Submitted:
01 January 2026
Posted:
02 January 2026
You are already at the latest version
Abstract
Dendrobium officinale (DO) is a traditional medicinal and edible plant whose polysaccharides help modulate gastrointestinal and metabolic functions. Fresh DO is commonly processed into “Fengdou” to prolong shelf life, but the effects of this processing on polysaccharide structure and bioactivity remain unclear. In this study, polysaccharides from fresh DO (FDOP) and Fengdou (DDOP) were isolated, purified, and comparatively characterized. Fourier transform infrared (FT-IR) analysis indicated similar functional groups and O-acetylated pyranosyl structures in both polysaccharides. Based on monosaccharide composition, methylation, and Nuclear Magnetic Resonance (NMR) analyses, both samples were identified as mannose-glucose heteropolysaccharides. However, FDOP was characterized by a higher mannose-to-glucose ratio (79.77:19.57) and molecular weight (187.1 kDa), as well as a more structurally diversified →4-linked backbone, whereas DDOP contained more glucose (68.74:30.94) and exhibited a lower molecular weight (125.1 kDa) and simplified backbone. In zebrafish models, both polysaccharides were found to alleviate loperamide-induced constipation and reduce lipid accumulation. DDOP showed stronger constipation-relieving activity, whereas FDOP exerted more pronounced hypolipidaemic effects, which may be attributed to its higher molecular weight, mannose enrichment, and more complex backbone structure. These findings provide a structural basis and theoretical support for developing DO-derived polysaccharides as functional food ingredients targeting constipation and dyslipidaemia.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Extraction and Purification of Dendrobium officinale Polysaccharides
2.2.1. Polysaccharide Extraction from Dendrobium officinale
2.2.2. Purification of DO Polysaccharides
2.3. Ultraviolet-Visible Spectroscopy (UV) Analysis
2.4. Molecular Weight Determination
2.5. FT-IR Analysis
2.6. Scanning Electron Microscopy (SEM)
2.7. Monosaccharide Analysis
2.8. Methylation Analysis
2.9. NMR Analysis
2.10. Animal Experimental Design
2.11. Constipation-Relieving Activity Test
2.12. Hypolipidemic Activity Test
2.12.1. In Vivo Lipid-Lowering Activity in Zebrafish
2.12.2. In Vitro Pancreatic Cholesterol Esterase (PCE) and Pancreatic Lipase (PL) Activity Test
2.13. Statistical Analysis
3. Results and Discussion
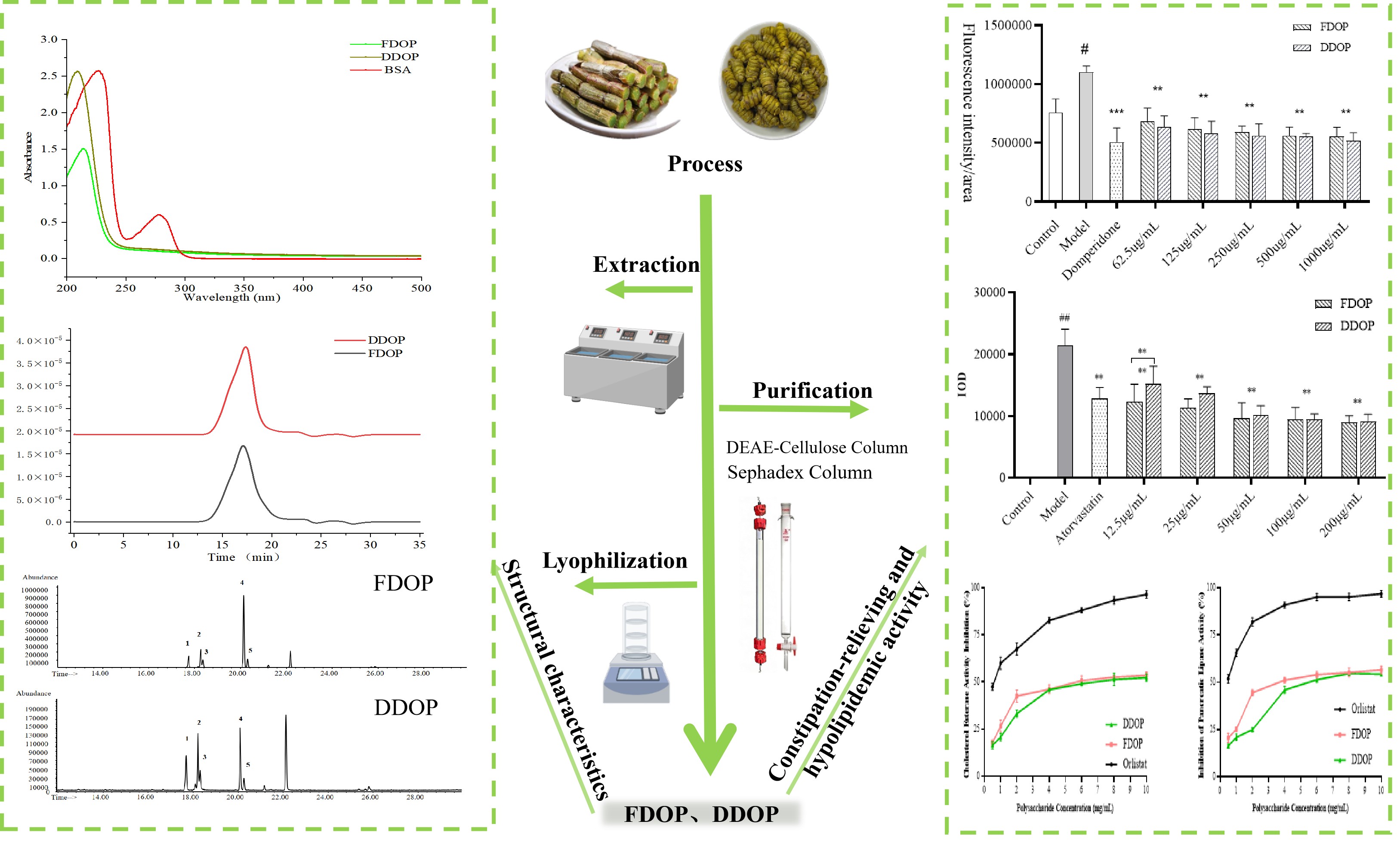
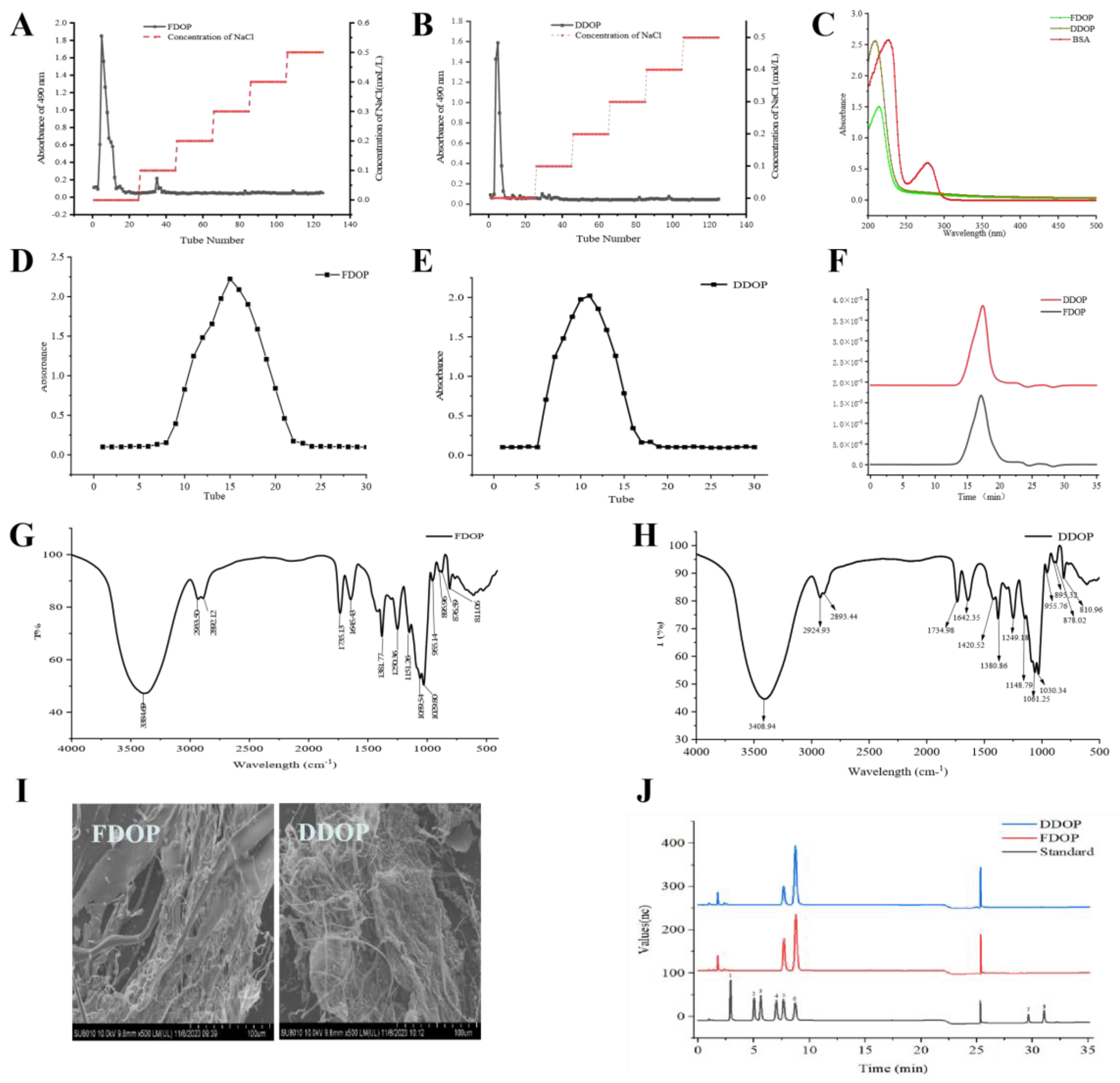
3.1. Isolation and Purification of Polysaccharides
3.2. UV Analysis
3.3. Molecular Weight Analysis
3.4. FT-IR Analysis
3.5. SEM
3.6. Analysis of Monosaccharide Composition
3.7. Methylation Analysis
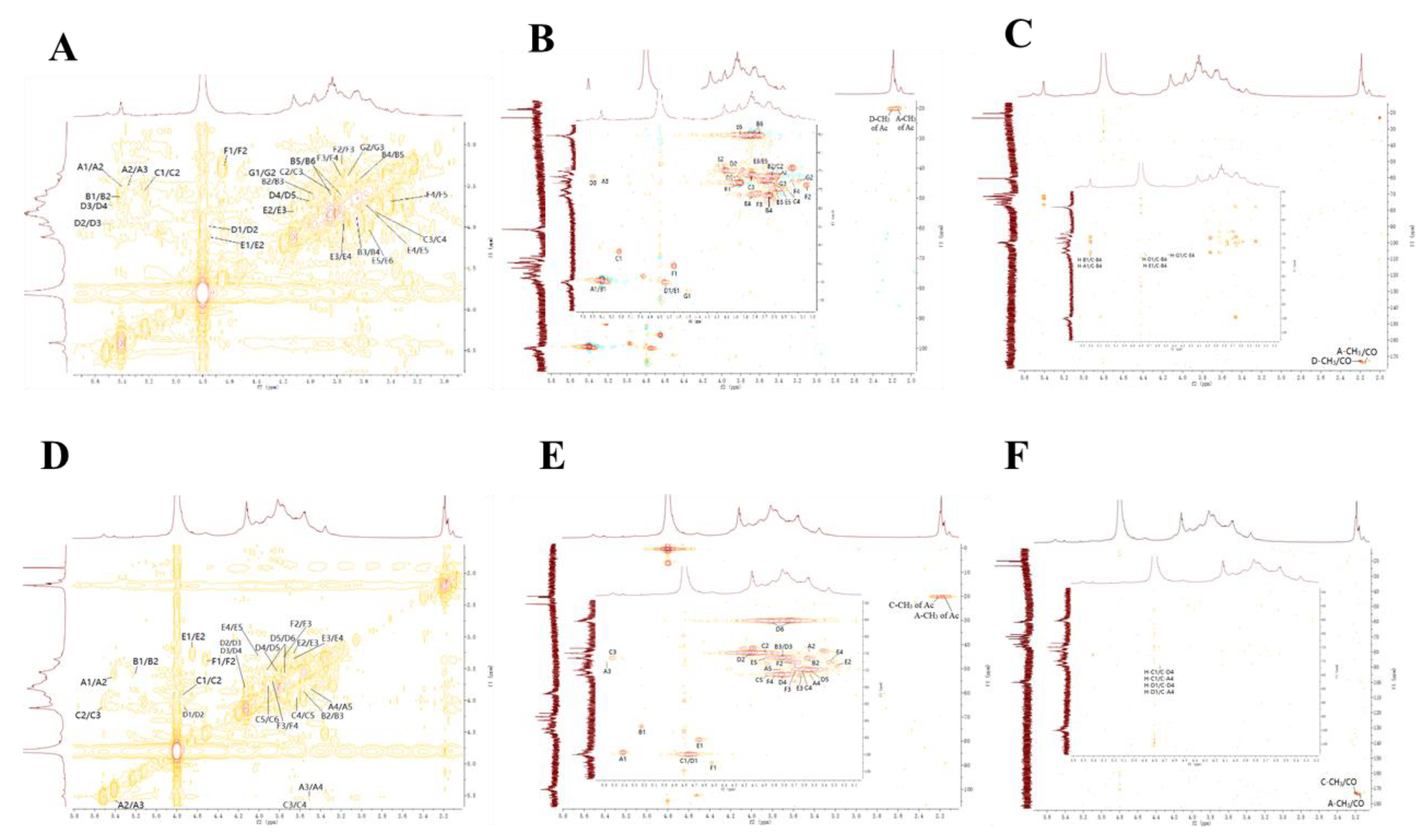
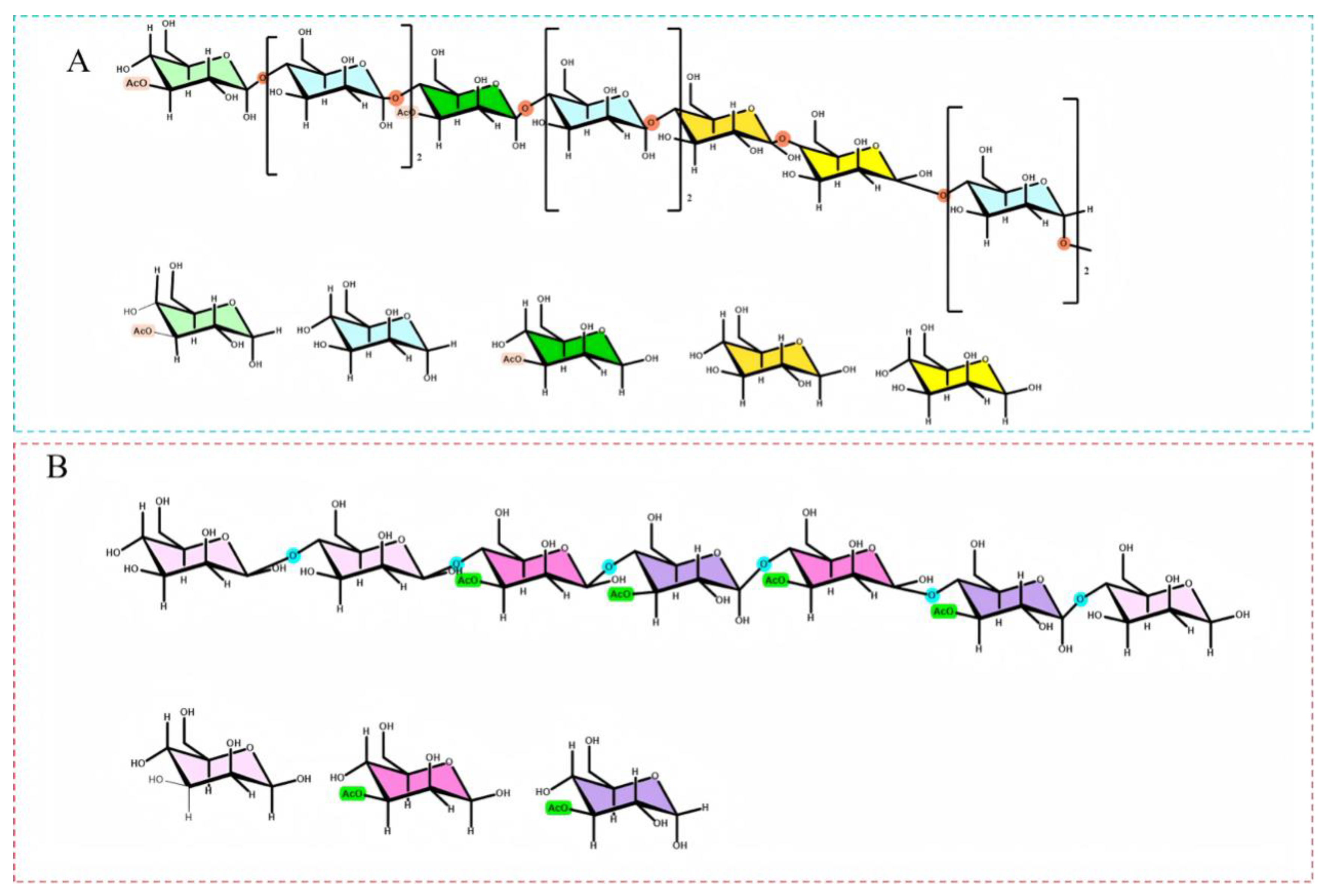
3.8. NMR Analysis
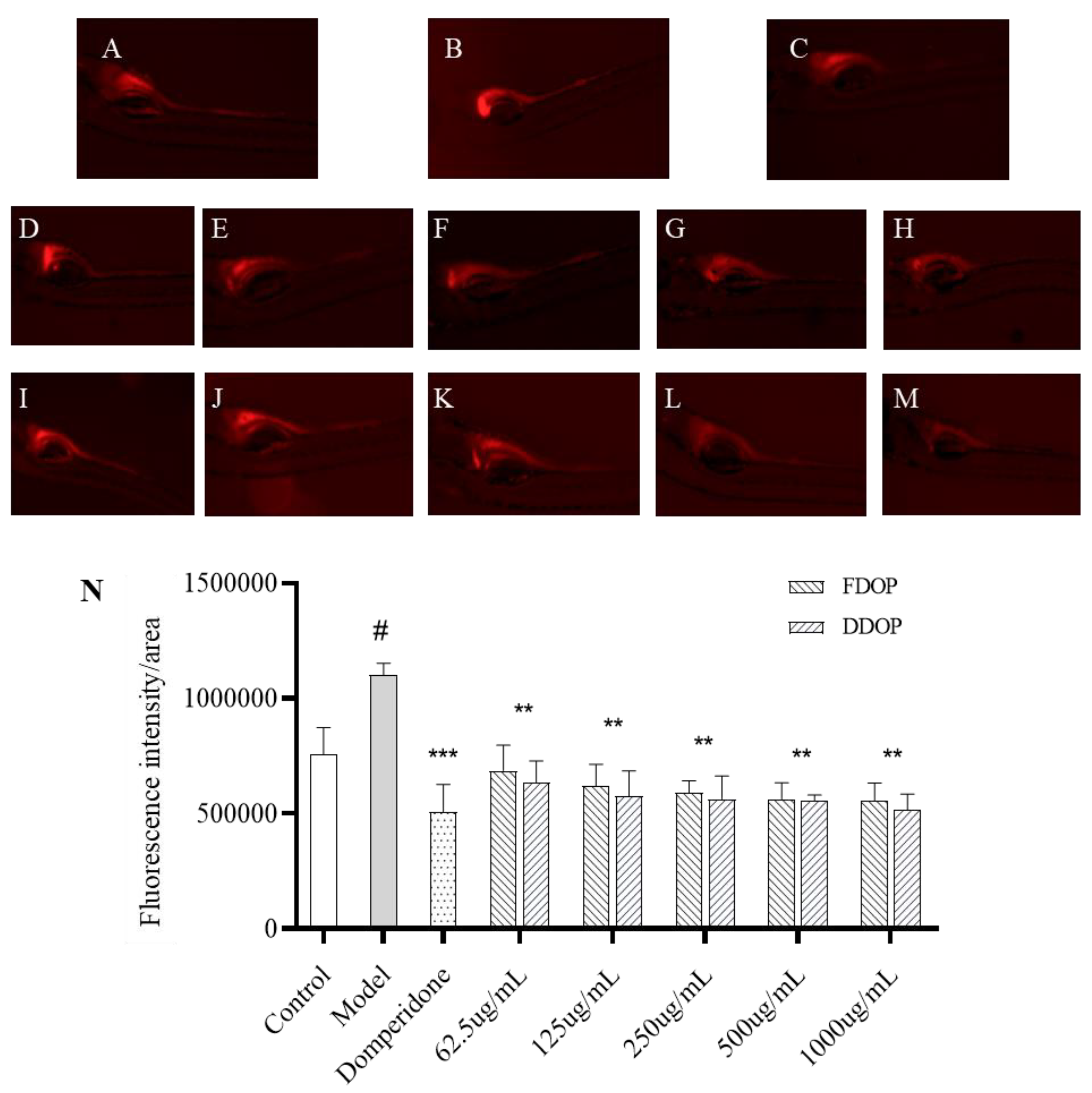
3.9. Evaluation of Constipation-Relieving Activity
3.10. Evaluation of Hypolipidemic Activity
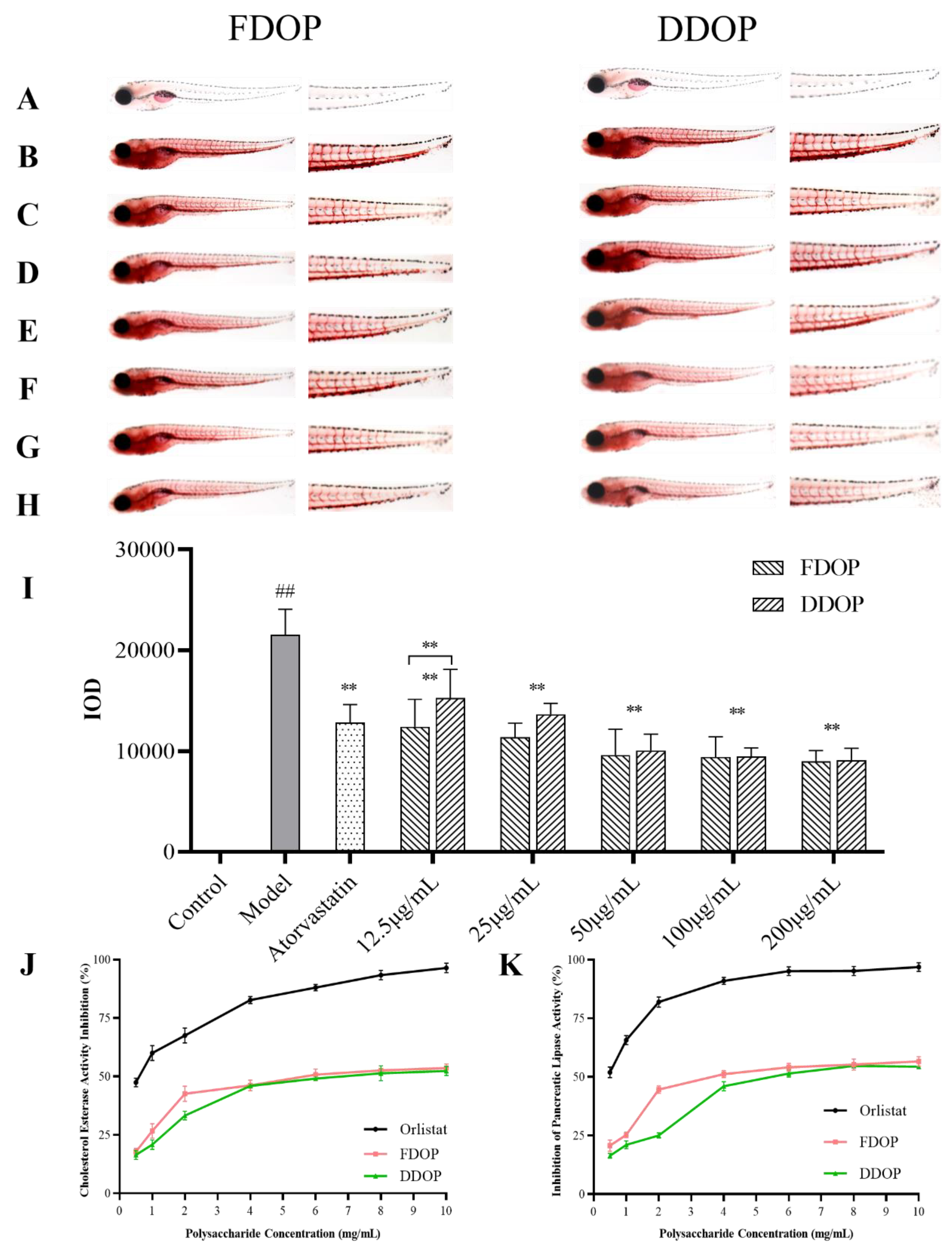
3.10.1. In Vivo Lipid-Lowering Activity Test
3.10.2. In Vitro Activity Assay
4. Conclusions
Supplementary Materials
Author Contributions
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bao, H.; Bao, H.; Wang, Y.; Wang, F.; Jiang, Q.; Li, H.; Ding, Y.; Zhu, C. Variations in cold resistance and contents of bioactive compounds among Dendrobium officinale Kimura et Migo Strains. Foods 2024, 13, 1467. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Fan, B.; Mu, Y.; Tong, L.; Lu, C.; Li, L.; Liu, J.; Sun, J.; Wang, F. Plant-Wide target metabolomics provides a novel interpretation of the changes in chemical components during Dendrobium officinale traditional processing. Antioxidants 2023, 12, 1995. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiang, Y.; Liu, Y.; Niu, Z.; Xue, Q.; Liu, W.; Ding, X. The large single-copy (LSC) region functions as a highly effective and efficient molecular marker for accurate authentication of medicinal Dendrobium species. Acta Pharm Sin B 2020, 10, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; He, Z.; Dou, P.; Wang, K.; Zhang, Y. Study on the intestinal metabolism and absorption of polysaccharides from Dendrobium officinale. Int. J. Biol. Macromol. 2025, 331, 148390. [Google Scholar] [CrossRef]
- Bu, Y.; Yin, B.; Qiu, Z.; Li, L.; Zhang, B.; Zheng, Z.; Li, M. Structural characterization and antioxidant activities of polysaccharides extracted from Polygonati rhizoma pomace. Food Chem X 2024, 23, 101778. [Google Scholar] [CrossRef]
- Cai, G.; Dong, H.; Liu, S.; Wu, W.; Yang, H. Comparative evaluation of the physiochemical properties, and antioxidant and hypoglycemic activities of Dendrobium officinale leaves processed using different drying techniques. Antioxidants 2023, 12, 1911. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, J.; Jin, H.; Gu, D.; Wang, Q.; Liu, Y.; Zan, K.; Fan, J.; Wang, R.; Wei, F.; Ma, S. Comparisons of physicochemical features and hepatoprotective potentials of unprocessed and processed polysaccharides from Polygonum multiflorum Thunb. Int. J. Biol. Macromol. 2023, 235, 123901. [Google Scholar] [CrossRef]
- Lu, M.; Chang, C.; Chao, C.; Hsu, Y. Structural changes, and anti-inflammatory, anti-cancer potential of polysaccharides from multiple processing of Rehmannia glutinosa. Int. J. Biol. Macromol. 2022, 206, 621–632. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, X.; Zhu, X.; Hua, Y. Chemical constituents, bioactivities, and pharmacological mechanisms of Dendrobium Officinale: A review of the past decade. J. Agric. Food Chem. 2023, 71, 14870–14889. [Google Scholar] [CrossRef]
- Wang, H.; Jin, H.; Dong, Y.; Wang, Z.; Wang, Y.; Wei, F. Structural characterization of Dendrobium huoshanense polysaccharides and its gastroprotective effect on acetic acid-induced gastric ulcer in mice. Int. J. Biol. Macromol. 2025, 311, 143361. [Google Scholar] [CrossRef]
- Cheng, H.; Xu, L.; Zhu, H.; Bu, T.; Li, Z.; Zhao, S.; Yang, K.; Sun, P.; Cai, M. Structural characterization of oligosaccharide from Dendrobium Officinale and its properties in vitro digestion and fecal fermentation. Food Chem. 2024, 460, 140511. [Google Scholar] [CrossRef]
- Fang, J.; Lin, Y.; Xie, H.; Farag, M.A.; Feng, S.; Li, J.; Shao, P. Dendrobium Officinale leaf polysaccharides ameliorated hyperglycemia and promoted gut bacterial associated SCFAs to alleviate type 2 diabetes in adult mice. Food Chem X 2022, 13, 100207. [Google Scholar]
- Zeng, H.; He, S.; Xiong, Z.; Su, J.; Wang, Y.; Zheng, B.; Zhang, Y. Gut microbiota-metabolic axis insight into the hyperlipidemic effect of lotus seed resistant starch in hyperlipidemic mice. Carbohydr. Polym. 2023, 314, 120939. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xu, S.; Li, B.; Zhang, B.; Chen, W.; Dai, D.; Liu, Z. Gut indigenous Ruminococcus gnavus alleviates constipation and stress-related behaviors in mice with loperamide-induced constipation. Food Funct. 2023, 14, 5702–5715. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Gao, M.; Ouyang, Z.; Li, Y.; Liu, D.; Zhu, M.; Sun, H.; Wang, J.; Chen, J.; et al. Research progress on the mechanism of action and screening methods of probiotics for lowering blood lipid levels. Foods 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Huang, J.; He, H.; Ju, X.; Ji, Y.; Deng, F.; Wang, Z.; He, R. Study on the hypolipidemic activity of rapeseed protein-derived peptides. Food Chem. 2023, 423, 136315. [Google Scholar] [CrossRef]
- Cao, Y.; Xu, F.; Xia, Q.; Liu, K.; Lin, H.; Zhang, S.; Zhang, Y. The peptide lltragl derived from Rapana venosa exerts protective effect against inflammatory bowel disease in zebrafish model by regulating multi-pathways. Mar. Drugs 2024, 22. [Google Scholar] [CrossRef]
- Kwiatkowska, I.; Hermanowicz, J.M.; Iwinska, Z.; Kowalczuk, K.; Iwanowska, J.; Pawlak, D. Zebrafish—An optimal model in experimental oncology. Molecules 2022, 27, 4223. [Google Scholar] [CrossRef]
- Lachowicz, J.; Szopa, A.; Ignatiuk, K.; Świąder, K.; Serefko, A. Zebrafish as an animal model in cannabinoid research. Int. J. Mol. Sci. 2023, 24, 10455. [Google Scholar] [CrossRef]
- Wu, X.; Li, W.; Li, S.; Zhu, S.; Pan, F.; Gu, Q.; Song, D. Hypolipidemic effect of polysaccharide from Sargassum fusiforme and its ultrasonic degraded polysaccharide on zebrafish fed high-fat diet. Int. J. Biol. Macromol. 2024, 276, 133771. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, Y.; Long, F.; Yang, D.; Huang, Y.; Han, Y.; Wu, Y.; Zhong, K.; Bu, Q.; Gao, H.; Huang, Y. Insight into structural characteristics of theabrownin from Pingwu Fuzhuan brick tea and its hypolipidemic activity based on the in vivo zebrafish and in vitro lipid digestion and absorption models. Food Chem. 2023, 404, 134382. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Tao, W.; Wang, M.; Liu, W.; Xing, J.; Yang, Y. Dendrobium Officinale Xianhu 2 polysaccharide helps forming a healthy gut microbiota and improving host immune system: An in vitro and in vivo study. Food Chem. 2023, 401. [Google Scholar] [CrossRef]
- Wu, Y.; Huo, Y.; Xu, L.; Xu, Y.; Wang, X.; Zhou, T. Purification, characterization and antioxidant activity of polysaccharides from Porphyra haitanensis. Int. J. Biol. Macromol. 2020, 165, 2116–2125. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wu, Z.; Li, M.; Cao, B.; Li, J.; Jiang, J.; Liu, H.; Zhang, Q.; Zhang, S. TCP80-1, a new levan-neoseries fructan from Tupistra chinensis baker rhizomes alleviates ulcerative colitis induced by dextran sulfate sodium in Drosophila melanogaster model. Food Res. Int. 2025, 203, 115860. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xing, N.; Jiao, Y.; Li, J.; Wang, T.; Zhang, Q.; Hu, X.; Li, C.; Kuang, W. An arabinan from Citrus Grandis fruits alleviates ischemia/reperfusion-induced myocardial cell apoptosis via the nrf2/keap1 and ire1/grp78 signaling pathways. Carbohydr. Polym. 2025, 347, 122728. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, J.; Yi, H.; Zhang, Z.; Zhang, L. Screening of intestinal peristalsis-promoting probiotics based on a zebrafish model. Food Funct. 2019, 10, 2075–2082. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhong, K.; Bai, J.-R.; Wu, Y.-P.; Zhang, J.-Q.; Gao, H. The biochemical characteristics of a novel fermented loose tea by Eurotium cristatum (MF800948) and its hypolipidemic activity in a zebrafish model. LWT-Food SCI Technol. 2020, 117, 108629. [Google Scholar] [CrossRef]
- Long, X.; Hu, X.; Xiang, H.; Chen, S.; Li, L.; Qi, B.; Li, C.; Liu, S.; Yang, X. Structural characterization and hypolipidemic activity of Gracilaria Lemaneiformis polysaccharide and its degradation products. Food Chem X 2022, 14. [Google Scholar] [CrossRef]
- Chang, S.; Lei, X.; Xie, Q.; Zhang, M.; Zhang, Y.; Xi, J.; Duan, J.; Ge, J.; Nian, F. In vitro study on antioxidant and lipid-lowering activities of tobacco polysaccharides. Bioresour. Bioprocessing 2024, 11. [Google Scholar] [CrossRef]
- Goudar, G.; Sharma, P.; Janghu, S.; Longvah, T. Effect of processing on barley β-glucan content, its molecular weight and extractability. Int. J. Biol. Macromol. 2020, 162, 1204–1216. [Google Scholar] [CrossRef]
- Cai, M.; Zhu, H.; Xu, L.; Wang, J.; Xu, J.; Li, Z.; Yang, K.; Wu, J.; Sun, P. structure, anti-fatigue activity and regulation on gut microflora in vivo of ethanol-fractional polysaccharides from Dendrobium Officinale. Int. J. Biol. Macromol. 2023, 234, 123572. [Google Scholar] [CrossRef] [PubMed]
- Kuang, M.-T.; Li, J.-Y.; Yang, X.-B.; Yang, L.; Xu, J.-Y.; Yan, S.; Lv, Y.-F.; Ren, F.-C.; Hu, J.-M.; Zhou, J. Structural characterization and hypoglycemic effect via stimulating glucagon-like peptide-1 secretion of two polysaccharides from Dendrobium Officinale. Carbohydr. Polym. 2020, 241, 116326. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Tang, Z.; Zhang, X.; Zhong, Y.; Yao, S.; Wang, L.; Lin, C.; Luo, X. Chemical properties and antioxidant activity of a water-soluble polysaccharide from Dendrobium Officinale. Int. J. Biol. Macromol. 2016, 89, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, X.; Sun, X.; Han, R.; Yu, N.; Liang, J.; Zhou, A. Comparison of the antioxidant activities and polysaccharide characterization of fresh and dry Dendrobium Officinale Kimura et Migo. Molecules 2022, 27, 6654. [Google Scholar] [CrossRef]
- Sims, I.M.; Carnachan, S.M.; Bell, T.J.; Hinkley, S.F.R. Methylation analysis of polysaccharides: Technical Advice. Carbohydr. Polym. 2018, 188, 1–7. [Google Scholar] [CrossRef]
- Liang, J.; Huang, Y.; Zhu, X.; Zhou, F.; Wu, T.; Jia, J.; Liu, X.; Kuang, H.; Xia, Y. Structural identification, rheological properties and immunological receptor of a complex galacturonoglucan from fruits of Schisandra chinensis (Turcz.) Baill. Carbohydr. Polym. 2024, 346, 122644. [Google Scholar] [CrossRef]
- Ciucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydrate Research 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Zhu, H.; Yi, X.; Jia, S.; Liu, C.; Han, Z.; Han, B.; Jiang, G.; Ding, Z.; Wang, R.; Lv, G. Optimization of three extraction methods and their effect on the structure and antioxidant activity of polysaccharides in Dendrobium huoshanense. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, H.; Fan, J.; Jia, S.; Yi, X.; Han, Z.; Wang, R.; Qiu, H.; Lv, G. Study on differences in structure and anti-inflammatory activity of polysaccharides in five species of Dendrobium. Polymers 2025, 17, 1164. [Google Scholar] [CrossRef]
- He, Y.; Li, L.; Chang, H.; Cai, B.; Gao, H.; Chen, G.; Hou, W.; Jappar, Z.; Yan, Y. Research progress on extraction, purification, structure and biological activity of Dendrobium officinale polysaccharides. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef]
- Butt, H.S.; Ulriksen, E.S.; Rise, F.; Wangensteen, H.; Duus, J.Ø.; Inngjerdingen, M.; Inngjerdingen, K.T. Structural elucidation of novel pro-inflammatory polysaccharides from Daphne mezereum L. Carbohydr. Polym. 2024, 324, 121554. [Google Scholar]
- Zeng, F.; Chen, W.; He, P.; Zhan, Q.; Wang, Q.; Wu, H.; Zhang, M. Structural characterization of polysaccharides with potential antioxidant and immunomodulatory activities from Chinese water chestnut peels. Carbohydr. Polym. 2020, 246, 116551. [Google Scholar] [CrossRef] [PubMed]
- Bi, Z.; Zhao, Y.; Hu, J.; Ding, J.; Yang, P.; Liu, Y.; Lu, Y.; Jin, Y.; Tang, H.; Liu, Y.; et al. A Novel Polysaccharide from Lonicerae Japonicae Caulis: Characterization and Effects on the Function of Fibroblast-like Synoviocytes. Carbohydr. Polym. 2022, 292, 119674. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Cheong, K.L.; Song, Z.; Shi, Y.; Huang, X. Structure and protective effect on UVB-induced keratinocyte damage of fructan from white garlic. Carbohydr. Polym. 2013, 92, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Li, J.; Zhang, J.; Liu, H.; Ye, Q.; Wang, Z. Structural characterization and antitumor activity of a polysaccharide from Dendrobium wardianum. Carbohydr. Polym. 2021, 269, 118253. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, Z.; Xue, X.; Luo, X.; Guo, Z.; Miao, S.; Zheng, B. Effect of fucoidan molecular weight on gut microbiota composition and anti-inflammatory activity after in vitro dynamic digestion and fermentation. Int. J. Biol. Macromol. 2025, 330, 147976. [Google Scholar] [CrossRef]
- Lai, X.; Xv, W.; Fu, S.; Li, S.; Fu, X.; Tan, C.; Wang, P.; Dou, Z.; Chen, C. The effect of the molecular weight of blackberry polysaccharides on gut microbiota modulation and hypoglycemic effect in vivo. Food Funct. 2024. [Google Scholar]
- Wang, Q.; Liang, J.; Liu, H. In Vitro Effects of four polysaccharides containing β-D-Glup on intestinal function. Int. J. Food Prop. 2019, 22, 1064–1076. [Google Scholar] [CrossRef]
- Anderson, J.W.; Siesel, A.E. Hypocholesterolemic effects of oat products. ADV EXP MED BIOL. 1990, 270, 17–36. [Google Scholar]
- Yin, J.; Nie, S.; Li, J.; Li, C.; Cui, S.W.; Xie, M. Mechanism of interactions between calcium and viscous polysaccharide from the seeds of Plantago asiatica L. J. Agric. Food Chem. 2012, 60, 7981–7987. [Google Scholar]
- Shen, H.; Wang, J.; Ao, J.; Ye, L.; Shi, Y.; Liu, Y.; Li, M.; Luo, A. The inhibitory mechanism of pentacyclic triterpenoid acids on pancreatic lipase and cholesterol esterase. Food Bioscience 2023, 51, 102341. [Google Scholar] [CrossRef]
- Chen, H.; Nie, Q.; Hu, J.; Huang, X.; Huang, W.; Nie, S. Metabolism amelioration of Dendrobium Officinale polysaccharide on type II diabetic rats. Food Hydrocoll. 2020, 102, 105582. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Y.; Zang, X.; Yang, Y.; Wang, W.; Zhang, J.; Que, Y.; Liang, F.; Wang, T.; Zhang, J.; Ma, H.; Guan, L. Physicochemical properties and fermentation characteristics of a novel polysaccharide degraded from Flammulina velutipes residues polysaccharide. Food Chem X 2024, 24, 102049. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, M.; Zhang, S.; Wang, Y.; Jiang, X. Structural characterization of polysaccharides from Cordyceps militaris and their hypolipidemic effects in high fat diet fed mice. RSC Adv. 2018, 8, 41012–41022. [Google Scholar] [CrossRef]
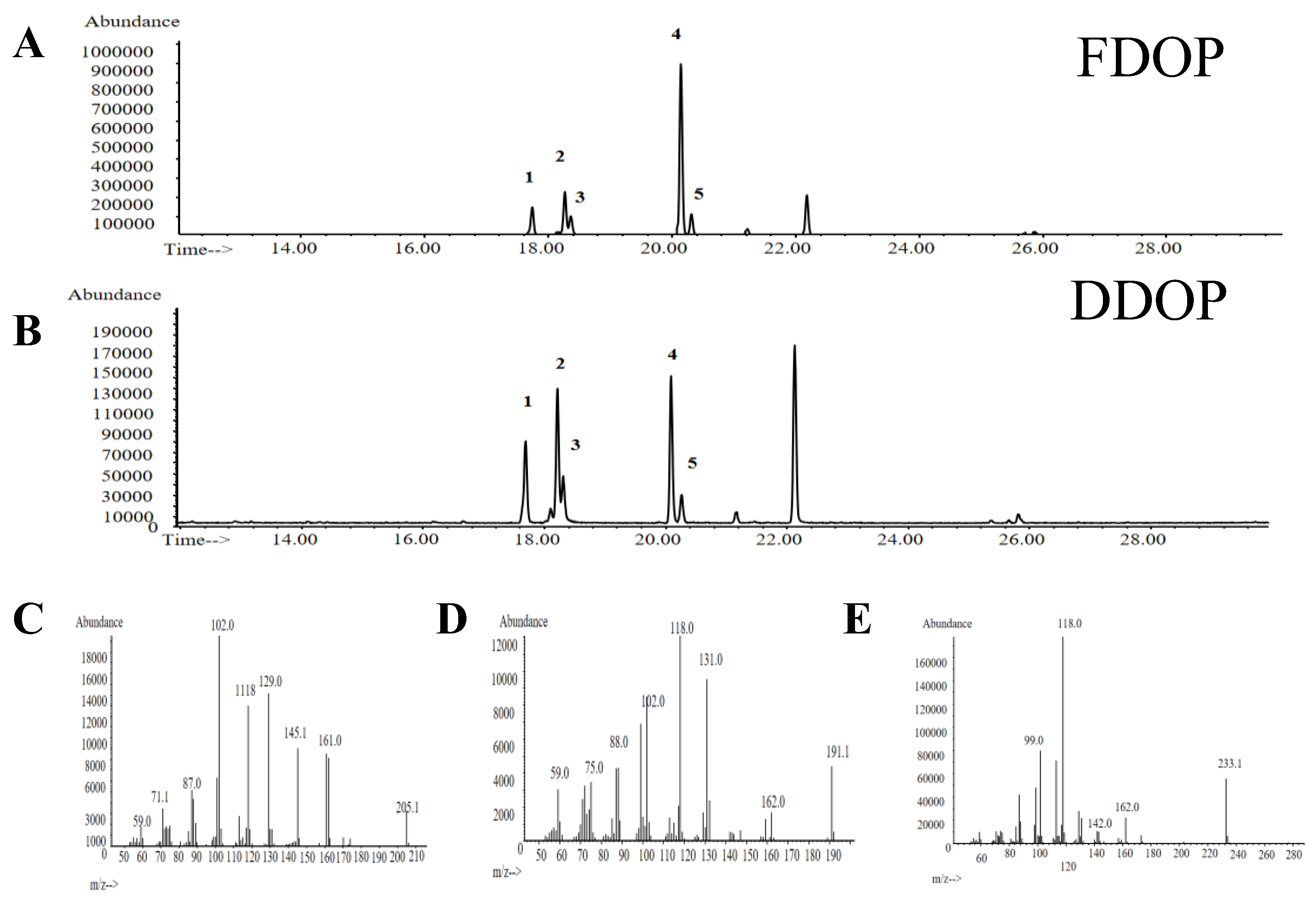

| No. | Methylation debris | Majorfragments (m/z) | Types of linkages | Molecular ratio % | |
| FDOP | DDOP | ||||
| 1 | 1,5-di-O-acetyl-(1-deuterio)-2,3,4,6-tetra-O-methyl glucitol | 59.0,71.7,87.0,102.0,111.8,129.0,145.1,161.0,205.1 | t-Glcp | 11.1 | 20.9 |
| 2 | 1,5-tri-O-acetyl-(1-deuterio)-2,3,6-tri-O-methyl-4-hydroxyl-manitol | 59.0,75.0,88.0,102.0,118.0,131.0,162.0,191.1 | 1,4-Manp(3-O-Ac) | 15.9 | 30.3 |
| 3 | 1,5-tri-O-acetyl-(1-deuterio)-2,3,6-tri-O-methyl-4-hydroxyl-glcitol | 59.0,75.0,88.0,102.0,118.0,131.0,162.0,191.1 | 1,4-Glcp(3-O-Ac) | 7.9 | 11.7 |
| 4 | 1,4,5-tri-O-acetyl-(1-deuterio)-2,3,6-tri-O-methyl-manitol | 99.0,118.0,142.0,162.0,233.1 | 1,4-Manp | 57.1 | 30.4 |
| 5 | 1,4,5-tri-O-acetyl-(1-deuterio)-2,3,6-tri-O-methyl-glucitol | 99.0,118.0,142.0,162.0,233.1 | 1,4-Glcp | 8.1 | 6.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



