Submitted:
28 November 2025
Posted:
01 December 2025
You are already at the latest version
Abstract
Abiotic stresses, including salt stress, drought, extreme temperature, heavy metal pollution, and waterlogging, interfere with the normal physiological activities of plants through multiple pathways. These stresses destroy the structure and function of cell membranes, inhibit enzyme activity, cause protein denaturation, and trigger oxidative stress. Such effects not only slow plant biomass accumulation, but also may initiate a series of secondary metabolic reactions, increasing the metabolic burden on plants. Abiotic stress poses a serious threat to agricultural production by reducing yields, while exerting profound negative impacts on ecosystem stability, causing many adverse effects. This review focuses on how Trichoderma promotes plant growth and nutrient uptake through multiple mechanisms under abiotic stress conditions. Additionally, it produces abundant secondary metabolites to activate the antioxidant system, thereby enhancing plant tolerance to abiotic stress and their defense capabilities. It can improve soil nutrient availability, repair agrochemical contaminated soil, promote crop growth, improve yield and quality, while reducing the use of chemical pesticides and lessening environmental impacts. Therefore, as a crucial soil microorganism,Trichoderma has great potential in alleviating crop abiotic stress. Through deep research and technological innovation, Trichoderma is expected to become an important tool for sustainable agricultural development.
Keywords:
Trichoderma
; abiotic stress
; salt stress
; antioxidant enzymes
; metabolites
1. Introduction
About 9 % of the global land is suitable for crop cultivation, with more than half of annual crops damaged by abiotic stresses. Such losses are likely to increase further with the intensification of global climate change [1]. Abiotic stresses include salt stress, extreme temperature, drought, waterlogging, heavy metal pollution, and nutrition stress, among which soil salinization, extreme temperature, and drought are the main adverse environmental conditions affecting crop growth [2]. Different responses caused by stress will seriously affect various life activities in plants, such as enzyme activity regulation, osmotic stress signal transduction, maintenance of cell membrane integrity. Ultimately, these disturbances impair plant growth and development, leading to reductions in crop yield and quality [3]. Salt stress not only disturbs the turgor pressure and chlorophyll content of plants, but also inhibits root growth, thereby reducing plant biomass [4,5]. Drought stress leads to the increase of plant osmotic factors and inhibit photosynthesis [6]. It is of great significance to enhance the tolerance of plants to abiotic stress. Microbial inoculants represent one effective solution to address soil ecological issues such as alleviating abiotic stresses, remediating soil and water pollution, and preventing plant diseases [7]. The application of microbial inoculants in plant cultivation enhances crop resilience to risks and disasters while promoting synergistic growth. Additionally, this approach helps repair the soil environmental damage caused by abiotic stress, so as to improve the agricultural quality and yield in a sustainable manner [8].
Trichoderma belongs to the Deuteromycetes subphylum and represents a common fungal genus in nature. Fungi exhibit broad adaptability to growth environments, engaging in interactions with higher plants, cyanobacteria, algae, insects, helminth, and other fungi, including intergrowth, symbiosis, antagonism, and parasitism [9]. Trichoderma indirectly promotes plant growth and enhances plant vitality via multiple pathways: by enhancing plant stress resistance, promoting the active absorption of nutrients by plants, participating in the bioremediation of contaminated rhizosphere soil, and providing plants with various secondary metabolites, enzymes, and defense-related proteins [10]. Previous studies found that Trichoderma was used as a direct biological control agent (BCA) in crop protection, and its indirect effects stimulated plant immunity, promoted plant growth, and improved the defense capacity against abiotic stresses [11]. When parasitizing or coming into contact with pathogenic bacteria, Trichoderma produces antibiotics and hydrolytic enzymes such as chitinase, β-1, 3-glucanase, lipase, and protease, which limit their growth and activity by degrading the cell walls of phytopathogenic fungi [12]. Trichoderma provides energy by metabolizing carbohydrates such as cellulose and chitin, and reduces soil pH by synthesizing and releasing organic acids such as gluconic acid [13]. Its secondary metabolites such as xylanase, peptide, terpene and pyranone promote plant growth and nutrient absorption, induce systemic resistance and control pathogens in the rhizosphere environment [14]. Therefore, as a beneficial microorganism to plants, Trichoderma contributes to the development of ecologically sustainable agriculture and provides theoretical support for the use of Trichoderma to alleviate the performance of crops under abiotic stress.
2. Trichoderma Species
Trichoderma is widely distributed in soil and plant rhizosphere, and produces various bioactive substances such as cell wall degrading enzymes, secondary metabolites, volatile organic compounds, and reactive oxygen species (ROS) in soil and plant rhizosphere, which plays an important role in crop protection [9]. Common biocontrol Trichoderma species include T.harzianum, T.viride, T.pseudokoningii, T.asperellum, T.longibrachiatum, T.polysporum, and T.brevicompactum [15]. Trichoderma forms symbionts with plant roots, stimulating root development and plant growth, activating seed germination, changing plant metabolic functions, increasing nutrient absorption and utilization, increasing crop yield, improving abiotic stress tolerance, and antagonizing pathogens [16]. Trichoderma rapidly colonizes and modifies the root structure through the interaction with plant roots, thus significantly affecting the metabolic function of plants. This interaction leads to changes in hormone levels, soluble sugars, phenolic compounds, and amino acid content within the plant, and affects photosynthetic rate, transpiration, and water content [17]. In addition, Trichoderma species exhibit high resistance to phytoalexins, phenols, aglycones, terpenoids, and flavonoids released by plants under abiotic stress. This enables Trichoderma to effectively exert its biocontrol and growth-promoting effects in the rhizosphere environment of plants [18].
3. Mechanisms of Trichoderma
There are three application mechanisms of Trichoderma in agricultural production: the first is to colonize in the soil, on plant roots, and in the rhizosphere, occupying physical space and preventing the propagation of pathogens; the second is to control plant pathogens through different mechanisms such as parasitism, antibiotic production, and induction of systemic resistance; the third is to stimulate root development and plant growth through growth-promoting effects [9]. The application of Trichoderma in the plant rhizosphere promotes the morphological characteristics of plants, such as root bud length, biomass, height, leaf number, tillers, branches, fruit, and so on [19]. When Trichoderma was inoculated, the seedling biomass, root structure index, soil nutrients, and soil enzyme activities of Pinus sylvestris var. mongolica annual seedlings were significantly increased [20]. Inoculation of Trichoderma on sugarcane improved the germination rate, tiller number, and yield of sugarcane [21]. When Trichoderma was inoculated on apples, the numbers of soil bacteria and fungi increased, and the relative contents of alkenes, ethers, and citrullines in root exudates increased [22]. More mechanisms and improvement effects of Trichoderma on alleviating plant abiotic stress are shown in Table 1 and Figure 1. Therefore, Trichoderma is widely used in agriculture as a biocontrol agents, biological agents, and soil remediation agents.
4. Application of Trichoderma in Crop Resistance to Abiotic Stress
4.1. Salt Stress
Soil salinization is a global issue, affecting approximately 9×108 hm2 of soil worldwide and resulting in annual agricultural losses of $27.3 billion.Salt stress causes ion toxicity when crops are subjected to it. High concentrations of sodium ions hinder the normal absorption of water and nutrients by plants, disrupting the ROS balance and ultimately leading to plant cell damage [43]. Therefore, salt stress affects seed germination and crop growth, resulting in reduced crop yield and quality [38].By inducing physiological protection mechanisms against oxidative damage within plants, Trichoderma further enhanced the vitality of tomato seedlings and improved the growth performance of tomatoes under osmotic stress, temperature stress, and salt stress conditions [39]. The application of Trichoderma on cucumber increased the root fresh weight by 92.55% and 84.86%, and the root dry weight by 75.71% and 53.31%, respectively, which promoted the growth of cucumber roots under high salt stress and reduced the adverse effects of salt stress on cucumber [40]. Under salt stress, a higher ethylene concentration inhibits root growth and affects the overall development of plants. The ethylene level in plants is regulated by the key enzyme 1-aminocyclopropane-1-carboxylic acid (ACC)-deaminase [41]. Trichoderma enhances salt tolerance of plants by producing IAA and increasing ACC-deaminase activity. IAA stimulates the synthesis of exogenous auxin, thereby promoting ethylene synthesis. However, the activity of ACC synthase (ACCD) in Trichoderma is increased, which decreases the availability of 1-aminocyclopropane-1-carboxylic acid (ACC) required for ethylene biosynthesis. Inoculation with Trichoderma can reduce the ethylene concentration in plants and alleviate the inhibitory effect of salt stress on root growth [42]. Liu et al. found that Trichoderma inoculated in a high salt environment effectively colonized the roots of cowpeas, enhanced photosynthesis by increasing the chlorophyll content, increased the soluble protein content, the activities of POD and CAT in seedlings, and decreased the MDA content (by 1.33% and 10.17%, respectively), reducing the ROS accumulation and salt stress damage. In addition, Trichoderma enhanced the salt tolerance of cowpeas by reducing Na+ accumulation, increasing K+ uptake in cowpeas, restoring the K+/Na+ balance, and alleviating ion toxicity [44].
Apart from ethylene, other plant hormones play a crucial role in regulating the growth and development of plants under salt stress. For example, salicylic acid (SA) activates the immune system of plants, enabling it to effectively resist various stressors, including salt stress. After inoculation with Trichoderma, the biomass, chlorophyll, and carotenoid contents of wheat seedlings increased significantly. Meanwhile, the activities of SOD, POD, and CAT increased by an average of 47.68%, 23.68%, and 38.65%, respectively. This result was closely related to the increase in endogenous SA content (the SA concentration increased by an average of 55.87% by Trichoderma treatment). SA enhanced the salt tolerance of wheat seedlings by activating phenylalanine ammonia-lyase (PAL) and the antioxidant enzyme system (such as SOD, POD, CAT), thus protecting cells and tissues from oxidative damage [45]. Trichoderma also interacted with Arabidopsis thaliana, mainly reflected in the growth and development of the plants. The number of lateral roots of the plants increased, and more auxin accumulated at the root tips. After inoculation with Trichoderma, the levels of ABA, L-proline, and ascorbic acid in Arabidopsis thaliana seedlings increased. This enhanced the ability of root exudates to remove Na+, thereby increasing the IAA level and the antioxidant and osmoprotective states of plants under salt stress. In addition, the auxin signal produced by Trichoderma improved the root branching of Arabidopsis thaliana, thus having a significant impact on plant biomass [46]. Under salt stress conditions, SOS3 perceived the Ca signal and activated SOS2, which then phosphorylated SOS1. This signaling pathway played a central role in plant response to salt stress and was one of the important molecular mechanisms for plants to adapt to a high salt environment [47]. Zhang et al. found that the transcription level of SOS1 was significantly higher than that of other groups after Trichoderma treatment [48]. In the high salinity environment, this gene plays a crucial role in regulating Na+ transport and effectively reduces the damage effect caused by Na+. In addition, the SOS1 gene also cooperates with the Na+/H+ exchanger in plants. When plants are under high salinity conditions, the two cooperate to regulate the cytoplasmic Na+ concentration and maintain intracellular ion balance, thereby enhancing the salt tolerance of plants.
4.2. Drought Stress
Drought is one of the key factors restricting global crop productivity, with an impact ranging from 9% to 10%. Under drought stress, a variety of physiological and biochemical functions of plants are disturbed, which leads to limited plant growth and decreased yield [49]. Trichoderma helps plants enhance their tolerance to drought stress by promoting antioxidant enzyme activity, the accumulation of secondary metabolites and the up-regulation of plant hormone levels. Consequently, it facilitates the growth of crops in drought-stressed soils, effectively improving the drought-resistance capabilities and yield levels of crops [50]. Inoculating Trichoderma under drought stress improves the root biomass, total biomass, nutrient content, and antioxidant enzyme activity of host plants [51]. The length of hypocotyls and roots increased after inoculation of Trichoderma in Brassica napus L., and the yield increased by 16.7% [52]. T.asperellum root dipping treatment improved the tobacco phenotype, increased the relative water content of leaves, enhanced the content of photosynthetic pigments, alleviated the accumulation of membrane lipid peroxidation products, increased the content of osmotic adjustment substances, and increased protective enzyme activity, thereby enhancing the defense ability of tobacco itself [53]. Inoculation of Trichoderma in Pinus massoniana seedlings enhanced the absorption of N, P, and K, effectively reducing the damage to cell membranes and needle tissue structure under drought stress. Meanwhile, it increased the activity of antioxidant enzymes and the content of osmoregulatory substances. In addition, Trichoderma increased the nutrient content of rhizosphere soil and the activities of invertase, phosphatase and urease. It also regulated plant physiological response and soil microbial community, simultaneously increased the rhizosphere soil contact area, secreted extracellular enzymes to activate soil nutrients, and effectively promoted the growth of Pinus massoniana seedlings under drought stress [54].
When plants are affected by drought stress, it leads to a decrease in leaf water potential, a reduction in turgor pressure, stomatal closure, and inhibition of cell growth and expansion [55]. The AQGP gene of T.harzianum represents a key role in improving plant tolerance to environmental stress by biotechnology. As reported by Vieira et al., the overexpression of AQGP of T.harzianum in tobacco improved water use efficiency and drought tolerance of tobacco. Under drought stress, AQGP transgenic lines showed higher fresh stem and root weight, suggesting that plants regulated their water content by altering their ability to retain water in stems and roots [56]. Bashyal et al. found that 1436 genes (57%) were upregulated and 1070 genes (42%) were downregulated in rice plants under drought stress treated with Trichoderma. Trichoderma delayed the adverse effects of drought stress on rice by upregulating the expression of photosynthetic genes, phenylpropanoid pathway genes, glutathione pathway related genes, and stress-enhanced osmotin [57]. Under drought stress, the contents of endogenous auxin, gibberellin, and cytokinin in plants usually decreased, while the contents of abscisic acid and ethylene tended to increase [58]. Trichoderma promoted plant growth and development by enhancing the ability of plants to absorb nutrients and water, and regulating plant hormones such as ethylene, ABA, CK, IAA, GA, and zeatin [59].
4.3. Extreme Temperature Stress
Global climate change causes the frequent occurrence of extreme temperatures, which seriously threatens crop production. When the temperature deviates from the optimal physiological range of plants, both heat stress caused by high temperatures and cold stress caused by low temperatures can cause irreversible damage to plant growth, development, and yield [60]. High temperature stress has become one of the key factors affecting plant growth and development [61]. It leads to the excessive production of ROS, lipid peroxidation, photoinhibition, protein denaturation and degradation, as well as RNA damage. These factors jointly trigger an imbalance in cellular homeostasis [62]. Cao et al. [63] discovered that inoculating Lanzhou lily with Trichoderma promoted the activities of the antioxidant enzyme system (SOD, CAT, and POD), increased the levels of heat-resistant related plant hormones (ABA, SA, and JA), raised the relative water content, elevated the chlorophyll content, and enhanced the net photosynthetic rate, thus reducing cell damage. Trichoderma effectively improved the heat tolerance of lily by promoting ABA synthesis and signal transduction. In addition, Trichoderma highly induced the DEGs of the HSF-HSP pathway. After long-term heat stress, the heat protection genes LzDREB2B, LzHsfA2a, LzMBF1c, LzHsp90 and LzHsp70 involved in the HSF-HSP signaling pathway were up-regulated. In tomato, Trichoderma enhanced heat resistance by regulating ROS metabolism, manifested as an increase in the activities of SOD, POD, PPO, and PAL, as well as an increase in proline and protein contents. Meanwhile, it maintained a relatively high PAL level to strengthen the plant’s defense response to high temperatures [64].
Chilling injury induces plant cold stress, which in turn affects plant growth. It causes a slowdown in plant growth, dehydration, leaf withering, and a reduction in the accumulation of dry matter in vegetative organs such as roots, stems, and leaves. Ultimately, this leads to a decline in plant yield or quality, and in severe cases, can even result in plant death [65]. The colonization of Trichoderma in tomato roots increased the photosynthesis and growth rate of plants, reduced the rate of membrane lipid peroxidation and electrolyte leakage, and increased the water content and proline accumulation in leaves. Meanwhile, the expression of transcription factors NAC1 and dehydrin TAS14 increased sharply, while P5CS expression declined with prolonged cold stress, thus alleviating the adverse effects of cold stress on tomato plants [34]. In addition, some volatile organic compounds released by Trichoderma species induced the expression of cold-related gene AtERD14. This indicated the ability of Trichoderma to mitigate the effects of cold stress on plants [66]. Cold stress can damage the enzyme activity, protein synthesis and cell membrane function of plants [67]. Afrouz et al. found that inoculation of maize seedlings with Trichoderma under cold conditions increased leaf length, root length, root volume and root dry weight, and significantly increased the contents of chlorophyll, carotenoids, and soluble proteins, as well as the activity of catalase. This indicated that exogenous application of Trichoderma enhanced the ability of maize to resist cold stress by optimizing the photosynthetic system, enhancing osmotic adjustment and antioxidant capacity [29].
4.4. Heavy Metal Stress
Industrial activities, sludge wastewater irrigation, and excessive use of chemical fertilizers have led to the accumulation and excess of heavy metals in soil, which not only causes damage to the soil ecosystem, but also threatens crops [68]. Heavy metal pollution destroys crop morphology, physiology and metabolic functions, including reducing ion homeostasis, interfering with photosynthesis, and reducing chlorophyll and carotenoid content [69]. Trichoderma alleviates the toxic effects of heavy metals on plants through multiple mechanisms, including valence state modification, intracellular/extracellular metal localization, metal adsorption and accumulation, and enhanced plant growth and nutrient uptake [70]. When Trichoderma alleviated heavy metal copper stress, the survival rate of onions increased under different copper concentrations. After inoculation with Trichoderma, the biomass in each treatment was mostly higher than that of non-inoculated ones. Moreover, physiological indicators such as the correlation coefficient of copper content in tissues, chlorophyll, MDA, and Pro also changed accordingly. The overall growth condition of onions was improved to a certain extent, reducing the negative impacts of copper stress [71]. Trichoderma contains various bioactive substances, such as polysaccharides and proteins. These substances bind to surrounding heavy metal ions through electrostatic adsorption, complexation, chelation, ion exchange, and covalent adsorption [9]. Polysaccharides play a crucial role in enhancing the tolerance of Trichoderma to lead (Pb2+). By increasing the proportion of uronic acid in the polysaccharide and the proportion of the main chain, the contact area between the polysaccharide and Pb2+ and the proportion of carboxyl groups can be expanded, thereby providing more binding sites for Pb2+, which helps reduce the amount of free Pb2+ and consequently mitigates heavy metal toxicity [72]. High concentrations of cadmium cause serious damage to plants, accompanied by symptoms such as yellowing, chlorosis, stem necrosis, stunted growth, and wilting poisoning. In sunflower plants inoculated with Trichoderma, the fresh weight, dry weight, plant height, and leaf area increased. Through quantitative analysis of glucose, fructose and starch, the results showed that the metabolic process of related carbohydrates was promoted, antioxidant capacity was significantly enhanced, and absorption of nutrients such as K, Ca, Mg, Fe, nitrate and P was also increased [73]. Trichoderma application modified soil structure by increasing porosity and surface area, thereby enhancing the immobilization capacity of heavy metals, and optimizing the function and synthesis of secondary metabolites such as total proteins and phenols [74]. By inoculating Trichoderma, the adverse effects of cadmium and lead toxicity on Vigna radiata (L.) were reduced, and its growth and development under metal stress were promoted [75]. The inoculation of Trichoderma in mine tailings soil contaminated by various heavy metals promoted the dissolution of heavy metals in mine tailings and removed a large amount of lead and other heavy metals from liquid media containing single and multiple metals [76]. Inoculation with Trichoderma enhanced the zinc absorption capacity of plants, effectively reduced the translocation of heavy metals to the aboveground parts, and reduced the level of oxidative stress, thereby promoting the healthy growth of hairy vetch and eliminating the toxic hazards of heavy metals to hairy vetch [77].
4.5. Waterlogging Stress
Under waterlogging stress, the stomata of plant leaves close, chlorophyll degrades, and the photosynthetic rate consequently declines. Meanwhile, waterlogging stress inhibits root respiration and induces the excessive accumulation of ROS such as H2O2 and O2-. These changes not only adversely affect the vegetative and reproductive growth of plants but further trigger cell death, ultimately leading to plant senescence [78]. T.harzianum enhanced the waterlogging tolerance of tomato seedlings by maintaining antioxidant status, regulating glucose metabolism and regulating the expression of key genes [79]. Niu et al. [80] also demonstrated that waterlogging priming induced stress memory by regulating stomatal conductance, maintaining ROS homeostasis, adjusting osmoregulatory substances, and regulating the expression of key genes mediated by H₂O₂. This alleviated the damage to tomato photosynthesis when waterlogging recurred. Trichoderma secreted plant hormones and secondary metabolites, enhanced the antioxidant system of plants, and affected physiological functions through polyamine production and regulation of gene expression to reduce the adverse effects of waterlogging stress [81,82,83,84,85,86,87,88,89,90].
5. Outlook
Trichoderma alleviates abiotic stress in plants through multiple mechanisms. Physiologically, it mitigates the adverse effects of abiotic stress by regulating stomatal aperture, increasing the relative water content of plants, and enhancing physiological parameters such as leaf number, root surface area, and root diameter. Biochemically, it activates the plant oxidative defense mechanisms, enhancing the ability to effectively remove ROS, reducing oxidative damage, and thus improving the plant resistance to abiotic stress. It also increases the contents of substances such as proline and sucrose, helping plants to lower the osmotic potential and maintain cellular water balance. Additionally, it produces or regulates the release of various plant hormones in plants, such as IAA and GA, and influences the composition of the rhizosphere microbial community, prompting the flora to produce cytokinins and antioxidants [91,92,93]. At the molecular level, it regulates the expression of plant-related genes and transmits signals through signal molecules such as transcription factors, providing support for plants to resist abiotic stress [94,95,96].
Although Trichoderma has a positive effect on coping with various abiotic stresses such as salt stress, drought stress, and extreme temperature stress, there are still some problems in its application. On the one hand, it is difficult to screen and isolate Trichoderma from extreme environments. On the other hand, the mechanism of the interaction between Trichoderma and plants is not yet fully understood. Moreover, the actual effect of field inoculation is unstable and restricted by various factors. For example, in experimental culture dishes, the colony growth of Trichoderma strains was inhibited by several fungi such as Aspergillus niger, Colletotrichum gloeosporioides, Alternaria alternata, and Fusarium. Also, the soil competitiveness of Trichoderma was tested in a corn field trial in Yakima, Washington State, USA. The results showed that the competitiveness of Trichoderma in the soil was limited, and it could not be detected a few weeks after planting [82].
In the future, it is necessary to further study the molecular mechanisms of interaction between Trichoderma and plants. Technologies such as multi-omics joint analysis should be used to explore the colonization, assembly, and growth patterns of Trichoderma in complex environments, and comprehensively analyze the mechanisms of Trichoderma in response to abiotic stress [97,98,99]. This is of great significance for understanding the molecular mechanisms of Trichoderma-plant interaction in promoting plant growth and alleviating plant stress. It is also necessary to optimize the preparation and application techniques of Trichoderma agents, explore Trichoderma strains with multiple functions, and reduce the limitations of single strain inoculation. When verifying the functions of strains, more experiments in natural environments should be carried out to improve the stability of the application effect of microbial agents in different environments. In addition, the synergistic effects of Trichoderma with other biological agents need to be explored. Trichoderma has the potential to be used in combination with low-concentration chemical pesticides, which improve the control effect on plant diseases without affecting the concept of sustainable and green development of agriculture. Trichoderma can also be combined with other biological agents to form biological agents beneficial to plant growth and disease resistance, which is widely applied in organic agricultural production. In the future, it is necessary to further explore the synergistic effect of Trichoderma and other biotic or abiotic factors to improve the mitigation effect on crop abiotic stress.
As an important soil microorganism, Trichoderma has great potential in alleviating abiotic stress in plants. Through in-depth research and technological innovation, Trichoderma is expected to become an important tool for sustainable agricultural development. It can improve soil nutrient availability, remediate agrochemical contaminated soil, promote crop growth, increase yield and quality, while reducing the use of chemical pesticides and minimizing the impact on the environment. With the deepening of the research on Trichoderma, it is believed that Trichoderma will play an increasingly important role in future agricultural production.
Author Contributions
Conceptualization, M.Z. and H.Z.; Data curation, M.Z. and H.Z.; Formal analysis, P.W.; Investigation, L.J. and Y.Y.; Project administration D.Z.; Supervision, D.Z.; Writing, M.Z. and H.Z. All authors have approved the final manuscript and agreed to the published version of the manuscript.
Funding
This work was supported by the Science and Technology Research Project of the Education Department of Hubei Province (No. D20241301).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zia, M.U.; Sambasivam, P.T.; Chen, D.; Bhuiyan, S.A.; Ford, R.; Li, Q. A carbon dot toolbox for managing biotic and abiotic stresses in crop production systems. EcoMat. 2024, 6, e12451. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Zhu, J.K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Cells Dev. 2020, 55, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, J.; Li, R.F.; Ge, Y.R.; Li, Y.F.; Li, R.L. Plants’ Response to Abiotic Stress: Mechanisms and Strategies. Int. J. Mol. Sci. 2023, 24, 10915. [Google Scholar] [CrossRef]
- Mahmud, F.M.A.; Islam, M.A.; Rubel, M.H.; Bhattacharya, D.; Ahmed, F. A sustainable methodological approach for mitigation of salt stress of rice seedlings in coastal regions: Identification of halotolerant rhizobacteria from Noakhali, Bangladesh and their impact. MethodsX 2024, 13, 102981. [Google Scholar] [CrossRef]
- Hao, R.; Zhou, W.; Li, J.; Luo, M.Q.; Scheres, B.; Guo, Y. On salt stress, PLETHORA signaling maintains root meristems. Cells Dev. 2023, 58, 1657–1669. [Google Scholar] [CrossRef]
- Yang, Z.J.; Strøbech, E.; Qiao, Y.J.; Konakalla, N.C.; Harris, P.; Peschel, G.; Agler-Rosenbaum, M.; Weber, T.; Andreasson, E.; Ding, L. Streptomyces alleviate abiotic stress in plant by producing pteridic acids. Nat. Commun. 2023, 14, 7398. [Google Scholar] [CrossRef]
- Ali, S.; Akhtar, M.S.; Siraj, M.; Zaman, W. Molecular Communication of Microbial Plant Biostimulants in the Rhizosphere Under Abiotic Stress Conditions. Int. J. Mol. Sci. 2024, 25, 12424. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.W.; Brettell, L.E.; Qiu, Z.G.; K.Singh, B. Microbiome-Mediated Stress Resistance in Plants. Trends Plant Sci. 2020, 25, 733–743. [Google Scholar] [CrossRef]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Enrique, M. Trichoderma: a multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramarkrishnan, M.; Landi, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The "Secrets" of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- TariqJaveed, M.; Farooq, T.; Al-Hazmi, A.S.; Hussain, M.D.; Rehman, A.U. Role of Trichoderma as a biocontrol agent (BCA) of phytoparasitic nematodes and plant growth inducer. J. Invertebr. Pathol. 2021, 183, 107626. [Google Scholar] [CrossRef]
- Kashyap, P.L.; Rai, P.; Srivastava, A.K.; Kumar, S. Trichoderma for climate resilient agriculture. World J. Microb. Biot. 2017, 33, 155. [Google Scholar] [CrossRef]
- Bononi, L.; Chiaramonte, J.B.; Pansa, C.C.; Moitinho; et al. Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. J. Archaeol Sci.-Rep. 2020, 10, 2858. [Google Scholar]
- Vinale, F.; Sivasithamparam, K. Beneficial effects of Trichoderma secondary metabolites on crops. Phytother. Res. 2020, 34, 2835–2842. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Druzhinina, I.S. In honor of John Bissett: authoritative guidelines on molecular identification of Trichoderma. Fungal divers. 2021, 107, 1–69. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Schmoll, M.; Esquivel-Ayala, B.A.; E.Gonzalez-Esquivel, C.; Rocha-Ramirez, V.; Larsen, J. Abiotic plant stress mitigation by Trichoderma species. Soil Ecol. Lett. 2024, 6, 240240. [Google Scholar] [CrossRef]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jolanta, J.S. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Najeeb, S.; Chen, J.; Wang, R.; Zhang, J.; Hou, J.M.; Liu, T. Insights into the molecular mechanism of Trichoderma stimulating plant growth and immunity against phytopathogens. J. Plant Physiol. 2023, 175, e14133. [Google Scholar] [CrossRef]
- Duan, X.; Zou, C.; Jiang, Y.; Yu, X.J.; Ye, X.L. Effects of Reduced Phosphate Fertilizer and Increased Trichoderma Application on the Growth, Yield, and Quality of Pepper. Plants 2023, 12, 2998. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R.Q. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Srivastava, S.N.; Singh, V.; Awasthi, S.K. Trichoderma induced improvement in growth, yield and quality of sugarcane. Sugar Tech 2006, 8, 166–169. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zhang, R.; Mao, Y.R.; Jiang, W.T.; Chen, X.S.; Shen, X.; Yin, C.M.; Mao, Z.Q. Effects of Trichoderma asperellum 6S-2 on Apple Tree Growth and Replanted Soil Microbial Environment. J. Fungi 2022, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Abdelmoaty, S.; Khandaker, M.M.; Mahmud, K.; Majrashi, A.; Alenazi, M.M.; Badaluddin, N.A. Influence of Trichoderma harzianum and Bacillus thuringiensis with reducing rates of NPK on growth, physiology, and fruit quality of Citrus aurantifolia. Braz. J. Biol. 2022, 82, e261032. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, Y.; Liu, C.; Chen, F.J.; Ge, H.L.; Tian, F.S.; Yang, T.W.; Ma, K.; Zhang, Y. Trichoderma harzianum mitigates salt stress in cucumber via multiple responses. Ecotox. Environ. Safe. 2019, 170, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Alfaro-Cuevas, R.; Lopez-Bucio, J. Trichoderma spp. Improve growth of Arabidopsis seedlings under salt stress through enhanced root development, osmolite production, and Na(+) elimination through root exudates. Mol. Plant Microbe. In. 2014, 27, 503–514. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, B.; Gan, Y. Seed Treatment with Trichoderma longibrachiatum T6 Promotes Wheat Seedling Growth under NaCl Stress Through Activating the Enzymatic and Nonenzymatic Antioxidant Defense Systems. Int. J. Mol. Sci. 2019, 20, 3729. [Google Scholar] [CrossRef]
- Mastouri, F.; Bjorkman, T.; Harman, G.E. Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistance to water deficit. Mol. Plant Microbe. In. 2012, 25, 1264–1271. [Google Scholar] [CrossRef]
- Scudeletti, D.; Crusciol, C.; Bossolani, J.W.; Moretti, L.G.; Momesso, L.; Brenda, S.T.; de Castro, S.G.Q.; De Oliveira, E.F.; Hungria, M. Trichoderma asperellum Inoculation as a Tool for Attenuating Drought Stress in Sugarcane. Front. Plant Sci. 2021, 12, 645542. [Google Scholar] [CrossRef]
- Afrouz, M.; Sayyed, R.Z.; Fazeli-Nasab, B.; Piri, R.; Almalki, W.H.; Fitriatin, B.N. Seed bio-priming with beneficial Trichoderma harzianum alleviates cold stress in maize. PeerJ 2023, 11, e15644. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, P.C.; Mishra, A.; Srivastava, S.; Chauhan, R.; Awasthi, S.; Mishra, S.; Dwivedi, S.; Tripathi, P.; Kalra, A.; Tripathi, R.D.; Nautiyal, C.S. Arsenic tolerant Trichoderma sp. reduces arsenic induced stress in chickpea (Cicer arietinum). Environ. Pollut. 2017, 223, 137–145. [Google Scholar] [CrossRef]
- Estevez-Geffriaud, V.; Vicente, R.; Vergara-Diaz, O.; Reinaldo, J.J.N.; Trillas, M.I. Application of Trichoderma asperellum T34 on maize (Zea mays) seeds protects against drought stress. Planta Daninha 2020, 252, 8. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma Applications on Strawberry Plants Modulate the Physiological Processes Positively Affecting Fruit Production and Quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Zhu, M.Y.; Su, H.Y.; Liu, X.; Li, S.X.; Zhi, Y.B.; Li, Y.X.; Zhang, J.D.; Thomas, S. Trichoderma asperellum boosts nitrogen accumulation and photosynthetic capacity of wolfberry (Lycium chinense) under saline soil stress. Tree Physiol. 2024, 44, tpad148. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanpour, A.; Salimi, A.; Ghanbary, M.A.T.; Pirdashti, H.; Dehestani, A. The effect of Trichoderma harzianum in mitigating low temperature stress in tomato (Solanum lycopersicum L.) plants. Sci. Hortic.-Ams Terdam 2017, 230, 134–141. [Google Scholar] [CrossRef]
- Yusnawan, E.; Taufiq, A.; Wijanarko, A.; Susilowati, D.M.; Praptana, R.H.; Chandra-Hioe, M.V.; Supriyo, A.; Inayati, A. Changes in Volatile Organic Compounds from Salt-Tolerant Trichoderma and the Biochemical Response and Growth Performance in Saline-Stressed Groundnut. Sustainability 2021, 13, 13226. [Google Scholar] [CrossRef]
- Ikram, M.; Ali, N.; Jan, G.; Iqbal, A.; Hamayun, M.; Jan, F.G.; Hussain, A.; Lee, I. Trichoderma reesei improved the nutrition status of wheat crop under salt stress. J. Plant Interact. 2019, 14, 590–602. [Google Scholar] [CrossRef]
- Wei, Y.L.; Yang, H.; Hu, J.D.; Li, H.M.; Zhao, Z.J.; Wu, Y.Z.; Li, J.S.; Zhou, Y.; Yang, K.; Yang, H.T. Trichoderma harzianum inoculation promotes sweet sorghum growth in the saline soil by modulating rhizosphere available nutrients and bacterial community. Front. Plant Sci. 2023, 14, 1258131. [Google Scholar] [CrossRef]
- Zhou, H.P.; Shi, H.F.; Yang, Y.Q.; Feng, X.X.; Chen, X.; Xiao, F.; Lin, H.H.; Guo, Y. Insights into plant salt stress signaling and tolerance. J. Genet. Genomics 2024, 51, 16–34. [Google Scholar] [CrossRef]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Wang, W.W.; Hu, Y.H.; Peng, Z.P.; Ren, S.; Xue, M.; Liu, Z.; Hou, J.; Xing, M.Y.; Liu, T. A novel salt-tolerant strain Trichoderma atroviride HN082102.1 isolated from marine habitat alleviates salt stress and diminishes cucumber root rot caused by Fusarium oxysporum. BMC Microbiol. 2022, 22, 67. [Google Scholar] [CrossRef]
- Brotman, Y.; Landau, U.; Cuadros-Inostroza, A.; Takayuki, T.; Fernie, A.; Chet, I.; Viterbo, A.; Willmitzer, L. Trichoderma-plant root colonization: escaping early plant defense responses and activation of the antioxidant machinery for saline stress tolerance. PLoS Pathog. 2013, 9, e1003221. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Ashraf, M. Microbial ACC-Deaminase: Prospects and Applications for Inducing Salt Tolerance in Plants. Crit. Rev. Plant Sci. 2010, 6, 360–393. [Google Scholar] [CrossRef]
- Gong, Z. Plant abiotic stress: New insights into the factors that activate and modulate plant responses. J. Integr. Plant biol. 2021, 63, 429–430. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, N.; Pang, Q.Y.; Khan, R.A.A.; Xu, Q.S.; Wu, C.D.; Liu, T. A Salt-Tolerant Strain of Trichoderma longibrachiatum HL167 Is Effective in Alleviating Salt Stress, Promoting Plant Growth, and Managing Fusarium Wilt Disease in Cowpea. J. Fungi 2023, 9, 304. [Google Scholar] [CrossRef]
- Boamah, S.; Zhang, S.W.; Xu, B.L.; Li, T.; Calderon-Urrea, A.; Tiika, R.J. Trichoderma longibrachiatum TG1 increases endogenous salicylic acid content and antioxidants activity in wheat seedlings under salinity stress. PeerJ 2022, 10, e12923. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Alfaro-Cuevas, R.; Lopez-Bucio, J. Trichoderma spp. Improve growth of Arabidopsis seedlings under salt stress through enhanced root development, osmolite production, and Na(+)elimination through root exudates. Mol. Plant Microbe In. 2014, 27, 503–514. [Google Scholar] [CrossRef]
- Zhao, S.S.; Zhang, Q.K.; Liu, M.Y.; Zhou, H.P.; Ma, C.; Wang, P.P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Zhang, S.; Gan, Y.; Xu, B. Mechanisms of the IAA and ACC-deaminase producing strain of Trichoderma longibrachiatum T6 in enhancing wheat seedling tolerance to NaCl stress. BMC Plant Biol. 2019, 19, 22. [Google Scholar]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nat. Mach. Intell. 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Reza Boorboori, M.; Zhang, H. The Mechanisms of Trichoderma Species to Reduce Drought and Salinity Stress in Plants. Phyton-Int. J. Exp. Bot. 2023, 92, 2261–2281. [Google Scholar] [CrossRef]
- He, C.; Wang, W.; Hou, J.; Li, X.E. Dark Septate Endophytes Isolated From Wild Licorice Roots Grown in the Desert Regions of Northwest China Enhance the Growth of Host Plants Under Water Deficit Stress. Front. Microbiol. 2021, 12, 522449. [Google Scholar] [CrossRef]
- Garstecka, Z.; Antoszewski, M.; Mierek-Adamska, A.; Krauklis, D.; Niedojadlo, K.; Kaliska, B.; Hrynkiewicz, K.; Dabrowska, G.B. Trichoderma viride Colonizes the Roots of Brassica napus L., Alters the Expression of Stress-Responsive Genes, and Increases the Yield of Canola under Field Conditions during Drought. Int. J. Mol. Sci. 2023, 24, 15349. [Google Scholar] [CrossRef]
- Singh, B.N.; Dwivedi, P.; Sarma, B.K.; Singh, G.S.; Singh, H.B. Trichoderma asperellum T42 Reprograms Tobacco for Enhanced Nitrogen Utilization Efficiency and Plant Growth When Fed with N Nutrients. Front. Plant Sci. 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Jiang, X.; Xu, H.Y.; Ding, G.J. Trichoderma longibrachiatum Inoculation Improves Drought Resistance and Growth of Pinus massoniana Seedlings through Regulating Physiological Responses and Soil Microbial Community. J. Fungi 2023, 9, 694. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 5, 100032. [Google Scholar] [CrossRef] [PubMed]
- Vieira, P.M.; Santos, M.P.; Andrade, C.M.; Souza-Neto, O.A.; Ulhoa, C.J.; Lima, A.; Francisco, J. Overexpression of an aquaglyceroporin gene from Trichoderma harzianum improves water-use efficiency and drought tolerance in Nicotiana tabacum. Plant Physiol. Indian J. Biochem. Bio. 2017, 121, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Bashyal, B.M.; Parmar, P.; Zaidi, N.W.; Aggarwal, R. Molecular Programming of Drought-Challenged Trichoderma harzianum-Bioprimed Rice (Oryza sativa L.). Front. Microbiol. 2021, 12, 655165. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Teng, R.M.; Liu, J.X.; Yang, Y.Z.; Lin, S.J.; Han, M.H.; Li, J.Y.; Zhuang, J. Identification and Analysis of Genes Involved in Auxin, Abscisic Acid, Gibberellin, and Brassinosteroid Metabolisms Under Drought Stress in Tender Shoots of Tea Plants. DNA Cell Biol. 2019, 38, 1292–1302. [Google Scholar] [CrossRef]
- Jaroszuk-Scisel, J.; Tyskiewicz, R.; Nowak, A.; Ozimek, E.; Majewska, M.; Hanaka, A.; Tyskiewice, K.; Pawlik, A.; Janusz, G. Phytohormones (Auxin, Gibberellin) and ACC Deaminase In Vitro Synthesized by the Mycoparasitic Trichoderma DEMTkZ3A0 Strain and Changes in the Level of Auxin and Plant Resistance Markers in Wheat Seedlings Inoculated with this Strain Conidia. Int. J. Mol. Sci. 2019, 20, 4923. [Google Scholar] [CrossRef]
- Huang, J.Y.; Zhao, X.B.; Bürger, M.; Chory, J.; Wang, X.C. The role of ethylene in plant temperature stress response. Trends Plant Sci. 2023, 28, 808–824. [Google Scholar] [CrossRef]
- Han, S.; Jiang, S.; Xiong, R.; Shanfique, K.; Zahid, K.R.; Wang, Y. Response and tolerance mechanism of food crops under high temperature stress: a review. Braz. J. Biol. 2022, 82, 253815-e253898. [Google Scholar] [CrossRef]
- Bokszczanin, K.L.; Fragkostefanakis, S.; Solanaceae, P.T.I.T. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 2013, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Sui, J.J.; Li, H.Y.; Yue, W.X.; Liu, T.; Hou, D.; Liang, J.H.; Wu, Z. Enhancing heat stress tolerance in Lanzhou lily (Lilium davidii var. unicolor) with Trichokonins isolated from Trichoderma longibrachiatum SMF2. Front. Plant Sci. 2023, 14, 1182977. [Google Scholar] [CrossRef]
- Tripathi, R.; Keswani, C.; Tewari, R. Trichoderma Koningii enhances tolerance against thermal stress by regulating ROS metabolism in tomato (Solanum lycopersicum L.) plants. J. Plant Interact. 2021, 16, 116–125. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.N.; Wang, L.C. Chilling and Drought Stresses in Crop Plants: Implications, Cross Talk, and Potential Management Opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- González-Pérez, E.; Ortega-Amaro, M.A.; Salazar-Badillo, F.B.; Bautisa, E.; Douter, J.B.; Juan, F. The Arabidopsis-Trichoderma interaction reveals that the fungal growth medium is an important factor in plant growth induction. Sci. Rep. 2018, 8, 16414–16427. [Google Scholar] [CrossRef]
- Kazemi-Shahandashti, S.S.; Maali-Amiri, R. Global insights of protein responses to cold stress in plants: Signaling, defence, and degradation. J. Plant Physiol. 2018, 226, 123–135. [Google Scholar] [CrossRef]
- Feng, D.; Wang, R.X.; Sun, X.A.; Liu, L.N.; Liu, P.; Tang, J.C.; Zhang, C.X.; Liu, H. Heavy metal stress in plants: Ways to alleviate with exogenous substances. Sci. Total Environ. 2023, 897, 165397. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.A.; Hao, Y.Y.; Shu, H.Y.; Mumtaz, M.A.; Cheng, S.H.; Alyemeni, M.N.; Ahmad, P.; Wang, Z. Melatonin enhanced the heavy metal-stress tolerance of pepper by mitigating the oxidative damage and reducing the heavy metal accumulation. J. Hazard. Mater. 2023, 454, 131468. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2015, 6, 1143. [Google Scholar] [CrossRef]
- Téllez Vargas, J.; Rodríguez-Monroy, M.; López Meyer, M.; Montes-Belmont, R.; Sepulveda-Jimenez, G. Trichoderma asperellum ameliorates phytotoxic effects of copper in onion (Allium cepa L.). Environ. Exp. Bot. 2017, 136, 85–93. [Google Scholar] [CrossRef]
- Sun, H.Q.; Meng, M.; Wu, L.R.; Zheng, X.M.; Zhu, Z.Y.; Dai, S.H. Function and mechanism of polysaccharide on enhancing tolerance of Trichoderma asperellum under Pb2+ stress. Int. J. Biol. Macromol. 2020, 151, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Abeed, A.H.A.; Mahdy, R.E.; Alshehri, D.; Hammami, I.; Eissa, M.A.; Latef, A.A.H.A.; Mahmoud, G.A.E. Induction of resilience strategies against biochemical deteriorations prompted by severe cadmium stress in sunflower plant when Trichoderma and bacterial inoculation were used as biofertilizers. Front. Plant Sci. 2022, 13, 1004173. [Google Scholar] [CrossRef] [PubMed]
- Amanullah, F.; Khan, W. Trichoderma asperellum L. Coupled the Effects of Biochar to Enhance the Growth and Physiology of Contrasting Maize Cultivars under Copper and Nickel Stresses. Plants 2023, 12, 958. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.; Ilyas, T.; Shahid, M.; Shafi, Z.; Tyagi, A.; Ali, S. Trichoderma Inoculation Alleviates Cd and Pb-Induced Toxicity and Improves Growth and Physiology of Vigna radiata (L.). ACS Omega 2024, 9, 8557–8573. [Google Scholar] [CrossRef]
- Babu, A.G.; Shea, P.J.; Oh, B.T. Trichoderma sp. PDR1-7 promotes Pinus sylvestris reforestation of lead-contaminated mine tailing sites. Sci. Total Environ. 2014, 476-477, 561–567. [Google Scholar] [CrossRef]
- Jam, E.; Khomari, S.; Ebadi, A.; Goli-Kalanpa, E.; Ghavidel, A. Correction to: Influences of peanut hull-derived biochar, Trichoderma harzianum and supplemental phosphorus on hairy vetch growth in Pb- and Zn-contaminated soil. Environ. Geochem. Hlth. 2023, 45, 9433–9434. [Google Scholar] [CrossRef]
- Pan, J.W.; Sharif, R.; Xu, X.W.; Chen, X.H. Mechanisms of Waterlogging Tolerance in Plants: Research Progress and Prospects. Front. Plant Sci. 2020, 11, 627331. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Alhaithloul, H.A.S.; Qari, S.H.; Soliman, M.H.; Hasanuzzaman, M. Pretreatment with Trichoderma harzianum alleviates waterlogging-induced growth alterations in tomato seedlings by modulating physiological, biochemical, and molecular mechanisms. Environ. Exp. Bot. 2020, 171, 103946. [Google Scholar] [CrossRef]
- Niu, L.F.; Jiang, F.L.; Yin, J.; Wang, Y.L.; Li, Y.K.; Yu, X.Q.; Song, X.M.; Ottosen, M.R.; Wu, Z.; Zhou, R.; Rosenqvist, E.; Mittler, R. ROS-mediated waterlogging memory, induced by priming, mitigates photosynthesis inhibition in tomato under waterlogging stress. Front. Plant Sci. 2023, 14, 1238108. [Google Scholar] [CrossRef]
- Rauf, M.; Awais, M.; Ud-Din, A.; Ali, K.; Gul, H.; Rahman, M.M.; Hamayun, M.; Arif, M. Molecular Mechanisms of the 1-Aminocyclopropane-1-Carboxylic Acid (ACC) Deaminase Producing Trichoderma asperellum MAP1 in Enhancing Wheat Tolerance to Waterlogging Stress. Front. Plant Sci. 2020, 11, 614971. [Google Scholar] [CrossRef]
- Redman, R.S.; Kim, Y.O.; Cho, S.; Mercer, M.; Rienstra, M.; Manglona, R.; Biaggi, T.; Zhou, X.G.; Chilvers, M.; Gray, Z.; Rodriguez, R.J.; Egamberdieva, D. A Symbiotic Approach to Generating Stress Tolerant Crops. Microorganisms 2021, 9, 920. [Google Scholar] [CrossRef]
- Zhang, D.; Chunyan, L.I.U.; Yujie, Y.A.N.G.; Qiangsheng, W.U.; Yeyun, L.I. Plant Root Hair Growth in Response to Hormones. Not. Bot. Horti Agrobot. 2019, 47, 278–281. [Google Scholar] [CrossRef]
- Sun, M.; Yuan, D.; Hu, X.; Zhang, D.; Li, Y. Effects of mycorrhizal fungi on plant growth, nutrient absorption and phytohormones levels in tea under shading condition. Not. Bot. Horti Agrobot. 2020, 48, 2006–2020. [Google Scholar] [CrossRef]
- Shu, B.; Xie, Y.; Zhang, F.; Zhang, D.; Liu, C.; Wu, Q.; Luo, C. Genome-wide identification of citrus histone acetyltransferase and deacetylase families and their expression in response to arbuscular mycorrhizal fungi and drought. J. Plant Interact. 2021, 16, 367–376. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, C.; Zhang, D.; Deng, X. Phosphorus-induced change in root hair growth is associated with IAA accumulation in walnut. Not. Bot. Horti Agrobot. 2021, 49, 12504. [Google Scholar] [CrossRef]
- Huang, X.; Gong, T.i.; Li, M.; Hu, C.; Zhang, D.; Sun, M. Root hair specification and its growth in response to nutrients. Not. Bot. Horti Agrobot. 2021, 49, 12258. [Google Scholar] [CrossRef]
- Sun, M.; Hui, X.; Tong, C.; Yuan, L.; Zhang, D. The Physiological and Molecular Responses of Exogenous Selenium to Selenium Content and Fruit Quality in Walnut. Phyton-Int. J. Exp. Bot. 2022, 92, 851–860. [Google Scholar] [CrossRef]
- Shu, B.; Xie, Y.; Zhang, F.; Zhang, D.; Liu, C.; Wu, Q.; Luo, C. Genome-wide identification of citrus calmodulin-like genes and their expression in response to arbuscular mycorrhizal fungi colonization and drought. Can. J. Plant Sci. 2022, 102, 112–123. [Google Scholar] [CrossRef]
- Jian, P.; Zha, Q.; Hui, X.; Tong, C.; Zhang, D. Research Progress of Arbuscular Mycorrhizal Fungi Improving Plant Resistance to Temperature Stress. Horticulturae 2024, 10, 855. [Google Scholar] [CrossRef]
- Lu, Q.; Jin, L.F.; Tong, C.L.; Liu, F.; Huang, B.; Zhang, D.J. Research Progress on the Growth-Promoting Effect of Plant Biostimulants on Crops. Phyton-Int. J. Exp. Bot. 2024, 93, 661–679. [Google Scholar] [CrossRef]
- Shi, G.; Zhou, X.; Tong, C.; Zhang, D. The Physiological and Molecular Mechanisms of Fruit Cracking Alleviation by Exogenous Calcium and GA3 in the Lane Late Navel Orange. Horticulturae 2024, 10, 1283. [Google Scholar] [CrossRef]
- Zhang, D.-J.; Tong, C.-L.; Wang, Q.-S.; Bie, S. Mycorrhizas Affect Physiological Performance, Antioxidant System, Photosynthesis, Endogenous Hormones, and Water Content in Cotton under Salt Stress. Plants 2024, 13, 805. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Y.; Tong, C.; Zhang, D. Effects and Mechanism of Auxin and Its Inhibitors on Root Growth and Mineral Nutrient Absorption in Citrus (Trifoliate Orange, Poncirus trifoliata) Seedlings via Its Synthesis and Transport Pathways. Agronomy 2025, 15, 719. [Google Scholar] [CrossRef]
- Hu, C.H.; Li, H.; Tong, C.L.; Zhang, D.J.; Lu, Y.M. Integrated transcriptomic and metabolomic analyses reveal the effect of mycorrhizal colonization on trifoliate orange root hair. Sci. Hortic. 2024, 336, 113429. [Google Scholar] [CrossRef]
- Zhang, D.; Zheng, J.; Yi, Q. Effects of Exogenous Trehalose on Plant Growth, Physiological and Biochemical Responses in Gardenia jasminoides Seedlings During Cold Stress. Horticulturae 2025, 11, 615. [Google Scholar] [CrossRef]
- Hu, C.; Zheng, Y.; Tong, C.; Zhang, D. Effects of exogenous melatonin on plant growth, root hormones and photosynthetic characteristics of trifoliate orange subjected to salt stress. Plant Growth Regul. 2022, 97, 551–558. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, Y.; Liu, C.; Zhang, F.; Hu, W.; Gong, S.; Wu, Q. Auxin modulates root-hair growth through its signaling pathway in citrus. Sci. Hortic. 2018, 236, 73–78. [Google Scholar] [CrossRef]
- Hu, C.; Yuan, S.; Tong, C.; Zhang, D.; Huang, R. Ethylene modulates root growth and mineral nutrients levels in trifoliate orange through the auxin-signaling pathway. Not. Bot. Horti Agrobot. 2023, 51, 13269. [Google Scholar] [CrossRef]
Figure 1.
The mechanisms of Trichoderma response to plant abiotic stress.
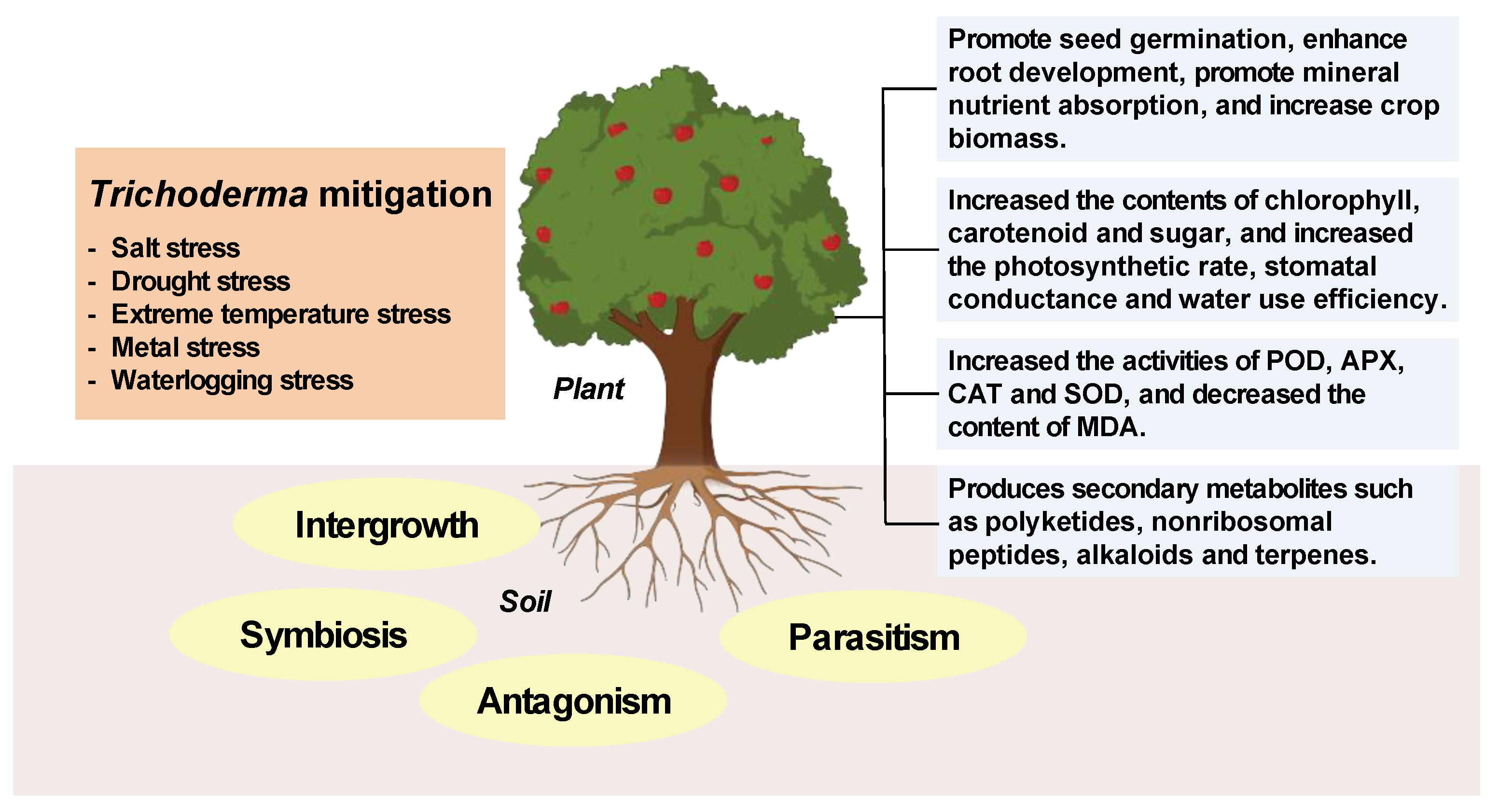

Table 1.
Mechanisms and effects of Trichoderma on pant stress.
| Crop | Trichoderma | Environment | Key function | Reference |
|---|---|---|---|---|
| Citrus aurantifolia | T. harzianum | No stress | The secretion of IAA and cytokinin increased the plant height, branch number, leaf area, and absolute growth rate. | [23] |
| Cucumber | T.harzianum | Salt stress | Increase the contents of proline, soluble sugars, soluble proteins, and chlorophyll. Enhance root activity, inhibit the absorption of Na+, and promote the absorption of K+. | [24] |
| Arabidopsis | T.viride | Salt stress | Enhance root development, improve plant IAA levels, and plant antioxidant capacity and osmotic protection. | [25] |
| Wheat | T. longibrachiatum | Salt stress | Promote seed germination, increase shoot and root weight, enhance Pro content, POD and APX enzyme activities, and reduce MDA content. | [26] |
| Tomato | T. harzianum | Drought stress | Increase the fresh and dry weights of roots and shoots, and enhance the enzymatic activities of SOD, CAT, and APX. | [27] |
| Sugarcane | T. harzianum | Drought stress | Increase the content of chlorophyll, carotenoids, sugar, improve photosynthesis rate, stomatal conductance, and water use efficiency. | [28] |
| Maize | T. harzianum | Cold stress | Produce growth regulators (such as auxin, cytokinin) to promote germination, root growth, and plant biomass. | [29] |
| Cicer arietinum | Trichoderma strain M-35 | Heavy metal stress | The methylation of arsenic in soil reduced the absorption of arsenic by plants, and the down-regulation of genes (MIPS, PGIP, CGG) enhanced the potential of plants to cope with As stress. | [30] |
| Maize | T. asperellum strain T34 | Drought stress | It improved grain P and C, grain number and dry weight, and increased leaf relative water content, water use efficiency, PSII maximum efficiency, and photosynthesis. | [31] |
| Strawberry | T. harzianum | No stress | Promote fruit development, increase yield, and enhance the accumulation of anthocyanins, and other antioxidants in fruits. | [32] |
| Lycium chinense | T. asperellum | Salt stress | Enhance the accumulation of nitrogen, dry matter, and biomass, and increase the activities of NR, NIR, and GS in roots and leaves. | [33] |
| Tomato | T. harzianum | Cold stress | It enhanced the expression of TAS14 (regulating ABA signaling pathway to enhance water retention capacity) and P5CS (catalyzes proline synthesis, maintains cell osmotic pressure), boosted the photosynthesis and growth rates, decreased the rate of lipid peroxidation and electrolyte leakage, and simultaneously increased the leaf water content and proline accumulation. | [34] |
| Groundnut | T. harzianum | Salt stress | IAA was produced, phenolic substances and flavonoids increased by 31% and 43%, photosynthesis was enhanced, chlorophyll content, bud and biomass weight were increased. | [35] |
| Wheat | T. reesei | Salt stress | Reduce the toxicity of ROS in cells and increase the content of IAA, GA, Ca, and K. | [36] |
| Sweet sorghum | T. harzianum | Salt stress | The yield, plant height, stem diameter and total sugar content in stem were increased by 35.52 %, 32.68 %, 32.09 %, and 36.82 %, respectively. | [37] |
Note: APX: ascorbate peroxidase; NR: nitrate reductase; NIR: nitrite reductase; GS: glutamine synthetase; SOD: superoxide dismutase; POD: peroxidase; CAT: catalase; Pro: proline; MDA: malondialdehyde; IAA: auxin; ABA: abscisic acid; GA: gibberellin; SA: salicylic acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



