1. Introduction
Breast cancer remains a major global health challenge, with triple-negative breast cancer (TNBC) representing one of its most aggressive and problematic subtypes. Defined by the lack of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) expression, TNBC accounts for approximately 15–20% of all breast cancer cases and is characterized by a high risk of early recurrence, metastasis, and poor overall survival [
1,
2]. The absence of established molecular targets renders TNBC patients insensitive to endocrine therapy and HER2-targeted agents, leaving conventional chemotherapy as the primary systemic treatment option [
2]. However, the clinical efficacy of chemotherapy is often compromised by dose-limiting toxicities and the rapid development of drug resistance [
3]. Consequently, the discovery of novel, potent, and more selective lead compounds that can address the unique vulnerabilities of TNBC is an urgent and critically important pursuit in oncology drug discovery [
4].
Nature, particularly microbial sources, has been an invaluable reservoir of anticancer agents. Among these, actinomycetes of the genus
Streptomyces are exceptionally prolific, responsible for producing a vast array of clinically used therapeutics, including antitumor antibiotics such as bleomycin, actinomycin D, and mitomycin [
5,
6,
7]. The advent of genome mining has revolutionized natural product discovery, enabling the systematic exploration of 'silent' or poorly expressed biosynthetic gene clusters (BGCs) in
Streptomyces strains, thereby unveiling their hidden chemical potential [
8,
9]. In this context, the unique ecosystems of Yunnan Province in China, a recognized biodiversity hotspot, offer a largely untapped reservoir of microbial diversity. The distinctive low-latitude and high-altitude environments of this region are conducive to the evolution of unique actinomycete lineages, presenting significant opportunities for the discovery of novel microorganisms and their bioactive metabolites [
10,
11].
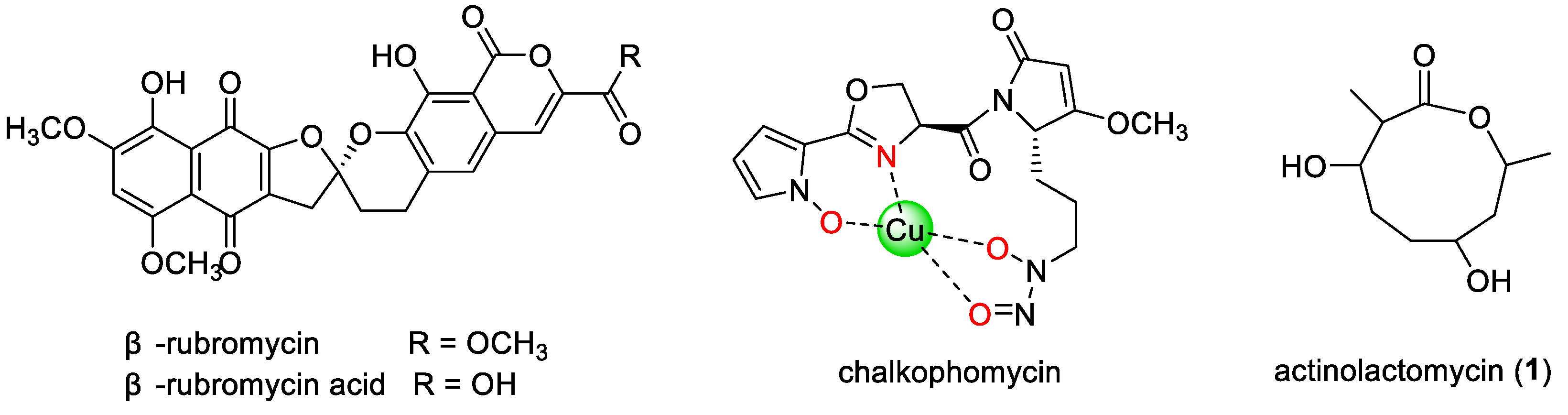
Streptomyces sp. CB00271, isolated from a pristine soil sample in Yunnan, has emerged as a particularly promising strain. Previous research has demonstrated its ability to produce the [
5,
6]-spiroketal-containing antitumor compound
β-rubromycin and a new derivative,
β-rubromycin acid (
Figure 1), with the latter exhibiting cytotoxicity across a panel of tumor cell lines [
12]. Subsequent optimization of fermentation conditions led to the discovery of chalkophomycin (
Figure 1), a structurally unprecedented copper-chelating metallophore that shows potent cytotoxicity against several cancer cell lines, significantly surpassing the efficacy of cisplatin [
13,
14]. Whole-genome sequencing of CB00271 further revealed an impressive biosynthetic capacity, harboring 45 BGCs, including 18 non-ribosomal peptide synthetase (NRPS) clusters, 6 NRPS–type I polyketide synthase (T1PKS) hybrid clusters, and 10 BGCs of unknown types, underscoring its potential as a prolific source of novel natural products [
15]. However, until now, actinolactomycin (ALM) had not been reported from this strain, nor had its potential against triple-negative breast cancer (TNBC) been systematically investigated.
Actinolactomycin (ALM,
Figure 1) is a structurally distinctive cyclic 2-oxonanonide natural product first reported from
Streptomyces flavoretus 18522 [
16]. It has demonstrated inhibitory activity against several human cancer cell lines (e.g., A2780, K562, HCT-15, A549, and HeLa) and has been shown to induce G
0/G
1 cell cycle arrest and apoptosis [
16,
17]. Mechanistic studies suggest that ALM exerts its effects by inhibiting the mammalian target of rapamycin (mTOR) signaling pathway and suppressing the translation of hypoxia-inducible factor 1α (HIF-1α) [
18]. Intriguingly, the mTOR–HIF-1α axis is critically implicated in the metabolic reprogramming, invasive phenotype maintenance, and therapy resistance of TNBC [
19,
20,
21], positioning ALM as a potential targeted therapeutic candidate for this aggressive subtype. Despite its promising bioactivity, ALM remains an exceedingly scarce natural product. To date, only four actinomycete strains capable of producing ALM have been reported worldwide, namely
Streptomyces flavoretus 18522 (from Yunnan soil, 2005) [
16], strain L39-3 (from Bohai Sea mud, 2010) [
17],
Streptomyces sp. K4 (from soil, 2024) [
22], and
Streptomyces pratensis TRM74023 (from rhizosphere soil of the Taklamakan Desert, 2025) [
21]. Its complete biosynthetic pathway also remains unelucidated, severely hindering further research and development. In this study, we report for the first time the isolation of actinolactomycin from
Streptomyces sp. CB00271 and provide a systematic evaluation of its potent and selective cytotoxicity against TNBC models, thereby introducing a new microbial source for this rare compound and highlighting its previously unreported anti-TNBC potential.
Given the pressing clinical challenge of TNBC and the scarcity of ALM, this study focused on the soil-derived Streptomyces sp. CB00271. We systematically investigated the secondary metabolites of this strain, leading to the isolation and structural elucidation of several compounds. The primary objective was to evaluate the antitumor activity of the rare component actinolactomycin against breast cancer models, with a particular emphasis on the refractory TNBC subtype. Our findings not only identify actinolactomycin as a highly potent anti-TNBC agent but also establish Streptomyces sp. CB00271 as a valuable microbial resource for this scarce compound, thereby providing a promising lead candidate and a solid foundation for the development of novel therapeutics against triple-negative breast cancer.
2. Results
2.1. Phylogenetic Analysis and Strain Identification of Streptomyces sp. CB00271
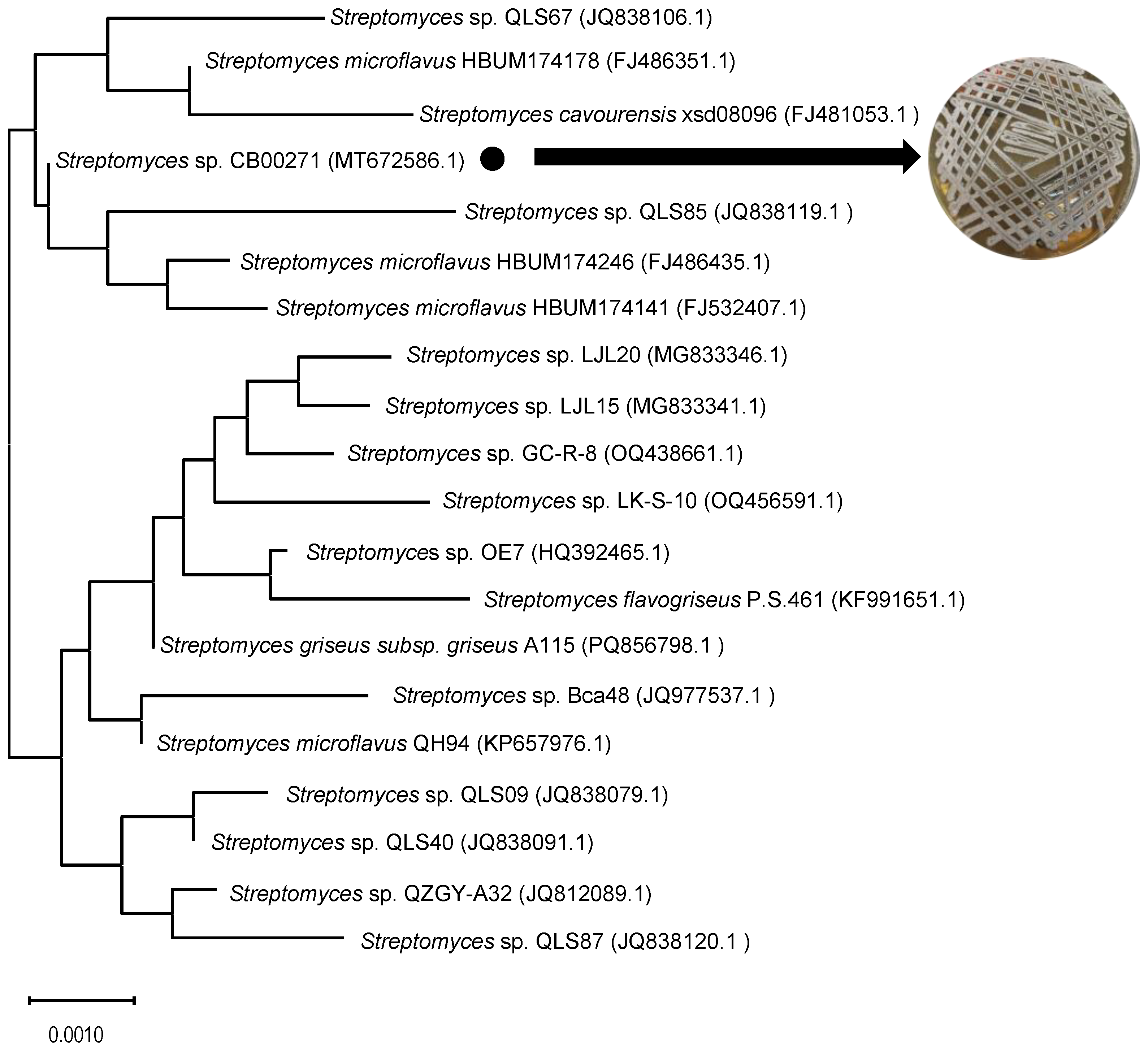
To confirm the taxonomic status of the soil-derived strain CB00271, we performed 16S rRNA gene sequencing and phylogenetic analysis. The nearly full-length 16S rRNA gene sequence (1,451 bp) was obtained and deposited in the GenBank database under accession number MT672586.1. BLAST analysis revealed that the sequence shared >99% identity with those of several known
Streptomyces species. A phylogenetic tree constructed using the neighbor-joining method demonstrated that strain CB00271 forms a stable clade with
Streptomyces sp. QLS85,
Streptomyces microfavus HBUM174246, and
Streptomyces microfavus HBUM174141 (
Figure 2), unambiguously assigning it to the genus
Streptomyces. Notably, this study identifies CB00271 as a novel microbial source of actinolactomycin-type natural products.
2.2. Isolation and Structural Elucidation of Secondary Metabolites
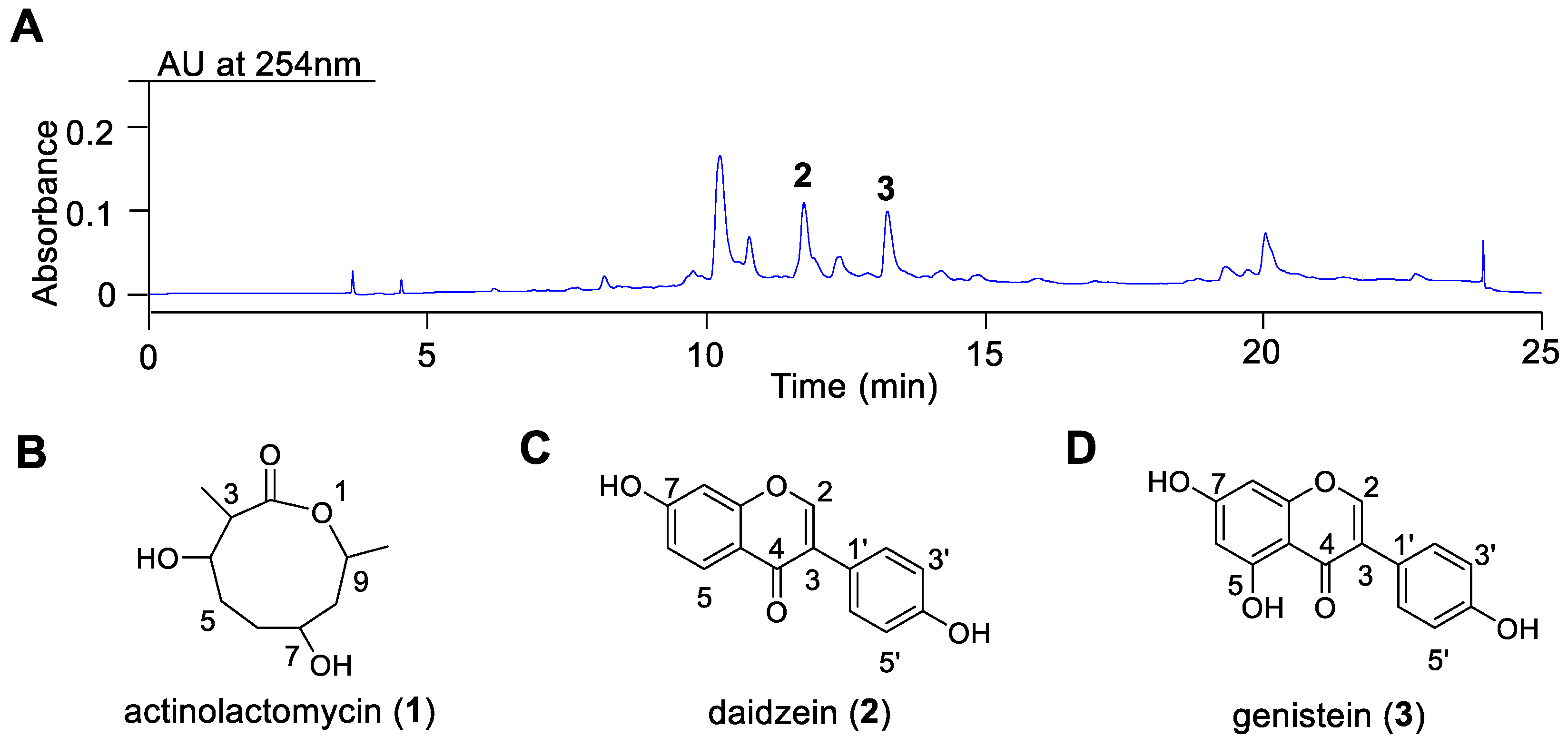
To investigate the chemical constituents of
Streptomyces sp. CB00271, large-scale fermentation (20 L) in M2 medium was performed, yielding 6.3 g of crude extract. The metabolic profile of the initial silica gel column fraction (Fr.1, 4.3 g) is shown in
Figure 3A. Subsequent purification of Fr.1 through repeated silica gel chromatography, recrystallization, and semi-preparative HPLC led to the isolation of three compounds:
1 (80 mg),
2 (1 mg), and
3 (1 mg). Their structures were unequivocally determined by comprehensive spectroscopic analysis, including HR-ESI-MS and NMR, and by comparison with literature data. The chemical structures of actinolactomycin (
1), daidzein (
2), and genistein (
3) are illustrated in
Figure 3B–D.
Compound 1 was obtained as a white crystalline powder, soluble in chloroform (CHCl
3). NMR spectra were recorded in deuterated chloroform (CDCl
3). The
1H-NMR (500 MHz, CDCl
3) data were as follows: (3H, 2 methyl protons of C9–CH
3 and C3–CH
3) of 1.19 d (
J = 6.2 Hz) and 1.04 d (
J = 6.9 Hz), respectively; (2H, 4 methylene protons of H-5a/b, H-6a/b, H-8a/b) of 1.88 m, 1.58 m, 1.44 m, 1.72 m; (1H, 4 methine protons of H-3, H-4, H-7, H-9) of 2.46 m, 3.97 dd (
J = 10.3, 4.7 Hz), 3.82 m, 4.94 m. The
13C-NMR (125 MHz, CDCl
3) data were: δC 174.21 (C-2), 80.01 (C-4), 76.32 (C-7), 69.02 (C-9), 45.21 (C-3), 42.24 (C-8), 31.36 (C-6), 28.12 (C-5), 20.48 (C-9–CH
3), 12.83 (C-3–CH
3). These spectroscopic data were consistent with those reported in the literature for actinolactomycin (ALM) [
12], leading to the identification of compound
1 as ALM. Due to the lack of a conjugated chromophore, this compound showed no significant UV absorption at 254 nm, explaining the absence of a corresponding peak in the chromatogram shown in
Figure 3A. A summary of the
1H and
13C NMR correlation data for compound
1 and the reference compound is provided in
Table 1.
Compound 2 was obtained as a light yellow amorphous powder, soluble in dimethyl sulfoxide (DMSO). HR-ESI-MS showed a pseudo-molecular ion peak at m/z 255.0651 [M+H]
+ (calcd. for C
15H
10O
4, 255.0652). The
1H-NMR (400 MHz, DMSO-
d6) data were as follows: (1H, aromatic proton H-2) of 8.23 s; (1H, aromatic proton H-5) of 7.90 d (
J = 8.8 Hz); (2H, aromatic protons H-2′/H-6′) of 7.37 d (
J = 8.7 Hz); (1H, aromatic proton H-6) of 6.85 dd (
J = 9.1,1.9 Hz); (1H, aromatic proton H-8) of 6.81 d (
J = 2.1 Hz); (2H, aromatic protons H-3′/H-5′) of 6.78 d (
J = 8.7 Hz). These data matched those reported in the literature for daidzein (7,4′-dihydroxyisoflavone) [
25], confirming its identity.
Compound 3 was obtained as a light yellow crystalline powder, soluble in DMSO. ESI-MS showed ions at m/z 271.01 [M+H]
+ (positive mode) and m/z 269.06 [M–H]
− (negative mode), suggesting a molecular formula of C
15H
10O
5. The 1H-NMR (400 MHz, DMSO-
d6) data were as follows: (1H, aromatic proton H-2) of 8.19 s; (2H, aromatic protons H-2′/H-6′) of 7.35 d (
J = 8.7 Hz); (2H, aromatic protons H-3′/H-5′) of 6.80 d (
J = 8.7 Hz); (1H, aromatic proton H-8) of 6.18 s; (1H, aromatic proton H-6) of 6.04 s. These data were consistent with those reported in the literature for genistein (4′,5,7-trihydroxyisoflavone) [
26], leading to its identification as genistein.
2.3. Cytotoxic Activity Evaluation of Compound 1
The cytotoxic potential of actinolactomycin (ALM,
1) was assessed against the human triple-negative breast cancer cell line MDA-MB-231 and the murine breast cancer cell line 4T1 using the CCK-8 assay, with cisplatin serving as a positive control. As summarized in
Table 2, after a 72-hour exposure, compound
1 demonstrated potent cytotoxic effects, with IC
50 values of 0.72 ± 0.12 μM in MDA-MB-231 cells and 0.15 ± 0.02 μM in 4T1 cells. Notably, the activity of ALM was approximately 9-fold and 31-fold higher than that of cisplatin (IC
50 = 6.37 ± 0.26 μM and 4.61 ± 1.30 μM, respectively), underscoring its superior potency in these in vitro models.
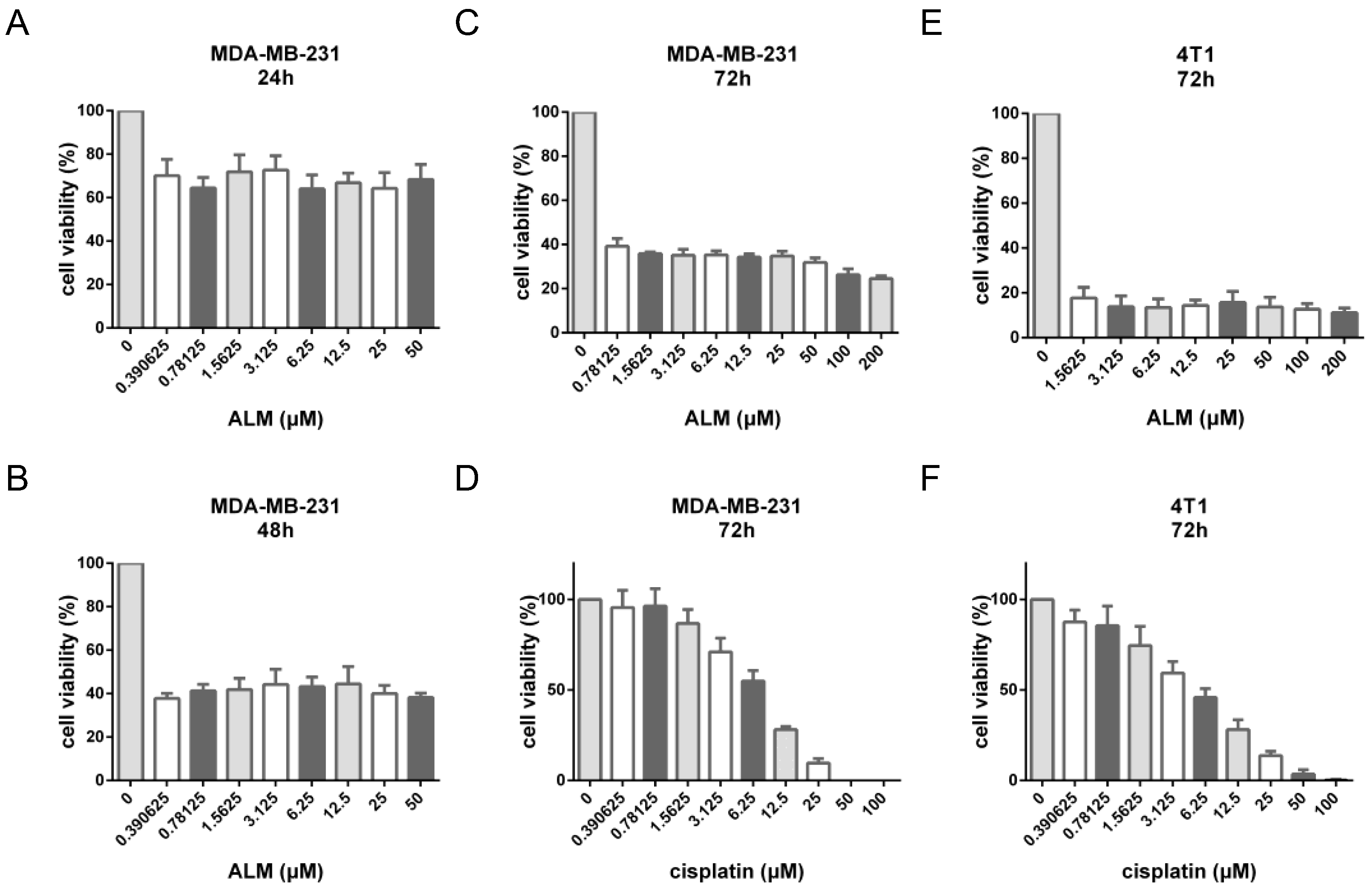
The inhibitory kinetics of compound
1 exhibited a distinct profile compared to the classical chemotherapeutic agent cisplatin. As illustrated in
Figure 4A–C, the inhibitory effect of ALM was clearly time-dependent, with a time-dependent decrease in cell viability over the 72-hour period. However, unlike cisplatin, which displayed a typical concentration-dependent response (
Figure 4D,F), compound
1 did not achieve complete inhibition (100%) within the tested concentration range (up to 200 μM), nor did it exhibit a classic concentration-effect relationship (
Figure 4C,E). This pattern suggests that ALM may not act primarily as a direct cytotoxic agent, but rather through mechanisms such as cell cycle arrest or the induction of apoptosis. Furthermore, the observed kinetics may indicate a relative selectivity of ALM towards actively proliferating cell subpopulations, with potentially limited effects on quiescent or drug-resistant cells.
3. Discussion
This study reports two primary findings with significant implications. First, we identify the soil-derived
Streptomyces sp. CB00271 as a novel and valuable microbial source for the exceedingly rare natural product actinolactomycin (ALM). To our knowledge, this strain represents only the fifth reported natural producer of ALM worldwide. Second, and more importantly, we provide the first systematic evaluation demonstrating ALM's exceptional cytotoxicity against triple-negative breast cancer (TNBC) models. ALM exhibited potent antiproliferative activity against both human MDA-MB-231 and murine 4T1 cells, with IC
50 values in the sub-micromolar range (0.72 and 0.15 μM, respectively). Its potency surpassed that of the conventional chemotherapeutic cisplatin by approximately 9-fold and 31-fold, respectively. Notably, this anti-TNBC activity is substantially higher than previously reported efficacies against other cancer types (e.g., IC
50 of 1.4-15.4 μM in K562 and A549 cells) [
15,
16], indicating a potential selective sensitivity toward the aggressive TNBC subtype.
The inhibitory kinetics of ALM revealed a distinct, non-classical profile. While its effect was strongly time-dependent, the dose-response curve reached a plateau without achieving complete (100%) inhibition, even at high concentrations. This pattern fundamentally diverges from the classic sigmoidal curve of direct DNA-damaging agents like cisplatin and is more characteristic of molecules that modulate specific cellular signaling pathways. This observation aligns with prior reports that ALM can induce G
0/G
1 cell cycle arrest and apoptosis [
17]. We propose that the plateau effect likely reflects heterogeneous cellular dependency on the pathway targeted by ALM, rather than merely indicating a resistant subpopulation. This hypothesis is strongly supported by the established molecular mechanism of ALM, which involves the inhibition of the mTOR-HIF-1α signaling axis [
18].
The proposed targeting of the mTOR-HIF-1α axis provides a compelling mechanistic rationale for ALM's observed potency against TNBC. This pathway is a master regulator of cellular metabolism, proliferation, and adaptation to stress, and its hyperactivation is a well-documented driver of the aggressive phenotype, metabolic reprogramming, and therapy resistance in TNBC [
19,
20]. Therefore, ALM’s activity may stem from a targeted disruption of this critical vulnerability in TNBC cells. Therefore, the inability to achieve complete cytostasis in vitro could be explained by the pharmacological challenge of fully ablating this robust survival pathway, a common feature of targeted therapeutics. These findings collectively position ALM not as a broad-spectrum cytotoxic agent, but as a promising precision-oriented lead compound that targets a key adaptive pathway in TNBC.
Beyond the biological activity of ALM, the genomic context of the producing strain,
Streptomyces sp. CB00271, significantly amplifies the impact of this discovery. Genomic analysis has revealed that this strain harbors a rich biosynthetic potential, with 45 putative biosynthetic gene clusters [
14]. This makes CB00271 not merely an alternative source, but a genetically tractable platform to address the critical supply bottleneck that has hindered ALM research. Future genome-mining and heterologous expression studies based on this strain are essential to elucidate the complete biosynthetic pathway, enable sustainable production, and facilitate structural diversification for optimized drug candidates.
This study has certain limitations that chart clear courses for future research. While we link ALM's activity to the mTOR-HIF-1α axis based on prior literature, its direct molecular target(s) within TNBC cells remain to be experimentally validated. Furthermore, the compelling in vitro efficacy reported here necessitates validation in in vivo TNBC models to assess its pharmacokinetic profile and antitumor effectiveness in a physiological context. Finally, given its targeted mechanism, exploring the synergistic potential of ALM with other targeted agents or standard chemotherapies represents a promising strategic direction. Addressing these points will be crucial for fully realizing the translational potential of ALM as a novel therapeutic candidate against TNBC.
4. Materials and Methods
4.1. Strain Source
The strain used in this study, Streptomyces sp. CB00271 (hereafter referred to as CB00271), was isolated from pristine soil samples collected in Yunnan Province, China. After isolation and purification in the laboratory, the strain was deposited in the China Center for Type Culture Collection (CCTCC) under the accession number CCTCC M 2020176.
4.2. Culture Media
The compositions of the media used were as follows:
G1 Solid Medium (g/L): Soluble starch 20, NaCl 0.5, KNO3 1, MgSO4·7H2O 0.5, FeSO4·7H2O 0.01, K2HPO4 0.5, agar 20; pH 7.0.
TSB Seed Medium (g/L): Tryptic Soy Broth 30; pH 7.0.
M2 Liquid Fermentation Medium (g/L): Soluble starch 15, cottonseed meal 5, CaCO3 2, CuSO4·5H2O 0.1, NaI 0.005; pH 7.1–7.2.
4.3. Reagents and Instruments
All analytical grade organic solvents, including petroleum ether, dichloromethane, ethyl acetate, and methanol, were purchased from Sinopharm Chemical Reagent Co., Ltd. (China). HPLC-grade acetonitrile was obtained from ACS (USA). Cisplatin was acquired from Beijing Solarbio Science & Technology Co., Ltd. (China). Diaion HP20 macroporous resin was from Mitsubishi Chemical Corporation (Japan). Silica gel (200–300 mesh) was from Qingdao Bangkai Hi-Tech Materials Co., Ltd. (China). The analytical column (AQ-C18, 5 µm, 4.6 mm × 250 mm) was from Waters Technologies (Shanghai) Ltd. The semi-preparative column (5 µm, 250 mm × 10 mm) was from Welch Materials, Inc. (Shanghai, China). The bacterial genomic DNA extraction kit was from Sangon Biotech (Shanghai) Co., Ltd. The CCK-8 cell proliferation assay kit was from Dojindo Molecular Technologies, Inc. (Shanghai, China).
Instrumentation included: NMR spectrometers (400/500 MHz, Bruker Corporation, Germany); a high-resolution mass spectrometer (Agilent Technologies, Inc., USA); HPLC, semi-preparative HPLC, and LC-MS systems (Waters Technologies (Shanghai) Ltd., China); a rotary evaporator (Hangzhou Yijie Technology Co., Ltd., China); a biochemical incubator (Shanghai Boxun Industry & Commerce Co., Ltd. Medical Equipment Factory, China); a shaking incubator (Shanghai Zhicheng Analytics Instrument Manufacture Co., Ltd., China); and a PCR instrument (Eppendorf AG, Germany).
4.4. Strain Identification
Total genomic DNA of the strain was extracted using the SteadyPure Bacterial Genomic DNA Extraction Kit [
23]. The 16S rRNA gene fragment was amplified via PCR using total DNA as the template and universal primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-TACGGYTACCTTGTTACGACTT-3′). The PCR product was purified by gel electrophoresis, and the target band was recovered using the Trelief® DNA Gel Extraction Kit. The purified product was sent to Tsingke Biotechnology Co., Ltd. (Beijing) for bidirectional sequencing. The resulting sequence was compared using the BLAST (Basic Local Alignment Search Tool) algorithm in the NCBI database. A phylogenetic tree was constructed using the neighbor-joining method in MEGA X software [
24].
4.5. Fermentation
Streptomyces sp. CB00271 was inoculated from a glycerol stock onto G1 solid medium and cultivated at 28 °C for 5–7 days. Colonies were then transferred to TSB seed medium and incubated at 28 °C with shaking at 200 rpm for 24–48 h. Subsequently, a 10% (v/v) inoculum was transferred into M2 liquid fermentation medium containing 3% (w/v) HP20 resin and cultivated at 30 °C with shaking at 220 rpm for 9 days.
4.6. Isolation and Purification of Secondary Metabolites
A total of 20 L of fermentation broth was collected. The HP20 resin was eluted with methanol, and the eluate was concentrated under reduced pressure. The resulting concentrate was sequentially extracted with petroleum ether and ethyl acetate. The ethyl acetate phase was concentrated to obtain a crude extract (6.3 g). This extract was subjected to silica gel (200–300 mesh) column chromatography and eluted with a stepwise gradient of ethyl acetate/methanol (10:1 to 0:1, v/v) to yield three fractions (Fr.1–Fr.3). Fr.1 was further separated by silica gel column chromatography using a gradient of dichloromethane/methanol (200:1 to 0:1, v/v), yielding six subfractions (Fr.1.1–Fr.1.6). Fr.1.4 was re-chromatographed on a silica gel column with a gradient of dichloromethane/methanol (200:1 to 0:1, v/v), resulting in four subfractions. Fr.1.4.3 afforded compound 1 (80 mg) after recrystallization. Fr.1.5 was subjected to silica gel column chromatography eluted with a gradient of dichloromethane/methanol (100:1 to 0:1, v/v), yielding three subfractions. Fr.1.5.2 was purified by semi-preparative HPLC under the following conditions: column, Welch Xtimate® C18 (5 µm, 250 mm × 10 mm i.d.); mobile phase A, 0.1% (v/v) formic acid in H2O; mobile phase B, 100% acetonitrile; gradient: 0–1 min, 95% A; 1–18 min, 95% A to 5% A; 18–20 min, 5% A; 20–20.5 min, 5% A to 95% A; 20.5–25 min, 95% A; flow rate, 2.0 mL/min; detection wavelength, 254 nm. This process yielded compound 2 (tR = 11.9 min, ~1 mg) and compound 3 (tR = 13.3 min, ~1 mg).
4.7. Structural Elucidation
The structures of the isolated compounds were determined using High-Resolution Electrospray Ionization Mass Spectrometry (HR-ESI-MS) and Nuclear Magnetic Resonance (NMR) spectroscopy.
4.8. Cytotoxicity Assay
The cytotoxicity of compound 1 against MDA-MB-231 and 4T1 cell lines was evaluated using the Cell Counting Kit-8 (CCK-8) assay, with cisplatin as the positive control. Cells were seeded into 96-well plates at a density of 3,000–5,000 cells per well and allowed to adhere for 24 h. Subsequently, the cells were treated with various concentrations of compound 1 for 24, 48, and 72 h. Then, 10 µL of CCK-8 reagent was added to each well, followed by incubation for 0.5–2 h. The absorbance was measured at 450 nm. Cell viability was calculated using the formula: (Aexperiment - Ablank) / (Acontrol - Ablank) × 100%. The IC50 values were determined by nonlinear regression analysis using GraphPad Prism 6.0 software.
5. Conclusions
In conclusion, this study makes two significant contributions to the field. First, we have identified the soil-derived Streptomyces sp. CB00271 as a novel and genetically rich microbial source for the scarce natural product actinolactomycin (ALM). Second, and more critically, we provide the inaugural systematic evidence that ALM exhibits exceptional and selective cytotoxicity against triple-negative breast cancer (TNBC) models, with potency surpassing the clinical standard cisplatin by an order of magnitude. Its unique, non-classical inhibitory kinetics align with a mechanism involving the targeted disruption of key oncogenic signaling pathways, potentially including the mTOR-HIF-1α axis, rather than direct DNA damage, positioning ALM as a promising precision-oriented lead compound against this aggressive subtype.
Critically, the discovery of ALM production in Streptomyces sp. CB00271 transcends the identification of a mere alternative source. The strain's sequenced genome and extensive biosynthetic potential establish it as a tangible platform to address the long-standing supply bottleneck through future genome-guided biomanufacturing. To fully realize the translational potential of ALM, subsequent research must prioritize the experimental validation of its direct molecular target(s) in TNBC, the assessment of its in vivo efficacy and pharmacokinetics, and the exploration of its synergy with existing therapeutic regimens.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: HR-ESI-MS spectra of 2; Figure S2: ESI-MS spectra of 2 and 3; Figure S3: UV spectra of 2 and 3; Figure S4: Crystal photo and thin-layer chromatography (TLC) for compound 1; Figure S5: 1H-NMR (500 MHz) spectrum of actinolactomycin (1) in CDCl3; Figure S6: 13C-NMR spectrum (125 MHz) spectrum of actinolactomycin (1) in CDCl3; Figure S7: 1H-NMR (400 MHz) spectrum of daidzein (2) in DMSO-d6; Figure S8: 1H-NMR (400 MHz) spectrum of genistein (3) in DMSO-d6; Figure S9: Cytotoxicity assays of ALM (1) and cisplatin.
Author Contributions
Conceptualization, B.G. (Bang Gong); methodology, B.G. (Bang Gong) and L.J. (Lin Jiang); software, B.G. (Bang Gong), J.P. (Jian Pan) and J.Z. (Jun Zhang); validation, B.G. (Bang Gong) and J.P. (Jian Pan); formal analysis, B.G. (Bang Gong); investigation, B.G. (Bang Gong) and L.J. (Lin Jiang); resources, B.G. (Bang Gong) and J.P. (Jian Pan); data curation, B.G. (Bang Gong) , J.P. (Jian Pan), J.Z. (Jun Zhang), S.R. (Shujuan Ren), and Y.T. (Yingnan Tang); writing—original draft preparation, B.G. (Bang Gong); writing—review and editing, B.G. (Bang Gong) and J.P. (Jian Pan); visualization, B.G. (Bang Gong) and J.P. (Jian Pan); supervision, L.J. (Lin Jiang); project administration, B.G. (Bang Gong); funding acquisition, B.G. (Bang Gong) and L.J. (Lin Jiang). All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Natural Science Foundation of Hunan Province (Grant No. 2025JJ80302), the Hunan Provincial Department of Education Science Research Project (Grant No. 22B0396 and 23B0988), and the “Liu Liang Academician Workstation” Mentorship Program (Grant No. 24YS004).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Barka, E. A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J. P.; Klenk, H. P.; Clément, C.; Ouhdouch, Y.; van Wezel, G. P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2015, 80(1), 1–43. [Google Scholar] [CrossRef]
- Otani, H.; Udwary, D. W.; Mouncey, N. J. Comparative and Pangenomic Analysis of the Genus Streptomyces. Sci. Rep. 2022, 12(1), 18909. [Google Scholar] [CrossRef]
- Katz, L.; Baltz, R. H. Natural Product Discovery: Past, Present, and Future. J. Ind. Microbiol. Biotechnol. 2016, 43(2-3), 155–176. [Google Scholar] [CrossRef] [PubMed]
- Gaudêncio, S. P.; Pathom-Aree, W. Editorial: Actinomycete Natural Products: Isolation, Structure Elucidation, Biological Activity, Biosynthesis, and Yield Improvement. Front. Chem. 2024, 12, 1471029. [Google Scholar] [CrossRef] [PubMed]
- Kavin Raja, M.; Subathra Devi, C. Recent Insights into Actinobacteria Research in Antimicrobial Resistance: A Review. Mol. Biol. Rep. 2025, 52(1), 683. [Google Scholar] [CrossRef]
- Genomic Mining of Ten Thousand Strains of Actinobacteria from Special Habitats in Yunnan. Yunnan Sci. Technol. Manage 2022, 35(6), 79. (in Chinese).
- Xu, L. H.; Yang, Y. R.; Jiang, C. L. A Study on the Ecological Distribution of Soil Actinomycete in Yunnan. Acta Microbiol. Sin. 1996, 36(3), 220–226. (in Chinese). [Google Scholar]
- Yi, L. W.; Kong, J. Q.; Xiong, Y.; Yi, S. R.; Gan, T.; Huang, C. S.; Duan, Y. W.; Zhu, X. C. Genome Mining of Streptomyces sp. CB00271 as a Natural High-Producer of β-Rubromycin and the Resulting Discovery of β-Rubromycin Acid. Biotechnol. Bioeng. 2021, 118(6), 2243–2254. [Google Scholar] [CrossRef]
- Gong, B.; Bai, E. H.; Feng, X. Q.; Yi, L. W.; Wang, Y. J.; Chen, X.; Zhu, X. C.; Duan, Y. W.; Huang, Y. Characterization of Chalkophomycin, a Copper(II) Metallophore with an Unprecedented Molecular Architecture. J. Am. Chem. Soc. 2021, 143(49), 20579–20584. [Google Scholar] [CrossRef]
- Gong, B. Discovery of Chalkophomycin, a Natural Copper(II) Metallophore with Antitumor Activity. Ph.D. Dissertation, Central South University, Changsha, China, 2022. (in Chinese). [Google Scholar]
- Yi, L. W. Genome Mining of Rubromycins Natural Products and the Biosynthetic Study of β-Rubromycin. Ph.D. Dissertation, Central South University, Changsha, China, 2021. (in Chinese). [Google Scholar]
- Han, B.; Cui, C. B.; Cai, B.; Ji, X. F.; Yao, X. S. Actinolactomycin, a New 2-Oxonanonidal Antitumor Antibiotic Produced by Streptomyces flavoretus 18522, and Its Inhibitory Effect on the Proliferation of Human Cancer Cells. Chin. Chem. Lett. 2005, 16(4), 471–474. [Google Scholar]
- Ren, H.; Cui, C. B.; Li, C. W.; Yao, Z. W.; Yang, M. Metabolites of Three Marine-Derived Microbial Strains and Their Antitumor Activity. J. Ocean Univ. China (Nat. Sci. Ed.) 2010, 40(5), 64–68. (in Chinese). [Google Scholar]
- Cheng, J. D.; Hu, L.; Yang, Z.; Suo, C. X.; Wang, Y. J.; Gao, P.; Cui, C. B.; Sun, L. C. 2-Oxonanonidal Antibiotic Actinolactomycin Inhibits Cancer Progression by Suppressing HIF-1α. Cells 2019, 8(5), 439. [Google Scholar] [CrossRef]
- Liu, L. H.; Bai, J.; Hu, L. X.; Jiang, D. Q. Hypoxia-Mediated Activation of Hypoxia-Inducible Factor-1α in Triple-Negative Breast Cancer: A Review. Medicine 2023, 102(43), e35493. [Google Scholar] [CrossRef]
- Srivastava, N.; Usmani, S. S.; Subbarayan, R.; Saini, R.; Pandey, P. K. Hypoxia: Syndicating Triple Negative Breast Cancer against Various Therapeutic Regimens. Front. Oncol. 2023, 13, 1199105. [Google Scholar] [CrossRef]
- Xiang, L. S.; Gilkes, D. M.; Chaturvedi, P.; Luo, W. B.; Hu, H. X.; Takano, N.; Liang, H. J.; Semenza, G. L. Ganetespib Blocks HIF-1 Activity and Inhibits Tumor Growth, Vascularization, Stem Cell Maintenance, Invasion, and Metastasis in Orthotopic Mouse Models of Triple-Negative Breast Cancer. J. Mol. Med. 2014, 92(2), 151–164. [Google Scholar] [CrossRef]
- Yao, H.; He, G. C.; Yan, S. C.; Chen, C.; Song, L. J.; Rosol, T. J.; Deng, X. Y. Triple-Negative Breast Cancer: Is There a Treatment on the Horizon? Oncotarget 2017, 8(1), 1913–1924. [Google Scholar] [CrossRef]
- Kandula, M.; Ch, K. K.; Ys, A. R. Molecular Mechanism and Targeted Therapy Options of Triple-Negative (ER, PgR, HER-2/neu) Breast Cancer: Review. World J. Oncol. 2013, 4(3), 137–141. [Google Scholar] [CrossRef]
- Tripp, A.; Poulogiannis, G. Banking on Metabolomics for Novel Therapies in TNBC. Cell Res. 2022, 32(5), 423–424. [Google Scholar] [CrossRef]
- Liu, C. W.; Chen, Y. H.; Liu, Y.; Chen, C.; Wang, T. C.; Liu, S. Y.; Luo, X. X. Analysis of Streptomyces TRM74023 Gene Cluster and Its Antibacterial Activity. Mod. Food Sci. Technol. (in Chinese). [CrossRef]
- Luo, W. G. Chemical Characterization of Three Microorganisms and Rumex crispus Seeds. Master’s Thesis, Ningxia Normal University, Guyuan, China, 2024. (in Chinese). [Google Scholar]
- Weisburg, W. G.; Barns, S. M.; Pelletier, D. A.; Lane, D. J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J. Bacteriol. 1991, 173(2), 697–703. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35(6), 1547–1549. [Google Scholar] [CrossRef]
- Xuan, Q. C.; Huang, R.; Miao, C. P.; Chen, Y. W.; Zhai, Y. Z.; Song, F.; Wang, T.; Wu, S. H. Secondary Metabolites of Endophytic Fungus Trichoderma sp. YM 311505 of Azadirachta indica. Chem. Nat. Compd. 2014, 50(1), 139–141. [Google Scholar] [CrossRef]
- Geng, Y.; Zhu, S. L.; Cheng, P.; Lu, Z. M.; Xu, H. Y.; Shi, J. S.; Xu, Z. H. Bioassay-Guided Fractionation of Ethyl Acetate Extract from Armillaria mellea Attenuates Inflammatory Response in Lipopolysaccharide (LPS) Stimulated BV-2 Microglia. Phytomedicine 2017, 26, 55–61. [Google Scholar] [CrossRef]
- Smeriglio, A.; Calderaro, A.; Denaro, M.; Laganà, G.; Bellocco, E. Effects of Isolated Isoflavones Intake on Health. Curr. Med. Chem. 2019, 26(27), 5094–5107. [Google Scholar] [CrossRef]
- Zhao, Q. X.; Zhao, M.; Parris, A. B.; Xing, Y.; Yang, X. H. Genistein Targets the Cancerous Inhibitor of PP2A to Induce Growth Inhibition and Apoptosis in Breast Cancer Cells. Int. J. Oncol. 2016, 49(3), 1203–1210. [Google Scholar] [CrossRef]
- Liu, Y. C.; Zou, T. B.; Wang, S. H.; Chen, H.; Su, D. J.; Fu, X. N.; Zhang, Q. Y.; Kang, X. M. Genistein-Induced Differentiation of Breast Cancer Stem/Progenitor Cells through a Paracrine Mechanism. Int. J. Oncol. 2016, 48(3), 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Berndt, S.; Issa, M. E.; Carpentier, G.; Cuendet, M. A Bivalent Role of Genistein in Sprouting Angiogenesis. Planta Med. 2018, 84(9-10), 653–661. [Google Scholar] [CrossRef] [PubMed]
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).