Submitted:
18 December 2025
Posted:
22 December 2025
You are already at the latest version
Abstract
The X-ray structural analysis of the N-terminal domain cavity from eleven tran-scription regulators of the FNR/CRP family (TFs) has shown several significant trends: (i) The conservancy of effector-binding Phosphate Binding Cassette features in three TFs suggests a closer connection among them than the one proposed based on amino acid sequences. (ii) The allosteric TFs display three clearly different activation mechanisms, which most likely evolved independently. (iii) There are also several non-allosteric TFs which do not bind any ligand; instead, the “allosteric” cavity is partially filled with buried salt bridges, (which is also the case for two allosteric apo TFs). These observations suggest that the non-allosteric TFs evolved from an allosteric counterpart and used salt bridges to fill the formally polar ligand-binding cavity. (iv) A comparison of the allosteric mecha-nisms and the nature of the effectors suggests possible evolutionary relationships among the relevant TFs. (v) O2-sensing TFs share some residues in the relevant N-terminal domain cavity and could have had an already non-allosteric common ancestor. There is a connection between some FNR/CRP TFs and N2 fixation and O2 metabolism; both gases were already metabolized by ancestors of cyanobacteria, over three billion years (3 Ga) ago.
Keywords:
transcription factor
; protein synthesis
; allostery
; fnr
; crp
; camp: x-ray structure
; salt bridge
; phosphate binding cassette
1. Introduction
The regulation of gene transcription is one of the most fundamental biological processes being essential for cell growth, reproduction and adaptation. One essential aspect of early life evolution was the need to preserve the genetic material from destruction by environmental causes. For instance, the enzymatic elimination of the 2’C-OH group in the ribose sugar, a component of RNA, to generate deoxyribose for DNA, removed problems related to the hydrolysis of the former. But it also restricted DNA to be an informational macromolecule.
Although DNA is less susceptible of undergoing chemical attack than RNA, it remains a large macromolecule that needs to be protected. This problem certainly worsened as the complexity of the (proto-)organisms increased. For instance, the DNA of the contemporary unicellular gram-negative prokaryote Escherichia coli (Ec) contains about 5.0x106 nucleotides corresponding to over 4,000 genes [1]. This circular chromosomal DNA not only has to be preserved; it also needs to be compacted over 1000-fold to be fitted inside the E. coli cell [2]. The necessary compaction of DNA, generally called supercoiling, has been proposed to be mediated by small basic proteins called Nucleoid-Associated Proteins (NAPs). Although NAPs are known to bind DNA with low-sequence specificity it has become apparent that they also regulate transcription [3]. This would be a case of exaptation [4,5] where some proteins, which initially bound DNA to supercoil it, later became regulating transcription factors (TFs). In fact, the formal distinction between TF and NAP has become blurred and these two protein classes are thought to form a continuous series going from highly specific ones, acting on few genes, to those that have a pervasive role in transcription [6].
2. Background
One well-studied group of TFs is the Fumarate Nitrate Reductase regulator (FNR)/cyclic AMP Receptor Protein (CRP) family. FNR/CRP TFs are homodimers, which monomers are composed of a versatile N-terminal domain (N-terD) and a C-terminal DNA-binding helix-turn-helix domain (DBD) connected by a long dimerization C-helix. The crystal structure of cAMP-bound EcCRP, one of the most studied TFs of this family, [7] is shown in Figure 1. The two types of domains of this family are found independently associated to proteins displaying a vast array of different functions. For instance, the allosteric phosphorylating protein kinase A undergoes significant conformational changes upon cAMP-binding [8]. The structural comparison of the cyclic nucleotide binding (CNB) domains of this kinase with a cyclic nucleotide-gated channel indicates that they share a C-helical region (Figure 3d in [8]) equivalent to the dimerization C-helix of FNR/CRP proteins (Figure 1).
Interestingly, in tetrameric protein kinases the two homologous CNB domains of each of the two regulatory subunits are linked by a C-helical stretch; however, these subunits do not form a homodimer connected by C-helices, as in FNR/CRP proteins [9]. This observation suggests that the fusion of the allosteric cAMP-binding and DNA-binding domains of FNR/CRP TFs, and their dimerization, took place after the evolution of cAMP-dependent enzymatic activities, such as the regulated phosphorylation by kinases in metabolic central processes.
Signal transduction within the FNR/CRP family is defined as a ‘one-component’ system, a configuration that is abundant in prokaryotic TFs [10]. These systems lack histidine kinase and response regulator domains, which are typical of more complex ‘two-component’ TFs [11]. This observation suggests that phosphorylation was not part of transcription regulation in very early metabolisms. One-component systems are considered to be ancestral to two-component systems, and it is possible that the former were the only class of signal transducers present in the last universal common ancestor (LUCA) [10].
Although DBDs, like the ones found in FNR/CRP proteins, are present in Archaea and Bacteria, their association with a ligand-binding N-terD is only found in the latter [12]. A very recent study by Krishnaswamy et al. has reported sequence and phylogenetic analyses concerning the evolution of the DBDs in the FNR/CRP family [12]. Besides binding to DNA, the DBD domain activates class 1 transcription by binding to the a C-terminal domain of RNA polymerase. The authors proposed that the ancestral DBDs were more similar to those found in a diverse class of “CRP-like” proteins distinct from the well-studied CRP and FNR TFs found in E. coli [12]. These conclusions do not agree with those reported earlier by Matsui et al. who, using a computer-based genomic “spectral clustering” analysis, postulated that an ancestral FNR protein, involved in nitrogen fixation, originated the contemporary FNR/CRP family [13].
The conflicting conclusions of these two studies reflect the inherent complications associated with building reliable prokaryotic phylogenetic trees; the problem is thought to be caused by both extensive horizontal gene transfer (HGT) and inaccuracies due to differences in evolutionary rates between linages [14].
In general, protein foldings are more conserved that amino acid sequences [15], and this is clearly the case for the FNR/CRP family. Consequently, it will make sense to compare the TFs of this family with available 3D X-ray structures to try and shed light on their evolution. Because of the inherently complex evolutionary pressure undergone by these regulatory proteins our analyses will be restricted to the cavity buried in the N-terD -which in EcCRP binds the allosteric cAMP ligand (Figure 1). The cAMP-EcCRP structure will be used as a reference when comparing the corresponding cavities of the other TFs.
3. Structural Similarities and Differences in the Cavity of the N-Terminal Domain
Our working hypothesis is that structural similarities in the N-terD “allosteric” cavity found among FNR/CRP TFs will help establishing their evolutionary relationships from a different perspective than the comparison of overall amino acid sequences.
3.1. The Allosteric Cavity in Members of the FNR/CRP Family
EcCRP. Although cAMP-bound EcCRP is involved in many aspects of carbon metabolism through allosteric transcription regulation it also displays DNA supercoiling activity [16,17]. Because apo-CRP can have a NAP-like global effect on gene expression, it has been suggested that this protein represents an intermediate stage in the evolution of a ligand-dependent allosteric TF from a NAP [17,18]. However, the apo form of EcCRP has its DBDs oriented very differently when compared to the holo form (Figure 2B) [19]. And apo-EcCRP binds to DNA with either low or no specificity.
The cAMP-dependent EcCRP allosteric activation mechanism has been proposed to be based on the H-bonds formed by the N6 group of the bound nucleotide with (cis) Thr128 from one C-helix and (trans) Ser129’ from the other [19]. These H-bonds bring the two C-helices closer together and closer to cAMP. This reorientation allows several hydrophobic residues on both helices to interact, which extends the length of the C-helices by 1.5 turns. These changes also modify the helical content and orientation of the DBDs placing them in the right configuration for cognate DNA binding (Figure 2B) [19]. The most conserved feature of the cAMP-binding cavity has been called the ‘phosphate binding cassette’ (PBC) [8] which, as we will see below, shares some structural elements with other TFs of the family. A second cAMP site is present in one of the monomers of the holo EcCRP (Figure 1 and Figure 2B) but it does not seem to play an in vivo allosteric role.
NtcA. Another example of allosteric regulation by a TF from the FNR/CRP family is the cyanobacterial nitrogen regulator NtcA [20]. The crystal structures of apo NtcA and in complex with its effector 2-oxoglutarate (2-OG) have been reported [21,22]. 2-OG is a metabolite of the Krebs cycle, a hub of cell metabolism of generally accepted great antiquity.
When nitrogen becomes scarce the reductive amination of 2-OG to generate glutamate is impaired and 2-OG accumulates; under these conditions NtcA binds this metabolite and positively regulates nitrogen assimilation [20]. The conformational transition between the apo and complexed states is much simpler in NtcA than in EcCRP (Figure 2); it mostly involves a relatively minor change in the orientation of the dimerization C-helices. This change is mainly caused by 2-OG binding to (cis) Arg129 from one C-helix and (trans) Glu134’ from the other.
Although NtcA is the only known member of the FNR/CRP family to be co-activated by another protein, its complex with 2-OG is similar to the cAMP-EcCRP complex ([22] and Figure 3A and 3B). Thus, one of the O atoms of the 5-COO- group of 2-OG and one of the O atoms of the phosphate group of cAMP show equivalent interactions with Arg88, and Arg83, which are H-bonded to the respective mainchain O atoms of Gly 76 and Gly 72. Interestingly, and as already noted by Zhao et al. [22], the helix-stabilizing EcCRP (trans) Ser129’-cAMP bond is stereochemically equivalent to the NtcA (trans) Glu134’-2-OG bond (Figure 3A and 3B). Thus, similar intermonomer interactions are found in the two TFs, although in NtcA the longer side chain of a Glu residue is required to reach the smaller 2-OG.
In EcCRP these motifs have been defined as part of the conserved PBC mentioned above [8]. Arg129 of NtcA, which forms a bidentate salt bridge with 2-OG, has a corresponding Arg124 in EcCRP; however, this residue makes a salt bridge interaction with Glu73 in a different orientation due to the presence of the cAMP adenine base (Figure 3A). Interestingly for the discussion below, in apo EcCRP this salt bridge is conserved but its orientation is very different (Figure 3C). Also, when holo and apo NtcA are compared, a salt bridge between R129 and (trans) Glu134’ is observed in the latter (Figure 3D).
Intriguingly, and based on amino acid sequence analyses, CRP and NtcA have been assigned to clearly different branches of the FNR/CRP superfamily [23]. However, unless one invokes a strong convergent evolutionary process, the observed similarities in their cavities should represent conserved features found in a common allosteric ancestor. The next point to address is the possible connection between that ancestor and the two contemporary effectors. Based on metabolic criteria cAMP seems to be a more versatile effector being involved in many types of stress responses [24]. Conversely, 2-OG, as a member of the Krebs cycle, is a precursor of the essential proteogenic Glu and Gln amino acids and it accumulates when the levels of biologically available nitrogen are low [20].
Another point to consider here is the allosteric mechanism elicited by each of these ligands. As shown in Figure 2B the activation of EcCRP involves major conformational changes. As mentioned above, this is not the case for NtcA where 2-OG fixation causes relatively minor readjustments that activate the TF towards DNA binding [22]. From a parsimonious viewpoint, it might then be concluded that NtcA should be closer to the common ancestor than EcCRP. What seems clear is that the evolutionary paths of the two proteins are directly connected.
CprK. Several bacterial species are able to conserve energy through halorespiration [25]. In the strictly anaerobic, gram-positive Desulfitobacterium spp. [26] the expression of halorespiratory genes is controlled by the allosteric CprK TF, which binds the o-chlorophenolacetic acid (OCPA) effector [27]. Several crystal structures of CprK have been reported with and without its ligand and hypotheses about the stereochemistry of the allosteric process have been proposed [28,29,30]. The halogenation of o-phenolacetate to yield OCPA is essential for its recognition by this TF and its activation [27]. The general conclusion is that halogenation favors the deprotonation of the o-phenolacetate -OH group, which is needed for the initially weak OCPA binding to one of the CprK monomers.
The crystal structure of the CprK-OCPA complex shows that the deprotonated phenoxide ion forms a salt bridge with the -NH3+ group of Lys133 from the cis C-helix (Figure 4). During that process, the ligand’s -Cl- ion docks to a pocket site formed by the two dimerization C-helices. This binding introduces several major conformational changes that greatly increase the affinity of the other CprK monomer for OCPA [28,30]. After the binding of the second OCPA, the DBDs of the holo CprK dimer are well oriented to bind its cognate DNA regions. This allosteric process has been defined as a case of “extreme positive cooperativity” [28]. Although OCPA binds to a site in CprK topologically equivalent to the sites of EcCRP for cAMP and NtcA for 2-OG, the residues involved in its binding are very different (Figure 4). It should also be mentioned that when CprK is in vitro exposed to O2 it forms an interdomain Cys11-Cys200’ disulfide bridge. The oxidized TF can still bind OCPA but is unable to recognize its cognate DNA. However, results obtained using the corresponding Cys → Ser mutants did not support a physiological role for the disulfide bridge in CprK expressed in aerobically-grown E. coli cells [31].
In summary, CprK may be a distinct, more recent addition to the FNR/CRP family. In fact, the comprehensive amino acid sequence analysis mentioned above has placed CprKs as a separate branch of this family [23]. In addition, several other observations agree with this notion. Firstly, phylogenetic and structural analyses suggest that there might be evolutionary and functional relationships between the six known families of halogenating enzymes and α/β hydrolases, acid phosphatases, peroxidases, chemotaxis phosphatases, oxidoreductases, and the SAM hydroxide adenosyltransferases, respectively [32]. This, in turn, suggests that each of the halogenating proteins has evolved from a pre-existing enzyme with similar catalytic properties. Secondly, halogen-containing compounds, such as OCPA, are mostly the polluting result of past and present industrial and agricultural human activities [28]. For this reason, it is not possible to rule out the relatively recent evolution of some of the halogenating enzymes in halogen-metabolizing bacteria to conserve energy in a respiration-type metabolism. Thirdly, the extreme cooperativity of the CprK allosteric mechanism is not found in other FNR/CRP family members [28].
3.2. Negative Allostery in the FNR/CRP Family
Clp. As mentioned above [12], and further discussed below, many of the functional and structural conclusions based on EcCRP, and by extension on NtcA, do not apply to all the members of the FNR/CRP family. Thus, effector binding can also have a negative effect on transcription regulation. The Clp TF from the plant pathogen Xanthomonas campestris (Xc) binds to cognate DNA without any ligand [33]. However, the binding of the bacterial allosteric inhibitor bis-(3’-5’)-cyclic di-guanosine monophosphate (c-d-GMP, Figure 5A) to XcClp causes its dissociation from DNA. The crystal structure of the apo form of this TF as determined by Chin et al. shows it adopts an intrinsically active conformation for DNA binding [33]. These authors have also explored possible c-d-GMP binding sites in XcClp by molecular docking using a home-made program. Their preferred model has c-d-GMP bound between the TF N-terD and DBD domains, close to the C-helix [33].
The negative allosteric mechanism displayed by XcClp [33,34] is the most unusual one reported so far for a member of the FNR/CRP family. Examination of the N-terD cavity shows that the orientation of the PBC Arg83-Gly72 and the Arg88-Gly76 motifs, involved in the respective binding of the acidic functions of cAMP and 2-OG (Figure 3), is also found in XcClp (Arg96-Gly87 motif, Figure 5B). In addition, the XcClp salt bridge between Glu88 and Arg144 is similar to the one found in EcCRP between Glu73 and Arg124 (Figure 5B). Furthermore, cAMP competed with c-d-GMP for binding to the related Clp from X. anoxopodis when added at mM concentrations [34,35], even though cAMP is not present in Xanthomonas [35]. This observation raises the possibility that the c-d-GMP effector, which has two phosphate groups chemically equivalent to the one found in cAMP (see Figure 2A and 5A), could bind close to the Arg96-Gly87 motif.
Because of its bulk, c-d-GMP binding would result in major conformational changes in Clp, plausibly causing its dissociation from DNA. Such changes would also complicate an in silico predicting c-d-GMP docking process. If this notion is correct the allosteric process of Clp would be almost the exact reverse of the one found in EcCRP.
3.3. Different Allostery in Other Members of the FNR/CRP Family
PrfA. In the gram-positive intracellular human pathogen Listeria monocytogenes the expression of the most virulent genes is regulated by PrfA [36]. This TF is capable of binding to a consensus DNA sequence with low affinity in the absence of an effector [37]. However, its activity increases in an intracellular environment and the crystal structure of the constitutively active G145S-PrfA variant showed the structuring of the helix-turn-helix motif of the DBDs by forming an H-bond between the Oγ of Ser145 and the O of Gly14 [37]. These observations strongly suggested that PrfA was an allosteric TF that would bind a specific effector. This hypothesis was confirmed when a glutathione synthase was shown to be essential for PrfA activity and reduced glutathione (GSH) increased it [38]. As reviewed in [39], glutathione is also known to play a central role in many other processes: redox buffering, response to oxidative stress, biosynthesis of iron–sulfur proteins, detoxification of metals and xenobiotics, redox signaling, apoptosis and sulfur storage and transport. Its role in the remodeling of the transcriptional program of the pathogen involves a redox 2 GSH ⇔ GSSG + 2 H+ equilibrium. When L. monocytogenes is phagocytized by a macrophage it is initially captured in a vacuole. There, both PrfA and GSH get oxidized (PrfA has four redox-active cysteine residues) and the oxidized TF can neither bind DNA nor be activated by GSSG. Next, the bacterium escapes from the oxidizing vacuole and both PrfA and bacterial GSSG get reduced. In addition, L. monocytogenes can also get reduced GSH from the macrophage cytoplasm; and under these conditions it colonizes the host cell [38].
The crystal structure of the protein clearly shows that GSH binds to a region in the N-terD different from the one described for EcCRP, NtcA and CprK (Figure 6). A structural comparison reveals that the region of PrfA corresponding to the cAMP-binding allosteric cavity of EcCRP is partially occupied by Gln61, Tyr63, Ile71 and Gln123 (the two Gln residues are H-bonded). Thus, although the cavity is topologically conserved [40], it is not fit to bind GSH. A superposition of cAMP-bound CRP and GSH-bound PrfA shows that the closest atoms of the two ligands are separated by about 4 Å. The conclusion is that allostery has evolved more than once in the FNR/CRP family [41].
Besides the structure of the G145S-PrfA variant [37] several other crystal structures of PrfA are available, including complexes with DNA and GSH [40,41]. GSH binds to a tunnel site between the N-terD and DBD domains of the monomer (Figure 6) and its thiol group is buried in a hydrophobic pocket (Figure 1 in ref. [41]). The effector binding causes conformational changes in the DBDs that are very similar to those found in the G145S-PrfA variant with or without bound GSH.
CooA. In bacteria such as Rhodospirillium rubrum (Rr), CO metabolism is controlled by the CooA TF that binds this gas to its heme b cofactor [42]. The first crystallographic report of CooA showed that the Fe2+ ion bound to heme b was axially coordinated by His77 and, rather surprisingly, by the terminal amine nitrogen of Pro2 from the other monomer [43]. A later report of the related Carboxydothermus hydrogenoformans (Ch) CooA structure with heme-bound imidazole (Im, Figure 7) indicated that, in vivo, CO should displace the trans Pro2 N-terminal ligand [44]. The functional properties of RrCooA and ChCooA, including different redox responses of their heme cofactors, such as the swap of an axial ligand, have been recently reviewed [45].
As expected for a member of the FNR/CRP family, CooA coordinates heme b in its N-terD cavity (Figure 7). However, the cofactor is not located at the place of either cAMP in EcCRP or GSH in PrfA. In fact, heme b binding requires the absence of a region corresponding to residues 72-82 in EcCRP; furthermore, a relative insertion at the N-terminal segment was needed for Pro2 to reach the heme of the other monomer [43].
Although trans monomer ligand binding effects are also found in EcCRP and NtcA (see above), they clearly differ from trans Pro2 binding to heme b in CooA. In the Im-bound ChCooA crystal structure (Figure 7) the -NH2 group of this residue is located 16 Å from the heme, which plane rotates 30° about the normal to the porphyrin ring relative to the inactive RrCooA structure [42]. In a new variant ChCooA structure, which has one monomer in the active state, it was found that the N-terminal segment was located between the heme-binding and DB domains [46]. In this bridging position, the N-terminal segment helps orienting the DBDs so that they adopt the DNA-binding configuration. This configuration has been defined as the ‘N-terminal Velcro model’ [47]. In many respects this allosteric member of the FNR/CRP family is the one that displays the most elaborate conformational changes upon CO binding.
3.4. Non-Allosteric Members of the FNR/CRP Family
SdrP/DdrI. Many TFs bypass effector binding completely. That is the case of SdrP from Thermus thermophilus [48] and DdrI from Deinococcus radiodurans [49,50], which are transcriptionally active without a ligand.
Although, based on the sequences of their ribosomal RNAs and proteins, these two extremophile organisms have evolved from a common ancestor, extensive specific gene loss and acquisition have rendered them clearly different [14,51]. T. thermophilus is a gram-negative thermophile that displays low resistance to ionizing radiation and desiccation whereas D. radiodurans is a gram-positive mesophile highly resistance to those two drastic environmental conditions.
In spite of the differences between these two extremophiles, the SdrP and DdrI crystal structures show that the site corresponding to where the cAMP adenine base binds in EcCRP is occupied by an Arg side chain involved in a salt bridge in both proteins (Figure 8A).
The (Tt) Arg43-Glu53 and (Dr) Arg55-Glu65 salt bridges help conferring a ‘cAMP-bound-like’ conformation to these CRPs. Notably, the residue equivalent to Arg43 and Arg55 in E. coli is Ser53 (Figure 8B).
Although it cannot be unambiguously determined, it seems structurally more parsimonious to conclude that cAMP-binding CRP predated the extremophile active apo-CRP versions because of its ubiquity [8]. After all, disabling the cAMP binding site would have required establishing the observed salt bridges at the N-terD cavities whereas optimizing cAMP binding to that site was probably a gradual, slow evolutionary process.
PgCRP. Another effector-independent TF has been described in the human oral pathogen Porphyromonas gingivalis (Pg) [52]. PgCRP is involved in biofilm formation and adhesion to, and invasion of, oral keratinocytes; its activity is thought to be regulated by its concentration in the bacterial cell. Like TtSdrP and DrDdrI, the PgCRP crystal structure is similar to the cAMP-EcCRP complex structure [53]. A superposition of these two TFs shows that the side chains of Arg77 and Arg88 of PgCRP occupy the flanks of the N-terD cavity (Figure 9A). They establish long-range interactions with main-chain carbonyl oxygens. Interestingly, the H-bond of Arg77 with the >C=O from Phe84 is structurally close to the one between Arg55 and the >C=O from Tyr62 in the D. radiodurans DdrI (Figure 9B); and to a lesser extent, between Arg43 and the >C=O from Tyr50 in T. thermophilus SdrP (not shown). These similarities are evolutionary intriguing.
As mentioned above, D. radiodurans is gram positive and strictly aerobic, T. thermophilus is gram negative and although aerobic, it can also function anaerobically, but P. gingivalis is gram negative and strictly anaerobic [54].
3.5. Non-Allosteric O2-Sensing Members of the FNR/CRP Family
FNRs/FixK2. Three other members of the family, which do not bind a ligand, are the O2-sensing [4Fe-4S]-FNRs from gram-negative [55] and gram-positive bacteria [56], and FixK2 from Bradyrhizobium japonicum [57].
The two types of iron-sulfur cluster-binding FNRs have very different sensing mechanisms. In FNRs from gram-negative bacteria a dimer → monomer transition, which results in loss of DNA binding, is caused by the disassembly upon O2 exposure of an N-terminally located [4Fe-4S] cluster (Figure 10B) [55]. Conversely, in the FNR from gram-positive bacteria, the [4Fe-4S] cluster is located in the C-terminal DBD; this region is expected to be altered upon O2 exposure which results in cluster decomposition, without causing a dimer-monomer transition [56].
FixK2 activates genes involved in nitrogen-fixation and in anoxic, microoxic and endosymbiotic processes through an O2-sensing mechanism, which does not depend on an iron-sulfur cluster. FixK2 has a very redox-sensitive Cys-SH group (Cys183), which gets oxidized by O2. Consequently, and in order to facilitate the manipulations, the crystallized species was the C183S variant [57].
DNA-bound dimeric FixK2 adopts a configuration that is very similar to the one displayed by the [4Fe-4S]-FNR from gram-negative Aliivibrio fischeri (Af) [58] (Figure 10). Because the crystal structure of uncomplexed FixK2 is not available [57] it is not known whether it displays a similar dimeric conformation. What seems clear is that holo AfFNR should have a DNA-binding dimeric conformation in solution as shown by the orientation of its DBDs in our crystal structure [58] (Figure 10B).
The AfFNR Cys-rich cluster-binding region has no visible counterpart in FixK2 (Figure 10B). In fact, both proteins display N-terminal disordered regions in their crystals (1-16 in AfFNR and 1-37 in FixK2) [57,58]. As expected, FixK2 lacks key residues that in FNR condition the dimer-monomer transition and loss of DNA binding that results from O2 exposure and [4Fe-4S] cluster disassembly [58].
Although the overall amino acid identity between AfFNR and FixK2 is about 25% three regions show significantly higher identities. As expected from their similar DNA binding sites, the respective helix-turn-helix DNA-binding sequences have 74% identity. Another region with a high identity score (62.5%) comprises the end of the C α-helix and half of the D α-helix. The third stretch with high identity (69.2%) lines the cavity structurally equivalent to the allosteric one where cAMP binds in EcCRP:

AfFNR and FixK2 have three homologous polar residues (FNR His93, Asp97 and Glu150 and FixK2 His86, Asp90 and Glu141) (Figure 10A) and one conservative change (FNR Gln155 and FixK2 Glu146) in that cavity. This degree of conservancy of internal charged and polar residues is considerable; it strongly suggests that the common ancestor of FNR and FixK2 was already not allosteric.
These two TFs have very different O2 sensing mechanisms. The FixK2 redox-active Cys183 thiolate mentioned above is located at the DBD. There, it reacts with radical oxygen species (ROS) to form either sulfenic, sulfinic, or sulfonic acid derivatives which hinder DNA binding [59]. This mechanism appears to be simpler than the dimer-monomer transition of gram-negative FNR suggesting that it may be older. AfFNR has two major insertions relative to FixK2: the three Cys [4Fe-4S] cluster binding loop (FNR res. 17-29) and the 180-190 loop. These two regions interact through van der Waals contacts and, upon cluster disassembly, the conformational change of the former loop would be transmitted to the DBD by the latter loop.
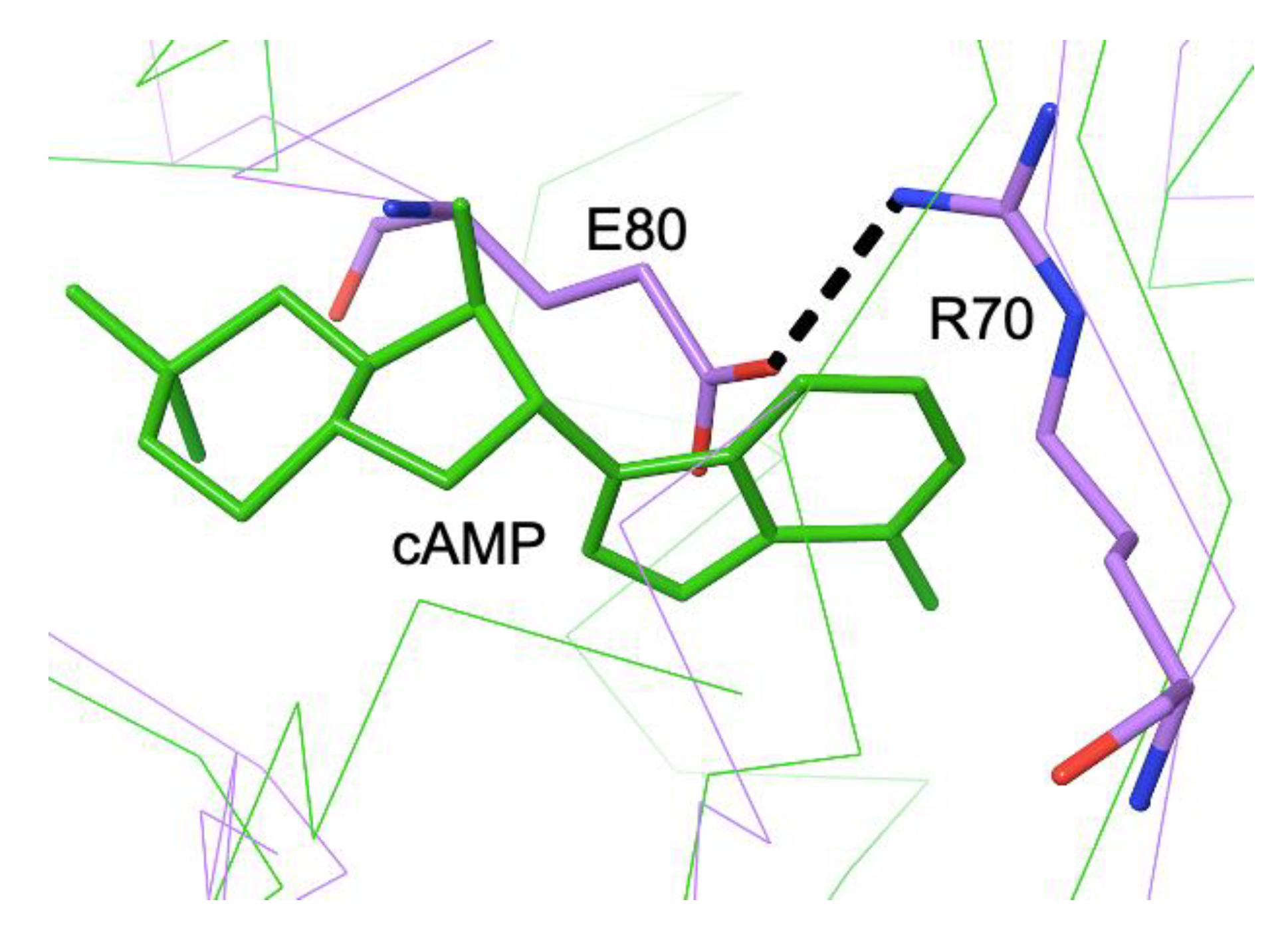
Although there is no X-ray structure available for an FNR from a gram-positive bacterium, an Alphafold model of the FNR from Bacillus subtilis (Bs) -downloaded from the Uniprot web site (AF052953.pdb) [60]- confirms the C-terminal position of the [4Fe-4S] cluster (not shown). Interestingly, the model also predicts the formation of a salt bridge between Arg70 and Glu80 in the N-terD cavity (Figure 11).
We have seen that FNR/CRP TFs have been subjected to intense evolutionary pressure to generate a large array of activities and mechanisms starting from a common rather simple two-domain protein. The structures reviewed above show that when a key metabolic function (such as the regulation of gene expression) needs to be adapted to changing conditions Nature can display a vast array of solutions
4. Conclusions
A few major points deserve to be noted: 1. Conservancy of effector-binding PBC features of EcCRP in NtcA and the PBC-like Arg-Gly motif in Clp, a case of negative allostery, suggests a closer connection among these TFs than the ones proposed based on overall amino acid sequences. 2. Although PrfA and CooA also use their N-terD for effector binding, their allosteric activation mechanisms are different from each other, and from the one mediated by cAMP in EcCRP; it may be then concluded that in the FNR/CRP family allostery independently evolved at least three times; 3. There are several non-allosteric TFs which do not bind any ligands; in SdrP, DdrI and the modelled BsFNR, the “allosteric” cavity has been partially filled with buried salt bridges, (also the case in apo EcCRP and apo NtcA). 4. These observations suggest that the non-allosteric TFs evolved from ligand-binding counterparts; not only allostery would have make more sense in metabolic regulation, also filling in a pre-existing ligand-binding polar cavity with salt bridge-forming residues seems simpler than the opposite. 5. The less involved allosteric mechanism of NtcA relative to EcCRP, and the metabolic role of 2-OG when compared to cAMP, suggest that 2-OG could have predated cAMP as an effector. 6. AfFNR and FixK2 have similar residues in the relevant N-terD cavity and could have had a common -already non-allosteric- ancestor. 7. A last point worth mentioning if the connection between some FNR/CRP TFs and N2 fixation (NtcA, FixK2) and O2 metabolism (FNRs, FixK2), which origins can be traced to 3.77 Ga [61] and 2.7-3.2 Ga [62], respectively. Both gases were already metabolized by ancestors of cyanobacteria.
Funding
This research received no external funding.
Acknowledgments
The author thanks Dr. Patricia Amara for critical reading of the manuscript and for preparing the figures with Maestro [63], and the Université Grenoble Alpes for administrative support.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
c-d-GMP: bis-(3’-5’)-cyclic di-guanosine monophosphate; CNB: cyclic nucleotide domain; DBD: DNA-binding domain, GSH: glutathione; N-terD: N-terminal domain; OCPA: o-chlorophenolacetic acid. TF: transcription factor; 2-OG: 2-oxoglutarate; PBC: phosphate binding cassette.
References
- Blattner, F.R.; Plunkett, G.; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The Complete Genome Sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Murphy, L.D.; Zimmerman, S.B. Condensation and Cohesion of λ DNA in Cell Extracts and Other Media: Implications for the Structure and Function of DNA in Prokaryotes. Biophys Chem 1995, 57, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Visweswariah, S.S.; Busby, S.J.W. Evolution of Bacterial Transcription Factors: How Proteins Take on New Tasks, but Do Not Always Stop Doing the Old Ones. Trends Microbiol. 2015, 23, 463–467. [Google Scholar] [CrossRef]
- Brosius, J. Exaptation at the Molecular Genetic Level. Sci China Life Sci 2019, 62, 437–452. [Google Scholar] [CrossRef]
- Gould, S.J. Exaptation: A Crucial Tool for an Evolutionary Psychology. J. Soc. Issues 1991, 47, 43–65. [Google Scholar] [CrossRef]
- Dorman, C.J.; Schumacher, M.A.; Bush, M.J.; Brennan, R.G.; Buttner, M.J. When Is a Transcription Factor a NAP? Curr. Opin. Microbiol. 2020, 55, 26–33. [Google Scholar] [CrossRef]
- McKay, D.B.; Steitz, T.A. Structure of Catabolite Gene Activator Protein at 2.9 A Resolution Suggests Binding to Left-Handed B-DNA. Nature 1981, 290, 744–749. [Google Scholar] [CrossRef]
- Berman, H.M.; Ten Eyck, L.F.; Goodsell, D.S.; Haste, N.M.; Kornev, A.; Taylor, S.S. The cAMP Binding Domain: An Ancient Signaling Module. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 45–50. [Google Scholar] [CrossRef]
- Weber, I.T.; Takio, K.; Titani, K.; Steitz, T.A. The cAMP-Binding Domains of the Regulatory Subunit of cAMP-Dependent Protein Kinase and the Catabolite Gene Activator Protein Are Homologous. Proc. Natl. Acad. Sci. U.S.A 1982, 79, 7679–7683. [Google Scholar] [CrossRef]
- Ulrich, L.E.; Koonin, E.V.; Zhulin, I.B. One-Component Systems Dominate Signal Transduction in Prokaryotes. Trends Microbiol. 2005, 13, 52–56. [Google Scholar] [CrossRef]
- Zschiedrich, C.P.; Keidel, V.; Szurmant, H. Molecular Mechanisms of Two-Component Signal Transduction. J. Mol. Biol. 2016, 428, 3752–3775. [Google Scholar] [CrossRef]
- Krishnaswamy, M.; Seshasayee, A.S.N. The Evolution of Function in the DNA Binding Domain of the CRP/FNR Family. Genome Biol Evol. 2025, 17(10), evaf182. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Tomita, M.; Kanai, A. Comprehensive Computational Analysis of Bacterial CRP/FNR Superfamily and Its Target Motifs Reveals Stepwise Evolution of Transcriptional Networks. Genome Biol Evol. 2013, 5, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, M.V.; Wolf, Y.I.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Daly, M.J.; Koonin, E.V.; Makarova, K.S. Comparative Genomics of Thermus Thermophilus and Deinococcus Radiodurans: Divergent Routes of Adaptation to Thermophily and Radiation Resistance. BMC Evol Biol 2005, 5, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-G.; Kim, S.-H. Evolution of Protein Structural Classes and Protein Sequence Families. Proc. Natl. Acad. Sci. U.S.A 2006, 103, 14056–14061. [Google Scholar] [CrossRef]
- Dinardo, S.; Voelkel, K.A.; Sternglanz, R.; Reynolds, A.E.; Wright, A. Escherichia coli DNA Topoisomerase I Mutants Have Compensatory Mutations in DNA Gyrase Genes. Cell 1982, 31, 43–51. [Google Scholar] [CrossRef]
- Heyde, S.A.H.; Frendorf, P.O.; Lauritsen, I.; Nørholm, M.H.H. Restoring Global Gene Regulation through Experimental Evolution Uncovers a NAP (Nucleoid-Associated Protein)-Like Behavior of Crp/Cap. mBio 2021, 12. [Google Scholar] [CrossRef]
- Hołówka, J.; Zakrzewska-Czerwińska, J. Nucleoid Associated Proteins: The Small Organizers That Help to Cope With Stress. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Sharma, H.; Yu, S.; Kong, J.; Wang, J.; Steitz, T.A. Structure of Apo-CAP Reveals That Large Conformational Changes Are Necessary for DNA Binding. Proc. Natl Acad Sci U S A 2009, 106, 16604–16609. [Google Scholar] [CrossRef]
- Vega-Palas, M.A.; Madueño, F.; Herrero, A.; Flores, E. Identification and Cloning of a Regulatory Gene for Nitrogen Assimilation in the Cyanobacterium Synechococcus sp. Strain PCC 7942. J Bacteriol 1990, 172, 643–647. [Google Scholar] [CrossRef]
- Forcada-Nadal, A.; Bibak, S.; Salinas, P.; Contreras, A.; Rubio, V.; Llácer, J.L. Structures of the Cyanobacterial Nitrogen Regulators NtcA and PipX Complexed to DNA Shed Light on DNA Binding by NtcA and Implicate PipX in the Recruitment of RNA Polymerase. Nucleic Acids Res. 2025, 53, gkaf096. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-X.; Jiang, Y.-L.; He, Y.-X.; Chen, Y.-F.; Teng, Y.-B.; Chen, Y.; Zhang, C.-C.; Zhou, C.-Z. Structural Basis for the Allosteric Control of the Global Transcription Factor NtcA by the Nitrogen Starvation Signal 2-Oxoglutarate. Proc Natl Acad Sci U S A 2010, 107, 12487–12492. [Google Scholar] [CrossRef] [PubMed]
- Körner, H.; Sofia, H.J.; Zumft, W.G. Phylogeny of the Bacterial Superfamily of Crp-Fnr Transcription Regulators: Exploiting the Metabolic Spectrum by Controlling Alternative Gene Programs. FEMS Microbiol. Rev. 2003, 27, 559–592. [Google Scholar] [CrossRef] [PubMed]
- Canaves, J.M.; Taylor, S.S. Classification and Phylogenetic Analysis of the cAMP-Dependent Protein Kinase Regulatory Subunit Family. J Mol Evol 2002, 54, 17–29. [Google Scholar] [CrossRef]
- Smidt, H.; Vos, W.M. de Anaerobic Microbial Dehalogenation. Annu. Rev. Microbiol. 2004, 58, 43–73. [Google Scholar] [CrossRef]
- Villemur, R.; Lanthier, M.; Beaudet, R.; Lépine, F. The Desulfitobacterium genus. FEMS Microbiol. Rev. 2006, 30, 706–733. [Google Scholar] [CrossRef]
- Smidt, H.; van Leest, M.; van der Oost, J.; de Vos, W.M. Transcriptional Regulation of the Cpr Gene Cluster in ortho-Chlorophenol-Respiring Desulfitobacterium dehalogenans. J. Bacteriol. 2000, 182, 5683–5691. [Google Scholar] [CrossRef]
- Levy, C.; Pike, K.; Heyes, D.J.; Joyce, M.G.; Gabor, K.; Smidt, H.; van der Oost, J.; Leys, D. Molecular Basis of Halorespiration Control by CprK, a CRP-FNR Type Transcriptional Regulator. Mol. Microbiol. 2008, 70, 151–167. [Google Scholar] [CrossRef]
- Kemp, L.R.; Dunstan, M.S.; Fisher, K.; Warwicker, J.; Leys, D. The Transcriptional Regulator CprK Detects Chlorination by Combining Direct and Indirect Readout Mechanisms. In Philos. Trans. R. Soc. London, Ser B: Biol. Sci.; 2013. [Google Scholar] [CrossRef]
- Joyce, M.G.; Levy, C.; Gábor, K.; Pop, S.M.; Biehl, B.D.; Doukov, T.I.; Ryter, J.M.; Mazon, H.; Smidt, H.; van den Heuvel, R.H.H.; et al. CprK Crystal Structures Reveal Mechanism for Transcriptional Control of Halorespiration*. J. Biol. Chem. 2006, 281, 28318–28325. [Google Scholar] [CrossRef]
- Gábor, K.; Veríssimo, C.S.; Cyran, B.C.; Ter Horst, P.; Meijer, N.P.; Smidt, H.; de Vos, W.M.; van der Oost, J. Characterization of CprK1, a CRP/FNR-Type Transcriptional Regulator of Halorespiration from Desulfitobacterium hafniense. J Bacteriol 2006, 188, 2604–2613. [Google Scholar] [CrossRef]
- Xu, G.; Wang, B.-G. Independent Evolution of Six Families of Halogenating Enzymes. PLoS One 2016, 11, e0154619. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.-H.; Lee, Y.-C.; Tu, Z.-L.; Chen, C.-H.; Tseng, Y.-H.; Yang, J.-M.; Ryan, R.P.; McCarthy, Y.; Dow, J.M.; Wang, A.H.-J.; et al. The cAMP Receptor-Like Protein CLP Is a Novel c-di-GMP Receptor Linking Cell–Cell Signaling to Virulence Gene Expression in Xanthomonas campestris. J. Mol. Biol. 2010, 396, 646–662. [Google Scholar] [CrossRef]
- Leduc, J.L.; Roberts, G.P. Cyclic di-GMP Allosterically Inhibits the CRP-like Protein (Clp) of Xanthomonas Axonopodis pv. citri. J Bacteriol 2009, 191, 7121–7122. [Google Scholar] [CrossRef] [PubMed]
- Gomelsky, M. Cyclic-Di-GMP-Binding CRP-Like Protein: A Spectacular New Role for a Veteran Signal Transduction Actor. J. Bact. 2009, 191, 6785–6787. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, T.; Leimeister-Wächter, M.; Domann, E.; Hartl, M.; Goebel, W.; Nichterlein, T.; Notermans, S. Coordinate Regulation of Virulence Genes in Listeria Monocytogenes Requires the Product of the prfA Gene. J Bacteriol 1992, 174, 568–574. [Google Scholar] [CrossRef]
- Eiting, M.; Hagelüken, G.; Schubert, W.-D.; Heinz, D.W. The Mutation G145S in PrfA, a Key Virulence Regulator of Listeria monocytogenes, Increases DNA-Binding Affinity by Stabilizing the HTH Motif. Mol. Microbiol. 2005, 56, 433–446. [Google Scholar] [CrossRef]
- Reniere, M.L.; Whiteley, A.T.; Hamilton, K.L.; John, S.M.; Lauer, P.; Brennan, R.G.; Portnoy, D.A. Glutathione Activates Virulence Gene Expression of an Intracellular Pathogen. Nature 2015, 517, 170–173. [Google Scholar] [CrossRef]
- Deponte, M. Glutathione Catalysis and the Reaction Mechanisms of Glutathione-Dependent Enzymes. Biochim. Biophys. Acta 2013, 1830, 3217–3266. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, H.; Zhu, Y.; Gao, P. Structural Insights into Glutathione-Mediated Activation of the Master Regulator PrfA in Listeria monocytogenes. Protein Cell 2017, 8, 308–312. [Google Scholar] [CrossRef]
- Hall, M.; Grundström, C.; Begum, A.; Lindberg, M.J.; Sauer, U.H.; Almqvist, F.; Johansson, J.; Sauer-Eriksson, A.E. Structural Basis for Glutathione-Mediated Activation of the Virulence Regulatory Protein PrfA in Listeria. Proc Natl Acad Sci U S A 2016, 113, 14733–14738. [Google Scholar] [CrossRef]
- Aono, S.; Nakajima, H.; Saito, K.; Okada, M. A Novel Heme Protein That Acts as a Carbon Monoxide-Dependent Transcriptional Activator in Rhodospirillum rubrum. Biochem. Biophys. Res. Commun. 1996, 228, 752–756. [Google Scholar] [CrossRef]
- Lanzilotta, W.N.; Schuller, D.J.; Thorsteinsson, M.V.; Kerby, R.L.; Roberts, G.P.; Poulos, T.L. Structure of the CO Sensing Transcription Activator CooA. Nat Struct Mol Biol 2000, 7, 876–880. [Google Scholar] [CrossRef]
- Komori, H.; Inagaki, S.; Yoshioka, S.; Aono, S.; Higuchi, Y. Crystal Structure of CO-Sensing Transcription Activator CooA Bound to Exogenous Ligand Imidazole. J. Mol. Biol. 2007, 367, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Dent, M.R.; Weaver, B.R.; Roberts, M.G.; Burstyn, J.N. Carbon Monoxide-Sensing Transcription Factors: Regulators of Microbial Carbon Monoxide Oxidation Pathway Gene Expression. J Bacteriol 205 e00332-22. [CrossRef] [PubMed]
- Borjigin, M.; Li, H.; Lanz, N.D.; Kerby, R.L.; Roberts, G.P.; Poulos, T.L. Structure-Based Hypothesis on the Activation of the CO-Sensing Transcription Factor CooA. Acta Crystallogr D Biol Crystallogr 2007, 63, 282–287. [Google Scholar] [CrossRef]
- Tripathi, S.; Poulos, T.L. Testing the N-Terminal Velcro Model of CooA Carbon Monoxide Activation. Biochemistry 2018, 57, 3059–3064. [Google Scholar] [CrossRef]
- Agari, Y.; Kashihara, A.; Yokoyama, S.; Kuramitsu, S.; Shinkai, A. Global Gene Expression Mediated by Thermus thermophilus SdrP, a CRP/FNR Family Transcriptional Regulator. Mol. Microbiol. 2008, 70, 60–75. [Google Scholar] [CrossRef]
- Agari, Y.; Kuramitsu, S.; Shinkai, A. X-Ray Crystal Structure of TTHB099, a CRP/FNR Superfamily Transcriptional Regulator from Thermus thermophilus HB8, Reveals a DNA-Binding Protein with No Required Allosteric Effector Molecule. Proteins 2012, 80, 1490–1494. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, J.; Gao, X.; Cao, Y.; Ye, S.; Chen, C.; Wang, L.; Xu, H.; Guo, M.; Zhang, D.; et al. cAMP-Independent DNA Binding of the CRP Family Protein DdrI from Deinococcus radiodurans. mBio 2024, 15, e01144-24. [Google Scholar] [CrossRef]
- Nishida, H.; Abe, R.; Nagayama, T.; Yano, K. Genome Signature Difference between Deinococcus radiodurans and Thermus thermophilus. Int. J. Evol. Biol 2012, 2012, 205274. [Google Scholar] [CrossRef]
- Śmiga, M.; Roszkiewicz, E.; Ślęzak, P.; Tracz, M.; Olczak, T. cAMP-Independent Crp Homolog Adds to the Multi-Layer Regulatory Network in Porphyromonas gingivalis. Front. Cell. Infect. Microbiol. 2025, 15. [Google Scholar] [CrossRef] [PubMed]
- Townsend, P.D.; Rodgers, T.L.; Glover, L.C.; Korhonen, H.J.; Richards, S.A.; Colwell, L.J.; Pohl, E.; Wilson, M.R.; Hodgson, D.R.W.; McLeish, T.C.B.; et al. The Role of Protein-Ligand Contacts in Allosteric Regulation of the Escherichia coli Catabolite Activator Protein*. J. Biol. Chem. 2015, 290, 22225–22235. [Google Scholar] [CrossRef] [PubMed]
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Bartova, J.; Janatova, T.; Prochazkova, J.; Duskova, J. Porphyromonas gingivalis: Major Periodontopathic Pathogen Overview. J Immunol Res 2014, 2014, 476068. [Google Scholar] [CrossRef] [PubMed]
- Kiley, P.J.; Beinert, H. Oxygen Sensing by the Global Regulator, FNR: The Role of the Iron-Sulfur Cluster. FEMS Microbiol. Rev. 1998, 22, 341–352. [Google Scholar] [CrossRef]
- Reents, H.; Gruner, I.; Harmening, U.; Böttger, L.H.; Layer, G.; Heathcote, P.; Trautwein, A.X.; Jahn, D.; Härtig, E. Bacillus subtilis Fnr Senses Oxygen via a [4Fe-4S] Cluster Coordinated by Three Cysteine Residues without Change in the Oligomeric State. Mol. Microbiol. 2006, 60, 1432–1445. [Google Scholar] [CrossRef]
- Bonnet, M.; Kurz, M.; Mesa, S.; Briand, C.; Hennecke, H.; Grütter, M.G. The Structure of Bradyrhizobium japonicum Transcription Factor FixK2 Unveils Sites of DNA Binding and Oxidation. J. Biol. Chem. 2013, 288, 14238–14246. [Google Scholar] [CrossRef]
- Volbeda, A.; Darnault, C.; Renoux, O.; Nicolet, Y.; Fontecilla-Camps, J.C. The Crystal Structure of the Global Anaerobic Transcriptional Regulator FNR Explains Its Extremely Fine-Tuned Monomer-Dimer Equilibrium. Sci Adv 2015, 1, e1501086. [Google Scholar] [CrossRef]
- Mesa, S.; Reutimann, L.; Fischer, H.-M.; Hennecke, H. Posttranslational Control of Transcription Factor FixK2, a Key Regulator for the Bradyrhizobium japonicum–Soybean Symbiosis. Proc. Natl. Acad. Sci. U.S.A. 2009, 106, 21860. [Google Scholar] [CrossRef]
- The UniProt Consortium UniProt: The Universal Protein Knowledgebase in 2025. Nucleic Acids Res 2025, 53, D609–D617. [CrossRef]
- Pi, H.-W.; Lin, J.-J.; Chen, C.-A.; Wang, P.-H.; Chiang, Y.-R.; Huang, C.-C.; Young, C.-C.; Li, W.-H. Origin and Evolution of Nitrogen Fixation in Prokaryotes. Mol Biol Evol 2022, 39, msac181. [Google Scholar] [CrossRef]
- Davín, A.A.; Woodcroft, B.J.; Soo, R.M.; Morel, B.; Murali, R.; Schrempf, D.; Clark, J.W.; Álvarez-Carretero, S.; Boussau, B.; Moody, E.R.R.; et al. A Geological Timescale for Bacterial Evolution and Oxygen Adaptation. Science 2025, 388, eadp1853. [Google Scholar] [CrossRef]
- Schrödinger Release 2019-2. Schrödinger, LLC: New York, NY, 2019.
Figure 1.
Ribbon depiction of one monomer from the dimeric EcCRP with allosteric cAMP bound (green) and complexed to DNA (blue), pdb code 2CGP. A second cAMP molecule bound between the cAMP-binding and DNA-binding domains is also found in the structure.
Figure 1.
Ribbon depiction of one monomer from the dimeric EcCRP with allosteric cAMP bound (green) and complexed to DNA (blue), pdb code 2CGP. A second cAMP molecule bound between the cAMP-binding and DNA-binding domains is also found in the structure.
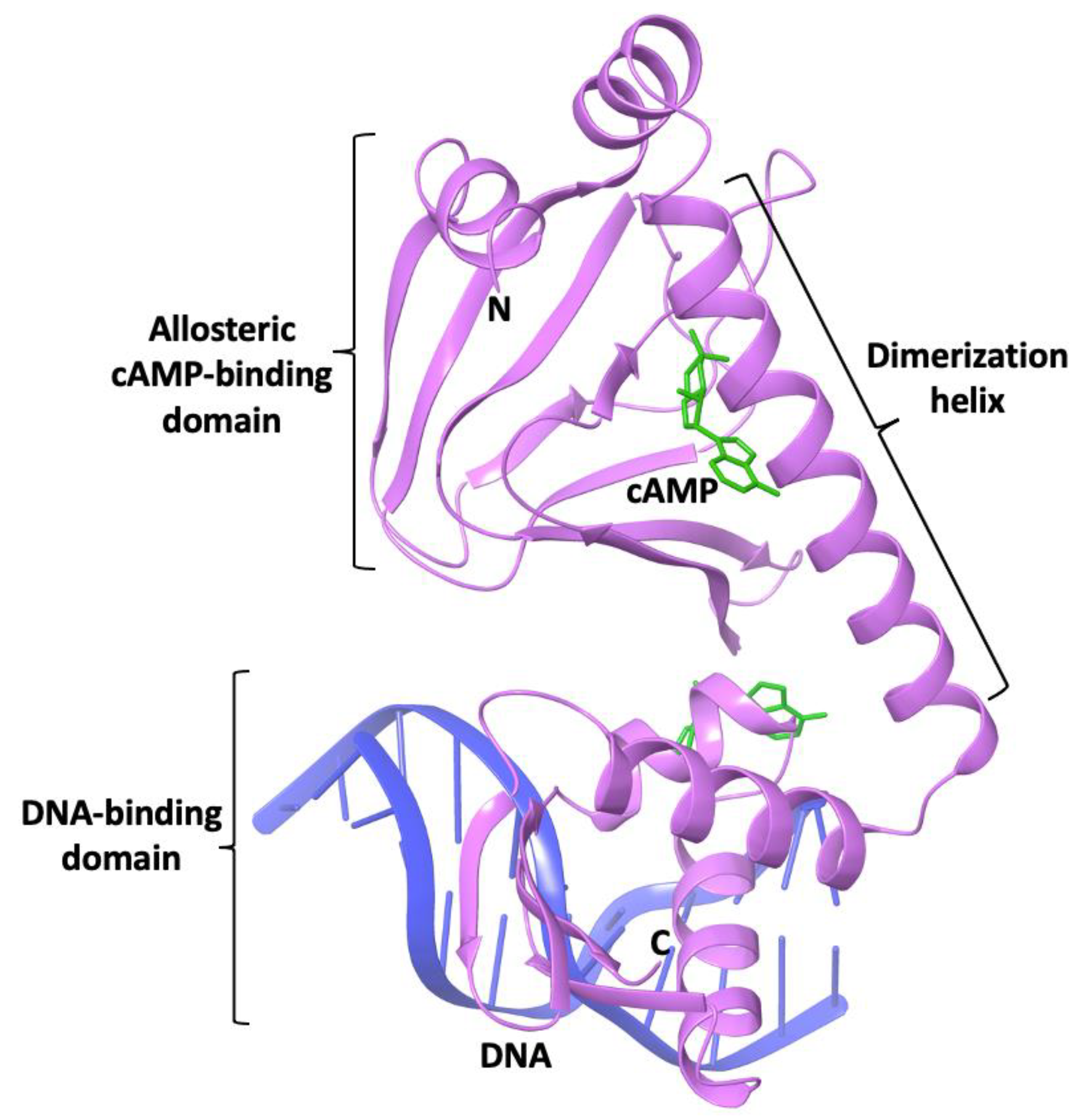
Figure 2.
A. The structure of cyclic AMP. B. Ribbon representation of the conformational change of dimeric EcCRP upon cAMP binding (pdb codes apo 49NH and holo 4HZF) [19]. DNA-binding domains (DBD) are indicated; cAMP molecules are depicted with green sticks.
Figure 2.
A. The structure of cyclic AMP. B. Ribbon representation of the conformational change of dimeric EcCRP upon cAMP binding (pdb codes apo 49NH and holo 4HZF) [19]. DNA-binding domains (DBD) are indicated; cAMP molecules are depicted with green sticks.
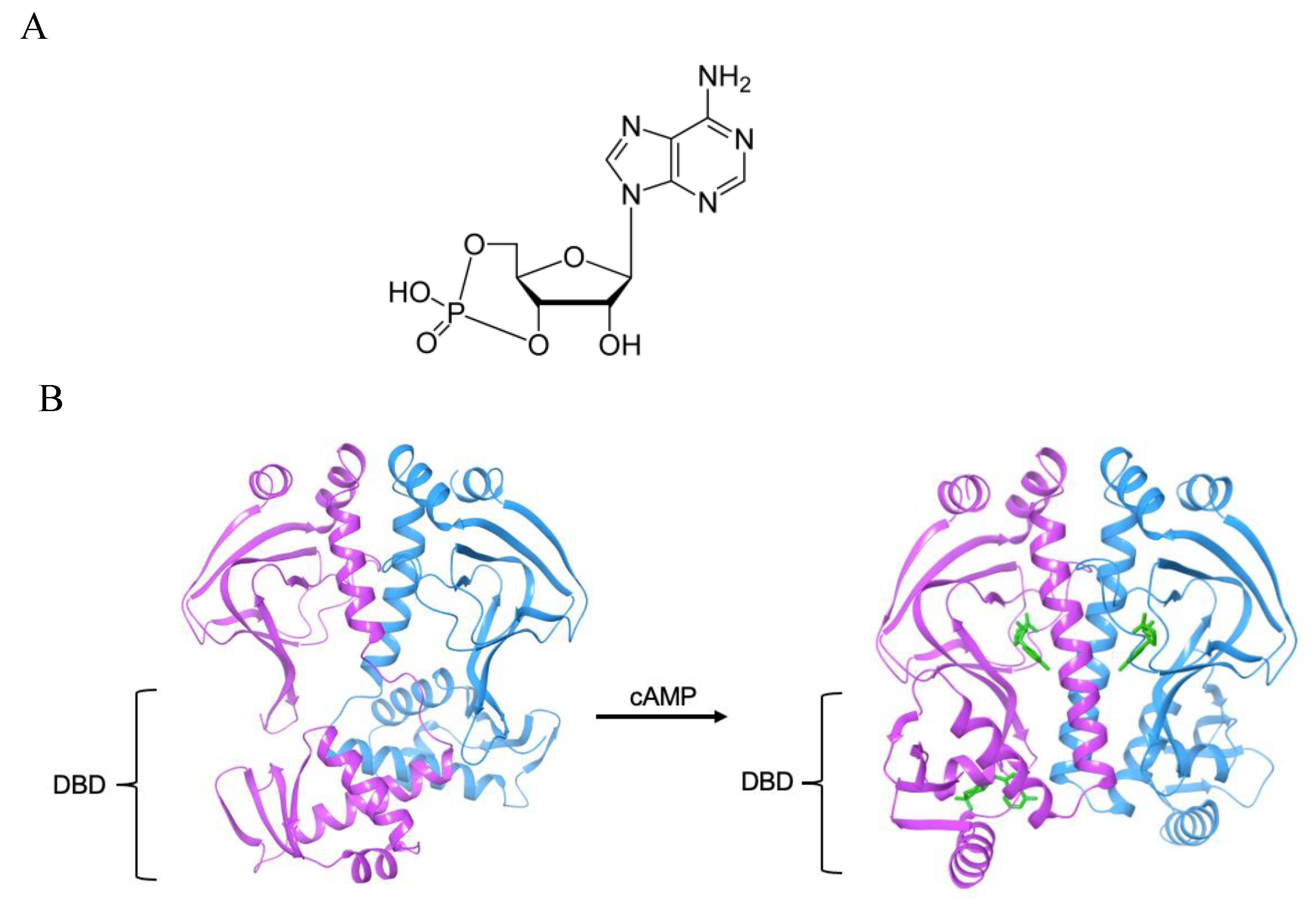
Figure 3.
Depiction of TF ligand-binding regions. A. holo EcCRP + cAMP (pdb 4HZF) and B. NtcA + 2-OG (pdb 3LA2). C. Superposition of holo EcCRP + cAMP (pdb 4HZF, dark grey C atoms) and apo EcCRP (pdb 4N9H, light grey C atoms). D. Superposition of holo NtcA (light grey C atoms, pdb 3LA2) into apo NtcA (dark grey C atoms, pdb 3LA7).
Figure 3.
Depiction of TF ligand-binding regions. A. holo EcCRP + cAMP (pdb 4HZF) and B. NtcA + 2-OG (pdb 3LA2). C. Superposition of holo EcCRP + cAMP (pdb 4HZF, dark grey C atoms) and apo EcCRP (pdb 4N9H, light grey C atoms). D. Superposition of holo NtcA (light grey C atoms, pdb 3LA2) into apo NtcA (dark grey C atoms, pdb 3LA7).
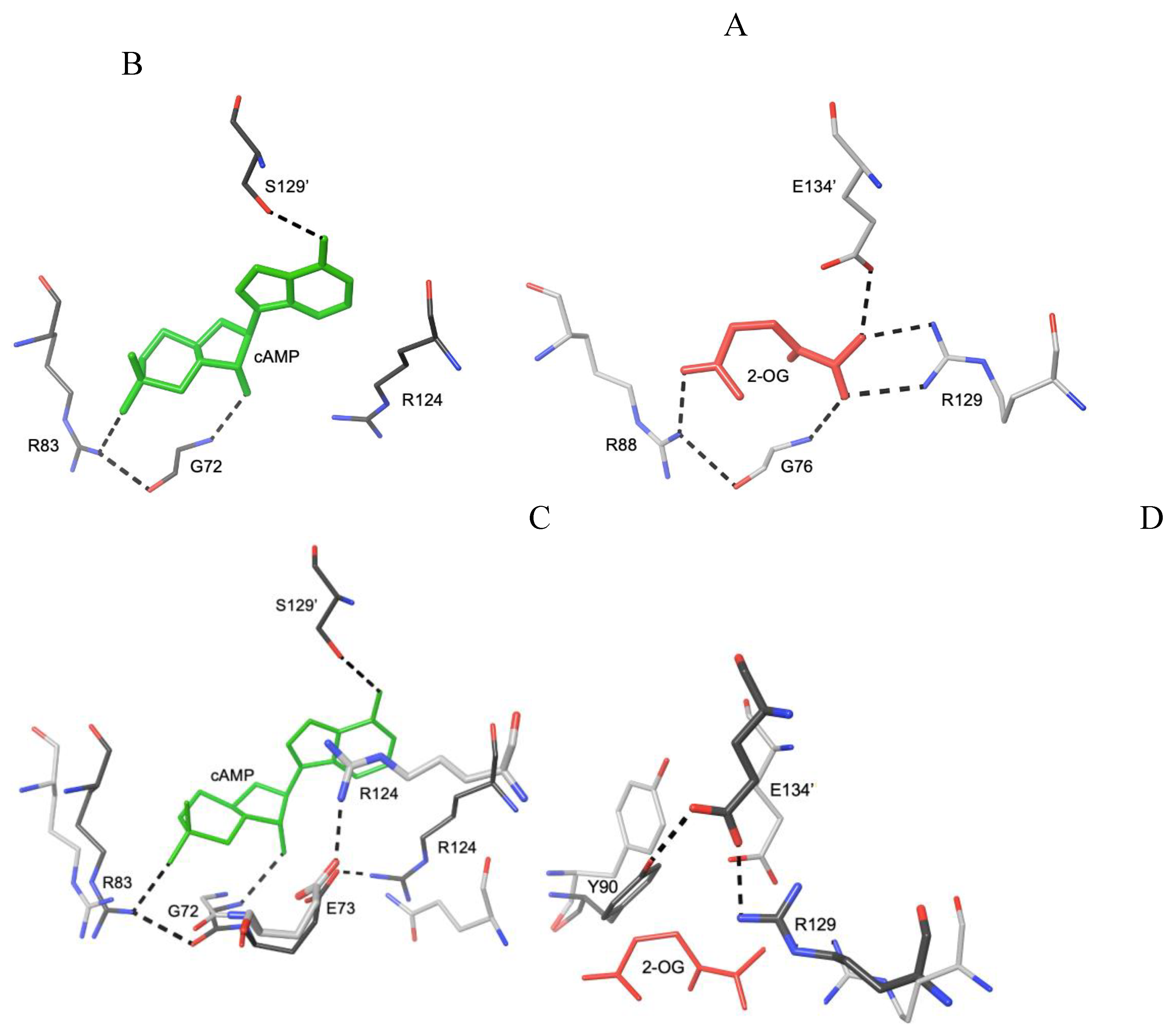
Figure 4.
Stereo depiction of OCPA binding to CprK (red C atoms and Cα tracing, green Cl atom, pdb 3E5U) superposed to cAMP-EcCRP (green cAMP and Cα tracing, pdb 4HZF). The bonding interactions of the ligand with Asn92, Lys86 and Lys133 are highlighted.
Figure 4.
Stereo depiction of OCPA binding to CprK (red C atoms and Cα tracing, green Cl atom, pdb 3E5U) superposed to cAMP-EcCRP (green cAMP and Cα tracing, pdb 4HZF). The bonding interactions of the ligand with Asn92, Lys86 and Lys133 are highlighted.
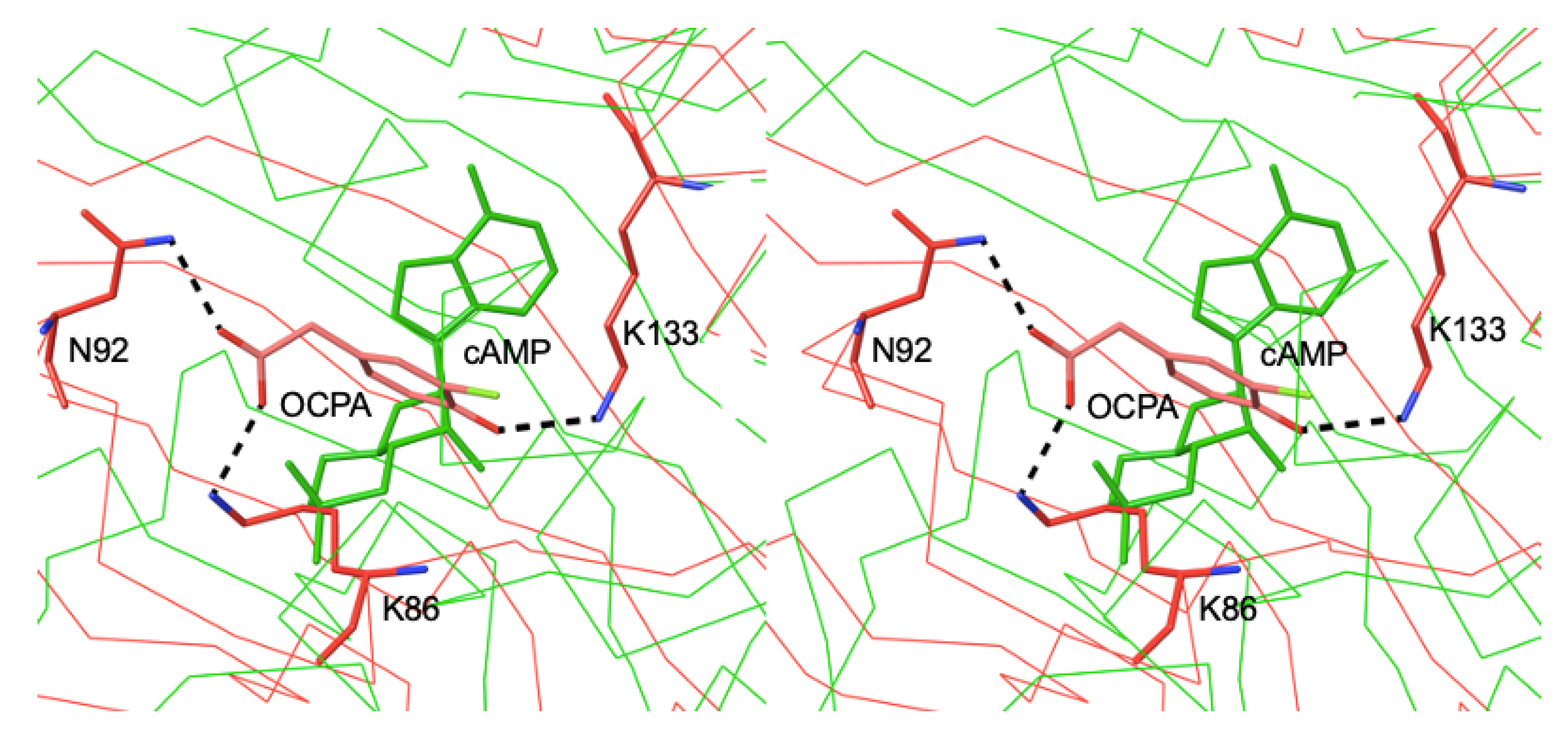
Figure 5.
A. bis-(3’-5’)-cyclic di-guanosine monophosphate. B. Superposition of holo EcCRP + cAMP (green C atoms and Cα tracing), pdb 4HZF, and apo XcClp (beige C bonds and Cα tracing), pdb 3IWZ.
Figure 5.
A. bis-(3’-5’)-cyclic di-guanosine monophosphate. B. Superposition of holo EcCRP + cAMP (green C atoms and Cα tracing), pdb 4HZF, and apo XcClp (beige C bonds and Cα tracing), pdb 3IWZ.
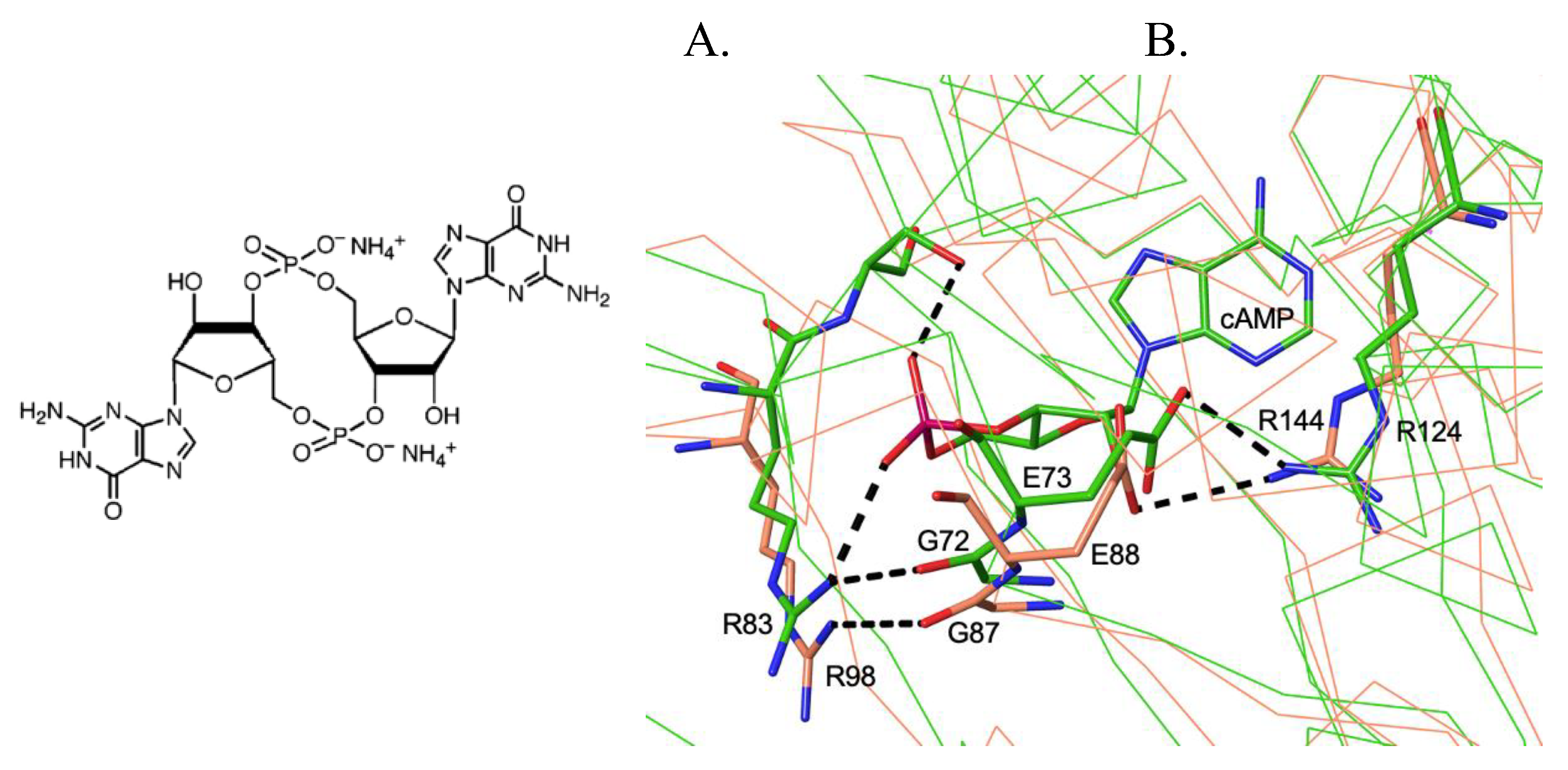
Figure 6.
Stereo view of the DNA-GSH-PrfA crystal structure (PrfA, magenta and orange ribbons, GSH green sticks and DNA blue and green ribbons, pdb code 5X6E) [40].
Figure 6.
Stereo view of the DNA-GSH-PrfA crystal structure (PrfA, magenta and orange ribbons, GSH green sticks and DNA blue and green ribbons, pdb code 5X6E) [40].
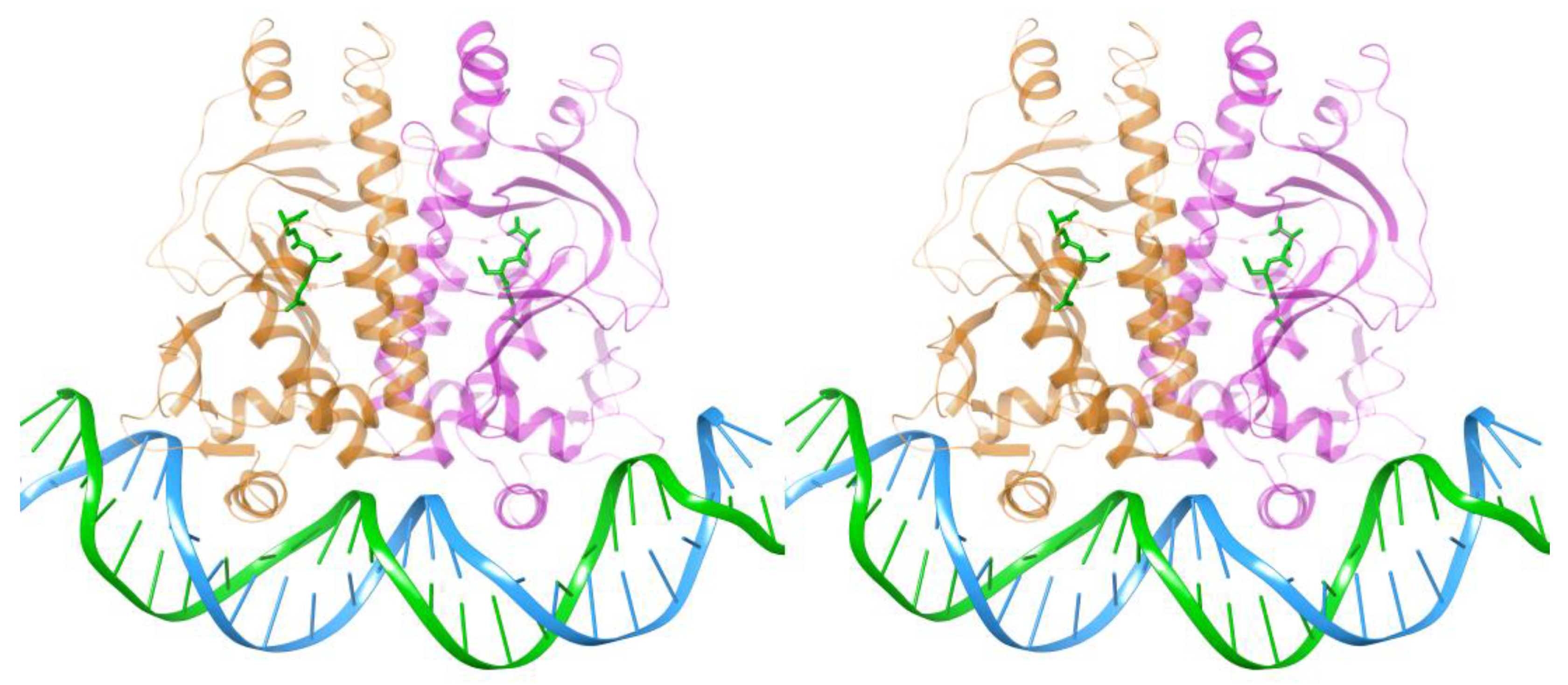
Figure 7.
Stereo depiction of the superposition of the ribbon tracings of EcCRP (pdb 4HZF, blue and orange ribbons) and ChCooA (pdb 2FMY, yellow and magenta ribbons). The cAMP and Im-bound heme b cofactors (green sticks) are labeled in the right-side monomers.
Figure 7.
Stereo depiction of the superposition of the ribbon tracings of EcCRP (pdb 4HZF, blue and orange ribbons) and ChCooA (pdb 2FMY, yellow and magenta ribbons). The cAMP and Im-bound heme b cofactors (green sticks) are labeled in the right-side monomers.
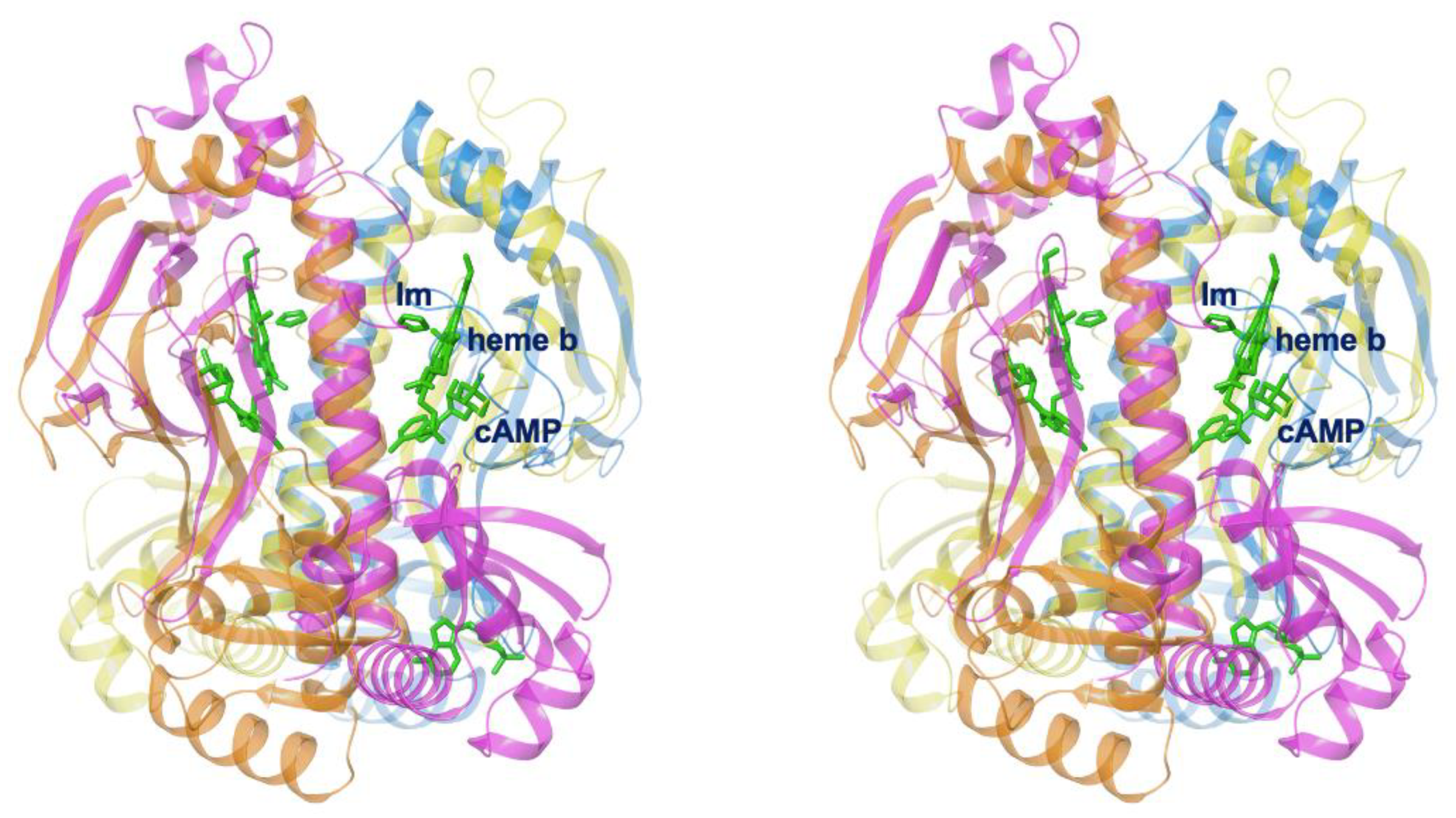
Figure 8.
A. Superposition of the Cα chain of holo EcCRP (pdb 4R8H) + cAMP (green sticks and Cα tracing) to T. thermophilus SdrP (pink C atoms and Cα tracing, pdb 3B02, left) and D. radiodurans DdrI (yellow C atoms and Cα tracing, pdb 8YZ7, right). Equivalent Arg43-Glu53 (left) and Arg55-Glu65 (right) salt bridges are highlighted. In addition, a H-bond between Glu66 and Tyr75 is observed in DrDdrI. The equivalent Glu54 and Tyr63 residues of TtSdrP do not interact in the crystal structure. B. Amino acid sequence alignment of the regions shown in A for T. thermophilus (Tt), D. radiodurans (Dr) and E. coli (Ec). Residues discussed in the text are highlighted (bold: strictly conserved ones).
Figure 8.
A. Superposition of the Cα chain of holo EcCRP (pdb 4R8H) + cAMP (green sticks and Cα tracing) to T. thermophilus SdrP (pink C atoms and Cα tracing, pdb 3B02, left) and D. radiodurans DdrI (yellow C atoms and Cα tracing, pdb 8YZ7, right). Equivalent Arg43-Glu53 (left) and Arg55-Glu65 (right) salt bridges are highlighted. In addition, a H-bond between Glu66 and Tyr75 is observed in DrDdrI. The equivalent Glu54 and Tyr63 residues of TtSdrP do not interact in the crystal structure. B. Amino acid sequence alignment of the regions shown in A for T. thermophilus (Tt), D. radiodurans (Dr) and E. coli (Ec). Residues discussed in the text are highlighted (bold: strictly conserved ones).
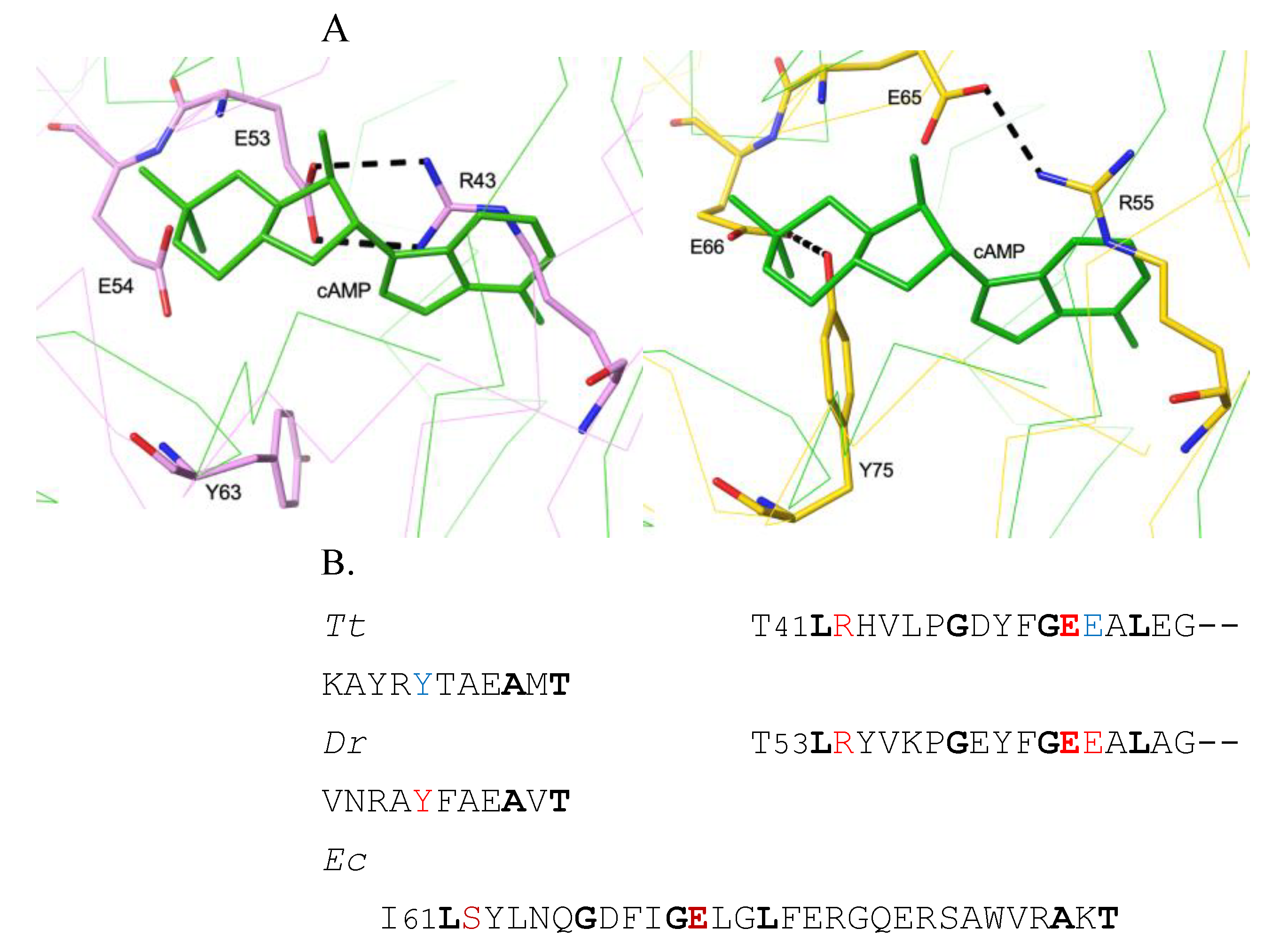
Figure 9.
A. Superposition of the Cα chain of holo EcCRP + cAMP (pdb 4HZF, green) to PgCRP (pdb 2GAU, blue)). B. Same as A plus DrCRP (pdb 8YZ7, yellow) oriented as in Figure 8A-right.
Figure 9.
A. Superposition of the Cα chain of holo EcCRP + cAMP (pdb 4HZF, green) to PgCRP (pdb 2GAU, blue)). B. Same as A plus DrCRP (pdb 8YZ7, yellow) oriented as in Figure 8A-right.
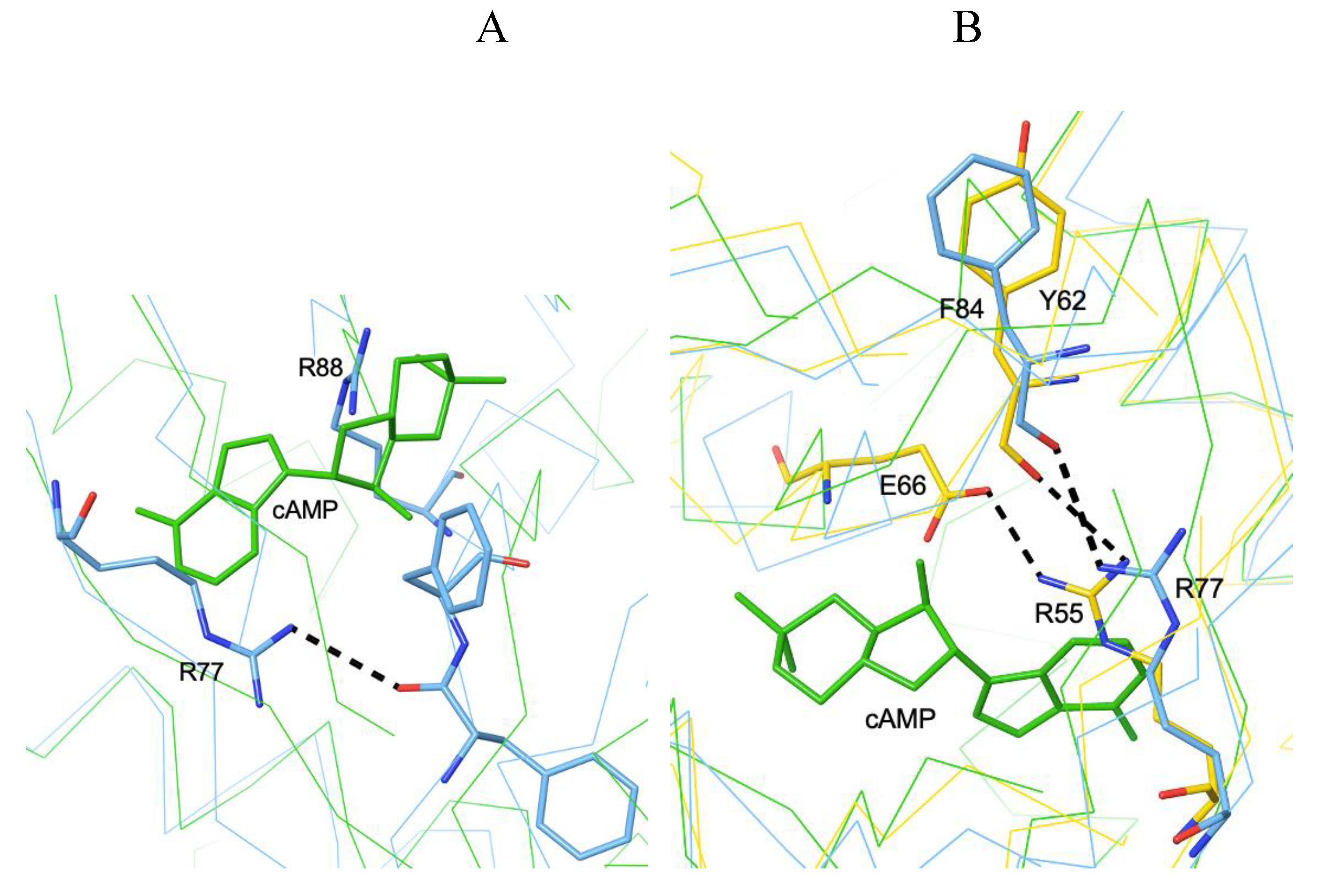
Figure 10.
A. AfFNR (left, pdb 9QLL) and FixK2 (right, pdb 4I2O) N-terD cavities. Respective residues His93, Asp97 and Glu150 and His86, Asp90 and Glu141 are labelled. The superposed cAMP-CRP complex is shown as a Cα chain + cAMP (green sticks). B. Stereo view of the superposition of dimeric AfFNR (yellow and red Cα ribbons, pdb code 9QLL) to the DNA-FixK2 complex (blue and green Cα ribbons, gray DNA strands, pdb code 4I2O) [4Fe-4S] clusters are represented with red Fe and yellow S atoms.
Figure 10.
A. AfFNR (left, pdb 9QLL) and FixK2 (right, pdb 4I2O) N-terD cavities. Respective residues His93, Asp97 and Glu150 and His86, Asp90 and Glu141 are labelled. The superposed cAMP-CRP complex is shown as a Cα chain + cAMP (green sticks). B. Stereo view of the superposition of dimeric AfFNR (yellow and red Cα ribbons, pdb code 9QLL) to the DNA-FixK2 complex (blue and green Cα ribbons, gray DNA strands, pdb code 4I2O) [4Fe-4S] clusters are represented with red Fe and yellow S atoms.
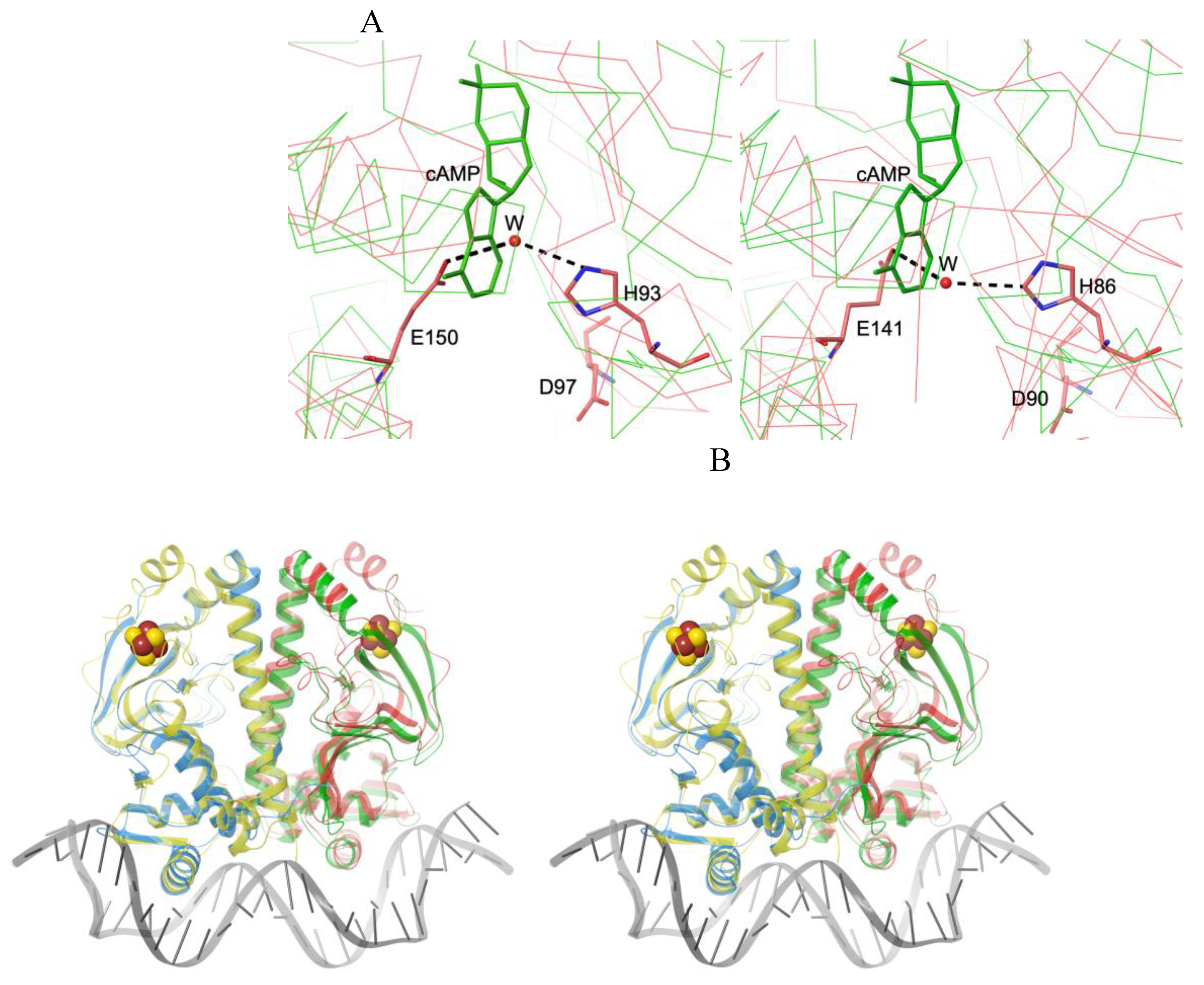
Figure 11.
Superposition of the Cα chain of holo EcCRP (pdb 4HZF) + cAMP (green) to Bacillus sp. gram-positive FNR (modeled AF052953.pdb, purple C bonds, see text).
Figure 11.
Superposition of the Cα chain of holo EcCRP (pdb 4HZF) + cAMP (green) to Bacillus sp. gram-positive FNR (modeled AF052953.pdb, purple C bonds, see text).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



