
Submitted:
16 December 2025
Posted:
17 December 2025
You are already at the latest version
Abstract
Homeostatic systems (nervous, immune, and endocrine) and their communications network are crucial for health and aging rate. The triple transgenic mice for Alzheimer´s disease (3xTg-AD) offer a valuable model to study the pathogenesis of AD, in which oxidative-inflammatory stress is involved. We previously reported behavioral and peritoneal leukocyte function alterations and oxidative-inflammatory stress in young female 3xTg-AD compared to age-matched non-transgenic (NTg) controls. However, it is not known how the deterioration of the homeostatic system interplay occurs at prodromal stages and the effects of sex. Here, we carried out an integrative analysis of the behavioral profile, splenic and thymic leukocyte functions, splen-ic oxidative-inflammatory state and plasmatic corticosterone in male and female 3xTg-AD at 4 months, compared to NTg counterparts. At this prodromal stage, char-acterized by anxiety-like behaviors and disrupted exploration, the 3xTg-AD mice ex-hibited reduced chemotaxis, natural killer activity, and lymphoproliferation—especially in the spleen. These changes were accompanied by lower anti-inflammatory and higher pro-inflammatory cytokine concentrations, and oxidative (higher oxidants and lower antioxidants) stress, indicating a breakdown of neuroimmune and redox-inflammatory homeostasis. Notably, several of these alterations displayed sex-dependent differences. These findings suggest that neuroimmune and redox-inflammatory dysfunctions emerge at the prodromal stage, preceding endocrine changes, and may act as early in-dicators of premature aging in Alzheimer’s pathology.
Keywords:
Alzheimer´s Disease
; 3xTg-AD mice
; prodromal stage
; premature aging
; neuroimmunoendocrine communications
; behavior
; immune functions
; oxidative-inflammatory stress
; spleen
; thymus
; sex effect
; males and females
1. Introduction
More than 30 million individuals suffer from Alzheimer´s disease (AD), the most prevalent chronic neurodegenerative disorder, which is primarily distinguished by extracellular amyloid beta plaques and intracellular neurofibrillary tangles, resulting in impaired synaptic function, neuronal loss, cognitive decline, and neuropsychiatric symptoms [1]. In the coming years, the burden of AD and its economic impact will dramatically increase with the aging of the population [2]. Despite decades of research, there are no effective therapies to deal with the many cellular and molecular alterations of this neurological disorder [3,4]. For this reason, an increasing number of studies have focused on the disclosure of AD pathology before the onset of dementia, trying to find prodromal peripheral markers that allow intervening before the disease is in an advanced stage [5,6,7].
In this context, we must consider the essential role of the homeostatic systems, such as the nervous, immune, and endocrine systems, and their communication to ensure adaptive mechanisms and, therefore, health maintenance. This neuroimmunoendocrine network is fundamental for successful aging and longevity. Thus, its deterioration, which is underpinned by an oxidative-inflammatory stress state, generated in part by the immune system, supposes a premature or accelerated aging as well as an increased risk of morbidity and mortality [8].
Currently, there is emerging research on the relevance of immunity in Alzheimer´s disease, contributing to neuroinflammation and oxidative stress, both within the central nervous system and in the periphery. Thus, the role of the peripheral immune system in this neurodegenerative disease seems evident. In fact, it has been proposed that the involvement of the bidirectional communication between peripheral immune cells and the central nervous system (CNS) accelerates neuroinflammation and cognitive decline [9,10,11,12,13]. Moreover, we have found that several functions of peripheral blood immune cells can serve as markers of AD progression [14].
The late clinical onset of AD in most affected people is a limitation for clinical research when deciphering the cellular and molecular changes at the preclinical (asymptomatic and prodromal) stages of the disease. For this reason, animal models, especially genetic mouse models of AD, are very useful for studying its pathogenesis. Although there are many mouse models of AD [15,16], the triple-transgenic mouse for Alzheimer’s disease (3xTg-AD), harboring PS1M146V, APPSwe, and tauP301L transgenes, represents an interesting model [17]. It mimics many critical hallmarks of AD neuropathology, such as both amyloid-beta and tau neuropathology in an age-dependent manner and disease-relevant brain regions, reproducing a similar temporal and anatomical specific profile observed in humans. This model presents synaptic and cholinergic deficits, the characteristic reactive gliosis inflammatory profile, as well as cognitive impairment [18,19,20,21,22,23,24,25] . In addition, since our first reports [26], we have consistently proposed the 3xTg-AD mouse as a model of special interest to study neuropsychiatric-like symptoms of dementia since it exhibits several behavioral alterations already before the early signs of intraneuronal beta-amyloid that worsen in an age-dependent manner along with changes at the neuroanatomical level [27].
The crosstalk between behavior (an indicator of nervous function) and the peripheral immune system plays a crucial role, particularly during the prodromal stages of disease [28]. In a previous longitudinal study carried out only in female 3xTg-AD mice we analyzed several functions of peritoneal immune cells [29,30], but the functional state of spleen and thymus leukocytes was assessed in older animals, at the end point [31,32]. Moreover, in this model, splenomegaly, histopathological changes and splenic dysfunction have been associated with advanced stages of disease [33,34,35]. However, the state and interplay of the components of the neuroimmunoendocrine system in these 3xTg-AD mice have not been studied in an integrated way at an early stage of the disease.
As mentioned before, oxidative stress has been considered a crucial central factor in the pathogenesis of AD, as a bridge that connects the different mechanisms of this disease [36], being an early event in AD progression. Thus, with respect to the corresponding NTg, peritoneal immune cells of 2-month-old 3xTg-AD female mice showed oxidative stress and higher lipid peroxidation [29,30]. Moreover, a higher presence of oxidants has been observed in the brain and serum [37,38] of these animals at 2-3 months of age. However, in several parameters, these differences disappear after 6 months of age. In addition, inflammatory stress (higher presence of pro-inflammatory than the anti-inflammatory compounds), a process associated with oxidative stress [39], is a common feature in both the brain and the peripheral immune system, contributing to pathogenesis and progression of AD [40]. In the 3xTg-AD mice, higher brain pro-inflammatory mediator expression has been observed at 6 and 12 months of age following parasitic infection [41]. In peritoneal immune cells of 3xTg-AD female mice, higher ratios of proinflammatory/anti-inflammatory concentrations in 3xTg-AD than in NTg mice at 2 and 4 months have been observed [30]. Nevertheless, the oxidative and inflammatory stress in immune organs at a prodromal stage of the disease is unknown. Finally, a relevant endocrine component in neuroimmune communication, the corticosterone hormone, which is also related to an inflammatory state, has been studied in 3xTg-AD mice at old ages [31] or in a social isolation stress response [42], but its plasma concentrations in these mice at 4 months of age have not been analyzed.
The typical age-related deterioration of the homeostatic systems and their communication, which is associated with an oxidative-inflammatory stress, is involved in the rate of aging and longevity [8] and can appear at a young age and in adults. Thus, in several murine models of premature aging, which is related to a shorter lifespan of these animals than that of their age-matched controls, premature alterations in behavioral responses, immunosenescence, and oxi-inflammaging have been observed [39,43]. However, in these young or adult animals that prematurely age, intervention with healthy lifestyle strategies allows their successful aging and greater longevity [39]. In this context, it is interesting to know the peripheral markers of immune and redox, and inflammatory state at early ages in 3xTg in order to have a better basis to test strategies to prevent or delay the symptoms of this disease.
Regarding sex-dependent differences observed in AD [44,45], these are relevant to neuroimmunoendocrine communication and the longevity of animals, including humans and mice [34,46]. In 3xTg-AD mice, sex differences have been observed in cerebral pathology, behavioral responses, as well as in several aspects of immunity and oxidative-inflammatory state [47,48,49,50,51,52].
For all the above-mentioned, the present work aimed to do an integrative study on the emergence of dysfunctions in the different components of the psychoneuroimmunoendocrine network in male and female 3xTg-AD mice at the prodromal stage (4 months of age) of the disease. Thus, their prodromal state was verified by several behavioral parameters, the endocrine system analyzing plasma corticosterone concentrations and weight of adrenal glands, and the immune system studying several leukocyte functions (chemotaxis, natural killer activity, lymphoproliferation, and cytokine release). Moreover, since the base neuroimmunoendocrine alterations are an oxidative-inflammatory stress, several markers of this stress were also measured.
2. Materials and Methods
2.1. Animals
Four-month-old male and female 3xTg-AD mice from the Spanish colony of homozygous 3xTg-AD mice, established in the Medical Psychology Unit, Universitat Autònoma de Barcelona, were used in the present study. The 3xTg-AD mouse strain harboring the familial AD mutations PS1M146V, APPswe, and tauP301L transgenes was genetically engineered at the University of California Irvine, as previously described [17,18]. The non-transgenic (NTg) mouse colony had the same genetic hybrid background (129 × C57BL6) as 3xTg-AD. Genotypes were confirmed by polymerase chain reaction (PCR) analysis of DNA obtained from tail biopsies. Three to five littermates of the same genotype and sex were maintained in Macrolon cages (Techniplast, Buguggiatta, Italy, 35 × 35 × 25 cm) under standard laboratory conditions of food and water ad libitum, temperature (22 ± 2 ºC) and humidity (50-60%), on a 12/12 hour light/dark cycle (lights on at 08:00). Estrous cycle of the female mice was not determined to avoid stressing the animals by the vaginal smear analyses. However, it is known that most female mice housed in groups in the same room spontaneously synchronize their estrous cycles [53]. This reduced the possibility of a hormone bias in the comparison of the different experimental groups.
2.2. Experimental design
Thirty-eight 3xTg-AD mice and sex- and age-matched NTg mice were assessed for behavior responses in the corner test (CT), open-field (OF), and T-maze (TM) [26], which were evaluated by direct observation and VideoTrack analysis system (ViewPoint Behavior Technology, Lyon, France). Experiments were conducted under dim white-light conditions (16-20 lux) from 10:00 a.m. to 1:00 p.m. Two days after the behavioral assessment, the animals were sacrificed by decapitation to obtain blood immediately, and the target samples (thymus, spleen, and adrenal glands) were removed and rapidly frozen, and stored at -80 ºC until the assays were performed.
All the experiments were conducted in accordance with the Spanish legislation on “Protection of Animals Used for Experimental and Other Scientific Purposes” and the EU Directive (2010/63/EU) on this subject, and the Ethical Commission of the Universitat Autònoma de Barcelona (Protocol Number CEEAH/00759 3588/DMAH 9452).
2.3. Prodromal behavioral profile
The prodromal behavioral state of animals was assessed by means of a battery of three tests:
Corner test (CT) for neophobia: The fear of a new home-cage was assessed by introducing the animal into the center of the standard cage (Macrolon, 35 × 35 × 25 cm) and counting the number of visited corners and rearing during a period of 30 s. Latency to the first rearing was also recorded [26].
Open-field test (OF): Mice were placed in the center of the apparatus (homemade, wooden, white, 50 × 50 × 25 cm high) and observed for 5 min [51]. Horizontal (crossings of 10 × 10 cm squares) and vertical (number of rearing) locomotor activities were recorded for each minute of the test. We also recorded the ethogram, the latency of the sequence of the following behavioral events: initial freezing (latency of initial movement), thigmotaxis or discrimination of unprotected/protected areas in the test (latency of leaving the central 10 × 10 cm square and that of entering in the peripheral ring 5 cm to the walls), and self-grooming behavior (latency, number, and duration of grooming). Emotionality (defecation and the presence/absence of urine) was also measured.
T-maze (TM): The spontaneous exploratory behavior and alternation of mice were tested in a T-shaped maze with “horizontal” arms 25 cm in length. Animals were placed inside the “vertical” arm of the maze (35 cm in length) with their head facing the end wall. The copying with stress strategies performance was evaluated by determining the time elapsed until the animal crossed (four-paw criteria) the intersection of the three arms [43]. The test was completed once the animal had finished exploring the two “horizontal arms”. The exploratory efficiency was measured by means of different variables: the total time invested to complete the maze exploration, the number of errors (revisiting an arm) made, and the ratio between arms (time needed to complete the test divided by the latency to reach the intersection). Emotionality (defecation and the presence/absence of urine) was also recorded.
2.4. Organometrics
The size (weight in mg) and relative size (% vs. body weight) of peripheral immunoendocrine organs, including the thymus, spleen, and adrenal glands, were recorded during the harvesting, before the analysis of peripheral immune functions and corticosterone assay [51].
2.5. Immune functions
2.5.1. Collection of lymphocyte suspensions
Spleen and thymus were removed aseptically, freed of fat, minced with scissors, and gently pressed through a mesh screen (Sigma-Aldrich, St. Louis, USA). The collection of lymphocytes was assessed according to the method previously described [43]. Spleen suspensions, due to their high concentration of erythrocytes, were centrifuged in a gradient of Ficoll-Hypaque (Sigma-Aldrich) with a density of 1.070g/mL. Cells were recollected from the interface and resuspended in complete medium [containing RPMI 1640 enriched with L-glutamine and phenol red and supplemented with 10% heat-inactivated (56ºC, 30 min) fetal calf serum (Hyclone, GE Healthcare, USA) and gentamicin (100 mg/mL, Sigma-Aldrich)]. Lymphocytes were washed, and their number was determined, and adjusted to 106 cells/mL. Cellular viability, routinely measured before and after each experiment by the Trypan-Blue exclusion test, was higher than 98% in all experiments. All incubations were performed at 37ºC in a humidified atmosphere of 5% CO2.
2.5.2. Chemotaxis assay
The induced mobility or chemotaxis of leukocytes was evaluated according to the method previously described [43]. Chambers with two compartments, separated by a 3 μm pore diameter filter (Millipore, Ireland), were used. Aliquots of 300 μL of splenic or thymic lymphocyte suspensions adjusted to 106 cells/mL in Hank’s medium were deposited in the upper compartment, and 400 μL of the chemoattractant agent formyl-methionyl-leucine-phenylalanine (f-met-leu-phe, Sigma-Aldrich) at a concentration of 10-8 M was deposited in the lower compartment. The chambers were incubated for 3 h, the filters were fixed and stained, and the number of lymphocytes on one-third of the lower face of the filter (corresponding to five scans of 5 mm) was counted using an optical microscope (×100 magnification) and recorded as Chemotaxis Index (C.I.).
2.5.3. Natural Killer (NK) cytotoxicity assay
The NK cell cytotoxicity, which is the main antitumoral protection of the organism, was measured by an enzymatic colorimetric assay (Cytotox 96 TM Promega, Boehringer Ingelheim, Germany) based on the determination of lactate dehydrogenase (LDH) released by the cytolysis of targets cells (YAC-1 cells from a murine lymphoma), using tetrazolium salts (ID) [43]. Briefly, target cells were seeded in 96-well U-bottom culture plates (Nunclon, Denmark) at 104 cells/well in 1640 RPMI medium without phenol red (Gibco). Effector cells (splenic or thymic lymphocytes suspensions adjusted to 106 cells/ mL) were added at 105 cells/well, obtaining an effector/target rate of 10/1. The plates were centrifuged at 250 g for 5 min to facilitate cell-to-cell contacts and were incubated for 4 h at 37°C. After incubation, LDH activity was measured in 50 μL/well by adding the enzyme substrate, with absorbance recorded at 490 nm. The results were expressed as the percentage of tumor cells killed (% lysis).
2.5.4. Lymphoproliferation assay
The proliferation capacity of lymphocytes was evaluated by a standard method, previously described [43]. The assay was assessed in basal and stimulated conditions using the mitogens concanavalin A (Con-A) and lipopolysaccharide (LPS). Aliquots of 200 μL of splenic or thymic leukocyte (mainly lymphocytes) suspensions adjusted to 106 cells/mL in complete medium were dispensed into 96-well plates (Nunclon, Denmark). 20 μL of complete medium (basal lymphoproliferation), Con-A or LPS (1 μg/mL, Sigma-Aldrich) was added to each well. After 48 h of incubation at 37°C in a sterile and humidified atmosphere of 5% CO2, 0.5 μCi 3H-thymidine (Hartmann Analytic, Germany) was added to each well. Previously, 100 μL of culture supernatant from each well was collected and stored at −80°C until used for harvesting in a semiautomatic harvester (Skatron Instruments, Norway). Thymidine uptake was then measured using a beta counter (LKB, Uppsala, Sweden) for 1 min. The results were calculated as 3H-thymidine uptake (counts per minute, c.p.m) for basal and stimulated (with mitogens) conditions and were also expressed as lymphoproliferation capacity (%), giving 100% to the c.p.m in basal conditions.
2.5.5. Cytokine concentrations in cultures of spleen leukocytes
The release of interleukin-2 (IL-2), interleukin-10 (IL10), interleukin-1beta (IL-1β) and tumor necrosis factor alpha (TNF-α) were measured in culture supernatants of splenic lymphocytes after 48 h of incubation in the presence of Con-A (1μg/mL) or LPS (1 μg/mL) as described above. Levels of IL-2, IL-10, IL-1β, and TNF-α were measured simultaneously by multiplex luminometry (MILLIPLEX® MAP Mouse Cytokine/Chemokine kit, Immunoassay, Millipore, Billerica, MA, USA). Each assay was run in duplicate. The minimum detectable concentration of IL-2, IL-10, IL-1β, and TNF-α was 3.2 pg/mL. The results were expressed as pg/mL.
2.6. Redox-inflammatory state
2.6.1. Glutathione peroxidase (GPx) activity
The GPx activity was spectrophotometrically evaluated in homogenates of the spleen according to the method described by Lawrence and Burk [54], with some modifications. Briefly, tissue samples of spleen were homogenized (50 mg/mL) with tampon phosphate (50 mM, pH 7.4), being maintained on ice during the entire process. Thereafter, homogenates were centrifuged at 3200g for 20 min at 4°C. Aliquots of the supernatants were measured in the spectrophotometer using cumene hydroperoxide (cumene-OOH, Sigma-Aldrich) as substrate, which carried out the oxidation of glutathione, which is regenerated by the addition of β-nicotinamide adenine dinucleotide phosphate, in its reduced form (β-NADPH, Sigma-Aldrich), in the presence of glutathione reductase (GR, 10 U/mL, Sigma-Aldrich). The reaction was monitored spectrophotometrically by measuring the decrease in absorbance at 340 nm every 40 s over 5 min. The results were expressed as international milliunits of enzymatic activity per mg of tissue (mU GPx/mg tissue).
2.6.2. Glutathione reductase (GR) activity
The GR activity was spectrophotometrically assessed in homogenates of the spleen according to the method described by Massey and Williams [55], with some modifications. Briefly, tissue samples of the spleen were homogenized in a tampon phosphate solution (50 mM, pH 7.4) containing EDTA (6.3 mM). Homogenates were centrifuged at 3200 g for 20 min at 4°C. Aliquots of the supernatants were measured in the spectrophotometer using the β-NADPH (6 mM) and oxidized glutathione (GSSG, 80 mM, Sigma-Aldrich) reagents. The reaction was monitored spectrophotometrically by measuring the decrease in absorbance at 340 nm every 40 s over a 4-minute period. The results were expressed as international milliunits of enzymatic activity per mg of tissue (mU GR/mg tissue).
2.6.3. Total glutathione (GSH) content
Total glutathione (GSH), the main non-enzymatic reducing agent of the organism, was assayed by the enzymatic recycling method previously described by monitoring the change in absorbance at 412 nm, with some modifications in organs [43]. Briefly, samples of spleen were homogenized (50 mg/mL) with 5% trichloroacetic acid (TCA, Panreac) and 0.01 N HCl (Panreac), being maintained on ice throughout the process. Homogenates were centrifuged at 3200 g for 5 min at 4°C. Aliquots of the supernatants were measured in the spectrophotometer using the following reaction mixture: 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB, 6 mM, Sigma-Aldrich), β-nicotinamide adenine dinucleotide phosphate, reduced form (β-NADPH, 0.3 mM, Sigma-Aldrich), and glutathione reductase (GR, 10 U/mL, Sigma-Aldrich). The reaction was monitored for 240 s, and the results were expressed as nanomoles of GSH per mg of tissue (nmol GSH/mg tissue).
2.6.4. Xanthine oxidase (XO) activity
The XO activity was assayed in homogenates of spleen by fluorescence using a commercial kit (Amplex Red Xanthine/Xanthine Oxidase Assay Kit, Molecular Probes, Paisley, UK) [43]. In the assay, XO catalyzes the oxidation of purine bases (xanthine) to uric acid and superoxide. The superoxide spontaneously degrades in the reaction mixture to H2O2, which in the presence of horseradish peroxidase (HRP), reacts stoichiometrically with Amplex Red reagent to generate the red-fluorescent oxidation product, resorufin. Tissue samples of the spleen were homogenized in phosphate buffer (50 mM, pH 7.4) containing EDTA (1 mM) and normalized according to total protein. The homogenates were centrifuged at 5000 g for 30 min at 4°C. Aliquots of the supernatants (50 µL) were incubated with 50 µL working solution of Amplex Red reagent (100 µM) containing HRP (0.4 U/mL) and xanthine (200 µM). After 30 minutes of incubation at 37 ºC, fluorescence measurement was performed using a microplate reader (Fluostar Optima, BMG Labtech, Biomedal, Spain) with excitation at 530 nm and emission detection at 595 nm. The XO supplied in the kit was used as the standard, and XO activity was measured by comparing the fluorescence of the samples with that of the standards. The protein content of the same samples was evaluated using the bicinchoninic acid protein assay kit protocol (Sigma-Aldrich), with serum albumin (BSA, Sigma-Aldrich) as the standard. The results were expressed as international milliunits (mU) of enzymatic activity per milligram of protein (mU XO/mg protein).
2.7. Plasma corticosterone.
Samples of about 1mL of whole trunk blood were collected at the time of sacrifice into heparinized tubes and centrifuged immediately at 10,000 ×g for 2min. The plasma obtained was stored at–20◦C. Corticosterone content (ng/mL) was analyzed using a commercial kit (Corticosterone EIA Immunodiagnostic Systems Ltd, Boldon, UK) and ELISA EMSReader MFV.2.9-0. 2.6. [51].
2.8. Statistical analysis
Statistical analysis was performed in SPSS IBM 29.0 (SPSS, Chicago, USA) and GraphPad Prism 10.1.1 (LLC, San Diego, CA, USA). The results are presented as the mean ± standard error (SEM). Normality of the samples and homogeneity of the variances were checked by the Kolmogorov–Smirnov test and Levene’s test, respectively. All tests were two-tailed. Differences were studied through two-way ANOVA. The factors were genotype and sex. The Tukey test was used for post-hoc comparisons when variances were homogeneous, whereas its counterpart, the Games-Howell analysis, was used in the case of unequal variances. In all cases, p < 0.05 was taken as statistically significant.
3. Results
3.1. Prodromal behavioral state of 4-month-old male and female 3xTg-AD mice
The study first verified the prodromal behavioral state of the 3xTg-AD as compared to sex-matched counterparts. Thus, the behavioral profile exhibited by the animals in the three behavioral tests was analyzed in terms of genotype, sex, and interaction effects, as summarized in Table 1. Thereafter, among the main variables showing statistically significant effects (as illustrated in Figure 1), the most sensitive variables to these factors were identified.
The first BPSD-like symptoms were already apparent in the 3xTg-AD mice. The sensitivity of the corner test to define the prodromal stage of 3xTg-AD mice was shown as a higher neophobia than the NTg mice, which could be measured not only by the latency (see Table 1, G**, p < 0.01) but also by a lower number of rearings (Table 1, G***, p < 0.001). As illustrated in Figure 1 a, this neophobia reached the highest levels in male 3xTg-AD mice as compared to all the other groups (**, p < 0.01 vs. NTg counterparts; ++ p < 0.01 vs. female counterparts).
When the animals were confronted with the open-field test, 3xTg-AD mice showed higher latencies in most of the behavioral events (Table 1, G*, p < 0.05). The incidence of bizarre behavior in the male and female 3xTg-AD mice was higher than in NTg mice (males, p < 0.001 and females, p < 0.5, respectively). Statistically significant differences in the time course of the vertical component (rearings) were recorded, mostly in females in minutes 2 and 3 (p < 0.05). Exhibition of emotional behaviors was lower in 3xTg-AD mice (Table 1, G*, p < 0.05) and male sex (S**, p < 0.01). Here and in the T-maze, NTg females showed a statistically significant higher number of defecation boli in the open-field test as compared to all the other groups (p < 0.05).
In the T-maze, genotype differences were observed in the latency to reach the intersection (Table 1, Figure 1h, p < 0.05). Post-hoc tests indicated that in this test were 3xTg-AD female mice who exhibited statistically significantly higher values compared to all other groups. The genotype effect can also be observed when the ratio between the time spent in the vertical and horizontal arms is considered (Table 1, p < 0.05), while the total time invested in completing the maze exploration (Table 1) did not differ between groups. A genotype × sex effect could be found in the number of errors (Figure 1i, Table 1, G × S*, p < 0.05). The emotional behavior showed sex differences with females doing more defecation boli and exhibiting a higher incidence of urination. This sex effect was primarily due to female NTg mice, which exhibited higher emotional behaviors compared to all other groups (Figure 1j, Table 1, p < 0.05).
Therefore, the factorial analysis (as detailed in Table 1) pointed out to the strong predominance of genotype factor, as it was observed in most of the variables studied, but also revealed their different magnitudes of sensitivity to these factors (Table 1, from G*, p < 0.05 to G***, p < 0.01). Sex differences (S**, p < 0.01) were found in emotional related variables such us the latency of vertical activity in the corner test (Figure 1a, Table 1, S** p < 0.01), the incidence of bizarre behaviors in the open field test (Figure 1b, Table 1, S** p < 0.01) and the defecatory behavior in the open field test and the T-maze (Figure 1g and Figure 1j, respectively; Table 1, S** p < 0.01). With regards to variables sensitive to one, both factors or their interaction, the time course of the vertical activity (Figure 1e) was sensitive to genotype (rearing on minute 2, and 3, both G*, p < 0.05) and sex (rearings on minute 4, S*, p < 0.05). Some variables also showed genotype per sex interaction effects (G × S*, p < 0.05 and G × S**, p < 0.01).
Among all the behavioral variables studied in the three behavioral tests to characterize the prodromal stage of 3xTg-AD mice, two were the most sensitive to genotype, sex and interaction effects: 1) The latency of rearing in the corner test (Figure 1a and Table 1), with genotype ( G**, p < 0.01), sex (S**, p < 0.01) and genotype per sex interaction effects (G × S*, p < 0.05) and male 3xTg-AD exhibiting statistically significant higher latencies than all the other groups (Figure 1a, post-hoc, * and +, both p < 0.05); 2) The incidence of bizarre behaviors in the open-field test, with genotype (G***, p < 0.001), sex (S**, p < 0.01) and genotype-by-sex (G × S**, p < 0.01) interaction effects, with male 3xTg-AD mice exhibiting statistically significant higher incidence than all the other groups (Figure 1a, post-hoc, *** p < 0.001 and ++, p < 0.01).
3.2. Changes in physical condition, organometrics, and the endocrine system are sex-dependent and also observed at 4 months of age in male and female 3xTg-AD.
The weight of animals as well as the size (weight in mg) and relative size (% vs. body weight) of peripheral immunoendocrine organs, thymus, spleen, and adrenal glands, were recorded before the analysis of peripheral immune functions and the endocrine system. The results (Table 2) confirmed sexual dimorphism in body weight, with males being heavier than females (p < 0.001). Additionally, the overweight phenotype described for 3xTg-AD mice [16,26] was already present at this age (genotype effect, p < 0.001) and was attributed to a higher relative weight of white adipose tissue (WAT) (genotype and sex effects, both p < 0.001). Genotype differences were also observed in the relative weight of immunological organs, with a lower weight of thymus and a higher weight of spleen in 3xTg-AD mice (genotype effect, both p < 0.001). The effect of sex was also observed in the weight of the thymus, which was smaller in male than female mice (p < 0.05).
At the endocrine level, although the weight of the adrenal glands did not differ between genotypes or sexes, a statistically significant genotype-by-sex effect (p < 0.05) was found, indicating a higher weight of this endocrine gland in male 3xTg-AD mice. A slight trend of higher plasma corticosterone concentrations in 3xTg-AD mice was observed, albeit it did not reach statistical significance (p > 0.05).
3.3. An impairment of the immune functions is presented in 4-month-old female and male 3xTg-AD mice
Several immune function parameters were studied in splenic and thymic leukocytes from female and male 3xTg-AD and NTg mice. The results are shown in Figure 2, Figure 3 and Figure 4 and Table 3 and Table 4. In general, at 4 months old, 3xTg-AD mice showed an impairment of the immune functions in their splenic and thymic leukocytes.
Regarding chemotaxis (Figure 2), our results showed that female and male 3xTg-AD mice had lower values of chemotactic index in their spleen (p<0.05 and p < 0.001 in females and males, respectively; Figure 2a) and thymus (p < 0.01 and p < 0.001 in females and males, respectively; Figure 2b) leukocytes, than those observed in gender-matched NTg mice. Regarding gender differences, the lower values of chemotaxis observed in leukocytes of 3xTg-AD mice were more pronounced in males than in females (p < 0.001). By contrast, male NTg mice showed higher chemotaxis index in their splenic leukocytes than NTg female mice (p < 0.05), whereas no gender differences were observed in thymic lymphocytes
An early impairment of NK cytotoxic activity (Figure 2) was also observed for spleen cells from female (p < 0.05) and male (p < 0.01) 3xTg-AD mice compared to gender-matched NTg (Figure 2c), whereas no statistical differences in NK activity were found in thymus lymphocytes between both groups of mice (Figure 2d). Regarding gender differences, both male NTg and 3xTg-AD mice exhibited higher NK activity in their splenic (p < 0.05, in NTg) and thymic (p < 0.001) leukocytes compared to female mice.
With respect to basal proliferation (Figure 3a.1-b.1), which is what takes place in the absence of a proliferative stimulus, there was a significantly lower value (p<0.01) in spleen cells from male 3xTg-AD mice as compared to male NTg mice, while no differences between 3xTg-AD and NTg female mice were observed (Figure 3a.1). Moreover, no statistical differences were found in basal proliferation of thymus lymphocytes between 3xTg-AD and NTg mice (Figure 3b.1). Interestingly, in both splenic and thymic leukocytes, male NTg mice showed higher basal proliferation (p < 0.001 in spleen; p < 0.01 in thymus) than female NTg mice. Similar results were also observed in thymic cells of male 3xTg-AD (p < 0.001) compared to female 3xTg-AD mice (Figure 3b.1).
The results of the lymphoproliferation after incubation with the mitogens LPS and ConA, which are specific for B and T lymphocytes, respectively, showed an impairment of this function in 3xTg-AD mice. In fact, significantly lower LPS-proliferative responses in spleen cells have been observed in 4-month-old female (p < 0.001; Figure 3a.2) and in thymic cells of female and male (p < 0.001; Figure 3b.2) 3xTg-AD mice than in NTg mice. With ConA, in the case of splenic leukocytes, these lower values of proliferative response were only observed in female 3xTg-Ad mice (p < 0.01; Figure 3a.3), but in thymic leukocytes in both female and male mice (p<0.001; Figure 3b.3). Interestingly, regarding gender differences, male NTg mice showed higher (p<0.01) and lower (p<0.05) LPS-proliferation percentage values in splenic and thymic leukocytes, respectively, than those observed in female animals. In the case of ConA-proliferative percentage, male NTg showed these lower (p<0.001) values in thymic cells. In male 3xTg-AD mice, LPS- and ConA-proliferative percentages of spleen leukocytes were higher (p<0.001) than in female, but in thymic cells these 3xTg-AD males showed lower (p<0.001) values in ConA-proliferation than females.
Cytokines are major mediators of the complex interactions among immune cells, being responsible for the development and resolution of the immune response. For this reason, we analyzed the concentrations of several cytokines, including IL-2, IL-10, IL-1β, and TNF-α, secreted ex vivo by splenic leukocytes cultured for 48 hours under conditions stimulated by Con A and LPS. In general, the results of our study showed that the release of these cytokines suffers impairments in 3xTg-AD mice, especially in females. Thus, as shown in Figure 4 and Table 4, under Con A- and LPS-stimulated conditions, lower concentrations of IL-2 and IL-10 were observed in female (Con A: p < 0.001 and p < 0.05, respectively; LPS: p < 0.01 for both) and male (ConA: p<0.05 and p < 0.001, respectively; LPS: p<0.001 in IL-10) 3xTg-AD mice in comparison to gender-matched NTg mice. Furthermore, under Con A-stimulated conditions, this was also accompanied by higher (p<0.001) release of IL-1β and TNF-α only in females 3xTg-AD (Figure 4c-4d). However, no differences were observed in these cytokines under stimulation with LPS (Table 4). Regarding gender differences, male NTg mice showed higher IL-1β, TNF-α and IL-10 concentrations (p<0.01, p<0.05 and p < 0.001, respectively) in ConA presence, and higher IL-10 (p<0.001) with LPS stimulation. However, male 3xTg-AD mice showed lower IL-1β and TNF-α concentrations under ConA-stimulated conditions (p < 0.001 and p < 0.01, respectively; Figure 4c-4d) and higher IL-2 and IL-10 concentrations with LPS (p<0.001. Table 4) than those observed in female 3xTg-AD mice.
The IL-10/TNF-α ratios under Con A-stimulated conditions (Figure 4e), showed in female and male 3xTg-AD mice lower values (p < 0.01 and p < 0.001, respectively), compared to their corresponding gender-matched NTg. Similarly, with LPS stimulation (Table 4) both female and male 3xTg-AD mice showed lower values (p<0.001) than those in the corresponding NTg mice. Moreover, under Con-A and LPS stimulation, both male NTg (p < 0.001) and 3xTg-AD (p < 0.05 with ConA and p<0.001 with LPS) mice presented a higher IL-10/TNF-α ratios than females.
3.4. An oxidative stress is present in the spleen of 4-month-old female and male 3xTg-AD mice
As shown in Figure 5 and Figure 3xTg-AD mice exhibited higher values of oxidative stress markers at 4 months of age, particularly in male mice, compared to NTg animals. In fact, the values of the antioxidant enzymes such as GPx and GR activities (Figure 5a-b), as well as of the GSH concentrations (Figure 5c), were lower in 4-month-old female (GPx: p < 0.01; GR: p < 0.05; GSH: p < 0.001) and male (GPx: p<0.01; GR and GSH: p < 0.001) 3xTg-AD mice than in NTg mice. Moreover, higher values of the pro-oxidant XO activity were found in female (p < 0.01) and male (p < 0.001) 3xTg-AD mice in comparison to NTg animals (Figure 5d). Finally, although the oxidative stress was more marked in male NTg mice, with lower values of GPx activity (p<0.05) and GSH concentration (p<0.001) than in female animals, in 3xAD-Tg, no statistical differences were observed in relation to sex
4. Discussion
According to our knowledge, this study is the first in which an integrative analysis of male and female 3xTg-AD mice at 4 months of age, when only intraneuronal immunoreactivity to beta-amyloid can be detected, shows that their prodromal behavioral profile aligns with the first alterations in several components of the neuroimmune network that prove the premature aging state of these animals. Moreover, the present work shows, also for the first time, a time lag in the alterations of the different components of the neuroimmunendocrine system that could suggest that behavioral and immune alterations precede endocrine ones in these transgenic animals.
It is known that one characteristic of the aging process is the deterioration of homeostatic systems (nervous, endocrine, and immune systems) and a crosstalk communication among them [8,39]. Moreover, the parameters analyzed in the present study, which are representatives of the state of these systems, suffer age-related deterioration [8,39] and, in several aspects, this is similar to that observed in 3xTg-AD at 15 months of age [31]. Therefore, we suggest that these animals exhibit premature aging, as they reach adulthood with neuroimmune characteristics typical of older animals, and distinct from those of their sex- and age-matched 4-month-old NTg counterparts. This premature deterioration in behavior and immunity is sex-specific, similarly to what occurs with the neuroimmunoendocrine impairment observed in male and female 15-month-old 3xTg-AD mice [31]. At this old age, which mimics advanced stages of Alzheimer´s disease [17,19] an accelerated deterioration of the neuroimmune system was observed, especially in males, demonstrating the relevance of this system in the etiopathogenesis of AD and its relation with the shorter life span of males than females [31]. Although there are differences in the alterations observed at 15 and at 4-month-old 3xTg-AD mice, the present results show that neuroimmune defects can be also observed at prodromic stages of disease. Moreover, corticosterone, a hormone very representative of the endocrine system state and related to stress response and the neuroimmunoendocrine system [42,56], didn´t present differences in its plasma concentrations in animals at 4 months of age. In contrast, the plasmatic values of corticosterone were higher in males at 15 months of age as compared to controls [31].
In order to study the underlying derangement of the neuroimmunoendocrine system in males and females, we first verified their prodromal status. Thus, we determined the level of behavioral disturbances of 3xTg-AD animals at 4 months of age, as compared with sex- and age-matched NTg mice. This allowed us to provide further evidence on the progressive derangement of the different components of the immune-endocrine function and to do so in both sexes. At the behavioral level, the results of the present study show that, at 4 months of age, when early signs of behavioral and psychological symptoms of dementia (BDSP)-like alterations are observed in the 3xTg-AD mice, as characterized by [26], a higher level of neophobia as measured by longer latency and lower number of rearing in the corner test. This was confirmed by changes in coping with stress strategies in the open-field and T-maze tests. This mouse model has provided evidence for the pathogenic role of intraneuronal Aβ in AD [26], and it has shown how this early event relates to other neuropathological aspects, such as the affectation of the basolateral amygdala and the early appearance of BPSD-like symptoms [27]. These early BPSD-like symptoms are mostly related to emotionality, stress coping strategies, and exploratory behaviors, resembling those observed in patients at the pre-clinical stages of the disease [57 Cerejeira et al 2012] At all pre-pathological stages, disinhibitory behavior and a lack of ability to cope with mild stressors, such as novelty, are common features of the 3xTg-AD behavioral phenotype [58] and worsen with the progression of the disease [18,23]. Thus, these results resemble the stronger behavioral impairment in 3xTg-AD at advanced stages of disease [31] and indicate that there are present already at prodromal stages of disease.
In all these tests, genotype was the predominant factor to explain the consistent delay in ethogram, that is, all the latencies of the behavioral events exhibited by both genders of 3xTg-AD mice (initial freezing, thigmotaxis, vertical exploratory activity, self-grooming behavior) and the consequent reduction in exploratory activity developed during the test as compared to NTg mice. Interestingly, several quantitative measures for exploratory efficiency in the T-maze indicate that they are, in fact, the result of qualitative differences in the behavioral patterns developed. Thus, both genotypes invested the same amount of time to complete the test, but this time corresponded to two different behavioral patterns, as evidenced by the ratio of time spent in the different arms and the number of errors. The poor ability to confront the test (higher latency to reach the intersection) exhibited by 3xTg-AD mice was followed by a short exploratory activity of the horizontal arms. In contrast, in the NTg mice, the ratio indicated a preference for exploring the two horizontal arms, which is the basis of the spontaneous alternation typical animals’ behavior in this maze [59] Therefore, it is essential to note that the lack of differences in total exploration time does not align with the qualitative differences in the behavioral profile, rendering this time variable a non-specific measure and necessitating caution when studied alone. In addition, it is interesting to note that the mean time to reach the intersection in the T-maze is a behavioral paradigm related to worse immunosenescence and reduced survival in prematurely aging mice (PAM) as compared to non-prematurely aging mice (NPAM) [8,43].
The higher number of defecation boli and the presence of urination in 3xTg-AD mice are considered to indicate changes in their emotionality [23]. We have previously reported such changes at 6, 7, 12, and 18 months of age, but not before, during young adulthood (2 months of age), nor at the early onset of the symptoms of the disease (4 months of age) [23,51]. In the present work, we found statistically significant differences in defecation between genotypes and sexes, which were primarily due to a higher number in NTg females. These post-hoc differences may indicate that female NTg mice are more sensitive to exhibiting the higher emotionality of female sex than 3xTg-AD mice at this age [51]. Some specific sex and genotype effects could be observed. Thus, on the one hand, male 3xTg-AD mice were more prone to exhibit BPSD-like alterations, such as neophobia in the corner test and bizarre behaviors in the open-field test. On the other hand, female 3xTg-AD mice were the ones to exhibit the most intense initial freezing behavior in the open-field test and the highest delay to reach the criteria (intersection) at the T-maze, which in both cases are known to be indicative of higher fearfulness and/or poor coping with stress strategies [39]. Other variables, such as the number of errors, also indicate differences between genotypes by sex in this respect. Overall, the behavioral data indicate BDSP-like alterations in 3xTg-AD mice at 4 months of age, during the early stages of the disease, and highlight the relevance of genotype effects underlying these alterations. In all the tests, horizontal and vertical exploratory activities were lower than in controls, although differences only reached statistical significance in the vertical component, which is the one known to be more sensitive to detect changes in exploratory activity [60].
The present work also demonstrates the relevance of BPSD-like symptoms and the peripheral immune system as early indicators of disease onset, before cognitive deficits and alterations in plasma glucocorticoid levels can be detected. As well, no sex differences have been described at the immunohistochemical level for intraneuronal βA and tau immunoreactivity at this age [25,26,27,61] which reinforces the value of the findings of BPSD-like behaviors and peripheral immune system as early indicators of the alterations during the onset of the disease.
The overweight described for 3xTg-AD mice, especially in males [26] was already present at this age, and it could be due to a higher relative weight of white adipose tissue. The organometrics of peripheral immune organs are relevant to understanding physical variation and changes associated with disease [28,31,51]. Thus, in 3xTg-AD mice, we have reported that both total weight and relative weight of peripheral immunoendocrine organs such as thymus, spleen, and adrenal glands correlate with the sex-dependent impairment of neuroimmunoendocrine network described at advanced stages of the disease, and even at 6 months of age [28,31,51]. The present work indicates that these changes can already be observed at early stages of the disease; however, immunological organs are more sensitive to exhibiting changes in their organometrics than the endocrine glands.
Regarding immune system functions analyzed in leukocytes from the spleen and thymus, including functions of innate immunity such as chemotaxis and antitumoral natural killer activity, and of adaptive immunity (lymphoproliferation in response to Con A and LPS mitogens), the results provide evidence that there is an impairment in 3xTg-AD mice at early stages of the disease (4 months of age) in males and females. Thus, both sexes of 3xTg-AD mice exhibited a lower chemotactic index in spleen and thymus leukocytes compared to NTg mice, with a more pronounced effect in males. The natural killer activity against tumor cells was lower in leukocytes from 3xTg-AD mice than in NTg animals, but these differences were only detectable in the spleen, and males were, again, more affected. It is known that these two functions, representatives of innate immunity, exhibit age-related changes. In general, these changes are shown as lower values in these functions, which represent worse innate defense mechanisms [8,14,46,62] in 3xTg-AD mice than in NTg animals.
A lower lymphoproliferative response to the mitogens Con A- and LPS, a typical adaptive immune function, was shown in 3xTg-AD mice in comparison with NTg mice, in the spleen (only in females) and in the thymus (in both sexes). This lower proliferation, a characteristic of immunosenescence [8,63], in spleen leukocytes from females could be due, at least in part, to the lower IL-2 release capacity of these cells, a cytokine involved in this proliferative response [64].
These functions altered in leukocytes from immune organs were also affected in peritoneal immune cells of female 3xTg-AD mice at the age of 4 months compared to NTg controls [29,30]. However, whereas the innate functions of chemotaxis and NK activity were also lower in female 3xTgAD than in NTg mice, similarly to that in leukocytes from immune organs, especially in the spleen, the proliferation of lymphocytes was higher in peritoneal leukocytes from transgenic animals than that of the female controls. These results show that in these transgenic mice, the differences with controls in the immune functions are different depending on the leukocyte location, at least in adaptive immunity. Nevertheless, in a model of prematurely aging mice (PAM), we have observed that all these immune functions in peritoneum of females showed lower values than in the corresponding non-prematurely aging mice (NPAM) controls, even in young adult animals of 4 months of age, and these PAM show lower lifespan than NPAM [8,39]. Moreover, these premature alterations detected in the peritoneum were also observed in immune organs, such as the spleen and thymus, of these PAM [43].
Regarding cytokines, the pro- and anti-inflammatory cytokines released by spleen leukocytes showed differences depending on sex and the mitogen used. Thus, with ConA, a mitogen more specific to T lymphocytes [30], in female 3xTg-AD, the concentrations of proinflammatory cytokines, such as IL-1β and TNF-α, were higher than in NTg. However, in males, the concentrations of IL-1β released by cells from 3xTg-AD were lower than those in NTg, whereas no differences were found in TNF-α. Nevertheless, since the amounts of the anti-inflammatory cytokine, IL-10, were lower in 3xTg-AD mice of both sexes, the ratio IL-10/ TNF-α, a good indicator of an appropriate balance anti-inflammatory/pro-inflammatory and successful aging and longevity [30, 65 Poulios 2025], were also lower in both sexes of 3xTg-AD than in the corresponding NTg animals. Similarly, with LPS, the lower concentrations of IL-10 in 3xTg-AD mice resulted in lower ratios of IL-10/TNF-α being observed in both males and females, as no differences in proinflammatory cytokines were noted. This loss of anti-inflammatory control, which could explain the incorrect immune response and the possible arrival of pro-inflammatory compounds in the brain, generating the neuroinflammation characteristic of these pathologies [7 Malarte], appears to occur at an early age in these 3xTg-AD mice. Moreover, an inflammatory stress was also observed, with a higher secretion of proinflammatory cytokines, such as IL-1β, and the ratios IL-1/IL-10 and IL-6/IL-10, in peritoneal leukocytes from 4-month-old 3xTg-AD females [30]. This peripheral inflammation has been linked to AD initiation and progression in preclinical and clinical studies [7, 40Xie et al., 2022]
In addition, it is known that the presence of both oxidative and inflammatory stress, especially in immune cells, underlies aging and neurodegenerative processes [8,14,29]. In this sense, we have also reported, in a previous study, premature oxidative stress in peritoneal immune cells from 4-month-old 3xTg-AD females. This stress was characterized by lower antioxidants and higher oxidants than their age- and sex-matched NTg counterparts, and higher lipid oxidative damage [29,30]. In the present work, the spleens of 3xTg-AD mice exhibited an oxidative stress situation compared to the corresponding NTg mice. Thus, antioxidant defenses such as the activity of the antioxidant enzymes of the glutathione cycle GPx and GR and the concentrations of the endogenous antioxidant glutathione reduced (GSH), which is related to the immune function [8], were lower in male and female 3xTg-AD mice than in those NTg counterparts. However, the activity of XO, a pro-oxidant enzyme, was higher in both sexes of those animals. In previous studies, carried out on peritoneal leukocytes from female 3xTg-AD mice at 4 months of age, lower concentrations of GSH [29] and higher activity of XO were also observed [30]. Again, male 3xTg-AD mice are most affected. In fact, although, in general, male, in comparison with female mice, show worst immune response and higher oxidative stress, which explains their lower lifespan [46], these sex differences are strain and age-dependent, being more evident in adult animals [46]. Thus, in spleen and thymus lymphocytes of male and female old 3xTg-AD mice no differences were found in GSH concentrations comparing with the corresponding values in NTg animals, although males shown lower concentrations of this antioxidant than females, at less in spleen [31].
Therefore, the results indicate that at this early stage of the disease, oxidative and inflammatory stresses are present. This fact, together with the premature immunosenescence, which is associated with oxidative-inflammatory stress [8,39], and also with the premature aging of several behavioral responses, confirms the premature aging of these 3xTg-AD mice and explains their shorter life span [30,31]. The present study confirms the importance of neuroimmune communication and the peripheral immunity in the AD pathogenesis [13]. In addition, since the results obtained in 3xTg-AD mice seem to be similar to these in human AD patients [14,32], our better knowledge on the interaction between the peripheral immune system and CNS as well as the state of each one of these homeostatic systems through evaluation of immune functions and of behavioral texts in these transgenic mice, could encourage the development of new therapeutic strategies. In fact, a treatment with an anti-TNFSF10 monoclonal antibody in 3xTg-AD mice restrain overshooting central and peripheral (spleen) inflammation by rebalancing the immune response, mitigating the progression of AD pathology in these animals [35]. Although the use of transgenic mice to research translational strategies in AD has been shown to be limited by several authors [66,67], 3xTg-AD mice have been proven useful, in this context, for many others [68,69,70,71].
5. Conclusions
In the 3xTg-AD mice, the impairment of the neuroendocrine-immune network becomes apparent at the prodromal stage of AD (4 months old), with sex differences. We also demonstrate that the different regulatory systems involved in this network exhibit distinct sensitivities in responding to these changes. This study demonstrates the relevance of BPSD-like symptoms, peripheral immune cell functions, and redox state as early indicators of disease onset in this animal model, before cognitive deficits and statistically significant changes in plasma corticosterone can be detected. Thus, we propose these biomarkers, identified at prodromal stages in the 3xTg-AD mice, as tools of interest at both a translational and clinical level, as they could be useful for risk assessment, early diagnosis, evaluation of disease progression, and assessment of treatment effectiveness in clinical trials. In addition, the alterations in behavior and immunity, as well as the oxidative-inflammatory stress observed in 3xTg-AD mice at this young age, confirm the premature aging of these animals, explaining their lower longevity.
Author Contributions
Conceptualization, L.G-L. and M.D.L.F.; methodology, L.G.-L, C.V. and R.M.; software, L.G.-L, C.V. and J.F.; formal analysis, L.G.-L., C.V., S..Q. and J.F.; investigation, L.G.-L, C.V. and R.M.; data curation, L.G., C.V.; writing—original draft preparation, L.G-L. and M.D.L.F; writing—review and editing, L.G.-L, M.D.L.F., J.F.; supervision, L.G-L. and M.D.L.F; project administration, L.G-L. and M.D.L.F.; funding acquisition, L-G-L. and M.D.L.F. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by FIS ISC3 PI10/00283, RETICEF (Red de Envejecimiento y Fragilidad) ISC3 RD06/0013/0003, Research Group of Translational Behavioral Neuroscience UAB-GE-260408, and the Research Group of UCM-CM (ENEROINN 910379).
Institutional Review Board Statement
All the experiments were conducted in accordance with the Spanish legislation on “Protection of Animals Used for Experimental and Other Scientific Purposes” and the EU Directive (2010/63/EU) on this subject, and the Ethical Commission of the Universitat Autònoma de Barcelona (Protocol Number CEEAH/00759 3588/DMAH 9452).
Data Availability Statement
Data will be available upon request to Lydia Giménez-Llort (lidia.gimenez@uab.cat).
Acknowledgments
We would like to thank Prof. Frank M. LaFerla (Department of Neurobiology and Behavior, University of California Irvine, CA, USA) for providing us with the progenitor animals.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Knopman, D.S.; Amieva, H.; Petersen, R.C.; Chételat, G.; Holtzman, D.M.; Hyman, B.T.; Nixon, R.A.; Jones, D.T. Alzheimer Disease. Nat. Rev. Dis. Primers 2021, 7, 33. [Google Scholar] [CrossRef]
- Feigin, V.L.; Vos, T.; Nichols, E.; Owolabi, M.O.; Carroll, W.M.; Dichgans, M.; Deuschl, G.; Parmar, P.; Brainin, M.; Murray, C. The Global Burden of Neurological Disorders: Translating Evidence into Policy. Lancet Neurol. 2020, 19, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.; Wahlund, L.O.; Westman, E. The Heterogeneity within Alzheimer’s Disease. Aging 2018, 10, 3058. [Google Scholar] [CrossRef] [PubMed]
- Livingston, G.; Huntley, J.; Liu, K. Y.; Costafreda, S. G.; Selbæk, G.; Alladi, S.; Ames, D.; Banerjee, S.; Burns, A.; Brayne, C.; Fox, N. C.; Ferri, C. P.; Gitlin, L. N.; Howard, R.; Kales, H. C.; Kivimäki, M.; Larson, E. B.; Nakasujja, N.; Rockwood, K.; Samus, Q.; Shirai, K.; Singh-Manoux, A.; Schneider, L.S.; Walsh, S.; Yao, Y.; Sommerlad, A.; Mukadam, N. Dementia prevention, intervention, and care: 2024 report of the Lancet standing Commission. Lancet 2024, 404, 572–628. [Google Scholar] [CrossRef] [PubMed]
- Lomi, M.; Geraci, F.; Del Seppia, C.; Dolciotti, C.; Del Carratore, R.; Bongioanni, P. Biomarker profile in peripheral blood cells related to Alzheimer´s disease. Mol Neurobiol. 2025, 8949–8964. [Google Scholar] [CrossRef]
- Yasumo, F.; Watanabe, A.; Kimura, Y.; Yamauchi, Y.; Ogata, A.; Ikenuma, H.; Abe, J.; Minami, H.; Nihashi, T.; Yokoi, K.; Shimoda, N.; Kasuga, K.; Ikeuchi, T.; Takeda, A.; Sakurai, T.; Ito, K.; Kato, T. Plasma IL-6 levels as a biomarker for behavioral changes in Alzheimer´s disease. Neuroimmunomodulation 2025, 1–21. [Google Scholar] [CrossRef]
- Malarte, M.L.; Chiotis, K.; Ioannou, K.; Rodriguez-Vieitez, E. Peripheral inflammation is associated with greater neuronal injury and lower episodic memory among late middle-aged adults. J Neurochem 2025, 169, e70222. [Google Scholar] [CrossRef]
- De la Fuente, M.; Miquel, J. An update of the oxidation-inflammation theory of aging: the involvement of the immune system in oxi-inflamm-aging. Curr Pharm Des 2009, 15, 3003–26. [Google Scholar] [CrossRef]
- McManus, R. M. The Role of Immunity in Alzheimer's Disease. Adv Biol 2022, 6, e2101166. [Google Scholar] [CrossRef]
- Long, C.; Fritts, A.; Broadway, J.; Brawman-Mintzer, O.; Mintzer, J. Neuroinflammation: A Driving Force in the Onset and Progression of Alzheimer's Disease. J Clin Med. 2025, 14, 331. [Google Scholar] [CrossRef]
- Heneka, M. T.; van der Flier, W. M.; Jessen, F.; Hoozemanns, J.; Thal, D. R.; Boche, D.; Brosseron, F.; Teunissen, C.; Zetterberg, H.; Jacobs, A. H.; et al. Neuroinflammation in Alzheimer disease. Nat Rev Immunol 2025, 25, 321–352. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Chen, S.; Zhu, J.; Ma, J.; Zhai, Y. The Emerging Role of Central and Peripheral Immune Systems in Neurodegenerative Diseases. Front Aging Neurosci 2022, 14, 872134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gao, Y.; Zhao, Y.; Huang, T. Y.; Zheng, Q.; Wang, X. Peripheral and central neuroimmune mechanisms in Alzheimer's disease pathogenesis. Mol Neurodegener 2025, 20, 22. [Google Scholar] [CrossRef]
- Vida, C.; Martinez de Toda, I.; Garrido, A.; Carro, E.; Molina, J. A.; De la Fuente, M. Impairment of Several Immune Functions and Redox State in Blood Cells of Alzheimer's Disease Patients. Relevant Role of Neutrophils in Oxidative Stress. Front Immunol 2018, 8, 1974. [Google Scholar] [CrossRef]
- Puzzo, D.; Gulisano, W.; Palmeri, A.; Arancio, O. Rodent Models for Alzheimer’s Disease Drug Discovery. Expert Opin. Drug Discov. 2015, 10, 703–711. [Google Scholar] [CrossRef]
- Sanchez-Varo, R.; Mejias-Ortega, M.; Fernandez-Valenzuela, J. J.; Nuñez-Diaz, C.; Caceres-Palomo, L.; Vegas-Gomez, L.; Sanchez-Mejias, E.; Trujillo-Estrada, L.; Garcia-Leon, J. A.; Moreno-Gonzalez, I.; Vizuete, M.; Vitorica, J.; Baglietto-Vargas, D.; Gutierrez, A. Transgenic Mouse Models of Alzheimer's Disease: An Integrative Analysis. Int J Mol Sci 2022, 23, 5404. [Google Scholar] [CrossRef]
- Oddo, S.; Caccamo, A.; Shepherd, J. D.; Murphy, M. P.; Golde, T. E.; Kayed, R.; Metherate, R.; Mattson, M. P.; Akbari, Y.; LaFerla, F. M. Triple-transgenic model of Alzheimer's disease with plaques and tangles: intracellular Abeta and synaptic dysfunction. Neuron 2003, 39, 409–21. [Google Scholar] [CrossRef]
- Oddo, S.; Caccamo, A.; Kitazawa, M.; Tseng, B. P.; LaFerla, F. M. Amyloid deposition precedes tangle formation in a triple transgenic model of Alzheimer's disease. Neurobiol Aging 2003, 24, 1063–70. [Google Scholar] [CrossRef]
- Oddo, S.; Caccamo, A.; Green, K. N.; Liang, K.; Tran, L.; Chen, Y.; Leslie, F. M.; LaFerla, F. M. Chronic nicotine administration exacerbates tau pathology in a transgenic model of Alzheimer's disease. Proc Natl Acad Sci U S A 2005, 102, 3046–51. [Google Scholar] [CrossRef]
- Oddo, S.; Caccamo, A.; Tran, L.; Lambert, M. P.; Glabe, C. G.; Klein, W. L.; LaFerla, F. M. Temporal profile of amyloid-beta (Abeta) oligomerization in an in vivo model of Alzheimer's disease. A link between Abeta and tau pathology. J Biol Chem 2006, 281, 1599–604. [Google Scholar] [CrossRef] [PubMed]
- Billings, L. M.; Oddo, S.; Green, K. N.; McGaugh, J. L.; LaFerla, F. M. Intraneuronal Abeta causes the onset of early Alzheimer's disease-related cognitive deficits in transgenic mice. Neuron 2005, 45, 675–88. [Google Scholar] [CrossRef]
- Janelsins, M. C.; Mastrangelo, M. A.; Oddo, S.; LaFerla, F. M.; Federoff, H. J.; Bowers, W. J. Early correlation of microglial activation with enhanced tumor necrosis factor-alpha and monocyte chemoattractant protein-1 expression specifically within the entorhinal cortex of triple transgenic Alzheimer's disease mice. J Neuroinflammation 2005, 2, 23. [Google Scholar] [CrossRef]
- Kitazawa, M.; Oddo, S.; Yamasaki, T. R.; Green, K. N.; LaFerla, F. M. Lipopolysaccharide-induced inflammation exacerbates tau pathology by a cyclin-dependent kinase 5-mediated pathway in a transgenic model of Alzheimer's disease. J Neurosci 2005, 25, 8843–53. [Google Scholar] [CrossRef]
- Sterniczuk, R.; Antle, M. C.; Laferla, F. M.; Dyck, R. H. Characterization of the 3xTg-AD mouse model of Alzheimer's disease: part 2. Behavioral and cognitive changes. Brain Res 2010, 1348, 149–55. [Google Scholar] [CrossRef]
- Belfiore, R.; Rodin, A.; Ferreira, E.; Velazquez, R.; Branca, C.; Caccamo, A.; Oddo, S. Temporal and regional progression of Alzheimer's disease-like pathology in 3xTg-AD mice. Aging Cell 2019, 18, e12873. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Blázquez, G.; Cañete, T.; Johansson, B.; Oddo, S.; Tobeña, A.; LaFerla, F. M.; Fernández-Teruel, A. Modeling behavioral and neuronal symptoms of Alzheimer's disease in mice: a role for intraneuronal amyloid. Neurosci Biobehav Rev 2007, 31, 125–47. [Google Scholar] [CrossRef]
- Muntsant, A.; Castillo-Ruiz, M. D. M.; Giménez-Llort, L. Survival Bias, Non-Lineal Behavioral and Cortico-Limbic Neuropathological Signatures in 3xTg-AD Mice for Alzheimer's Disease from Premorbid to Advanced Stages and Compared to Normal Aging. Int J Mol Sci 2023, 24, 13796. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Torres-Lista, V.; De la Fuente, M. Crosstalk between behavior and immune system during the prodromal stages of Alzheimer's disease. Curr Pharm Des 2014, 20, 4723–32. [Google Scholar] [CrossRef] [PubMed]
- Maté, I.; Cruces, J.; Giménez-Llort, L.; De la Fuente, M. Function and redox state of peritoneal leukocytes as preclinical and prodromic markers in a longitudinal study of triple-transgenic mice for Alzheimer's disease. J Alzheimers Dis 2015, 43, 213–26. [Google Scholar] [CrossRef] [PubMed]
- Ceprián, N.; Martínez de Toda, I.; Maté, I.; Garrido, A.; Gimenez-Llort, L.; De la Fuente, M. Prodromic Inflammatory-Oxidative Stress in Peritoneal Leukocytes of Triple-Transgenic Mice for Alzheimer's Disease. Int J Mol Sci 2024, 25, 6976. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Arranz, L.; Maté, I.; De la Fuente, M. Gender-specific neuroimmunoendocrine aging in a triple-transgenic 3xTg-AD mouse model for Alzheimer's disease and its relation with longevity. Neuroimmunomodulation 2008, 15, 331–43. [Google Scholar] [CrossRef] [PubMed]
- St-Amour, I.; Bosoi, C. R.; Paré, I.; Ignatius Arokia Doss, P. M.; Rangachari, M.; Hébert, S. S.; Bazin, R.; Calon, F. Peripheral adaptive immunity of the triple transgenic mouse model of Alzheimer's disease. J Neuroinflammation 2019, 16, 3. [Google Scholar] [CrossRef]
- Yang, S. H.; Kim, J.; Lee, M. J.; Kim, Y. Abnormalities of plasma cytokines and spleen in senile APP/PS1/Tau transgenic mouse model. Sci Rep 2015, 5, 15703. [Google Scholar] [CrossRef]
- Fraile-Ramos, J.; Reig-Vilallonga, J.; Giménez-Llort, L. Glomerular Hypertrophy and Splenic Red Pulp Degeneration Concurrent with Oxidative Stress in 3xTg-AD Mice Model for Alzheimer's Disease and Its Exacerbation with Sex and Social Isolation. Int J Mol Sci 2024, 25, 6112. [Google Scholar] [CrossRef]
- Cantone, A.F.; Burgaletto, C.; Di Benedetto, G.; Gaudio, G.; Giallongo, C.; Caltabiano, R.; Broggi, G.; Bellanca, C. M.; Cantarella, G.; Bernardini, R. Rebalancing immune interactions within the brain-spleen axis mitigates neuroinflammation in an aging mouse model of Alzheimer´s disease. J Neuroimmune Pharmacol 2025, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Guo, J.; Ye, X. Y.; Xie, Y.; Xie, T. Oxidative stress: The core pathogenesis and mechanism of Alzheimer's disease. Ageing Res Rev 2022, 77, 101619. [Google Scholar] [CrossRef]
- Torres-Lista, V.; Parrado-Fernández, C.; Alvarez-Montón, I.; Frontiñán-Rubio, J.; Durán-Prado, M.; Peinado, J.R.; Johansson, B.; Alcaín, F.J.; Giménez-Llort, L. Neophobia, NQO1 and SIRT1 as premorbid and prodromal indicators of AD in 3xTg-AD mice. Behav Brain Res. 2014, 271, 140–6. [Google Scholar] [CrossRef]
- Shen, L.; Chen, Y.; Yang, A.; Chen, C.; Liao, L.; Li, S.; Ying, M.; Tian, J.; Liu, Q.; Ni, J. Redox Proteomic Profiling of Specifically Carbonylated Proteins in the Serum of Triple Transgenic Alzheimer's Disease Mice. Int J Mol Sci 2016, 17, 469. [Google Scholar] [CrossRef]
- De la Fuente, M. Oxidation and inflammation in the immune and nervous systems, a link between aging and anxiety. In Handbook of immunosenescence; Springer, 2019; pp. pp 1–31. [Google Scholar]
- Xie, J.; Van Hoecke, L.; Vandenbroucke, R.E. The Impact of Systemic Inflammation on Alzheimer's Disease Pathology. Front Immunol. 2022, 12, 796867. [Google Scholar] [CrossRef]
- Montacute, R.; Foley, K.; Forman, R.; Else, K. J.; Cruickshank, S. M.; Allan, S. M. Enhanced susceptibility of triple transgenic Alzheimer's disease (3xTg-AD) mice to acute infection. J Neuroinflammation 2017, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Munstant, A.; Giménez-Llort, L. Crosstalk of Alzheimer's disease-phenotype, HPA axis, splenic oxidative stress and frailty in late-stages of dementia, with special concerns on the effects of social isolation: A translational neuroscience approach. Front Aging Neurosci. 2022, 14, 969381. [Google Scholar] [CrossRef]
- Garrido, A.; Cruces, J.; Ceprián, N.; Corpas, I.; Tresguerres, J. A.; De la Fuente, M. Social environment improves immune function and redox state in several organs from prematurely aging female mice and increases their lifespan. Biogerontology 2019, 20, 49–69. [Google Scholar] [CrossRef]
- Barnes, L.L.; Wilson, R.S.; Bienias, J.L.; Schneider, J.A.; Evans, D.A.; Bennett, D.A. Sex differences in the clinical manifestations of Alzheimer disease pathology. Arch Gen Psychiatry 2005, 62, 685–91. [Google Scholar] [CrossRef]
- Ferretti, M.; Iulita, M.F.; Cavedo, E.; Chiesa, P. A.; Schumacher Dimech, A.; Santuccione Chadha, A.; Baracchi, F.; Girouard, H.; Misoch, S.; Giacobini, E.; Depypere, H.; Hampel, H. & Women’s Brain Project and the Alzheimer Precision Medicine Initiative. Sex differences in Alzheimer disease - the gateway to precision medicine. Nature reviews. Neurology 2018, 14, 457–469. [Google Scholar] [CrossRef]
- Suarez, L. M.; Diaz-Del Cerro, E.; Felix, J.; Gonzalez-Sanchez, M.; Ceprian, N.; Guerra-Perez, N.; Novelle, M. G.; Martinez de Toda, I.; De la Fuente, M. Sex differences in neuroimmunoendocrine communication. Involvement on longevity. Mech Ageing Dev 2023, 211, 111798. [Google Scholar] [CrossRef]
- Yang, J. T.; Wang, Z. J.; Cai, H. Y.; Yuan, L.; Hu, M. M.; Wu, M. N.; Qi, J. S. Sex Differences in Neuropathology and Cognitive Behavior in APP/PS1/tau Triple-Transgenic Mouse Model of Alzheimer's Disease. Neurosci Bull 2018, 34, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, M.; Mian, M.F.; Michalski, B.; Azam, A.B.; Ma, D.; Salwierz, P.; Christopher, A.; Rosa, E.; Zovkic, I.B.; Forsythe, P. Sex-dependent differences in spontaneous autoimmunity in adult 3xTg-AD mice. J Alzheimer’s Dis. 2018, 63, 1191–205. [Google Scholar] [CrossRef] [PubMed]
- Kane, A.E.; Shin, S.; Wong, A.A.; Fertan, E.; Faustova, N.S.; Howlett, S.E.; Brown, R.E. Sex differences in healthspan predict lifespan in the 3xTg-AD mouse model of Alzheimer’s disease. Front Aging Neurosci 2018, 10, 172. [Google Scholar] [CrossRef]
- Dennison, J. L.; Ricciardi, N. R.; Lohse, I.; Volmar, C. H.; Wahlestedt, C. Sexual Dimorphism in the 3xTg-AD Mouse Model and Its Impact on Pre-Clinical Research. J Alzheimers Dis 2021, 80, 41–52. [Google Scholar] [CrossRef]
- Baeta-Corral, R.; De la Fuente, M.; Giménez-Llort, L. Sex-dependent worsening of NMDA-induced responses, anxiety, hypercortisolemia, and organometry of early peripheral immunoendocrine impairment in adult 3xTg-AD mice and their long-lasting ontogenic modulation by neonatal handling. Behav Brain Res 2023, 438, 114189. [Google Scholar] [CrossRef]
- Barber, A. J.; Del Genio, C. L.; Swain, A. B.; Pizzi, E. M.; Watson, S. C.; Tapiavala, V. N.; Zanazzi, G. J.; Gaur, A. B. Age, sex and Alzheimer's disease: a longitudinal study of 3xTg-AD mice reveals sex-specific disease trajectories and inflammatory responses mirrored in postmortem brains from Alzheimer's patients. Alzh Res Ther. 2024, 16, 134. [Google Scholar] [CrossRef]
- Meziane, H.; Ouagazzal, A. M.; Aubert, L.; Wietrzych, M.; Krezel, W. Estrous cycle effects on behavior of C57BL/6J and BALB/cByJ female mice: implications for phenotyping strategies. Genes Brain Behav 2007, 6, 192–200. [Google Scholar] [CrossRef]
- Lawrence, R. A.; Burk, R. F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem Biophys Res Commun 1976, 71, 952–8. [Google Scholar] [CrossRef]
- Massey, V.; Williams, C. H., Jr. On the reaction mechanism of yeast glutathione reductase. J Biol Chem 1965, 240, 4470–80. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, B.; Ao, H. Corticosterone effects induced by stress and immunity and inflmmation: mechanisms of communication. Front Endocrinol. 2025, 16, 1448750. [Google Scholar] [CrossRef]
- Cerejeira, J; Lagarto, L; Mukaetova-Ladinska, EB. Behavioral and psychological symptoms of dementia. Front Neurol. 2012, 3, 73. [Google Scholar] [CrossRef]
- Benaroya-Milshtein, N.; Hollander, N.; Apter, A.; Kukulansky, T.; Raz, N.; Wilf, A.; Yaniv, I.; Pick, C. G. Environmental enrichment in mice decreases anxiety, attenuates stress responses and enhances natural killer cell activity. Eur J Neurosci 2004, 20, 1341–7. [Google Scholar] [CrossRef]
- Deacon, R.M.J.; Rawlins, J.N.P. T-Maze Alternation in the Rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Ferré, S.; Martínez, E. Effects of the systemic administration of kainic acid and NMDA on exploratory activity 861 in rats. Pharmacol Biochem Behav 1995, 51, 205–10. [Google Scholar] [CrossRef]
- Clinton, L. K.; Billings, L. M.; Green, K. N.; Caccamo, A.; Ngo, J.; Oddo, S.; McGaugh, J. L.; LaFerla, F. M. Age-dependent sexual dimorphism in cognition and stress response in the 3xTg-AD mice. Neurobiol Dis 2007, 28, 76–82. [Google Scholar] [CrossRef]
- Kim, HH; Dixit, V D. Metabolic regulation of immunological aging. Nat Aging 2025, 5, 1425–1440. [Google Scholar] [CrossRef]
- Fukushima, Y; Ueno, R; Minato, N; Hattori, M. Senescence-associates T cells in immunosenescence and disease. Int Immunol. 2025, 37, 143–152. [Google Scholar] [CrossRef]
- Abbas, AK; TRotta, E; Simeonov, D R; Marson, A; Bluestone, JA. Revisiting IL-1: Biology and therapeutic prospects. Sci Immunol 2018, 3, eaat1482. [Google Scholar] [CrossRef] [PubMed]
- Poulios, P; Skampouras, S; Piperi, Ch. Deciphering the role of cytokines in aging: Biomarker potential and effective targeting. Mech Ageing Dev. 2025, 224, 112036. [Google Scholar] [CrossRef] [PubMed]
- Sonsalla, M.M.; Lamming, D.W. Geroprotective interventions in the 3xTg mouse model of Alzheimer’s disease. GeroScience 2023, 45, 1343–1381. [Google Scholar] [CrossRef]
- Lopes, PA; Guil-Guerrero, JL. Beyond Transgenic Mice: Emerging Models and Translational Strategies in Alzheimer's Disease. Int J Mol Sci. 2025, 26, 5541. [Google Scholar] [CrossRef]
- Dubal, D. B.; Broestl, L.; Worden, K. Sex and gonadal hormones in mouse models of Alzheimer's disease: what is relevant to the human condition? Biol Sex Differ 2012, 3, 24. [Google Scholar] [CrossRef] [PubMed]
- Fertan, E.; Rodrigues, G. J.; Wheeler, R. V.; Goguen, D.; Wong, A. A.; James, H.; Stadnyk, A.; Brown, R. E.; Weaver, I. C. G. Cognitive Decline, Cerebral-Spleen Tryptophan Metabolism, Oxidative Stress, Cytokine Production, and Regulation of the Txnip Gene in a Triple Transgenic Mouse Model of Alzheimer's Disease. Am J Pathol. 2019, 189, 1435–1450. [Google Scholar] [CrossRef]
- Ochi, S.; Iga, J. I.; Funahashi, Y.; Yoshino, Y.; Yamazaki, K.; Kumon, H.; Mori, H.; Ozaki, Y.; Mori, T.; Ueno, S. I. Identifying Blood Transcriptome Biomarkers of Alzheimer's Disease Using Transgenic Mice. Mol Neurobiol 2020, 57, 4941–4951. [Google Scholar] [CrossRef]
- Kapadia, M.; Mian, M. F.; Ma, D.; Hutton, C. P.; Azam, A.; Narkaj, K.; Cao, C.; Brown, B.; Michalski, B.; Morgan, D.; Forsythe, P.; Zovkic, I. B.; Fahnestock, M.; Sakic, B. Differential effects of chronic immunosuppression on behavioral, epigenetic, and Alzheimer's disease-associated markers in 3xTg-AD mice. Alzheimer's Res Ther. 2021, 13, 30. [Google Scholar] [CrossRef]
Figure 1.
Prodromal behavioral profile of 4-month-old male (M) and female (F) non-transgenic (NTg) and 3xTg-AD mice. Results are mean ± SEM. Corner test: (a) Latency of vertical activity (rearings); Open-field test: (b) bizarre incidence (%); (c) latency to leave the center; (d) latency of grooming; (e) vertical activity (number of rearings in each minute of the test); (f) total vertical activity (total number of rearings), (g) number of defecation boli. T-maze: (h) Latency to reach the intersection; (i) number of errors; (j) number of defecation boli. Experimental groups: NTg mice: 11 females; 9 males; 3xTg-AD: 8 females; 10 males. Statistics: * p < 0.05, ** p < 0.01 and *** p < 0.01 vs. the corresponding sex-matched NTg group; + p < 0.05, ++ p < 0.01 vs. the corresponding females.
Figure 1.
Prodromal behavioral profile of 4-month-old male (M) and female (F) non-transgenic (NTg) and 3xTg-AD mice. Results are mean ± SEM. Corner test: (a) Latency of vertical activity (rearings); Open-field test: (b) bizarre incidence (%); (c) latency to leave the center; (d) latency of grooming; (e) vertical activity (number of rearings in each minute of the test); (f) total vertical activity (total number of rearings), (g) number of defecation boli. T-maze: (h) Latency to reach the intersection; (i) number of errors; (j) number of defecation boli. Experimental groups: NTg mice: 11 females; 9 males; 3xTg-AD: 8 females; 10 males. Statistics: * p < 0.05, ** p < 0.01 and *** p < 0.01 vs. the corresponding sex-matched NTg group; + p < 0.05, ++ p < 0.01 vs. the corresponding females.
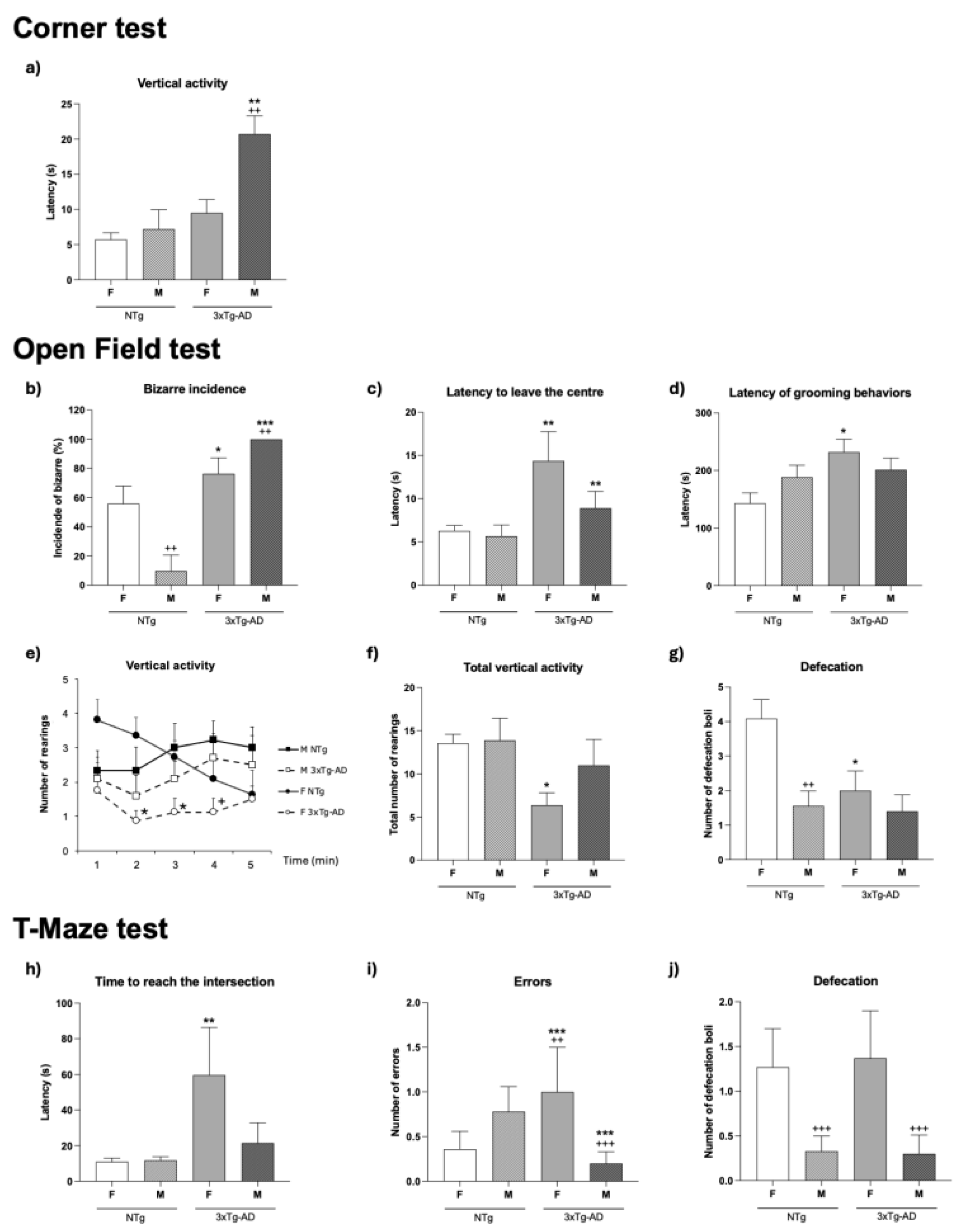
Figure 2.
Chemotactic index of spleen (a) and thymus (b) leukocytes, and Natural killer (NK) cell activity (% lysis of tumor cells) of spleen (c) and thymus (d) leukocytes in 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic for Alzheimer's Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8–11 values corresponding to the same number of subjects (11 NTg female; 9 NTg male; 8 3xTg-AD female; 10 3xTg-AD male). Each value is the mean of duplicate assays. Statistics: * p < 0.05, ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched group; + p < 0.05, ++ p < 0.01, +++ p < 0.001 vs. the corresponding females.
Figure 2.
Chemotactic index of spleen (a) and thymus (b) leukocytes, and Natural killer (NK) cell activity (% lysis of tumor cells) of spleen (c) and thymus (d) leukocytes in 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic for Alzheimer's Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8–11 values corresponding to the same number of subjects (11 NTg female; 9 NTg male; 8 3xTg-AD female; 10 3xTg-AD male). Each value is the mean of duplicate assays. Statistics: * p < 0.05, ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched group; + p < 0.05, ++ p < 0.01, +++ p < 0.001 vs. the corresponding females.
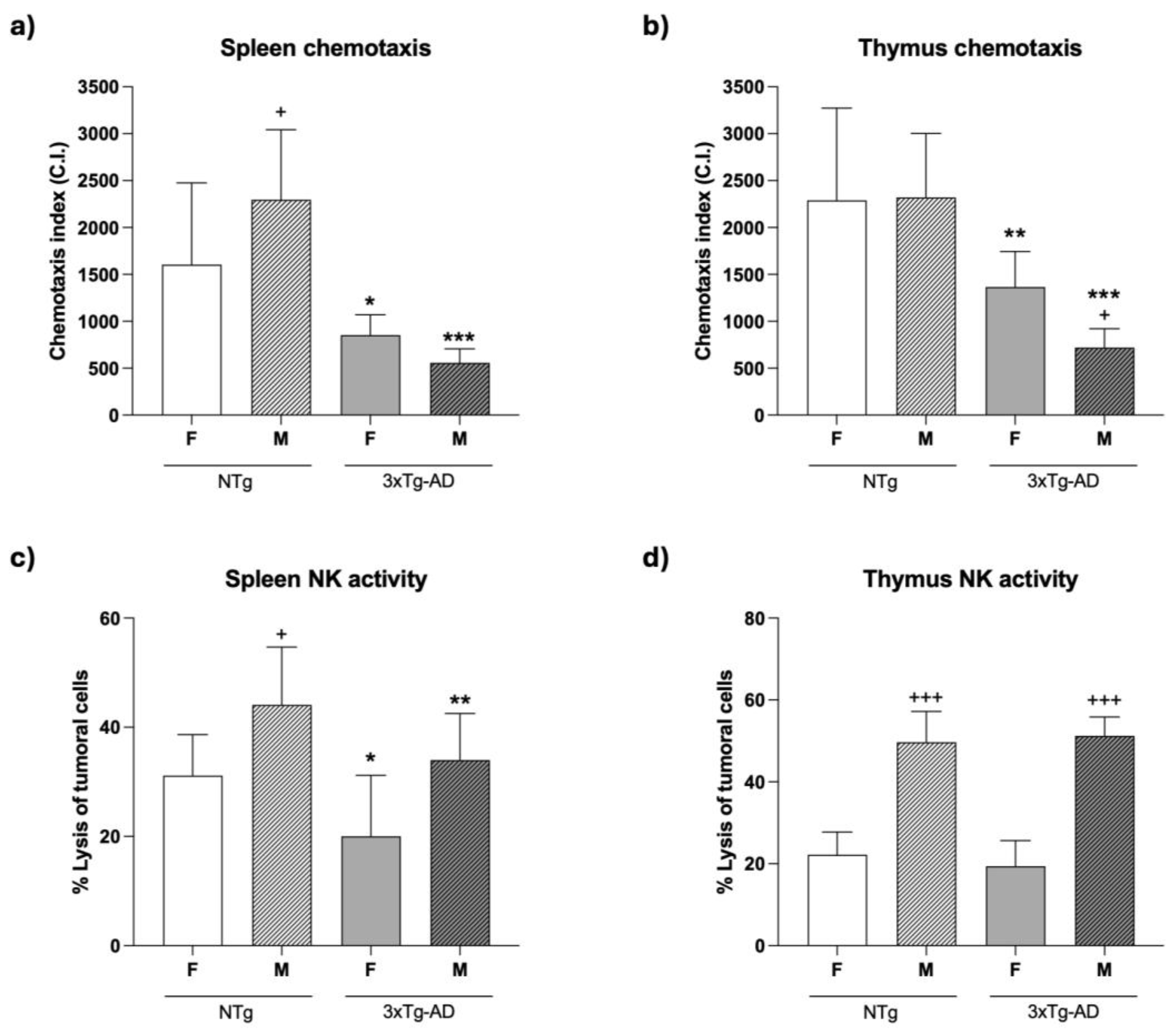
Figure 3.
Proliferation in basal (counts per minute; cpm) and concanavalin A (Con A) and lipopolysaccharide (LPS) stimulated (% stimulation) conditions of spleen (a) and thymus (b) leukocytes, in 4-month-old female (F) and male (M) non-trasngenic (NTg) and triple transgenic for Alzheimer's Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8–11 values corresponding to the same number of subjects (11 NTg females; 9 NTg males; 8 3xTg-AD females; 10 3xTg-AD males). Each value is the mean of triplicate assays. Statistics: ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched group; + p < 0.05, ++ p < 0.01, +++ p < 0.001 vs. the corresponding females.
Figure 3.
Proliferation in basal (counts per minute; cpm) and concanavalin A (Con A) and lipopolysaccharide (LPS) stimulated (% stimulation) conditions of spleen (a) and thymus (b) leukocytes, in 4-month-old female (F) and male (M) non-trasngenic (NTg) and triple transgenic for Alzheimer's Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8–11 values corresponding to the same number of subjects (11 NTg females; 9 NTg males; 8 3xTg-AD females; 10 3xTg-AD males). Each value is the mean of triplicate assays. Statistics: ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched group; + p < 0.05, ++ p < 0.01, +++ p < 0.001 vs. the corresponding females.
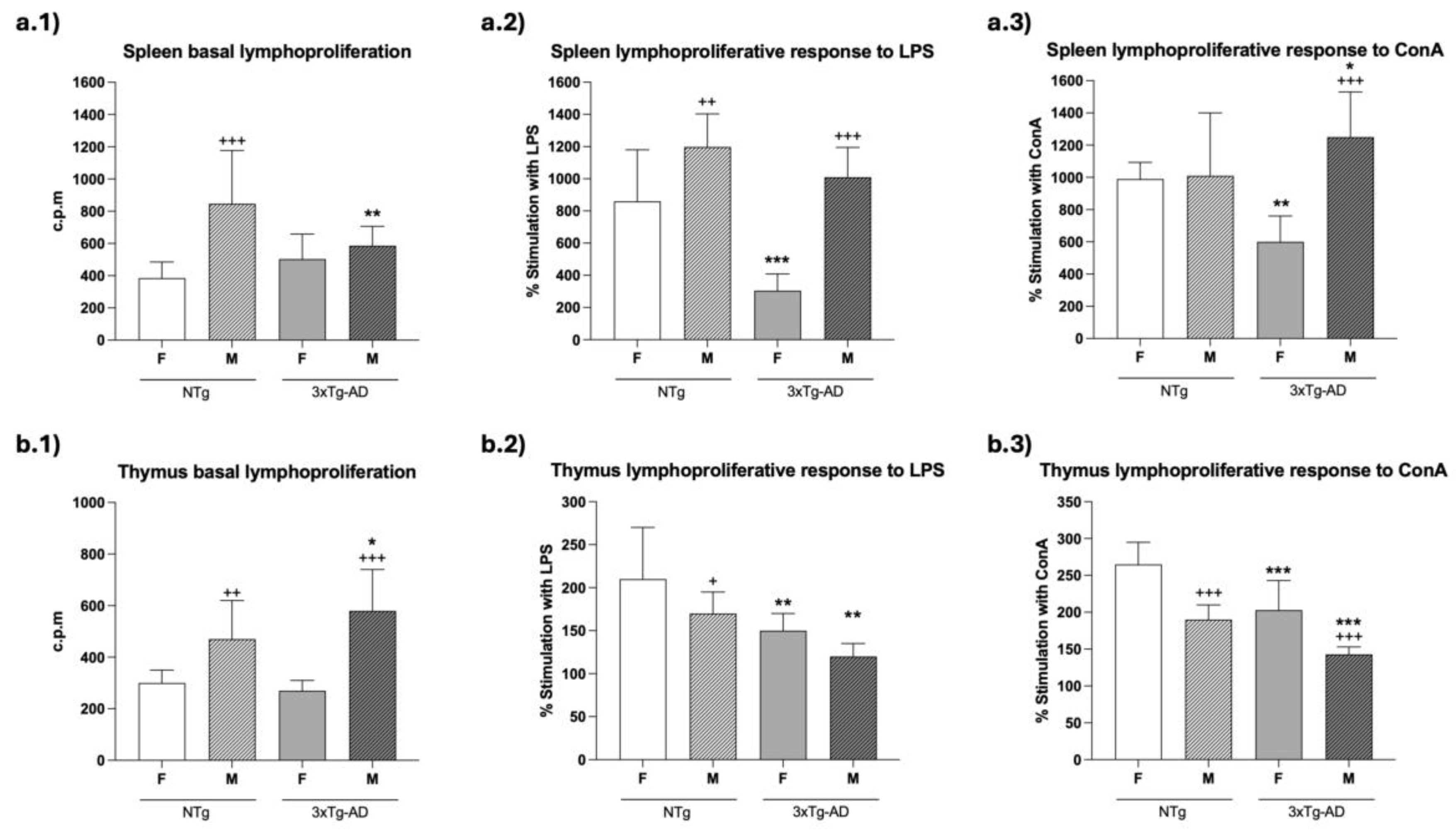
Figure 4.
Concentrations of IL-2 (a), IL-10 (b), IL-1β (c) and TNF-α (d) in supernatants of cultures of spleen leukocytes in response to concanavaline A (Con A, 1 µg/mL) in 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic for Alzheimer Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8 values corresponding to the same number of subjects in each group of mice. Each value is the mean of duplicate assays. Statistics: * p < 0.05, ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched group; + p < 0.05, ++ p < 0.01, +++ p < 0.001 vs. the corresponding females.
Figure 4.
Concentrations of IL-2 (a), IL-10 (b), IL-1β (c) and TNF-α (d) in supernatants of cultures of spleen leukocytes in response to concanavaline A (Con A, 1 µg/mL) in 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic for Alzheimer Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8 values corresponding to the same number of subjects in each group of mice. Each value is the mean of duplicate assays. Statistics: * p < 0.05, ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched group; + p < 0.05, ++ p < 0.01, +++ p < 0.001 vs. the corresponding females.
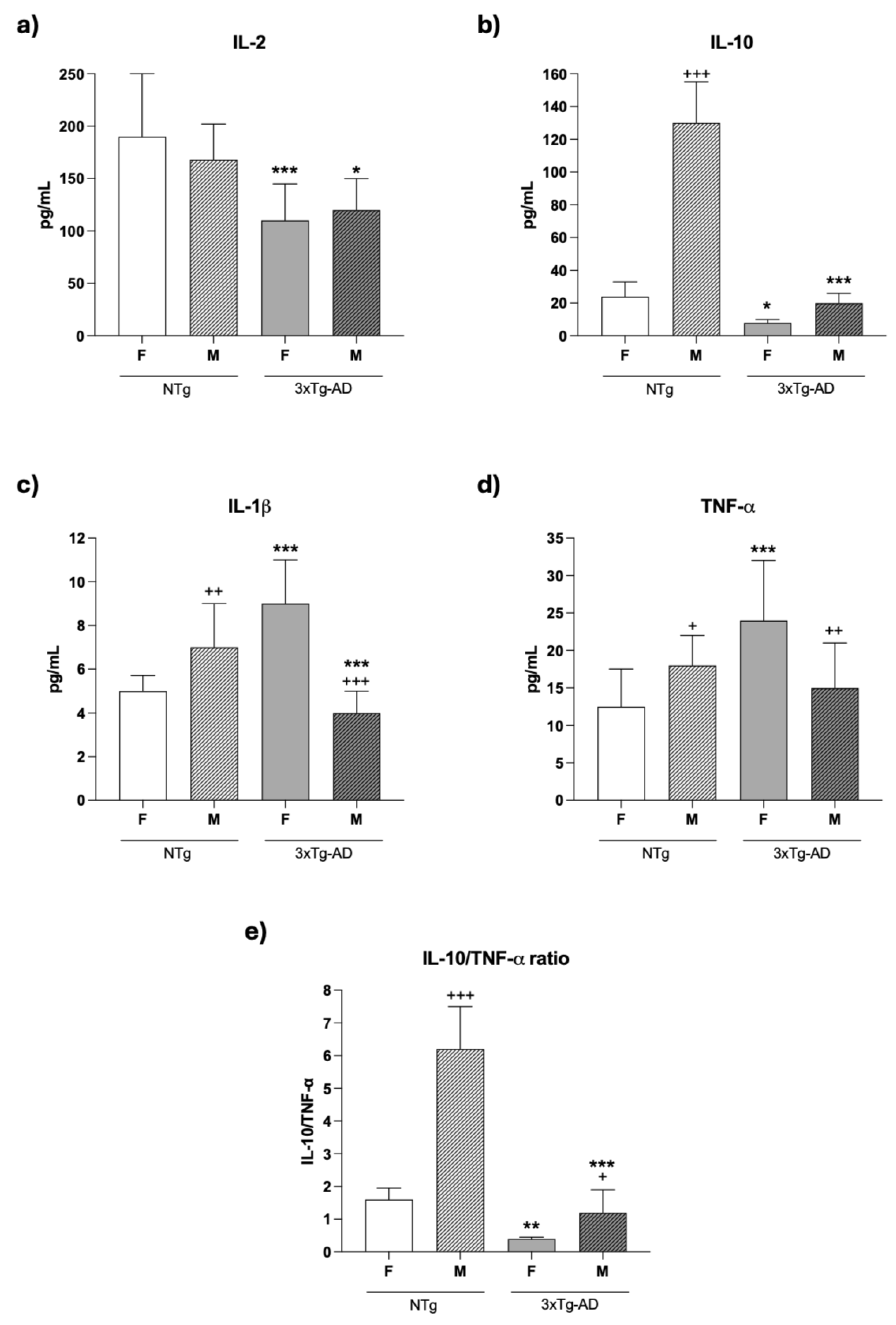
Figure 5.
Glutathione peroxidase (GPx) activity (a), glutathione reductase (GR) activity (b), total reduced glutathione (GSH) concentrations (c) and xanthine oxidase (XO) activity (d) in homogenates of spleens from 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic for Alzheimer’s Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8–11 values (the mean of duplicate assays) corresponding to the same number of subjects (11 NTg female; 9 NTg males; 8 3xTg-AD females; 10 3xTg-AD males). Statistics: * p < 0.05, ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched young group; ++ p < 0.01 vs. the corresponding females.
Figure 5.
Glutathione peroxidase (GPx) activity (a), glutathione reductase (GR) activity (b), total reduced glutathione (GSH) concentrations (c) and xanthine oxidase (XO) activity (d) in homogenates of spleens from 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic for Alzheimer’s Disease (3xTg-AD) mice. Each column represents the mean ± SEM of 8–11 values (the mean of duplicate assays) corresponding to the same number of subjects (11 NTg female; 9 NTg males; 8 3xTg-AD females; 10 3xTg-AD males). Statistics: * p < 0.05, ** p < 0.01, *** p < 0.001 vs. the corresponding gender-matched young group; ++ p < 0.01 vs. the corresponding females.
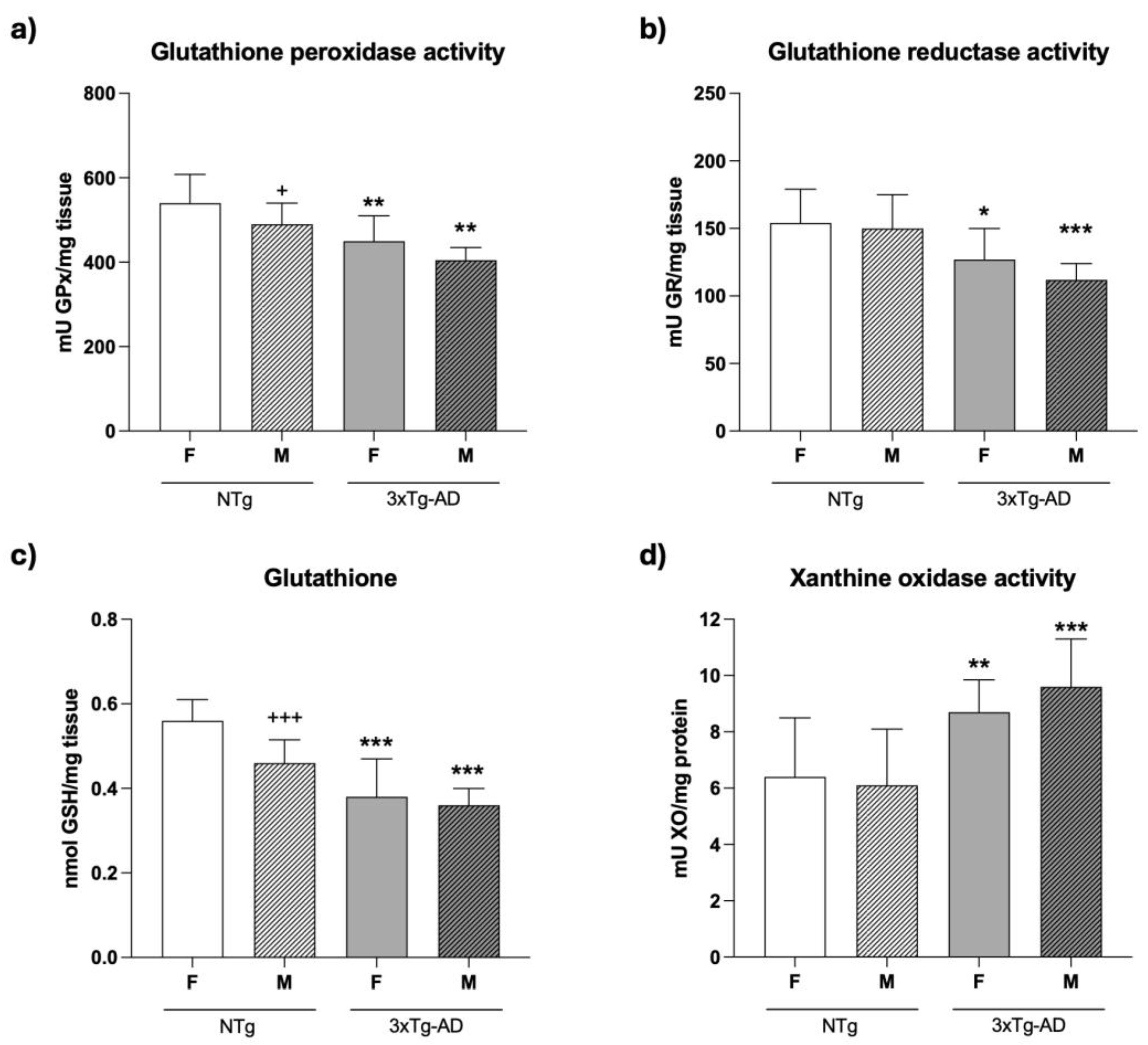

Table 1.
Behavioral profile of 3xTg-AD mice indicates BPSD-like behaviors in both sexes already at 4 months of age.
Table 1.
Behavioral profile of 3xTg-AD mice indicates BPSD-like behaviors in both sexes already at 4 months of age.
| BEHAVIORAL TESTS | NTg | 3xTg-AD | Statistical Factor Analysis G, genotype S, Sex |
||
|---|---|---|---|---|---|
| Females (n=11) | Males (n=9) |
Females (n=8) |
Males (n=10) |
||
| CORNER TEST | |||||
| Sequence of behavioral events | |||||
| Vertical activity (latency, s) | see Fig. 1a | G**,S**, G×S* | |||
| Locomotor activity | |||||
| Horizontal activity (corners, n) | 13.73 ± 0.98 | 11.89 ± 1.29 | 11.25 ± 2.66 | 8.20 ± 1.42 | n.s.,n.s.,n.s. |
| Vertical activity (rearings, n) | 6.36 ± 0.47 | 7.22 ± 1.26 | 4.38 ± 1.02 | 2.30 ± 0.63 | G***, n.s., n.s. |
| OPEN FIELD TEST | |||||
| Sequence of behavioral events | |||||
| Initial movement (latency, s) | 1.36 ± 0.28 | 0.33 ± 0.17 | 2.50 ± 0.53 a | 1.10 ± 0.38 | G*, n.s., n.s. |
| Incidence of bizarre behavior (%) | see Fig. 1b | G***, S**, G×S** | |||
| Exit from the centre (latency, s) | see Fig. 1c | G**, n.s., n.s. | |||
| Entrance to periphery (latency, s) | 15.36 ± 2.04 | 49.4 ± 16.5 | 48.8 ± 27.6 | 24.2 ± 5.05 | n.s., n.s., G×S** |
| Vertical activity (latency, s) | 32.18 ± 2.88 | 46.2 ± 10.2 | 72.1 ± 27.1 a | 74.6 ± 21.0 | G*, n.s., n.s. |
| Self-grooming (latency, s) | see Fig. 1d | G*, n.s., n.s. | |||
| Locomotor activity | |||||
| Vertical activity per minute (rearings, n) |
see Fig. 1e | G*, n.s., n.s./ G*(r2,r3) S*(r4) n.s. |
|||
| Total vertical activity (rearings, n) |
see Fig. 1f | n.s., n.s., n.s. | |||
| Self-grooming behavior | |||||
| Self-grooming episodes (n) | 1.36 ± 0.20 | 1.33 ± 0.37 | 0.75 ± 0.25 | 1.20 ± 0.25 | n.s., n.s., n.s. |
| Total self-grooming duration (s) | 4.55 ± 0.73 | 4.00 ± 0.85 | 2.50 ± 0.76 | 4.40 ± 0.79 | n.s., n.s., n.s. |
| Emotional behaviors | |||||
| Defecation boli (n) | see Fig. 1g | G*, S**, n.s. | |||
| Urination (incidence) | 0.27 ± 0.14 | 0 ± 0 | 0.25 ± 0.16 | 0.30 ± 0.15 | n.s.,n.s.,n.s. |
| T-MAZE | |||||
| Sequence of behavioral events | |||||
| Reaching intersection (latency, s) | see Fig. 1h | G*, n.s., n.s. | |||
| Exploratory efficiency (time, s) | 44.55 ± 14.3 | 56.0 ± 12.6 | 88.0 ± 27.3 | 36.3 ± 12.0 | n.s., n.s., n.s. |
| Ratio Time in the arms (s) | 4.31 ± 0.77 | 4.90 ± 0.72 | 2.86 ± 0.63 a | 2.69 ± 0.37 a | G**, n.s., n.s. |
| Errors (n) | see Fig. 1i | G***, n.s., G×S* | |||
| Emotional behaviors | |||||
| Defecation boli (n) | see Fig. 1j | n.s., S**, n.s. | |||
| Urination (incidence) | 0.55 ± 0.21 | 0 ± 0 | 0.12 ± 0.12 | 0.10 ± 0.10 | n.s., n.s., n.s. |
Results are the mean ± SEM of 8–11 animals. Statistics: Factor analysis: * p < 0.05, ** p < 0.01, and *** p < 0.001 vs. NTg (Genotype effects, G), or female mice (Sex effects, S), or in the comparison to genotype × sex effects (G × S); n.s. non-statistical differences. Post-hoc analysis: a p < 0.05 vs. NTg group with the same sex; b p < 0.05 vs. female group with the same genotype; * vs. all the groups. NTg: non-transgenic mice; 3xTg-AD: triple transgenic mice for Alzheimer's disease.
Table 2.
Physical condition, organometrics and endocrine system in 4-month-old female and male 3xTg-AD and NTg mice.
Table 2.
Physical condition, organometrics and endocrine system in 4-month-old female and male 3xTg-AD and NTg mice.
| NTg | 3xTg-AD | Statistics Factor Analysis G, genotype S, sex |
|||
|---|---|---|---|---|---|
| Females (n=11) | Males (n=9) |
Females (n=8) | Males (n=10) |
||
| Physical condition | |||||
| Body Weight (g) | 21.7 ± 1.00 | 26.7 ± 0.50 b | 26.3 ± 0.60 a | 34.5 ± 0.90 a,b | G*** , S***, n.s. |
| % WAT in body weight | 1.01 ± 0.06 | 0.80 ± 0.03 | 1.39 ± 0.04 a | 1.22 ± 0.14 a | G*** , S***, n.s. |
| Peripheral immune system | |||||
| % Thymus in body weight | 0.18 ± 0.06 | 0.12 ± 0.01 | 0.15 ± 0.00 | 0.10 ± 0.00 | G*** , S** , n.s. |
| % Spleen in body weight | 0.38 ± 0.01 | 0.32 ± 0.01 | 0.52 ± 0.02 a | 0.45 ± 0.07 a | G*** , S** , n.s. |
| Endocrine system | |||||
| Adrenal glands (mg) | 8.2 ± 0.2 | 7.40 ± 0.5 | 7.3 ± 0.2 | 9.4 ± 0.6 | n.s., n.s., G × S** |
| Plasma corticosterone (ng/mL) | 35.5 ± 8.3 | 28.9 ± 7.2 | 43.3 ± 5.7 | 42.6 ± 7.8 | n.s., n.s., n.s. |
Results are the mean ± SEM of 8–11 animals. Statistics: Factor analysis, **p <0.01 and *** p <0.001 compared to NTg (G, genotype), or female mice (S, sex), or in the comparison to genotype x sex (GxS); n.s. non-statistical differences. Post-hoc: a p < 0.05 vs NTg group with the same sex; b p < 0.05 vs. female group with the same genotype. F: females; M: males; NTg: non-transgenic mice; 3xTg-AD: triple transgenic mice for Alzheimer's disease. WAT: white adipose tissue.
| Statistics Factor Analysis G, genotype S, sex |
||
|---|---|---|
| Spleen Chemotaxis (C.I.) | G*** , n.s., G × S* | |
| Thymus Chemotaxis (C.I.) | G*** , n.s., n.s. | |
| Spleen NK Activity (% Lysis) | G***, n.s., G × S* | |
| Thymus NK Activity (% Lysis) | n.s., S***, n.s. | |
| Spleen basal lymphoproliferation (c.p.m.) | n.s., S***, G × S** | |
| Spleen lymphoproliferative response to LPS (% Stimulation) | G*** , S***, G × S* | |
| Spleen lymphoproliferative response to ConA (% Stimulation) | n.s., S***, G × S*** | |
| Thymus basal lymphoproliferation (c.p.m.) | n.s., S***, n.s. | |
| Thymus lymphoproliferative response to LPS (% Stimulation) | G*** , S**, n.s. | |
| Thymus lymphoproliferative response to ConA (% Stimulation) | G*** , S***, n.s. | |
| IL-2 (pg/ml) | G*** , n.s., n.s. | |
| IL-10 (pg/ml) | G*** , S***, G × S*** | |
| IL-1β (pg/ml) | n.s., S**, G × S*** | |
| TNF-α (pg/ml) | G* , n.s., G × S*** | |
| IL-10/TNF-α ratio | G***, S***, G × S*** |
Statistics: Factor analysis, * p < 0.05, ** p < 0.01, *** p <0.001 compared to NTg (G, genotype), or female mice (S, sex), or in the comparison to genotype x sex (GxS); n.s. non-statistical differences. C.I.: Chemotaxis index. NK: Natural Killer. LPS: Lipopolysaccharide. ConA: Concanavaline A. c.p.m.: Counts per minute.
Table 4.
Concentrations of IL-2, IL-10, IL-1β, TNF-α, and TNF-α/IL-10 ratio in supernatants of cultures of splenic leukocytes in response to lipopolysaccharide (LPS) from 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic Alzheimer's Disease (3xTgAD) mice.
Table 4.
Concentrations of IL-2, IL-10, IL-1β, TNF-α, and TNF-α/IL-10 ratio in supernatants of cultures of splenic leukocytes in response to lipopolysaccharide (LPS) from 4-month-old female (F) and male (M) non-transgenic (NTg) and triple transgenic Alzheimer's Disease (3xTgAD) mice.
| NTg | 3xTg-AD | Statistics Factor Analysis G, genotype S, sex |
|||
|---|---|---|---|---|---|
| Females (n=11) | Males (n=9) |
Females (n=8) | Males (n=10) |
||
| IL-2 (pg/ml) | 33 ± 13 | 37 ± 15 | 13 ± 4 aa | 43 ± 16 bbb | n.s., S***, G × S***. |
| IL-10 (pg/ml) | 80 ± 15 | 306 ± 78 bbb | 20 ± 5 aa | 112 ± 40 aaa, bbb | G***,S***, G × S*** |
| IL-1β (pg/ml) | 5 ± 1 | 6 ± 2 | 5 ± 1 | 6 ± 2 | n.s., n.s., n.s. |
| TNF-α (pg/ml) | 18 ± 6 | 18 ± 4 | 17 ± 5 | 16 ± 4 | n.s., n.s., n.s. |
| IL-10/TNF-α ratio | 5.23 ± 0.92 | 14.24 ± 1.95 bbb | 1.34 ± 0.44 aaa | 6.30 ± 1.49 aaa, bbb | G***,S***, G × S*** |
Results are the mean ± SEM of 8–11 animals. Statistics: Factor analysis, *** p <0.001 compared to NTg (G, genotype), or female mice (S, sex), or in the comparison to genotype x sex (GxS); n.s. non-statistical differences. Post-hoc: aa p < 0.01, aaa p < 0.001 vs NTg group with the same sex; bbb p < 0.001 vs. female group with the same genotype. F: females; M: males; NTg: nontransgenic mice; 3xTg-AD: triple transgenic mice for Alzheimer's disease.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



