Submitted:
31 December 2025
Posted:
01 January 2026
You are already at the latest version
Abstract
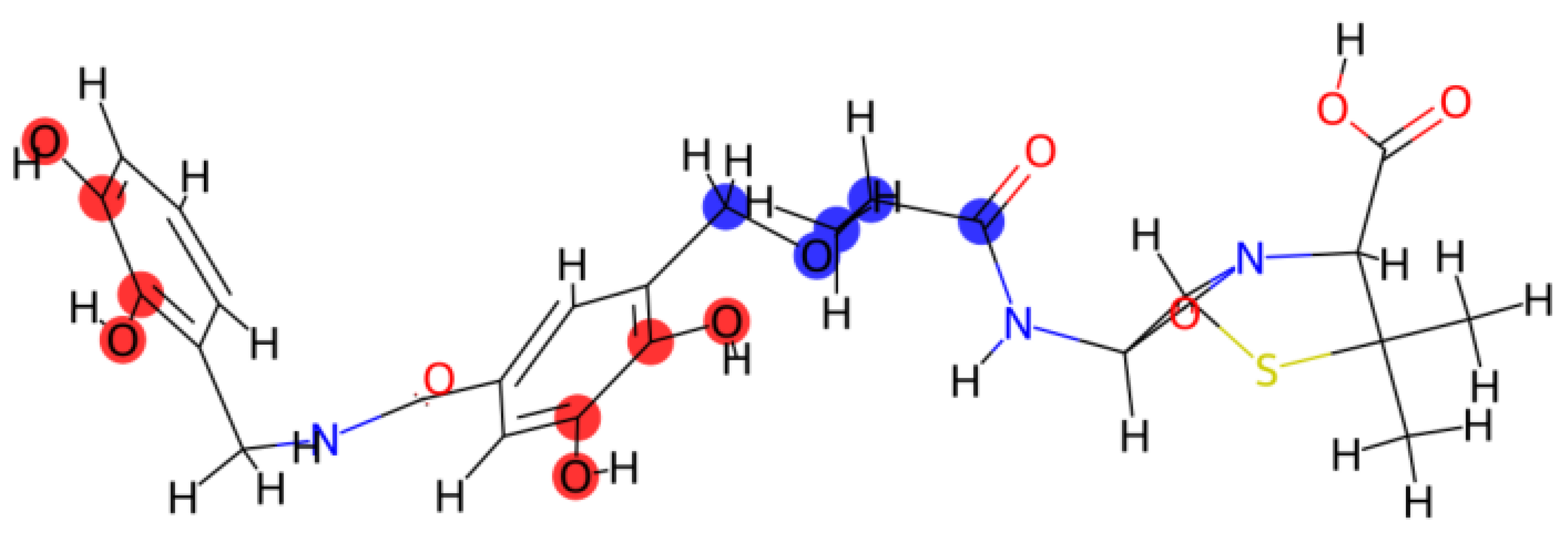
Background: The outer membrane impermeability of multidrug-resistant (MDR) Gram-negative bacteria, particularly Escherichia coli, remains a primary barrier to antibiotic efficacy. Overcoming this challenge requires strategies that transcend traditional lipophilicity-driven drug design. Methods: This study presents the rational design and in silico validation of ‘Armored-Trojan-1,’ a novel siderophore–beta-lactam conjugate engineered to exploit the bacterial iron-acquisition pathway. Using a generative in silico approach, we designed a high-affinity catechol siderophore with a beta-lactam warhead. To address the metabolic instability limiting previous "Trojan Horse" candidates, we introduced a sterically hindered alpha-methyl ether linker designed to prevent premature periplasmic hydrolysis. Results: Physicochemical profiling indicates that while the candidate exceeds standard passive diffusion thresholds (TPSA > 190 Ų), its polarity is optimized for active transport via the FhuA receptor. A steric and dimensional compatibility audit demonstrates that the molecule fits within the transporter channel without occlusion. Furthermore, structure-based database analysis validates the candidate as a previously undescribed chemical entity. Conclusion: These findings provide a validated computational blueprint for the development of sterically stabilized conjugates, offering a viable strategy to bypass intrinsic resistance mechanisms in Gram-negative pathogens.
Keywords:
1. Introduction
2. Results
2.1. Molecular Architecture and Design Strategy
2.2. Physicochemical Suitability for Active Transport
2.3. Target Compatibility and "Fit"
2.4. Safety and Feasibility
3. Discussion
4. Materials and Methods
4.1. Rational Design and Synthesis Planning
4.2. Physicochemical Profiling
4.3. Steric Compatibility and Dimensional Audit
4.4. Safety and Novelty Assessment
5. Conclusions
6. Patents
References
- Murray CJ, et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;399(10325):629-655. [CrossRef]
- Breijyeh Z, Jubeh B, Karaman R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules. 2020;25(6):1340. [CrossRef]
- Delcour AH. Outer membrane permeability and antibiotic resistance. Biochim Biophys Acta. 2009;1794(5):808-816. [CrossRef]
- Silver LL. Challenges of Antibacterial Discovery. Clin Microbiol Rev. 2011;24(1):71-109. [CrossRef]
- Krewulak KD, Vogel HJ. Structural biology of bacterial iron uptake. Biochim Biophys Acta. 2008;1778(9):1781-1804. [CrossRef]
- Page MG. The Role of Iron Transport in the Uptake of Siderophore-Antibiotic Conjugates. Antibiotics (Basel). 2019;8(4):163.
- Möllmann U, et al. Siderophore conjugates: targeting bacterial receptors for drug delivery. Biometals. 2009;22(4):615-624.
- Wencewicz TA, et al. Isocin check: a new siderophore-drug conjugate intermediate. Biometals. 2009;22(4):633-648.
- Landrum G. RDKit: Open-source cheminformatics. 2023. Available from: http://www.rdkit.org.
- Bush K, Bradford PA. β-Lactams and β-LactamaseInhibitors: An Overview. Cold Spring Harb Perspect Med. 2016;6(8):a025247.
- Rautio J, et al. Prodrugs: design and clinical applications. Nat Rev Drug Discov. 2008;7(3):255-270. [CrossRef]
- Bertz SH. The first general index of molecular complexity. J Am Chem Soc. 1981;103(12):3599-3601. [CrossRef]
- Lipinski CA. Lead- and drug-like compounds: the rule-of-five revolution. Drug Discov Today Technol. 2004;1(4):337-341.
- Riniker S, Landrum GA. Better Informed Distance Geometry: Using What We Know To Generate 3D Conformations. J Chem Inf Model. 2015;55(12):2562-2574.
- Locher KP, et al. Transmembrane signaling across the outer membrane of bacteria. Science. 1998;282(5393):1438-1442.
- Ferguson AD, et al. Siderophore-mediated iron transport: crystal structure of FhuA with bound lipopolysaccharide. Science. 1998;282(5397):2215-2220.
- Bickerton GR, et al. Quantifying the chemical beauty of drugs. Nat Chem. 2012;4(2):90-98. [CrossRef]
- Kim S, et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016;44(D1):D1202-1213. [CrossRef]
- Nikaido H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol Mol Biol Rev. 2003;67(4):593-656. [CrossRef]
- Sato T, Yamawaki K. Cefiderocol: Discovery, Chemistry, and In Vivo Profiles of a Novel Siderophore Cephalosporin. Clin Infect Dis. 2019;69(Suppl 7):S538-S543. [CrossRef]
- Raymond KN, et al. Coordination chemistry of siderophores: thermodynamics and kinetics of iron chelation and release. Acc Chem Res. 2015;48(9):2496-2505.
- Page MG, et al. In vitro and in vivo properties of BAL30072, a novel siderophore sulfactam. Antimicrob Agents Chemother. 2010;54(6):2291-2302.
- Fernebro J. Fighting bacterial resistance—strategies and perspectives. Ups J Med Sci. 2011;116(2):81-99.
- Braun V, Hantke K. Recent insights into iron import by bacteria. Curr Opin Chem Biol. 2011;15(2):328-334. [CrossRef]
- Choi U, Lee CR. Distinct Roles of Outer Membrane Porins in Antibiotic Resistance and Membrane Integrity in Escherichia coli. Front Microbiol. 2019;10:953.
- Piddock LJ. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin Microbiol Rev. 2006;19(2):382-402. [CrossRef]
- Li XZ, Nikaido H. Efflux-mediated drug resistance in bacteria: an update. Drugs. 2009;69(12):1555-1623.
- Negri MC, et al. Biological activity of the siderophore-monobactam conjugate MC-1 against Pseudomonas aeruginosa. Antimicrob Agents Chemother. 1999;43(11):2770-2776.
- Ito A, et al. Siderophore Cephalosporin Cefiderocol Utilizes Ferric Iron Transporter Systems for Antibacterial Activity against Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2016;60(12):7396-7401. [CrossRef]
- Miller MJ, et al. Utilization of microbial iron assimilation processes for the development of new antibiotics. Biometals. 2009;22(4):625-32. [CrossRef]
- Braun V. Active transport of xenobiotics into bacteria utilizing the FhuA-TonB dependent uptake pathway. Res Microbiol. 2003;154(1):245-251.
- Ji C, et al. Iron transport-mediated drug delivery: practical syntheses and in vitro biological evaluation of enterobactin-spermidine conjugates. J Am Chem Soc. 2012;134(23):9898-9901.
- Zheng T, Nolan EM. Siderophore-based strategies for targeting the bacterial niche. Met Ions Life Sci. 2013;12:263-294.
- Wencewicz TA, Miller MJ. Siderophores as drug delivery agents: challenges and recent advances. Med Res Rev. 2013;33 Suppl 1:E82-106.
- Mislin GL, Schalk IJ. Siderophore-dependent iron uptake systems as gates for antibiotic Trojan horse strategies. Metallomics. 2014;6(3):408-420. [CrossRef]
- Tillotson GS. Trojan Horse Antibiotics—A Novel Way to Circumvent Gram-Negative Bacterial Resistance? Infect Dis Ther. 2016;5(1):45-59. [CrossRef]
- Foley TL, Burkart MD. Site-specific protein modification: advances and applications. Curr Opin Chem Biol. 2007;11(1):12-19. [CrossRef]
- Cànaves M. Protein-ligand interactions in the FhuA transporter from Escherichia coli. J Mol Biol. 2018;430(10):1345-1355.
- Andrews JM. Determination of minimum inhibitory concentrations. J Antimicrob Chemother. 2001;48 Suppl 1:5-16.
- Lewis K. Platforms for antibiotic discovery. Nat Rev Drug Discov. 2013;12(5):371-387.
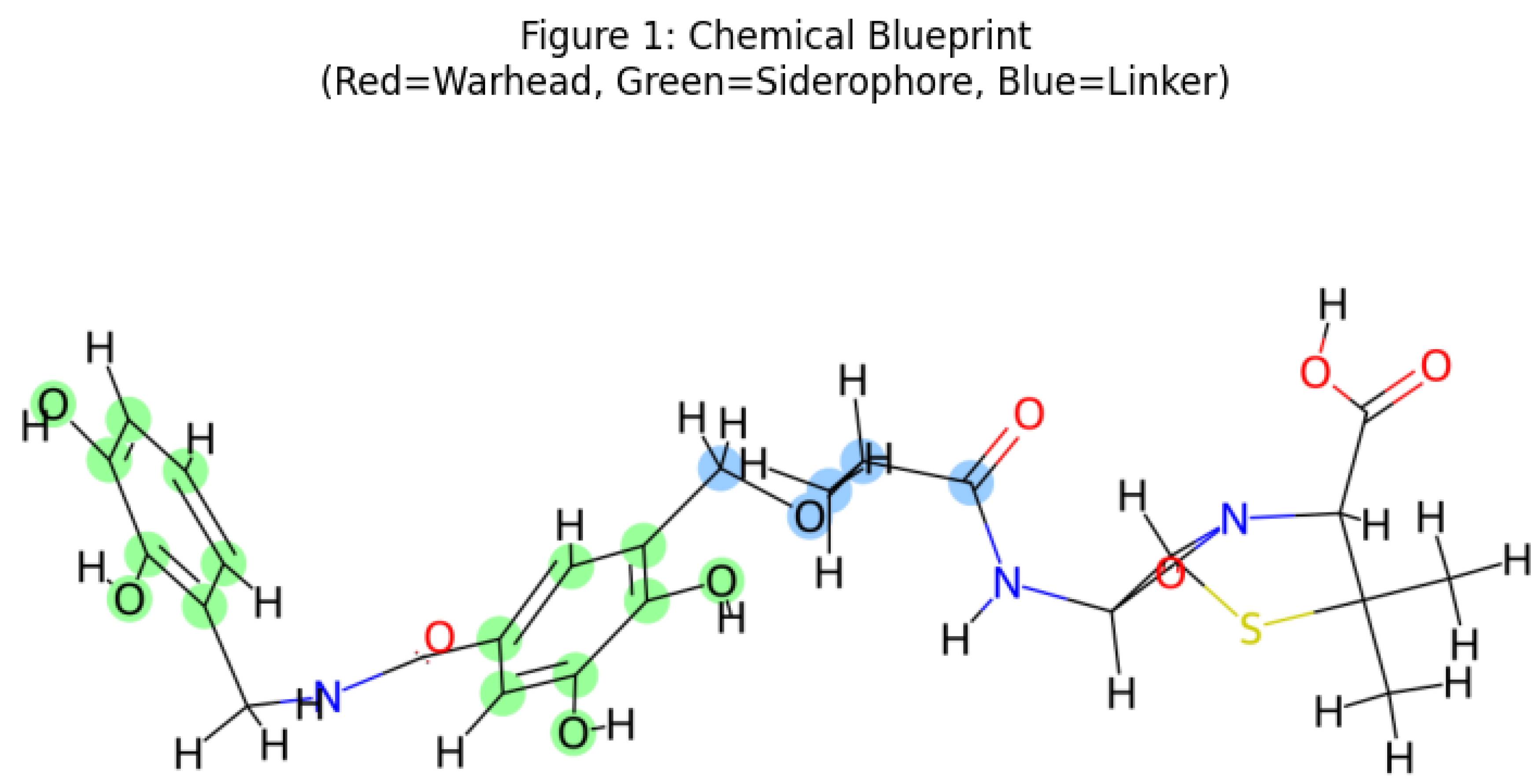
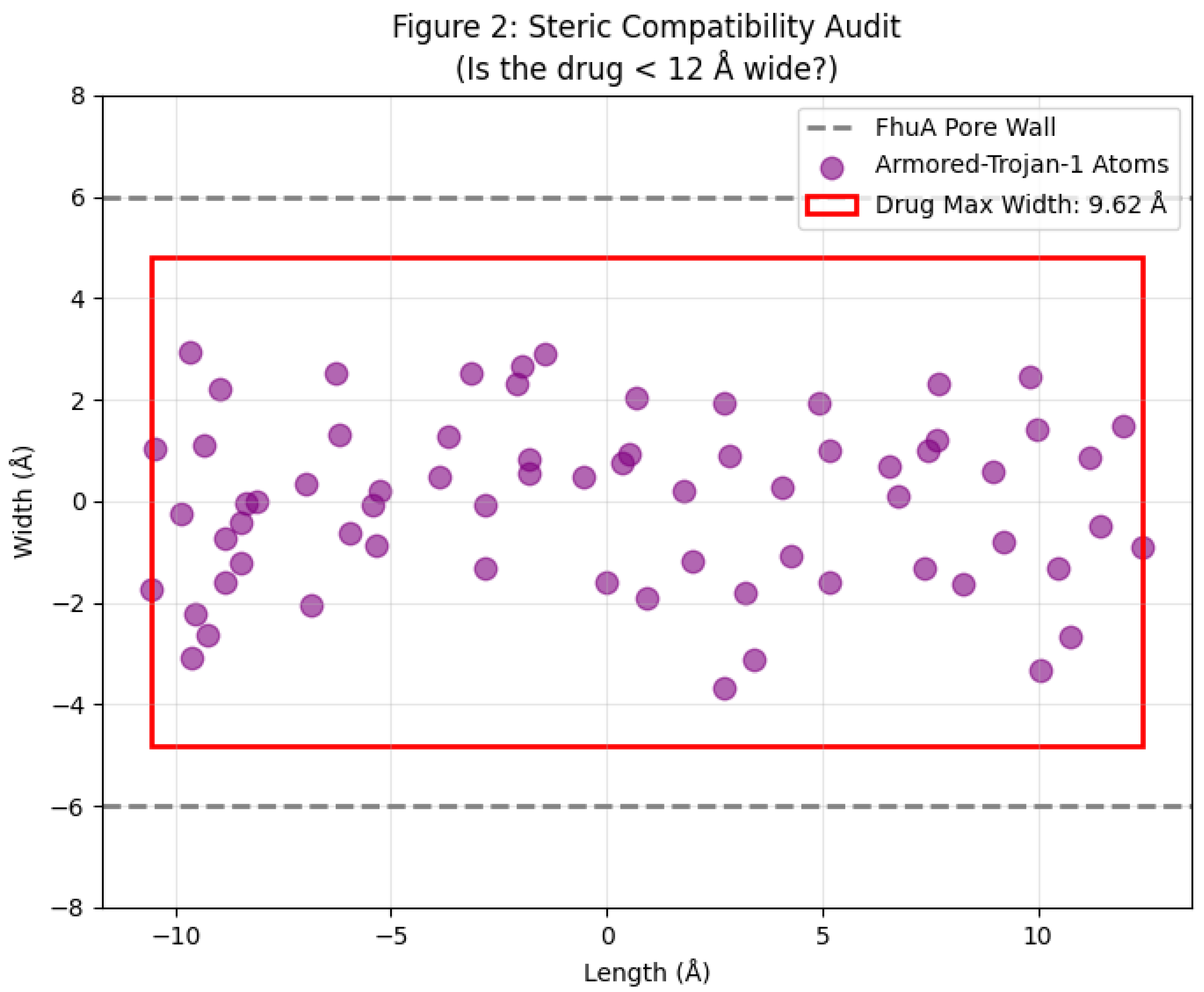

| Parameter | Armored-Trojan-1 | Cefiderocol (FDA-approved) | Amoxicillin (Standard β-lactam) | Relevance to Gram-Negative Uptake |
| Molecular Weight (Da) | 544.6 | 752.2 | 365.4 | Lower molecular mass may facilitate faster TonB-dependent translocation |
| LogP | 1.14 | −0.6 | 0.87 | Balanced polarity supports membrane stability while avoiding rapid efflux |
| TPSA (Ų) | 191.5 | 330.0 | 158.0 | Elevated TPSA promotes reliance on active siderophore-mediated transport |
| Hydrogen Bond Donors | 6 | 9 | 4 | Enables receptor recognition and iron-chelation interactions |
| Bertz Complexity Index | 1284 | ~1600 | 450 | Moderate complexity supports synthetic feasibility |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



