
Submitted:
02 January 2026
Posted:
04 January 2026
You are already at the latest version
Abstract
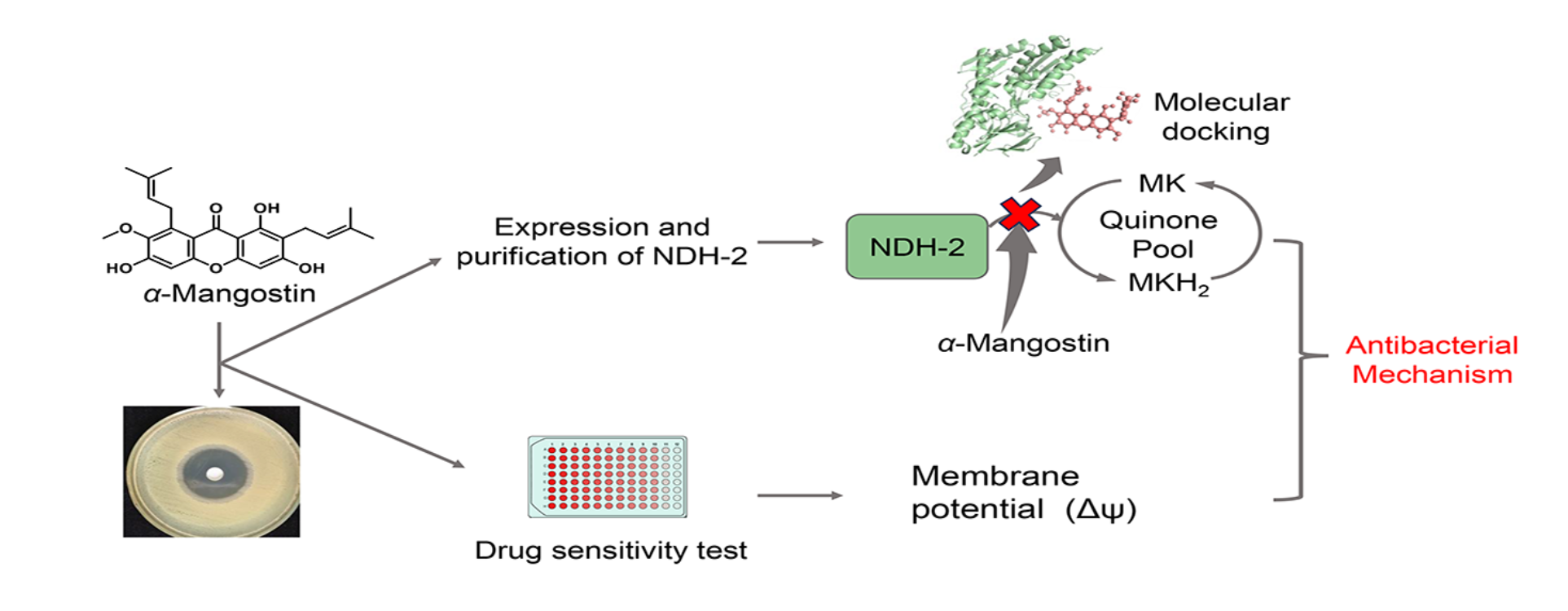
α-Mangostin, a natural product from Garcinia mangostana L, presents most antibacterial activity in plant flavonoids against Staphylococcus aureus so far. Recently, it was reported that the quinone pool is a key target of α-mangostin against Gram-positive bacteria. To further confirm this and investigate the detail of α-mangostin killing S. aureus, the interactions between α-mangostin and a key enzyme as type II NADH:quinone oxidoreductase (NDH-2), together with possible non-enzymatic mechanisms, were explored. Through the enzyme kinetic inhibition experiments, it was found that α-mangostin mainly competes with the menaquinone-binding sites of NDH-2, and the half-maximal inhibitory concentration (IC50) of α-mangostin on NDH-2 is 4.95 μM. Fluorescence analyses indicated that α-mangostin can spontaneously bind to NDH-2 to form an α-mangostin–NDH-2 complex. Subsequently, molecular simulation further indicated that α-mangostin can dock to the menaquinone-binding sites of NDH-2. Another, non-enzymatic mechanism showed that α-mangostin can cause membrane potential depolarization and disrupt the proton motive force balance, thereby promoting the cell-membrane destruction of S. aureus. These results suggest that α-mangostin mainly can interact with the amino acid residues at the menaquinone-binding pocket of NDH-2 to block the electron transfer at the quinone pool in the respiratory chain of S. aureus, and which will hinder the energy supply and promote its incidental effect on membrane disruption, ultimately leading to the death of S. aureus. This once again proves that the quinone pool is a key target of plant flavonoids against Gram-positive bacteria.
Keywords:
α-mangostin
; NDH-2
; respiratory chain
; menaquinone
; mechanism
; flavonoid
1. Introduction
The emergence of multidrug-resistant (MDR) bacteria has brought severe challenges to the global health. Methicillin-resistant Staphylococcus aureus (MRSA) is one of the main MDR bacteria causing mortality, and it is the predominant source of bloodstream, skin, and soft tissue infections [1,2]. Although some antibiotics have been approved for the treatment of MRSA infection, the discovery of new drugs especially from natural products with unique antimicrobial mechanisms remains essential for treating infections caused by MRSA [3].
Plant flavonoids, widely distributed in various plants and demonstrated favorable safety profiles, have been drawing increasing attention to their antibacterial potential. Some of them not only show stronger antibacterial activity than clinical antibiotics, but also present synergistic effects in combination with antimicrobial agents and even reverse antibiotic resistance [4,5]. α-Mangostin (Figure 1A) is a flavonoid, belonging to the subclass of xanthones, from Garcinia mangostana L [6]. It presents most antibacterial activity in plant flavonoids against Staphylococcus aureus so far, and has been studied as an antibacterial agent against MRSA since the 1980s [7,8]. It is indicated the cell membrane is an important target of α-mangostin against MRSA [9], but its effects on the respiratory chain were not involved. Recently, a research suggests that a key target for plant flavonoids inhibiting Gram-positive bacteria is the quinone pool of the respiratory chain, and which likely involves multiple mechanisms including some enzyme and non-enzyme inhibitions [10].
The quinone pool is a key hub of the electron transfer on the respiratory chain of S. aureus. In front of the quinone pool, NDH-2 is a key enzyme that facilitates the electron transfer from NADH to menaquinone [11,12]. Namely, it can catalyze the reduction of menaquinone, and which is the rate limiting step of electron transfer in the respiratory chain. The interaction between NDH-2 and the quinone pool helps sustain the NADH/NAD⁺ redox balance, ultimately driving ATP synthesis and ensuring the energy supply of S. aureus [13,14]. As NDH-2 is the sole NADH:quinone oxidoreductase in the respiratory chain of S. aureus, the inhibition to NDH-2 can eventually lead to bacterial cell death [15]. Simultaneously, preliminary molecular docking shows that α-mangostin can bind to the active pocket of NDH-2. Therefore, here the interactions between α-mangostin and NDH-2 of S. aureus, together with possible non-enzymatic mechanisms, were explored for the detail targeting the quinone pool of this flavonoid with most antibacterial activity against Gram-positive bacteria.
2. Materials and Methods
2.1. Materials, Chemicals, and Reagents
Escherichia coli BL21(DE3) and the pET-28a(+) vector were purchased from Tsingke Biotechnology Co., Ltd. (Beijing, China). The strain of MRSA ATCC 33592 and S.aureus ATCC 25923 was obtained from the American Type Culture Collection (Manassas, VA, USA). α-Mangostin (purity >98%, HPLC) was sourced from Push Bio-technology Co., Ltd. (Chengdu, China). Ni²⁺-NTA agarose resin was acquired from QIAGEN (Hilden, Germany). FAD (>98%, HPLC), NADH (>98%, HPLC), 3,3’-dipropylthiadicarbocyanine iodide DisC3(5) (>98%, HPLC) and menaquinone (MK) (>98%, HPLC) were all purchased from Sangon Biotech Co., Ltd. (Shanghai, China). Protein markers for gel analysis were obtained from Thermo Fisher Scientific (Waltham, MA, USA). Most chemical reagents used for NDH-2 expression, gel analysis, and Western blotting were supplied by Sangon Biotech Co., Ltd. (Shanghai, China), Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China), or Xilong Scientific Co., Ltd. (Shantou, China). Mueller-Hinton Broth (MHB) was used for antimicrobial susceptibility testing. α-Mangostin was dissolved in dimethyl sulfoxide (DMSO) and subsequently diluted to a final DMSO concentration of <2.5% (v/v) or lower.
2.2. Antimicrobial Susceptibility Assay
Following the standard procedures described by the Clinical and Laboratory Standards Institute (CLSI) [16], logarithmic phase bacterial cultures were diluted with Mueller-Hinton Broth (MHB) to a final concentration of approximately 1.0 × 106 CFU/mL. The susceptibility of S. aureus ATCC 25923 and MRSA ATCC 33592 to α-mangostin was then determined using the microbroth dilution method in 96-well plates. After incubating the 96-well plates at 35 °C for 24 h in an incubator, 20 μL of 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide (MTT, 4.0 mg/mL) was added to each well. The minimum inhibitory concentration (MIC) was defined as the lowest concentration of the compound that completely inhibited bacterial growth in the wells, determined by the absence of color change when bacterial growth was fully observed in the blank control wells [17]. Based on the MIC results, wells with α-mangostin concentrations at different multiples of the MIC were selected. A 100 μL aliquot of bacterial suspension was aspirated from each selected well and spread onto solid medium plates, which were then incubated overnight. The minimum bactericidal concentration (MBC) was determined according to the number of bacterial colonies grown on the solid medium plates.
2.3. Expression and Purification of the Enzyme
NDH-2 was expressed in E. coli BL21(DE3)/pET28a(+)-NDH-2. E. coli BL21(DE3)/pET28a(+)-NDH-2 was cultured on Luria-Bertani (LB) agar plates at 37 °C. NAH-2 single colony was picked and inoculated into fresh LB broth containing kanamycin (50 μg/mL), followed by incubation in a shaker at 37 °C and 180 rpm for 12 h. Subsequently, the culture was diluted 1:100 into fresh LB broth supplemented with kanamycin (50 μg/mL) and incubated at 37 °C until the optical density at 600 nm (OD600) reached 0.6–0.8. After cooling on ice for 15 min, isopropyl β-D-1-thiogalactopyranoside (IPTG) (Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China) was added to a final concentration of 0.4 mM. The culture was then induced for expression in a shaker at 25 °C and 150 rpm for 10–12 h. After induction, bacterial cells were harvested by low temperature centrifugation. Bacterial cells were resuspended in lysis buffer (50 mM Tris, 500 mM NaCl, 5% glycerol, 1 mM PMSF, pH 8.0) and lysed by ultrasonic disruptor. The supernatant was collected by centrifugation at 6000g for 25 min and loaded onto a Ni²⁺-NTA-agarose gel pre-equilibrated with buffer, washed with washing buffers containing different concentrations of imidazole. The purity of the purified protein was verified by SDS-PAGE, and the protein was identified by Western blotting. Using bovine serum albumin (BSA) as the standard, the protein concentration was determined with a BCA kit (Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China).
2.4. Determination of NDH-2 Activity
The oxidation rate of NADH was determined in the presence of different concentrations of substrates. The 200 μL reaction mixture contained 20 μM FAD, 0.5 μg NDH-2, 5% glycerol, 150 mM NaCl, and 50 mM Tris-HCl buffer (pH 7.5). Under the incubation condition of 30 °C, enzyme kinetic parameters were measured by adding MK (0–60 μM) or NADH (0–450 μM). The reaction was initiated by the addition of NADH. The absorbance was monitored at 340 nm using multifunctional microplate reader to track the reaction progress (NADH ε = 6.22 mM⁻¹ cm⁻¹).
2.5. Inhibition of NDH-2 by α-Mangostin
The assay was performed using multifunctional microplate reader by measuring the absorbance at 340 nm. The reaction system was referenced to Section 2.4 of this research. To evaluate the inhibitory effect of α-mangostin on NDH-2, the enzyme was pre-incubated with various concentrations of α-mangostin in the reaction mixture for 2 min with 40 μM MK, and the reaction was initiated by the addition of 300 μM NADH. To analyze the inhibitory mechanism of α-mangostin on NDH-2, different concentrations of substrate MK (0–60 μM) were added to the reaction mixture. The enzymatic reaction rates were measured at varying substrate concentrations in the presence of α-mangostin (0, 1, 2 and 4 μg/mL). Lineweaver-Burk plots were generated to determine the inhibition type and inhibition constant (Kᵢ) of α-mangostin against NDH-2.
2.6. Fluorescence Spectroscopy Analysis of the Interaction between α-Mangostin and NDH-2
Referring to the experimental method described by Zilong Luo et al. [18], fluorescence spectra of the interaction between α-mangostin and NDH-2 were measured using an FL970 fluorescence spectrophotometer (Taikang Instruments Co., Ltd., Shanghai, China). The slit width was set to 2.5 nm, the scan speed to 1200 nm/min, and the sampling interval to 0.2 nm.
The reaction system was prepared in PBS buffer (pH 7.4), containing 0.5 mg/mL NDH-2 and 20 μM FAD, with final concentrations of α-mangostin at 0, 1, 2, 4, 8, 10, 15 and 20 μg/mL, incubating the mixture at 298 K and 310 K for 5 min. The excitation wavelength was set to 280 nm, and fluorescence spectra were scanned in the range of 290–600 nm. Background fluorescence from the PBS buffer was subtracted, and the maximum emission wavelength and fluorescence intensity were recorded.
For the synchronous fluorescence spectra of the samples, measurements were performed at 298 K and 310 K. The excitation wavelength range was set to 250–500 nm, wavelength differences (Δλ) between emission and excitation wavelengths of 15 nm and 60 nm. The maximum emission wavelength and fluorescence intensity were recorded.
2.7. Molecular Docking
Molecular docking between α-mangostin and NDH-2 was performed using AutoDock Tools 1.5 [19]. The crystal structure of NDH-2 (PDB ID: 5NA1) was downloaded from the RCSB PDB online database (https://www.rcsb.org/ (accessed on 1 September 2025)), and all crystal water molecules and metal ions were removed from the protein structure. The three dimensional structure of α-mangostin (PubChem SID: 433967754) was retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/ (accessed on 1 September 2025)) and imported into Chem3D 22.2 for energy minimization. The active pocket of NDH-2 was predicted using the DrugRep tool [20]. The grid size was set to 32 Å × 20 Å × 32 Å, with the center coordinates of the docking box at X (14.8 Å), Y (14.2 Å), and Z (-24.7 Å), and a grid spacing of 0.375 Å. AutoDock Tools 1.5 was used to calculate the potential docking conformations of α-mangostin with NDH-2. The docking conformation with the lowest binding free energy was selected for binding mode analysis. The results were imported into PyMOL 2.6 and LigPlot+ 2.2.9 for further visualization and analysis, and to generate 3D and 2D docking interaction diagrams [21,22].
2.8. Measurement of Membrane Potential
The method described by Zhiqiang Wang et al. [23] was used with minor modifications. A single colony of MRSA ATCC 33592 was inoculated into MHB. Bacterial cells were harvested by centrifugation, washed with PBS buffer (pH 7.4), and resuspended in PBS buffer (plus 20 mM glucose). In a black 96-well plate, bacterial suspension and DisC₃(5) (1 μM) were added, followed by incubation at 37 °C in the dark for 10 min. After monitoring until the baseline stabilized, different concentrations of α-mangostin (2, 4, 8 and 16 × MIC) was added. After incubation for 30 min, the fluorescence intensity was measured using multifunctional microplate reader with an excitation wavelength of 622 nm and an emission wavelength of 670 nm.
3. Results
3.1. Minimum Inhibitory Concentrations (MICs) and Minimum Bactericidal Concentrations (MBCs)
The MICs of α-mangostin against S. aureus ATCC 25923 and MRSA ATCC 33592 was determined using the microbroth dilution method. The MICs of α-mangostin against S. aureus ATCC 25923 and MRSA ATCC 33592 are 2 μg/mL and 0.5 μg/mL, respectively. Based on the MIC results, their MBCs were further measured. The MBCs of α-mangostin against S. aureus ATCC 25923 and MRSA ATCC 33592 are 64 μg/mL and 4 μg/mL, respectively. These results suggest that α-mangostin has strong antibacterial activity against S. aureus.
3.2. Protein Purification and Enzymatic Characterization
To investigate the effect of α-mangostin on NDH-2, we successfully achieved in vitro expression of NDH-2 through transformation in E. coli BL21(DE3) followed by IPTG induction. SDS-PAGE analysis (Figure 2A) revealed a distinct protein band at approximately 43 kDa. Furthermore, Western blot analysis (Figure 2B) confirmed the expression of the target protein. Due to the protein carries a 6 × His tag, Ni²⁺-NTA agarose affinity chromatography combined with concentrator ultrafiltration was employed to obtain a highly purified protein sample that suitable for subsequent experiments. UV spectrophotometric analysis of the protein sample showed no absorption peak at 450 nm, indicating that the heterologously expressed NDH-2 did not contain FAD, which an essential cofactor in the NDH-2 protein structure. Fortunately, the addition of FAD to the reaction system allowed the enzymatic reaction to proceed normally.
The natural oxidant for NDH-2 in the membrane of S. aureus is menaquinone [12]. Therefore, in this experiment, menaquinone was used as the oxidant to evaluate the NADH:menaquinone oxidoreductase activity of purified NDH-2 in the presence of NADH, and several enzymatic kinetic characteristics were determined (Figure 2C–F). Under the conditions of 30 °C and pH 7.5, the NADH:menaquinone oxidoreductase activity dependent on either NADH or MK followed simple Michaelis-Menten kinetics. The Km value for NADH was 157.95 ± 42.71 μM, and for MK was 19.26 ± 1.91 μM, which is consistent with previously reported values [24]. The successful determination of these enzymatic kinetic parameters indicates that NDH-2 effectively oxidizes NADH to NAD⁺. Thus, the purified NDH-2 can be utilized for investigating the antibacterial mechanism of α-mangostin.
3.3. Inhibitory Effect of α-Mangostin on NDH-2 Activity
The relationship between the relative activity of NDH-2 and different concentrations of α-mangostin is shown in Figure 3A. Data analysis revealed that the half maximal inhibitory concentration (IC50) of α-mangostin for NDH-2 is 2.03 ± 0.09μg/mL (4.95 μM). HQNO, a known effective inhibitor of NDH-2, has a reported IC50 of 7.3 ± 1.2 μM [25]. Therefore, α-mangostin can be considered to exhibit stronger inhibitory efficacy against NDH-2 in vitro compared to the positive control, suggesting its potential as a promising drug candidate which targets NDH-2.
To further elucidate the inhibitory mechanism of α-mangostin on NDH-2, enzyme kinetic inhibition assays were performed (Figure 3B). The fitted curves for different concentrations of α-mangostin intersect in the second quadrant, and with increasing inhibitor concentration, the Kₘ value gradually increases while the Vₘₐₓ value decreases. These results indicate that α-mangostin acts as a mixed type inhibitor of NDH-2. The inhibitor constant (Ki), also known as the dissociation constant of the enzyme inhibitor complex, reflects the binding affinity between the enzyme and the inhibitor. A lower Kᵢ value indicates stronger inhibition. Experimental results show that the competitive inhibition constant (Kic) of α-mangostin for NDH-2 is 1.73 μM, and the noncompetitive inhibition constant (Kin) is 5.70 μM. The significantly lower Kic value compared to Kin suggests that the competitive inhibition mode predominates in the mixed type inhibition, indicating that α-mangostin can interact with both free NDH-2 and the NDH-2–MK complex, but exhibits stronger binding to free NDH-2.
3.4. Analysis of the Binding Properties Between α-Mangostin and NDH-2
The intrinsic fluorescence properties of proteins, attributed to amino acids such as tryptophan (Trp) and tyrosine (Tyr), are highly sensitive to changes in their microenvironments. Therefore, fluorescence spectroscopy can be employed to analyze the binding characteristics and interactions between proteins and drug molecules [26]. With the excitation wavelength fixed at 280 nm, the fluorescence emission spectrum of NDH-2 was measured. As shown in Figure 4A,B, the maximum fluorescence intensity peak of NDH-2 appeared near 335 nm. Under incubation conditions at 298 K and 310 K, the fluorescence intensity of NDH-2 gradually decreased with increasing concentrations of α-mangostin, indicating that α-mangostin interacts with Trp and Tyr residues in NDH-2 in a manner that quenches their fluorescence.
To further investigate the quenching mechanism of NDH-2 by α-mangostin, we calculated the quenching constant (Ksv), bimolecular quenching rate constant (Kq), binding constant (Ka), number of binding sites (n), enthalpy change (ΔH), entropy change (ΔS), and Gibbs free energy (ΔG) based on fluorescence spectra obtained at 298 K and 310 K. The results are summarized in Table 1, and the linear fitting curves are shown in Figure 4C,D. As seen in Table 1, the Ksv values for the quenching of NDH-2 by α-mangostin decrease with increasing temperature. Additionally, the Kq values exceed the maximum dynamic quenching constant (2.0 × 1010 L/mol/s), indicating that the quenching mechanism is static quenching. During this process, a α-mangostin–NDH-2 complex is formed, and the stability of this complex decreases with rising temperature, suggesting that the binding affinity between α-mangostin and NDH-2 weakens as temperature increases. The values of n at both 298 K and 310 K are greater than 0.5, indicating a single binding site for α-mangostin on NDH-2. This suggests the presence of a specific binding site for α-mangostin in the NDH-2, which may be located within the enzyme’s active center. Furthermore, the Ka for the interaction between α-mangostin and NDH-2 are all below 104 L·mol⁻¹, indicating weak binding. As temperature increases, the Ka values decrease, which aligns with the theoretical expectation that higher temperatures reduce the stability of protein–quencher complexes and lead to lower binding constants in static quenching. This result further confirms that the binding affinity between α-mangostin and NDH-2 diminishes with increasing temperature. In addition, in the binding process between α-mangostin and NDH-2, the values of ΔG, ΔH, and ΔS are all negative, indicating that the binding is spontaneous. Hydrogen bonds and van der Waals forces play dominant roles in the interaction. Moreover, the negative ΔH value signifies an exothermic reaction, which contributes to the weakening of binding and the observed decrease in Ka. This finding is consistent with the conclusions drawn above.
To investigate the microenvironmental information of Tyr or Trp residues after the binding of α-mangostin to NDH-2, synchronous fluorescence spectra at different α-mangostin concentrations were measured, with Δλ between emission and excitation wavelengths set at 15 nm and 60 nm, respectively. The results (Figure 4E) show that as the concentration of α-mangostin increases, all maximum emission wavelengths (λmax) exhibit a blue shift. This indicates that α-mangostin specifically binds to NDH-2, leading to a decrease in the polarity and freedom of the microenvironment around the Trp and Tyr residues of the protein. It is thus inferred that the binding site is located in the hydrophobic region of the protein, where α-mangostin interacts stably with NDH-2 through hydrophobic interactions and hydrogen bonding, which subsequently supported by molecular docking experiments. At 298 K, the blue shifts in λmax for both Δλ = 15 nm and Δλ = 60 nm were approximately 1 nm, with no significant difference observed. In contrast, at 310 K, the blue shift in λmax under Δλ = 15 nm (approximately 1.2–5.6 nm) was greater than that under Δλ = 60 nm (approximately 0.4 nm). This suggests that the increase in temperature weakens the binding between α-mangostin and NDH-2, resulting in enhanced hydrophobicity of the protein’s microenvironment. Furthermore, the data indicate that α-mangostin has a more pronounced effect on Tyr residues than on Trp residues.
3.5. Interaction Pattern Between α-Mangostin and NDH-2
Through molecular docking, we further investigated the binding site of α-mangostin with NDH-2. The docking results (Figure 5) indicate that α-mangostin binds to the active center of NDH-2, with the docking site located the MK - binding pocket. Both the 2D and 3D docking diagrams clearly illustrate the types and strengths of interactions between α-mangostin and the amino acid residues in the binding region. Specifically, α-mangostin forms hydrogen bonds with two amino acids on NDH-2: a phenolic hydroxyl group forms a hydrogen bond with Thr169 (bond length: 2.75 Å), and the ketone carbonyl group forms a hydrogen bond with Ala319 (bond length: 2.74 Å). Additionally, hydrophobic interactions are observed with Tyr15, Asp302, Thr48, Thr318, Tyr347, Thr352, Phe168, Phe111, and Ala47. This hydrophobic cavity largely overlaps with the conserved cavity on the FAD si-side of NDH-2, which has been characterized as the quinone-binding site [14]. Therefore, these results further confirm that α-mangostin primarily competes with MK for binding at the active center of NDH-2.
3.6. Influence of Membrane Potential
DisC3(5) is a membrane potential-sensitive cyanine dye whose distribution between bacterial cells and the culture medium depends on the cytoplasmic membrane potential. When the dye enters cells, self-fluorescence quenching occurs due to increased concentration. Disruption of the cytoplasmic membrane structure leads to membrane potential dissipation, resulting in the release of the dye into the culture medium and a subsequent increase in fluorescence intensity. Specifically, the degree of membrane potential reduction is proportional to the increase in fluorescence intensity, and this change can be detected using a fluorescence spectrophotometer [27]. Addition of α-mangostin at different concentrations to bacterial suspensions induced an increase in DisC3(5) fluorescence intensity, indicating membrane potential collapse and the release of accumulated DisC3(5). This concentration dependent relationship is shown in Figure 6, low concentrations of α-mangostin caused a gradual loss of membrane potential, while the fluorescence signal under high concentrations of α-mangostin exhibited a strong and immediate increase. These results demonstrate that α-mangostin significantly disrupts the membrane potential of S. aureus, inducing a depolarization effect.
4. Discussion
As a plant-derived flavonoid, α-mangostin exhibits broad pharmacological activities and has been demonstrated to possess potent inhibitory activity against Gram-positive bacteria [28]. The above results suggest that α-mangostin can compete with menaquinone to bind the active sites of NDH-2 of S. aureus, blocking the electron transfer into the quinone pool of the respiratory chain, affecting energy supply, and ultimately leading to bacterial autolysis. It is confirmed that the quinone pool in the respiratory chain is a major target of plant flavonoids against Gram-positive bacteria [10]. Therefore, here this conclusion is further approved.
In Gram-positive bacteria, menaquinone, as an electron receptor on the cell membrane, is the only quinone that performs electron transfer. After accepting the electrons from NDH-2, it is reduced to MKH₂ which will transfer the electrons to the next enzyme in the respiratory chain, and this process also plays an important role for maintaining the energy for the proton gradient [12,29,30]. The analyses of fluorescence spectroscopy indicated that a significant quenching for the intrinsic fluorescence of NDH-2 occurred when incubated with α-mangostin. This quenching presented remarkably a concentration dependent pattern which is consistent with the Stern-Volmer equation, and accompanied by blue shift in fluorescence peak positions and changes in fluorescence lifetime. This indicates there is direct binding between α-mangostin and NDH-2. By fitting the data, general parameters such as the binding constant, number of binding sites, enthalpy change, entropy change, and Gibbs free energy were obtained, indicating that the binding affinity decreases with the increase of temperature. This confirms that the quenching is static in nature, suggesting a spontaneous process of α-mangostin binding NDH-2.
Previous studies indicated that MK-4 or the MK extract from S. aureus can attenuate the antibacterial activity of plant flavonoids against S. aureus, and their antibacterial activities continuously decrease with the increase in the interfering concentrations of MK-4 and the MK extract, especially for α-mangostin [10]. Here enzyme kinetic inhibition assays indicate that α-mangostin can directly binds to NDH-2 of S. aureus. These findings together suggest that α-mangostin likely can target the menaquinone-binding sites of NDH-2 to lead to the blocking of the binding of MK and NDH-2, rather than inhibit the biosynthesis of MK. Namely, α-mangostin can compete with MK for the menaquinone-binding sites of NDH-2 of S. aureus. This is also confirmed by the results of molecular docking showing that α-mangostin can interact with the amino acid residues of MK-binding pocket in NDH-2 of S. aureus.
It was reported that there are two independent binding sites in NDH-2, for the bindings of MK and NADH, respectively [14]. This research reveals that α-mangostin can bind to the MK-binding site of this protein. Simultaneously, under conditions where MK is saturated, the double-reciprocal plot of NADH as a substrate exhibits a nonlinear pattern in the presence of α-mangostin. Therefore, it can be inferred that α-mangostin specifically competes for the MK-binding site without interfering the binding of NADH. This is also supported by previous work which confirms that the engagement of drug to MK-binding sites of NDH-2 does not directly affect the NADH binding process and that two binding sites are independent and functionally segregated [31].
Furthermore, this research showed that α-mangostin can cause the collapse of the membrane potential of S. aureus, and this is in accordance with previous report that α-mangostin can disrupt the cytoplasmic membrane and alter the proton motive force [9]. However, its low hemolysis rate on sheep red blood cells [32], with only 25.86% of red blood cells lysed at 32 μg/mL. Since α-mangostin has strong antibacterial activity with the MICs ranged from 0.5 to 2 μg/mL, these facts indicated that its effects on cytoplasmic membrane and proton motive force are probably incidental or at least nonessential. After all, the blocking of electron transfer in the respiratory chain also can probably lead to these effects as mentioned above [33,34]. Together with the consideration that menaquinone is the sole quinone in the quinone pool of S. aureus, it was deduced that α-mangostin mainly competes for the menaquinone-binding sites of NDH-2 to block the electron transfer at the quinone pool in the respiratory chain, hindering the energy supply of S. aureus, and which will also promote its effect on membrane disruption, ultimately leading to a rapid bactericidal effect. Since α-mangostin is a representative of plant flavonoids with most antibacterial activity against S. aureus, these finding once again proves that the quinone pool is a key target of plant flavonoids against Gram-positive bacteria, but also provides a new antibacterial mechanism of plant flavonoids.
5. Conclusions
In summary, this research demonstrates that α-mangostin mainly can interact with the amino acid residues at the menaquinone-binding pocket of NDH-2 to block the electron transfer at the quinone pool in the respiratory chain of S. aureus. This will hinder the energy supply and promote its incidental effect on membrane disruption, and ultimately leading to the death of S. aureus. Furthermore, this further supports the conclusion that the quinone pool is the key target of plant flavonoids inhibiting Gram-positive bacteria.
Author Contributions
Meifang Zhang: Methodology, Writing – original draft, Software, Formal analysis, Investigation, Data curation, Visualization. Jianing Hu: Data curation, Investigation, Methodology. Yu Wang: Investigation, Visualization. Liaolongyan Luo: Investigation, Visualization. Ganjun Yuan: Writing – review & editing, Conceptualization, Investigation, Methodology, Software, Validation, Formal analysis, Data curation, Resources, Project administration, Funding acquisition, Supervision.
Funding
This research was funded by the National Natural Science Foundation of China, grant numbers 82360691 and 82073745.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this research. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- GBD 2021 Antimicrobial Resistance Collaborators. Global burden of bacte-rial antimicrobial resistance 1990-2021: a systematic analysis with forecaststo 2050. Lancet 2024, 404(10459), 1199–1226. [Google Scholar] [CrossRef]
- Talan, DA; Krishnadasan, A; Gorwitz, RJ; Fosheim, GE; Limbago, B; Albrecht, V; et al. Comparison of Staphylococcus aureus from skin and soft-tissue infections in US emergency department patients, 2004 and 2008. Clin Infect Dis. 2011, 53, 144–9. [Google Scholar] [CrossRef]
- Kurosu, M; Siricilla, S; Mitachi, K. Advances in MRSA drug discovery: where are we and where do we need to be? Expert Opin Drug Discov 2013, 8, 1095–116. [Google Scholar] [CrossRef]
- Cushnie, TPT; Lamb, AJ. Antimicrobial activity of flavonoids. Int J Antimicrob Agents 2005, 26, 343–56. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, TLA; Bhattacharya, D. Antimicrobial activity of quercetin: An ap-proach to its mechanistic principle. Molecules 2022, 27, 2494. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y; Guan, T; Wang, S; Zhou, C; Wang, M; Wang, X; et al. Novel xanth-one antibacterials: Semi-synthesis, biological evaluation, and the action me-chanisms. Bioorganic & Medicinal Chemistry 2023, 83, 117232. [Google Scholar] [CrossRef]
- Iinuma, M; Tosa, H; Tanaka, T; Asai, F; Kobayashi, Y; Shimano, R; et al. Antibacterial activity of xanthones from guttiferaeous plants against methicil-lin-resistant Staphylococcus aureus. J Pharm Pharmacol. 1996, 48, 861–5. [Google Scholar] [CrossRef]
- Sakagami, Y; Iinuma, M; Piyasena, KGNP; Dharmaratne, HRW. Antibacterial activity of alpha-mangostin against vancomycin resistant Enterococci (VRE) and synergism with antibiotics. Phytomedicine 2005, 12, 203–8. [Google Scholar] [CrossRef] [PubMed]
- Koh, JJ; Qiu, S; Zou, H; Lakshminarayanan, R; Li, J; Zhou, X; et al. Rapid bactericidal action of alpha-mangostin against MRSA as an outcome of membrane targeting. Biochim Biophys Acta 2013, 1828, 834–44. [Google Scholar] [CrossRef]
- Zhang, L; Yan, Y; Zhu, J; Xia, X; Yuan, G; Li, S; et al. Quinone pool, a key target of plant flavonoids inhibiting Gram-Positive Bacteria. Molecules 2023, 28, 4972. [Google Scholar] [CrossRef]
- Kaila, VRI; Wikström, M. Architecture of bacterial respiratory chains. Nat Rev Microbiol 2021, 19, 319–30. [Google Scholar] [CrossRef]
- Kurosu, M; Begari, E. Vitamin K2 in electron transport system: are enzymes involved in vitamin K2 biosynthesis promising drug targets? Molecules 2010, 15, 1531–53. [Google Scholar] [CrossRef]
- Sena, FV; Sousa, FM; Pereira, AR; Catarino, T; Cabrita, EJ; Pinho, MG; et al. The two alternative NADH:quinone oxidoreductases from Staphylococcus aureus: Two players with different molecular and cellular roles. Microbiol Spectr. 2024, 12, e0415223. [Google Scholar] [CrossRef]
- Sena, FV; Batista, AP; Catarino, T; Brito, JA; Archer, M; Viertler, M; et al. Type-II NADH:quinone oxidoreductase from Staphylococcus aureus has two distinct binding sites and is rate limited by quinone reduction. Mol Microbiol. 2015, 98, 272–88. [Google Scholar] [CrossRef] [PubMed]
- Schurig-Briccio, LA; Yano, T; Rubin, H; Gennis, RB. Characterization of the type 2 NADH:menaquinone oxidoreductases from Staphylococcus aureus and the bactericidal action of phenothiazines. Biochim Biophys Acta 2014, 1837, 954–63. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory and Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, Approved Standards, CLSI Document M07-A10. Clinical and Laboratory and Standards Institute: Wayne, PA, USA, 2024.
- Xu, X; Xu, L; Yuan, G; Wang, Y; Qu, Y; Zhou, M. Synergistic combinationof two antimicrobial agents closing each other’s mutant selection windows to prevent antimicrobial resistance. Sci Rep. 2018, 8, 7237. [Google Scholar] [CrossRef]
- Luo, Z; Li, P; Zhang, D; Zhu, J; Yuan, G; et al. A novel antimicrobial mechanism of Azalomycin F acting on lipoteichoic acid synthase and cell envelope. Molecules 2024, 29, 856. [Google Scholar] [CrossRef]
- Morris, GM; Huey, R; Lindstrom, W; Sanner, MF; Belew, RK; et al. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J Comput Chem. 2009, 30, 2785–91. [Google Scholar] [CrossRef]
- Gan, JH; Liu, JX; Liu, Y; Chen, SW; Dai, WT; Xiao, ZX; et al. DrugRep: an automatic virtual screening server for drug repurposing. Acta Pharmacol Sin. 2023, 44, 888–96. [Google Scholar] [CrossRef]
- Schrödinger, LLC. The PyMOL molecular graphics system, Version 1.8 2015.
- Laskowski, RA; Swindells, MB. LigPlot+: multiple ligand-protein interaction diagrams for drug discovery. J Chem Inf Model. 2011, 51, 2778–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y; Jia, Y; Yang, K; Li, R; Xiao, X; Zhu, K; et al. Metformin restores tetracyclines susceptibility against multidrug resistant bacteria. Advanced Science 2020, 7, 1902227. [Google Scholar] [CrossRef]
- Mogi, T; Matsushita, K; Murase, Y; Kawahara, K; Miyoshi, H; Ui, H; et al. Identification of new inhibitors for alternative NADH dehydrogenase (NDH-II). FEMS Microbiol Lett. 2009, 291, 157–61. [Google Scholar] [CrossRef] [PubMed]
- Petri, J; Shimaki, Y; Jiao, W; Bridges, HR; Russell, ER; Parker, EJ; et al. Structure of the NDH-2 - HQNO inhibited complex provides molecular insight into quinone-binding site inhibitors. Biochim Biophys Acta Bioenerg. 2018, 1859, 482–90. [Google Scholar] [CrossRef]
- Xi, L; Wang, Y; He, Q; Zhang, Q; Du, L. Interaction between Pin1 and its natural product inhibitor epigallocatechin-3-gallate by spectroscopy and molecular dynamics simulations. Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy 2016, 169, 134–43. [Google Scholar] [CrossRef] [PubMed]
- Wu, M; Hancock, REW. Interaction of the cyclic antimicrobial cationic peptide bactenecin with the outer and cytoplasmic membrane. Journal of Biological Chemistry 1999, 274, 29–35. [Google Scholar] [CrossRef]
- Górecka, H; Guźniczak, M; Buzalewicz, I; Ulatowska-Jarża, A; Korzekwa, K; Kaczorowska, A. Alpha-mangostin: A review of current research on its potential as a novel antimicrobial and anti-biofilm ggent. Int J Mol Sci. 2025, 26(11), 5281. [Google Scholar] [CrossRef] [PubMed]
- Paudel, A; Hamamoto, H; Panthee, S; Sekimizu, K. Menaquinone as a potential target of antibacterial agents. Drug Discov Ther. 2016, 10, 123–8. [Google Scholar] [CrossRef]
- Boersch, M; Rudrawar, S; Grant, G; Zunk, M. Menaquinone biosynthesis inhibition: a review of advancements toward a new antibiotic mechanism. RSC Adv. 2018, 8, 5099–105. [Google Scholar] [CrossRef]
- Yano, T; Rahimian, M; Aneja, KK; Schechter, NM; Rubin, H; Scott, CP. M-ycobacterium tuberculosis Type II NADH-menaquinone oxidoreductase cat-alyzes electron transfer through a two-site ping-pong mechanism and has two quinone-binding sites. Biochemistry 2014, 53, 1179–90. [Google Scholar] [CrossRef]
- Song, M; Liu, Y; Li, T; Liu, X; Hao, Z; Shen, J; et al. Plant natural flavonoids against multidrug resistant pathogens. Advanced Science 2021, 8, 2100749. [Google Scholar] [CrossRef]
- Geberetsadik, G; Inaizumi, A; Nishiyama, A; Yamaguchi, T; Hamamoto, H; Panthee, S; et al. Lysocin E targeting menaquinone in the membrane of my-cobacterium tuberculosis is a promising lead compound for antituberculosisdrugs. Antimicrob Agents Chemother. 2022, 66(9), e0017122. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, H; Urai, M; Ishii, K; Yasukawa, J; Paudel, A; Murai, M; et al. Lysocin E is a new antibiotic that targets menaquinone in the bacterial membrane. Nat Chem Biol. 2015, 11, 127–33. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) Chemical structure of α-mangostin. (B) Diagram of α-mangostin acting on NDH-2 in the respiratory chain of S. aureus.
Figure 1.
(A) Chemical structure of α-mangostin. (B) Diagram of α-mangostin acting on NDH-2 in the respiratory chain of S. aureus.
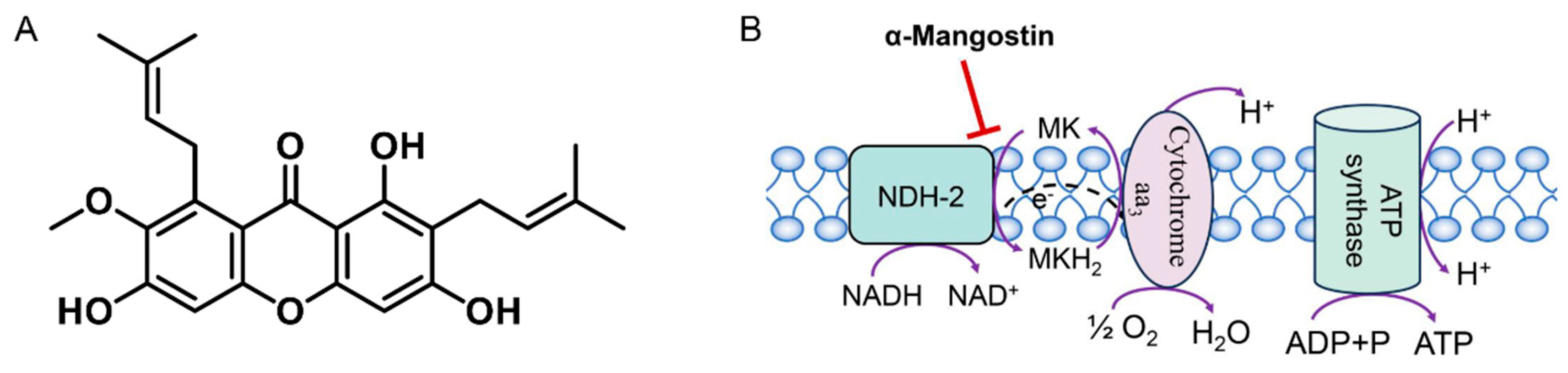
Figure 2.
Heterologous expression and enzymatic characterization of NDH-2. (A) SDS-PAGE of purified NDH-2: 1: Uninduced without IPTG; 2: Supernatant; 3: Precipitate; 4: Flow through; 5: 20 mM imidazole; 6: 75 mM imidazole; 7: 100 mM imidazole. (B) Western blot of NDH-2: a: Purified protein; b~d: Supernatant. (C, D) Michaelis-Menten plots of the enzyme against different substrate concentrations. (E, F) Lineweaver-Burk plots of NDH-2.
Figure 2.
Heterologous expression and enzymatic characterization of NDH-2. (A) SDS-PAGE of purified NDH-2: 1: Uninduced without IPTG; 2: Supernatant; 3: Precipitate; 4: Flow through; 5: 20 mM imidazole; 6: 75 mM imidazole; 7: 100 mM imidazole. (B) Western blot of NDH-2: a: Purified protein; b~d: Supernatant. (C, D) Michaelis-Menten plots of the enzyme against different substrate concentrations. (E, F) Lineweaver-Burk plots of NDH-2.
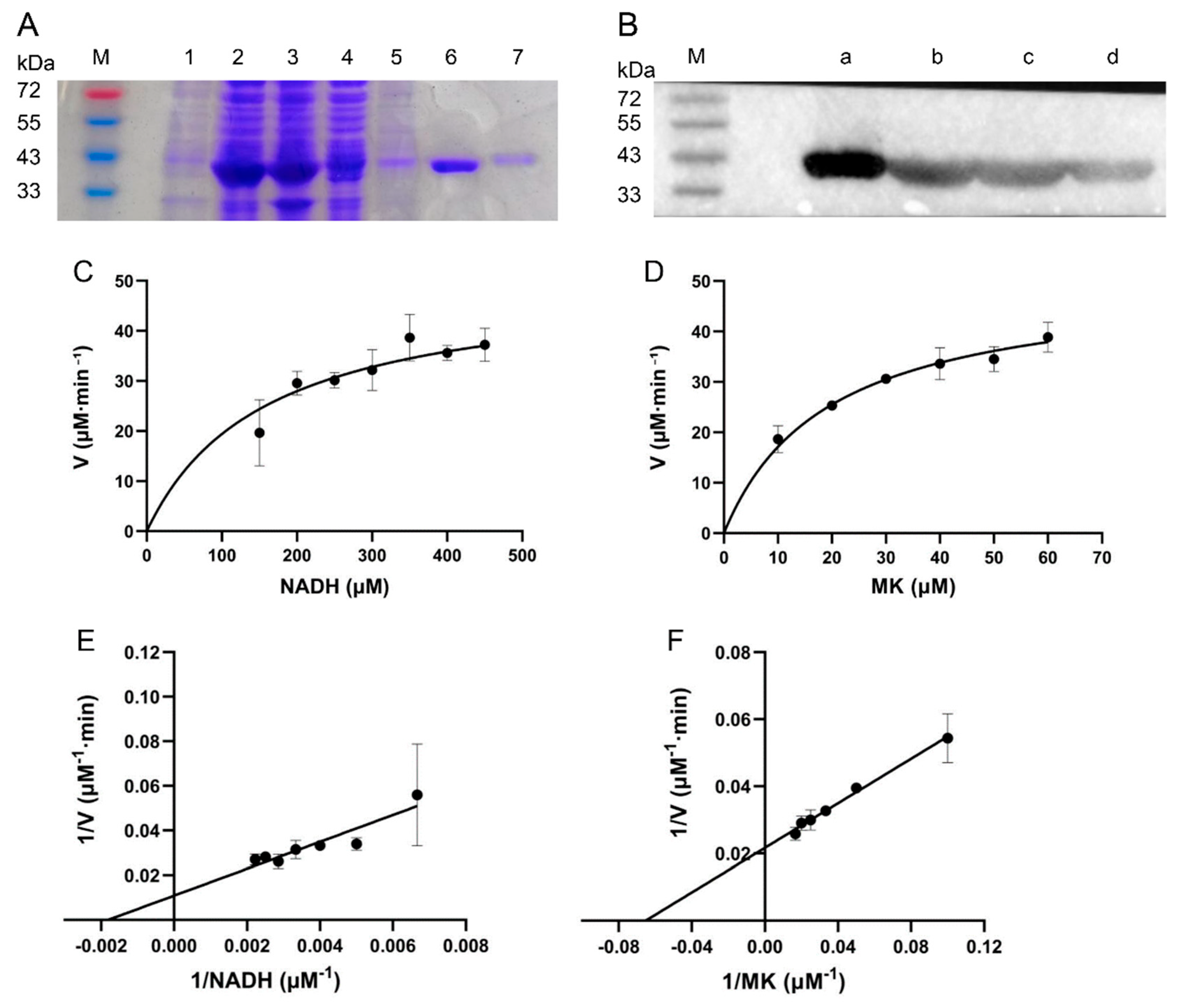
Figure 3.
Inhibitory effect of α-mangostin on NDH-2. (A) Effect of α-mangostin on NDH-2 activity in the presence of 40 μM MK. (B) Kinetic analysis of NDH-2 inhibition by α-mangostin (Lineweaver–Burk plot).
Figure 3.
Inhibitory effect of α-mangostin on NDH-2. (A) Effect of α-mangostin on NDH-2 activity in the presence of 40 μM MK. (B) Kinetic analysis of NDH-2 inhibition by α-mangostin (Lineweaver–Burk plot).
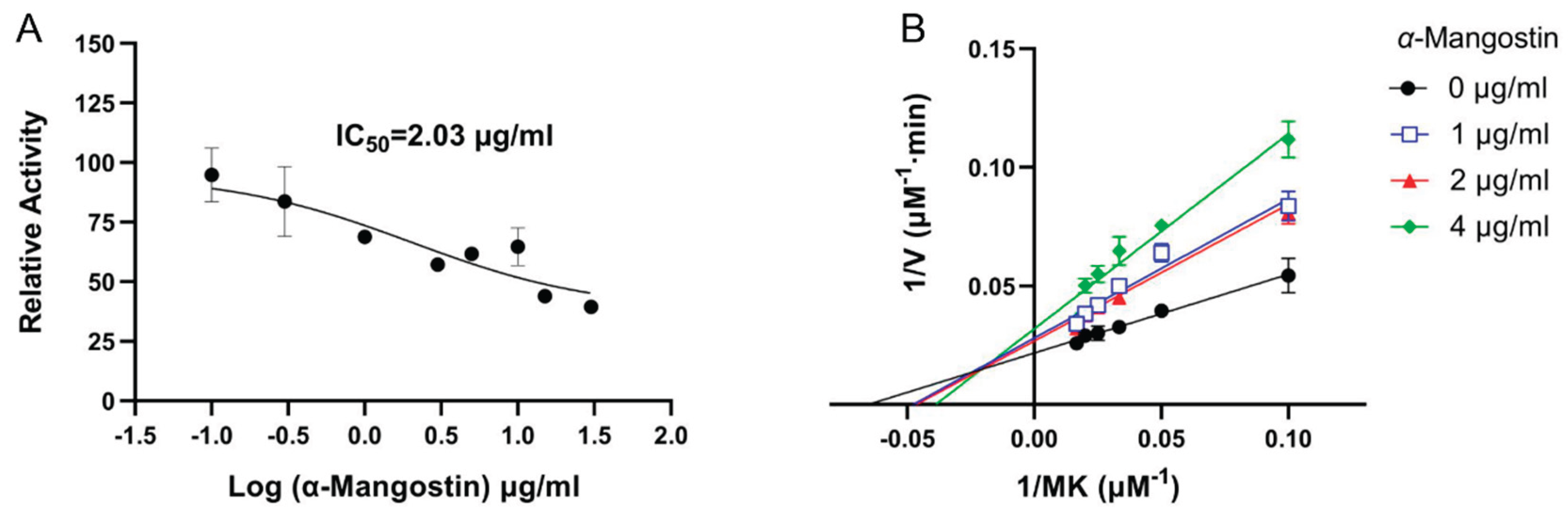
Figure 4.
Fluorescence analysis diagram of α-mangostin quenching NDH-2. (A, B) Fluorescence emission spectra of NDH-2 quenched by α-mangostin at different temperatures. (C) Stern-Volmer plots of α-mangostin quenching NDH-2 at 298K and 310K. (D) Double logarithmic plots of α-mangostin quenching NDH-2 at 298K and 310K. (E) Synchronous fluorescence spectra of α-mangostin quenching NDH-2 Tyr and Trp at 298K and 310K respectively (Δλ = 15 nm for Tyr, Δλ = 60 nm for Trp).
Figure 4.
Fluorescence analysis diagram of α-mangostin quenching NDH-2. (A, B) Fluorescence emission spectra of NDH-2 quenched by α-mangostin at different temperatures. (C) Stern-Volmer plots of α-mangostin quenching NDH-2 at 298K and 310K. (D) Double logarithmic plots of α-mangostin quenching NDH-2 at 298K and 310K. (E) Synchronous fluorescence spectra of α-mangostin quenching NDH-2 Tyr and Trp at 298K and 310K respectively (Δλ = 15 nm for Tyr, Δλ = 60 nm for Trp).
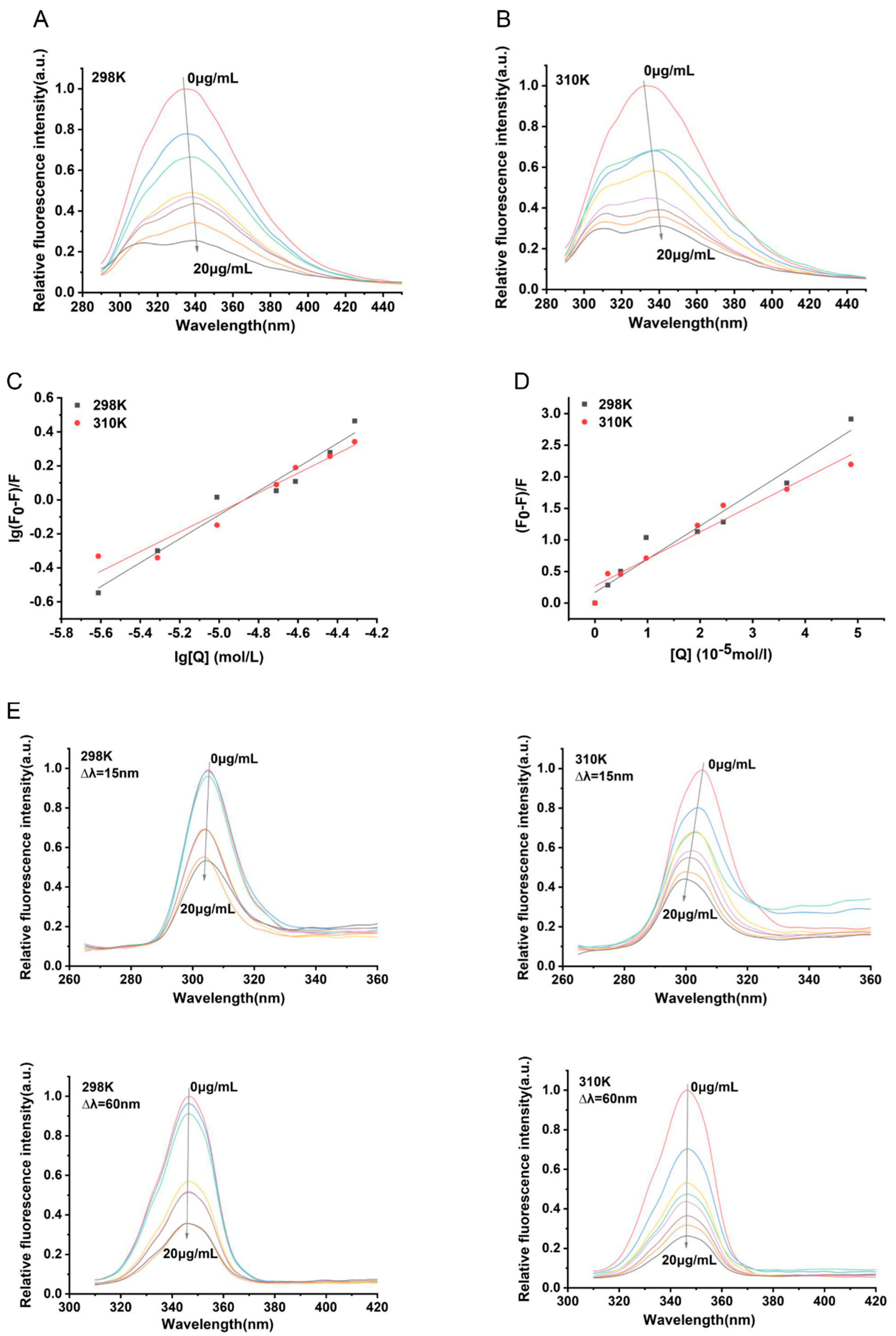
Figure 5.
2D and 3D structures of molecular docking between α-mangostin and NDH-2 (PDB ID: 5NA1).
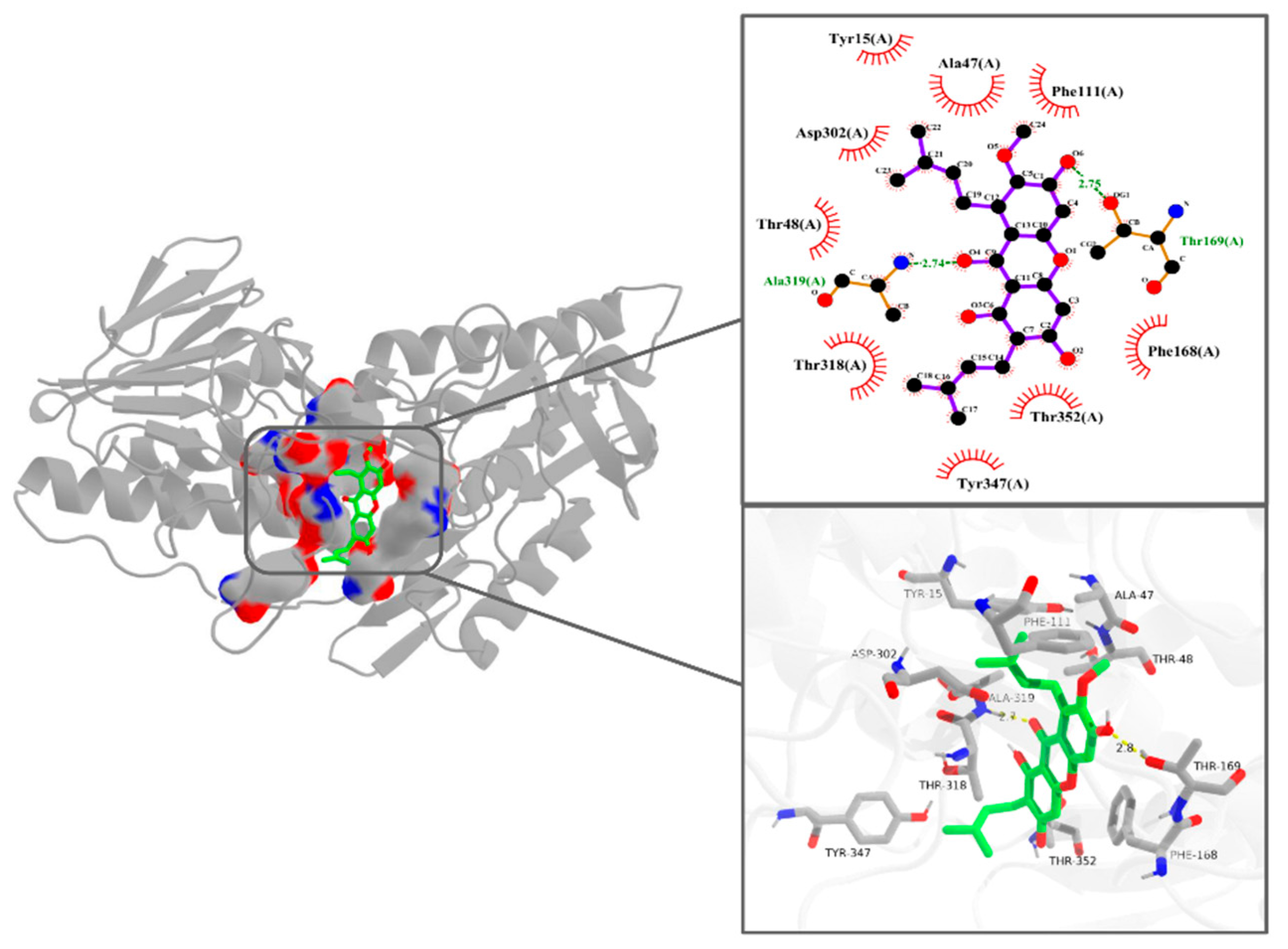
Figure 6.
Depolarization of MRSA membrane potential by α-mangostin at concentrations of 2, 4, 8 and 16 × MIC. Experiments were performed as three biologically independent replicates, and data are presented as mean ± SD (n = 3).
Figure 6.
Depolarization of MRSA membrane potential by α-mangostin at concentrations of 2, 4, 8 and 16 × MIC. Experiments were performed as three biologically independent replicates, and data are presented as mean ± SD (n = 3).

Table 1.
Parameters for the Interaction Between α-Mangostin and NDH-2 Obtained from Fluorescence Quenching at 298 K and 310 K a.
Table 1.
Parameters for the Interaction Between α-Mangostin and NDH-2 Obtained from Fluorescence Quenching at 298 K and 310 K a.
| Parameters | 298 K | 310 K |
|---|---|---|
| Ksv (×104 L·mol-1) | 5.275 ± 0.045 | 4.273 ± 0.045 |
| Kq (×1012 L·mol-1) | 5.275 ± 0.045 | 4.273 ± 0.037 |
| Ra | 0.979 | 0.978 |
| Ka (L·mol-1) | 2.649×103 | 646.725 |
| N | 0.702 ± 0.065 | 0.577 ± 0.059 |
| Rb | 0.979 | 0.975 |
| ΔH (kJ·mol-1) | -90.239 | -90.239 |
| ΔG (kJ·mol-1) | -19.528 | -16.680 |
| ΔS (kJ·mol-1·K-1) | -0.237 | -0.237 |
a Ksv, quenching constant; Kq, bimolecular quenching rate constant; Ka, binding constant; n, number of binding sites; ΔH, enthalpy change; ΔG, Gibbs free energy change; ΔS, entropy change; Ra and Rb are the Pearson correlation coefficients for Ksv, Ka and n.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



