
Submitted:
07 December 2025
Posted:
08 December 2025
You are already at the latest version
Abstract
The microbiota–gut–brain axis (GBA) is a bidirectional communication system integrating neural, endocrine, immune, and metabolic pathways to coordinate gastrointestinal and central nervous system (CNS) function. As the majority of human microbes reside in the gastrointestinal tract, their metabolic activity and immunomodulatory roles are essential for systemic homeostasis, cognitive processes, mood regulation, and neuroprotection. Increasing evidence highlights the vagus nerve as a primary conduit of microbiota–brain signaling, with germ-free mice demonstrating reduced vagal tone that is restored following microbial colonization. Key microbial metabolites—including short-chain fatty acids (SCFAs), bile acids, indoles, and neurotransmitter precursors—activate vagal afferents, regulate neuroimmune responses, and influence microglial maturation and synaptic function. Disruption of microbial community structure, or dysbiosis, is now recognized as a contributing factor in numerous neurological and psychiatric disorders. Neurodegenerative diseases such as Alzheimer’s disease (AD) and Parkinson’s disease (PD) exhibit distinct microbial alterations, chronic neuroinflammation, and impaired gut and blood–brain barrier integrity. PD patients, for example, consistently show increased Enterobacteriaceae and reduced Prevotellaceae. Dysbiosis also contributes to mood disorders by disrupting microbial production of GABA, serotonin, and tryptophan metabolites. Although microbial biomarkers vary across studies due to disease heterogeneity, integrative multi-omics approaches have begun identifying shared metabolic patterns and therapeutic targets. Microbiota-derived metabolites such as butyrate show neuroprotective and anti-inflammatory effects, while fecal microbiota transplantation (FMT), prebiotics, probiotics, and next-generation probiotics (NGPs) are emerging as promising modulators of the GBA. Advances in metagenomics, metabolomics, and machine-learning–based patient profiling further support the development of personalized, microbiome-driven interventions for neurological disorders.
Keywords:
gut–brain axis (GBA)
; gut microbiota
; neurodegenerative disorders
; dysbiosis
; short-chain fatty acids (SCFAs)
; vagus nerve
; neuroinflammation
; Parkinson’s disease (PD)
; Alzheimer’s disease (AD)
; microglia
; fecal microbiota transplantation (FMT)
1. Introduction
1.1. Overview of Neurodegenerative Diseases (NDDs)
1.1.1. Definition and Epidemiology
Neurodegenerative diseases (NDDs) represent a growing global health crisis, driven by the aging population, and the combination of environmental factors and changes in lifestyle. These debilitating conditions—including Alzheimer’s disease (AD), Parkinson’s disease (PD), and Multiple System Atrophy (MSA)—are characterized by the progressive loss of structure or function of neurons and impose a significant burden on healthcare systems worldwide [1]. For instance, a comprehensive analysis of neurological disorders in the United States from 1990 to 2017 revealed alarming trends in morbidity and mortality associated with NDDs [2]. The prevalence of AD, for example, is projected to triple globally by 2050, while PD cases are expected to double by 2030. These statistics underscore the urgent need for innovative approaches to understand, prevent, and treat these debilitating conditions [3].
The rising incidence of NDDs highlights the inadequacy of current therapeutic strategies, many of which focus narrowly on managing symptoms rather than addressing underlying causes. This gap in understanding has spurred intense interest in novel, non-traditional pathways that link the gastrointestinal system to the central nervous system (CNS) [4]. Recent advances suggest that the gut microbiota—the complex community of microorganisms residing in the digestive tract—plays a pivotal and previously overlooked role in modulating NDD pathogenesis, offering new avenues for early intervention and precision medicine [5].
1.1.2. Key Pathophysiological Hallmarks
At the core of NDD progression lies neuroinflammation, a chronic inflammatory process within the CNS that contributes to neuronal damage and loss. This inflammatory cascade is a shared hallmark of diseases like AD, PD, and MSA. However, emerging evidence points to the gut–brain axis (GBA) as a critical mediator of this neuroinflammation and a key player in the ‘synaptic’ communication between the gut and the brain [6]. Dysregulation of the GBA can exacerbate neurodegeneration by promoting systemic inflammation, oxidative stress, mitochondrial dysfunction, and impaired neural signaling. For instance, microbial metabolites produced in the gut, such as short-chain fatty acids (SCFAs), have been shown to modulate microglial activity and influence the expression of pro-inflammatory cytokines (Figure 1), thereby shaping the CNS microenvironment [7].
Likewise, dysbiosis—a term used to describe an imbalance or disruption in the gut microbiota—has been increasingly implicated in the pathogenesis of NDDs. Dysbiosis can lead to increased intestinal permeability (“leaky gut”), enabling bacterial products like lipopolysaccharides (LPS) to enter systemic circulation [8]. These microbial-derived factors activate Toll-like receptors (TLRs) on immune cells, triggering inflammatory pathways that ultimately affect brain health. Understanding the direct and indirect interplay between gut dysbiosis, the GBA and neuroinflammation is therefore essential for unveiling the synapses underlying NDDs and developing effective, targeted therapies [9].
1.1.3. Specific NDDs Associated with Microbiome Dysbiosis
Recent research has established a strong correlation between gut microbiota abnormalities and the onset and progression of NDDs. Below are some examples of how dysbiosis influences specific neurodegenerative conditions:
- Parkinson’s Disease (PD): Altered gut microbiota composition has been linked to both motor and non-motor symptoms of PD. Studies suggest that microbial dysbiosis precedes the clinical manifestation of the disease, making it a potential biomarker for early detection [10]. Key findings include an increased abundance of Enterobacteriaceae, which is associated with inflammation and oxidative stress, and reduced populations of Prevotellaceae, which play a role in mucin production and gut barrier maintenance. These microbial imbalances contribute to neuroinflammation, alpha-synuclein aggregation, and the progression of PD [11].
- Alzheimer’s Disease (AD): Changes in gut microbial communities have been associated with amyloid-beta deposition and cognitive decline, implicating the gut microbiota in AD pathogenesis. Dysbiosis leads to reduced production of SCFAs, which are critical for maintaining gut barrier integrity and preventing the translocation of pro-inflammatory molecules like LPS into systemic circulation [12]. Elevated levels of LPS in the blood have been linked to neuroinflammation and amyloid-beta accumulation, suggesting that targeting the gut microbiome could mitigate AD progression [13,74].
- Multiple System Atrophy (MSA): Preliminary findings indicate that gut dysbiosis may contribute to neuroinflammation and autonomic dysfunction in MSA. While research in this area is still in its infancy, the observed microbial alterations provide valuable insights into the role of the gut-brain axis in this rare but devastating condition [14].
Taken together, the growing body of evidence underscores the gut microbiota’s pivotal role in the pathogenesis of NDDs. By identifying microbial signatures and their functional contributions, researchers can develop targeted interventions aimed at restoring gut homeostasis and mitigating neurodegeneration [9]. This represents a promising frontier for early diagnosis and personalized treatment strategies.
1.2. The Gut Microbiome: Definition and Importance
1.2.1. Definition of Microbiota and Microbiome
The human gut harbors a complex and dynamic ecosystem of microorganisms, collectively referred to as the microbiota (Figure 2). This community includes bacteria (the dominant members), archaea, fungi (primarily yeasts), viruses/phages, and protists [15]. The collective genetic material and functional potential of these microbes, including their metabolic products, constitute the microbiome a term often used interchangeably with microbiota in contemporary literature, but which technically refers to the entire habitat and its genetic capacity [16].
The gut microbiota, concentrated primarily in the colon, is one of the most biodiverse ecosystems in the human body, housing approximately 1013 microbial cells per gram of intestinal content. Due to its profound and essential influence on host physiology [17], it is frequently described as the “second genome”. Advances in metagenomics sequencing have revealed that the gut microbiome contains vastly more genes (an estimated 150 times more) than the human genome, highlighting its immense metabolic and functional potential through encoded enzymes crucial for processes like carbohydrate metabolism and vitamin synthesis [18].
1.2.2. Role in Homeostasis
Dysbiosis refers to a state of imbalance or disruption in the composition and function of the gut microbiota. Key features include reduced microbial diversity, a loss of beneficial microbial species (like SCFA producers), and the overgrowth of potentially pathogenic microbes [19]:
- Immune Regulation: The microbiota educates and modulates the immune system, promoting tolerance to commensal microbes while defending against pathogens. It also primes innate and adaptive immunity, ensuring a balanced response to infections and preventing autoimmune reactions [20].
- Metabolic Activities: Gut microbes facilitate the breakdown of complex carbohydrates, fiber, and other indigestible compounds, producing beneficial metabolites such as SCFAs. These metabolites serve as energy sources for colonocytes, regulate gene expression, and exert anti-inflammatory effects. Additionally, the microbiota synthesizes essential vitamins, including B vitamins and vitamin K, which are critical for various physiological processes [21].
- Colonization Resistance: By outcompeting harmful pathogens for nutrients and attachment sites, the gut microbiota provides a protective barrier against infections. This colonization resistance is particularly important for preventing the overgrowth of opportunistic pathogens like Clostridioides difficile [22].
- Energy Balance and Intestinal Homeostasis: The microbiota influences energy harvest, fat storage, and the maintenance of gut barrier integrity. It prevents systemic inflammation caused by bacterial translocation, thereby safeguarding overall health [23].
1.2.3. Dysbiosis
Dysbiosis refers to an imbalance or disruption in the composition and function of the gut microbiota. Key features of dysbiosis include reduced microbial diversity, loss of beneficial microbial species, and overgrowth of potentially pathogenic microbes. These disruptions can trigger a cascade of adverse effects, including chronic inflammation, metabolic dysfunction, and increased susceptibility to diseases [24]. Dysbiosis has been implicated in a wide range of conditions, from gastrointestinal disorders like irritable bowel syndrome (IBS) and inflammatory bowel disease (IBD) to systemic illnesses, including obesity, type 2 diabetes, cardiovascular disease, and neurodegenerative disorders [25].
For examples, in Table 1, and recent studies have shown that dysbiosis in individuals consuming a Western diet characterized by high fat, sugar, and processed foods—leads to reduced SCFA production and increased levels of trimethylamine-N-oxide (TMAO), a metabolite linked to cardiovascular disease. Similarly, antibiotic use has been shown to reduce microbial diversity, leading to long-term alterations in gut microbiota composition and function. These findings emphasize the importance of maintaining a healthy gut microbiome for overall well-being (26, 101).
1.3. The Gut–Brain Axis (GBA): Concept and Relevance
1.3.1. Conceptual Framework
The Gut–Brain Axis (GBA) represents a bidirectional communication network linking the gastrointestinal (GI) tract to the central nervous system (CNS). This intricate system integrates multiple pathways, including neural, endocrine, and immunological routes, to coordinate gut health with host physiology and neurological function [46].
- Neural Pathways: The vagus nerve serves as a primary conduit for transmitting signals between the gut and brain. Recent research has demonstrated that microbial metabolites such as SCFAs and bile acids activate vagal afferent neurons in a receptor-dependent manner, enabling the gut microbiome to modulate chemosensory signals transmitted to the brain [47].
- Endocrine Pathways: Hormones and neurotransmitters produced by gut microbes influence CNS activity. For example, serotonin—a neurotransmitter critical for mood regulation—is predominantly synthesized in the gut, where it interacts with enterochromaffin cells and vagal afferents [48].
- Immunological Pathways: Cytokines and other immune mediators modulate neuroinflammation and brain function. Dysbiosis can disrupt this balance, leading to systemic inflammation and neuroinflammation [49].
Additionally, the GBA connects the CNS, autonomic nervous system (ANS), enteric nervous system (ENS), and hypothalamic-pituitary-adrenal (HPA) axis, creating a comprehensive regulatory network.
1.3.2. Emerging Role
The GBA plays a pivotal role in maintaining systemic health and regulating neurological function. It influences processes such as satiety, digestion, mood, and behavior. Recent studies have highlighted the GBA’s involvement in psychiatric disorders, including anxiety and depression, as well as its contribution to neurodegenerative diseases [50]. For instance, dysbiosis-induced systemic inflammation has been linked to the development of depressive symptoms, while altered gut microbiota composition has been associated with cognitive decline in AD patients [51].
Understanding the mechanisms underlying antibiotic-induced dysbiosis and its systemic effects via the GBA is critical for developing novel therapeutic strategies. For example, probiotics, prebiotics, fecal microbiota transplantation (FMT), and dietary interventions have shown promise in restoring gut homeostasis and alleviating symptoms of NDDs [5,9,52].
1.3.3. Dysbiosis via the GBA
Alterations in the gut microbiota, whether due to antibiotics, diet, or environmental factors, can disrupt GBA signaling, leading to systemic inflammation and neuroinflammation. These changes have been linked to the pathophysiology of psychiatric and neurodegenerative disorders [53], emphasizing the importance of the gut microbiota in CNS health. By leveraging tools like microbiome profiling and multi-omics integration, researchers can identify microbial signatures and develop personalized interventions to target the gut-brain axis effectively (Table 2) [54].
2. Composition and Structure of the Gut Microbiome
The human gut microbiome—a term often used interchangeably with “microbiota” but technically referring to the collective genetic material of microorganisms residing in the gut—represents one of the most complex and dynamic ecosystems known to science. This intricate microbial community is not only vast in diversity and density but also plays a pivotal role in maintaining host health [54]. Recent advances in metagenomics, transcriptomics, and multi-omics integration have deepened our understanding of its structure, composition, and functional significance. Below, we synthesize current knowledge on the key components, anatomical distribution, variability, and factors influencing the gut microbiome [55].
2.1. Key Microbial Components and Genetic Scale
The microbial community in the gut consists of trillions of symbiotic microbial cells. This complex community includes not only bacteria, but also archaea (single-celled organisms without nuclei), fungi (mostly yeasts), viruses/phages, and protozoa/ microbial eukaryotes. The collective genetic material of these microbes (the microbiome) is immense, estimated to be 150 times greater than the human genome [56].
In terms of density:
- The human gut microbiota represents the highest species diversity and quantity of microorganisms in the body.
- The number of microbial cells in the gut is estimated to be 10–100 trillion.
- The microbial density in the colon reaches up to 1012 cells per gram of intestinal material [57].
2.2. Dominant Bacterial Phyla and Genera
While thousands of bacterial species exist in the gut, the community is dominated by a few major phyla, which are highly conserved across nearly all individuals.
| Phylum | General Characteristics and Abundance |
| Firmicutes | Along with Bacteroidetes, they make up more than 90% of the entire gut population. Firmicutes alone constitute approximately 70% of the total gut bacteria population [58]. |
| Bacteroidetes | The second predominant phylum, making up about 23% of the total gut bacteria. They are highly conserved across individuals. [59] |
The gut environment is also favorable to the growth of bacteria from other primary phyla, including Actinobacteria, Proteobacteria, Fusobacteria, Verrucomicrobia, and Cyanobacteria [60].
At the genus level, several groups are highly prevalent:
- Species belonging to the Firmicutes group, such as Ruminococcus, Eubacterium, and Clostridium, are highly prevalent.
- Other dominant genera in the large intestine include Bacteroides, Bifidobacterium, Propionibacterium, Peptostreptococcus, and Ruminococcus [61].
- The microbial environment is overwhelmingly dominated by anaerobic bacteria, accounting for about 99% of the bacteria in the gut [62].
2.3. Anatomical Distribution of Microbiota
The distribution, quantity, and specific type of microorganisms vary significantly throughout the gastrointestinal tract due to differing physiology, pH, substrate availability, and oxygen partial pressure.
| Location | Characteristics and Dominant Taxa |
| Stomach and Small Intestine | These areas have a lower species count and diversity compared to the colon. The most abundant phyla here tend to be Proteobacteria and Firmicutes, which can withstand lower pH and other factors. Commonly found genera include Prevotella, Veillonella, and Streptococcus [63]. |
| Large Intestine (Colon) | This region is home to the most dense and diverse microbial community. Anaerobic conditions here provide an ideal environment for anaerobic growth. Most microbial biomass and metabolism occur in the luminal contents of the large intestine [64]. |
2.4. Variability and Factors Influencing Composition
The gut microbiome is highly individualized, shaped by a complex interplay of intrinsic and extrinsic factors. Understanding this variability is critical for developing personalized therapeutic strategies.
- Inter-Individual Variability: Although the major phyla (Firmicutes and Bacteroidetes) are highly conserved in virtually all individuals, the relative proportions of these two phyla can vary dramatically among healthy people, ranging from 10% to 90%. The variation in flora between individuals is undeniable [65].
- Host Factors: The nature of the symbiotic relationship is intricately influenced by age, genetic makeup, the host’s immune status, and environmental factors.
- Diet: Diet is a significant factor in shaping the gut microbiota. Dietary choices, such as adherence to plant-based diets versus the Western diet, profoundly affect the community structure [66].
- Disease and Dysbiosis: Dysbiosis, characterized by a reduction in beneficial species or a decrease in overall diversity, is associated with the onset and progression of numerous diseases, including infectious diseases, metabolic disorders (like obesity and type 2 diabetes), autoimmune diseases, and cancer [67].
- Stability: While the structure and function of gut microbiota tend to be stable in adult humans, events such as antibiotic treatments and diseases can cause alterations. However, the notion that an individual’s microbiome will be universally stable is one of the common pitfalls in clinical expectations [68].
The genetic material contained within the bacterial community is specifically referred to as the microbiome. Advancements in tools like metagenomic sequencing allow for the genomic and functional analysis of this community, characterizing not only the taxonomy but also the functional potential encoded by these genes.
3. The Gut–Brain Axis: Communication Mechanisms
The Gut–Brain Axis (GBA) represents a sophisticated bidirectional communication network that intricately links the gastrointestinal (GI) tract to the central nervous system (CNS). This axis integrates multiple pathways, including neural, humoral (endocrine/metabolite), and immunological mechanisms, to coordinate gut health with host physiology and neurological function [80]. By bridging the gap between the gut microbiome and brain activity, the GBA has emerged as a pivotal player in understanding systemic health and disease pathogenesis. Recent advances have shed light on the molecular and physiological underpinnings of this axis, providing new insights into how microbial signals influence brain function and behavior [81].
3.1. Neural Pathways (Vagus Nerve and ENS)
The neural pathway provides rapid communication between the gut and the brain, utilizing the body’s nervous systems.
a. Vagus Nerve Function:
At the core of the GBA lies the vagus nerve, which serves as a critical conduit for transmitting signals between the gut microbiome and the brain. This cranial nerve facilitates rapid communication by conveying chemosensory signals from the gut lumen to the CNS [82]. Recent studies have provided compelling evidence of the vagus nerve’s role in gut microbiome brain signaling (Table 3), addressing a long-standing gap in understanding this complex axis. Notably, research has demonstrated a clear causal relationship between gut microbiota composition and vagal nerve activity, underscoring the importance of microbial modulation in neural signaling [82].
For instance, germ-free mice—animals raised without any gut bacteria—exhibit significantly lower vagal nerve activity compared to their colonized counterparts. Remarkably, this reduced vagal tone is reversible upon the introduction of gut bacteria, highlighting the dynamic interplay between microbes and neural pathways [83]. Furthermore, experiments involving antibiotics have revealed that disrupting the gut microbiota acutely decreases vagal activity. Restoring the gut environment by reintroducing intestinal fluids from normal mice restores vagal activity, whereas fluids from germ-free mice fail to elicit the same effect. These findings confirm the existence of a direct gut-to-brain pathway mediated by the vagus nerve, with activation extending to neurons in the brainstem [84].
b. Enteric Nervous System (ENS):
The GBA’s neural pathway is mediated by both the vagus nerve and the ENS, often referred to as the “second brain.” In addition to the vagus nerve, ENS often referred to as the “second brain” plays a crucial role in mediating the GBA’s neural pathways. The ENS comprises two main components [85]:
- Meissner’s Plexus: Located in the intestinal submucosa, this network regulates secretory functions.
- Auerbach’s Plexus: Found in the muscular layer of the gut, it governs peristalsis and other motor functions [86].
The ENS is densely interconnected by neural fibers and closely linked with gut-associated lymphoid tissue (GALT), forming a complex regulatory network. This intricate interplay between the ENS and gut bacteria is essential for maintaining homeostasis within the GBA. Dysregulation of this interaction has been implicated in disorders such as irritable bowel syndrome (IBS) and neurodegenerative diseases [87], emphasizing the ENS’s role as a key mediator of gut–brain communication.
3.2. Humoral and Metabolic Pathways
Microbial metabolites represent a cornerstone of the GBA, acting as molecular messengers that extend the influence of gut microbes far beyond the gastrointestinal tract. These small molecules regulate systemic pathways and exert effects on distant organs, including the brain. Among the most well-studied microbial metabolites are short-chain fatty acids (SCFAs) [88], bile acids, and indoles, all of which play critical roles in modulating host health and neural function.
a. Microbial Metabolites as Signals:
The gut microbiota produces a wide array of secondary metabolites that extend their effects far beyond the gastrointestinal tract, influencing systemic pathways and distant organs. Specific microbial metabolites, including short-chain fatty acids (SCFAs), bile acids, and 3-indoxyl sulfate, activate vagal afferent neurons in a receptor-dependent manner [89]. Serial perfusion of each metabolite class activates both shared and distinct neuronal subsets with varied response kinetics.
b. Short-Chain Fatty Acids (SCFAs):
SCFAs namely, butyrate, propionate, and acetate—are generated through the fermentation of dietary fiber and complex carbohydrates by the gut microbiota. The main bacterial phyla involved in SCFA production are Firmicutes and Bacteroidetes [90].
- SCFAs represent a valuable source of energy for host cells.
- SCFAs (alongside bile acids) are key activators of vagal neurons.
- SCFA acetate promotes the proliferation of human neural progenitor cells.
SCFAs have diverse effects, including regulation of gene expression, apoptosis, and maintaining the integrity of the gastrointestinal epithelium. Butyrate, in particular, is crucial for promoting host cell differentiation, inflammation control, apoptosis, and intestinal mucosal integrity [91].
c. Neurotransmitters:
Gut bacteria synthesize various neuroactive compounds that directly affect CNS function, behavior, and mood. These include serotonin, acetylcholine, dopamine, and gamma-aminobutyric acid (GABA). Both serotonin and GABA are critical for mood regulation. Probiotics have also been linked to neurotransmitters like serotonin, GABA, histamine, noradrenaline, and adrenaline [92].
d. Bile Acids and Indoles: Modulators of Systemic Health
Symbiotic intestinal bacteria mediate the conversion of primary bile acids (BAs) to secondary BAs, which can enter systemic circulation and influence glucose homeostasis by interacting with nuclear receptors. Secondary BAs also possess antimicrobial properties, shaping gut microbiota composition and protecting the host from pathogens. Alterations in bile acid metabolism, driven by changes in gut microbial composition, have been linked to dyslipidemia and metabolic disorders [93].
Similarly, indole derivatives such as 3-indoxyl sulfate are regulated by the gut microbiome and activate vagal afferent neurons. These compounds contribute to systemic communication within the GBA, further emphasizing the gut microbiota’s role in maintaining host health [94].
e. Trimethylamine-N-Oxide (TMAO): A Double-Edged Sword
The gut microbiota metabolizes dietary choline, lecithin, and L-carnitine to produce trimethylamine (TMA), which is subsequently oxidized in the liver to form trimethylamine-N-oxide (TMAO). While TMAO plays a role in lipid metabolism, elevated levels in the blood have been associated with an increased risk of cardiovascular disease and stroke. This duality highlights the complex balance between beneficial and detrimental microbial metabolites in systemic health [95].
Acting as a critical mediator of the gut-brain axis, TMAO circulates in human blood and cerebrospinal fluid. Notably, high concentrations of TMAO are detrimental to the neurovascular unit (NVU), which is the functional collective of the cerebrovascular system’s vascular and glial cells, neurons, and extracellular matrix [95].

Table 3.
Key Signaling and Regulatory Molecules.
| Component | Origin | Function/Role in Disease | Source(s) |
|---|---|---|---|
| Short-Chain Fatty Acids (SCFAs) (e.g., Butyrate, Acetate) | Produced by gut bacteria via fermentation of dietary fibers. | Protective: Maintain integrity of the mucosal barrier and tight junctions. Flow to the liver via portal circulation, reducing hepatic inflammatory injury. Drive NLRP6 inflammasomes. | [29,40,75] |
| Lipopolysaccharide (LPS) | Component of microbe-derived Pathogen-Associated Molecular Patterns (PAMPs). | Inflammatory/Pathogenic: Recognised by TLR receptors, activating NF-κB. Flows via the portal vein to the liver, activating Kupffer cells and initiating immunoregulatory programs. | [13,74] |
| Bile Acids (BAs) / Secondary BAs | Primary BAs synthesized in the liver; converted to Secondary BAs (SBAs) by intestinal microorganisms. | Metabolic/Immune Modulators: SBAs have weaker NK cell activation ability than primary BAs, potentially favoring tumor proliferation. | [93,94] |
3.3. Immune and Inflammatory Pathways
The intricate interaction between the gut microbes and the host immune system is pivotal for maintaining immune homeostasis and regulating inflammatory processes.
a. Neuroinflammation:
The GBA is thought to be associated with neuroinflammation, a process leading to the loss of neurons typical of neurodegenerative diseases (NDDs) like Alzheimer’s disease (AD) and Parkinson’s disease (PD). The host microbiota constantly controls the maturation and function of microglia in the CNS. Disruptions in the gut microbial balance can lead to a dysregulated immune response, resulting in chronic inflammation [96].
b. LPS and Endotoxin Translocation:
Dysbiosis can compromise the structural stability and functional balance of the intestinal barrier [97].
- LPS (Lipopolysaccharide): Microbe-derived factors, including LPS, a structural part of Gram-negative bacteria, are recognized as Pathogen-Associated Molecular Patterns (PAMPs) by Pattern Recognition Receptors (PRRs).
- Increased Intestinal Permeability (“Leaky Gut”): Dysbiosis leads to intestinal barrier destruction. Increased gut permeability enhances the absorption of LPS. Bacterial products and PAMPs enter the portal circulation and access the liver.
- Inflammatory Activation: PAMPs, such as LPS, bind to Toll-like receptors (TLRs) (e.g., TLR4) on immune cells, such as Kupffer cells in the liver. This binding activates inflammatory pathways. The binding of LPS to TLR4 on intestinal epithelial cells, triggering the TLR4–MyD88 pathway, also stimulates the NF-κB pathway [97].
c. Cytokine Release:
This systemic activation leads to the release of pro-inflammatory mediators [98].
- Toxic Effect on CNS: The synthesis of pro-inflammatory cytokines can be induced by LPS. Pro-inflammatory cytokines have a toxic effect on the central nervous system (CNS).
- Specific Cytokines: Activation of Kupffer cells and other immune cells promotes the release of inflammatory mediators, such as Tumor Necrosis Factor-alpha (TNF-α) and Interleukin-6 (IL-6). These are major contributors to the inflammatory cascade [98].
4. Microbiota Dysbiosis in Specific Neurodegenerative Diseases
The involvement of the gut microbiota in neurodegenerative diseases (NDDs) has become a focal point of emerging research, driven by the realization that alterations in microbial community structure (dysbiosis) and function can profoundly influence the central nervous system (CNS) through the gut–brain axis. This section explores the role of dysbiosis in specific neurodegenerative and neurological disorders, highlighting microbial signatures, metabolite influences, and therapeutic implications [99].
4.1. Alzheimer’s Disease (AD)
a. Microbial Signatures:
The pathogenesis of Alzheimer’s disease (AD) is increasingly linked to pathogenic bacteria, such as Mycobacterium tuberculosis, Staphylococcus aureus, and Salmonella spp., which may contribute to neuroinflammation and amyloid-beta deposition. Dysbiosis, characterized by an overgrowth of harmful microbiota and a loss of beneficial species, plays a critical role in AD progression. This imbalance disrupts immune homeostasis and promotes systemic inflammation, which can exacerbate neurodegeneration [100].
b. Metabolite Influence:
Microbial metabolites significantly modulate cognitive function and neurological health in AD. Short-chain fatty acids (SCFAs), particularly butyrate, are key products of gut microbial fermentation with neuroprotective properties. Butyrate regulates gene expression, reduces Neuroinflammation, and enhances mitochondrial function, offering potential therapeutic applications for AD. Additionally, SCFAs play a crucial role in maintaining gut barrier integrity, preventing the translocation of pro-inflammatory molecules like lipopolysaccharides (LPS) into systemic circulation [13,14,100].
4.2. Parkinson’s Disease (PD)
a. Microbial Alterations:
A growing body of evidence underscores the gut microbiota’s pivotal role in Parkinson’s disease (PD) pathogenesis. Changes in gut microbiota composition have been observed to precede the onset of motor and non-motor symptoms in PD patients. Key findings include [101]:
- Increased abundance of Enterobacteriaceae, which is associated with inflammation and oxidative stress [11].
b. Therapeutic Relevance:
Short-chain fatty acids (SCFAs) produced by microbial fermentation exhibit neuroprotective effects and modulate immune responses, making them promising therapeutic candidates for PD. Additionally, Fecal Microbiota Transplantation (FMT)—a procedure aimed at restoring a healthy microbial community—has shown preliminary success in preclinical studies. FMT has been explored as a means to reverse dysbiosis and alleviate symptoms in PD patients, although further clinical trials are needed to validate its efficacy [27,28,29,30,31].
4.3. Other Neurological and Nervous System Diseases
a. Associated Disorders: Dysbiosis has been implicated in various neurological and nervous system diseases, including [102]:
- Parkinson’s Disease (PD): As discussed earlier, PD exhibits distinct microbial alterations.
- Multiple Sclerosis (MS): MS is characterized by autoimmune-mediated damage to the CNS, with gut microbiota playing a role in modulating immune responses. Meta-analyses have highlighted the link between microbial dysbiosis and MS progression.
- Autism Spectrum Disorders (ASD): Altered gut microbiota composition has been associated with behavioral and cognitive symptoms in ASD, emphasizing the gut–brain axis’s role in neurodevelopmental disorders [102].
b. Heterogeneity of Microbial Signatures: A comprehensive meta-analysis comparing microbial alterations across broad disease categories revealed notable differences in microbial signatures:
- Autoimmune Diseases (e.g., IBD, RA, SLE): These conditions exhibit consistent microbial changes, including enrichment of genera such as Enterococcus, Veillonella, Streptococcus, and Lactobacillus, alongside depletion of beneficial taxa like Ruminococcus, Faecalibacterium, and Butyricicoccus [69,70].
- Nervous System Diseases (e.g., MS, PD): In contrast, microbial alterations in nervous system diseases appear more heterogeneous, with no consistent markers identified across the category. This heterogeneity suggests that the microbiomes associated with NDDs are highly individualized and influenced by factors such as genetics, environmental exposures, and disease stage [2,9].
The lack of consistent microbial signatures in nervous system diseases highlights the complexity of these disorders and underscores the need for personalized approaches in microbiome-targeted therapies.
5. Mechanisms of Molecular Action
The gut microbiota exerts a profound influence on host physiology and neurological health through the production of bioactive metabolites, modulation of barrier integrity, and regulation of inflammatory signaling pathways. These mechanisms highlight the intricate interplay between the gut microbiota and the central nervous system (CNS), mediated via the gut–brain axis [23,81,88].
5.1. Role of SCFAs and Neuroactive Metabolites
a. Epigenetic Regulation: SCFAs Modulate Epigenetic Mechanisms
Microbial metabolites, particularly short-chain fatty acids (SCFAs), play a pivotal role in epigenetic regulation by modulating DNA methylation and histone modifications. This modulation offers a novel therapeutic avenue for reprogramming disease-associated epigenetic alterations [90].
i. Histone Modification and Gene Transcription:
SCFAs exert their epigenetic effects by inhibiting histone deacetylase (HDAC) activity, thereby promoting histone acetylation. This process influences gene transcription, leading to the activation or repression of specific genes [103].
- Butyrate as an HDAC Inhibitor: Butyrate is a potent HDAC inhibitor that suppresses the activities of HDAC1 and HDAC2. This inhibition can activate tumor suppressor genes while repressing oncogenes, reducing the risk of tumorigenesis. Butyrate has also been shown to promote apoptosis and inhibit cancer cell proliferation. Additionally, SCFAs modulate T-cell receptor signaling in cytotoxic CD8+ T cells, enhancing anti-PD-1 anti-tumor efficacy [104].
b. Neural Communication: Microbial Metabolites Activate Vagal Afferent Neurons
Microbial metabolites produced in the intestinal lumen regulate neural communication along the gut–brain axis, particularly through the vagus nerve [7,47].
i. Vagal Neuron Activation:
SCFAs, bile acids, and 3-indoxyl sulfate are key microbial metabolites that stimulate vagal afferent neurons in a receptor-dependent manner [48].
- Chemosensory Signals: These metabolites enable the microbial modulation and transmission of chemosensory signals from the gut to the brain [47].
- Target Specificity: Serial perfusion of each metabolite class activates both shared and distinct neuronal subsets within the vagus nerve, demonstrating varied response kinetics. Metabolite-induced increases in vagal activity correspond with the activation of brainstem neurons, confirming a direct gut-to-brain communication pathway [26].
ii. Neurotransmitters:
5.2. Impact on Barrier Integrity (Gut and CNS)
a. Intestinal Permeability: Dysbiosis Damages the Intestinal Barrier
The gut barrier serves as a critical interface between the intestinal lumen and systemic circulation, preventing the translocation of harmful substances such as bacteria, endotoxins, and antigens. Dysbiosis compromises this barrier’s structural stability (Table 4) and functional balance [23,29,30].
i. Damage Mechanisms:
- Direct mucolytic activity, acetaldehyde production by microbiota from ethanol metabolism, or activation of the NF-κB pathway by bacterial products can damage the intestinal barrier [97].
- Increased intestinal permeability, commonly referred to as “leaky gut,” facilitates the translocation of bacterial products like lipopolysaccharide (LPS) and other Pathogen-Associated Molecular Patterns (PAMPs) into systemic circulation [105].
b. SCFA Protection: SCFAs are Vital for Preserving the Gut Barrier
SCFAs play a crucial role in maintaining the integrity and functionality of the intestinal barrier.
ii. Gut Barrier Integrity:
- SCFAs regulate the balance between the function and morphology of the mucosal barrier, protect tight junctions, and maintain barrier permeability [23].
- SCFA-Specific Roles: Butyrate is particularly critical for promoting cell regeneration and maintaining intestinal barrier function. Supplementation with SCFAs can restore a damaged intestinal barrier.
- Therapeutic Relevance: Probiotic strains such as Lactobacillus improve intestinal barrier integrity, reducing bacterial translocation across the mucosa and mitigating systemic inflammation [69].
c. CNS Consequences: Systemic Inflammation is Linked to CNS ToxicityThe translocation of PAMPs and inflammatory signals from a compromised gut barrier leads to systemic inflammation, which directly impacts the CNS.
iii. Inflammatory Cascade:
- In the liver, PAMPs like LPS bind to Toll-like receptors (TLRs) on immune cells, such as Kupffer cells, activating inflammatory pathways via the NF-κB pathway [105].
- This activation induces the production of pro-inflammatory mediators, including Tumor Necrosis Factor-alpha (TNF-α) and Interleukin-6 (IL-6) [98].
iv. Neuroinflammation and CNS Toxicity:
- Pro-inflammatory cytokines have toxic effects on the CNS, contributing to neuroinflammation and neurodegeneration. For example, endogenous alcohol (produced by gut bacteria) and LPS translocation are hypothesized to exacerbate conditions like nonalcoholic steatohepatitis (NASH) and neurodegenerative diseases [6,9,11].
v. Protection through SCFAs:
6. Therapeutic Approaches Targeting the Microbiome
Microbiome-driven therapeutics represent a rapidly evolving field aimed at addressing complex diseases, including neurodegenerative disorders (NDDs), by correcting gut microbial imbalances (dysbiosis) and modulating host–microbe interactions. These interventions leverage the gut microbiota’s profound influence on systemic health through the gut–brain axis (GBA), offering novel strategies for disease prevention and treatment [106].
6.1. Probiotics, Prebiotics, and Synbiotics
a. Probiotics:
Probiotics are live microorganisms, primarily bacteria and yeast, that confer health benefits when consumed in adequate amounts.
Examples and Composition:
Common probiotic strains include reclassified Lactobacillus genera (e.g., Lacticaseibacillus, Limosilactobacillus), Bifidobacterium species, and Saccharomyces boulardii. For instance, Bifidobacterium bifidum has demonstrated efficacy in improving tumor control and enhancing anti-PD-L1 therapy in both mice and humans [107].
i. Mechanisms of Action:
- Immune Modulation: Probiotics regulate immune responses by influencing cytokine production and promoting an anti-inflammatory state. They also modulate immune cells and intestinal epithelia, mitigating dysbiosis-related illnesses [108].
- Barrier Integrity: Strains like Lactobacillus strengthen the intestinal barrier, reducing bacterial translocation and systemic inflammation.
- Antimicrobial Production: Probiotics produce antimicrobial agents (e.g., bacteriocins) that inhibit pathogenic microbes [107].
- Neurotransmitter Influence: Probiotics synthesize neurotransmitters such as serotonin, GABA, and dopamine, impacting mood regulation and neuroimmunological conditions [109].
ii. Next-Generation Probiotics (NGPs):
NGPs, derived directly from the human gut microbiota, offer enhanced adaptability. Promising candidates include Faecalibacterium prausnitzii and Akkermansia muciniphila, which exhibit anti-inflammatory properties and potential roles in managing chronic diseases [110].
iii. Clinical Trials (Non-NDD):
Probiotics have successfully managed conditions like irritable bowel syndrome (IBS), inflammatory bowel disease (IBD), and antibiotic-associated diarrhea. In psychiatric contexts, they have reduced depressive symptoms in patients with major depressive disorder. In oncology, specific strains like Clostridium butyricum CBM 588 have improved progression-free survival (PFS) in immunotherapy-treated cancer patients [111].
b. Prebiotics:
Prebiotics are non-digestible food components, typically carbohydrates (e.g., fructans, glucans), that selectively stimulate beneficial gut bacteria.
- Mechanism: Prebiotics serve as substrates for beneficial microbes like Bifidobacterium and Lactobacilli, promoting the production of short-chain fatty acids (SCFAs) [112].
- Relevance: Prebiotics enhance gut health and systemic well-being by supporting microbial balance, digestion, and immune function. For example, dietary fiber supplementation increases sensitivity to PD-1 inhibitors by suppressing regulatory T cells (Tregs) [113].
- Limitations: The efficacy of prebiotics depends on the presence of sufficient target bacteria, requiring strict control of inter-individual variability.
c. Synbiotics:
Synbiotics combine probiotics and prebiotics to achieve synergistic effects.
- Synergistic Benefits: The prebiotic component supports the survival and activity of probiotic strains, enhancing therapeutic outcomes.
- Clinical Application (Non-NDD): Synbiotics reduce the incidence of necrotizing enterocolitis and improve outcomes in ulcerative colitis (UC). They are also explored for cancer prevention [111].
6.2. Fecal Microbiota Transplantation (FMT)
Application:
FMT involves transferring intestinal microbiota from a healthy donor to a recipient’s colon, restoring microbial diversity and function. It is highly effective for treating refractory and recurrent Clostridioides difficile infections (CDI), achieving success rates of 90–92% [114].
- Neurodegenerative Context: FMT is emerging as a promising therapy for neurological diseases, including Parkinson’s disease (PD) and Multiple Sclerosis (MS) [23].
- Mechanism in Health: FMT introduces diverse beneficial microbes that compete with pathogenic bacteria, restoring metabolic activities like bile acid and SCFA production [31].
- Cancer Context: FMT from donors responsive to immune checkpoint inhibitors (ICIs) enhances antitumor immune responses, overcoming resistance to therapies like anti-PD-1 in melanoma patients [113].
6.3. Dietary and Lifestyle Modulation
a. Dietary Patterns:
- Plant-based and Mediterranean Diets (MD): These diets promote gut health and prevent non-communicable diseases (NCDs). Adherence to the MD is associated with a healthy microbiota profile and increased SCFA production [115].
- SCFA Production: High-fiber diets enhance bacterial fermentation, producing beneficial metabolites like SCFAs.
- TMAO Modulation: Plant-based diets lower trimethylamine-N-oxide (TMAO) levels, reducing inflammation and cardiovascular risks. Conversely, Western diets increase TMAO, inflammation, and metabolic dysfunction [95].
- Obesity/Metabolism: Chronic adherence to the MD normalizes gut microbiota actions, paralleling reductions in insulin resistance in obese patients [66].
b. Lifestyle Factors:
Exercise, alongside diet and genetics, influences gut microbiome composition and function.
6.4. Microbiome Engineering and Precision Therapies
a. Microbiome Profiling and Multi-omics:
Advanced tools like genomics, transcriptomics, metabolomics, and microbiomics decipher disease-microbiome-immunity interactions [54].
- Metabolomics: Focuses on small molecules and metabolic products, providing insights into microbial activities and clinical outcomes.
- Computational Approaches: Machine learning and AI identify microbial biomarkers indicative of disease, enabling personalized interventions. Network analysis uncovers causal relationships, guiding precision medicine [116].
b. Engineered Microbiomes:
Microbiome engineering manipulates microbial communities for therapeutic benefits.
- Synthetic Biology: Engineered microbes (e.g., E. coli, Lactococcus lactis) perform tasks like biosensing, metabolic production, and immune modulation [117].
- CRISPR Technology: CRISPR-based editing enables precise microbiome manipulation, targeting antibiotic-resistant pathogens or delivering anti-cancer agents.
- Targeting Specific Pathogens: Engineered E. coli detects opportunistic pathogens (e.g., Pseudomonas aeruginosa) and releases targeted bacteriocins, combating infections effectively [117].
7. Future Directions and Conclusion
7.1. Research Gaps and Technological Advancement
Future research must focus on addressing significant knowledge gaps and leveraging advanced technologies to transition microbiome science into effective clinical solutions.
a. Need for Longitudinal Studies:
There is a critical requirement for large, longitudinal human studies to monitor microbial changes over the disease course. Currently, the long-term effects of microbiome modulation on treatment efficacy and side effects often remain largely unknown, especially for long-term human treatments. Longitudinal cohort studies are essential, as the development of dynamic network modeling approaches, which can analyze changes over time, hinges on the generation of longitudinal cohorts to fill this data gap.
b. Causal Inference:
A major objective is advancing beyond mere correlation to establish definitive causal links between specific microbial actions and Neurodegenerative Disorder (NDD) pathogenesis. Most historical case–control studies did not attempt to identify the causal microbes, and whether gut microbiome alterations are causal, consequential. Grasping these essential causal links between microbiome components is crucial for understanding disease mechanisms and formulating appropriate treatment strategies.
c. Systems Biology:
This field demands the extensive employment of network analysis and multi-omics integration to fully understand complex ecological dynamics and predict microbial behavior. Advances in technologies such as genomics, transcriptomics, metabolomics, and microbiomics are gradually helping to decipher the intricate connections between disease, immunity, and the microbiome [54].
- Network Analysis: Network modeling approaches facilitate the exploration of microbial interactions, the consolidation of various forms of data, and the identification of key players, offering a cohesive framework for investigating the complex dynamics of the gut microbiome. Specifically, the application of causal networks can be used to scrutinize relationships relying on observed microecology data or deployed to evaluate the impact of deliberate interventions.
- Multi-omics Integration: Multi-omics network construction provides a multifaceted approach, aggregating data from disparate sources (such as metagenomics, metatranscriptomics, and metabolomics) to offer a panoramic view of crucial units. Integrating multi-omics time-series microbiome data to construct dynamic Bayesian networks (DBNs) is a common method that helps dynamically monitor the ever-evolving gut microecological environments [55].
7.2. Clinical Translation and Implementation Challenges
Translating promising findings into routine clinical practice faces several hurdles concerning standards, safety, and individual variability.
- Standardization and Safety: Establishing robust regulatory frameworks and ethical guidelines is paramount to ensure patient safety and treatment efficacy. Translational applications are often hindered by regulatory hurdles and a lack of standardized protocols. For interventions like FMT, standardized preparation and long-term safety assessment are needed, as its long-term effectiveness and stability remain unclear [118].
- Personalization: Clinicians must overcome the challenge of high inter-individual variability in microbiomes. The gut microbiome exhibits significant heterogeneity in inter-individual differences, which strongly influences therapeutic response rates. This variability suggests that a “one size fits all” approach is often insufficient, necessitating the development of personalized therapeutic strategies guided by individual microbial profiles [119].
8. Conclusions
8.1. Recap Key Insight:
The sources indicate that the gut microbiota functions as a critical immune-metabolic regulator, profoundly influencing systemic health and neurological function through the Gut–Brain Axis (GBA). The GBA is a bidirectional communication network linking the gastrointestinal tract with the Central Nervous System (CNS). Research has confirmed a clear gut-to-brain pathway, revealing that the gut microbiome regulates metabolites (such as short-chain fatty acids and bile acids) that activate the vagus nerve, enabling the microbial modulation of chemosensory signals transmitted to the brain. Dysbiosis in this ecosystem has been associated with neurological diseases such as Alzheimer’s and Parkinson’s.
8.2. Outlook:
Microbiome-based approaches hold transformative potential to become the cornerstone of precision medicine. By leveraging tools like microbiome profiling and metagenomic data, these approaches offer a way to identify individual-specific microbial signatures, paving the way for personalized and effective diagnostic and therapeutic solutions for neurodegenerative disorders and other complex conditions. Discovering potent and stable gut bacteria as effective markers is a crucial area of investigation that can predict clinical response and provide new therapeutic targets.
8.3. Call to Action:
There is an urgent need for interdisciplinary research integrating microbiology, genomics, immunology, and personalized medicine. The full realization of the potential held by microbial communities for global health improvement necessitates a joint effort from multidisciplinary fields to develop a comprehensive, systematic, safe, and stable therapeutic system.
Funding
This research received no external funding.
Institutional Review Board Statement
The study did not involve human or animal subjects; therefore, ethical approval was not required.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Acknowledgments
The authors are grateful to the Deanship of Scientific (PSAU/2024/R/1445), Prince Sattam bin Abdulaziz University, Al-Kharj, Saudi Arabia for its support and encouragement in conducting the research and publishing this report.
Conflicts of Interest
There is no conflict of interests between authors.
References
- Menezes, A.A.; Shah, Z.A. A Review of the Consequences of Gut Microbiota in Neurodegenerative Disorders and Aging. Brain Sci. 2024, 14, 1224. [Google Scholar] [CrossRef]
- Gupta, M.K.; Chauhan, K.; Bhardwaj, S.; Srivastava, R. Innovative Interventions: Postbiotics and Psychobiotics in Neurodegenerative Disease Treatment. Probiotics Antimicrob. Proteins 2025, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Wang, S.; Xu, C.; Zhou, X.; Lian, X.; He, L.; Li, K. Gut microbiota, pathogenic proteins and neurodegenerative diseases. Front. Microbiol. 2022, 13, 959856. [Google Scholar] [CrossRef] [PubMed]
- Bicknell, B.; Liebert, A.; Borody, T.; Herkes, G.; McLachlan, C.; Kiat, H. Neurodegenerative and Neurodevelopmental Diseases and the Gut-Brain Axis: The Potential of Therapeutic Targeting of the Microbiome. Int. J. Mol. Sci. 2023, 24, 9577. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.; Galaction, A.I.; Turnea, M.; Blendea, C.D.; Rotariu, M.; Poștaru, M. Redox Homeostasis, Gut Microbiota, and Epigenetics in Neurodegenerative Diseases: A Systematic Review. Antioxidants 2024, 13, 1062. [Google Scholar] [CrossRef]
- Koumpouli, D.; Koumpouli, V.; Koutelidakis, A.E. The Gut–Brain Axis and Neurodegenerative Diseases: The Role of Nutritional Interventions Targeting the Gut Microbiome—A Systematic Review. Appl. Sci. 2025, 15, 5558. [Google Scholar] [CrossRef]
- Guo, B.; Zhang, J.; Zhang, W.; Chen, F.; Liu, B. Gut microbiota-derived short chain fatty acids act as mediators of the gut–brain axis targeting age-related neurodegenerative disorders: a narrative review. Crit. Rev. Food Sci. Nutr. 2023, 65, 265–286. [Google Scholar] [CrossRef]
- Kalyan, M.; Tousif, A.H.; Sonali, S.; Vichitra, C.; Sunanda, T.; Praveenraj, S.S.; Ray, B.; Gorantla, V.R.; Rungratanawanich, W.; Mahalakshmi, A.M.; et al. Role of Endogenous Lipopolysaccharides in Neurological Disorders. Cells 2022, 11, 4038. [Google Scholar] [CrossRef]
- De Luca, R.; Arrè, V.; Nardone, S.; Incerpi, S.; Giannelli, G.; Trivedi, P.; Anastasiadou, E.; Negro, R. Gastrointestinal microbiota and inflammasomes interplay in health and disease: a gut feeling. Gut 2025, 75, 161–175. [Google Scholar] [CrossRef]
- Nair, A.T.; Ramachandran, V.; Joghee, N.M.; Antony, S.; Ramalingam, G. Gut Microbiota Dysfunction as Reliable Non-invasive Early Diagnostic Biomarkers in the Pathophysiology of Parkinson’s Disease: A Critical Review. J. Neurogastroenterol. Motil. 2018, 24, 30–42. [Google Scholar] [CrossRef]
- Liang, F.; Chen, C.-Y.; Li, Y.-P.; Ke, Y.-C.; Ho, E.-P.; Jeng, C.-F.; Lin, C.-H.; Chen, S.-K. Early Dysbiosis and Dampened Gut Microbe Oscillation Precede Motor Dysfunction and Neuropathology in Animal Models of Parkinson’s Disease. J. Park. Dis. 2022, 12, 2423–2440. [Google Scholar] [CrossRef]
- Marizzoni, M.; Cattaneo, A.; Mirabelli, P.; Festari, C.; Lopizzo, N.; Nicolosi, V.; Mombelli, E.; Mazzelli, M.; Luongo, D.; Naviglio, D.; et al. Short-Chain Fatty Acids and Lipopolysaccharide as Mediators Between Gut Dysbiosis and Amyloid Pathology in Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 78, 683–697. [Google Scholar] [CrossRef]
- Marizzonia, M.; Cattaneob, A.; Mirabellic, P.; Festaria, C.; Lopizzob, N.; Nicolosia, V.; Frisonig, G.B. Short-Chain Fatty Acids and Lipopolysaccharide as Mediators Between Gut Dysbiosis and Amyloid Pathology in Alzheimer’s Disease. Handbook of Microbiome and Gut-Brain-Axis in Alzheimer’s Disease 2022, 9, 97. [Google Scholar]
- Luo, Y.; Yang, N.; Yang, W.; Chen, B.; Zhu, S.; Wu, Y.; Wang, Q. Autonomic dysfunction in multiple system atrophy: from pathophysiology to clinical manifestations. Annals of Medicine 2025, 57, 2488111. [Google Scholar] [CrossRef]
- Moran, N.A.; Ochman, H.; Hammer, T.J. Evolutionary and ecological consequences of gut microbial communities. Annual Review of Ecology, Evolution, and Systematics 2019, 50, 451–475. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.C.; Charles, T.; Schloter, M. Microbiome definition re-visited: old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.G.; Wanjari, U.R.; Kannampuzha, S.; Murali, R.; Namachivayam, A.; Ganesan, R.; Gopalakrishnan, A.V. The implication of mechanistic approaches and the role of the microbiome in polycystic ovary syndrome (PCOS): a review. Metabolites 2023, 13, 129. [Google Scholar] [CrossRef]
- Rolig, A.S.; Cech, C.; Ahler, E.; Carter, J.E.; Ottemann, K.M. The degree of Helicobacter pylori-triggered inflammation is manipulated by preinfection host microbiota. Infection and immunity 2013, 81, 1382–1389. [Google Scholar] [CrossRef]
- Hrncir, T. Gut microbiota dysbiosis: triggers, consequences, diagnostic and therapeutic options. Microorganisms 2022, 10, 578. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Harrison, O.J. Homeostatic immunity and the microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.J.; Zhang, W. Role of dietary nutrients in the modulation of gut microbiota: a narrative review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef]
- Seekatz, A.M.; Safdar, N.; Khanna, S. The role of the gut microbiome in colonization resistance and recurrent Clostridioides difficile infection. Therapeutic Advances in Gastroenterology 2022, 15, 17562848221134396. [Google Scholar] [CrossRef]
- Spiljar, M.; Merkler, D.; Trajkovski, M. The immune system bridges the gut microbiota with systemic energy homeostasis: focus on TLRs, mucosal barrier, and SCFAs. Frontiers in immunology 2017, 8, 1353. [Google Scholar] [CrossRef]
- Belizário, J.E.; Faintuch, J.; Garay-Malpartida, M. Gut microbiome dysbiosis and immunometabolism: new frontiers for treatment of metabolic diseases. Mediators of inflammation 2018, 2018, 2037838. [Google Scholar] [CrossRef]
- Toor, D.; Wasson, M.K.; Kumar, P.; Karthikeyan, G.; Kaushik, N.K.; Goel, C.; Prakash, H. Dysbiosis disrupts gut immune homeostasis and promotes gastric diseases. International journal of molecular sciences 2019, 20, 2432. [Google Scholar] [CrossRef]
- Mohsenzadeh, A.; Pourasgar, S.; Mohammadi, A.; Nazari, M.; Nematollahi, S.; Karimi, Y.; Elahi, R. The gut microbiota and cardiovascular disease: Exploring the role of microbial dysbiosis and metabolites in pathogenesis and therapeutics. Life Sciences 2025, 123981. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, N.N.; Forry, S.P.; Servetas, S.L.; Hunter, M.E.; Dootz, J.N.; Dunkers, J.P.; Jackson, S.A. Measuring microbial community-wide antibiotic resistance propagation via natural transformation in the human gut microbiome. bioRxiv 2024, 2024–11. [Google Scholar] [CrossRef]
- Majumdar, A.; Siva Venkatesh, I.P.; Basu, A. Short-chain fatty acids in the microbiota–gut–brain axis: role in neurodegenerative disorders and viral infections. ACS chemical neuroscience 2023, 14, 1045–1062. [Google Scholar] [CrossRef]
- LaGreca, M.; Hutchinson, D.R.; Skehan, L. The microbiome and neurotransmitter activity. The Journal of Science and Medicine 2021, 3. [Google Scholar] [CrossRef]
- Pandit, S.S.; Meganathan, P.; Vedagiri, H. Harmonizing gut microbiota dysbiosis: Unveiling the influence of diet and lifestyle interventions. Metabolism Open 2025, 100384. [Google Scholar] [CrossRef]
- Redondo-Useros, N.; Nova, E.; González-Zancada, N.; Díaz, L.E.; Gómez-Martínez, S.; Marcos, A. Microbiota and lifestyle: a special focus on diet. Nutrients 2020, 12, 1776. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Saunders, C.; Sanossian, N. Food, gut barrier dysfunction, and related diseases: A new target for future individualized disease prevention and management. Food science & nutrition 2023, 11, 1671–1704. [Google Scholar]
- Nisa, P.; Kirthi, A.V.; Sinha, P. Microbiome-based approaches to personalized nutrition: from gut health to disease prevention. Folia Microbiologica 2025, 1–18. [Google Scholar] [CrossRef]
- Kociszewska, D.; Chan, J.; Thorne, P.R.; Vlajkovic, S.M. The link between gut dysbiosis caused by a high-fat diet and hearing loss. International Journal of Molecular Sciences 2021, 22, 13177. [Google Scholar] [CrossRef] [PubMed]
- Al Tarraf, A. Impact of polyphenols and feeding rhythms on the immunomodulation properties of the probiotic bacteria in the gastro-intestinal tract. Doctoral dissertation, Université Bourgogne Franche-Comté, 2021. [Google Scholar]
- Green, J.E.; Davis, J.A.; Berk, M.; Hair, C.; Loughman, A.; tle, D.; Marx, W. Efficacy and safety of fecal microbiota transplantation for the treatment of diseases other than Clostridium difficile infection: a systematic review and meta-analysis. Gut microbes 2020, 12, 1854640. [Google Scholar] [CrossRef]
- Renesteen, E.; Boyajian, J.L.; Islam, P.; Kassab, A.; Abosalha, A.; Makhlouf, S.; Prakash, S. Microbiome Engineering for Biotherapeutic in Alzheimer’s Disease Through the Gut–Brain Axis: Potentials and Limitations. International Journal of Molecular Sciences 2025, 26, 5351. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: paradigm for next-generation beneficial microorganisms. Nature Reviews Gastroenterology & Hepatology 2022, 19, 625–637. [Google Scholar]
- Yousefi, Y.; Baines, K.J.; Vareki, S.M. Microbiome bacterial influencers of host immunity and response to immunotherapy. Cell Reports Medicine 2024, 5. [Google Scholar] [CrossRef]
- Wilson, DM, III; Cookson; Van Den Bosch, L.; Zetterberg, H.; Holtzman, DM.; Dewachter, I. Hallmarks of neurodegenerative diseases. Cell 2023, 186, 693–714. [Google Scholar] [CrossRef]
- 42; Wingo, TS.; Liu, Y.; Gerasimov, ES.; Vattathil, SM.; Wynne, ME.; Liu, J.; (). Shared mechanisms across the major psychiatric and neurodegenerative diseases. Nat Commun. 2022, 13, 4314. [Google Scholar] [CrossRef]
- Voet, S.; Srinivasan, S.; Lamkanfi, M.; van Loo, G. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO Mol Med. 2019, 43 11, e10248. [Google Scholar] [CrossRef] [PubMed]
- Miller, AL.; Bessho, S.; Grando, K.; Tükel, Ç. Microbiome or infections: amyloid-containing biofilms as a trigger for complex human diseases. Front Immunol 2021, 12, 638867. [Google Scholar] [CrossRef]
- Round, JL.; Mazmanian, SK. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci U S A 2010, 107, 12204–9. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, I.; Dey, S.; Raut, A.J.; Katta, S.; Sharma, P. Exploring the Gut-Brain Axis: A Comprehensive Review of Interactions Between the Gut Microbiota and the Central Nervous System. International Journal for Multidisciplinary Research 2024, 6, 1. [Google Scholar]
- Ni Dhonnabhain, R.; Xiao, Q.; O’Malley, D. Aberrant gut-to-brain signaling in irritable bowel syndrome-the role of bile acids. Frontiers in Endocrinology 2021, 12, 745190. [Google Scholar] [CrossRef]
- Makris, A.P.; Karianaki, M.; Tsamis, K.I.; Paschou, S.A. The role of the gut-brain axis in depression: endocrine, neural, and immune pathways. Hormones 2021, 20, 1–12. [Google Scholar] [CrossRef]
- Solanki, R.; Karande, A.; Ranganathan, P. Emerging role of gut microbiota dysbiosis in neuroinflammation and neurodegeneration. Frontiers in Neurology 2023, 14, 1149618. [Google Scholar] [CrossRef]
- Gautam, P.; Vishwakarma, R.K.; Pathak, A. The Gut-Brain Axis (GBA): Implications for Brain Longevity. In Rejuvenating the Brain: Nutraceuticals, Autophagy, and Longevity; Springer Nature Singapore: Singapore, 2025; pp. 219–268. [Google Scholar]
- Samarani, M.; Loberto, N.; Murdica, V.; Schiumarini, D.; Prioni, S.; Prinetti, A.; Aureli, M. Glycohydrolases in the central nervous system: the role of GBA2 in the neuronal differentiation. SpringerPlus 2015, 4 Suppl 1, P42. [Google Scholar] [CrossRef]
- Hasan, A.; Scuderi, S.A.; Capra, A.P.; Giosa, D.; Bonomo, A.; Ardizzone, A.; Esposito, E. An Updated and Comprehensive Review Exploring the Gut–Brain Axis in Neurodegenerative Disorders and Neurotraumas: Implications for Therapeutic Strategies. Brain Sciences 2025, 15, 654. [Google Scholar] [CrossRef]
- Gautam, P.; Vishwakarma, R.K.; Pathak, A. The Gut-Brain Axis (GBA): Implications for Brain Longevity. In Rejuvenating the Brain: Nutraceuticals, Autophagy, and Longevity; Springer Nature Singapore: Singapore, 2025; pp. 219–268. [Google Scholar]
- García-López, R.; Pérez-Brocal, V.; Moya, A. Beyond cells–The virome in the human holobiont. Microbial Cell 2019, 6, 373. [Google Scholar] [CrossRef]
- Xia, Y.; Sun, J. Applied Microbiome Statistics: Correlation, Association, Interaction and Composition; Chapman and Hall/CRC, 2024. [Google Scholar]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Chen, Z.S. Microbiota in health and diseases. Signal transduction and targeted therapy 2022, 7, 135. [Google Scholar] [CrossRef]
- Zhu, B.; Wang, X.; Li, L. Human gut microbiome: the second genome of human body. Protein & cell 2010, 1, 718–725. [Google Scholar]
- Colella, M.; Charitos, I.A.; Ballini, A.; Cafiero, C.; Topi, S.; Palmirotta, R.; Santacroce, L. Microbiota revolution: How gut microbes regulate our lives. World journal of gastroenterology 2023, 29, 4368. [Google Scholar] [CrossRef] [PubMed]
- Piccioni, A.; Rosa, F.; Manca, F.; Pignataro, G.; Zanza, C.; Savioli, G.; Candelli, M. Gut microbiota and clostridium difficile: what we know and the new frontiers. International Journal of Molecular Sciences 2022, 23, 13323. [Google Scholar] [CrossRef]
- Doytchinov, V.V.; Dimov, S.G. Microbial community composition of the Antarctic ecosystems: Review of the bacteria, fungi, and archaea identified through an NGS-based metagenomics approach. Life 2022, 12, 916. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Vedantam, G.; Hecht, D.W. Antibiotics and anaerobes of gut origin. Current Opinion in Microbiology 2003, 6, 457–461. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S. Human colonic microbiota: ecology, physiology and metabolic potential of intestinal bacteria. Scandinavian Journal of Gastroenterology 1997, 32(sup222), 3–9. [Google Scholar] [CrossRef]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cellular and Molecular Life Sciences 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Magne, F; Gotteland, M; Gauthier, L; Zazueta, A; Pesoa, S; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nature Reviews Immunology 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Moya, A.; Ferrer, M. Functional redundancy-induced stability of gut microbiota subjected to disturbance. Trends in microbiology 2016, 24, 402–413. [Google Scholar] [CrossRef]
- Bellocchi, C.; Fernández-Ochoa, Á.; Montanelli, G.; Vigone, B.; Santaniello, A.; Quirantes-Piné, R.; Beretta, L. Identification of a shared microbiomic and metabolomic profile in systemic autoimmune diseases. Journal of Clinical Medicine 2019, 8, 1291. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, G.; Cao, H.; Yu, D.; Fang, X.; de Vos, W.M.; Wu, H. Gut dysbacteriosis and intestinal disease: mechanism and treatment. Journal of applied microbiology 2020, 129, 787–805. [Google Scholar] [CrossRef] [PubMed]
- Syromyatnikov, M.; Nesterova, E.; Gladkikh, M.; Smirnova, Y.; Gryaznova, M.; Popov, V. Characteristics of the gut bacterial composition in people of different nationalities and religions. Microorganisms 2022, 10, 1866. [Google Scholar] [CrossRef]
- Shapiro, J.; Cohen, N.A.; Shalev, V.; Uzan, A.; Koren, O.; Maharshak, N. Psoriatic patients have a distinct structural and functional fecal microbiota compared with controls. The Journal of dermatology 2019, 46, 595–603. [Google Scholar] [CrossRef]
- Scher, J.U.; Ubeda, C.; Artacho, A.; Attur, M.; Isaac, S.; Reddy, S.M.; Abramson, S.B. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis & rheumatology 2015, 67, 128–139. [Google Scholar]
- Das, B.K. Altered gut microbiota in hepatocellular carcinoma: Insights into the pathogenic mechanism and preclinical to clinical findings. Apmis 2022, 130, 719–740. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Greten, T.F. Gut microbiome in HCC Mechanisms, diagnosis and therapy. Journal of hepatology 2020, 72, 230–238. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, T.; Tu, X.; Huang, Y.; Zhang, H.; Tan, D.I.; Fang, W. Gut microbiome affects the response to anti-PD-1 immunotherapy in patients with hepatocellular carcinoma. Journal for immunotherapy of cancer 2019, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Wu, C.J.; Hung, Y.W.; Lee, C.J.; Chi, C.T.; Lee, I.C.; Huang, Y.H. Gut microbiota and metabolites associate with outcomes of immune checkpoint inhibitor–treated unresectable hepatocellular carcinoma. Journal for immunotherapy of cancer 2022, 10, e004779. [Google Scholar] [CrossRef]
- Laface, C.; Lauricella, E.; Ranieri, G.; Ambrogio, F.; Maselli, F.M.; Parlagreco, E.; Numico, G. HCC and Immunotherapy: The Potential Predictive Role of Gut Microbiota and Future Therapeutic Strategies. Onco 2025, 5, 9. [Google Scholar] [CrossRef]
- Moe, K.T.; Tan, K.S.W. Mechanistic insights on microbiota-mediated development and progression of esophageal cancer. Cancers 2024, 16, 3305. [Google Scholar] [CrossRef]
- Aljeradat, B.; Kumar, D.; Abdulmuizz, S.; Kundu, M.; Almealawy, Y.F.; Batarseh, D.R.; Weinand, M. Neuromodulation and the gut–brain axis: therapeutic mechanisms and implications for gastrointestinal and neurological disorders. Pathophysiology 2024, 31, 244–268. [Google Scholar] [CrossRef]
- Ortlek, B.E.; Akan, O.B. Modeling and Analysis of SCFA-Driven Vagus Nerve Signaling in the Gut-Brain Axis via Molecular Communication. IEEE Transactions on Molecular, Biological, and Multi-Scale Communications, 2025. [Google Scholar]
- López-Ojeda, W.; Hurley, R.A. The Vagus Nerve and the Brain-Gut Axis: Implications for Neuropsychiatric Disorders. The Journal of Neuropsychiatry and Clinical Neurosciences 2024, 36, 278–282. [Google Scholar] [CrossRef]
- Longo, S.; Rizza, S.; Federici, M. Microbiota-gut-brain axis: relationships among the vagus nerve, gut microbiota, obesity, and diabetes. Acta diabetologica 2023, 60, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Jameson, K. Vagal interoception of microbial metabolites from the small intestinal lumen; University of California: Los Angeles, 2023. [Google Scholar]
- Aljeradat, B.; Kumar, D.; Abdulmuizz, S.; Kundu, M.; Almealawy, Y.F.; Batarseh, D.R.; Weinand, M. Neuromodulation and the gut–brain axis: therapeutic mechanisms and implications for gastrointestinal and neurological disorders. Pathophysiology 2024, 31, 244–268. [Google Scholar] [CrossRef]
- Neckel, P.H. Annotated translation of Georg Meissner’s first description of the submucosal plexus. Neurogastroenterology & Motility 2023, 35, e14480. [Google Scholar]
- Oyovwi, M.O.; Ajayi, A.F. A comprehensive review on immunological mechanisms amd gut-brain pathways linking gut health and neurological disorders. Discover Medicine 2025, 2, 1–34. [Google Scholar] [CrossRef]
- Majumdar, A.; Siva Venkatesh, I.P.; Basu, A. Short-chain fatty acids in the microbiota–gut–brain axis: role in neurodegenerative disorders and viral infections. ACS chemical neuroscience 2023, 14, 1045–1062. [Google Scholar] [CrossRef] [PubMed]
- Jabbari Shiadeh, S.M.; Chan, W.K.; Rasmusson, S.; Hassan, N.; Joca, S.; Westberg, L.; Ardalan, M. Bidirectional crosstalk between the gut microbiota and cellular compartments of brain: Implications for neurodevelopmental and neuropsychiatric disorders. Translational Psychiatry 2025, 15, 278. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Zhang, J.; Zhang, W.; Chen, F.; Liu, B. Gut microbiota-derived short chain fatty acids act as mediators of the gut–brain axis targeting age-related neurodegenerative disorders: a narrative review. Critical reviews in food science and nutrition 2025, 65, 265–286. [Google Scholar] [CrossRef]
- Rauf, A.; Khalil, A.A.; Rahman, U.U.; Khalid, A.; Naz, S.; Shariati, M.A.; Rengasamy, K.R.R. Recent advances in the therapeutic application of short-chain fatty acids (SCFAs): An updated review. Critical Reviews in Food Science and Nutrition 2022, 62, 6034–6054. [Google Scholar] [CrossRef] [PubMed]
- Dezfouli, M.A.; Rashidi, S.K.; Yazdanfar, N.; Khalili, H.; Goudarzi, M.; Saadi, A.; Kiani Deh Kiani, A. The emerging roles of neuroactive components produced by gut microbiota. Molecular Biology Reports 2025, 52, 1. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; Okafor, C.D.; Patterson, A.D. Bile acids and the gut microbiota: metabolic interactions and impacts on disease. Nature Reviews Microbiology 2023, 21, 236–247. [Google Scholar] [CrossRef]
- Ostrakhovitch, E.A.; Ono, K.; Yamasaki, T.R. Metabolomics in Parkinson’s Disease and Correlation with Disease State. Metabolites 2025, 15, 208. [Google Scholar] [CrossRef]
- Liu, J.; Ge, P.; Luo, Y.; Sun, Z.; Luo, X.; Li, H.; Chen, H. Decoding TMAO in the Gut-Organ Axis: From Biomarkers and Cell Death Mechanisms to Therapeutic Horizons. Drug Design, Development and Therapy 2025, 19, 3363–3393. [Google Scholar] [CrossRef]
- Krsek, A.; Schleicher, L.M.S.; Jagodic, A.; Baticic, L. Nanomedicine-Driven Modulation of the Gut–Brain Axis: Innovative Approaches to Managing Chronic Inflammation in Alzheimer’s and Parkinson’s Disease. International Journal of Molecular Sciences 2025, 26, 9178. [Google Scholar] [CrossRef]
- Guo, B.; Zhang, J.; Zhang, W.; Chen, F.; Liu, B. Gut microbiota-derived short chain fatty acids act as mediators of the gut–brain axis targeting age-related neurodegenerative disorders: a narrative review. Critical reviews in food science and nutrition 2025, 65, 265–286. [Google Scholar] [CrossRef]
- Bicknell, B.; Liebert, A.; Borody, T.; Herkes, G.; McLachlan, C.; Kiat, H. Neurodegenerative and neurodevelopmental diseases and the gut-brain axis: the potential of therapeutic targeting of the microbiome. International Journal of Molecular Sciences 2023, 24, 9577. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Bonfili, L.; Wei, T.; Eleuteri, A.M. Understanding the gut–brain axis and its therapeutic implications for neurodegenerative disorders. Nutrients 2023, 15, 4631. [Google Scholar] [CrossRef]
- Jain, N. The molecular interplay between human and bacterial amyloids: Implications in neurodegenerative diseases. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 2024, 1872, 141018. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Scuderi, S.A.; Capra, A.P.; Giosa, D.; Bonomo, A.; Ardizzone, A.; Esposito, E. An Updated and Comprehensive Review Exploring the Gut–Brain Axis in Neurodegenerative Disorders and Neurotraumas: Implications for Therapeutic Strategies. Brain Sciences 2025, 15, 654. [Google Scholar] [CrossRef]
- Moțățăianu, A.; Șerban, G.; Andone, S. The role of short-chain fatty acids in microbiota–gut–brain cross-talk with a focus on amyotrophic lateral sclerosis: a systematic review. International journal of molecular sciences 2023, 24, 15094. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Q.; Zhang, S.; Liu, J.; Fan, X.; Han, B.; Ai, X. Microbial short chain fatty acids: Effective histone deacetylase inhibitors in immune regulation. International journal of molecular medicine 2026, 57, 16. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, K.; Wei, J.; Ding, Y.; Wang, X.; Hou, H.; Cao, H. Gut microbiota-derived short-chain fatty acids regulate gastrointestinal tumor immunity: a novel therapeutic strategy? Frontiers in immunology 2023, 14, 1158200. [Google Scholar] [CrossRef]
- Choi, H.; Lee, D.; Mook-Jung, I. Gut microbiota as a hidden player in the pathogenesis of Alzheimer’s disease. Journal of Alzheimer’s Disease 2022, 86, 1501–1526. [Google Scholar] [CrossRef]
- Bicknell, B.; Liebert, A.; Borody, T.; Herkes, G.; McLachlan, C.; Kiat, H. Neurodegenerative and neurodevelopmental diseases and the gut-brain axis: the potential of therapeutic targeting of the microbiome. International Journal of Molecular Sciences 2023, 24, 9577. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Latiano, T.; Pazienza, V. Microbiota manipulation by probiotics administration as emerging tool in cancer prevention and therapy. Frontiers in oncology 2020, 10, 679. [Google Scholar] [CrossRef]
- Deng, M.; Li, X.; Wu, H.; Xue, D.; Wang, Y.; Guo, R.; Li, H. The current status and prospects of gut microbiota combined with PD-1/PD-L1 inhibitors in the treatment of colorectal cancer: a review. BMC gastroenterology 2025, 25, 380. [Google Scholar] [CrossRef]
- Sharma, R., Gupta, D., Mehrotra, R., & Mago, P. Psychobiotics: the next-generation probiotics for the brain. Current microbiology 2021, 78, 449–463. Olajide, T.S., & Ijomone, O.M. (2025). Targeting gut microbiota as a therapeutic approach for neurodegenerative diseases. Neuroprotection.
- Tomita, Y.; Sakata, S.; Imamura, K.; Iyama, S.; Jodai, T.; Saruwatari, K.; Sakagami, T. Association of Clostridium butyricum therapy using the live bacterial product CBM588 with the survival of patients with lung cancer receiving chemoimmunotherapy combinations. Cancers 2023, 16, 47. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.; Choi, T.G.; Kim, S.S. Role of short chain fatty acids in epilepsy and potential benefits of probiotics and prebiotics: targeting “health” of epileptic patients. Nutrients 2022, 14, 2982. [Google Scholar] [CrossRef]
- Ashaolu, T.J.; Ashaolu, J.O.; Adeyeye, S.A. Fermentation of prebiotics by human colonic microbiota in vitro and short-chain fatty acids production: a critical review. Journal of Applied Microbiology 2021, 130, 677–687. [Google Scholar] [CrossRef]
- Ijaz, M.U.; Akbar, A.; Ashraf, A.; Yeni, D.K.; Naz, H.; Shahid, M. Microbiome and Neurological Disorders. In Human Microbiome: Techniques, Strategies, and Therapeutic Potential; Springer Nature Singapore: Singapore, 2024; pp. 273–301. [Google Scholar]
- Khoruts, A.; Staley, C.; Sadowsky, M.J. Faecal microbiota transplantation for Clostridioides difficile: mechanisms and pharmacology. Nature Reviews Gastroenterology & Hepatology 2021, 18, 67–80. [Google Scholar]
- Dominguez, L.J.; Di Bella, G.; Veronese, N.; Barbagallo, M. Impact of Mediterranean diet on chronic non-communicable diseases and longevity. Nutrients 2021, 13, 2028. [Google Scholar] [CrossRef] [PubMed]
- Krsek, A.; Schleicher, L.M.S.; Jagodic, A.; Baticic, L. Nanomedicine-Driven Modulation of the Gut–Brain Axis: Innovative Approaches to Managing Chronic Inflammation in Alzheimer’s and Parkinson’s Disease. International Journal of Molecular Sciences 2025, 26, 9178. [Google Scholar] [CrossRef]
- Nazir, A.; Hussain, F.H.N.; Raza, A. Advancing microbiota therapeutics: The role of synthetic biology in engineering microbial communities for precision medicine. Frontiers in Bioengineering and Biotechnology 2024, 12, 1511149. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, C.A.; Kassam, Z.; Daw, J.; Smith, M.B.; Kelly, C.R. The regulation of fecal microbiota for transplantation: An international perspective for policy and public health. Clinical Research and Regulatory Affairs 2015, 32, 99–107. [Google Scholar] [CrossRef]
- Shukla, V.; Singh, S.; Verma, S.; Verma, S.; Rizvi, A.A.; Abbas, M. Targeting the microbiome to improve human health with the approach of personalized medicine: Latest aspects and current updates. Clinical Nutrition ESPEN 2024, 63, 813–820. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanisms of Gut Microbiota Influence on Neurodegeneration via the Gut-Brain Axis.
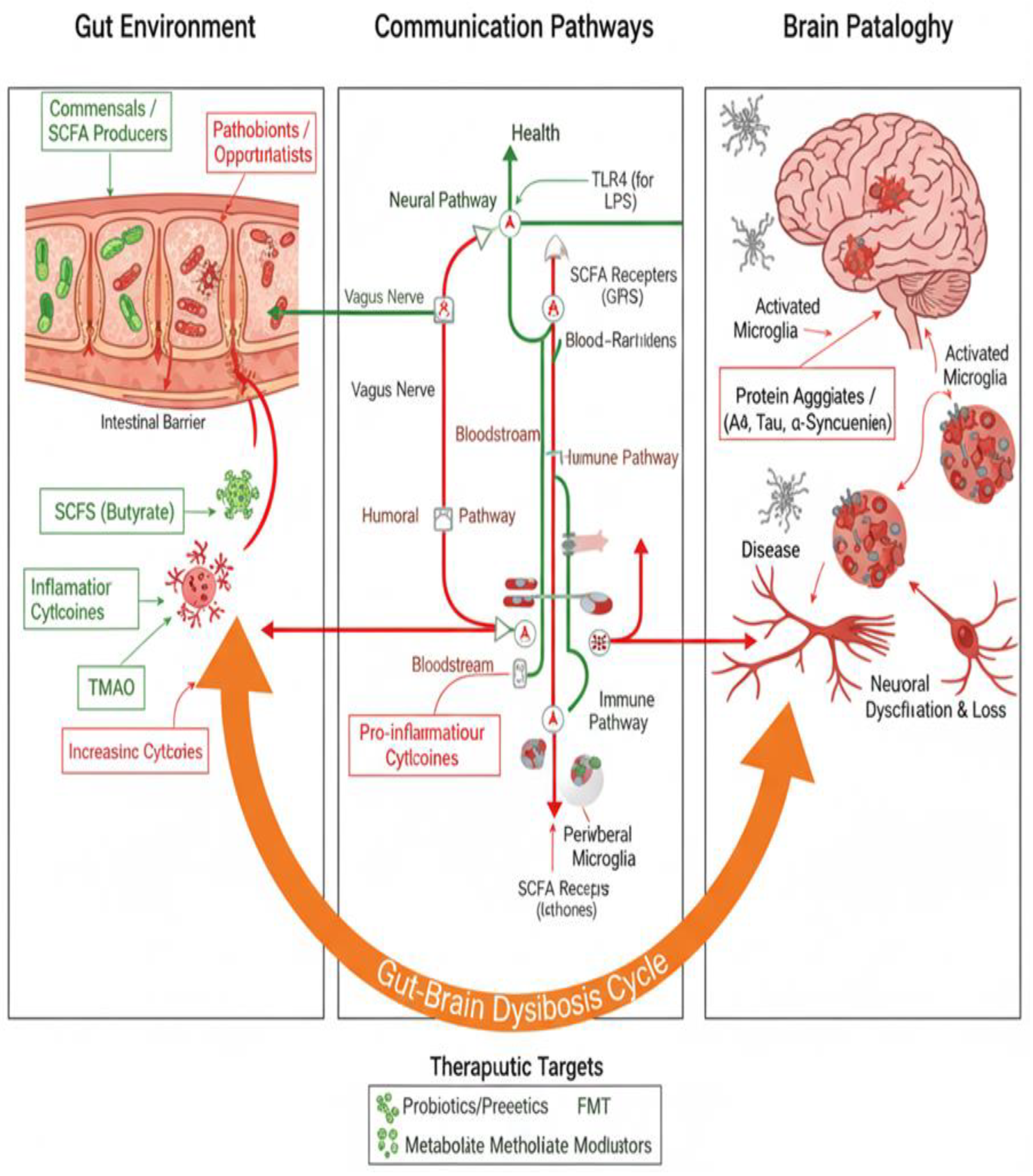
Figure 2.
Name of Human microbiota composition in different locations and body parts ([56] Nature).
Figure 2.
Name of Human microbiota composition in different locations and body parts ([56] Nature).
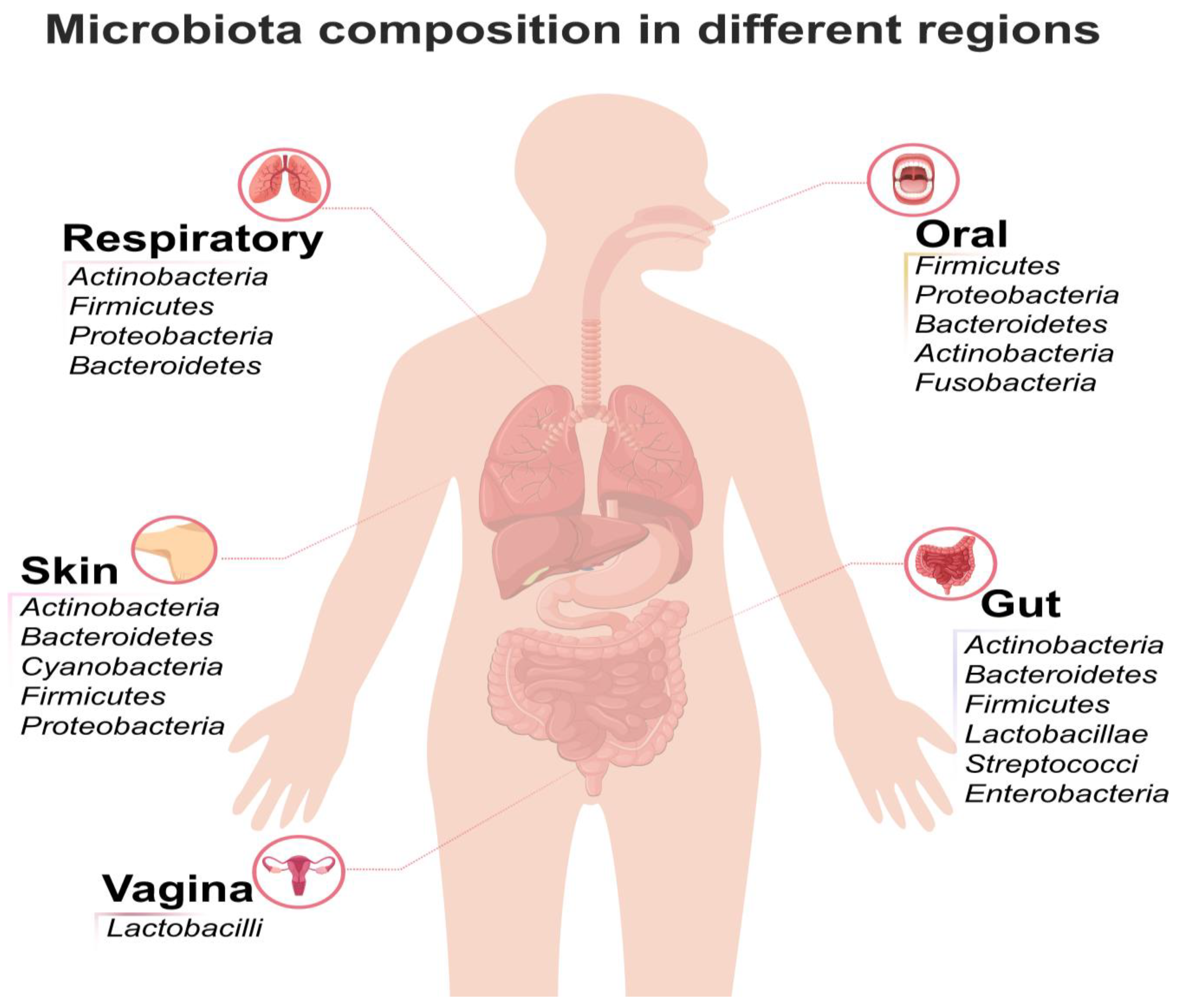
Table 1.
Overview of the Gut Microbiome, Its Functions, Associated Diseases, and Therapeutic Interventions.
Table 1.
Overview of the Gut Microbiome, Its Functions, Associated Diseases, and Therapeutic Interventions.
| Category | Details | Examples/Evidence |
|---|---|---|
| Composition of Gut Microbiome | - Dominant phyla: Firmicutes and Bacteroidetes. - Other phyla: Proteobacteria, Actinobacteria, Fusobacteria, Cyanobacteria. - Highly individualized; varies based on diet, age, genetics, and lifestyle. |
- Firmicutes and Bacteroidetes account for ~90% of gut bacteria [27]. - Over 35,000 bacterial species identified [28]. |
| Functions of Gut Microbiome | - Immune Modulation: Educates immune cells, maintains tolerance. - Nutrient Metabolism: Digests complex carbs, produces SCFAs. - Gut Barrier Integrity: Prevents harmful substances from entering bloodstream. - Gut-Brain Axis: Influences mental health. |
- SCFAs (butyrate, propionate, acetate) have anti-inflammatory properties [29]. - Gut microbiome produces neurotransmitters like serotonin [30]. |
| Causes of Dysbiosis | - Antibiotics: Disrupt microbial balance. - Diet: High in processed foods, low in fiber. - Lifestyle: Stress, lack of sleep, sedentary behavior. - Infections: Bacterial or viral infections. |
- Antibiotic use reduces microbial diversity [31]. - High-fat diets promote harmful bacteria [32]. |
| Diseases Linked to Dysbiosis | - Inflammatory Disorders: IBD, IBS, psoriasis. - Cancer: Influences tumor growth and therapy efficacy. - Metabolic Disorders: Obesity, type 2 diabetes. - Mental Health: Anxiety, depression, neurodegenerative diseases. |
- IBD patients show reduced microbial diversity [33]. - Dysbiosis linked to obesity [34]. - Gut-brain axis implicated in depression [35]. |
| Therapeutic Interventions | - Probiotics: Live bacteria that confer health benefits. - Prebiotics: Non-digestible fibers that promote beneficial bacteria. - FMT: Transplants fecal matter from healthy donors. - Dietary Interventions: High-fiber diets, specific herbs. |
- Probiotics like Lactobacillus improve gut barrier function [36]. - FMT has a 90% success rate in treating C. difficile infections [37]. -Pseudostellaria heterophylla improves gut health [38]. |
| Specific Bacteria and Their Roles | - Akkermansia muciniphila: Improves metabolic health, enhances immunotherapy. - Bifidobacterium: Promotes antitumor immunity, treats IBD. - Faecalibacterium prausnitzii: Anti-inflammatory, produces butyrate. |
- Akkermansia improves insulin sensitivity [39]. - Bifidobacterium enhances anti-PD-1 immunotherapy [40]. |
| Gut Microbiome and Cancer | - Influences tumor microenvironment (TME). - Modulates immune responses to immunotherapy. - Specific bacteria enhance or inhibit tumor growth. |
- Bifidobacterium improves response to anti-PD-1 therapy in melanoma [41]. - Dysbiosis linked to colorectal cancer [42]. |
| Gut Microbiome and Mental Health | - Produces neurotransmitters (serotonin, GABA). - Influences mood, anxiety, and depression. - Linked to neurodegenerative diseases like Alzheimer’s. |
- Gut-brain axis implicated in anxiety and depression [41]. - SCFAs reduce neuroinflammation [43]. |
| Ethical Considerations for FMT | - Informed Consent: Patients must understand risks and benefits. - Donor Screening: Rigorous screening to prevent disease transmission. - Long-Term Safety: Lack of long-term data on FMT outcomes. |
- FMT has been associated with rare cases of severe infections [44]. - Ethical concerns include donor selection and regulatory oversight [45]. |
Table 2.
Specific Gut Microbiota Alterations and Disease Signatures.
| Disease/Clinical Context | Microbial Taxa (Genus/Phylum/Species) | Observed Alteration | Clinical Implication/Mechanism | Source(s) |
|---|---|---|---|---|
| Autoimmune Diseases (Shared Patterns) | Enterococcus, Veillonella, Streptococcus, Lactobacillus, Erysipelotrichaceae incertae sedis | Enriched (Increased abundance in patients) | These alterations are notably consistent and strong, indicating a higher homogeneity of microbial markers across different autoimmune diseases compared to other categories (cancer, metabolic, nervous system disease). Enrichment of Enterococcus may drive organ-specific and systemic autoimmunity. | [69,70] |
| Autoimmune Diseases (Shared Patterns) | Ruminococcus, Faecalibacterium, Alistipes, Roseburia, Coprococcus, Oscillibacter (among others) | Depleted (Decreased abundance in patients) | Characteristically depleted genera noted across meta-analyses of autoimmune diseases. | [71] |
| Psoriasis | Firmicutes | Higher prevalence | Characteristic of unique fecal microbial communities observed in psoriasis patients. | [72] |
| Psoriasis | Bacteroidetes | Lower prevalence | Characteristic of unique fecal microbial communities observed in psoriasis patients. | [72} |
| Psoriasis | Ruminococcus and Megasphaera (Firmicutes) | Most significant difference in abundance | Identified as having significantly different abundances in psoriasis cases compared to controls. | [73] |
| Hepatocellular Carcinoma (HCC) Progression | LPS-producing Klebsiella spp. and Haemophilus spp. | Increased | Increased LPS levels boost HCC progression by stimulating the NF-κB pathway and producing pro-inflammatory cytokines, leading to hepatic inflammation and oxidative damage. | [74] |
| HCC Progression | Butyrate (Microbial metabolite) | Decreased | Promotes the development of HCC by leading to impaired energy supply to the intestinal mucosa and collapse of regulatory programs in host cells. | [75] |
| HCC Immunotherapy Response (Anti-PD-1) | Akkermansia muciniphila and Ruminococcaceae spp. | Higher abundance (in responsive patients) | Fecal samples from immunotherapy-responsive HCC patients showed a higher abundance of these taxa compared to non-responders, suggesting they are effective markers for predicting clinical response and survival benefit. | [76] |
| HCC Immunotherapy Response (Anti-PD-1) | Lachnoclostridium, Lachnospiraceae, and Veillonella | Predominant (in patients in objective remission) | Found to be predominant in HCC patients who achieved objective remission before receiving immunotherapy. | [77] |
| HCC Immunotherapy Response (Anti-PD-1) | Prevotella 9 | Significantly enriched (in progressive disease) | Associated with progressive forms of HCC disease after immunotherapy. Patients depleted in Prevotella 9 and enriched in Lachnoclostridium showed better Overall Survival (OS) and Progression-Free Survival (PFS). | [78] |
| Esophageal Cancer (ESCC/EAC) | Streptococcus, Veillonella, Prevotella, Porphyromonas gingivalis, Fusobacteria | More abundant | Found to be more common in patients with Esophageal Squamous-Cell Carcinoma (ESCC) and Esophageal Adenocarcinoma (EAC). P. gingivalis and F. nucleatum LPSs stimulate the NF-κB pathway via the TLR-4/MyD88 cascade. | [79] |
Table 4.
Structural Compartments and Boundaries (The Axes) [89].
Table 4.
Structural Compartments and Boundaries (The Axes) [89].
| Component | Description | Relevance to the Axis/Diagram | Source(s) |
|---|---|---|---|
| Gut Lumen/Microbiota | The ecosystem containing trillions of microorganisms, including bacteria, viruses, and fungi. | Source of microbial factors (PAMPs) and metabolites (SCFAs, BAs) that drive systemic signaling. | [56] |
| Intestinal Barrier / Epithelial Cells (IECs) | The physical boundary, including the protective mucus layer and tight junctions, crucial for maintaining symbiosis. | Integrity disruption allows microbial factors (LPS, PAMPs) to translocate into the host circulation. | [22,27] |
| Enteric Nervous System (ENS) / Vagus Nerve | The nervous component facilitating bidirectional communication, including vagal afferent neurons. | Essential, activity-based evidence for the Gut–Brain Axis (GBA). Vagal activity is regulated by select microbial metabolites in the small intestine. | [47,48] |
| Portal Circulation / Hepatic Portal Vein | The anatomical link connecting the gut to the liver (Gut–Liver Axis). | Transport route for gut-derived products (LPS, SCFAs, BAs) directly to the liver. | [93] |
| Liver | Target organ for systemic immune activation; contains resident immune cells (Kupffer cells). | Site where LPS activates Kupffer cells. Involved in primary bile acid synthesis and metabolism. | [97,98] |
| Central Nervous System (CNS) / Brain | Target organ modulated by the GBA; contains brainstem neurons. | Receives metabolite-induced chemosensory signals transmitted via the vagus nerve. | [47,82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



