Submitted:
19 December 2025
Posted:
23 December 2025
You are already at the latest version
Abstract
Immuno modulation and metabolism are crucial for survival, with host metabolites and microbiota influencing immune responses. This study explores the role of immune stimulation on insulin sensitivity, lipid management, and glucose regulation, highlighting tissue-specific effects in muscles, liver, adipose tissue, and the gut. It also examines human milk-derived bioactives on early metabolism and immunity, as well as how microbial substrates, postbiotics, and dysbiosis impact immune function and contribute to metabolic diseases, including obesity and inflammation. Prenatal immunometabolism changes significantly impact pregnancy outcomes and are linked to long-term metabolic issues. This review explores various factors, including the microbiome, autophagy, epigenetics, organokine signaling, and immunological dysfunction from undernutrition, along with advancements in metabolomics. It assesses therapeutic strategies aimed at restoring metabolic-immune balance, particularly focusing on anti-inflammatory and nutritional interventions such as IL-1β antagonism, omega-3 fatty acids, and intermittent fasting. The findings highlight the importance of understanding immune-metabolic interactions to improve health and develop personalized treatments for metabolic syndrome and related disorders.
Keywords:
Introduction
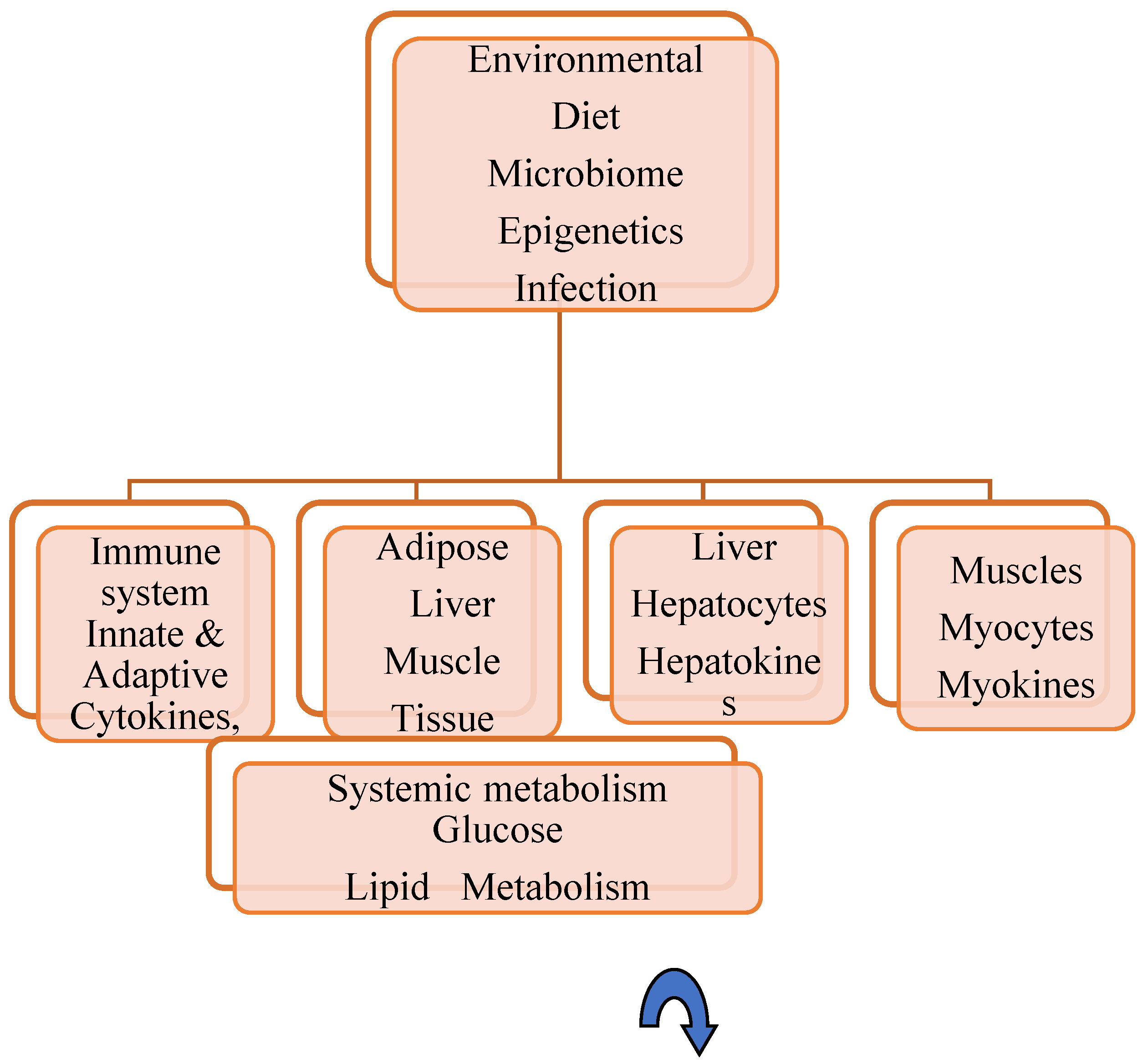
Metabolic–Immune Hemostasis
Human Milk-Based Neonatal Metabolic- Immune Programming
Pregnancy Immunometabolic Outcomes
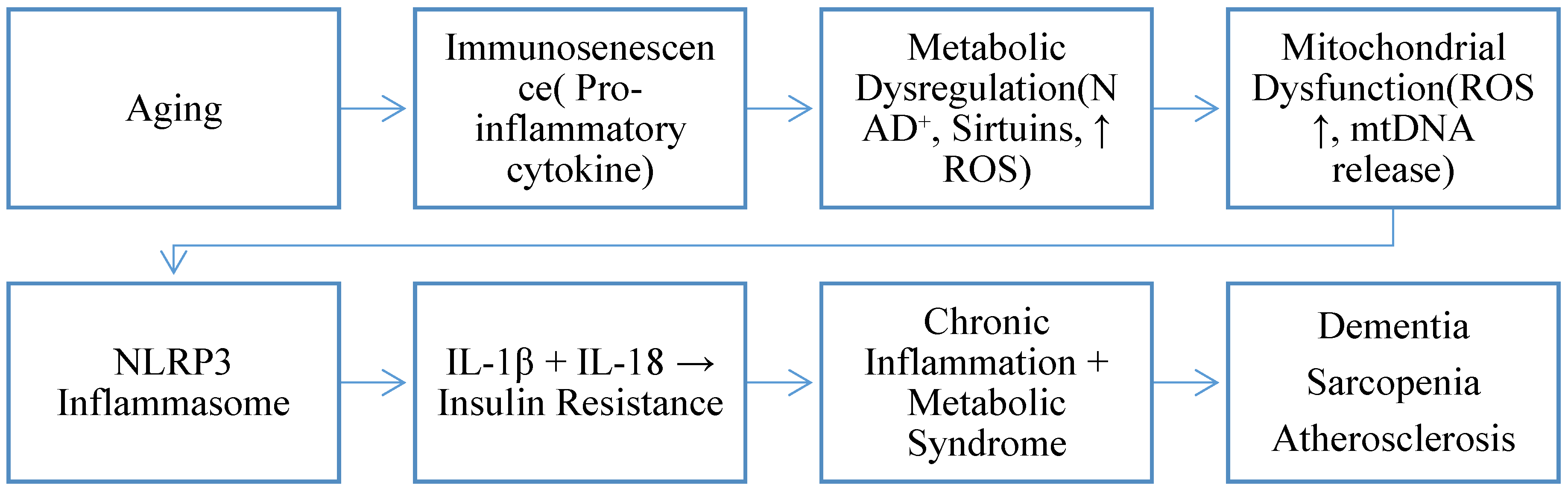
Aging-Related Immune–Metabolic Stress
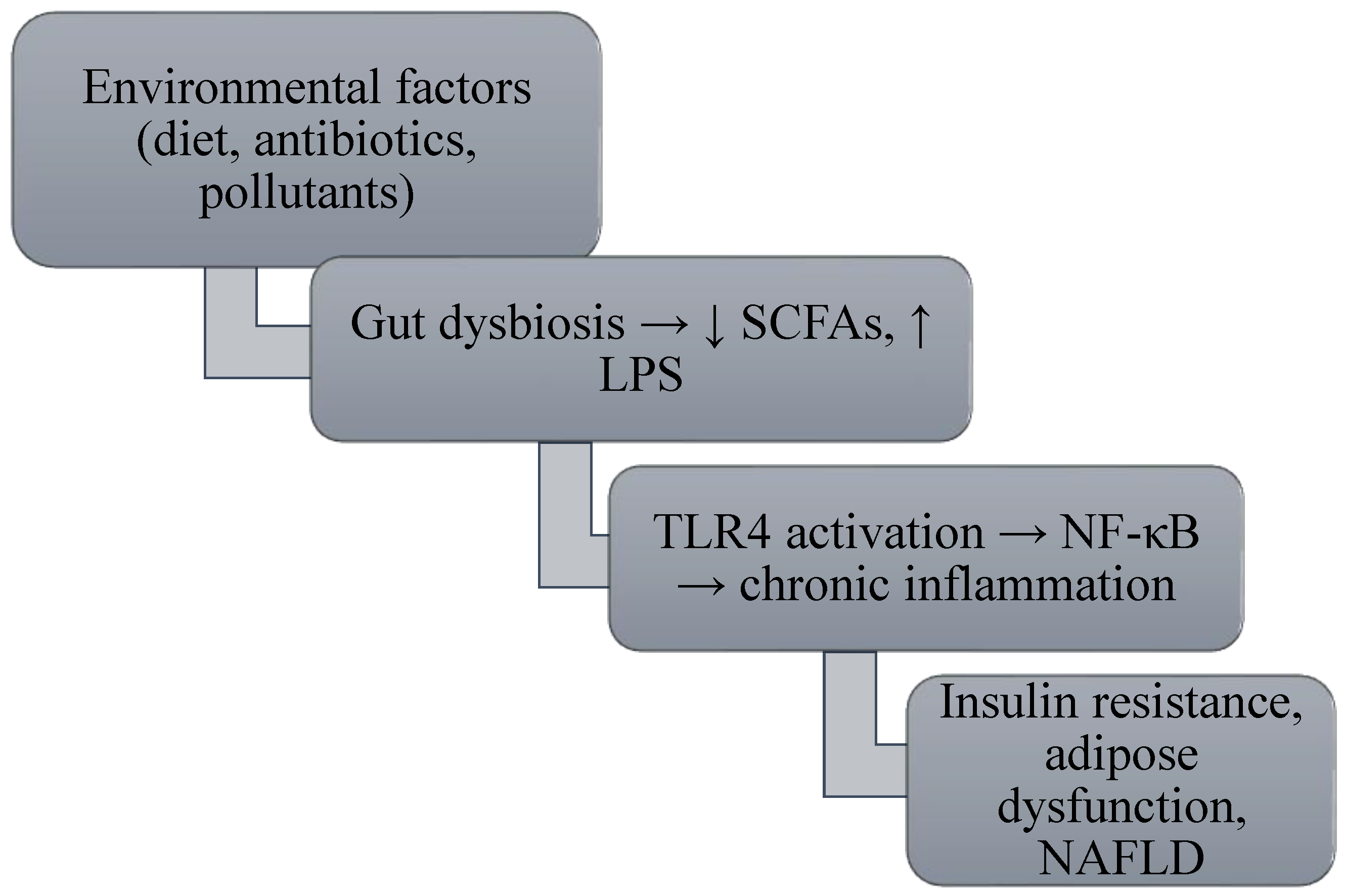
Microbiota–Autophagy Dynamics
Genomic–Epigenetic Immunometabolism
Immunometabolic Adipose Stress
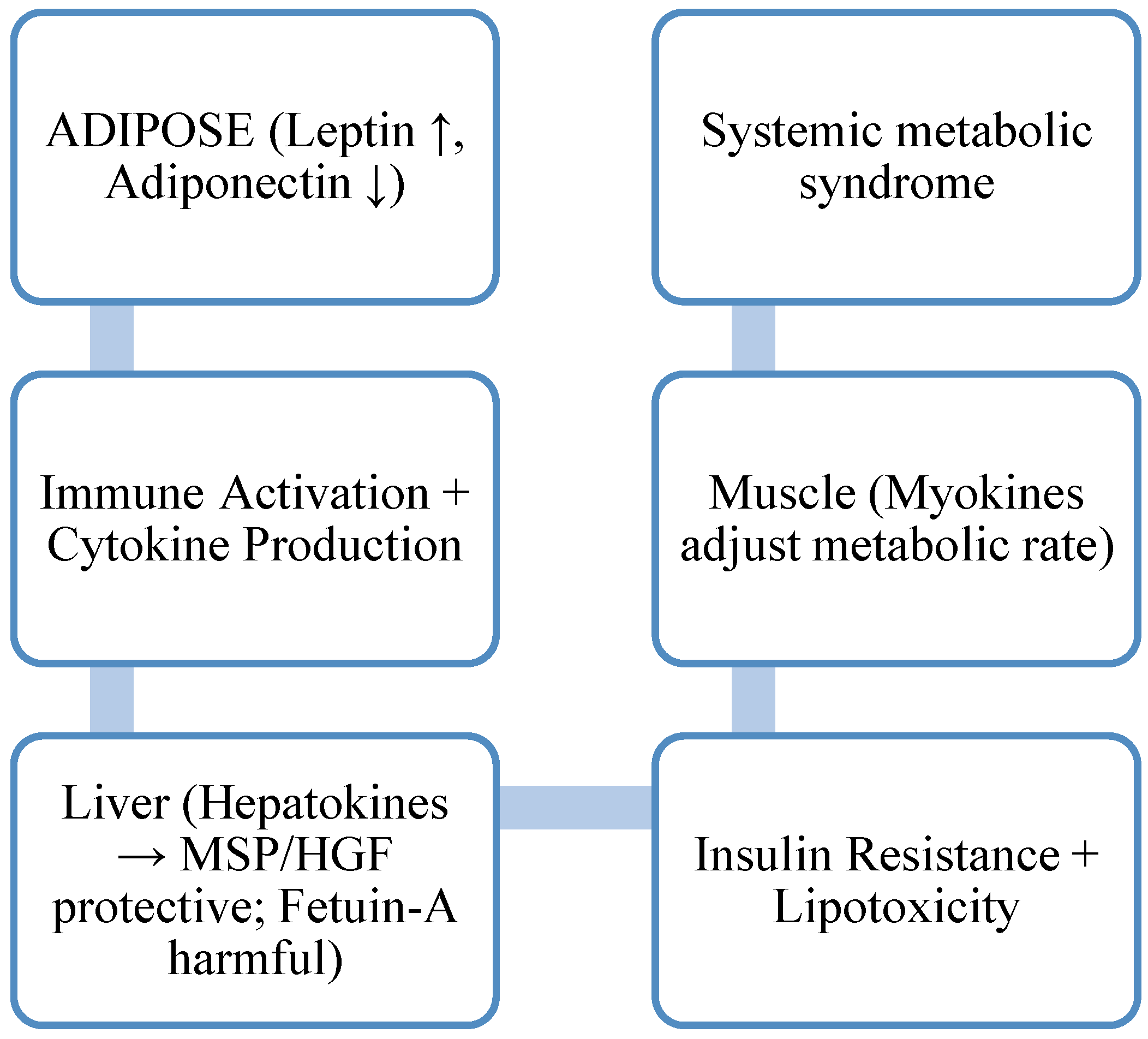
Organs' Endocrine Interaction in Metabolic Syndrome
Childhood Cachexia and Immune Impact
Precision Metabolomics in Malnutrition
Translating Anti-Inflammatory Strategies into Therapy
Key Insights and Emerging Directions
References
- Arrieta, M.-C.; Stiemsma, L. T.; Amenyogbe, N.; Brown, E. M.; Finlay, B. The intestinal microbiome in early life: Health and disease. Frontiers in Immunology 2014, 5, 427. [Google Scholar] [CrossRef] [PubMed]
- Amin, T.; Mercer, J. G. Hunger and satiety mechanisms and their potential exploitation in the regulation of food intake. Current Obesity Reports 2016, 5, 106–112. [Google Scholar] [CrossRef]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P. M. Maternal immunological adaptation during normal pregnancy. Frontiers in Immunology 2020, 11(7), 11, [DOI not found]. [Google Scholar] [CrossRef]
- Alzaid, F.; Fagherazzi, G.; Riveline, J. P.; et al. Immune cell–adipose tissue crosstalk in metabolic diseases with a focus on type 1 diabetes. Diabetologia 2025, 68(?), 1616–1631. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A. L. Human milk composition: Nutrients and bioactive factors. Pediatric Clinics of North America 2013, 60(1), 49–74. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host & Microbe 2015, 17(5), 690–703. [Google Scholar] [CrossRef]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22(9), 1147–1162. [Google Scholar] [CrossRef]
- Brandtzaeg, P. Secretory IgA: Designed for anti-microbial defense. Frontiers in Immunology 2013, 4, 222. [Google Scholar] [CrossRef]
- Buck, M. D.; Sowell, R. T.; Kaech, S. M.; Pearce, E. L. Metabolic instruction of immunity. Cell 2015, 161(1), 63–76. [Google Scholar] [CrossRef]
- Baffy, G. Kupffer cells in non-alcoholic fatty liver disease: The emerging view. Journal of Hepatology 2009, 51(1), 212–223. [Google Scholar] [CrossRef] [PubMed]
- Bindra, R.; Thakur, P.; Hussain, M. E. Role of inflammatory signaling pathways in obesity-induced insulin resistance. Journal of Molecular Endocrinology 2013, 51(3), R157–R171. [Google Scholar] [CrossRef]
- Becker, T.; Loch, G.; Beyer, M.; Zinke, I.; Aschenbrenner, A. C.; Carrera, P.; Inhester, T.; Schultze, J. L.; Hoch, M. FOXO-dependent regulation of innate immune homeostasis. Nature 2010, 463(7279), 369–373. [Google Scholar] [CrossRef]
- Cavelti-Weder, C.; Babians-Brunner, A.; Keller, C.; Stahel, M. A.; Kurz-Levin, M.; Zayed, H.; Donath, M. Y. Effects of IL-1 antagonism on glycemic control in type 2 diabetes: A randomized, double-blind, placebo-controlled trial. Diabetes Care 2012, 35(10), 2069–2076. [Google Scholar]
- Cox, L. M.; Blaser, M. J. Antibiotics in early life and obesity. Nature Reviews Endocrinology 2015, 11(3), 182–190. [Google Scholar] [CrossRef]
- Canani, R. B.; Cirillo, P.; Terrin, G. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World Journal of Gastroenterology 2011, 17(12), 1519–1528. [Google Scholar] [CrossRef]
- Canfora, E. E.; Jocken, J. W.; Blaak, E. E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nature Reviews Endocrinology 2015, 11(10), 577–591. [Google Scholar] [CrossRef]
- Carrasco, D.; Del Campo, R.; Castillo, F. Polyamines in human milk and their influence on infant growth and development. Nutrients 2018, 10(8), 1065. [Google Scholar] [CrossRef]
- Camacho-Morales, A.; Caba, M.; García-Juárez, M.; Caba-Flores, M. D.; Viveros-Contreras, R.; Martínez-Valenzuela, C. Breastfeeding contributes to physiological immune programming in the newborn. Frontiers in Pediatrics 2021, 9, 744104. [Google Scholar] [CrossRef] [PubMed]
- Carr, L. E.; Weir, T. L.; Sanderson, C. J. Role of human milk bioactives on infants’ gut and immune system maturation. Frontiers in Immunology 2021, 12, 604080. [Google Scholar] [CrossRef]
- Ciastek, B.; Kapłon, K.; Domaszewski, P. A comprehensive perspective on the biological effects of intermittent fasting and periodic short-term fasting: A promising strategy for optimizing metabolic health. Nutrients 2025, 17(13), 2061. [Google Scholar] [CrossRef] [PubMed]
- de la Calle-Fabregat, C.; Morante-Palacios, O.; Ballestar, E. Understanding the relevance of DNA methylation changes in immune differentiation and disease. Genes 2020, 11(1), 110. [Google Scholar] [CrossRef] [PubMed]
- Dhondge, R. H.; Agrawal, S.; Patil, R.; Kadu, A.; Kothari, M. A comprehensive review of metabolic syndrome and its role in cardiovascular disease and type 2 diabetes mellitus: Mechanisms, risk factors, and management. Cureus 2024, 16(8), e67428. [Google Scholar] [CrossRef]
- Donovan, S. M.; Comstock, S. S. Human milk oligosaccharides influence neonatal mucosal and systemic immunity. Annals of Nutrition & Metabolism 2016, 69 Suppl. 2, 42–51. [Google Scholar] [CrossRef]
- Donath, M. Y. Targeting inflammation in the treatment of type 2 diabetes: Time to start. Nature Reviews Drug Discovery 2014, 13(6), 465–476. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Abdellatif, M.; Zimmermann, A. Dietary spermidine for lowering blood pressure and extending lifespan. Nature Medicine 2016, 22(12), 1428–1438. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune–metabolic viewpoint for age-related diseases. Nature Reviews Endocrinology 2018, 14(10), 576–590. [Google Scholar] [CrossRef]
- Ferrannini, E.; Balkau, B.; Coppack, S. W.; Dekker, J.; Mari, A.; Nolan, J.; Walker, M.; Natali, A. Effect of tumor necrosis factor-α neutralization on insulin sensitivity in humans. Diabetes Care 2007, 30(10), 2489–2494. [Google Scholar]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. Journals of Gerontology Series A: Biological Sciences and Medical Sciences 2014, 69 Suppl 1, S4–S9. [Google Scholar] [CrossRef]
- Fang, H.; Rodrigues E-Lacerda, R.; Barra, N. G.; Zada, D. K.; Robin, N.; Mehra, A.; Schertzer, J. D. Postbiotic impact on host metabolism and immunity provides therapeutic potential in metabolic disease. Endocrine Reviews 2025, 46(1), 60–79. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. Journal of Allergy and Clinical Immunology 2005, 115(5), 911–919. [Google Scholar] [CrossRef]
- Greenberg, A. S.; Obin, M. S. Obesity and the role of adipose tissue in inflammation and metabolism. American Journal of Clinical Nutrition 2006, 83(2), 461S–465S. [Google Scholar] [CrossRef]
- Guimarães, G. R.; Almeida, P. P.; Santos, L. de O.; Rodrigues, L. P.; de Carvalho, J. L.; Boroni, M. Hallmarks of aging in macrophages: Consequences to skin inflammaging. Cells 2021, 10(6), 1323. [Google Scholar] [CrossRef]
- Gerede, A.; Danavasi, M.; Stavros, S.; Potiris, A.; Zikopoulos, A.; Moustakli, E.; Skentou, C.; Domali, E.; Nikolettos, N.; Eleftheriades, M. Obesity and pregnancy: Impact on childbirth timing, delivery mode, and maternal recovery: An update. Medical Sciences 2025, 13(3), 182. [Google Scholar] [CrossRef]
- Géminard, C.; Rulifson, E. J.; Léopold, P. Remote control of insulin secretion by fat cells in Drosophila. Cell Metabolism 2009, 10(3), 199–207. [Google Scholar] [CrossRef] [PubMed]
- Goldfine, A. B.; Silver, R.; Aldhahi, W.; Cai, D.; Tatro, E.; Lee, J.; Shoelson, S. E. Use of salsalate to target inflammation in the treatment of insulin resistance and type 2 diabetes. Clinical and Translational Science 2008, 1(1), 36–43. [Google Scholar] [CrossRef] [PubMed]
- Gopal, P. K.; Merenstein, D.; Ouwehand, A. Glycoconjugates in human milk: Implications for the neonatal gut microbiota. Current Opinion in Biotechnology 2008, 19(2), 150–157. [Google Scholar] [CrossRef]
- Gopalakrishna, K. P.; Hand, T. W. Influence of maternal milk on the neonatal intestinal microbiome. Gut Microbes 2020, 11(3), 1–20. [Google Scholar] [CrossRef]
- Gregor, M. F.; Hotamisligil, G. S. Inflammatory mechanisms in obesity. Annual Review of Immunology 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed]
- Hennet, T.; Borsig, L. Effects of host glycosylation on gut microbiota. Nature Reviews Gastroenterology & Hepatology 2016, 13(5), 259–271. [Google Scholar] [CrossRef]
- Haffner, S. M.; Greenberg, A. S.; Weston, W. M.; Chen, H.; Williams, K.; Freed, M. I. Effect of rosiglitazone on nontraditional cardiovascular disease markers. Circulation 2002, 106(6), 679–684. [Google Scholar] [CrossRef]
- Hirosumi, J.; Tuncman, G.; Chang, L. A central role for JNK in obesity and insulin resistance. Nature 2002, 420(6913), 333–336. [Google Scholar] [CrossRef]
- Hotamisligil, G. S. Inflammation and metabolic disorders. Nature 2006, 444(7121), 860–867. [Google Scholar] [CrossRef]
- Hofer, S. J.; Daskalaki, I.; Bergmann, M.; Friščić, J.; Zimmermann, A.; Mueller, M. I.; Abdellatif, M.; Nicastro, R.; Masser, S.; Durand, S.; Nartey, A.; Waltenstorfer, M.; Enzenhofer, S.; Faimann, I.; Gschiel, V.; Bajaj, T.; Niemeyer, C.; Gkikas, I.; Pein, L.; Cerrato, G.; Pan, H.; Liang, Y.; Tadic, J.; Jerkovic, A.; Madeo, F. Spermidine is essential for fasting-mediated autophagy and longevity. Nature Cell Biology 2024, 26, 1571–1584. [Google Scholar] [CrossRef]
- Hu, T.; Liu, C.-H.; Lei, M.; Zeng, Q.; Li, L.; Tang, H.; Zhang, N. Metabolic regulation of the immune system in health and diseases: Mechanisms and interventions. Signal Transduction and Targeted Therapy 2024, 9, 268. [Google Scholar] [CrossRef]
- Hu, C.; Xuan, Y.; Zhang, X.; Liu, Y.; Yang, S.; Yang, K. Immune cell metabolism and metabolic reprogramming. Molecular Biology Reports 2022, 49(10), 9783–9795. [Google Scholar] [CrossRef]
- Hotamisligil, G. S. Inflammation and metabolic disorders. Nature 2006, 444(7121), 860–867. [Google Scholar] [CrossRef]
- Kim, S.; Goel, R.; Kumar, A. Imbalance in gut microbiota contributes to hypertension in hypertensive rats: The role of gut inflammation. Physiological Genomics 2018, 50(6), 1–13. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165(6), 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Kazankov, K.; Jørsboe, E.; Andersen, K. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nature Reviews Gastroenterology & Hepatology 2019, 16, 145–159. [Google Scholar] [CrossRef]
- Khalafi, M.; Habibi Maleki, A.; Mojtahedi, S.; Fernandez, M. L.; et al. The effects of intermittent fasting on inflammatory markers in adults: A systematic review and pairwise and network meta-analyses. Nutrients 2025, 17(15), 2388. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Mishra, A.; Gautam, P.; Feroz, Z.; Vijayaraghavalu, S.; Likos, E. M.; Shukla, G. C.; Kumar, M. Metabolic pathways, enzymes, and metabolites: Opportunities in cancer therapy. Cancers 2022, 14(21), 5268. [Google Scholar] [CrossRef]
- Oh, D. Y.; Talukdar, S.; Bae, E. J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W. J.; Watkins, S. M.; Olefsky, J. M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142(5), 687–698. [Google Scholar] [CrossRef]
- Jordan, S.; Tung, N.; Casanova-Acebes, M.; Chang, C.; Cantoni, C.; Zhang, D.; Wirtz, T. H.; Naik, S.; Rose, S. A.; Brocker, C. N.; Savage, P. A.; Yewdell, J. W. Dietary intake regulates the circulating inflammatory monocyte pool. Cell 2019, 178(5), 1102–1114.e17. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and aging: Signaling pathways and intervention therapies. Signal Transduction and Targeted Therapy 2023, 8, 239. [Google Scholar] [CrossRef]
- Mosca, A.; Panera, N.; Maggiore, G.; Alisi, A. From pregnant women to infants: Non-alcoholic fatty liver disease is a poor inheritance. Journal of Pediatric Gastroenterology and Nutrition 2020, 73(6), 1590–1592. [Google Scholar] [CrossRef]
- Nagano, N.; Morioka, I. Glucose and lipid metabolic aberrations in preterm and low birth weight infants: Associations with lifestyle-related diseases. In Pediatrics International; Advance online publication, 2025. [Google Scholar] [CrossRef]
- Nguyen, K. H. H.; Le, N. V.; Nguyen, P. H.; Nguyen, H. H. T.; Hoang, D. M.; Huynh, C. D. Human immune system: Exploring diversity across individuals and populations. Heliyon 2025, 11(2), e41836. [Google Scholar] [CrossRef] [PubMed]
- Kalupahana, N. S.; Claycombe, K. J.; Moustaid-Moussa, N. Anti-inflammatory and metabolic effects of omega-3 fatty acids. Annual Review of Nutrition 2011, 31, 153–176. [Google Scholar]
- Larsen, C. M.; Faulenbach, M.; Vaag, A.; Ehses, J. A.; Donath, M. Y.; Mandrup-Poulsen, T. Interleukin-1 receptor antagonist in type 2 diabetes mellitus. New England Journal of Medicine 2007, 356(15), 1517–1526. [Google Scholar] [CrossRef]
- Liu, C.; Li, X. Role of leptin and adiponectin in immune response and inflammation. International Immunopharmacology 2025, 161, 115082. [Google Scholar] [CrossRef]
- Lumeng, C. N.; Saltiel, A. R. Inflammatory links between obesity and metabolic disease. Journal of Clinical Investigation 2011, 121(6), 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Lokossou, G. A. G.; Kouakanou, L.; Schumacher, A.; Zenclussen, A. C. Human breast milk: From food to active immune response with disease protection in infants and mothers. Frontiers in Immunology 2022, 13, 849012. [Google Scholar] [CrossRef]
- Le Cosquer, G.; Vergnolle, N.; Motta, J.-P. Gut microb-aging and its relevance to frailty aging. Microbes and Infection 2024, 26(3), Article 105309. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Cai, D. Defective hypothalamic autophagy directs the central pathogenesis of obesity via the IKKβ/NF-κB pathway. Cell 2011, 146(1), 47–59. [Google Scholar] [CrossRef]
- Mishra, P. K.; Talukdar, I.; Balaji, K. N. Environmental pollutants and the gut microbiome: An emerging threat to immune homeostasis and metabolic health. Frontiers in Immunology 2021, 12, 700656. [Google Scholar] [CrossRef]
- Maedler, K.; Sergeev, P.; Ris, F.; et al. IL-1β triggers β-cell apoptosis via NF-κB activation and NO production. Diabetes 2002, 51(4), 1086–1095. [Google Scholar] [CrossRef]
- Ofei, F.; Hurel, S.; Newkirk, J.; Sopwith, M.; Taylor, R. Effects of an engineered human anti–TNF-α antibody on insulin sensitivity and glycemic control in NIDDM. Diabetes 1996, 45(7), 881–885. [Google Scholar] [CrossRef]
- Matsushima, G.; Yanase, Y.; Nakagawa, T.; Goda, M.; Ozawa, K.; Hosoi, T. Endoplasmic reticulum stress and unfolded protein response in immune cell function. Frontiers in Immunology 2025, 16, 1694102. [Google Scholar] [CrossRef] [PubMed]
- McNelis, K., Yodoshi, T., Divanovic, S., Gandhi, C., Kim, J. H., Anton, C. G., Trout, A. T., & Mouzaki, M. (2021). Hepatic steatosis in infancy: The beginning of pediatric nonalcoholic fatty liver disease? JPGN Reports, 2(3), e113. Meo, S. A., & Hassain, A. (2016). Metabolic physiology in pregnancy. Journal of the Pakistan Medical Association, 66(9 Suppl 1), S8–S10.
- Moldenhauer, L. M.; Hull, M. L.; Foyle, K. L.; McCormack, C. D.; Robertson, S. A. Immune-metabolic interactions and T cell tolerance in pregnancy. Journal of Immunology 2022, 209(8), 1426–1436. [Google Scholar] [CrossRef]
- Ma, Y. Mitochondrial dysfunction and inflammation in metabolic disorders. Journal of Molecular Medicine 2020, 98(6), 781–789. [Google Scholar] [CrossRef]
- Madeo, F.; Carmona-Gutierrez, D.; Hofer, S. J.; Kroemer, G. Spermidine in health and disease. Science 2018, 359(6374), eaan2788. [Google Scholar] [CrossRef]
- Man, A. L.; Bertelli, E.; Rentini, S.. Age-associated modifications of intestinal barrier function and gut microbiota in mice. Aging Cell 2015, 14(6), 1093–1103. [Google Scholar] [CrossRef]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose tissue in obesity-related inflammation and insulin resistance: Cells, cytokines, and chemokines. ISRN Inflammation 2013, 2013, 139239. [Google Scholar] [CrossRef]
- Meex, R. C. R.; Watt, M. J. Hepatokines: Linking nonalcoholic fatty liver disease and insulin resistance. Nature Reviews Endocrinology 2017, 13(9), 509–520. [Google Scholar] [CrossRef]
- Ramos-Lopez, O.; Milagro, F. I.; Riezu-Boj, J. I.; Martínez, J. A. Epigenetic signatures underlying inflammation: An interplay of nutrition, physical activity, metabolic diseases, and environmental factors for personalized nutrition. Inflammation Research 2021, 70, 29–49. [Google Scholar] [CrossRef]
- O’Toole, P. W.; Jeffery, I. B. Gut microbiota and aging. Science 2015, 350(6265), 1214–1215. [Google Scholar] [CrossRef]
- Olszak, T.; An, D.; Zeissig, S. Microbial exposure during early life regulates the immune system via the microbiota. Science 2012, 336(6080), 489–493. [Google Scholar] [CrossRef] [PubMed]
- Newburg, D. S.; Walker, W. A. Protection of the neonate by the innate immune system of developing gut and of human milk. Pediatric Research 2007, 61(1), 2–8. [Google Scholar] [CrossRef] [PubMed]
- Najjar, R. S. The impacts of animal-based diets in cardiovascular disease development: A cellular and physiological overview. Journal of Cardiovascular Development and Disease 2023, 10(7), 282. [Google Scholar] [CrossRef]
- O’Neill, L. A. J.; Pearce, E. L. Immunometabolism governs dendritic cell and macrophage function. Nature Reviews Immunology 2016, 16(9), 553–565. [Google Scholar] [CrossRef] [PubMed]
- Oh, D. Y.; Talukdar, S.; Bae, E. J.; Imamura, T.; Morinaga, H.; Fan, W.; Olefsky, J. M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142(5), 687–698. [Google Scholar] [CrossRef]
- Rodríguez-Morales, P.; Franklin, R. A. Macrophage phenotypes and functions: Resolving inflammation and restoring homeostasis. Trends in Immunology 2023, 44(12), 986–998. [Google Scholar] [CrossRef]
- Ryan, D. G.; O’Neill, L. A. J. Krebs cycle rewired for macrophage and dendritic cell effector functions. Nature Reviews Immunology 2020, 20(6), 425–437. [Google Scholar] [CrossRef]
- Sajdel-Sulkowska, E. M. The impact of maternal gut microbiota during pregnancy on fetal gut–brain axis development and life-long health outcomes. Microorganisms 2023, 11(9), 2199, [DOI not given in your list — DOI not found]. [Google Scholar] [CrossRef] [PubMed]
- Salinas, M. L.; Mulakala, B. K.; Davidson, L. A.; Cai, J. J.; Donovan, S. M.; Chapkin, R. S.; Yeruva, L. Single-cell transcriptomics reveals that human milk feeding shapes neonatal immune cell interleukin signaling pathways in a nonrandomized clinical trial. The American Journal of Clinical Nutrition 2025, 122(1), 196–207, [DOI not given — DOI not found]. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Rodrigues, P. R. S.; Zaher, S.; Davies, L. C.; Ghazal, P. Immune-metabolic adaptations in pregnancy: A potential stepping-stone to sepsis. eBioMedicine 2022, 86, 104337. [Google Scholar] [CrossRef]
- Stern, J. H.; Rutkowski, J. M.; Scherer, P. E. Adiponectin, leptin, and fatty acids in the maintenance of metabolic homeostasis through adipose tissue crosstalk. Cell Metabolism 2016, 23(5), 770–784. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Zheng, M.; Liang, W.; Ouyang, L.; Wang, S. 2′-Fucosyllactose synbiotics with Bifidobacterium bifidum to improve intestinal transcriptional function and gut microbiota in constipated mice. Food Research International 2025, 217, 116840. [Google Scholar] [CrossRef]
- Shi, H.; Kokoeva, M. V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J. S. TLR4 links innate immunity and fatty acid–induced insulin resistance. Journal of Clinical Investigation 2006, 116(11), 3015–3025. [Google Scholar] [CrossRef]
- Stienstra, R.; Joosten, L. A. B.; Koenen, T. The inflammasome-mediated caspase-1 activation controls adipocyte differentiation and insulin sensitivity. Cell Metabolism 2011, 12(6), 593–605. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Inflammaging: Disturbed interplay between autophagy and inflammasomes. Aging 2012, 4(3), 166–175. [Google Scholar] [CrossRef] [PubMed]
- Thevaranjan, N.; Puchta, A.; Schulz, C.; et al. Age-associated microbial dysbiosis promotes systemic inflammation and inflammaging. Cell Host & Microbe 2017, 21(2), 155–166. [Google Scholar] [CrossRef]
- Parrettini, S.; Caroli, A.; Torlone, E. Nutrition and metabolic adaptations in physiological and complicated pregnancy: Focus on obesity and gestational diabetes. In Frontiers in Endocrinology; Frontiers+1, 2020; Volume 11. [Google Scholar] [CrossRef]
- Tang, W. H. W.; Kitai, T.; Hazen, S. L. Gut microbiota in cardiovascular health and disease. Journal of Clinical Investigation 2017, 127(1), 22–31. [Google Scholar] [CrossRef]
- Vijay-Kumar, M.; Aitken, J. D.; Carvalho, F. A. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science 2010, 328(5975), 228–231. [Google Scholar] [CrossRef]
- Vassilakou, T. Childhood malnutrition: Time for action. Children 2021, 8(2), 103. [Google Scholar] [CrossRef]
- Wang, B.; Brand-Miller, J.; Brockman, R. Maternal milk oligosaccharides, infant gut microbiota, and growth. Nutrients 2021, 13(6), 1921. [Google Scholar] [CrossRef]
- Wensveen, F. M.; Valentić, S.; Šestan, M.; Turk Wensveen, T.; Polić, B. The “Big Bang” in obese fat: Cytokines and adipose tissue expansion. European Journal of Immunology 2019, 49(10), 1467–1476. [Google Scholar] [CrossRef]
- Youm, Y.-H.; Grant, R. W.; McCabe, L. R. Canonical Nlrp3 inflammasome links systemic low-grade inflammation to functional decline in aging. Cell Metabolism 2013, 18(4), 519–532. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J. Y.; Groer, M.; Dutra, S. V. O.; Sarkar, A.; McSkimming, D. I. Gut microbiota and immune system interactions. Microorganisms 2020, 8(10), 1587. [Google Scholar] [CrossRef]
- You, M.; Xie, Z.; Zhang, N.; Zhang, Y.; Xiao, D.; Liu, S.; Zhuang, W.; Li, L.; Tao, Y. Signaling pathways in cancer metabolism: Mechanisms and therapeutic targets. Signal Transduction and Targeted Therapy 2023, 8, 142. [Google Scholar] [CrossRef]
- Yen, F.-S.; Lee, Y.-H.; Kuo, P.; Wu, M.-C.; Leong, P.-Y. Anti-inflammatory mechanisms of antidiabetic therapies: Metabolic regulation and inflammatory pathways. In International Journal of Rheumatic Diseases; Advance online publication, 2025. [Google Scholar] [CrossRef]
- Yki-Järvinen, H. Thiazolidinediones. New England Journal of Medicine 2004, 351(11), 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Konstantopoulos, N.; Lee, J.; Hansen, L.; Li, Z.-W.; Karin, M.; Shoelson, S. E. Reversal of obesity- and diet-induced insulin resistance with salicylates or targeted disruption of IKKβ. Science 2001, 293(5535), 1673–1677. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Buonfiglio, F.; Li, J.; Pfeiffer, N.; Gericke, A. Mechanisms underlying vascular inflammaging: Current insights and potential treatment approaches. Aging and Disease 2024, 16(4), 1889–1917. [Google Scholar] [CrossRef] [PubMed]
- Zhernakova, A.; Yassour, M.; Hall, L. J.; Collado, M. C. Unlocking the power of human milk and infant feeding: Understanding how nutrition and early microbiota interaction shapes health programming. Cell Host & Microbe 2025, 33(6), 820–835. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, G.; Zhang, H.; Karin, M.; Bai, H.; Cai, D. Hypothalamic IKKβ/NF-κB and ER stress link overnutrition to aging. Cell 2021, 135(1), 61–73. [Google Scholar] [CrossRef]


| Mechanism | Molecular Players | Effect on Metabolism | References |
|---|---|---|---|
| Inflammasome Activation | NLRP3, IL-1β, caspase-1 | Insulin resistance; lipid dysregulation | (Stienstra et al., 2011; Youm et al., 2013) |
| TLR Signaling | LPS, TLR4, MyD88 | Chronic inflammation impaired insulin signaling | (Shi et al., 2006; Vijay-Kumar et al., 2010) |
| Metabolic Reprogramming of Immune Cells | mTOR, HIF-1α, AMPK | Controls macrophage polarity, T-cell subsets | (O’Neill & Pearce, 2016; Buck et al., 2015) |
| SCFA-Mediated Signaling | Butyrate, GPR41/43 | Improves glucose tolerance; enhances anti-inflammatory immunity | (Canfora et al., 2015; Koh et al., 2016) |
| Cytokine Regulation of Insulin Pathways | TNF-α, IL-6, IFN-γ | Inhibits IRS-1, decreases GLUT4 translocation | (Hotamisligil, 2006; Wensveen et al., 2019) |
| Mitochondrial ROS Generation | ROS, Unfolded protein respose , ER stress | Drives chronic low-grade inflammation | (Gregor & Hotamisligil, 2011; Ma et al., 2020) |
| Component | Immune Effects | Metabolic Effects | Function | References |
|---|---|---|---|---|
| IgA | Protects mucosal surfaces; prevents pathogen colonization | Indirect metabolic stability via reduced inflammation | Central in passive immunity | Brandtzaeg, 2013; Gopalakrishna & Hand, 2020 |
| Cytokines (e.g., IL-10–inducing factors) | Promotes anti-inflammatory environment | Reduces adipose inflammation, protects liver from fat accumulation | LNFP III promotes IL-10 | Hennet & Borsig, 2016; Donovan & Comstock, 2016 |
| HMOs (e.g., sialylated HMOs, 2′-FL, LNFP III) | Promote Bifidobacterium; enhance gut barrier; influence immune tolerance | Influence weight gain, tissue metabolism, musculoskeletal growth | Levels vary with maternal health | Bode, 2012; Wang et al., 2021 |
| Polyamines | Support intestinal lymphocyte maturation | Essential for healthy gut development | Lower in milk from obese mothers | Canani et al., 2011; Carrasco et al., 2018 |
| Glycoconjugates | Support gut wall maturation; defense | Influence metabolic development | Strong neonatal impact | Newburg & Walker, 2007; Gopal et al., 2008 |
| Microbial modulators | Shape infant microbiota composition | Affect short chain fatty acid production, energy balance | Support early colonization | Arrieta et al., 2014; Bäckhed et al., 2015 |
| Aging Change | Mechanism | Main Effect | Disease Connection | References |
|---|---|---|---|---|
| ↓ Autophagy | ROS ↑ → NLRP3 inflammasome activation | IL-1β / IL-18 ↑ | Metabolic syndrome; inflammaging | (Franceschi et al., 2018; Salminen et al., 2012) |
| Hypothalamic autophagy loss | IKKβ activation → inflammatory signaling | Insulin resistance | Obesity; type 2 diabetes | (Zhang et al., 2021; Meng & Cai, 2011) |
| ↓ Spermidine | ↓ Autophagy → ROS accumulation | Increased cellular stress | Accelerated aging | (Eisenberg et al., 2016; Madeo et al., 2018) |
| Gut dysbiosis | ↓ Faecalibacterium prausnitzii, ↓ Bifidobacterium | Pro-inflammatory cytokines ↑ | Hypertension; chronic inflammation | (O’Toole & Jeffery, 2015; Kim et al., 2018) |
| Leaky gut | LPS translocation into circulation | Systemic inflammation ↑ | Inflammaging; frailty | (Thevaranjan et al., 2017; Man et al., 2015) |
| Tissue | Inflammatory Mechanism | Outcome | References |
|---|---|---|---|
| Adipose | M1 macrophages; TNF-α | ↑ Lipolysis; ↑ Free fatty acids | (Hotamisligil, 2006; Lumeng & Saltiel, 2011) |
| Liver | Kupffer cell activation | NAFLD; ↑ VLDL secretion | (Baffy, 2009; Kazankov et al., 2019) |
| Muscle | NF-κB; JNK signaling | ↓ Glucose uptake | (Hirosumi et al., 2002; Bindra et al., 2013) |
| Pancreas | IL-1β–mediated β-cell dysfunction | ↓ Insulin secretion | (Maedler et al., 2002; Donath, 2014) |
| Strategy | Molecular Target | Mechanistic Action | Clinical Outcome | Limitations | References |
|---|---|---|---|---|---|
| TNF-α Blockade | TNF-α → TNFR1/2 | ↓ NF-κB activation; ↓ macrophage inflammation | Minimal improvement in insulin sensitivity; slight glucose reduction | Weak metabolic effect; heterogeneity in response | Ofei et al. (1996); Ferrannini et al., 2007) |
| IL-1β Antagonism | IL-1β → IL-1R | ↓ β-cell apoptosis; ↑ insulin secretion; ↓ systemic inflammation | Improved glucose homeostasis; moderate HbA1c & fasting glucose reduction | Cost; variable long-term benefits | Larsen et al. 2007; Cavelti-Weder et al. 2012. |
| Salicylates (e.g., high-dose aspirin) | IKKβ/NF-κB pathway | Blocks inflammatory signaling; ↓ hepatic glucose output | HbA1c reduction in T2D; ↓ systemic inflammation | Benefits limited to specific patient subgroups | Yuan et al., 2001; Goldfine et al., 2008) |
| Thiazolidinediones (TZDs) | PPAR-γ (nuclear receptor) | ↑ adiponectin; ↓ macrophage infiltration; ↑ Treg cells; ↑ FGF21 | Markedly improved insulin sensitivity; ↓ adipose inflammation | Weight gain, edema, variable patient response | Yki-Järvinen 2004: Haffner et al. (2002) |
| Omega-3 (EPA/DHA) | GPR120 (anti-inflammatory receptor) | ↓ macrophage chemotaxis; ↓ cytokines; ↑ insulin signaling | Improved insulin sensitivity; better glucose tolerance | Dose-dependent; inconsistent trial outcomes | Oh et al. 2010); Kalupahana et al. (2011) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.



