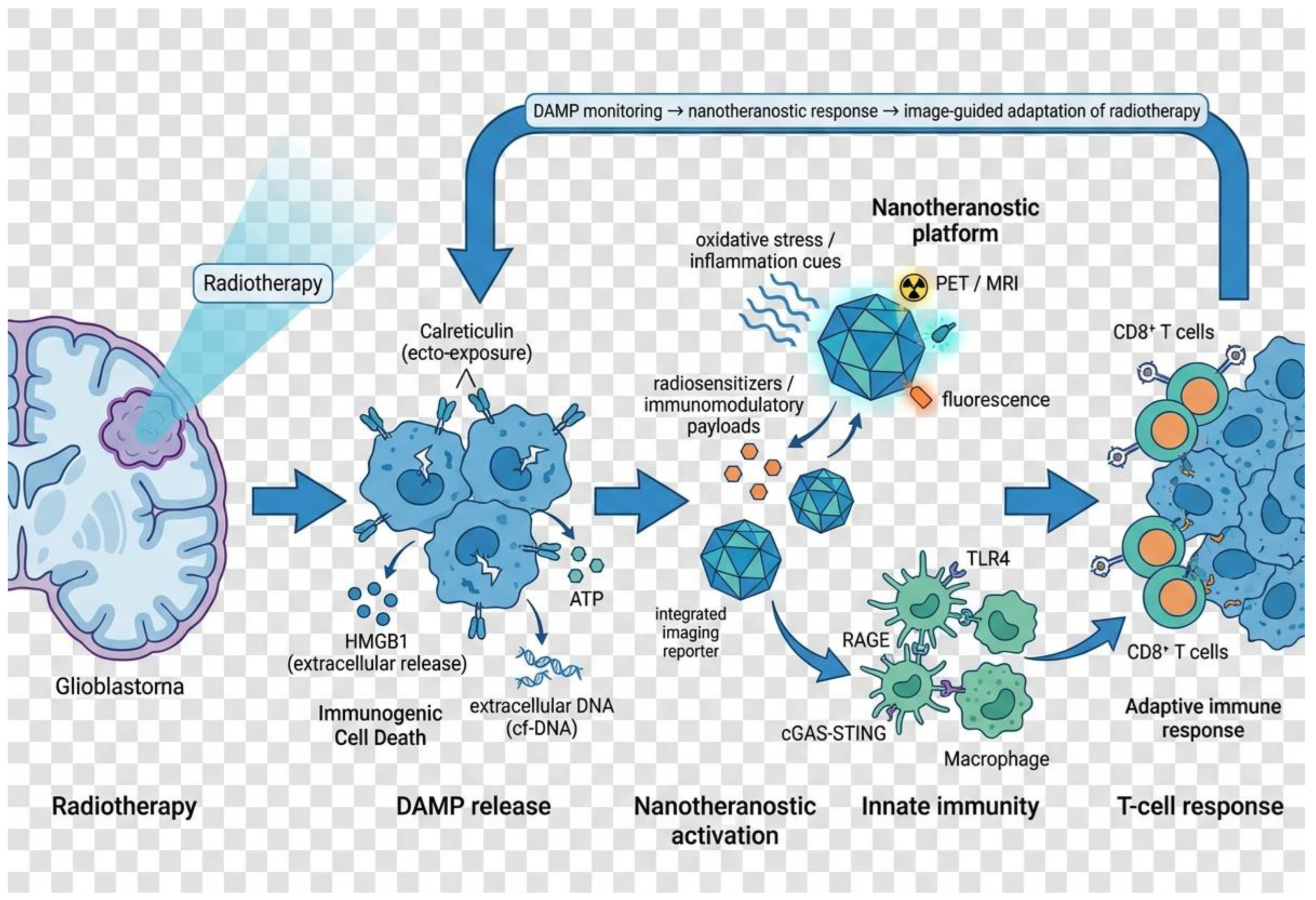

Table 1.
Overview of radiation modalities and their reported DAMP responses.
| Radiation modality | Main DAMP(s) reported | Timing of detection | Model/clinical setting | Key findings | Evidence for modality-specific differences in DAMPs |
| Photons (clinical IMRT + CRT) | HMGB1 (plasma) | Weekly during RT; post-treatment follow-up | HNSCC patients receiving definitive radiochemotherapy | Serial plasma sampling showed dynamic rise of HMGB1 over the course of RT, with patterns associated with outcome and toxicity. | No qualitative difference in DAMP species vs. other modalities; clinical data only for photons. Quantitative behaviour reflects tumor burden, inflammation and toxicity, not unique photon biology. [1] |
| Photons (fractionated, preclinical) | HMGB1, HSP70 and other ICD-associated DAMPs | ~24–72 h after irradiation; repeated pulses with fractionation | Glioma and other tumor cell lines; small-animal models | Fractionated photon RT induces pulse-like release of HMGB1/HSP70 after each fraction, consistent with ICD kinetics. | Same DAMP panel as for other RT types; no evidence of photon-specific DAMPs. Differences are dose-, time- and cell-line–dependent. [2,3] |
| γ-rays vs. BNCT (high-LET neutrons) | HMGB1 (cell supernatant, plasma) | In vitro: ~24 h; in vivo: peak ~day 3 | Xenograft model of BNCT vs. γ-irradiation | BNCT induced early extracellular HMGB1 release in vitro and a marked plasma HMGB1 peak around day 3 in mice; γ-rays also induced HMGB1 but in a less pronounced fashion. | Same DAMP (HMGB1) for γ and BNCT. Some quantitative enhancement with BNCT, but no new DAMP species; LET mainly affects magnitude and kinetics. [4] |
| Photons vs. protons (in vitro) | Ecto-calreticulin (ecto-CRT) | 12, 24, 48 h (max at ~48 h) | Multiple human tumor cell lines, including U251 glioma | Photon, proton and carbon-ion beams all increased ecto-CRT in a time- and dose-dependent manner. At iso-doses, photons and protons showed very similar CRT exposure; carbon ions required lower doses to reach comparable or higher CRT. | No qualitative difference in DAMP (CRT) between photons and protons. High-LET carbon ions show stronger CRT at lower doses, but the DAMP itself is the same. [5] |
| Photons vs. protons (in vitro) | Ecto-CRT, MHC I, ICAM-1, tumor antigens (immunogenic modulation) | 48–72 h after RT | Multiple tumor cell lines | Proton and photon irradiation produced a shared immunogenic modulation signature, including increased CRT and antigen presentation, rendering cells more sensitive to T cell killing. | Authors explicitly conclude that tumor cells surviving proton or photon RT share a common immunogenic profile; no DAMP unique to one modality. Differences are not consistent enough to claim modality-specific DAMP biology. [6] |
| Carbon ions (high LET) vs. X-rays | HMGB1 | 24–72 h after RT | Various human cancer cell lines | Carbon-ion beams induce HMGB1 release at levels comparable to X-rays; subsequent work shows HMGB1 release increases with LET, but remains the same molecule. | High-LET RT increases amount and persistence of HMGB1, but does not introduce new DAMPs; pattern is quantitative, not qualitative. [7,8] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



