
Submitted:
30 December 2025
Posted:
31 December 2025
You are already at the latest version
Abstract
Background/Objectives: Many products that claim to have anti-aging effects have been reported, but their relative potency is not clear. In this study, in vitro replicative lifespan extension (RLE) activity of various groups of physiologically active substances was compared by updated “overlay method”. Methods: Human dermal and periodontal ligament fibroblasts (HDFa, HPLF) were inoculated into the inner 60 wells of 96-well microplate, surround by sterile water to prevent the water evaporation. At Day 1 and Day 8, the cells were overlayed with wide ranges of concentrations (0.01~100 µM) of samples without medium change. Viable cell number was measured by MTT method at Day 15, and then corrected for the variation of cell growth due to the location of inoculated cells. RLE value was calculated as the maximum cell proliferation rate relative to the control. Results: Cell density of HDFa and HPLF at subculture decreased with the passage number, and their growth was stopped at 56 or 85 population doubling levels (PDLs), respectively. Hydrocortisone showed the highest RIE values among six hormones, followed by 3 plant extracts, sodium ascorbate and quercetin. On the other hand, other antioxidants, chlorogenic acid, phenylpropanoids, vanilloids, bacterial products showed little or no RLE effects. However, for HPLF cells, hydrocortisone did not show RLE effects while oxytocin showed slight stimulation. Conclusions: When differences in proliferation due to cell seeding position were corrected, biphasic dose response curve of most of the compounds significantly reduced. The present study suggests the significant role of hormone for the regulation of long-term aging process.
Keywords:
anti-aging
; replicative life span
; overlay quantification method
; hormones
; antioxidants
; hormesis
1. Introduction
Due to the increase of lifespans, the proportion of elderly people has increased. As aging progresses, oxidation, inflammation, and cellular dysfunction occur, leading to the onset of various diseases and death [1,2]. Traditional medicines, natural products and even daily supplements may prevent age-related diseases, but their relative anti-aging potential has never been investigated. Human primary cultured cells are used as a model of cellular aging because they cease proliferation after approximately 50±10 divisions due to DNA damage, and deviation from the cell cycle. During replicative phase (phase I), cellular metabolic activity, intracellular NAD+ concentration, telomere length, extracellular matrix (e.g., collagen and elastin) and histone H1 content gradually decline, while cellular and nuclear volume increased [3,4,5] (Figure 1). At the terminal phase (phase II), senescent cells exhibit a senescence-associated secretory phenotype (SASP) and affect the surrounding environment by secreting various physiologically active substances and inducing inflammation and tissue remodeling [6]. Associated β-galactosidase (SA-βgal) has been used as a marker of terminal phase of aging, since this marker was expressed by larger senescent cells that occupy very small percentage of total population, but not smaller pre-senescent fibroblasts [7,8]. We found that β-galactosidase can be detected only in dying or irreversibly damaged cells, that cannot be evaluated by RLE (Figure 1). The phase I process is overwhelmingly longer than the phase II process, and we thought that this length of phase I may reflect “Healthy life expectancy” that refers to the period during which one can live independently and healthily without restrictions on daily life. And we got the idea that establishing a method to accurately measure this replicative life span extension (RLE) could be used to search for substances that extend healthy lifespan.
Until now, the colony formation method has been used to quantify the extension of life span of human fibroblasts, based on the tight association of aging progression and reduction of colony forming activity. However, there are colonies of various morphological types (dense, mixed and diffuse) formed by skin fibroblasts. During in vitro aging, diffuse colonies, that contain higher number of SA-βgal positive senescent cells, and lower number of Ki67 positive rapidly grown cell, increases [9]. This heterogeneity of colony size hinders the accurate measurement of RLE activity. This urges us to establish a new quantitative method of the RLE activity using 96-well microplate [8]. After the publication, however, we noticed that two additional restrictive conditions have to be added for the accurate quantification of the RLE: first, the number of inoculated cells have to be within a quantitative range at Day 15, and second, the variation in cell proliferation rate due to the position of the inoculated cells must be corrected.
In this study, we first updated the RLE quantification method as described above, then identified hydrocortisone as the positive control that showed the highest RLE activity for human dermal fibroblasts (HDFa). Next, using hydrocortisone as positive control, the RLE activity of various physiologically active compounds including antioxidants with anti-aging properties were investigated. We also examined their effect on periodontal ligament fibroblasts (HLPF) for their future dental application.
2. Materials and Methods
2.1. Chemicals
Hydrocortisone (MW=362.5), estradiol (MW=272.4), resveratrol (MW=228.1), curcumin (MW=368.4), quercetin (MW=302.2), sodium ascorbate (MW=198.1), vanillic acid (MW=168.1), vanillin (MW=152.2), p-coumaric acid, caffeic acid (MW=180.1), trans-ferulic acid (MW=194.2), chlorogenic acid (MW=354.3),were purchased from Tokyo Chemical Industry Co., Ltd., Tokyo, Japan; hydrocortisone sodium succinate (Solu-Cortef) (MW=485.5) from Takeda Pharmaceutical Co. Ltd., Osaka, Japan; fetal bovine serum (FBS) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) from Sigma-Aldrich Inc., St. Louis, MO, USA. Dimethyl sulfoxide, modified Eagle’s medium (DMEM), penicillin-streptomycin solution (×100), and 0.25% trypsin-1 mM EDTA-4Na, testosterone, melatonin, oxytocin, were purchased from Fujifilm Wako Pure Chemical Ind., Osaka, Japan, while 96-microwell plates were purchased from Techno Plastic Products AG, Trasadingen, Switzerland [10].
2.2. Preparation of Plant Extracts
Taheebo tea was purchased from Konishi Pharmaceutical Co., Ltd. (Osaka, Japan). 10 g of tea was weighed out and placed in a tea bag in a heat-resistant pot containing 1 liter of water. The tea was brought to a boil, then reduced to low heat and brewed for 30 minutes. After making the extract isotonic by adding phosphate buffer solution (PBS(-)) powder, it was sterilized by filtration through a 0.45 µm membrane filter. Twenty Japanese pine cone of Pinus parviflora Sieb et Zucc was extracted with 4L boiling water, and made isotonic, and then sterilized similarly. The alkaline extract of Sasa sp. (Sasa Health®) (SE) was provided from Daiwa Biological Research Institute Co., Ltd., Kanagawa, Japan [11].
2.3. Preparation of Bacterial Cells and Supenatant
2.3.1. Intestinal Bacteria
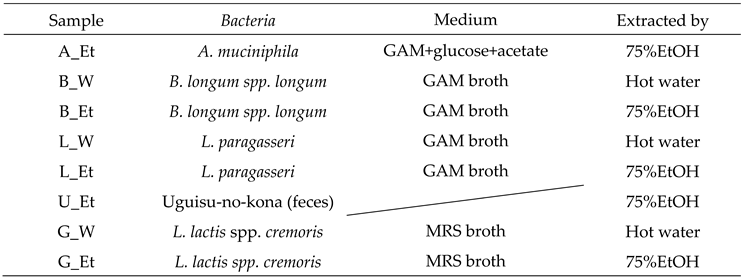
Microorganisms at late logarithmic growth phase or early stationary phase were used. Akkermansia muciniphila (grown in modified GAM+glucose+acetate), Bifidobacterium longum spp. longum and Lactobacillus paragasseri (both grown in GAM broth) were grown under anaerobic condition. Lactococcus lactis spp. cremoris (gown in MRS broth) was grown under aerobic condition. The culture medium was centrifuged (6,000 x g, 5 min) to collect the bacterial pellet. The collected bacterial cells were frozen and stored at -80 °C. The freeze-dried product was crushed and extracted with either pure water or 75% (v/v) ethanol (Table 1), according to previous reports [12. 13, 14]. Only L. lactis spp. cremoris was treated with gold nanoparticle after removing the medium components by centrifugation. Freeze-dried feces (intestinal bacterial community) of Japanese bush warbler (Uguisu-no-kona) was directly extracted with 75% ethanol. All samples were filtered through Millipore filter (0.22 μm) for the sterilization.
2.3.2. Oral bacteria
Candida albicans strains SC5314 (MYA-2876) was cultured in YEPD broth under aerobic conditions at 37 °C. Porphyromonas gingivalis (ATCC 33277) was cultured anaerobically in GAM broth supplemented with 0.5 mg/ml hemin and 5 mg/ml menadione using AnaeroPack systems. The supernatants were collected and centrifuged again at 15,000 rpm for 30 min at 4 °C. The resulting supernatants were filtered (0.22 μm) and ultracentrifuged at 100,000 × g.
2.4. Cell culture
Human dermal fibroblast, adult (HDFa) from adult skin (catalog number: C0135C; Thermo Fisher Scientific, Waltham, MA, USA), human periodontal ligament fibroblast (HPLF) (purchased from SCR ScienCell Research Laboratories, Carlsbad, CA, USA) were cultured at 37 °C in regular culture medium [DMEM supplemented with 10% heat (56 °C, 30 min) inactivated FBS, 100 U/ml penicillin G, and 100 μg/ml streptomycin sulfate] in a humidified incubator (MCO-170 AICUVD-P; Panasonic Healthcare Co., Ltd., Gunma, Japan) with 5% CO2 [10]. For subculture, HDFa cells were harvested using 0.25% trypsin-EDTA and seeded at a 1:4 ratio once a week, with a medium change after 4 days. On the other hand, HPLF cells were seeded at 1:3 ratio twice a week considering the rapid cell growth (Figure 2A). With continued passage in this manner, the cell density gradually decreased, and proliferation ceased after 56 or 85 divisions (Figure 2B).
2.5. Quantification of Replicative lifespan elongation (RLE) activity
Replicative lifespan elongation (RLE) activity was quantified by the updated “Overlay” method. Before seeding the cells, 275 μl of sterile water was added to the outer periphery of the 96-microwell plate (Becton Dickinson Labware, Franklin Lakes, NJ, USA) to minimize the water evaporation and bacterial infection during cell culture. Thus, 60 µL of 32~256 cells (depending on the PDL and type of cell) were inoculated onto inner 60 wells of the plate. After 24 h, complete cell attachment was achieved, and 40 μl of fresh medium containing different concentrations of sample was added, without medium change to prevent the cell detachment. After incubation for 7 days, cells were again overlaid with 100 µl of fresh medium with samples. Cells were incubated for a further 7 days (for a total of 14 days). The relative viable cell number at Day 15 was then determined by the MTT method, and expressed as the absorbance at 560 nm (A560) [8](Figure 3A). The A560 value of control cells was usually in the range of 0.15 to 0.45 to keep the linearity of viability (Figure 2C). Since cells near the center showed more vigorous cell proliferation than the cells in the periphery (Figure 3B, insert), the difference of growth rate (Δ)(up to 25%) depending on the location of cell inoculation was corrected. The replicative lifespan elongation (RLE) was calculated by the following equation: RIE=(b/a) x 100, where a is absorbance at 560 nm (A560) of control cells, and b is that of the maximum cell number of treated cells at the optimal concentrations of the sample (Figure 3B).
2.4. Statistical Analysis
Experimental data are expressed as the mean ± standard deviation of sextuple determinations. Bonferroni’s post-hoc analysis was performed after one-way ANOVA to compare the means of 10 samples including control, using the SPSS program for Windows version 22, and p<0.05 was considered as significant.
3. Results
3.1. Requirement for Accurate Calculation of Replicative Lifespan Extension (RLE) Activity
3.1.1. Inoculation of Optimal Concentration of Cells to Allow Qualitative RLE Measurement
After long-term culture (2 weeks), it is necessary to bring the cell number within a range that permits the quantitative measurement. For HDFa cells, A560 value at Day 15 should be between 0.1 to 0.42, that corresponds to 16 to 1024 cells/well (Figure 2C, upper panel; Supplementary Figure S1) [15].
Also, for HPLF cells, linearity was kept similarly between A560 value from 0.1 to 0.6, but optimal inoculation number shifted to lower range (16 to 64 PDL) due to their rapid growth when using young cells (25 PDL, 40%lifespan) as compared with HDFa cells. If an excess of cells are inoculated, the number of cells will fall outside the range of quantitative measurement, making it impossible to accurately determine the cell number (Figure 2C, lower panel). When older HPLF used, more numbers of cells should be inoculated to fall into the linear range.
3.1.2. Correction for Proliferation Errors Due to the Position of Cell Inoculation Is Essential
We found that the viable cell number increased gradually when the position of cell inoculation approached to the center of the place. The difference of viable cell number between these two points was approximately 25% on the average of 72 expeimets (Figure 3B). This artifactual variation should be corrected in each well. If this correction was not performed, all cells would exhibit a biphasic hormesis curve even without addition of test samples, leading to the misconception as if sample promotes proliferation.
3.2. RLE activity of anti-aging candidates against HDFa cells
3.2.1. Hormones
Among 7 hormones, both water-soluble and lipophilic hydrocortisone, but not succinate (supplementary Figure S1, cited from [11] with permission), exhibited the highest replicative lifespan extension (RLE) effects on human dermal fibroblasts (HDFa). The RLE effects of other hormones (testosterone, estradiol, cholecalciferol, melatonin, and oxytocin) were negligible (Figure 4, supplementary Table S1A). Hydrocortisone, either hydrophilic or lipophilic, can be used as a positive control for the search of new RLE substances for HDFa cells. When the treatment time was halved to 7 days, the RLE effect of hydrocortisone was also halved (Supplementary Figure S2), indicating that RLE effect of hydrocortisone increases continuously for at least 2 weeks.
3.2.2. Antioxidants
Among eight anti-oxidants, only sodium ascorbate and quercetin showed some RLE effects, but not so potent as hydrocortisone. Other antioxidants such as curcumin, resveratrol, astaxanthin, docosahexaenoic acid (DHA), coenzyme Q10 and epigallocatechin gallate (EGCG) were essentially inactive (Supplementary Figure S3, Supplementary Table S1B) [16].
3.2.3. Chlorogenic acid, phenylpropanoids and vanilloids
Since we reported that coffee extract showed very high RLE activity against HDFa cells [10], we investigated here the RLE activity of main coffee component. In contrast to our expectation, chlorogenic acid and caffeic acid, and other related phenylpropanoids (ferulic acid, p-coumaric acid), and vanilloids (vanillin and vanillic acid) showed little or no RLE effects on HDFa cells (Figure 5, Supplementary Table S1C)
3.2.4. Plant extract
Taheebo tea showed comparable RLE activity with hydrocortisone in HDFa cells, but the effective concentration range was slightly narrower. Pine cone of Pinus parviflora Sieb. et Zucc.and Sasa sp. Extracts showed comparable RLE value (Supplementary Figure S4A. B). Only Sasa sp. showed eal RLE activity of HPLF (Figure S4, Table S1D)[11].
3.2.5. Bacterial secretion
Six of eight intestinal bacteria, and 1 out of four oral bacteria showed minor RLE activity, although their effects are much lower than hydrocortisone (Figure 6, Supplementary Table S1E).
4. Discussion
During the establishment of the updated assay for RLE activity, we had noticed that the following four conditions must be met for accurate calculation of RLE value,: (i) preparation of single-cell suspension to prevent the peeling off the aggregated cells during long culture and minimize the variation of the data as small as possible, (ii) inoculation of an appropriate number of cells to allow the quantitative measurement of viable cell number, (iii) overlay of fresh medium containing samples to prevent the nutritional depletion and the cell detachment by the medium change, and (iv) the correction of different cell growth in the different wells of 96-microwell plate (Figure 3).
The present study, performed under these four restricted conditions, demonstrated that hydrocortisone showed the highest RIE values among 7 hormones against HDFa cells, followed by several plant extract such as taheebo tea, pine cone and Sasa sp, whereas most of antioxidants, chlorogenic acid, phenylpropanoids, vanilloid and bacterial culture supernatants showed little or no RLE effects. These data were summarized in Figure 7 and supplementary Table S1). Many drugs are known to exhibit a biphasic dose-response curve, so-called hormesis, over a wide concentration range [17]. However, such a biphasic dose response of antioxidants significantly diminished, if the artificial effect of the increased cell growth in the center of plates were subtracted. The reproducibility of the present findings should be investigated in the long term culture experiments with various types of cells.
Initially, we examined the RLE activity of various physiological chemicals using human dermal fibroblast (HDFa) cells. Dermal fibroblasts are the main cell type present in skin connective tissue (dermis), and play an important role in cutaneous wound healing [18]. Hydrocortisone, the endogenous glucocorticoids in human, is released through the hypothalamic-pituitary-adrenal axis in response to various stressors. The in vitro anti-aging effect of hydrocortisone changes depending on the type of vertebrate cell lines. It has been reported to prolongs the lifespan of human lung fibroblast by the reduction of its high affinity binding sites [19], while it accelerated skin aging [20] (PMID: 28793178), and cause skin atrophy and inhibit collagen production [21]. We also found that hydrocortisone did not clearly show RLE activity in human periodontal ligament fibroblast (HPLF) (Figure 4C and Figure 5C), Oh the other hand, oxytocin showed week but significant RLE effect on HPLF (Figure 4C). Since oxytocin is strongly expressed in oral cells, such as periodontal ligament fibroblast [22] and pulp cells [23], suggesting that the connection of RLE activity and receptor expression.
It has been reported that oxidation products accumulate with age, and that oxidative stress accelerates aging [1,2]. However, this study provided surprising result that antioxidants showed very low or no RLE activity.
Short period (1 day) treatment of human dermal papilla cells with higher concentrations (50-100 µM) of resveratrol has been reported to downregulate the expression of aging markers such as β-galactosidase and senescence-associated secretory phenotype (SASP) [24]. On the other hand, a longer time (2 weeks or more) treatment with lower concentrations (0.2-20 µM) of resveratrol showed weak RLE effect on human fetal lung fibroblast MRC-5 (2 PDLs), no effect on human foreskin fibroblasts FB0603, but rather shortened the life span of human ear skin fibroblasts H8F2p 25 LM and human foreskin fibroblasts Hs68 [25]. Recent study demonstrated that nitroxide 4-hydroxy-TEMPO, ergothioneine, and Trolox extended the replicative lifespan (40 ± 1 population doublings (PD)) by 7 ± 2, 4 ± 1, and 3 ± 1 PD, suggesting the possible involvement of the elevation of the mitochondrial membrane potential in the prolongation of the replicative lifespan, whereas coumaric acid, curcumin and resveratrol did not affect the replicative lifespan [26]. These date are consistent with the present study that showed no significant RLE activity of resveratrol oh HDFa cells.
One possible reason is that antioxidants and polyphenols are unstable under long culture condition, possibly due to their decomposition and/or production of hydrocortisone in the presence of oxygen [27,28]. To solve this problem, the addition of catalase (that degrade hydrogen peroxide) may ameliorate the harmful effect of oxidative stress is achieved by antioxidant enzymes (Superoxide dismutases-SODs, catalase, glutathione peroxidase-GPx) [29]. Citric acid contributes to the stabilization of vitamin C, so it is expected that the antioxidant power will be increased if the two compounds are used together at low concentrations that do not cause toxicity [30].
We have previously reported that lignin degradation products such as phenylpropanoid (caffeic acid, ferulic acid and p-coumaric acid) and especially vanilloids (vanillic acid, vanillin) showed higher anti-UVC activity than vitamin C, due to higher stability than sodium ascorbate [31]. This suggests such substances having high anti-UVC activity (short effect) do not always show RLE activity (long effect).
Several plant extracts such as Taheebo tea, pine cone and Sasa sp. show higher RLE effect than their components. Combination of several components may may potentiate their RLE effect. In dentistry, the hot water-extracted herbal preparations (so-called Kampo Medicines), Rikkosan is used for the treatment of toothache [32], and Hangeshashinto for the treatment of stomatitis [33]. Sakagami et al. demonstrated that Rikkosan exhibited RLE effects on HDFa cells, to comparable with that achieved by hydrocortisone, but its optimal concentration range is narrow. On the other hand, Hangeshashinto did not exhibit the RLE activity, due to the presence of cytotoxic effects in constituent plant extracts. The removal of cytotoxic substances may elevate the specific activity of these two Kampo Medicines (The 67th Annual Meeting of Japanese Association for Oral Biology, 2025 September). We found that several intestinal bacteria showed slightly higher RLE activity than on HDFa cells than oral bacteria. Their effects on human oral cells such as gingival fibroblast, pulp cells as well as HPLF are under way.
5. Conclusions
We have developed the updated “overlay method” that can measure RLE activity of a large number of samples at the same time. The determination of RLE values by MTT method is based on mitochondrial enzyme activities such as succinate dehydrogenase and NAD(P)H-dependent dehydrogenase, which reflects the number of cells and cellular metabolic activity [34]. Since there is a close relationship between mitochondrial dysfunction and senescence [35], this system may be useful for developing drugs that extend healthy lifespan. In this study, hydrocortisone exerted an RLE effect in skin cells but inhibited it in periodontal ligament cells, suggesting the existence of a tissue-specific regulatory mechanism. In contrast, the low RLE values of many antioxidants may be due to the short-term nature of their effects, suggesting the importance to establish the strategy how to stabilize them. Furthermore, because various plant extracts contain a large number of active ingredients, a strong synergistic effect may be observed. Conversely, if cytotoxic substances are present, removing them may enhance their activity. The present method can also be used at the purification and identification step of new substances that extend replicative lifespan.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Linearity of RLE effect of hydrocortisone was maintained between 0.2 to 1.0 of A560 in HDFa cells.; Figure S2: Hydrocortisone succinate, but not succinic acid, showed potent RLE effect on HDFa cells; Figure S3: Most of anti-oxidants, except quercetin and sodium ascorbate, showed little or no RLE effect on HDFa cells; Figure S4: Taheebo, pine cone and and Sasa sp. extracts showed potent RLE effect on HDFa cells.; Table S1: RLE activities of hormones, anti-oxidants, polyphenols, plant and bacterial exudates on HDFa and HPLF.:
Author Contributions
HS, AMA, TH, MA and HA performed experiments. HS wrote the article. HK and YK reviewed the article. TT designed and interpreted the experimental results. FI, KB, MI and SY edited the article. All Authors read and approved the final version of the article.
Funding
This research was funded by in part by Japan Medical Herb Association (Sakagami) and Research fund based on the agreement between Meikai University and Autonomous University of the State of Mexico.
Institutional Review Board Statement
Not applicable for studies not involving humans or animals.
Informed Consent Statement
Not applicable for studies not involving humans.
Acknowledgments
The Authors thank Dr. Hiroshi Kadokura and Prof. Megumi Inomata for the supply of HDFa and HPLF cells.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| RLE | Replicative Lifespan Elongation |
| PDL | Population doubling level |
| HDFa | Human dermal fibroblast |
| HPLF | Periodontal ligament fibroblast |
References
- Moldogazieva, NT; Mokhosoev, IM; Mel’nikova, TI; Porozov, YB; Terentiev, AA. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. In Oxid Med Cell Longev;Review; PMC article, 14 Aug 2019; Volume 2019, p. 3085756. [Google Scholar] [CrossRef] [PubMed]
- Rea, IM; Gibson, DS; McGilligan, V; McNerlan, SE; Alexander, HD; Ross, OA. Age and Age-Related Diseases: Role of Inflammation Triggers and Cytokines. In Front Immunol;Review; PMC article, 9 Apr 2018; Volume 9. [Google Scholar] [CrossRef] [PubMed]
- Ogrodnik, M. Cellular aging beyond cellular senescence: Markers of senescence prior to cell cycle arrest in vitro and in vivo. Aging Cell Epub. 2021, 20(4), e13338. [Google Scholar] [CrossRef] [PubMed]
- Kim, DJ; Chang, SS; Lee, J. Anti-Aging Potential of Substance P-Based Hydrogel for Human Skin Longevity. Int J Mol Sci 2019, 20(18), 4453. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, Y; Sakagami, H; Yamada, M. Histone H1 in G1 arrested, senescent and Werner syndrome fibroblasts. From: Werner syndrome and human aging; Salk, D, Fujisawa, Y, Martin, GM, Eds.; Plenum Publishing Corporation, 1985; pp. 373–389. [Google Scholar]
- Coppé, J-P; Desprez, P-Y; Krtolica, A; Campisi, J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol 2010, 5, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Dimri, GP; Lee, X; Basile, G; Acosta, M; Scott, G; Roskelley, C; Medrano, EE; Linskens, M; Rubelj, I; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci U S A 1995, 92(20), 9363–7. [Google Scholar] [CrossRef]
- Biran, A; Zada, L; Abou Karam, P; Vadai, E; Roitman, L; Ovadya, Y; Porat, Z; Krizhanovsky, V. Quantitative identification of senescent cells in aging and disease. Aging Cell 2017, 16(4), 661–671. [Google Scholar] [CrossRef]
- Zorin, V; Zorina, A; Smetanina, N; Kopnin, P; Ozerov, IV; Leonov, S; Isaev, A; Klokov, D; Osipov, AN. Diffuse colonies of human skin fibroblasts in relation to cellular senescence and proliferation. Aging 2017, 9(5), 1404–1413. [Google Scholar] [CrossRef]
- Acra, AM; Sakagami, H; Uota, S; Yoshihara, M; Kito, S; Izawa, M; Ohtaka, Y; Nakaya, G; Koga-Ogawa, Y; Nobesawa, T; Ueda, D; Suzuki, R. Quantification of In Vitro Replicative Lifespan Elongation Activity of Pharmaceuticals, Natural Products and Radiation Using the “Overlay” Method. In Vivo 2025, 39(5), 2534–2548. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H; Horiuchi, M; Suguro, M; Uota, S; Toeda, K; Oizumi, T; Iwama, Y. Search for anti-aging substances: Superiority of Taheebo tea and specificity of Sasahealth®. New Food Industry 2025, 67(10), 566–572. [Google Scholar]
- Yuan, H; Shi, J; Gu, C; Yuan, J; Huang, C; Li, X; Zhou, K; Qi, J. Akkermansia muciniphila: A next-generation gut probiotic supporting neurorepair and functional recovery. Neural Regen Res Epub ahead of print. 2025. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y; Hou, Y; Lao, X. The Role of Akkermansia muciniphila in Disease Regulation. Probiotics Antimicrob Proteins 2025, 17(4), 2027–2038. [Google Scholar] [CrossRef] [PubMed]
- Anand, a; Sato, M; Aoyagi, H. Screening of phosphate-accumulating probiotics for potential Use in chronic kidney disorder. Food Science and Technology Research 2019, 25(1), 89–96. [Google Scholar] [CrossRef]
- Harada, T; Sakagami, H; Acra, AM; Asami, R; Uota, S; Sakiyama, K; Iwasa, F. Comparative study of the replicative life span extension effects of hydrocortisone, quercetin, and astaxanthin. New Food Industry 2025, 67(7), 395–399. [Google Scholar]
- Harada, T; Sakagami, H; Acra, AM; Asami, R; Uota, S; Sakiyama, K; Iwasa, F. Re-evaluation of the replicative life span extension effects of antioxidants. New Food Industry 2025, 67(9), 507–512. [Google Scholar]
- Calabrese, EJ. Paradigm lost, paradigm found: the re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environ Pollut 2005, 138(3), 379–411. [Google Scholar] [CrossRef] [PubMed]
- Marta, AK. Agnes SKl:Isolation and Culture of Human Dermal Fibroblasts. Methods Mol Biol 2019, 1993, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Rosner, BA; Cristofalo, VJ. Hydrocortisone: a specific modulator of in vitro cell proliferation and aging. Mech Ageing Dev 1979, 9(5-6), 485–96. [Google Scholar] [CrossRef]
- Nam, J.-J.; Min, J.-E.; Son, M.-H.; Jin-Hwan Oh, J.-H.; Kang, S. Ultraviolet- and infrared-induced 11 beta-hydroxysteroid dehydrogenase type 1 activating skin photoaging is inhibited by red ginseng extract containing high concentration of ginsenoside Rg3(S). Photodermatol Photoimmunol Photomed 2017, 33(6), 311–320. [Google Scholar] [CrossRef]
- Hall, L; Hart, R. Role of corticosteroids in skin physiology and therapeutic potential of an 11β-HSD1 inhibitor: A review. Int J Dermatol 2024, 63(4), 443–454. [Google Scholar] [CrossRef]
- Ge, B; Liu, H; Liang, Q; Shang, L; Wang, T; Ge, S. Oxytocin facilitates the proliferation, migration and osteogenic differentiation of human periodontal stem cells in vitro Arch Oral Biol. 2019, 99, 126–133. [Google Scholar] [CrossRef]
- Kato, Y; Yokose, S. Oxytocin facilitates dentinogenesis of rat dental pulp cells. J Endod 2021, 47(4), 592–599. [Google Scholar] [CrossRef]
- Ei, ZZ; Srithawirat, T; Chunhacha, P; Chaotham, C; Arunmanee, W; Phookphan, P; Chanvorachote, P. Resveratrol shows potent senescence reversal in experimental cellular models of particular matter 2.5-induced cellular senescence in human dermal papilla cells. In Vivo 2024, 38(2), 665–673. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I. Bartosz G: Effect of antioxidants on the fibroblast replicative lifespan in vitro. Oxid Med Cell Longev 2020, 2020, 6423783. [Google Scholar] [CrossRef]
- Bartosz, G; Pieńkowska, N; Sadowska-Bartosz, I. Effect of Selected Antioxidants on the In Vitro Aging of Human Fibroblasts. Int J Mol Sci. 2024, 25(3), 1529. [Google Scholar] [CrossRef]
- Tajima, N; Takasaki, M; Fukamachi, H; Igarashi, T; Nakajima, Y; Arakawa, H. Determination of reactive oxygen generated from natural medicines and their antibacterial activity. J Pharm Anal 2016, 6(4), 214–218. [Google Scholar] [CrossRef]
- Arakawa, H; Tsuruoka, K; Ohno, K; Tajima, N; Nagano, H. Development of a highly sensitive chemiluminescent assay for hydrogen peroxide under neutral conditions using acridinium ester and its application to an enzyme immunoassay. Luminescence 2014, 29(4), 374–7. [Google Scholar] [CrossRef]
- Jomova, K; Raptova, R; Alomar, SY; Alwasel, SH; Nepovimova, E; Kuca, K; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch Toxicol;Epub 2023, 97(10), 2499–2574. [Google Scholar] [CrossRef] [PubMed]
- Crawford, RD. Proposed role for a combination of citric acid and ascorbic acid in the production of dietary iron overload: a fundamental cause of disease. Biochem Mol Med 1995. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H; Amano, S; Uota, S; Tanuma, SI; Inomata, M; Shindo, A; Kusano, M; Kikkawa, Y; Horiuchi, M; Ooka, T. Prominent Anti-UVC Activity of Lignin Degradation Products. In Vivo 2022, 36(6), 2689–2699. [Google Scholar] [CrossRef]
- Itagaki, T; Nakamura, K; Tanabe, T; Shimura, T; Nakai, Y; Sakata, KI; Sato, J; Kitagawa, Y. Rikkosan’s Short-Term Analgesic Effect on Burning Mouth Syndrome: A Single-Arm Cohort Study. Biomedicines 2024, 12(5), 1013. [Google Scholar] [CrossRef] [PubMed]
- Takata, T; Moriya, J; Miyazawa, K; Inoue, S; Yamada, S; Han, J; Yang, Q; Guo, X; Mizuta, S; Nakahashi, T; Onai, N; Nakano, H; Masauji, T; Motoo, Y. Potential of Natural Products in Hangeshashinto Water Extract on the Direct Suppression of Stomatitis Induced by Intra-/Extracellular Advanced Glycation End-Products Review. Int J Mol Sci. 2025, 26(18), 9118. [Google Scholar] [CrossRef] [PubMed]
- Präbst, K; Engelhardt, H; Ringgeler, S; Hübner, H. Basic Colorimetric Proliferation Assays: MTT, WST, and Resazurin. Methods Mol Biol. 2017, 1601, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C; Macchi, C; D’Alonzo, C; Venturin, M; Ruscica, M; Corsini, A; Battaglia, C; Bellosta, S. Simvastatin ameliorates senescence-induced mitochondrial dysfunction in vascular smooth muscle cells. Atherosclerosis 2025, 403, 119176. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Expression of age-related characteristics during the in vitro aging process of human fibroblast model.
Figure 1.
Expression of age-related characteristics during the in vitro aging process of human fibroblast model.
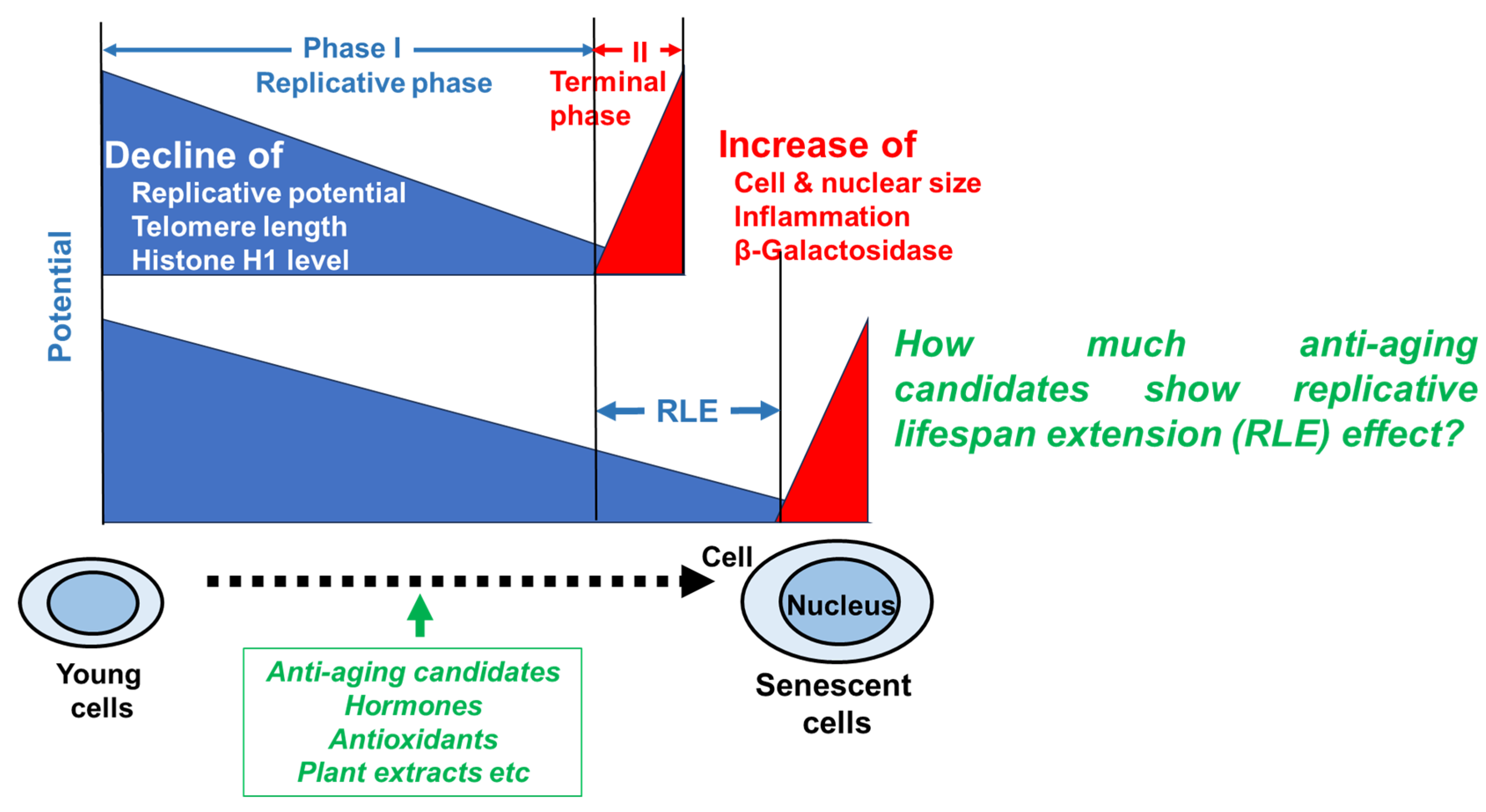
Figure 2.
Subculture schedule of HDFa and HPLF cells (A) to monitor the aging-associated decline of viable cell number (B) and the range of linearity of viable cell number at Day 15.
Figure 2.
Subculture schedule of HDFa and HPLF cells (A) to monitor the aging-associated decline of viable cell number (B) and the range of linearity of viable cell number at Day 15.
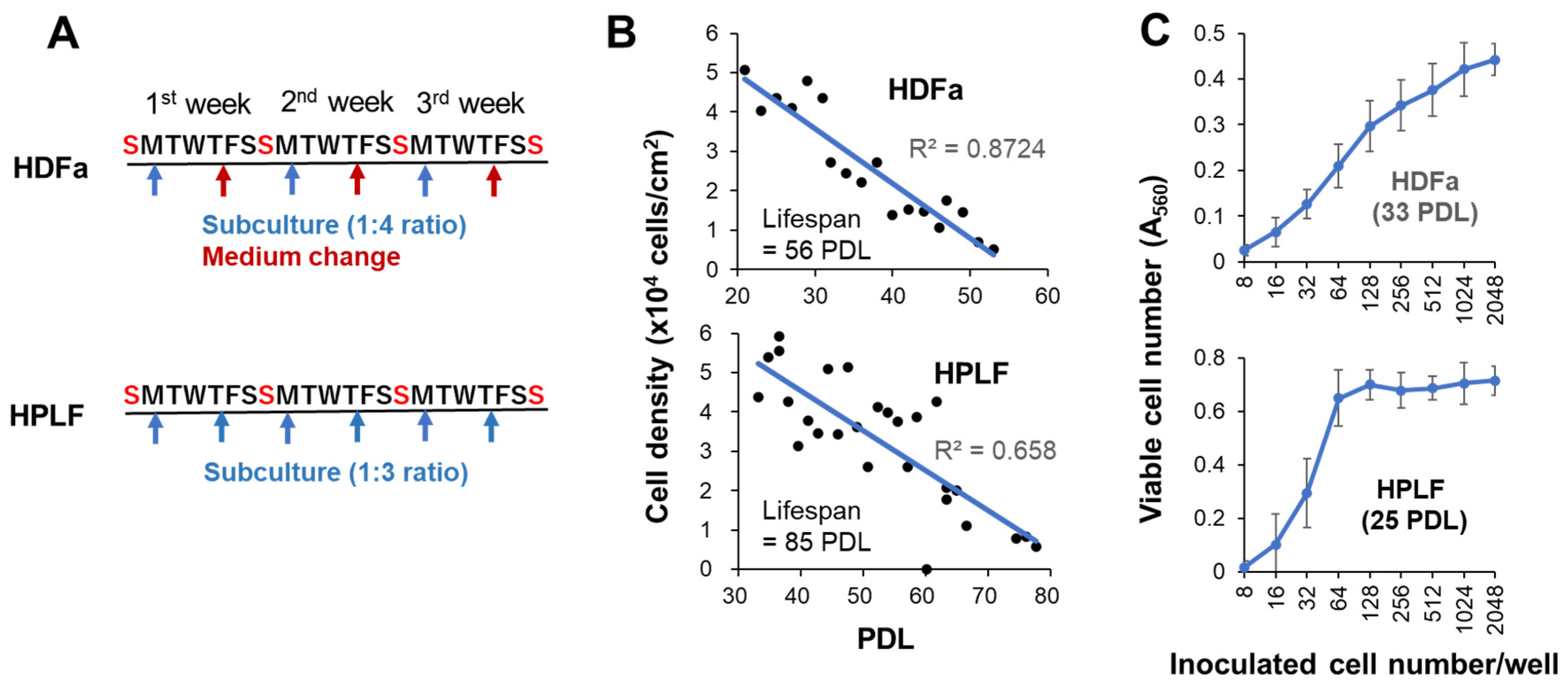
Figure 3.
The determination of replicative lifespan elongation (RLE) by updated “overlay method”. Cells were inoculated into the inner 60 wells of a 96-microwell plate, and the test samples were treated for 14 days without (control) or with increasing concentrations of samples from the left to right (n=6), with the sample layer added during the incubation. After culturing for two weeks, the viable cell count was measured (A). Since cells grow more vigorously closer to the center of the plate, the variation in cell number due to location was corrected for in each experiment (B and insert).
Figure 3.
The determination of replicative lifespan elongation (RLE) by updated “overlay method”. Cells were inoculated into the inner 60 wells of a 96-microwell plate, and the test samples were treated for 14 days without (control) or with increasing concentrations of samples from the left to right (n=6), with the sample layer added during the incubation. After culturing for two weeks, the viable cell count was measured (A). Since cells grow more vigorously closer to the center of the plate, the variation in cell number due to location was corrected for in each experiment (B and insert).
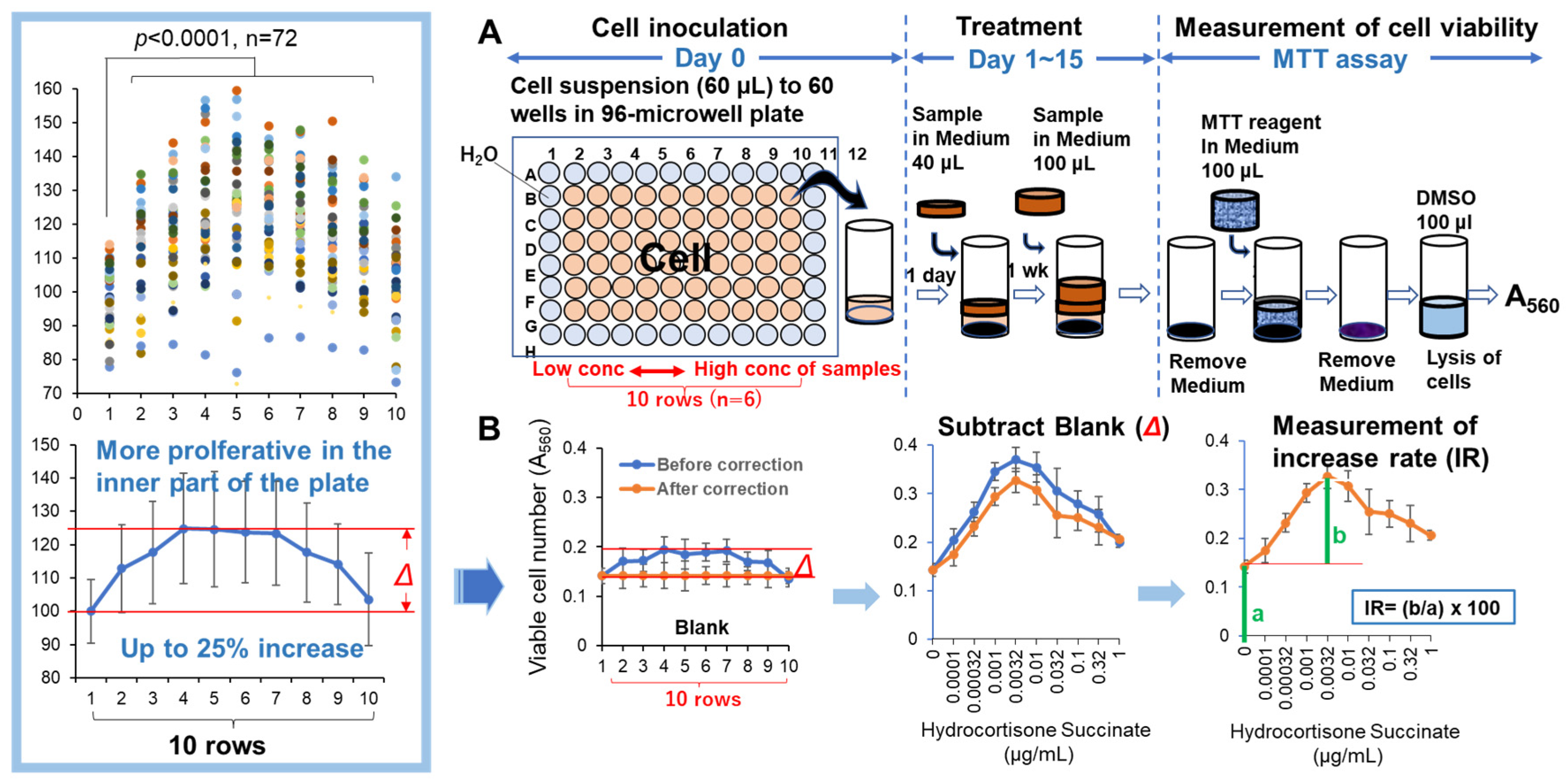
Figure 4.
RLE activity of hormones against human dermal (HDFa) and periodontal ligament fibroblasts (HPLF). Each point represents mean ± S.D. (n=6). * p < 0.05 vs. control (Bonferroni’s post-test). RLE values are presented in Table S1A.
Figure 4.
RLE activity of hormones against human dermal (HDFa) and periodontal ligament fibroblasts (HPLF). Each point represents mean ± S.D. (n=6). * p < 0.05 vs. control (Bonferroni’s post-test). RLE values are presented in Table S1A.
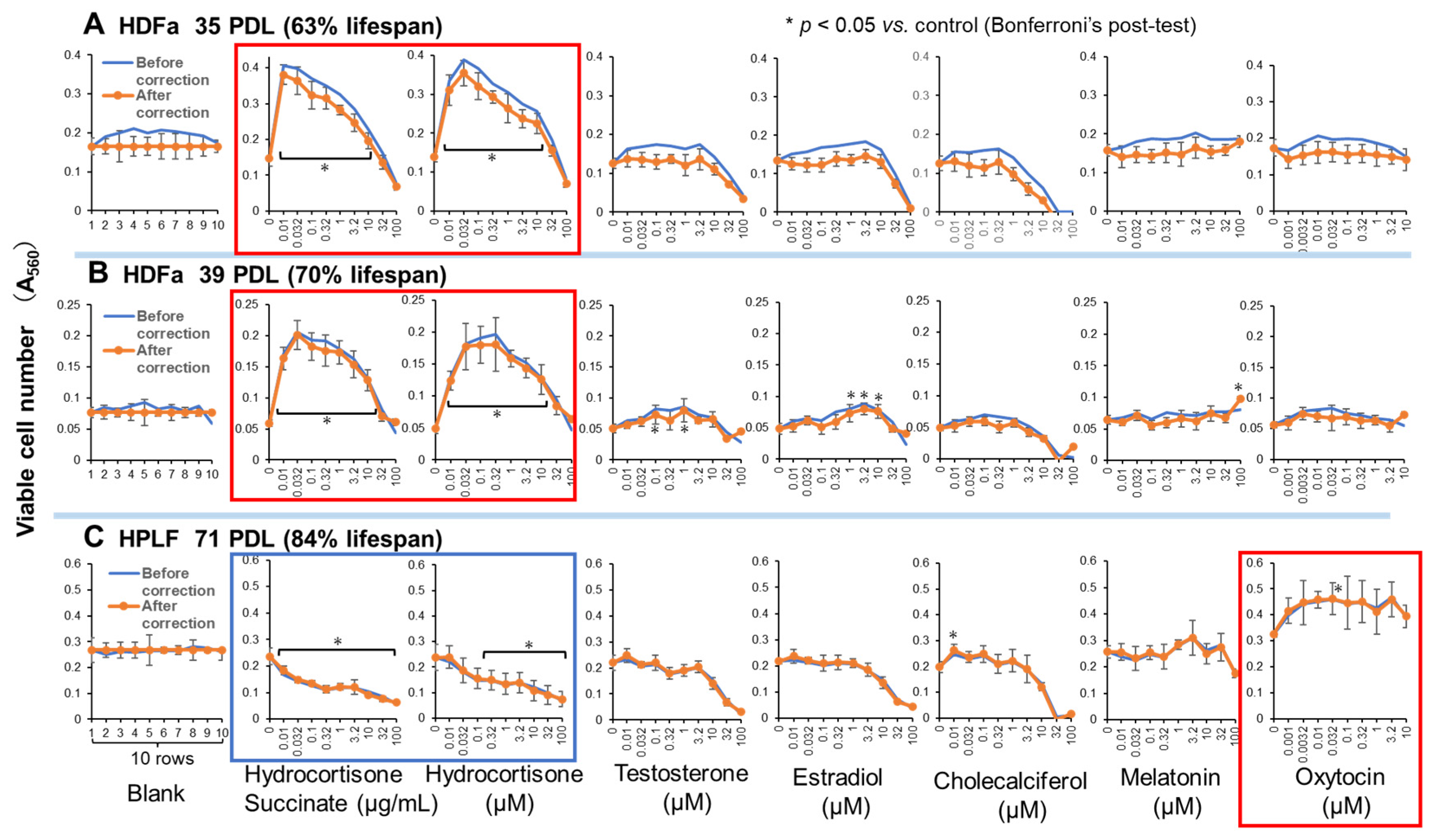
Figure 5.
RLE activity of phenylpropanoids and vanilloids on HDFa (A, B) and HPLF (C). Each point represents mean ± S.D. (n=6). * p < 0.05 vs. control (Bonferroni’s post-test).
Figure 5.
RLE activity of phenylpropanoids and vanilloids on HDFa (A, B) and HPLF (C). Each point represents mean ± S.D. (n=6). * p < 0.05 vs. control (Bonferroni’s post-test).
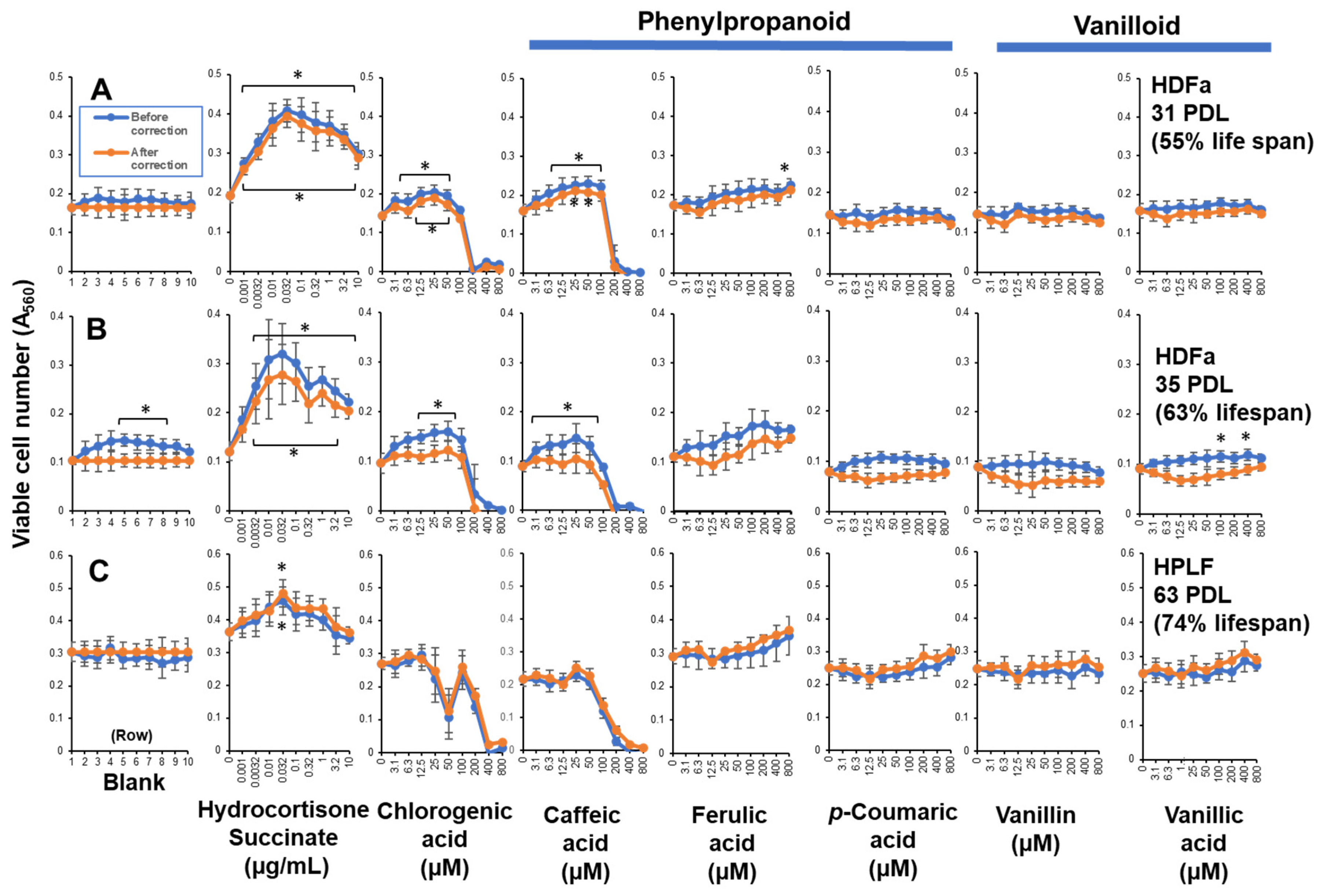
Figure 6.
RLE activity of bacterial culture supernatants. Each point represents mean ± S.D. (n=6). * p < 0.05 vs. control (Bonferroni’s post-test). RLE values are presented in Table S1E.
Figure 6.
RLE activity of bacterial culture supernatants. Each point represents mean ± S.D. (n=6). * p < 0.05 vs. control (Bonferroni’s post-test). RLE values are presented in Table S1E.
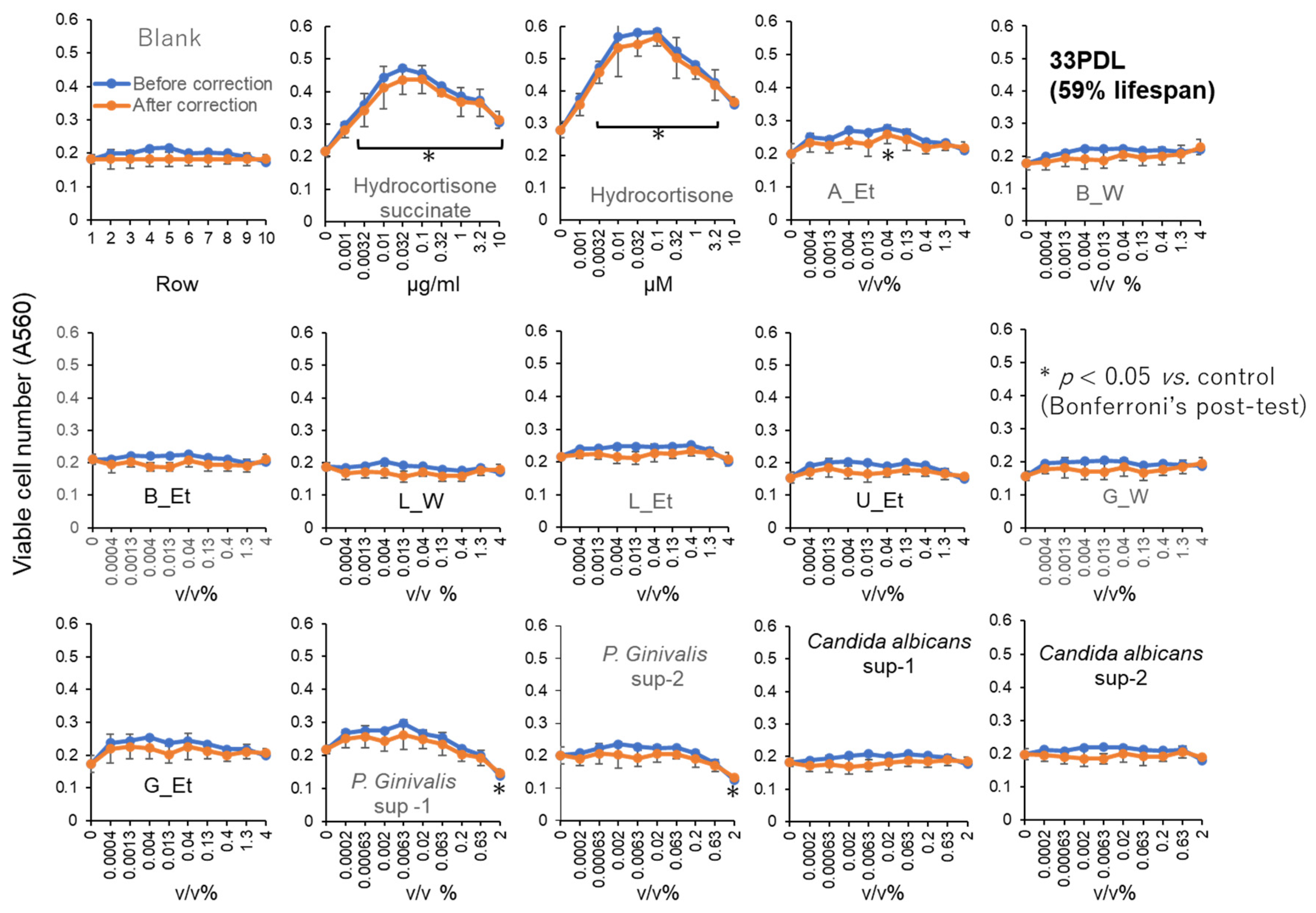
Figure 7.
Relative potency of RLE activities of test samples on HDFa and HPLF. These data were derived from Figure 4, Figure 5 and Figure 6, and supplemental Figure S3 and S4, Table S1. Exact values (except for bacterial sections) are the mean of two independent experiments (each performed with 6 technical replicates). Data of antioxidants and plant extract were cited from [16] and [11], respectively, with permission.
Figure 7.
Relative potency of RLE activities of test samples on HDFa and HPLF. These data were derived from Figure 4, Figure 5 and Figure 6, and supplemental Figure S3 and S4, Table S1. Exact values (except for bacterial sections) are the mean of two independent experiments (each performed with 6 technical replicates). Data of antioxidants and plant extract were cited from [16] and [11], respectively, with permission.
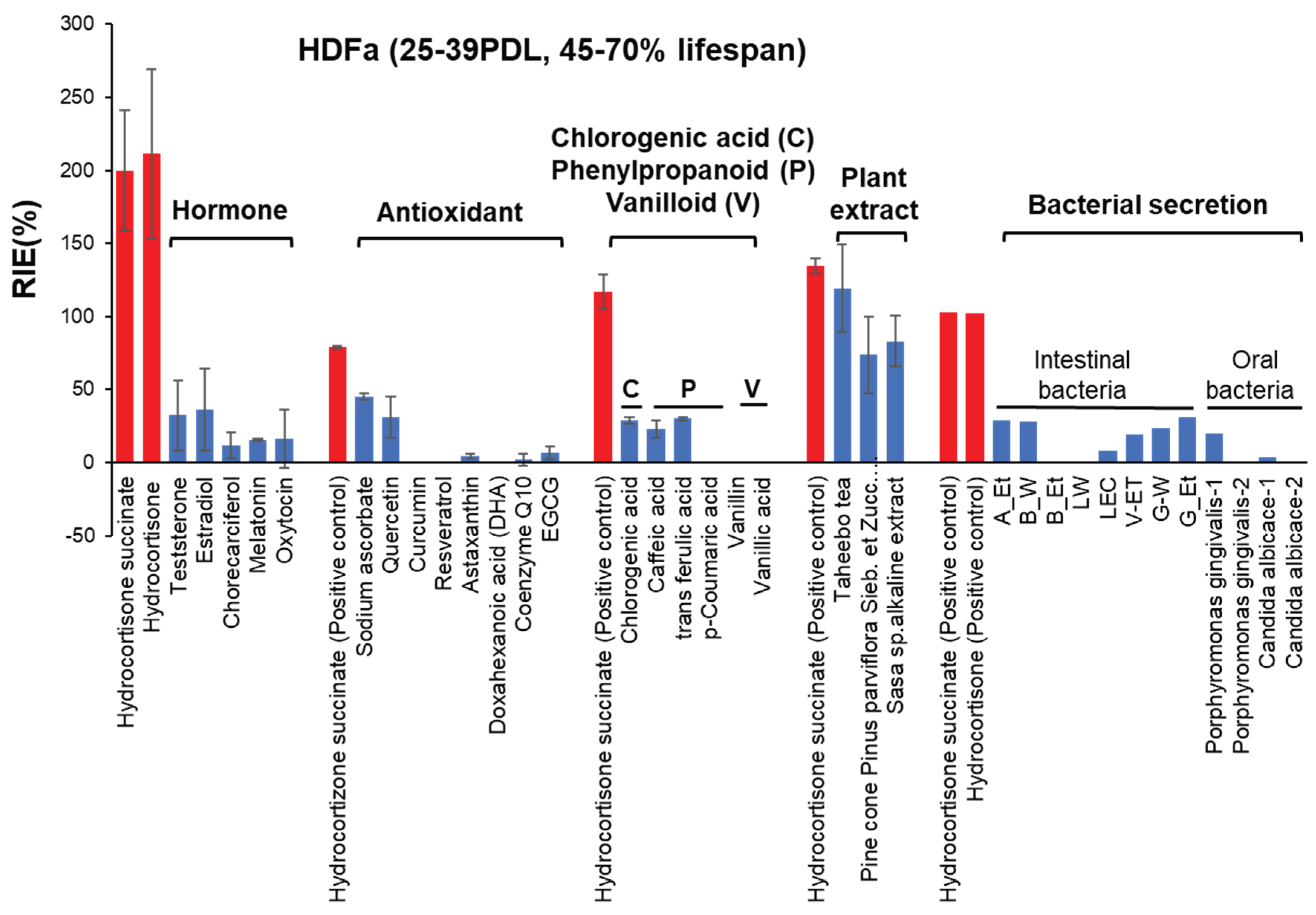

Table 1.
Culture and extraction conditions of bacteria used in the present study.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



