Submitted:
11 December 2025
Posted:
12 December 2025
You are already at the latest version
Abstract
Endophytic and marine-derived fungi represent prolific and structurally innovative sources of steroidal natural products. In recent years, extensive chemical investigations of diverse fungal taxa—including Aspergillus, Penicillium, and numerous other genera—have revealed an extraordinary variety of steroids and steroid-like metabolites featuring unprecedented carbon skeletons, unusual ring rearrangements, heterocyclic fusions, and hybrid architectures. These metabolites encompass ergostane-, lanostane-, pregnane-, and abeo-type derivatives; secosteroids; polyoxygenated and polycyclic frameworks; meroterpenoid–steroid hybrids; and rare Diels–Alder adducts. Many of these compounds exhibit significant biological activities, including cytotoxic, anti-inflammatory, antimicrobial, antiviral, immunosuppressive, antioxidant, enzyme-inhibitory, larvicidal, neuroprotective, and herbicidal effects. Several metabolites, such as cordycepsterols, citristerones, rubensteroid A, anicequol, and various hybrid steroidal structures, demonstrate potent inhibition of key molecular targets (e.g., COX-2, NF-κB, PTP1B, AChE, NO production) and show promise as leads for anticancer, anti-infective, anti-neuroinflammatory, and metabolic disease therapeutics. Collectively, the rapidly expanding diversity of fungal steroids underscores the remarkable biosynthetic capabilities of fungi and highlights their continued potential as reservoirs of structurally novel and biologically valuable natural products. This review summarizes recent discoveries and structural classes of fungal-derived steroids, emphasizing their chemical diversity, biosynthetic features, and bioactivity profiles.
Keywords:
microorganisms
; bacteria
; fungal endophytes
; steroids
; activity
1. Introduction
Endophytic fungi are microorganisms that inhabit plant tissues for all or part of their life cycle without causing apparent disease. This symbiotic association is often mutualistic, conferring a range of physiological and ecological benefits to the host plant. Endophytes are highly diverse and have been documented in nearly all plant species, colonizing roots, stems, leaves, flowers, and seeds [1,2,3].
The interaction between endophytic fungi and their hosts is particularly compelling due to the multifaceted advantages these microorganisms provide. In exchange for shelter and nutrients, endophytic fungi promote plant growth by enhancing biomass accumulation, height, root development, and the uptake of water and nutrients. These effects are mediated through mechanisms such as the production of phytohormones and the solubilization of essential nutrients, including phosphorus and nitrogen. Endophytes also improve plant tolerance to abiotic stresses—such as drought, salinity, and extreme temperatures—by regulating osmotic balance, modulating hormone levels, and synthesizing antioxidant compounds. Furthermore, several endophytic fungi function as biological control agents, protecting plants against pathogens, insects, and herbivores through the production of antimicrobial and insecticidal secondary metabolites [4,5,6,7,8,9].
Among endophytic fungi, species belonging to the genera Aspergillus [10,11,12] and Penicillium [13,14,15] are particularly noteworthy. These genera are ubiquitous in nature and are recognized for their remarkable capacity to biosynthesize structurally diverse and pharmacologically active secondary metabolites, including steroids, terpenoids, alkaloids, and polyketides. Many of these compounds have demonstrated significant biological activities, such as anti-inflammatory, antibacterial, cytotoxic, and antitumor effects [16,17,18,19,20,21,22].
This review summarizes recent advances in the discovery, structural diversity, and biological activity of bioactive and structurally unusual steroids produced by microbial sources.
2. The Genus Aspergillus
Aspergillus (Ascomycota) is a genus of filamentous fungi characterized by predominantly asexual reproduction. Species within this genus are cosmopolitan and ecologically versatile, playing important roles in natural ecosystems as well as in various sectors of the human economy. Owing to their capacity to produce a wide array of extracellular enzymes and organic acids, Aspergillus species have long attracted scientific interest for their biotechnological and industrial applications. They also synthesize numerous secondary metabolites of relevance to biotechnology and natural product discovery [23,24,25,26].
Despite their beneficial uses, several Aspergillus species are pathogenic to plants and animals. In humans, aspergillosis refers collectively to diseases caused by members of this genus. Systemic Aspergillus infections generally occur in immunocompromised individuals, and the rising incidence of invasive aspergillosis has placed an increasing burden on clinical healthcare systems [26,27,28].
1.1. Steroid Production in Aspergillus aculeatus
Recent work by Yue and colleagues [29] reported the isolation of novel steroids from the endophytic fungus Aspergillus aculeatus. This strain was isolated from the internal tissues of a surface-sterilized leaf of Pseudostellaria heterophylla, collected in Liaoning Province, China. Ps. heterophylla, known as “hai er shen” or “tai zi shen” and commonly referred to as false chickweed, is a eudicot species in the family Caryophyllaceae. It is widely used in traditional Chinese medicine as a qi tonic and a yin-nourishing herb [30,31].
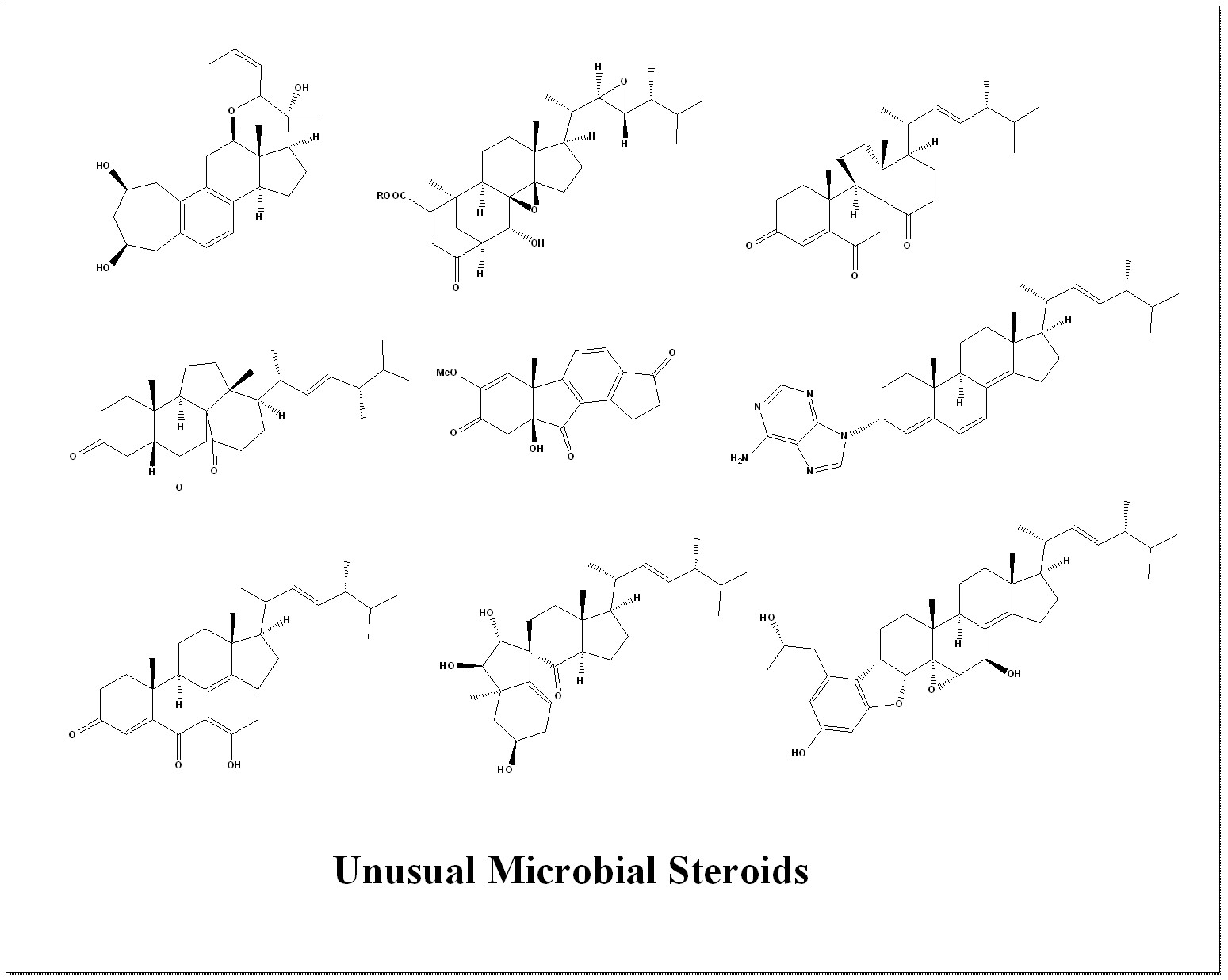
Six new steroids (compounds 1–5 and 7, the structures are shown in Figure 1) were isolated from an ethyl acetate extract obtained after incubating the fungal strain on a rice substrate for 15 days. Structural elucidation was accomplished through comprehensive spectroscopic analyses and comparison with previously reported data. These compounds were identified as camphoratin L (1), camphoratin K (2), camphoratin M (3), camphoratin O (4), camphoratin N (5), and camphoratin P (7). The anti-inflammatory properties of compounds 1, 2, 4, 6, and 8 were assessed by measuring nitric oxide (NO) production in LPS-stimulated RAW264.7 macrophages. Compounds 1, 2, and 6 exhibited moderate inhibitory effects on NO release, indicating potential anti-inflammatory activity [29,32].
Several known steroids were also identified: (22E,24S)-5α,8α-epidioxy-24-ethylcholesta-6,22-dien-3β-ol (6), previously isolated from the mushroom Lactarium volemus [33]; ergosta-7,22-dien-6β-methanoxy-3β,5α-diol (8), previously reported in Aspergillus awamori [34]; and cerevisterol (9), known from Penicillium brasilianum [35].
Further investigation of other fractions from A. aculeatus revealed a broader diversity of steroidal metabolites. HSQC spectral datasets were exported as CSV files and analyzed using the SMART system to predict potential structural types. The computational analysis indicated that the fractions likely contain additional steroids (compounds 10–24), corresponding to structures present in existing spectral databases. Thus, it is reasonable to infer that A. aculeatus also produces steroids (10–24) [29,32].
1.2. Steroids from other Aspergillus Species
Penicysteroid C (25), a polyoxygenated steroid, was isolated from a co-culture of Aspergillus niger and Streptomyces pyomogenus AS63D grown on rice medium under solid-state fermentation. This compound demonstrated antimicrobial and cytotoxic activities, highlighting the potential of interspecies co-cultivation strategies for activating otherwise silent biosynthetic pathways [36].
Aspersteroid A (26, Figure 2), a highly rearranged 1(10→6)-abeo-18,22-cyclosterol, together with two additional 18,22-cyclosterols (27 and 28), was isolated from the culture extract of Aspergillus ustus NRRL 275. Sterol 26 features a highly unusual carbon framework, comprising a 6/6/6/5/5 fused-ring system. Its proposed biosynthetic pathway involves A-ring scission, sequential 1,2-shifts, and subsequent C-18/C-22 cyclization. Notably, aspersteroid A exhibited potent immunosuppressive and antimicrobial activities [37].
An endophytic strain Aspergillus sp. 1022LEF, inhabiting the internal tissues of a red marine alga, produced a novel polyketide–terpene hybrid metabolite, tennessenoid A (29). This compound represents an unprecedented steroid–sorbicillinoid adduct linked through a C–C bond, a rare structural motif in natural products. Tennessenoid A showed broad-spectrum antifungal activity against Sclerotium rolfsii, Fusarium oxysporum, Fusarium sp., Coniella diplodiella, Physalospora piricola, Fusarium graminearum, Alternaria mali, Colletotrichum orbiculare, and Alternaria porri [38].
Spectasterols A–E (30–34), aromatic ergosterols with distinctive ring architectures, were isolated from Aspergillus spectabilis. Sterols 30 and 31 contain a 6/6/6/5/5 ring system with an additional cyclopentene moiety, whereas compounds 32 and 33 possess an unusual 6/6/6/6 system generated through D-ring expansion mediated by 1,2-alkyl shifts. Compound 33 demonstrated cytotoxic activity (IC₅₀ = 6.9 μM) and induced cell cycle arrest and apoptosis in HL-60 cells. It also exhibited anti-inflammatory effects by reducing COX-2 expression at both transcriptional and protein levels and by inhibiting the nuclear translocation of NF-κB p65 [39].
Two structurally unusual naturally occurring Diels–Alder adduct steroids, ergosterdiacids A and B (35 and 36), featuring a 6/6/6/6/5 pentacyclic system, were isolated from a mangrove-derived Aspergillus sp. Both compounds inhibited Mycobacterium tuberculosis protein tyrosine phosphatase B (MptpB), with IC₅₀ values of 15.1 and 30.1 μM, respectively, acting through a noncompetitive mechanism. Additionally, both sterols displayed potent anti-inflammatory activity, suppressing nitric oxide production at 4.5 and 3.6 μM, respectively [40].
From Aspergillus nidulans, two previously undescribed 30-norlanostane triterpenoids, nidulanoids A and B, were isolated along with an ergostane-type steroid featuring an unusual Δ17,20 double bond, designated as (17E,22E,24R)-3β,5α-dihydroxyergosta-7,17,22-trien-6,16-dione (37), and a pregnane derivative, (7Z,9Z,17Z)-2α,3β-dihydroxypregna-7,9,17(20)-trien-18-al (38).
Nidulanoids A and B represent the first natural examples of 30-norlanostane triterpenoids bearing a C9 side-chain at C-17 and a hemiacetal moiety linking C-3 and C-19, suggesting their role as intermediates between lanostanes and conventional steroids. Compound 37 is notable for its atypical Δ17,20 unsaturation, while 38 is the first reported C-21 steroid containing an aldehyde group at C-13. Among these metabolites, compound 38 displayed moderate cytotoxicity against PC12 rat neuronal cancer cells (IC₅₀ = 7.34 μM). Collectively, these findings expand the structural diversity of triterpenoid and steroidal metabolites produced by A. nidulans, and highlight compound 38 as a promising anticancer lead [41].
A sterol derivative, ergosterimide B (39), was isolated from the rice fermentation broth of Aspergillus tubingensis YP-2. This newly characterized compound exhibited weak cytotoxicity, with IC₅₀ = 11.05 μM against tested cell lines [42].
The deep-sea fungus Aspergillus unguis IV17-109 yielded an ergostane-type sterol, aspersterol A (40), distinguished by a rare side chain featuring conjugated Δ17 and Δ22 double bonds. Aspersterol A is an uncommon anthra-steroid bearing a hydroxyl group at C-2 and demonstrated broad cytotoxicity across six cancer cell lines, with a GI₅₀ value of 3.4 μM [43].
A fungal strain, 1901NT-1.40.2, isolated from a Cliona sp. sponge collected at 8–10 m depth in Nha Trang Bay, Vietnam, was identified as Aspergillus subramanianii. This strain produced an undescribed ergostane-type triterpenoid, aspersubrin A (41), representing a new addition to the structural repertoire of Aspergillus metabolites [44].
From the marine-derived endophytic fungus Aspergillus ochraceus MCCC 3A00521, a new ergostane-type sterol, ochrasterone (42), was isolated. This species, an established sterol producer, is preserved in the Marine Culture Collection of China [45].
Finally, a new sterol, aspersterol E (43), was obtained from an endophytic Aspergillus species associated with Hibiscus tiliaceus (Malvaceae, see Figure 3). Compound 43 exhibited cytotoxicity toward MFC mouse pre-gastric cancer cells (IC₅₀ = 153.43 μM) [46]. In addition, a newly isolated Aspergillus species from soil in Qalubiya Governorate, Egypt, displayed strong antimicrobial activity. Chemical analysis revealed the presence of a polyhydroxysterol (44), which demonstrated potent antimicrobial effects [47].
An endophytic fungal strain was isolated from the fruit of the mangrove tree Avicennia marina, collected at Kilo 17, Safaga, Red Sea, Egypt. From this isolate, identified as Aspergillus versicolor, a new epoxy-ergostane sterol named versicolor (45) was obtained as a minor metabolite from fermented rice cultures. Versicolor exhibited inhibitory activity against the SARS-CoV-2 3CL protease (3CLpro), with an IC₅₀ value of 2.168 ± 0.09 μM, indicating its potential as a candidate inhibitor of SARS-CoV-2 3CLpro [48].
A second marine-derived fungus, Aspergillus sp. ZJUT223, was isolated from seawater collected in the Grant Trough near the Marceau Trough and identified through ITS sequencing. Extraction was performed using ethanol followed by purification with ethyl acetate. From this strain, a new steroid, ganodermanic acid (46), was isolated [49].
Additionally, a new 18,22-cyclosterol, aspersteroline A (47), together with a structurally related analogue (48), was obtained from the soil-derived Aspergillus versicolor QC812. Both compounds showed moderate cytotoxic activity against the HL-60 human leukemia cell line [50].
Two oxygenated ergostane-type steroids—one new compound, 3β-hydroxy-5α,6β-methoxyergosta-7,22-dien-15-one (49), and one known analogue, ergosta-6,22-dien-3β,5α,8α-triol (50)—were isolated from crude extracts of a marine sponge-derived Aspergillus sp. Notably, compound 49 represents a marine ergostane-type steroid featuring two methoxy groups at C-5 and C-6 and demonstrated antibacterial activity against Staphylococcus aureus [51].
3. The Genus Penicillium
Penicillium is a globally distributed genus of blue–green molds belonging to the kingdom Fungi. Species within this genus reproduce asexually (anamorphic or deuteromycetous forms) and play major ecological roles as decomposers of organic matter. Many species cause destructive spoilage in the food industry and produce a broad range of mycotoxins, while others function as prolific producers of industrial enzymes or are recognized as common indoor allergens. Although DNA sequencing is essential for accurate species identification, the lack of a comprehensive and validated reference database continues to challenge Penicillium taxonomy [52,53,54].
With the adoption of the “one fungus, one name” principle under the International Code of Nomenclature for algae, fungi, and plants, the generic boundaries of Penicillium have been revised, incorporating species previously assigned to genera such as Chromocleista, Eladia, Eupenicillium, Torulomyces, and Thysanophora. Together, these taxa form a broader monophyletic clade. Reflecting the numerous new species described in recent years, the list of accepted Penicillium species has been updated and currently includes 354 recognized species. This revision also includes new combinations involving Aspergillus crystallinus, A. malodoratus, and A. paradoxus, all of which have been reassigned to Penicillium section Paradoxa [55,56,57].
To increase the taxonomic utility of the updated species list, information is provided on MycoBank numbers, live ex-type cultures, and GenBank accession numbers for ITS, β-tubulin, calmodulin, and RPB2 sequences, thereby supplying a verified reference set for researchers working with this genus. Standardized protocols for species description and identification are also recommended to improve reliability and reproducibility across laboratories [58,59,60,61,62,63].
3.1. Steroidal Metabolites from Penicillium
A new steroid, persteroid (51, see Figure 4), was isolated from the marine-derived Penicillium sp. ZYX-Z-143. Persteroid exhibited inhibitory activity against protein tyrosine phosphatase 1B (PTP1B), with an IC₅₀ value of 46 μM, and strongly suppressed nitric oxide (NO) production in LPS-stimulated RAW264.7 macrophages, suggesting potential anti-inflammatory properties [64].
Another new steroid, penivariod A (52), was isolated from Penicillium variabile EN-394, an endophytic strain obtained from the marine red alga Rhodomela confervoides. Penivariod A demonstrated potent antimicrobial activity, particularly against Escherichia coli and Pseudomonas aeruginosa, with MIC values of 1.0 and 2.0 μg/mL, respectively [65].
A series of unusual C₂₅ steroids (53–66), characterized by a distinctive bicyclo[4.4.1] A/B ring system, were isolated from an antitumor mutant strain of Penicillium purpurogenum G59 AD-1-2. The isolated metabolites included antineocyclocitrinols A (53) and B (54), and 23-O-methylantineocyclocitrinol (55), all featuring a bicyclo[4.4.1] A/B framework with a Z-configured Δ20,22 double bond. Additional C₂₅ steroids—neocyclocitrinols A (57), B (56), C (59), and D (58), threo-23-O-methylneocyclocitrinol (60), erythro-23-O-methyl-neo-cyclocitrinol (61), 24-epi-cyclocitrinol (62), cyclocitrinol (63), 20-O-methyl-24-epi-cyclocitrinol (64), 20-O-methylcyclocitrinol (65), and isocyclocitrinol B (66)—were also identified. All compounds displayed varying degrees of cytotoxicity against multiple human cancer cell lines, highlighting the pharmacological potential of this structurally unique class of C₂₅ steroids [66].
Two new C₂₃-steroid derivatives, cyclocitrinic acid A (67) and cyclocitrinic acid B (68), were isolated from the mangrove-derived fungus Penicillium sp. SCSIO 41429. Cyclocitrinic acid B demonstrated moderate pancreatic lipase inhibition, with an IC₅₀ value of 32 μM, and exhibited both pancreatic lipase inhibitory and antioxidant properties [67].
From the plant-associated fungus Penicillium fellutanum, an unusual clathrate-type meroterpenoid, isoatlantinone A (69), along with two new steroids, acrocalysterols E (70) and F (71), was isolated. Isoatlantinone A is notable for its highly oxygenated meroterpenoid structure featuring a unique caged bioxatetracyclo-[6.3.2.0¹,⁶.0¹,¹²]-tridecane ring system. All isolates were screened for antifungal and cytotoxic activities, among which compound 71 exhibited potent cytotoxicity toward HCC-1806 human breast cancer cells (IC₅₀ = 18.15 ± 1.05 μM). These findings highlight P. fellutanum as a promising source of structurally novel and bioactive metabolites [68].
Chemical investigation of Penicillium oxalicum 2021CDF-3, an endophytic fungus associated with marine red algae, led to the discovery of a new polyoxygenated ergostane steroid, peniciloxatone A (72). This compound showed cytotoxic activity against FADU and HepG2 cell lines, with IC₅₀ values of 9.5 and 18.1 μM, respectively [69].
A new steroid with strong antibacterial activity, rubensteroid A (73), along with its decarboxylated analogue, solitumergosterol A (74), was isolated from the Magellan Seamount-derived fungus Penicillium rubens AS-130. Rubensteroid A features a rare 6/6/6/6/5 pentacyclic ring system, proposed to originate from a [4+2] Diels–Alder cycloaddition between 14,15-didehydroergosterol (14-DHE) and maleic acid or maleimide, followed by decarboxylation. Compound 73 demonstrated potent antibacterial activity against Escherichia coli and Vibrio parahaemolyticus, both with MIC values of 0.5 μg/mL [70].
From the lichen-associated fungus Penicillium aurantiacobrunneum, two new sterols—(20S)-hydroxy-24(28)-dehydrocampesterol (75) and 7α-methoxy-8β-hydroxy-paxisterol (76)—were obtained. Sterol 75 showed cytotoxicity against the HPAC pancreatic adenocarcinoma epithelial cell line (IC₅₀ = 17.76 ± 5.35 μM) [71].
A new cytotoxic steroid, 16α-methylpregna-17α,19-dihydroxy-(9,11)-epoxy-4-ene-3,18-dione-20-acetoxy (77), was isolated from Penicillium citrinum SCSIO 41017, associated with the sponge Callyspongia sp. This compound displayed moderate cytotoxicity against MCF-7 human breast cancer cells, with IC₅₀ values of 13.5–18.0 μM [72].
From mangrove sediments collected in the Dongzhaigang Mangrove Reserve (Hainan, China), strain ABC190807 of Penicillium brefeldianum was isolated. Its EtOAc extract exhibited potent larvicidal activity against Aedes aegypti third-instar larvae (LC₅₀ = 0.089 mg/mL). A novel purinyl steroid, ergosta-4,6,8(14),22-tetraen-3-(6-amino-9H-purin-9-yl) (78), was isolated from this extract [73].
Sterolic acid (79, see Figure 5), an unusual sterol, was isolated from a deep-sea sediment-derived Penicillium sp. This metabolite features a diepoxy moiety within its A-ring and an oxabicyclo[2.2.2]octane system—structural motifs previously known only from plant-derived natural products. Sterolic acid additionally contains a carboxylic acid group at C-27, further distinguishing it from typical fungal sterols [74].
Penicillitone (80), a sterol with a rare tetracyclic skeleton, was obtained from Penicillium purpurogenum. Penicillitone exhibited notable cytotoxicity toward multiple cancer cell lines, including A549 (IC₅₀ = 5.57 μM), HepG2 (IC₅₀ = 4.44 μM), and MCF-7 (IC₅₀ = 5.98 μM), with adriamycin serving as a positive control [75].
A novel inhibitor of anchorage-independent tumor cell growth was isolated from the culture broth of Penicillium aurantiogriseum TP-F0213. The compound, identified as 16-acetoxy-3,7,11-trihydroxyergost-22-en-6-one (81), known as anicequol, possesses an ergostane-type carbon skeleton with substituent configurations 3β, 5α, 7β, 11β, 16β, and 24S. Anicequol inhibited anchorage-independent proliferation of DLD-1 human colon cancer cells with an IC₅₀ of 1.2 μM, while showing substantially lower activity against anchorage-dependent growth (IC₅₀ = 40 μM) [76].
A new polyoxygenated steroid, penicisteroid A (82), was isolated from the culture extract of Penicillium chrysogenum QEN-24S, an endophytic strain from an unidentified Laurencia species (see Figure 6) of marine red algae [77].
The steroid 8(14),22E-dien-3β,5α,6β,7α-tetraol (83) was isolated from a Penicillium sp. associated with South Pole sea moss. This compound showed anticancer activity toward HepG2 liver cancer cells, with an IC₅₀ of 10.4 μg/mL [78].
Two novel naturally occurring [4+2] Diels–Alder cycloaddition ergosteroids (84 and 85) were isolated from Penicillium herquei. These compounds represent the first known steroidal cycloadducts formed with 1,4,6-trimethyl-1,6-dihydropyridine-2,5-dione or 4,6-dimethyl-1,6-dihydropyridine-2,5-dione [79].
Several novel steroids—citristerones A (86), B (87), D (88), E (89), and a new series of 23,24-diol-containing ergosterols (90)—along with three known analogues, were isolated from Penicillium citrinum TJ507, an endophytic strain from Hypericum wilsonii. Citristerone B exhibited exceptional anti-neuroinflammatory activity (IC₅₀ = 0.60 μM) in LPS-stimulated BV-2 microglial cells. Further mechanistic studies revealed that citristerone B markedly reduced NO and cytokine release, inhibited TNF-α, iNOS, and NF-κB expression, and suppressed ROS accumulation [80].
Three andrastin-type meroterpenoids, hemiacetalmeroterpenoids A–C (91–93), were isolated from the mangrove-soil-derived Penicillium sp. N-5. Hemiacetalmeroterpenoid A (91) possesses a unique, highly congested 6,6,6,6,5,5 hexacyclic skeleton and exhibited strong antimicrobial activity against Penicillium italicum and Colletotrichum gloeosporioides (MIC = 6.25 μg/mL) [81].
Two novel nortriterpenoids (94 and 95) were isolated from the endophytic fungal strain Penicillium ochrochloron SWUKD4.1850, collected from healthy Kalmia angustifolia in Yunnan Province, China. Compound 95 represents the first naturally occurring 27-nor-3,4-secocycloartane shinortriterpenoid and exhibited moderate cytotoxicity toward HL-60, SMMC-7721, and MCF-7 cell lines (IC₅₀ = 6.5–17.8 μM) [82].
From Penicillium expansum WTJP1, isolated from Aconitum carmichaelii, a previously undescribed compound named expansinin (96) was discovered. Expansinin represents the first naturally occurring conjugate of an indole alkaloid and an ergosteroid, and was evaluated for cytotoxicity against five human cancer cell lines [83].
Scabrosteroid A (97), a novel steroidal heterodimer, was isolated from Penicillium scabrosum FXI744. This compound represents the first example of a naturally occurring pyrrolidinone–ergosterol hybrid, linked via a C-3/C-3′ bond. Scabrosteroid A inhibited NO production (IC₅₀ = 9.5 μM) and showed moderate immunosuppressive activity [84].
Penicildiones A (98) and B (99), two new steroids, were isolated from the soft-coral-derived fungus Penicillium sp. SCSIO 41201 cultured in 1% NaCl potato dextrose broth [85].
Finally, isopenicins A (100) and C (101)—novel meroterpenoids with unprecedented terpenoid–polyketide hybrid skeletons—were isolated from Penicillium sp. sh18. Compound 102 was identified as a potent inhibitor of the Wnt/β-catenin signaling pathway [86].
4. Steroids Produced by Miscellaneous Microorganisms
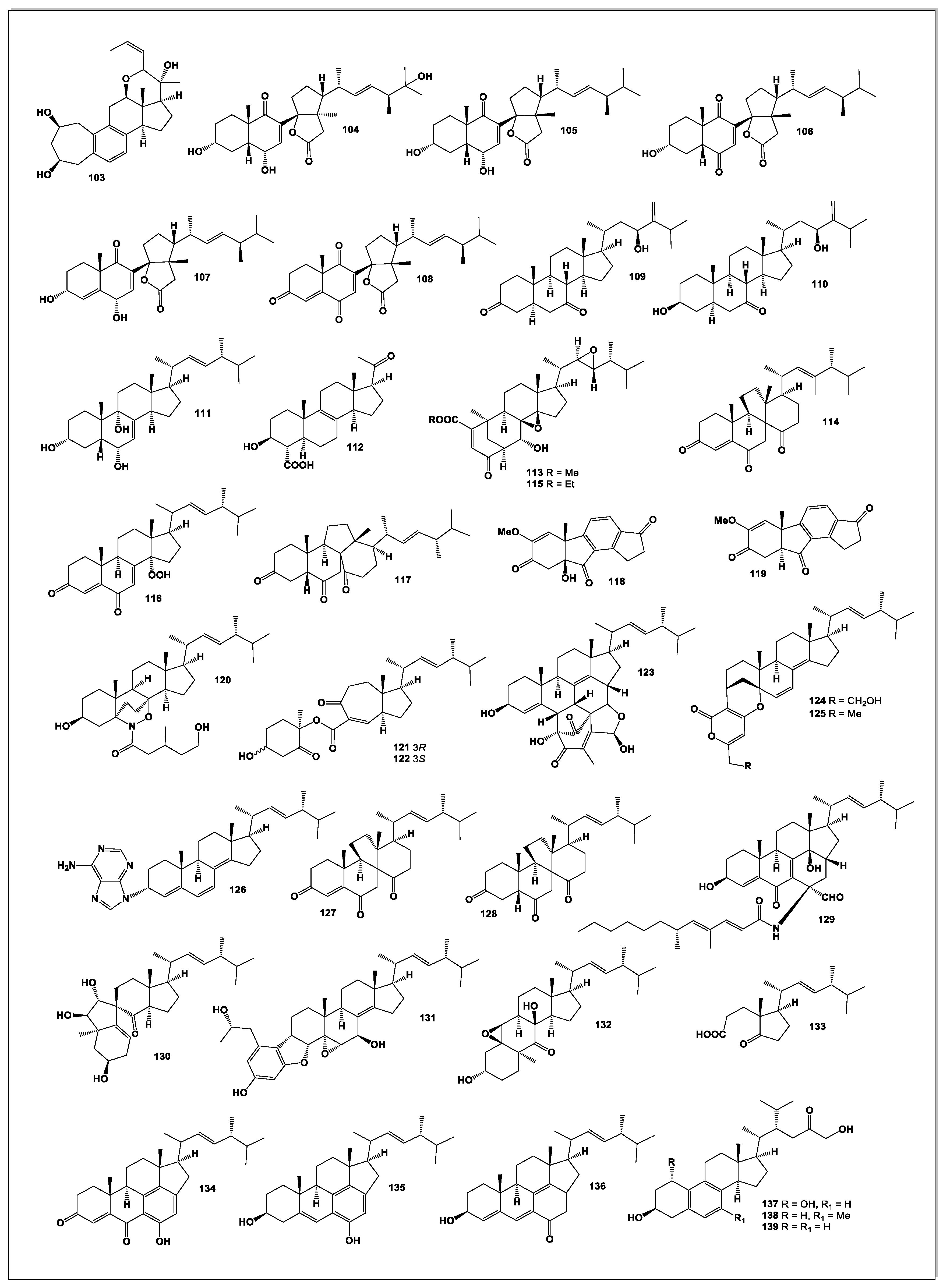
A structurally unique C₂₅ steroid, phomarol (103, see Figure 7), was isolated from a cultured Phoma sp. derived from the giant jellyfish Nemopilema nomurai. Phomarol is distinguished by a seven-membered carbocyclic A ring [1(10→19)abeo], an aromatic B ring, and a cyclized side chain forming a fused pentacyclic skeleton [87].
From the mangrove endophytic fungus Talaromyces sp. SCNU-F0041, a rare 9,11-secosteroid—cyclosecosteroid A (104)—was isolated. This compound exhibited moderate acetylcholinesterase (AChE) inhibitory activity, with an IC₅₀ value of 46 μM [88].
Four new 9,11-secosteroid-derived γ-lactones—altersteroids A (105), B (106), C (107), and D (108)—were obtained from cultures of the ascomycete Alternaria sp. These metabolites contain a γ-lactone fused to the steroid D ring at C-13/C-14. Among them, compound 107 demonstrated moderate cytotoxicity against several tumor cell lines and induced apoptosis in A549 cells; notably, it displayed equal potency against both cisplatin-sensitive MB49 and cisplatin-resistant MB49 CisR cells (IC₅₀ = 12.7 μM) [89].
Two new steroids, acrocalysterols A (109) and B (110), were isolated from Acrocalymma sp., an endophytic fungus obtained from the tender stems of Sinomenium acutum. Compound 110 displayed potent cytotoxicity toward HeLa, HCC-1806, and RKO cancer cell lines, with IC₅₀ values ranging from 18.37 to 19.64 μM [90].
A new ergostane-type steroid, (22E)-3α,6α,9α-ergosta-7,22-diene-3,6,9-triol (111), was isolated from Periconia pseudobyssoides K5 grown on fermented unpolished rice. This endophytic fungus was isolated from a deciduous tree Toona sureni. Compound 111 inhibited heme polymerization with an IC₅₀ of 8.24 ± 0.03 mg/mL [91].
Investigation of the endophyte Phaeosphaeria spartinae, associated with the marine red alga Ceramium sp., yielded spartopregnenolone (112), a metabolite combining features of triterpenes and steroids. Its structure—including a Δ⁸,⁹ double bond, a C-4 carboxyl group, and an acetylated side chain—identifies it as a 4α-carboxy-8,9-pregnene derivative representing a transitional form in triterpene-to-steroid biosynthesis [92].
Two new functionalized ergostane-type steroids, phomopsterones A (113) and B (114), were isolated from Phomopsis sp. TJ507A. Phomopsterone A features an unprecedented rearranged bicyclo[3.3.1]nonane motif formed through B-ring scission and subsequent 180° rotation of the A ring. Phomopsterone B exhibited anti-inflammatory activity [93].
Phytochemical analysis of Stereum hirsutum fermentation broth led to the identification of ten steroids, including two previously unreported ones—steresterones A (115) and B (116)—and compound 117. Compound 117 displayed significant anti-proliferative activity, with IC₅₀ values as low as 2.3 μM [94].
Two unusual steroid-like metabolites, asterogynin A (118) and asterogynin B (119), were obtained from an endophytic fungus isolated from the small palm Asterogyne martiana [95].
From Trichoderma koningiopsis, an endophytic fungus isolated from the gut of a centipede collected on Tongji campus, a steroidal derivative (120) was isolated. Named trichosterol A, it represents the first naturally occurring steroid–alkaloid hybrid containing a rare 6/6/6/5/6 pentacyclic skeleton incorporating an unusual 1,2-oxazine moiety. Compound 120 exhibited notable herbicidal activity against Medicago sativa, highlighting its potential as a natural bioherbicide lead molecule [96].
Two novel steroids—microascusteroids A and B (121 and 122)—were isolated from the marine-derived fungus Microascus sp. SCSIO 41821. These metabolites are unusual 5,6-seco-9,10-seco ergostane derivatives featuring a rearranged C ring. Both compounds moderately inhibited NO production in LPS-stimulated RAW 264.7 cells and showed mild inhibitory effects on PTP1B as well as cytotoxicity against H1975 and HepG2 cancer cells [97].
Chaeglobol A (123), obtained from Chaetomium globosum HBU-45, possesses a highly unusual octacyclic 6/6/6/5/6/5/6/5 skeleton. Its biosynthesis is proposed to involve a [4+2] cycloaddition followed by enzymatic cyclization. Chaeglobol A inhibited Botryosphaeria dothidea by disrupting cell membrane integrity and inducing oxidative damage [98].
Three unprecedented hybrid steroids—striasteroids A–C (124–126)—were isolated from Striaticonidium cinctum SCSIO 41432. Striasteroids A and B feature a new 6/6/6/6/6/5 hexacyclic framework forming polyketide–steroid hybrids bridged by an oxabicyclo[3.3.1]nonane moiety. Striasteroid C (126), the first naturally occurring adenine–steroid hybrid, showed strong neuraminidase inhibition (IC₅₀ = 6.15 ± 1.15 μM) [99].
The sponge-derived fungus Gymnacella dankaliensis produced two exceptionally unusual steroids, dankasterones A (127) and B (128), when cultured in a modified malt extract medium containing soluble starch instead of glucose. Gymnasterone A (129) was obtained from standard malt extract medium. All steroids except 129 inhibited growth of murine P388 cancer cells, and dankasterone A also showed potent activity against human cancer cell lines [100].
Two undescribed steroids, bipolarsterols A (130) and B (131), were isolated from pathogenic fungus Bipolaris oryzae, along with nine known congeners. Bipolarsterol A represents the first example of a 19(10→5)-abeo-7(8→9)-abeo-ergostane featuring a spiro[4.5]decan-6-one system, while bipolarsterol B is a new member of the rare steroid–phenylpropanoid hybrid class [101].
Matsutakone (132), a novel sterol featuring an unprecedented polycyclic ring system, along with a new norsteroid, matsutoic acid (133), was isolated from the fruiting bodies of Tricholoma matsutake. Bioassay results demonstrated that both compounds exhibited inhibitory activity against acetylcholinesterase, with compound 132 displaying an IC₅₀ value of 20.9 μM [102].
Three unusual C₃₀ ergosterols—cordycepsterols A–C (134–136)—were isolated from the medicinal fungus Cordyceps militaris. These sterols possess an uncommon 6/6/6/5/6 pentacyclic skeleton that is hypothesized to arise from a canonical ergosterol framework through extension by two additional skeletal carbons. All three metabolites showed potent inhibition of nitric oxide production, with IC₅₀ values of 3.0, 0.9, and 2.3 μM, respectively. At 5 μM, they significantly suppressed the secretion of tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6), underscoring their promising anti-inflammatory potential and highlighting C. militaris as a valuable source of bioactive sterols [103].
Mangrove endophytic fungi, as extremophilic microorganisms, are known to produce diverse and biologically active secondary metabolites. A strain of Dothiorella sp. ZJQQYZ-1, isolated from the mangrove plant Kandelia candel, yielded six metabolites, including three new benzofuran derivatives and three new steroids: phomosterol C (137), phomosterol B (138), and phomosterol A (139). Among these, phomosterol A exhibited significant anti-inflammatory activity with an IC₅₀ value of 4.6 μM. Mechanistic studies further revealed that compound 139 effectively suppressed the protein expression of inducible nitric oxide synthase (iNOS) in LPS-stimulated RAW264.7 macrophages [104].
5. Conclusion
Fungal organisms—particularly endophytic, marine-derived, and extremophilic species—continue to demonstrate exceptional biosynthetic richness, yielding steroids with unprecedented structural diversity. The discoveries summarized in this review highlight an expanding repertoire of ergostane, lanostane, pregnane, abeo-rearranged, secosteroid, and hybrid scaffolds, many of which feature rare ring systems, extensive oxygenation, or heterocyclic fusions not previously observed in nature. These structural innovations frequently translate into notable biological activities, including cytotoxic, anti-inflammatory, antimicrobial, neuroactive, and enzyme-inhibitory effects, positioning fungal steroids as promising leads for drug discovery and agricultural applications. As advances in isolation techniques, spectroscopy, genome mining, and coculture strategies continue to evolve, the rate of discovering novel steroidal metabolites is likely to accelerate. Overall, fungi represent a largely untapped reservoir of chemically and biologically compelling steroids, underscoring their value in natural product research and their potential contribution to future therapeutic development.
Author Contributions
Conceptualization, V.M.D.; methodology, V.M.D.; software, A.O.T.; investigation, V.M.D.; resources, V.M.D.; writing—original draft preparation, A.O.T. and V.M.D.; writing—review and editing, A.O.T. and V.M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gao, Y.; Xu, Y.; Dong, Z.; Guo, Y.; Luo, J.; Wang, F.; Zou, X. Endophytic fungal diversity and its interaction mechanism with medicinal plants. Molecules 2025, 30, 1028. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic fungi in forest trees: are they mutualists? Fungal Biology Reviews 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Saxena, S.; Dufossé, L.; Deshmukh, S.K.; Chhipa, H.; Gupta, M.K. Endophytic fungi: a treasure trove of antifungal metabolites. Microorganisms 2024, 12, 1903. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O. Azo dyes and the microbial world: synthesis, breakdown, and bioactivity. Microbiology Research 2025, 16, 100. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Ayilara, M.S.; Akinola, S.A.; Babalola, O.O. Biocontrol mechanisms of endophytic fungi. Egyptian J. Biological Pest Control 2022, 32, 46. [Google Scholar] [CrossRef]
- Dos Reis, J.B.A.; Lorenzi, A.S.; do Vale, H.M.M. Methods Used for the Study of Endophytic Fungi: A Review on Methodologies and Challenges, and Associated Tips. Arch. Microbiol. 2022, 204, 675. [Google Scholar] [CrossRef]
- Omomowo, I.O.; Amao, J.A.; Abubakar, A.; Ogundola, A.F.; Ezediuno, L.O.; Bamigboye, C.O. A Review on the Trends of Endophytic Fungi Bioactivities. Sci. Afr. 2023, 20, e01594. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C. Endophytic Fungi: A Tool for Plant Growth Promotion and Sustainable Agriculture. Mycology 2022, 13, 39–55. [Google Scholar] [CrossRef]
- Bogas, A.C.; Cruz, F.P.N.; Lacava, P.T.; Sousa, C.P. Endophytic Fungi: An Overview on Biotechnological and Agronomic Potential. Braz. J. Biol. 2024, 84, e258557. [Google Scholar] [CrossRef]
- Li, H.; Fu, Y.; Song, F. Marine Aspergillus: A Treasure Trove of Antimicrobial Compounds. Mar. Drugs 2023, 21, 277. [Google Scholar] [CrossRef]
- Tian, F.; Woo, S.Y.; Lee, S.Y.; Park, S.B.; Zheng, Y.; Chun, H.S. Antifungal Activity of Essential Oils and Plant-Derived Natural Compounds against Aspergillus flavus. Antibiotics 2022, 11, 1727. [Google Scholar] [CrossRef]
- Jangid, H.; Garg, S.; Kashyap, P.; Karnwal, A.; Shidiki, A.; Kumar, G. Bioprospecting of Aspergillus sp. as a Promising Repository for Anti-Cancer Agents: A Comprehensive Bibliometric Investigation. Front. Microbiol. 2024, 15, 1379602. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, A.C.; Ogawa, C.Y.; Rodrigues, L.C.; de Medeiros, L.S.; Veiga, T.A.M. Penicillium Genus as a Source for Anti-Leukemia Compounds: An Overview from 1984 to 2020. Leuk. Lymphoma 2021, 62, 2079–2093. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Fu, Y.; Song, F.; Xu, X. Recent Updates on the Antimicrobial Compounds from Marine-Derived Penicillium Fungi. Chem. Biodiversity 2023, 20, e202301278. [Google Scholar] [CrossRef]
- Jahan, I.; Wang, Y.; Li, P.; Hussain, S.; Song, J.; Yan, J. Comprehensive Analysis of Penicillium sclerotiorum: Biology, Secondary Metabolites, and Bioactive Compound Potential. J. Agric. Food Chem. 2024, 72, 9555–9566. [Google Scholar] [CrossRef]
- Donova, M.V.; Egorova, O.V. Microbial Steroid Transformations: Current State and Prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef]
- Averina, O.V.; Zorkina, Y.A.; Yunes, R.A.; Kovtun, A.S.; Ushakova, V.M.; Morozova, A.Y.; et al. Bacterial Metabolites of Human Gut Microbiota Correlating with Depression. Int. J. Mol. Sci. 2020, 21, 9234. [Google Scholar] [CrossRef]
- Russell, W.R.; Hoyles, L.; Flint, H.J.; Dumas, M.E. Colonic Bacterial Metabolites and Human Health. Curr. Opin. Microbiol. 2013, 16, 246–254. [Google Scholar] [CrossRef]
- De Carvalho, C.C.; Fernandes, P. Production of Metabolites as Bacterial Responses to the Marine Environment. Mar. Drugs 2010, 8, 705–727. [Google Scholar] [CrossRef]
- Audrain, B.; Farag, M.A.; Ryu, C.M.; Ghigo, J.M. Role of Bacterial Volatile Compounds in Bacterial Biology. FEMS Microbiol. Rev. 2015, 39, 222–233. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Hydrobiological Aspects of Fatty Acids: Unique, Rare, and Unusual Fatty Acids Incorporated into Linear and Cyclic Lipopeptides and Their Biological Activity. Hydrobiology 2022, 1, 331–432. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Terent’ev, A.O.; Baranin, S.V. Boronosteroids as Potential Antitumor Drugs: A Review. Tumor Discov. 2025, 3, 025290069. [Google Scholar] [CrossRef]
- Bennett, S.E.B.J.W. An Overview of the Genus Aspergillus. In The Aspergilli; 2007; pp. 23–34. [Google Scholar]
- Krijgsheld, P.; Bleichrodt, R.V.; Van Veluw, G.J.; Wang, F.; Müller, W.H.; Dijksterhuis, J.; Wösten, H.A.B. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef]
- Schuster, E.; Dunn-Coleman, N.; Frisvad, J.C.; Van Dijck, P.W. On the Safety of Aspergillus niger: A Review. Appl. Microbiol. Biotechnol. 2002, 59, 426–435. [Google Scholar] [CrossRef]
- Xue, M.; Qu, Z.; Moretti, A.; Logrieco, A.F.; Chu, H.; Zhang, Q.; et al. Aspergillus Mycotoxins: The Major Food Contaminants. Adv. Sci. 2025, 12, 2412757. [Google Scholar] [CrossRef]
- Zakaria, L. An Overview of Aspergillus Species Associated with Plant Diseases. Pathogens 2024, 13, 813. [Google Scholar] [CrossRef]
- Sun, Z.; Wu, Y.; Long, S.; Feng, S.; Jia, X.; Hu, Y.; Zeng, B. Aspergillus oryzae as a Cell Factory: Research and Applications in Industrial Production. J. Fungi 2024, 10, 248. [Google Scholar] [CrossRef]
- Yue, F.; Yang, Y.H.; Liu, L.; Shen, Y.; Li, Q.M.; Li, M.; Pei, Y.H. Steroids Isolated from Endophytic Fungus Aspergillus aculeatus. SSRN Preprint 2025. [Google Scholar] [CrossRef]
- Hu, D.J.; Shakerian, F.; Zhao, J.; Li, S.P. Chemistry, pharmacology and analysis of Pseudostellaria heterophylla: a mini-review. Chinese Medicine 2019, 14, 21. [Google Scholar] [CrossRef]
- Lei, Z.; Luan, F.; Zou, J.; Zhang, X.; Zhai, B.; Xin, B.; Shi, Y. Traditional uses, phytochemical constituents, pharmacological properties, and quality control of Pseudostellaria heterophylla (Miq.) Pax. J. Ethnopharmacol. 2025, 337, 118871. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Zhang, R.H.; Liu, L.Y.; Yang, Y.H.; Shen, Y.; Li, Q.M.; Pei, Y.H. Steroids, peniciversiols and aspergilosidols from endophytic fungus Aspergillus sp. TZS-Y4. Fitoterapia 2025, 106650. [Google Scholar] [CrossRef]
- Yue, J.; Chen, S.; Lin, Z.; Sun, H. Sterols from the fungus Lactarium volemus. Phytochemistry 2001, 56, 801–806. [Google Scholar] [CrossRef]
- Gao, H.; Hong, K.; Zhang, X.; Liu, H.; Wang, N.; Zhuang, L.; Yao, X. New steryl esters of fatty acids from the mangrove fungus Aspergillus awamori. Helv. Chim. Acta 2007, 90, 1165–1178. [Google Scholar] [CrossRef]
- Tang, H.; Zhang, Q.; Li, H.; Gao, J. Antimicrobial and allelopathic metabolites produced by Penicillium brasilianum. Nat Prod. Res. 2015, 29, 345–348. [Google Scholar] [CrossRef]
- Abdel-Razek, A.S.; Hamed, A.; Frese, M.; Sewald, N.; Shaaban, M. Penicisteroid C: New polyoxygenated steroid produced by co-culturing of Streptomyces piomogenus with Aspergillus niger. Steroids 2018, 138, 21–25. [Google Scholar] [CrossRef]
- Liu, L.; Duan, F.F.; Gao, Y.; Peng, X.G.; Chang, J.L.; Chen, J.; Ruan, H.L. Aspersteroids A–C, three rearranged ergostane-type steroids from Aspergillus ustus NRRL 275. Organic Letters 2021, 23, 9620–9624. [Google Scholar] [CrossRef]
- Zhao, D.-L.; Wang, H.-S.; Gao, L.-W.; Zhang, P. Tennessenoid A, an Unprecedented Steroid−Sorbicillinoid Adduct From the Marine-Derived Endophyte of Aspergillus sp. Strain 1022LEF. Front. Mar. Sci. 2022, 9, 923128. [Google Scholar] [CrossRef]
- Wei, M.; Huang, L.; Li, Q.; Qiao, X.; Zhao, Z.; Yin, J.; Zhang, Y. Spectasterols, Aromatic Ergosterols with 6/6/6/5/5, 6/6/6/6, and 6/6/6/5 Ring Systems from Aspergillus spectabilis. J. Nat. Prod. 2023, 86, 1385–1391. [Google Scholar] [CrossRef]
- Liu, Z.; Dong, Z.; Qiu, P.; Wang, Q.; Yan, J.; Lu, Y.; She, Z. Two new bioactive steroids from a mangrove-derived fungus Aspergillus sp. Steroids 2018, 140, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zheng, Y.; Fu, A.; Wei, M.; Kang, X.; Chen, C.; Zhang, Y. 30-norlanostane triterpenoids and steroid derivatives from the endophytic fungus Aspergillus nidulans. Phytochemistry 2022, 201, 113257. [Google Scholar] [CrossRef]
- Yu, S.; Zhu, Y.X.; Peng, C.; Li, J. Two new sterol derivatives isolated from the endophytic fungus Aspergillus tubingensis YP-2. Nat. Prod. Res. 2021, 35, 3277–3284. [Google Scholar] [CrossRef]
- Cao, V.A.; Kwon, J.H.; Kang, J.S.; Lee, H.S.; Heo, C.S.; Shin, H.J. Aspersterols A–D, ergostane-type sterols with an unusual unsaturated side chain from the deep-sea-derived fungus Aspergillus unguis. J. Nat. Prod. 2022, 85, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Khmel, O.O.; Yurchenko, A.N.; Trinh, P.T.H.; Ngoc, N.T.D.; Trang, V.T.D.; Khanh, H.H.N.; Yurchenko, E.A. Secondary Metabolites of the Marine Sponge-Derived Fungus Aspergillus subramanianii 1901NT-1.40. 2 and Their Antimicrobial and Anticancer Activities. Marine Drugs 2025, 23, 353. [Google Scholar] [CrossRef]
- Tong, Z.; Xiao, X.; Lu, Y.; Zhang, Y.; Hu, P.; Jiang, W.; Hu, L. New metabolites from Aspergillus ochraceus with antioxidative activity and neuroprotective potential on H2O2 insult SH-SY5Y cells. Molecules 2021, 27, 52. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Wang, M.M.; Tian, J.; Hu, B.H.; Lu, Y.N.; Yang, C.M.; Feng, R.Z. Chemical constituents from the endophytic fungus Aspergillus sp. S3 of Hibiscus tiliaceus. J. Asian Nat. Prod. Res. 2025, 27, 292–300. [Google Scholar] [CrossRef]
- Abdel-Razek, A.S.; El-Ghonemy, D.H.; Shaaban, M. Production and purification of bioactive compounds with potent antimicrobial activity from a novel terrestrial fungus Aspergillus sp. DHE 4. Biocatal. Agricul. Biotechnol. 2020, 28, 101726. [Google Scholar] [CrossRef]
- Elsbaey, M.; Ibrahim, M.A.; Hegazy, M.E.F. Versisterol, a new endophytic steroid with 3CL protease inhibitory activity from Avicennia marina (Forssk.) Vierh. RSC advances 2022, 12, 12583–12589. [Google Scholar] [CrossRef]
- Ying, Y.; Chen, X.; Zhan, S.; Yang, S.; Liu, J.; Shan, W.; Wang, R. Ganodermanic acid, a new steroidal compound derived from the marine fungus Aspergillus sp. ZJUT223. Nat. Prod. Res. 2025, 21, 1–8. [Google Scholar] [CrossRef]
- Weng, J.; Li, S.; Ma, R.; Shi, Q.; Meng, X.; Zhou, G.; Li, H. New secondary metabolites from the soil-derived Aspergillus versicolor QC812. Steroids 2025, 109629. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.M.; Zhang, Y.W.; Feng, F.J.; Huang, G.B.; Lv, Y.H.; Zhang, Z.Y.; Ding, L.J. Antibacterial oxygenated ergostane-type steroids produced by the marine sponge-derived fungus Aspergillus sp. J. Asian Nat. Prod. Res. 2024, 26, 548–554. [Google Scholar] [CrossRef]
- Peterson, S.W. Aspergillus and Penicillium identification using DNA sequences: barcode or MLST? Applied Microbiol. Biotechnol. 2012, 95, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Susca, A. Penicillium species and their associated mycotoxins. Mycotoxigenic Fungi: Methods and Protocols 2016, 11, 107–119. [Google Scholar] [CrossRef]
- Visagie, C.M.; Yilmaz, N.; Kocsubé, S.; Frisvad, J.C.; Hubka, V.; Samson, R.A.; Houbraken, J. A review of recently introduced Aspergillus, Penicillium, Talaromyces and other Eurotiales species. Studies in Mycology 2024, 107, 1–66. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Studies in Mycology 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Ashtekar, N.; Anand, G.; Thulasiram, H.V.; Rajeshkumar, K.C. Genus Penicillium: advances and application in the modern era. New and Future Developments in Microbial Biotechnology and Bioengineering 2021, 201–213. [Google Scholar] [CrossRef]
- Kozlovskii, A.G.; Zhelifonova, V.P.; Antipova, T.V. Fungi of the genus Penicillium as producers of physiologically active compounds. Applied Biochem. Microbiol. 2013, 49, 1–10. [Google Scholar] [CrossRef]
- Chávez, R.; Bull, P.; Eyzaguirre, J. The xylanolytic enzyme system from the genus Penicillium. J. Biotechnol. 2006, 123, 413–433. [Google Scholar] [CrossRef]
- Meskini, M.; Rami, M.R.; Tavakoli, R.; Salami, M. Molecular Markers in Diagnostics of Fungi and Fungal Mycotoxins: A Narrative Review. J. Infection Public Health 2025, 103073. [Google Scholar] [CrossRef]
- Mady, M.; Haggag, E. Review on fungi of genus Penicillium as a producers of biologically active polyketides. J. Advanced Pharmacy Res. 2020, 4, 33–45. [Google Scholar] [CrossRef]
- Chemmam, D.A.; Bourzama, G.; Chemmam, M. The genus Penicillium: Ecology, secondary metabolites and biotechnological applications. Biol. Aujourd’hui 2025, 219, 69–87. [Google Scholar] [CrossRef]
- Rodrigues, F.Z.; Marcelão, C.V.P.; Silva, J.J.D.; Taniwaki, M.H. Fungi and mycotoxins in Brazilian artisanal cheese. Brazilian J. Food Technol. 2025, 28, e2024131. [Google Scholar] [CrossRef]
- Alghamdi, S.S.; Alamre, J.H.; Alsubait, A.; Alanzi, A.R.; Aldawish, B.S.; Althobiti, F.; Mohammed, A.E. Bioactive Compounds From Saudi Arabian Fungi: A Systematic Review of Anticancer Potential. Clinical Pharmacology: Advances and Applications 2025, 291–304. [Google Scholar] [CrossRef]
- Dai, L.T.; Yang, L.; Wang, Z.P.; Guo, J.C.; Ma, Q.Y.; Xie, Q.Y.; Zhao, Y.X. Persteroid, a new steroid from the marine-derived fungus Penicillium sp. ZYX-Z-143. Nat. Prod. Res. 2024, 1–8. [Google Scholar] [CrossRef]
- Zhang, F.Z.; Li, X.M.; Meng, L.H.; Wang, B.G. A new steroid with potent antimicrobial activities and two new polyketides from Penicillium variabile EN-394, a fungus obtained from the marine red alga Rhodomela confervoides. J. Antibiot. 2024, 77, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.W.; Cui, C.B.; Li, C.W.; Wu, C.J. Three new and eleven known unusual C25 steroids: Activated production of silent metabolites in a marine-derived fungus by chemical mutagenesis strategy using diethyl sulphate. Marine Drugs 2014, 12, 1545–1568. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, C.; Cai, J.; Chen, Y.; Zhu, Y.; Yang, B.; Tao, H. Two C23-Steroids and a New Isocoumarin Metabolite from Mangrove Sediment-Derived Fungus Penicillium sp. SCSIO 41429. Marine Drugs 2024, 22, 393. [Google Scholar] [CrossRef]
- Lin, L.B.; Wang, M.N.; Hu, J.Y.; Han, R.; Yang, X.; Shi, W.; Xiao, J. An Unusual Meroterpenoid and Two New Steroids From Fungus Penicillium fellutanum and Their Bioactivities. Chem. Biodiver 2025, 22, e202403443. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Song, W.; Li, H.; Cao, L. Peniciloxatone A, a New Polyoxygenated Ergostane Steroid Isolated from the Marine Alga-Sourced Fungus Penicillium oxalicum 2021CDF-3. Rec. Nat. Prod. 2024, 18, 699–704. [Google Scholar] [CrossRef]
- Ying, Z.; Li, X.M.; Wang, B.G.; Li, H.L.; Meng, L.H. Rubensteroid A, a new steroid with antibacterial activity from Penicillium rubens AS-130. J. Antibiot. 2023, 76, 563–566. [Google Scholar] [CrossRef]
- Lindsay, C.A.; Tan, C.Y.; Krishnan, D.; Uchenik, D.; Eugenio, G.D.A.; Salinas, E.D.; Rakotondraibe, H.L. Steroids and Epicoccarines from Penicillium aurantiancobrunneum. Phytochemistry Letters 2024, 63, 79–86. [Google Scholar] [CrossRef]
- Salendra, L.; Lin, X.; Chen, W.; Pang, X.; Luo, X.; Long, J.; Yang, B. Cytotoxicity of polyketides and steroids isolated from the sponge-associated fungus Penicillium citrinum SCSIO 41017. Nat. Prod. Res. 2021, 35, 900–908. [Google Scholar] [CrossRef]
- Hou, Z.M.; Yu, S.Q.; Tao, M.; Xia, C.B.; Xia, Y.L.; Wu, X.F.; Dong, C.Z. New Purinyl-Steroid and Other Constituents from the Marine Fungus Penicillium brefeldianum ABC190807: Larvicidal Activities against Aedes aegypti. J. Chem. 2021, 2021, 6640552. [Google Scholar] [CrossRef]
- Lindsay, C.A.; Kinghorn, A.D.; Rakotondraibe, H.L. Bioactive and unusual steroids from Penicillium fungi. Phytochemistry 2023, 209, 113638. [Google Scholar] [CrossRef]
- Xue, J.; Wu, P.; Xu, L.; Wei, X. Penicillitone, a potent in vitro anti-inflammatory and cytotoxic rearranged sterol with an unusual tetracycle core produced by Penicillium purpurogenum. Organic Letters 2014, 16, 1518–1521. [Google Scholar] [CrossRef]
- Igarashi, Y.; Sekine, A.; Fukazawa, H.; Uehara, Y.; Yamaguchi, K.; Endo, Y.; Oki, T. Anicequol, a novel inhibitor for anchorage-independent growth of tumor cells from Penicillium aurantiogriseum Dierckx TP-F0213. J. Antibiot. 2002, 55, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.S.; Li, X.M.; Li, C.S.; Proksch, P.; Wang, B.G. Penicisteroids A and B, antifungal and cytotoxic polyoxygenated steroids from the marine alga-derived endophytic fungus Penicillium chrysogenum QEN-24S. Bioorg. Med. Chem. Letters 2011, 21, 2894–2897. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tian, L.; Huang, J.; Li, W.; Pei, Y.H. Cytotoxic sterols from marine-derived fungus Pennicillium sp. Nat. Prod. Res. 2006, 20, 381–384. [Google Scholar] [CrossRef]
- Deng, M.; Xiao, Y.; Wang, S.; Zhang, M.; Qiao, Y.; Huang, S.; Zhou, X. Penicimides A and B, two novel diels–alder [4+2] cycloaddition ergosteroids from Penicillium herquei. Bioorg. Chem. 2024, 143, 107025. [Google Scholar] [CrossRef]
- Zhang, Y.; Kang, J.; Zhou, Q.; Chen, M.; Zhang, J.; Shi, Z.; Zhang, Y. Discovery of 23, 24-diols containing ergosterols with anti-neuroinflammatory activity from Penicillium citrinum TJ507. Bioorg. Chem. 2024, 150, 107575. [Google Scholar] [CrossRef]
- Chen, T.; Yang, W.C.; Li, T.B.; Yin, Y.H.; Liu, Y.F.; Wang, B.; She, Z.G. Hemiacetalmeroterpenoids A–C and astellolide Q with antimicrobial activity from the marine-derived fungus Penicillium sp. N-5. Mar. Drugs 2022, 20, 514. [Google Scholar] [CrossRef]
- Hu, J.; Qin, D. Diversity and Biological Activity of Secondary Metabolites Produced by the Endophytic Fungus Penicillium ochrochlorae. Fermentation 2025, 11, 394. [Google Scholar] [CrossRef]
- Li, C.; He, Z.; Wen, Z.; Sun, G.; Tang, X.; Yin, T.; Cai, L. Expansinine, a novel indole alkaloid–ergosteroid conjugate from the endophytic fungus Penicillium expansum of Aconitum carmichaelii. RSC Advances 2025, 15, 14283–14288. [Google Scholar] [CrossRef]
- Li, Z.; Qin, C.L.; Zhou, T.; Li, W.P.; Hu, J.Y.; Zhou, Y.H.; Ruan, H.L. Scabrosteroids A–D, Four Pyrrolidinone–Ergosterol Heterodimers from Penicillium scabrosum FXI744. Org. Lett. 2025, 27(24), 6434–6438. [Google Scholar] [CrossRef]
- Jie-Yi, L.O.N.G.; Jun-Feng, W.A.N.G.; Sheng-Rong, L.I.A.O.; Xiu-Ping, L.I.N. Four new steroids from the marine soft coral-derived fungus Penicillium sp. SCSIO41201. Chinese J. Nat. Med. 2020, 18, 250–255. [Google Scholar] [CrossRef]
- Tang, J.W.; Kong, L.M.; Zu, W.Y.; Hu, K.; Li, X.N.; Yan, B.C.; Puno, P.T. Isopenicins A–C: two types of antitumor meroterpenoids from the plant endophytic fungus Penicillium sp. sh18. Organic Letters 2019, 21, 771–775. [Google Scholar] [CrossRef]
- La Kim, E.; Li, J.L.; Hong, J.; Yoon, W.D.; Kim, H.S.; Liu, Y.; Jung, J.H. An unusual 1 (10→ 19) abeo steroid from a jellyfish-derived fungus. Tetrahedron Lett. 2016, 57, 2803–2806. [Google Scholar] [CrossRef]
- Li, J.; Chen, C.; Fang, T.; Wu, L.; Liu, W.; Tang, J.; Long, Y. New steroid and isocoumarin from the mangrove endophytic fungus Talaromyces sp. SCNU-F0041. Molecules 2022, 27, 5766. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Zhang, Y.; Wang, C.; Si, S.; Liu, X.; Che, Y. Altersteroids A–D, 9, 11-Secosteroid-Derived γ-Lactones from an Alternaria sp. J. Nat. Prod. 2023, 86, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Liu, J.; Lin, L.B.; Hu, J.Y.; Wu, G.W.; Fan, P.H.; Wang, X.L. Acrocalysterols A and B, two new steroids from endophytic fungus Acrocalymma sp. Phytochemistry Letters 2022, 48, 77–80. [Google Scholar] [CrossRef]
- Azhari, A.; Naini, A.A.; Harneti, D.; Wulandari, A.P.; Mulyani, Y.; Purbaya, S.; Supratman, U. New steroid produced by Periconia pseudobyssoides K5 isolated from Toona sureni (Meliaceae) and its heme polymerization inhibition activity. J. Asian Nat. Prod. Res. 2023, 25, 1117–1124. [Google Scholar] [CrossRef]
- Elsebai, M.F.; Kehraus, S.; König, G.M. Caught between triterpene-and steroid-metabolism: 4α-Carboxylic pregnane-derivative from the marine alga-derived fungus Phaeosphaeria spartinae. Steroids 2013, 78, 880–883. [Google Scholar] [CrossRef]
- Hu, Z.; Wu, Y.; Xie, S.; Sun, W.; Guo, Y.; Li, X.N.; Zhang, Y. Phomopsterones A and B, two functionalized ergostane-type steroids from the endophytic fungus Phomopsis sp. TJ507A. Org. Lett. 2017, 19, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Z.; Han, K.Y.; Li, Z.H.; Feng, T.; Chen, H.P.; Liu, J.K. Cytotoxic ergosteroids from the fungus Stereum hirsutum. Phytochemistry Lett. 2019, 30, 143–149. [Google Scholar] [CrossRef]
- Cao, S.; Ross, L.; Tamayo, G.; Clardy, J. Asterogynins: secondary metabolites from a Costa Rican endophytic fungus. Org. Lett. 2010, 12, 4661–4663. [Google Scholar] [CrossRef]
- Mo, S.; Yang, J.; Zhao, J.; Wang, J.; Zhang, Y.; Hu, Z. Trichosterol A, a unique 6/6/6/5/6-fused steroid-alkaloid hybrid with bioherbicidal activity from Trichoderma koningiopsis. Org. Biomol. Chem. 2025, 23, 10280–10284. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Q.; Yao, F.H.; Luo, L.X.; Wu, Y.H.; Qi, S.H. Microascusteroids A and B, two 5, 6-seco-9, 10-seco steroids with a C-ring rearranged ergostane skeleton from the marine-derived fungus Microascus sp. SCSIO 41821. Fitoterapia 2025, 183, 106537. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Du, H.F.; Zhang, Y.H.; Liu, Z.Q.; Qi, X.Q.; Luo, D.Q.; Cao, F. Chaeglobol A, an unusual octocyclic sterol with antifungal activity from the marine-derived fungus Chaetomium globosum HBU-45. Chinese Chem. Lett. 2025, 36, 109858. [Google Scholar] [CrossRef]
- Chen, C.; Cai, J.; Yang, C.; Tao, H.; Tang, L.; Liu, Y.; Zhou, X. Striasteroids A–C, Three Hybrid Steroids with Neuraminidase Inhibitory Activities from a Marine-Derived Striaticonidium cinctum. Org. Lett. 2025, 27, 3737–3741. [Google Scholar] [CrossRef]
- Amagata, T.; Tanaka, M.; Yamada, T.; Doi, M.; Minoura, K.; Ohishi, H.; Numata, A. Variation in cytostatic constituents of a sponge-derived Gymnascella dankaliensis by manipulating the carbon source. J. Nat. Prod. 2007, 70, 1731–1740. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, X.; Zhang, X.; Liu, Y.; Li, Q.; Mao, L.; Zhu, H. Bipolarsterol A, a steroid with an unprecedented 6/5/6/5 carbon skeleton from the phytopathogenic fungus Bipolaris oryzae. Org. Chem. Frontiers 2025, 12, 1474–1484. [Google Scholar] [CrossRef]
- Zhao, Z.Z.; Chen, H.P.; Wu, B.; Zhang, L.; Li, Z.H.; Feng, T.; Liu, J.K. Matsutakone and matsutoic acid, two (nor) steroids with unusual skeletons from the edible mushroom Tricholoma matsutake. J. Org. Chem. 2017, 82, 7974–7979. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.A.; Li, H.F.; Huang, S.S.; Jin, T.; Xie, W.Z.; He, J.; Feng, T. Cordycepsterols A–C, Anti-Inflammatory C30 Ergosterols with a 6/6/6/5/6-Fused Ring System from Cordyceps militaris. J. Nat. Prod. 2025, 88, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhu, J.; Mu, R.; Wang, C.; Sun, Y.; Qian, B.; Chen, Y. Metabolites with Anti-Inflammatory Activities Isolated from the Mangrove Endophytic Fungus Dothiorella sp. ZJQQYZ-1. Microorganisms 2025, 13, 890. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
New steroids produced by Ascomycetes of the genus Aspergillus.
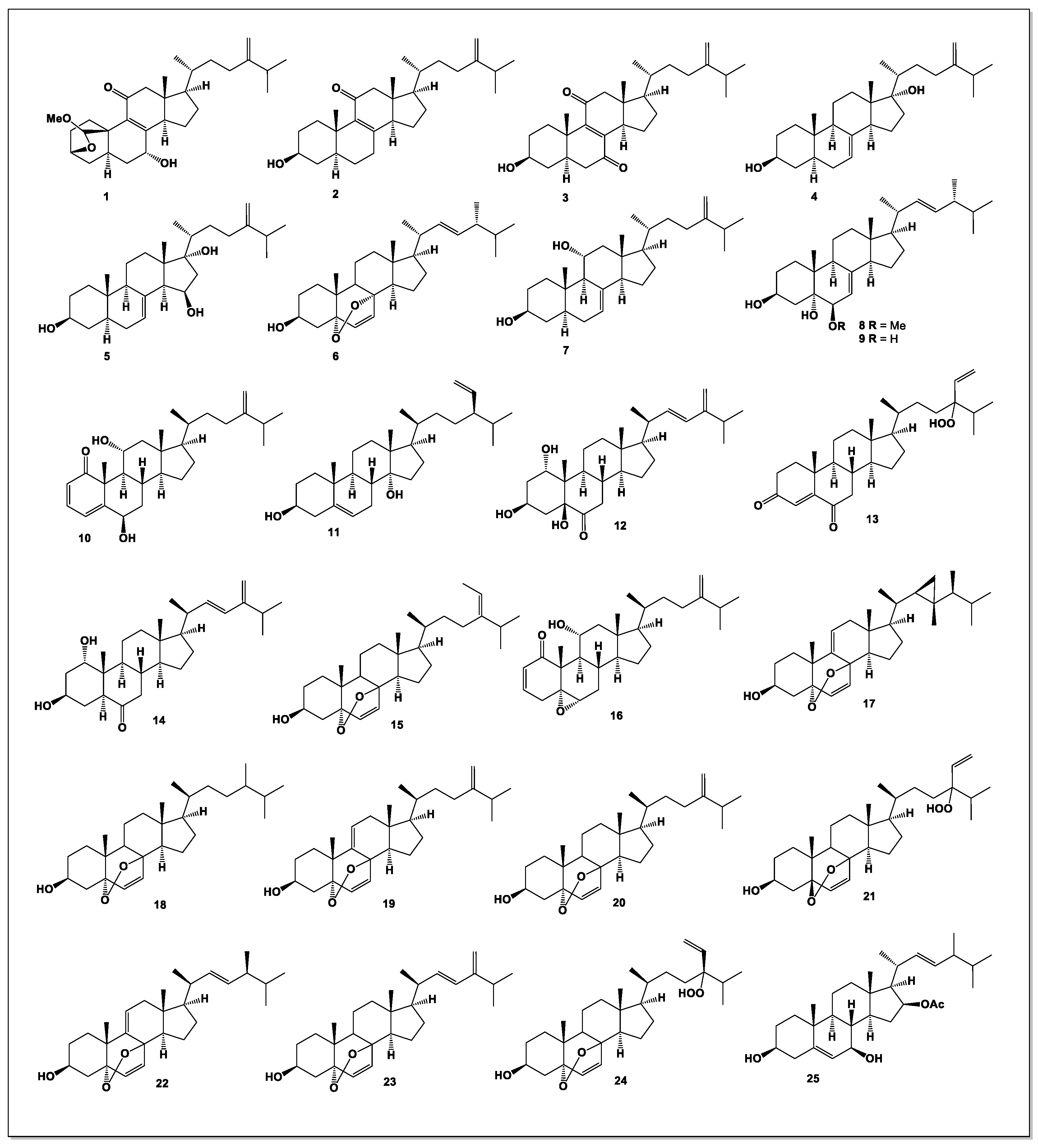
Figure 2.
Bioactive rare and uncommon steroids derived from fungal endophytes.
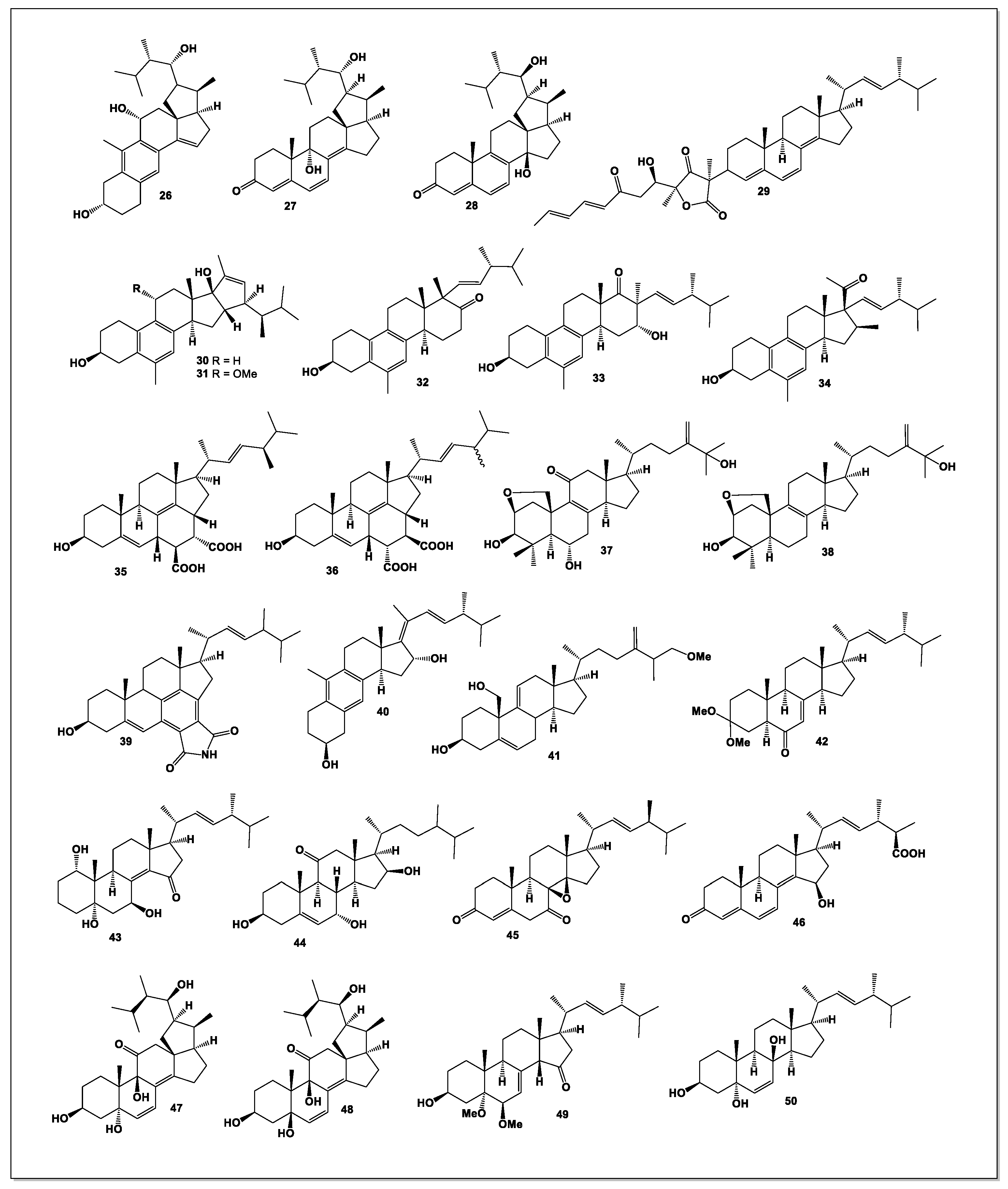
Figure 3.
Fungal endophytes related to the genus Aspergillus, which are producers of active steroids, are found in a variety of organisms, which are presented below. a) Pseudostellaria heterophylla (Tai Zi Shen or False Starwort) is a vital Chinese medicinal herb, known as “lung ginseng,” prized for its mild, ginseng-like properties, strengthening spleen, boosting qi, moistening lungs, and supporting immunity, often used for fatigue, poor appetite, and post-illness weakness, with its roots containing bioactive compounds like steroids and saponins, cultivated in China and used in health foods. b) Cliona is a genus of boring sponges, known for their ability to excavate tunnels and chambers within calcium carbonate substrates, such as limestone, coral, and mollusk shells. They are found worldwide and play a significant role in marine ecosystems by recycling calcium carbonate and shaping benthic habitats. c) Hibiscus tiliaceus, or Beach Hibiscus, is a versatile plant used in traditional medicine for fevers, coughs, chest congestion, diarrhea, and skin issues like abscesses, with leaves, flowers, bark, and sap all utilized for ailments like infections, inflammation, and as laxatives, possessing antioxidant and antimicrobial properties. d) The fruits of Avicennia marina (Grey Mangrove) are green, oval capsules, about 20-25 mm in diameter, with a short beak, developing from creamy-yellow flowers and often germinating on the tree before falling to be dispersed by water. They are rich in nutrients, offering high caloric value and essential proteins, fats, and carbs, and are used traditionally for food and medicine due to their strong antioxidant properties and bioactive compounds, though high doses might have mild effects on liver/kidneys.
Figure 3.
Fungal endophytes related to the genus Aspergillus, which are producers of active steroids, are found in a variety of organisms, which are presented below. a) Pseudostellaria heterophylla (Tai Zi Shen or False Starwort) is a vital Chinese medicinal herb, known as “lung ginseng,” prized for its mild, ginseng-like properties, strengthening spleen, boosting qi, moistening lungs, and supporting immunity, often used for fatigue, poor appetite, and post-illness weakness, with its roots containing bioactive compounds like steroids and saponins, cultivated in China and used in health foods. b) Cliona is a genus of boring sponges, known for their ability to excavate tunnels and chambers within calcium carbonate substrates, such as limestone, coral, and mollusk shells. They are found worldwide and play a significant role in marine ecosystems by recycling calcium carbonate and shaping benthic habitats. c) Hibiscus tiliaceus, or Beach Hibiscus, is a versatile plant used in traditional medicine for fevers, coughs, chest congestion, diarrhea, and skin issues like abscesses, with leaves, flowers, bark, and sap all utilized for ailments like infections, inflammation, and as laxatives, possessing antioxidant and antimicrobial properties. d) The fruits of Avicennia marina (Grey Mangrove) are green, oval capsules, about 20-25 mm in diameter, with a short beak, developing from creamy-yellow flowers and often germinating on the tree before falling to be dispersed by water. They are rich in nutrients, offering high caloric value and essential proteins, fats, and carbs, and are used traditionally for food and medicine due to their strong antioxidant properties and bioactive compounds, though high doses might have mild effects on liver/kidneys.

Figure 4.
Steroidal hormones derived from Penicillium species.
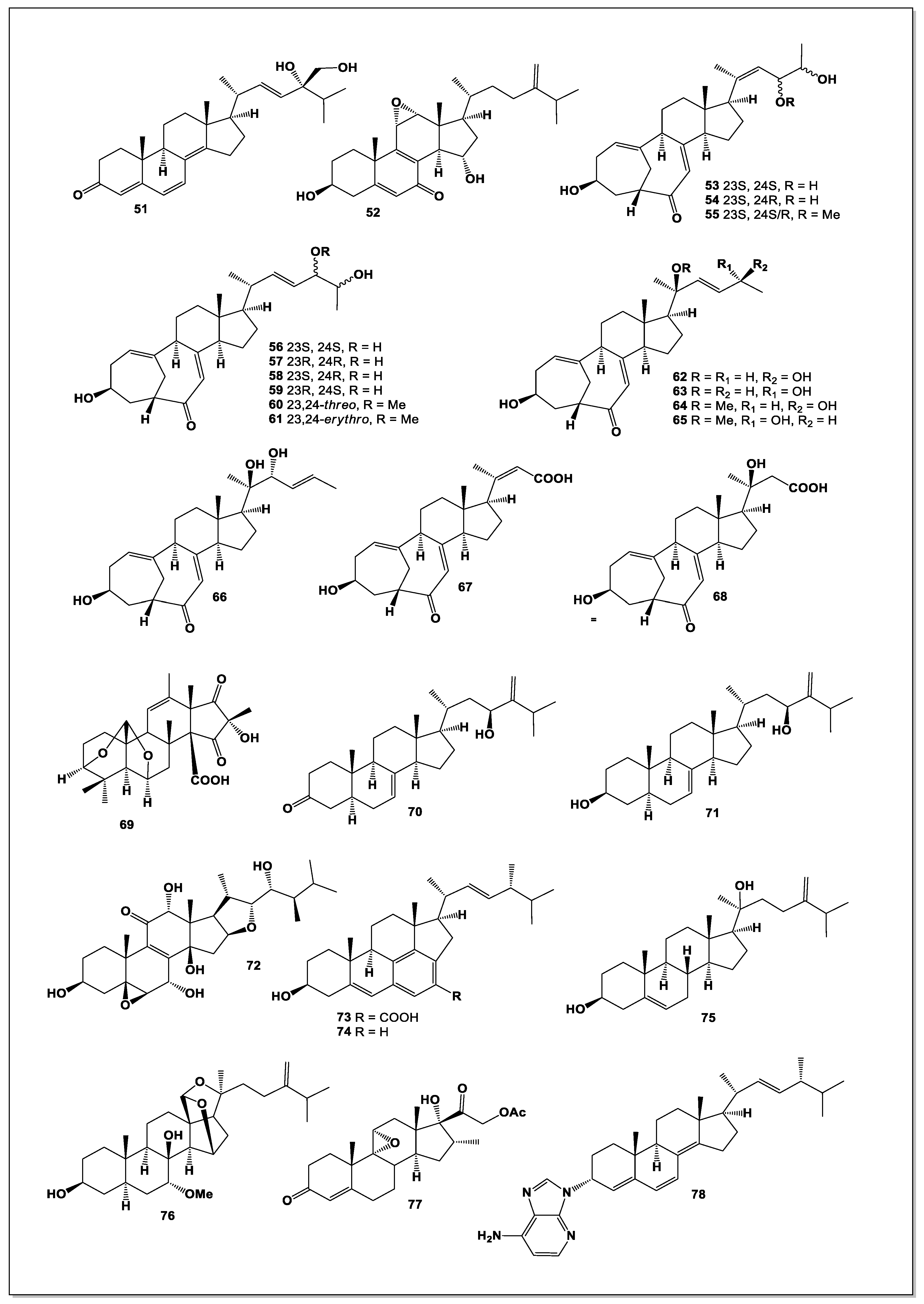
Figure 5.
Steroid and their unusual derivatives derived from Penicillium species.
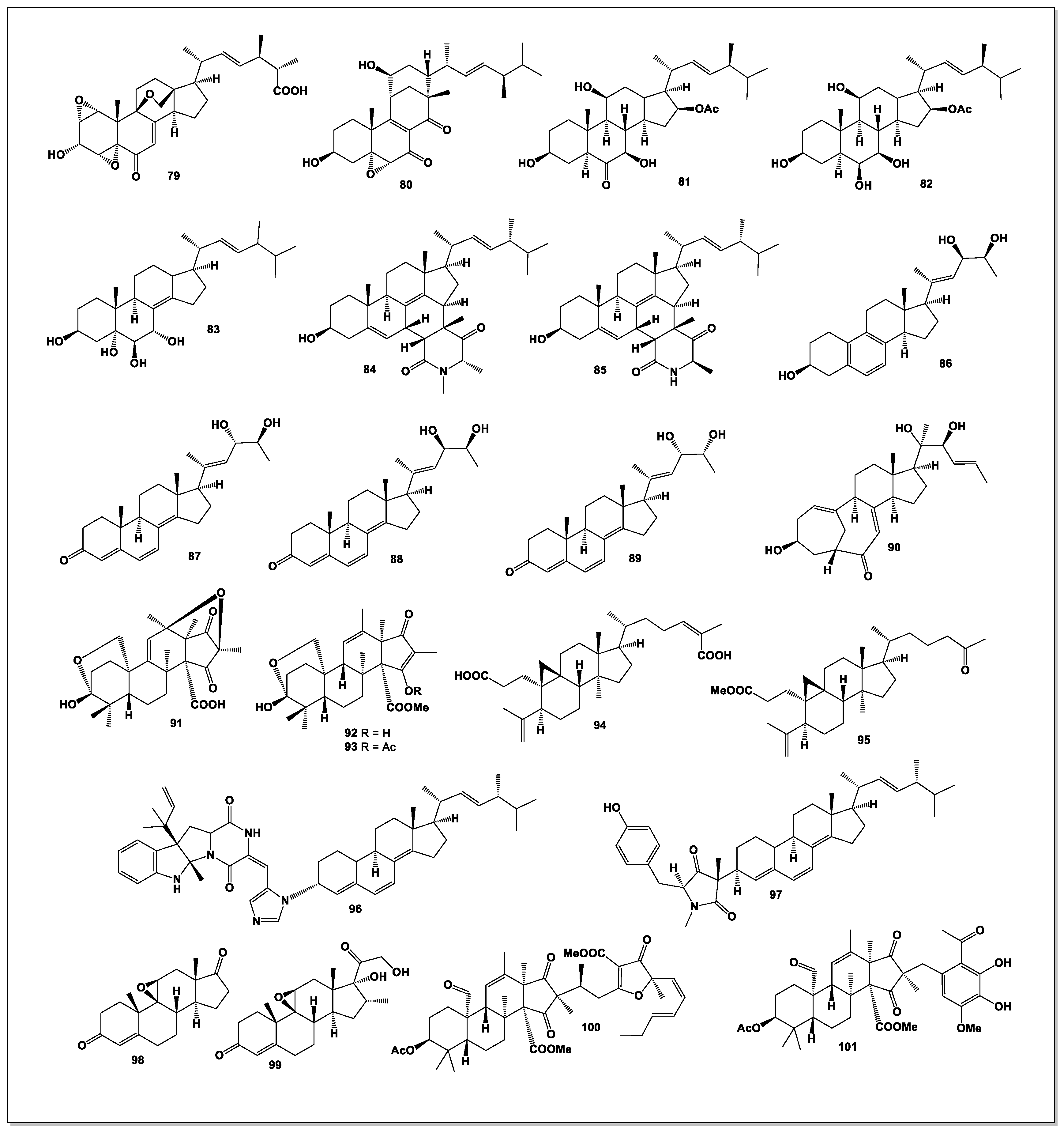
Figure 6.
Fungal endophytes related to the genus Penicillium, which are producers of active steroids, are found primarily in marine red algae, sea mosses, and sea sponges, some of which are presented below. a) Rhodomela confervoides is a common species of brownish-red, bushy marine red algae (seaweed) that is widely distributed in intertidal pools. It belongs to the family Rhodomelaceae. b) The genus Laurencia comprises approximately 130 to 137 taxonomically accepted species of marine red algae worldwide. These species are widely distributed in tropical, subtropical, and temperate waters and are known for producing a large number of secondary metabolites with various biological activities. c) South Pole (Antarctic) sea moss refers to the unique, hardy mosses that thrive in Antarctica’s extreme coastal environments, acting as vital mini-ecosystems in the icy desert, growing slowly in green carpets or banks, absorbing sun for warmth, drying out to survive winter, and providing habitat for tiny creatures like tardigrades, with their growth patterns also serving as key indicators of climate change. d) The genus Callyspongia encompasses a wide variety of marine sponges found in tropical coral reef ecosystems worldwide, known for their diverse forms and colors. They are filter feeders and play an important ecological role in reef communities.
Figure 6.
Fungal endophytes related to the genus Penicillium, which are producers of active steroids, are found primarily in marine red algae, sea mosses, and sea sponges, some of which are presented below. a) Rhodomela confervoides is a common species of brownish-red, bushy marine red algae (seaweed) that is widely distributed in intertidal pools. It belongs to the family Rhodomelaceae. b) The genus Laurencia comprises approximately 130 to 137 taxonomically accepted species of marine red algae worldwide. These species are widely distributed in tropical, subtropical, and temperate waters and are known for producing a large number of secondary metabolites with various biological activities. c) South Pole (Antarctic) sea moss refers to the unique, hardy mosses that thrive in Antarctica’s extreme coastal environments, acting as vital mini-ecosystems in the icy desert, growing slowly in green carpets or banks, absorbing sun for warmth, drying out to survive winter, and providing habitat for tiny creatures like tardigrades, with their growth patterns also serving as key indicators of climate change. d) The genus Callyspongia encompasses a wide variety of marine sponges found in tropical coral reef ecosystems worldwide, known for their diverse forms and colors. They are filter feeders and play an important ecological role in reef communities.

Figure 7.
Steroids and their unusual derivatives derived from miscellaneous microorganisms.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



