
Submitted:
28 November 2025
Posted:
28 November 2025
You are already at the latest version
Abstract
As a three-dimensional in vitro model, organoid technology represents a revolutionary breakthrough in precision medicine. By harnessing the self-organizing capabilities of stem cells within biomimetic extracellular matrices, it enables the generation of miniature tissues that recapitulate key structural and functional characteristics of their source organs[1]. Conventional two-dimensional cell cultures lack tissue architecture and microenvironmental cues, whereas animal models are hindered by interspecies differences and inadequate representation of human pathological heterogeneity[2]. By effectively addressing these limitations, organoids have emerged as powerful platforms that are highly representative of human physiology and disease processes in oncology, genetic disorders, and infectious diseases. They demonstrate significant potential for use in drug screening, toxicity assessment, and the development of personalized treatment strategies[3, 4]. Although challenges such as limited vascularization, lack of standardized culture protocols, and ethical considerations remain, the integration of multidisciplinary approaches such as AI-assisted analysis, organ-on-a-chip systems, and 3D bioprinting, together with increasing policy support and industrial advancement, is accelerating the clinical translation of organoid technology[5]. In this review, the construction strategies for and applications of organoid models are systematically summarized, and their value and limitations in disease modeling, precision medicine, and preclinical research is highlighted. Finally, future development pathways driven by multidisciplinary collaboration and standardization are outlined.
Keywords:
organoids
; disease modeling
; personalized medicine
; 3D cell culture
; drug screening
; clinical translation
1. Introduction
Preclinical research serves as the cornerstone of drug development and disease mechanism elucidation. However, traditional two-dimensional (2D) cell culture and animal models have long been relied upon and exhibit increasingly evident limitations in translational medical research [6]. The 2D culture systems lack the support of an extracellular matrix and three-dimensional spatial organization and fail to recapitulate the physiological microenvironment and cell-cell interactions of native tissues [7]. Owing to fundamental interspecies differences in drug metabolism, immune responses, and pathological progression, animal models have significantly compromised abilities to predict human physiology and disease processes [8]. In the study of highly heterogeneous diseases such as cancer, traditional models struggle to maintain the genomic diversity and cellular subpopulation architecture of primary tumors [9], whereas animal models cannot fully replicate immune regulatory networks and stromal signaling exchanges within the human tumor microenvironment [10]. These inherent shortcomings of conventional models are considered major contributing factors to the eventual failure of approximately 90% of drugs that enter clinical trials [6].
To overcome these bottlenecks, organoid technology has emerged as a groundbreaking three-dimensional in vitro model system. Organoids are derived from adult stem cells or pluripotent stem cells and, through self-organization within biomimetic materials mimicking the native extracellular matrix, they form miniature tissues that possess the key cell types, spatial architecture, and physiological functions of the source organ [11]. Since pioneering work on intestinal organoids in 2009, this technology has achieved significant breakthroughs across multiple organ systems and disease models [12]. In particular, the successful establishment of patient-derived tumor organoid (PDTO) models, which have high retention of the histological features, mutational spectra, and drug response heterogeneity of primary tumors, has substantially increased the physiological relevance and predictive accuracy of preclinical studies [13,14]. Organoids not only demonstrate unique advantages in modeling organ development, disease mechanisms, and drug metabolism but also provide a novel platform for high-throughput drug screening and the development of personalized treatment strategies [15].
This review systematically outlines the comprehensive applications and developmental trajectory of organoid technology in biomedical research. It first delves into the role of organoids in disease modeling, covering key areas such as genetic disorders, cancer biology, and infectious disease mechanisms. It subsequently focuses on analyzing the application prospects of organoids to translational medicine, including drug toxicity assessment, prediction of individualized therapeutic efficacy, and tissue engineering strategies in regenerative medicine (Figure 1). Lastly, it examines the core challenges currently faced by this technology, such as limited vascularization and model standardization issues, and offers perspectives on the role of emerging technologies such as organ-on-a-chip systems and 3D bioprinting—in the context of multidisciplinary integration—in advancing the functionalization and clinical translation of organoids. By synthesizing recent advances from basic biology, engineering, and clinical medicine, this review aims to provide a systematic reference for researchers, facilitating the transition of organoid technology from basic research to clinical application.
2. Organoids and Organ-on-a-Chip Systems
2.1. Organoids Systems
With the continuous advancement of science and technology, tumor models have evolved from traditional two-dimensional (2D) cell culture systems to more physiologically relevant three-dimensional (3D) models, such as tumor spheroids and organoids. Accumulating evidence has demonstrated that patient-derived models offer significant advantages in recapitulating tumor complexity and predicting clinical responses [16]. Patient-derived organoids (PDOs), which are derived from the three-dimensional culture of patient tumor cells in vitro, not only retain the molecular characteristics and heterogeneity of the primary tumor but also partially mimic its structural and functional properties [17,18]. Thus, PDOs show great potential use in preclinical personalized drug evaluation.
Organoids are typically generated from induced pluripotent stem cells (iPSCs) or adult stem cells (ASCs) and can self-organize into 3D cultures that replicate the complex spatial architecture and physiological functions of the native tissue [19,20]. Compared with conventional 2D cultures, organoids exhibit organ-specific cell types and spatial organization, retain a full genetic background, and support long-term in vitro culture, making them highly promising for disease modeling and therapy research [21,22,23].
The value of PDOs is particularly evident in modeling organ physiology and pathology. A key advantage lies in their biological fidelity: by preserving the cellular heterogeneity and microenvironmental features of primary tumors, organoids can closely simulate in vivo pathophysiological conditions. Studies have shown that such models maintain inter- and intratumor heterogeneity and accurately reconstruct 3D tissue structures through cell-cell and cell-matrix interactions [18]. For example, PDOs retain critical tumor biological traits and can reconstruct the tumor microenvironment (TME) through co-culture systems, thereby overcoming the limitations of 2D culture—such as loss of spatial information—and the species-specific discrepancies inherent in animal models [24,25].
In translational medicine, PDOs demonstrate remarkable clinical predictive power. Numerous studies have used organoids for drug screening, development, and personalized assessment. For instance, organoid models can predict clinical responses to therapies for gastroesophageal, colorectal, and breast cancers with high accuracy [26,27]. They are also used to evaluate the efficacy of structurally similar compounds and the synergistic effects of drug combinations [28]. Notably, the concordance between drug sensitivity testing in organoids and actual patient treatment responses exceeds 80% [29,30], outperforming traditional animal models in terms of predictive accuracy [31]. Furthermore, by integrating 3D printing and organ-on-a-chip technologies, organoid models can aid in surgical planning and lesion localization, further promoting clinical translation [32].
2.2. Organ-on-a-Chip Systems
Organ-on-a-chip technology is an advanced, microfluidic-based in vitro model designed to overcome the limitations of conventional organoids in mimicking the in vivo microenvironment. Through the culture of cells and tissues within microscale channels, this technology enables precise control over physiological conditions such as fluid flow, mechanical forces, and tissue-tissue interactions, thereby more accurately recapitulating organ-level physiological and pathological states than other models [33]. Organ-on-a-chip systems not only retain the genetic properties of the parent tissues but also integrate multiple tissue interactions and dynamic microenvironments, providing more reliable outcomes for drug evaluation. These systems have been successfully established for various tissues, including the liver, intestines, skin, and reproductive system, demonstrating broad prospects as critical tools for drug screening and biological analysis [34,35,36,37].
In terms of pathophysiological simulation, organ-on-a-chip platforms use microfluidic perfusion systems to emulate biophysical and biochemical stimuli such as vascular perfusion, mechanical stress, and concentration gradients. These capabilities support gas exchange, metabolic activity, and tissue-specific functions. For instance, lung chips can be used to model targeted viral infection processes and evaluate drug effects on alveolar-capillary barrier function [38], whereas cardiovascular chips can be used to study cellular communication disorders and oxidative stress responses in ischemia-reperfusion injury [39]. Moreover, by incorporating patient-specific cells and 3D-bioprinted anatomical structures, such chips can be used to establish pathology models closely linked to clinical phenotypes, significantly increasing their translational value [40].
A key challenge and focus in the development of organ-on-a-chip technology is the construction of vascular and immune microenvironments. Since most tissues require vascular networks to maintain nutrient supply and waste removal when they exceed 400 μm in thickness in vitro, researchers are dedicated to developing strategies for achieving functional vascularization within chips [41,42]. Examples include inducing endothelial sprouting and directed migration through mechanical and biochemical signals in hydrogels or co-culturing fibroblasts and tumor cells to form vascularized microtumors [43,44]. Moreover, progress has been made in simulating immune microenvironments—for instance, using microfluidic devices to investigate interactions between tumor biopsy tissues and infiltrating lymphocytes dynamically, providing new platforms for immunotherapy research [45,46].
To improve resource utilization and model reproducibility, novel technologies such as droplet microfluidics have been introduced into the organ-on-a-chip field. These tools enable high-throughput and standardized organoid culture through the generation of uniform droplets, significantly increasing sample utilization efficiency and experimental throughput [47]. Furthermore, gradient generators integrated into chip designs support the efficient evaluation of multi-concentration drugs and combination therapies. In the future, integrating chips into automated multi-well plate platforms is expected to achieve end-to-end standardization and automation from culture to detection [48].
The integration of biosensing and imaging technologies further increases the analytical power of organ-on-a-chip systems [49]. Electrochemical sensors enable real-time monitoring of key parameters such as pH [50], oxygen partial pressure [51], and glucose levels [52], dynamically reflecting the metabolic status of organoids. Advanced bioimaging techniques allow for high-resolution, non-destructive visualization of structure and function, revealing internal dynamic changes within organoids [53]. Combining multi-organ chip systems with bioprinting and sensing technologies can simulate whole-body drug metabolism processes, overcoming the limitations of single-organ models and advancing personalized medicine and new drug development [54]. With significant advantages in simulating human physiology and drug responses, organ-on-a-chip technology is poised to play an increasingly central role in precision medicine and translational research through further integration of engineering innovations and bioinformatic methods [55].
2.3. Organoid Revolution
Organoid technology has emerged as a revolutionary breakthrough in biomedicine, providing a powerful tool to bridge the gap between traditional tumor models and clinical research [11]. Conventional drug development relies heavily on in vitro cell lines and animal models, which have significant limitations: On the one hand, the drug development process is time-consuming and costly, taking, on average, more than ten years from discovery to market approval, with substantial resources expended during animal testing and clinical trial phases [56]; on the other hand, these models fail to accurately mimic the heterogeneity of human tumors, the complexity of the tumor microenvironment (TME), and authentic therapeutic responses, leading to high clinical failure rates among drug candidates. For example, approximately 86% of liver cancer drugs fail in clinical trials, largely because of interspecies physiological differences and inadequate recapitulation of the TME [57].
As three-dimensional (3D) culture systems, organoids preserve the histoarchitectural features, genomic profiles, and cellular heterogeneity of primary tumors, significantly improving predictive accuracy for drug sensitivity and disease mechanisms [13]. In the case of liver cancer, PDOs not only simulate glandular structures and specific genetic mutations but also replicate liver-specific drug metabolism functions, thereby overcoming the predictive biases inherent in animal models due to metabolic disparities [58]. Studies have shown that the drug sensitivity prediction accuracy of organoids is greater than 35%. Their advantages include precise retention of individualized mutational profiles, support for co-culture systems to reconstruct fibrotic and immune microenvironments, and the enabling of efficient drug screening and mechanistic studies.
In addressing species-specific differences, organoids demonstrate three major breakthroughs: First, functional substitutability—for instance, human liver organoids successfully simulate hepatitis C virus (HCV) infection-induced transcriptional reprogramming, a process that cannot be replicated in mouse models [59]. Second, dynamic monitoring capabilities—vascularized organoid-on-a-chip platforms—allow for real-time tracking of drug distribution kinetics, with vascular leakage features being significantly more strongly correlated with clinical imaging data than they are in animal models [60]. Third, immune interaction modeling-co-culture systems of organoids with tumor-infiltrating lymphocytes (TILs) achieve an AUC value of 0.92 in predicting the response to PD-1 inhibitors and are markedly superior to xenograft models [61,62].
Technological innovations are further advancing organoid standardization and clinical translation. Novel micro-patterned agarose scaffolds enable the generation of up to 8,000 homogeneous organoids in a single batch, increasing the high-throughput screening capacity by 20-fold [63]. Nanofiber hydrogel culture systems increase the expression of drug metabolism genes by 4.2-fold, more closely resembling human physiological conditions than other models [64]. These advancements have prompted numerous pharmaceutical companies to incorporate organoids into their preclinical evaluation systems.
The clinical translation pathway for organoids is now approaching a closed-loop process: biopsy samples can be used to establish PDO biobanks within 2–3 weeks, enabling simultaneous genomic sequencing and drug library screening [65]. Clinical studies have demonstrated that organoid-guided treatment strategies significantly improve objective response rates, with drug resistance prediction specificity reaching 91%, thereby avoiding ineffective treatments and reducing individual medical costs [66]. Although challenges remain in vascularization and immune microenvironment modeling, the integration of novel technologies such as organ-on-a-chip systems, single-cell sequencing, and CRISPR screening is continuously driving progress [67]. With the ongoing refinement and interdisciplinary integration of organoid culture technologies, it is anticipated that within three years, the use of organoids will enable the full-course prediction of individualized therapies, shorten drug development cycles from 12 years to 7 years, and ultimately usher in an “organoid-guided era” of precision medicine [68].
3. Advancing Organoid Technology: Culture and Quality Control
3.1. Breakthroughs in the Core Culture Technology of Organoids
Owing to its undefined composition and batch-to-batch variability, conventional mouse-derived Matrigel has become a major limiting factor in organoid standardization [69]. In recent years, synthetic hydrogel systems have enabled biomimetic reconstruction of the spatiotemporal distribution of bioactive ligands [70]. These materials promote the progression of the extracellular matrix microenvironment through precise control of mechanical properties, viscoelasticity, and adaptable physicochemical characteristics and can incorporate growth factor sustained-release systems, significantly improving culture reproducibility and long-term stability. Notably, by preserving tissue-specific ECM components, decellularized extracellular matrix (dECM) hydrogels support the polarized growth and functional maturation of liver and kidney organoids, with gene expression profiles showing greater than 85% similarity to those of in vivo tissues [71].
Microfluidic organ-on-a-chip technology addresses the core limitation of nutrient/oxygen gradient diffusion in static culture by constructing 3D microvascular networks for co-culture with organoids [72]. For example, co-culturing human umbilical vein endothelial cells (HUVECs) with organoids on a chip can result in the formation of functional vasculature within seven days, increasing renal organoid filtration function threefold [73]. Moreover, functionalized scaffolds guide the directed extension of dorsal root ganglion neurons and improve the efficiency of synaptic formation [74,75,76]. These advances propel organoids from a cellular scale toward a tissue-level model, offering new paradigms for drug permeability testing and neurological disease modeling.
Microfluidic technology simulates in vivo physiological conditions through dynamic perfusion systems, significantly increasing organoid survival and functional maturation. Multi-organ-chips achieve functional coupling of organoids via fluidic interconnection, modeling systemic drug pharmacokinetics and toxicity responses [54]. For instance, intestinal absorption followed by hepatic metabolism and renal excretion of a drug can be continuously monitored on-chip, providing a more accurate toxicity assessment model for drug development [77].
To address issues of organoid size/morphology heterogeneity and low throughput associated with manual operation, droplet-based microfluidics enables large-scale standardized production by generating uniformly sized organoid precursor droplets. Bioreactor-based large-scale suspension culture under controlled agitation significantly improves yield and batch-to-batch consistency [78]. Automated platforms integrating robotic liquid handling and AI-driven image analysis achieve end-to-end automation from culture and drug dosing to phenotypic detection, greatly increasing experimental reproducibility and throughput [79].
Single-cell multi-omics technologies allow for the systematic resolution of cellular heterogeneity and differentiation trajectories in organoids, providing molecular guidance for culture optimization [80]. CRISPR-Cas9 technology enables precise editing of the organoid genome, and genome-wide gRNA library screening identifies key genes involved in tumorigenesis and cell fate determination [81]. Integrating multi-omics data with CRISPR screening results in the establishment of a complete genotype-to-phenotype validation pipeline, offering functional evidence for precision medicine [15].
Label-free techniques such as optical coherence tomography (OCT) and Raman spectroscopic imaging enable real-time monitoring of organoid morphology, structure, and metabolic status [82]. The integration of microfluidic chips with biosensors allows for real-time detection of key metabolic indicators, including glucose levels, oxygen levels, and pH. For example, wide-field optical redox imaging measures the autofluorescence intensities of NADPH and FAD to assess the treatment response in organoids without labels [83].
Transplantation of human organoids under the renal capsule or subcutaneously into immunodeficient mice promotes vascularization and functional maturation through the host environment [84]. Xeno-symbiotic models provide a unique platform for studying interactions between human cells and the host microenvironment, although they require concomitant immunosuppression to mitigate rejection [85] (Figure 2). In summary, breakthroughs in organoid culture technology span matrix design, microenvironment construction, scalable production, functional analysis, and real-time monitoring. Together, these advances are transforming organoids from simple 3D aggregates into highly physiologically relevant microphysiological systems, providing powerful tools for disease modeling, drug screening, and personalized medicine [86].
3.2. Quality Assessment and Standardization of Organoids
The reliability and reproducibility of organoid technology largely depend on the refinement of its quality assessment system and the implementation of standardized practices [15]. A comprehensive and systematic quality evaluation must cover three core dimensions—morphology, function, and genetic stability—to ensure that organoids accurately mimic the physiological and pathological characteristics of the source tissue, thereby providing a robust and reliable platform for scientific research and clinical translation [87].
Morphological assessment is the primary step in evaluating organoid quality and focuses on the fidelity of three-dimensional structure, size uniformity, and cellular composition [87]. Histological staining and light microscopy can be used to determine whether organoids form organ-specific spatial structures, such as crypt-villus units in intestinal organoids [12], tubule-like structures in kidney organoids [88], or layered features in brain organoids [89]. Quantitative imaging techniques can be used to measure the diameter, volume, and luminal ratio systematically and calculate within-batch and between-batch coefficients of variation to assess culture stability [47]. Immunohistochemistry or immunofluorescence is used to verify whether the spatial distribution of key cell types faithfully recapitulates the original tissue architecture [81]. Standardization efforts should focus on establishing organ-specific morphological scoring systems and minimum structural thresholds as well as promoting automation and uniformity in imaging analysis pipelines [90].
Functional assessment is critical for evaluating the biological activity and predictive value of organoids [11]. Organ-specific functional markers must be employed: liver organoids should be assessed for albumin secretion, urea synthesis, and cytochrome P450 enzyme activity [58], and intestinal organoids should be evaluated for mucus secretion, hormone response, and epithelial barrier function [91]. Additionally, drug response tests should demonstrate dose-dependent effects consistent with clinical patient responses to support personalized drug screening. Electrophysiological properties, such as rhythmic beating in cardiac organoids and synchronized neuronal firing in brain organoids, are also important evaluation criteria [92,93]. Standardization practices should involve defining organ-specific functional test panels and establishing activity thresholds for key indicators [94].
Genetic stability is the cornerstone of long-term culture and the reliable application of organoids, particularly for tumor organoids and long-term passaged models [13]. Whole-exome sequencing can be used to monitor accumulated copy number variations and single-nucleotide variants during passaging, whereas single-cell DNA sequencing helps reveal subclonal evolution [95]. For tumor organoids, regular validation of the consistency of the driver gene mutation status with that of the primary tumor is essential [96]. Epigenetic stability, such as passaging-induced drift in DNA methylation profiles, should also be systematically evaluated [97]. Standardization efforts should include clearly defining maximum passage numbers, establishing intervals for regular genetic monitoring, and employing highly sensitive methods for the quantitative tracking of critical mutations [78].
To advance the standardization of organoid technology, an international consensus framework is needed. First, experimental protocols—including tissue dissociation, matrix selection, culture medium formulation, and passaging methods—should be standardized through unified standard operating procedures to increase reproducibility [98]. In data reporting, the FAIR principles (findable, accessible, interoperable, and reusable) should be adopted to ensure complete disclosure of culture conditions, passage history, and analytical workflows, thereby promoting data reusability and transparency [99]. Furthermore, a tiered quality certification system should be established to distinguish between “research-grade” and “clinical-grade” organoids, accommodating differential quality requirements across various application scenarios [100]. In summary, organoid quality assessment is a multidimensional and systematic endeavor that relies on the integration of morphological, functional, and genetic indicators [101]. The current core challenge lies in defining organ-specific “minimum essential indicator sets” and validating their effectiveness through multicenter collaborations [102]. Moving forward, there is an urgent need to develop automated, high-throughput quality control platforms to facilitate the transition of organoids from experimental tools to standardized biological reagents.
4. Clinical Translation of Organoids
4.1. Application of Organoids in Clinical Diagnostics
As three-dimensional miniature organ models, organoids demonstrate two core values in clinical diagnostics: assisting pathological subtyping and developing early diagnostic biomarkers. In pathological subtyping, organoids serve as critical tools for personalized treatment decision-making by preserving the histological features and molecular heterogeneity of primary tumors (Figure 3). For example, in colorectal cancer, patient-derived organoids (PDOs) can simulate clinical chemotherapy responses through in vitro drug sensitivity testing. Studies have shown that the variation in the sensitivity of organoids to oxaliplatin/5-FU regimens closely aligns with actual patient treatment responses [103]. Such “organoid drug sensitivity profiles” not only guide chemotherapy selection but also help identify primarily resistant subpopulations—for instance, compared with wild-type organoids, TP53-mutant organoids exhibit significantly greater resistance to irinotecan [104]. Furthermore, the abnormal bile acid metabolism observed in liver cancer organoids provides a quantitative indicator for pathological grading [105], whereas after their morphological consistency with that of primary tumors is validated via H&E staining, gastric cancer organoids can assist in Lauren classification [106]. More cutting-edge explorations integrate artificial intelligence into organoid manufacturing processes and use deep learning to analyze correlations between organoid morphological features and pathological subtypes, enabling the automated classification of ovarian cancer organoids [107].
In the field of early diagnostic biomarker development, organoid-derived biomarkers demonstrate unique advantages. Brain organoid models reveal early abnormal tau protein phosphorylation patterns in Alzheimer’s disease, offering a novel in vitro validation platform for the ATN(I) classification system [108]. Notably, compared with traditional cell models, exosomes secreted by organoids carry organ-specific molecular signatures: the combination of miR-21-5p and miR-100-5p in intestinal organoid exosomes can differentiate Crohn’s disease from ulcerative colitis [109], while the EMT-related miRNA profile in thyroid cancer organoid exosomes can predict tumor invasiveness earlier than traditional cell models [110]. Kidney organoid studies have shown that injury markers dynamically induced by esculentoside A strongly correlate with those in preclinical animal models, confirming that these organoids can serve as reliable screening systems for nephrotoxicity biomarkers [111]. Through reconstructing biliary networks, liver organoids have for the first time captured the early biliary epithelial marker ANXA4, which is associated with primary sclerosing cholangitis, in an in vitro setting [112]. These findings support the use of organoids from mere disease models to “biomarker incubators,” as their three-dimensional microenvironments more authentically simulate in vivo intercellular communication and paracrine regulation [15].
Technological integration further expands diagnostic applications. CRISPR-edited knockout organoids can rapidly validate the pathological importance of candidate biomarkers, while microfluidic chip-cultured vascularized organoids enable dynamic monitoring of the secretion patterns in metastasis-related markers [113]. Through proteomic analysis, recent advances in brain injury organoids have revealed novel biomarker combinations, such as GFAP/IL-8, whose diagnostic efficacy surpasses that of traditional imaging-based classification [114]. Despite challenges such as insufficient vascularization, the diagnostic value of organoids in precision medicine is widely recognized—they not only serve as “living biobanks” that preserve patient-specific pathological features but also accelerate translational research through high-throughput screening [115]. With the integration of technologies such as single-cell sequencing, organoid-driven “spatiotemporal omics diagnostics” are expected to reshape clinical practice paradigms in the future [116].
4.2. Applications of Organoids in Disease Modeling
Organoid technology has emerged as a vital tool in disease modeling, demonstrating strong application potential across multiple medical fields. Its core value lies in its ability to simulate the structure and function of human organs strongly, providing a precise platform for mechanistic studies, drug screening, and personalized medicine [15]. In oncology, patient-derived organoids (PDOs) preserve the heterogeneity, tissue architecture, and molecular characteristics of primary tumors through three-dimensional culture, making them key models in cancer research [13]. In colorectal cancer, PDOs can successfully predict patient resistance to chemotherapeutic agents such as irinotecan and oxaliplatin with an accuracy exceeding 85%. In breast cancer, PDO models based on ER/PR/HER2 expression profiles can guide combined strategies of endocrine therapy and targeted drugs [27]. Liver cancer organoids not only recapitulate the histological features of tumors but also simulate the evolution of drug resistance following sorafenib treatment, thereby informing personalized treatment plans [58]. Furthermore, the integration of organoids with microfluidic chips increases drug permeability and microenvironment controllability, significantly strengthening their utility in anticancer therapy development [113].
Organoids have shown outstanding performance in the study of genetic disease mechanisms and validating treatments. In cystic fibrosis (CF), bronchial organoids differentiated from induced pluripotent stem cells (iPSCs) have been successfully used for functional validation of CFTR gene repair and for evaluating the efficacy of CFTR modulators through ion transport assays [117]. In neurodevelopmental research, brain organoids have overcome the limitations of traditional models: Zika virus infection models replicate the process of virus-specific attacks on neural progenitor cells, leading to microcephaly, and confirm the protective role of interferon lambda-1 [89]. Furthermore, brain organoids with SHANK3 gene mutations associated with autism exhibit abnormal synapse formation, offering new insights into disease mechanisms [118]. Organoids are also used to model rare diseases such as Rett syndrome and alpha-1 antitrypsin deficiency. When combined with CRISPR-Cas9 gene editing, they enable the exploration of pathogenic mechanisms and the validation of treatment strategies [119].
Organoids provide physiologically relevant models for studying host-pathogen interactions in infectious diseases. The mechanism through which the rotavirus invades epithelial cells by disrupting the tight junction protein ZO-1 [120] has been revealed in intestinal organoids, while the pathway through which the Helicobacter pylori CagA protein induces apoptosis has been confirmed in gastric organoids. In COVID-19 research, type II alveolar cells have been identified as primary targets of SARS-CoV-2 infection in alveolar organoids because of their high ACE2 receptor expression, and it has been demonstrated that viral replication can be inhibited by remdesivir [121]. Co-culture systems of lung organoids and immune cells have successfully simulated cytokine storms, providing a standardized platform for screening anti-inflammatory drugs [122]. Additionally, the mechanism of norovirus invasion via the CD300lf receptor [123] has been elucidated in intestinal organoids, and the dynamic process of Zika virus transmission along synapses leading to microcephaly has been replicated in brain organoids [124].
Liver organoids have proven highly valuable in metabolic studies of non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH). The induction of fatty acid overload recapitulates typical pathological features such as lipid accumulation, inflammatory responses, and the upregulation of the fibrosis marker α-SMA [125]. In diabetes research, through glucose-stimulated insulin secretion (GSIS) assays, the detrimental effects of SLC30A8 gene variants on β-cell function have been confirmed, and the proliferative effects of GLP-1 receptor agonists have been evaluated [126]. The development of vascularized islet organoids has further improved the physiological relevance of insulin secretion, increasing the predictive power of the model [127].
4.3. Application of Organoids in Therapeutic Development and Personalized Medicine
Organoid technology is rapidly emerging as a pivotal tool in therapeutic development and personalized medicine. By closely mimicking the physiological and pathological features of human organs, it provides a robust platform for drug screening, toxicity testing, and individualized treatment strategies. Owing to their three-dimensional architecture and retention of original tissue functions, organoids have been widely adopted to establish a new generation of high-throughput drug screening platforms. For instance, lung cancer organoids cultured under standardized conditions enable rapid drug screening, with results that strongly correlate with clinical responses [128]. Organoid-based screening models not only improve the predictive accuracy but also significantly reduce the reliance on animal testing in drug development. In fact, the U.S. FDA has recognized organoids as an alternative to animal models for preclinical drug sensitivity testing [129]. This approach is equally applicable to rare disease research—such as using patient-specific brain organoids to screen potential drugs for mitochondrial disorders [130] or employing iPSC-derived intestinal organoids to model infections and identify antimicrobial agents—demonstrating broad potential across diverse disease contexts [131].
In the context of drug metabolism and toxicity evaluation, liver organoids accurately express drug-metabolizing enzymes such as those in the CYP450 family, substantially increasing the reliability of hepatotoxicity prediction [132]. Recent studies have shown that biliary organoids can be used to assess drug-induced bile duct injury, effectively addressing a gap in traditional toxicology models [112]. Similarly, by reconstituting tubule function, kidney organoids enable sensitive detection of nephrotoxic biomarkers such as KIM-1, with data showing a correlation of greater than 85% with clinical kidney injury [133]. These advances underscore the central role of organoids in drug safety assessment and hold promise for considerably reducing the use of animal experiments [33].
Patient-derived organoids (PDOs) retain the heterogeneity and microenvironmental features of primary tumors, offering direct evidence for personalized treatment selection. For example, gastric cancer PDOs can be subject to complete sensitivity testing of chemotherapeutic agents such as 5-FU within 48 hours, with results showing up to 92% concordance with actual patient treatment responses [30]. In colorectal cancer, PDOs outperform genetic testing methods in predicting resistance to irinotecan or oxaliplatin [28]. Moreover, co-culture systems combining organoids and immune cells provide a novel tool for evaluating immunotherapy efficacy, enabling the prediction of PD-1 inhibitor efficacy by simulating tumor-immune interactions—thereby supporting patient stratification and treatment optimization [134]. Representative cases have demonstrated that the response of metastatic colorectal cancer PDOs to the FOLFOX regimen significantly correlates with patient progression-free survival (PFS) [135]. In organoids resistant to oxaliplatin, overexpression of the ERCC1 gene has been observed, which is consistent with findings in patient clinical samples. Such studies not only validate the utility of PDOs as “patient avatars” but also indicate their potential to help in avoiding ineffective treatments, thereby reducing health care costs [136].
Organoids provide a highly physiologically relevant platform for functional validation in gene editing and cell therapy. In genetic disease research, intestinal organoids are used to evaluate the efficacy of CFTR gene repair, in which the lumen formation capacity directly reflects the functional recovery of the channel [137]. In diabetes research, the CRISPR-Cas9-mediated knockout of the GLP-1R gene in pancreatic organoids revealed the critical role of the receptor in β-cell regeneration [126]. Furthermore, liver cancer organoids successfully model tumor evolution through targeted editing of driver genes such as TP53, offering an ideal system for developing targeted gene therapies [138].
Organoids also play important roles in evaluating the safety of cell therapies. For example, bone marrow organoids can simulate cytokine release syndrome (CRS) induced by CAR-T-cell therapy, with IL-6 secretion levels being significantly correlated with clinical toxicity grading in patients [139]. Brain organoids are used to assess the potential off-target effects of GD2-CAR-T cells on normal neural tissue, providing key insights for treatment safety and optimization [140]. These applications considerably shorten drug development timelines and increase the predictive power of preclinical evaluation.
4.4. Barriers in Translation
While organoid technology has demonstrated significant potential in biomedical research and clinical translation, its development and application continue to face a series of technical and practical challenges. These challenges primarily revolve around physiological relevance, standardization and reproducibility, cost and clinical integration barriers, as well as ethical and regulatory considerations [15]. First, most current organoid systems lack functional vascular networks and a complete immune cell composition, limiting their ability to mimic the true in vivo microenvironment. The absence of a vasculature results in insufficient nutrient supply and hypoxia in core regions, restricting organoid growth and functional maturation, such as impaired filtration in kidney organoids. Although strategies such as hypoxia-inducible factor (HIF-1α) modulation, bioprinting, or co-culture with endothelial cells can partially induce vascularization, the perfusion efficiency and stability of these structures remain far from physiological levels. Moreover, most existing organoid models fail to incorporate immune components, making it difficult to recapitulate key pathophysiological processes such as tumor immune evasion or chronic inflammation [141]. While technologies such as microfluidic chips can simulate hemodynamic forces and improve microenvironment control, the spatial distribution and dynamic recruitment mechanisms of immune cells still require further optimization [33].
Second, the reproducibility of organoid cultures is affected by variability in cell sources, medium composition, matrix materials, and differentiation protocols, leading to significant differences between batches—even within the same laboratory [142]. For instance, regionalized assembloids of brain organoids still lack unified operational protocols and quality assessment criteria. Standardization challenges span multiple aspects, including culture systems, environmental control, and functional evaluation [94]. Although artificial intelligence can provide data-driven optimization of culture protocols, the absence of international consensus standards severely limits data comparability and clinical translatability. Technical fluctuations in automated large-scale culture—such as variations in 3D printing precision and fluid control stability—further exacerbate batch inconsistency.
Lastly, the specialized culture media, cytokines, and advanced equipment required for organoid construction and maintenance result in high costs, but extended culture periods limit their utility in acute disease research. From a clinical translation perspective, organoid technology faces the following dual hurdles: technically limited scalability and unvalidated systemic integration with host tissues [112]. From a regulatory standpoint, organoids lack unified good manufacturing practice (GMP) production standards, efficacy evaluation systems, and regulatory frameworks—whether intended as therapeutic products or screening tools. Additionally, the application of organoids in pharmacokinetic-pharmacodynamic (PK-PD) modeling remains challenging because of their systemic complexity [143].
As the complexity of organoids increases, ethical concerns are gaining attention. For example, brain organoids exhibiting spontaneous neural electrical activity have sparked debate regarding the boundaries of “consciousness” and their moral status [144]. While the application of gene editing technologies in organoids facilitates disease mechanism research, potential misuse—such as germline editing or chimera generation—also raises ethical risks [145]. Furthermore, patient-derived organoids involve issues such as individual privacy protection, commercial use rights of samples, and species boundary considerations in animal transplantation. There is an urgent need for interdisciplinary dialog to establish the corresponding ethical guidelines and legal regulations [146].
On the regulatory front, organoid technology faces outdated frameworks and a lack of standards. Internationally, there is still no clear classification or approval pathway for organoid-based products, whether used as disease models, drug screening tools, or therapeutics [55]. Clinical validation of companion diagnostic organoids is also progressing slowly because of the absence of unified analytical performance standards. Advancing the standardization and clinical translation of organoid technology requires the establishment of an internationally coordinated regulatory science system and the adoption of mechanisms such as “regulatory sandboxes” to accelerate safe and effective application [147]. In summary, realizing the full clinical potential of organoid technology will require further breakthroughs in vascularization and immune modeling, standardization, cost reduction, and ethical-regulatory alignment. Multidisciplinary collaboration and the integration of innovative technologies are essential for addressing these challenges.
5. Future Directions: Integration of AI and Sequencing Technologies
5.1. AI-Powered Organoid Analysis
Although organoid technology has made significant strides in in vitro culture, physiological simulation, and drug evaluation—as driven by advanced platforms such as microfluidics—the complexity of growth and differentiation mechanisms, drug response pathways, and associated data analysis challenges remain major bottlenecks hindering clinical translation. The deep integration of artificial intelligence (AI) with organoid models is promising for substantially increasing the efficiency and reliability of organoid construction, phenotypic interpretation, and clinical application [148].
Organoid research often relies on techniques such as multiplex immunofluorescence imaging for functional assessment. However, the inherent multifocality, heterogeneity, and three-dimensional structural complexity of organoids make it difficult for traditional image analysis methods to achieve rapid and accurate spatial feature extraction and quantification [81]. AI methods, particularly deep learning-based frameworks such as convolutional neural networks, provide effective tools for high-throughput, automated morphological quantification and pharmacodynamic evaluation of organoids [149]. For example, combined with bright-field or label-free optical coherence tomography (OCT) imaging, machine learning models enable non-invasive, real-time monitoring and quantitative analysis of dynamic changes in organoids, significantly improving the objectivity and reproducibility of experimental results. Generative adversarial networks (GANs) further support the reconstruction of fine 3D spatial structures from unlabeled images, avoiding sample damage caused by traditional staining methods and enabling long-term experiments [150].
Beyond morphological analysis, AI has demonstrated a strong ability to integrate multimodal data from organoids. The massive amount of multi-omics data generated during organoid experiments requires efficient and standardized analytical tools. AI methods can be used to combine these datasets to identify cellular subpopulation heterogeneity within organoids, to track functional evolution trajectories, and to correlate molecular and phenotypic responses following drug treatment [151]. Furthermore, AI-driven quality control modules help improve the stability and reproducibility of organoids, providing support for their standardization and clinical translation [152].
In the context of personalized medicine, the linkage between organoids and patient clinical data has opened new paradigms for disease modeling and treatment prediction. AI-predictive models integrate multi-omics features of organoids with patient clinical information to construct patient-specific drug response profiles [153]. Machine learning algorithms can identify biomarkers of drug sensitivity and predict the efficacy of chemotherapy or targeted therapy, which is particularly valuable in studies of highly heterogeneous cancers or rare diseases. Furthermore, reinforcement learning frameworks can simulate dynamic treatment scenarios and iteratively optimize dosing strategies on organoid platforms, thereby shortening drug screening cycles and reducing reliance on animal testing and costly clinical studies. Notably, the U.S. FDA has introduced new guidelines allowing certain drugs to bypass animal testing before entering clinical trials—a policy shift that further underscores the importance of organoids and organ-on-a-chip models as alternative systems. The integration of AI with organ-on-chip technology will enable real-time monitoring and closed-loop feedback, allowing for the dynamic adjustment of treatment strategies and advancing precision medicine from “static prediction” to “dynamic intervention”. In the future, as single-cell sequencing costs decline and edge computing technologies advance, distributed AI systems are expected to enable the real-time analysis of organoid data and clinical decision support [154]. Overall, the synergistic innovation of AI and organoid technology will profoundly transform existing paradigms in disease modeling, drug development, and personalized therapy, accelerating the translation from basic research to clinical applications.
5.2. Application of Organoid Technology in Single-Cell and Long-Read Sequencing
Organoid technology, when integrated with single-cell and long-read sequencing, demonstrates unprecedented spatiotemporal resolution, with its core value reflected in two key dimensions: dynamic monitoring and multi-omics integration. In terms of dynamic monitoring, real-time genomic analysis based on long-read sequencing technologies such as Nanopore is revolutionizing the application of organoid models in studying tumor evolution. For example, clonal dynamics in patient-derived tumor organoids under chemotherapeutic pressure can be captured using single-cell whole-genome sequencing to detect low-frequency drug-resistant mutations. The unique continuous reading capability of long-read sequencing enables the resolution of structural variations and the coordinated evolution of epigenetic modifications. Studies have confirmed that microfluidic chip systems can automate the entire process of organoid culture, digestion, and single-cell isolation. When combined with real-time Nanopore sequencing, this approach allows the accumulation of mutations in genes associated with chemotherapy resistance to be tracked, with single-base resolution even identifying subclonal-level chimeric mutations [155]. Furthermore, using combinatorial indexing strategies, dynamic single-cell multi-omics monitoring simultaneously captures transcriptomic and chromatin accessibility data, revealing spatiotemporal correlations between WNT/β-catenin pathway activation and open chromatin regions in drug-resistant cells [156]. This dynamic perspective provides a cellular molecular clock for understanding the evolution of tumor heterogeneity.
At the multi-omics integration level, the synergistic application of spatial transcriptomics and single-cell sequencing overcomes the spatial limitations associated with studying microenvironmental interactions within organoids. For instance, spatial transcriptomics can locate specific niches in intestinal organoids that interact with symbiotic microorganisms, while the combined use of single-cell immunomics and microbiome analysis deciphers the negative feedback mechanism between antimicrobial peptide secretion by goblet cells and the spatial distribution of the microbiota [157]. State-of-the-art algorithms, which integrate single-cell transcriptomics, chromatin accessibility, and spatial proteomics data through deep learning frameworks, have successfully mapped metabolic-immune interaction networks specific to liver lobule zones in hepatic organoids [158]. Notably, multi-omics integration has also revealed conservation differences in developmental regulation: single-cell multi-omics analysis of human kidney organoids revealed that, compared with those in adult kidneys, renal tubular cells in organoids exhibit broader open chromatin regions and fetal-like enhancer-promoter interaction patterns—findings functionally validated in proximal tubule cell lines via CRISPR interference experiments. These findings suggest that organoids may more closely resemble developing tissue states than mature tissue states [159].
Technical challenges and solutions coexist. In dynamic monitoring, distinguishing endogenous mutations in organoids from artificial variations introduced during in vitro culture requires paired normal tissue sequencing. Multi-omics integration faces issues such as data sparsity and batch effects, which emerging algorithms address by using generative adversarial networks to simulate real data distributions and optimize analytical workflows [152]. In the future, single-cell multi-omics platforms combining synthetic biology and live-cell imaging are expected to enable four-dimensional reconstruction of organoid development and pathological processes. These advances will propel organoids from static models to dynamic systems, ultimately serving the personalized predictive needs of precision medicine [48].
6. Future Perspectives
Organoid technology is currently at a critical juncture, transitioning from basic research to clinical translation. Its future development will heavily rely on interdisciplinary innovation, the establishment of standardized systems, and deeper integration into clinical applications. Although challenges remain, this technology has the potential to reshape existing paradigms in disease research, drug development, and precision medicine [142]. Further advancements in organoid technology will require deep integration with engineering, computational science, and biotechnology. Artificial intelligence and machine learning will significantly enhance the analysis of high-throughput imaging and single-cell multi-omics data from organoids, enabling automated phenotypic recognition, drug response prediction, and mechanistic exploration. The integration of organ-on-a-chip and biosensing technologies will allow for the simulation of interorgan interactions and complex microenvironments, while real-time monitoring of metabolic activity, oxygen consumption, and electrophysiological parameters will provide critical data for dynamic pharmacodynamic evaluation [33]. Future efforts should be focused on developing more biocompatible biomimetic scaffold materials and addressing the integration of cross-scale, multi-modal data to improve both the physiological relevance of organoids and the quality of the data generated.
Clinical translation of organoids urgently requires internationally unified culture standards, quality evaluation systems, and regulatory frameworks. Regulatory agencies have begun recognizing the value of organoids in drug toxicity testing and personalized medicine—for instance, using standardized organoid protocols for hepatotoxicity assessment. Promoting industrialization will depend on addressing three core issues: the standardization of culture processes, the construction of quality control systems, and the development of large-scale production technologies [129]. Collaboration among industry, academia, and research institutions is key to reducing costs, accelerating technology adoption, and establishing international consensus guidelines.
As foundational infrastructures for precision medicine, organoid biobanks are already playing important roles in various cancer fields [115]. Optimizing cryopreservation and revival techniques, along with establishing sample-sharing platforms, will expand their coverage of diseases and populations. The combination of dynamic intervention systems and real-time monitoring technologies is expected to enable precise regulation and continuous monitoring of organoid function, providing closed-loop feedback for individualized therapies [48]. However, insufficient vascularization and a lack of immune microenvironments remain major technical bottlenecks, necessitating solutions through co-culture systems, tissue engineering, and organ-on-a-chip technologies [112].
In the future, organoids are expected to serve as “living biological reagents,” fundamentally transforming research approaches to disease modeling, drug development, and regenerative medicine [87]. Their value will be demonstrated in three main aspects: replacing animal models to enable human-specific disease studies; integrating multi-omics data to reveal disease mechanisms and predict therapeutic targets; and serving as cell sources for tissue repair in regenerative medicine [80]. Ultimately, organoid technology will drive the transition of medicine toward an integrated precision system encompassing “patient-organoid-clinical” synergy. However, as organoid complexity increases, ethical concerns regarding the boundaries of “consciousness” and the implications of gene editing are becoming increasingly prominent, underscoring the urgent need for interdisciplinary collaboration to develop corresponding ethical guidelines and regulatory frameworks [48,160].
7. Conclusions
As three-dimensional physiologically relevant models, organoid technology has significantly overcome the limitations of traditional two-dimensional cell cultures and animal models—such as species-specific differences and the absence of a true microenvironment—by closely mimicking the structural complexity, cellular heterogeneity, and microenvironmental features of human organs. This breakthrough has successfully bridged a critical gap between basic research and clinical translation. Organoids demonstrate exceptional capabilities in recapitulating tumor pathological behavior and patient-specific drug responses, providing a highly reliable platform for predicting drug efficacy and toxicity. For instance, colorectal cancer organoids can accurately replicate individual tumor biology, thereby accelerating the translation from mechanistic research to clinical intervention. Furthermore, studies involving organoid transplantation have preliminarily demonstrated their potential use in tissue repair and regenerative medicine [112].
Organoids have not only profoundly advanced our understanding of disease mechanisms but have also brought revolutionary changes to personalized medicine. Patient-derived organoids have been used to model a wide range of pathological mechanisms successfully, from genetic disorders and infectious processes to cancer progression [161]. In personalized medicine, organoid platforms enable patient-specific prediction of drug responses, guiding the optimization of clinical chemotherapy, targeted therapy, and immunotherapy regimens. Moreover, by integrating iPSC-derived models and multi-omics technologies, organoids further support end-to-end integration from disease modeling to the development of individualized treatment strategies, offering tailored solutions for rare and complex diseases [162].
Despite their considerable translational potential, the widespread clinical adoption of organoid technology still faces several challenges. Technically, insufficient standardization of culture systems leads to batch-to-batch variability, limited functional maturity, and the absence of vascularization and innervation, underscoring the urgent need to establish unified quality assessment systems across laboratories. Further advancements in organoid technology will require deep interdisciplinary integration [33]. Moreover, regulatory and industrial development remains lagging, necessitating the improvement of ethical frameworks, the promotion of clinical trial validation, and the formulation of international consensus guidelines. Moving forward, global collaboration and multidisciplinary cooperation—encompassing bioengineering, clinical medicine, and data science—will be essential to address these technical and administrative bottlenecks, ultimately enabling the scalable and standardized translation of organoids from experimental systems to clinical tools [86].
In summary, organoid technology bridges the gap between traditional preclinical models and human trials. Leveraging their high physiological relevance, organoids exhibit tremendous potential for elucidating disease mechanisms, enabling personalized drug screening, and optimizing regenerative treatment strategies. Future research should focus on multidisciplinary technology integration, the establishment of standardized systems, and the exploration of clinical translation pathways. Only by systematically addressing the core “3R” challenges—reproducibility, regulatory frameworks, and real-time monitoring—can the revolutionary role of organoids in precision medicine be fully realized, transforming them from research tools into clinical decision-support systems [55].
Author Contributions
Xue Zhifeng and Yang Runze: conceptualization, investigation, writing – original draft. Han Luo and Yaling Liu: supervision, validation, writing – review & editing. All authors have read and agreed to the published version of the manuscript.
Funding
No funding.
Acknowledgments
We thank our supervisors, [Han Luo and Yaling Liu], for their expert guidance and encouragement.
Conflicts of Interest
The authors declare no competing interests.
Abbreviations
| 2D | Two-Dimensional |
| 3D | Three-Dimensional |
| PDTO | Patient-Derived Tumor Organoid |
| iPSCs | Induced Pluripotent Stem Cells |
| ASCs | Adult Stem Cells |
| PDOs | Patient-Derived Organoids |
| TME | Tumor Microenvironment |
| HUVECs | Human Umbilical Vein Endothelial Cells |
| NVUs | Neurovascular Units |
| dECM | Decellularized Extracellular Matrix |
| OCT | Optical Coherence Tomography |
| NADPH | Nicotinamide Adenine Dinucleotide Phosphate |
| FAD | Flavin Adenine Dinucleotide |
| CF | Cystic Fibrosis |
| CFTR | Cystic Fibrosis Transmembrane Conductance Regulator |
| TILs | Tumor-Infiltrating Lymphocytes |
| NAFLD | Non-Alcoholic Fatty Liver Disease |
| NASH | Non-Alcoholic Steatohepatitis |
| GSIS | Glucose-Stimulated Insulin Secretion |
| FDA | U.S. Food and Drug Administration |
| CRS | Cytokine Release Syndrome |
| CAR-T | Chimeric Antigen Receptor T-Cell |
| HIF-1α | Hypoxia-Inducible Factor-1α |
| GMP | Good Manufacturing Practice |
| PK-PD | Pharmacokinetic-Pharmacodynamic |
| AI | Artificial Intelligence |
| GANs | Generative Adversarial Networks |
| FAIR | Findable, Accessible, Interoperable, Reusable |
| AUC | Area Under the Curve |
| siRNA | Small Interfering RNA |
| mRNA | Messenger RNA |
| DNA | Deoxyribonucleic Acid |
References
- Han, X.; Cai, C.; Deng, W.; et al. Landscape of human organoids: Ideal model in clinics and research. Innovation (Camb) 2024, 5, 100620. [Google Scholar] [CrossRef]
- Mishra, I.; Gupta, K.; Mishra, R.; et al. An Exploration of Organoid Technology: Present Advancements, Applications, and Obstacles. Curr Pharm Biotechnol 2024, 25, 1000–1020. [Google Scholar] [CrossRef]
- Song, X.; Hou, K.; Zhou, H.; et al. Liver organoids and their application in liver cancer research. Regen Ther 2024, 25, 128–137. [Google Scholar] [CrossRef]
- Tao, B.; Li, X.; Hao, M.; et al. Organoid-Guided Precision Medicine: From Bench to Bedside. MedComm 2025, 6, e70195. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, B.; Zinocker, S.; Holm, S.; et al. Organoids in the Clinic: A Systematic Review of Outcomes. Cells Tissues Organs 2023, 212, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, L.; Kirk, R. High drug attrition rates--where are we going wrong? . Nat Rev Clin Oncol 2011, 8, 189–190. [Google Scholar] [CrossRef]
- Langhans, S.A. Three-Dimensional in Vitro Cell Culture Models in Drug Discovery and Drug Repositioning. Front Pharmacol 2018, 9, 6. [Google Scholar] [CrossRef]
- Mak, I.W.; Evaniew, N.; Ghert, M. Lost in translation: animal models and clinical trials in cancer treatment. Am J Transl Res 2014, 6, 114–118. [Google Scholar]
- Gilbert, P.M.; Weaver, V.M. Cellular adaptation to biomechanical stress across length scales in tissue homeostasis and disease. Semin Cell Dev Biol 2017, 67, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Maman, S.; Witz, I.P. A history of exploring cancer in context. Nat Rev Cancer 2018, 18, 359–376. [Google Scholar] [CrossRef]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat Rev Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef]
- Vlachogiannis, G.; Hedayat, S.; Vatsiou, A.; et al. Patient-derived organoids model treatment response of metastatic gastrointestinal cancers. Science 2018, 359, 920–926. [Google Scholar] [CrossRef]
- Kim, J.; Koo, B.; Knoblich, J.A. Human organoids: model systems for human biology and medicine. Nat Rev Mol Cell Biol 2020, 21, 571–584. [Google Scholar] [CrossRef]
- Verduin, M.; Hoeben, A.; De Ruysscher, D.; et al. Patient-Derived Cancer Organoids as Predictors of Treatment Response. Front Oncol 2021, 11, 641980. [Google Scholar] [CrossRef]
- Chen, K.; Li, Y.; Wang, B.; et al. Patient-derived models facilitate precision medicine in liver cancer by remodeling cell-matrix interaction. Front Immunol 2023, 14, 1101324. [Google Scholar] [CrossRef]
- Battistini, C.; Cavallaro, U. Patient-Derived In Vitro Models of Ovarian Cancer: Powerful Tools to Explore the Biology of the Disease and Develop Personalized Treatments. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Song, K.; Wang, Y.; Wang, Y.; et al. Advances in Thyroid Organoids Research and Applications. Endocr Res 2024, 49, 86–91. [Google Scholar] [CrossRef]
- Ryu, J.; Seong, Y.; Lee, E.; et al. Applications and research trends in organoid based infectious disease models. Sci Rep 2025, 15, 25185. [Google Scholar] [CrossRef]
- Jiang, X.; Oyang, L.; Peng, Q.; et al. Organoids: opportunities and challenges of cancer therapy. Front Cell Dev Biol 2023, 11, 1232528. [Google Scholar] [CrossRef]
- Choi, S.Y.; Kim, T.H.; Kim, M.J.; et al. Validating Well-Functioning Hepatic Organoids for Toxicity Evaluation. Toxics 2024, 12. [Google Scholar] [CrossRef]
- Zhang, H.; Qu, P.; Liu, J.; et al. Application of human cardiac organoids in cardiovascular disease research. Front Cell Dev Biol 2025, 13, 1564889. [Google Scholar] [CrossRef]
- Bittman-Soto, X.S.; Thomas, E.S.; Ganshert, M.E.; et al. The Transformative Role of 3D Culture Models in Triple-Negative Breast Cancer Research. Cancers (Basel) 2024, 16. [Google Scholar] [CrossRef]
- Dionne, O.; Sabatie, S.; Laurent, B. Deciphering the physiopathology of neurodevelopmental disorders using brain organoids. Brain 2025, 148, 12–26. [Google Scholar] [CrossRef]
- Crespo, M.; Vilar, E.; Tsai, S.; et al. Colonic organoids derived from human induced pluripotent stem cells for modeling colorectal cancer and drug testing. Nat Med 2017, 23, 878–884. [Google Scholar] [CrossRef]
- Sachs, N.; de Ligt, J.; Kopper, O.; et al. A Living Biobank of Breast Cancer Organoids Captures Disease Heterogeneity. Cell 2018, 172, 373–386. [Google Scholar] [CrossRef]
- Ooft, S.N.; Weeber, F.; Dijkstra, K.K.; et al. Patient-derived organoids can predict response to chemotherapy in metastatic colorectal cancer patients. Sci Transl Med 2019, 11. [Google Scholar] [CrossRef]
- Driehuis, E.; van Hoeck, A.; Moore, K.; et al. Pancreatic cancer organoids recapitulate disease and allow personalized drug screening. Proc Natl Acad Sci U S A 2019, 116, 26580–26590. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, X.; Yang, L.; et al. Patient-Derived Organoids Predict Chemoradiation Responses of Locally Advanced Rectal Cancer. Cell Stem Cell 2020, 26, 17–26. [Google Scholar] [CrossRef]
- Wensink, G.E.; Elias, S.G.; Mullenders, J.; et al. Patient-derived organoids as a predictive biomarker for treatment response in cancer patients. NPJ Precis Oncol 2021, 5, 30. [Google Scholar] [CrossRef]
- Sontheimer-Phelps, A.; Hassell, B.A.; Ingber, D.E. Modelling cancer in microfluidic human organs-on-chips. Nat Rev Cancer 2019, 19, 65–81. [Google Scholar] [CrossRef]
- Ingber, D.E. Human organs-on-chips for disease modelling, drug development and personalized medicine. Nat Rev Genet 2022, 23, 467–491. [Google Scholar] [CrossRef]
- Lee-Montiel, F.T.; George, S.M.; Gough, A.H.; et al. Control of oxygen tension recapitulates zone-specific functions in human liver microphysiology systems. Exp Biol Med (Maywood) 2017, 242, 1617–1632. [Google Scholar] [CrossRef]
- Shah, P.; Fritz, J.V.; Glaab, E.; et al. A microfluidics-based in vitro model of the gastrointestinal human-microbe interface. Nat Commun 2016, 7, 11535. [Google Scholar] [CrossRef]
- Xiao, S.; Coppeta, J.R.; Rogers, H.B.; et al. A microfluidic culture model of the human reproductive tract and 28-day menstrual cycle. Nat Commun 2017, 8, 14584. [Google Scholar] [CrossRef]
- Schimek, K.; Hsu, H.; Boehme, M.; et al. Bioengineering of a Full-Thickness Skin Equivalent in a 96-Well Insert Format for Substance Permeation Studies and Organ-On-A-Chip Applications. Bioengineering (Basel) 2018, 5. [Google Scholar] [CrossRef]
- Si, L.; Bai, H.; Rodas, M.; et al. A human-airway-on-a-chip for the rapid identification of candidate antiviral therapeutics and prophylactics. Nat Biomed Eng 2021, 5, 815–829. [Google Scholar] [CrossRef]
- Mathur, A.; Loskill, P.; Shao, K.; et al. Human iPSC-based cardiac microphysiological system for drug screening applications. Sci Rep 2015, 5, 8883. [Google Scholar] [CrossRef]
- Peletier, M.; Zhang, X.; Klein, S.; et al. Multicellular 3D models to study myocardial ischemia-reperfusion injury. Front Cell Dev Biol 2024, 12, 1494911. [Google Scholar] [CrossRef]
- Rajan, S.A.P.; Aleman, J.; Wan, M.; et al. Probing prodrug metabolism and reciprocal toxicity with an integrated and humanized multi-tissue organ-on-a-chip platform. Acta Biomater 2020, 106, 124–135. [Google Scholar] [CrossRef]
- Auger, F.A.; Gibot, L.; Lacroix, D. The pivotal role of vascularization in tissue engineering. Annu Rev Biomed Eng 2013, 15, 177–200. [Google Scholar] [CrossRef]
- Tan, T.; Mouradov, D.; Lee, M.; et al. Unified framework for patient-derived, tumor-organoid-based predictive testing of standard-of-care therapies in metastatic colorectal cancer. Cell Rep Med 2023, 4, 101335. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, Y.; Qin, X.; et al. From cell spheroids to vascularized cancer organoids: Microfluidic tumor-on-a-chip models for preclinical drug evaluations. Biomicrofluidics 2021, 15, 61503. [Google Scholar] [CrossRef]
- Sobrino, A.; Phan, D.T.T.; Datta, R.; et al. 3D microtumors in vitro supported by perfused vascular networks. Sci Rep 2016, 6, 31589. [Google Scholar] [CrossRef]
- Jenkins, R.W.; Aref, A.R.; Lizotte, P.H.; et al. Ex Vivo Profiling of PD-1 Blockade Using Organotypic Tumor Spheroids. Cancer Discov 2018, 8, 196–215. [Google Scholar] [CrossRef]
- Brandenberg, N.; Hoehnel, S.; Kuttler, F.; et al. High-throughput automated organoid culture via stem-cell aggregation in microcavity arrays. Nat Biomed Eng 2020, 4, 863–874. [Google Scholar] [CrossRef]
- Novak, R.; Ingram, M.; Marquez, S.; et al. Robotic fluidic coupling and interrogation of multiple vascularized organ chips. Nat Biomed Eng 2020, 4, 407–420. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Aleman, J.; Shin, S.R.; et al. Multisensor-integrated organs-on-chips platform for automated and continual in situ monitoring of organoid behaviors. Proc Natl Acad Sci U S A 2017, 114, E2293–E2302. [Google Scholar] [CrossRef]
- Manjakkal, L.; Dervin, S.; Dahiya, R. Flexible potentiometric pH sensors for wearable systems. RSC Adv 2020, 10, 8594–8617. [Google Scholar] [CrossRef]
- Moya, A.; Ortega-Ribera, M.; Guimera, X.; et al. Online oxygen monitoring using integrated inkjet-printed sensors in a liver-on-a-chip system. Lab Chip 2018, 18, 2023–2035. [Google Scholar] [CrossRef]
- Wang, H.; Lang, Q.; Liang, B.; et al. Electrochemical Glucose Biosensor Based on Glucose Oxidase Displayed on Yeast Surface. Methods Mol Biol 2015, 1319, 233–243. [Google Scholar] [PubMed]
- Perni, M.; Galvagnion, C.; Maltsev, A.; et al. A natural product inhibits the initiation of alpha-synuclein aggregation and suppresses its toxicity. Proc Natl Acad Sci U S A 2017, 114, E1009–E1017. [Google Scholar] [CrossRef]
- Szucs, D.; Fekete, Z.; Guba, M.; et al. Toward better drug development: Three-dimensional bioprinting in toxicological research. Int J Bioprint 2023, 9, 663. [Google Scholar] [CrossRef]
- Low, L.A.; Mummery, C.; Berridge, B.R.; et al. Organs-on-chips: into the next decade. Nat Rev Drug Discov 2021, 20, 345–361. [Google Scholar] [CrossRef]
- Sun, D.; Gao, W.; Hu, H.; et al. Why 90% of clinical drug development fails and how to improve it? . Acta Pharm Sin B 2022, 12, 3049–3062. [Google Scholar] [CrossRef]
- Dagogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat Rev Clin Oncol 2018, 15, 81–94. [Google Scholar] [CrossRef]
- Broutier, L.; Mastrogiovanni, G.; Verstegen, M.M.; et al. Human primary liver cancer-derived organoid cultures for disease modeling and drug screening. Nat Med 2017, 23, 1424–1435. [Google Scholar] [CrossRef]
- Meyers, N.L.; Ashuach, T.; Lyons, D.E.; et al. Hepatitis C virus infects and perturbs liver stem cells. mBio 2023, 14, e131823. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, S.; Yang, Z.; et al. Construction of vascularized liver microtissues recapitulates angiocrine-mediated hepatocytes maturation and enhances therapeutic efficacy for acute liver failure. Bioact Mater 2025, 50, 525–539. [Google Scholar] [CrossRef]
- Niazi, V.; Parseh, B. Organoid models of breast cancer in precision medicine and translational research. Mol Biol Rep 2024, 52, 2. [Google Scholar] [CrossRef]
- Dixon, T.A.; Muotri, A.R. Advancing preclinical models of psychiatric disorders with human brain organoid cultures. Mol Psychiatry 2023, 28, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jiang, S.; Gu, L.; et al. High-throughput bioengineering of homogenous and functional human-induced pluripotent stem cells-derived liver organoids via micropatterning technique. Front Bioeng Biotechnol 2022, 10, 937595. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Ueyama-Toba, Y.; Yokota, J.; et al. Efficient hepatocyte differentiation of primary human hepatocyte-derived organoids using three dimensional nanofibers (HYDROX) and their possible application in hepatotoxicity research. Sci Rep 2024, 14, 10846. [Google Scholar] [CrossRef]
- Gout, J.; Ekizce, M.; Roger, E.; et al. Pancreatic organoids as cancer avatars for true personalized medicine. Adv Drug Deliv Rev 2025, 224, 115642. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Zhang, S.; Ma, D.; et al. Targeting SphK2 Reverses Acquired Resistance of Regorafenib in Hepatocellular Carcinoma. Front Oncol 2020, 10, 694. [Google Scholar] [CrossRef]
- Kong, D.; Mourtzinos, A.; Heegsma, J.; et al. Growth differentiation factor 7 autocrine signaling promotes hepatic progenitor cell expansion in liver fibrosis. Stem Cell Res Ther 2023, 14, 288. [Google Scholar] [CrossRef]
- El-Khobar, K.E.; Sukowati, C.H.C. Updates on Organoid Model for the Study of Liver Cancer. Technol Cancer Res Treat 2023, 22, 2071071318. [Google Scholar] [CrossRef]
- Hughes, C.S.; Postovit, L.M.; Lajoie, G.A. Matrigel: a complex protein mixture required for optimal growth of cell culture. Proteomics 2010, 10, 1886–1890. [Google Scholar] [CrossRef]
- Gjorevski, N.; Sachs, N.; Manfrin, A.; et al. Designer matrices for intestinal stem cell and organoid culture. Nature 2016, 539, 560–564. [Google Scholar] [CrossRef]
- Giobbe, G.G.; Michielin, F.; Luni, C.; et al. Functional differentiation of human pluripotent stem cells on a chip. Nat Methods 2015, 12, 637–640. [Google Scholar] [CrossRef]
- Lee, S.H.; Sung, J.H. Organ-on-a-Chip Technology for Reproducing Multiorgan Physiology. Adv Healthc Mater 2018, 7. [Google Scholar] [CrossRef]
- Homan, K.A.; Gupta, N.; Kroll, K.T.; et al. Flow-enhanced vascularization and maturation of kidney organoids in vitro. Nat Methods 2019, 16, 255–262. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, W.; Shao, Q.; et al. Pentapeptide IKVAV-engineered hydrogels for neural stem cell attachment. Biomater Sci 2021, 9, 2887–2892. [Google Scholar] [CrossRef]
- Qiao, F.; Zou, Y.; Bie, B.; et al. Dual siRNA-Loaded Cell Membrane Functionalized Matrix Facilitates Bone Regeneration with Angiogenesis and Neurogenesis. Small 2024, 20, e2307062. [Google Scholar] [CrossRef]
- Ge, J.; Wang, Y.; Li, Q.; et al. Trends and challenges in organoid modeling and expansion with pluripotent stem cells and somatic tissue. PeerJ 2024, 12, e18422. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.C.; Than, N.; Park, S.J.; et al. Bioengineered human gut-on-a-chip for advancing non-clinical pharmaco-toxicology. Expert Opin Drug Metab Toxicol 2024, 20, 593–606. [Google Scholar] [CrossRef]
- Hofer, M.; Lutolf, M.P. Engineering organoids. Nat Rev Mater 2021, 6, 402–420. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, K.; Iversen, P.W.; Schumacher, D.; et al. Assay Establishment and Validation of a High-Throughput Screening Platform for Three-Dimensional Patient-Derived Colon Cancer Organoid Cultures. J Biomol Screen 2016, 21, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.; Boutros, M.; Camp, J.G.; et al. The Organoid Cell Atlas. Nat Biotechnol 2021, 39, 13–17. [Google Scholar] [CrossRef]
- Dekkers, J.F.; Alieva, M.; Wellens, L.M.; et al. High-resolution 3D imaging of fixed and cleared organoids. Nat Protoc 2019, 14, 1756–1771. [Google Scholar] [CrossRef]
- Ishikura, M.; Muraoka, Y.; Hirami, Y.; et al. Adaptive Optics Optical Coherence Tomography Analysis of Induced Pluripotent Stem Cell-Derived Retinal Organoid Transplantation in Retinitis Pigmentosa. Cureus 2024, 16, e64962. [Google Scholar] [CrossRef]
- Gil, D.A.; Deming, D.; Skala, M.C. Patient-derived cancer organoid tracking with wide-field one-photon redox imaging to assess treatment response. J Biomed Opt 2021, 26. [Google Scholar] [CrossRef]
- Das, S.; Sahoo, S.; Pattanaik, S.; et al. Advancing breast cancer research: a comprehensive review of in vitro and in vivo experimental models. Med Oncol 2025, 42, 316. [Google Scholar] [CrossRef] [PubMed]
- Souto, E.P.; Dobrolecki, L.E.; Villanueva, H.; et al. In Vivo Modeling of Human Breast Cancer Using Cell Line and Patient-Derived Xenografts. J Mammary Gland Biol Neoplasia 2022, 27, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. COVID-19: organoids go viral. Nat Rev Mol Cell Biol 2020, 21, 355–356. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Huch, M. Disease modelling in human organoids. Dis Model Mech 2019, 12. [Google Scholar] [CrossRef]
- Takasato, M.; Er, P.X.; Chiu, H.S.; et al. Kidney organoids from human iPS cells contain multiple lineages and model human nephrogenesis. Nature 2015, 526, 564–568. [Google Scholar] [CrossRef]
- Qian, X.; Nguyen, H.N.; Song, M.M.; et al. Brain-Region-Specific Organoids Using Mini-bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef]
- Boretto, M.; Maenhoudt, N.; Luo, X.; et al. Patient-derived organoids from endometrial disease capture clinical heterogeneity and are amenable to drug screening. Nat Cell Biol 2019, 21, 1041–1051. [Google Scholar] [CrossRef]
- Jalili-Firoozinezhad, S.; Gazzaniga, F.S.; Calamari, E.L.; et al. A complex human gut microbiome cultured in an anaerobic intestine-on-a-chip. Nat Biomed Eng 2019, 3, 520–531. [Google Scholar] [CrossRef]
- Giacomelli, E.; Meraviglia, V.; Campostrini, G.; et al. Human-iPSC-Derived Cardiac Stromal Cells Enhance Maturation in 3D Cardiac Microtissues and Reveal Non-cardiomyocyte Contributions to Heart Disease. Cell Stem Cell 2020, 26, 862–879. [Google Scholar] [CrossRef]
- Trujillo, C.A.; Gao, R.; Negraes, P.D.; et al. Complex Oscillatory Waves Emerging from Cortical Organoids Model Early Human Brain Network Development. Cell Stem Cell 2019, 25, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Mead, B.E.; Safaee, H.; et al. Engineering Stem Cell Organoids. Cell Stem Cell 2016, 18, 25–38. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.; Ha, G.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Tiriac, H.; Belleau, P.; Engle, D.D.; et al. Organoid Profiling Identifies Common Responders to Chemotherapy in Pancreatic Cancer. Cancer Discov 2018, 8, 1112–1129. [Google Scholar] [CrossRef]
- Huang, S. Non-genetic heterogeneity of cells in development: more than just noise. Development 2009, 136, 3853–3862. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: mechanism and applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.J.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci Data 2016, 3, 160018. [Google Scholar] [CrossRef]
- Castiglione, H.; Madrange, L.; Baquerre, C.; et al. Towards a quality control framework for cerebral cortical organoids. Sci Rep 2025, 15, 29431. [Google Scholar] [CrossRef]
- Fatehullah, A.; Tan, S.H.; Barker, N. Organoids as an in vitro model of human development and disease. Nat Cell Biol 2016, 18, 246–254. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Chung, M.; Fioret, B.; et al. Lung organoids: current uses and future promise. Development 2017, 144, 986–997. [Google Scholar] [CrossRef]
- Xiang, D.; He, A.; Zhou, R.; et al. Building consensus on the application of organoid-based drug sensitivity testing in cancer precision medicine and drug development. Theranostics 2024, 14, 3300–3316. [Google Scholar] [CrossRef] [PubMed]
- Madorsky Rowdo, F.P.; Xiao, G.; Khramtsova, G.F.; et al. Patient-derived tumor organoids with p53 mutations, and not wild-type p53, are sensitive to synergistic combination PARP inhibitor treatment. Cancer Lett 2024, 584, 216608. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cao, J.; Wang, G.; et al. Circulating bile acid profiles characteristics and the potential predictive role in clear cell renal cell carcinoma progression. BMC Nephrol 2025, 26, 206. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, Y.; Zhang, Y.; et al. Clinical value and influencing factors of establishing stomach cancer organoids by endoscopic biopsy. J Cancer Res Clin Oncol 2023, 149, 3803–3810. [Google Scholar] [CrossRef]
- Thorel, L.; Elie, N.; Morice, P.; et al. Automated Scoring to Assess RAD51-Mediated Homologous Recombination in Ovarian Patient-Derived Tumor Organoids. Lab Invest 2025, 105, 104097. [Google Scholar] [CrossRef]
- Raja, W.K.; Mungenast, A.E.; Lin, Y.; et al. Self-Organizing 3D Human Neural Tissue Derived from Induced Pluripotent Stem Cells Recapitulate Alzheimer’s Disease Phenotypes. PLoS One 2016, 11, e161969. [Google Scholar] [CrossRef]
- Wang, M.; Li, R.; Sheng, S.; et al. Combination therapy using intestinal organoids and their extracellular vesicles for inflammatory bowel disease complicated with osteoporosis. J Orthop Translat 2025, 53, 26–36. [Google Scholar] [CrossRef]
- Nairon, K.G.; Nigam, A.; Khanal, T.; et al. RCAN1.4 regulates tumor cell engraftment and invasion in a thyroid cancer to lung metastasis-on-a-chip microphysiological system. Biofabrication 2024, 17. [Google Scholar] [CrossRef]
- Astashkina, A.I.; Mann, B.K.; Prestwich, G.D.; et al. Comparing predictive drug nephrotoxicity biomarkers in kidney 3-D primary organoid culture and immortalized cell lines. Biomaterials 2012, 33, 4712–4721. [Google Scholar] [CrossRef]
- Sampaziotis, F.; Muraro, D.; Tysoe, O.C.; et al. Cholangiocyte organoids can repair bile ducts after transplantation in the human liver. Science 2021, 371, 839–846. [Google Scholar] [CrossRef]
- Nashimoto, Y.; Okada, R.; Hanada, S.; et al. Vascularized cancer on a chip: The effect of perfusion on growth and drug delivery of tumor spheroid. Biomaterials 2020, 229, 119547. [Google Scholar] [CrossRef]
- Bellotti, C.; Samudyata, S.; Thams, S.; et al. Organoids and chimeras: the hopeful fusion transforming traumatic brain injury research. Acta Neuropathol Commun 2024, 12, 141. [Google Scholar] [CrossRef]
- van de Wetering, M.; Francies, H.E.; Francis, J.M.; et al. Prospective derivation of a living organoid biobank of colorectal cancer patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef]
- Yang, R.; Yu, Y. Patient-derived organoids in translational oncology and drug screening. Cancer Lett 2023, 562, 216180. [Google Scholar] [CrossRef] [PubMed]
- Geurts, M.H.; de Poel, E.; Amatngalim, G.D.; et al. CRISPR-Based Adenine Editors Correct Nonsense Mutations in a Cystic Fibrosis Organoid Biobank. Cell Stem Cell 2020, 26, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Mao, Y.; Han, X.; et al. EGR1 Regulates SHANK3 Transcription at Different Stages of Brain Development. Neuroscience 2024, 540, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.; Kolahi, K.S.; Du, Y.; et al. A CRISPR/Cas9-Engineered ARID1A-Deficient Human Gastric Cancer Organoid Model Reveals Essential and Nonessential Modes of Oncogenic Transformation. Cancer Discov 2021, 11, 1562–1581. [Google Scholar] [CrossRef]
- Childs, C.J.; Poling, H.M.; Chen, K.; et al. Coordinated differentiation of human intestinal organoids with functional enteric neurons and vasculature. Cell Stem Cell 2025, 32, 640–651. [Google Scholar] [CrossRef]
- Youk, J.; Kim, T.; Evans, K.V.; et al. Three-Dimensional Human Alveolar Stem Cell Culture Models Reveal Infection Response to SARS-CoV-2. Cell Stem Cell 2020, 27, 905–919. [Google Scholar] [CrossRef]
- Salahudeen, A.A.; Choi, S.S.; Rustagi, A.; et al. Progenitor identification and SARS-CoV-2 infection in human distal lung organoids. Nature 2020, 588, 670–675. [Google Scholar] [CrossRef]
- Ettayebi, K.; Kaur, G.; Patil, K.; et al. Insights into human norovirus cultivation in human intestinal enteroids. mSphere 2024, 9, e44824. [Google Scholar] [CrossRef]
- Watanabe, M.; Buth, J.E.; Vishlaghi, N.; et al. Self-Organized Cerebral Organoids with Human-Specific Features Predict Effective Drugs to Combat Zika Virus Infection. Cell Rep 2017, 21, 517–532. [Google Scholar] [CrossRef]
- Ouchi, R.; Togo, S.; Kimura, M.; et al. Modeling Steatohepatitis in Humans with Pluripotent Stem Cell-Derived Organoids. Cell Metab 2019, 30, 374–384. [Google Scholar] [CrossRef]
- Balboa, D.; Saarimaki-Vire, J.; Borshagovski, D.; et al. Insulin mutations impair beta-cell development in a patient-derived iPSC model of neonatal diabetes. Elife 2018, 7. [Google Scholar] [CrossRef]
- Shahjalal, H.M.; Abdal Dayem, A.; Lim, K.M.; et al. Generation of pancreatic beta cells for treatment of diabetes: advances and challenges. Stem Cell Res Ther 2018, 9, 355. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Mun, H.; Sung, C.O.; et al. Patient-derived lung cancer organoids as in vitro cancer models for therapeutic screening. Nat Commun 2019, 10, 3991. [Google Scholar] [CrossRef] [PubMed]
- Adashi, E.Y.; O’Mahony, D.P.; Cohen, I.G. The FDA Modernization Act 2.0: Drug Testing in Animals is Rendered Optional. Am J Med 2023, 136, 853–854. [Google Scholar] [CrossRef]
- Le, S.; Petersilie, L.; Inak, G.; et al. Generation of Human Brain Organoids for Mitochondrial Disease Modeling. J Vis Exp 2021. [Google Scholar]
- Nakamoto, N.; Sasaki, N.; Aoki, R.; et al. Gut pathobionts underlie intestinal barrier dysfunction and liver T helper 17 cell immune response in primary sclerosing cholangitis. Nat Microbiol 2019, 4, 492–503. [Google Scholar] [CrossRef]
- Robertson, M.J.; Soibam, B.; O’Leary, J.G.; et al. Recellularization of rat liver: An in vitro model for assessing human drug metabolism and liver biology. PLoS One 2018, 13, e191892. [Google Scholar] [CrossRef]
- Guo, H.; Deng, N.; Dou, L.; et al. 3-D Human Renal Tubular Organoids Generated from Urine-Derived Stem Cells for Nephrotoxicity Screening. ACS Biomater Sci Eng 2020, 6, 6701–6709. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, K.K.; Cattaneo, C.M.; Weeber, F.; et al. Generation of Tumor-Reactive T Cells by Co-culture of Peripheral Blood Lymphocytes and Tumor Organoids. Cell 2018, 174, 1586–1598. [Google Scholar] [CrossRef]
- Ganesh, K.; Wu, C.; O’Rourke, K.P.; et al. A rectal cancer organoid platform to study individual responses to chemoradiation. Nat Med 2019, 25, 1607–1614. [Google Scholar] [CrossRef]
- Chen, G.; Gong, T.; Wang, Z.; et al. Colorectal cancer organoid models uncover oxaliplatin-resistant mechanisms at single cell resolution. Cell Oncol (Dordr) 2022, 45, 1155–1167. [Google Scholar]
- Berical, A.; Lee, R.E.; Lu, J.; et al. A multimodal iPSC platform for cystic fibrosis drug testing. Nat Commun 2022, 13, 4270. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhang, X.; Ge, Q.; et al. CRISPR/Cas9-mediated knockout of HBsAg inhibits proliferation and tumorigenicity of HBV-positive hepatocellular carcinoma cells. J Cell Biochem 2018, 119, 8419–8431. [Google Scholar] [CrossRef] [PubMed]
- Logun, M.; Wang, X.; Sun, Y.; et al. Patient-derived glioblastoma organoids as real-time avatars for assessing responses to clinical CAR-T cell therapy. Cell Stem Cell 2025, 32, 181–190. [Google Scholar] [CrossRef]
- Moghimi, B.; Muthugounder, S.; Jambon, S.; et al. Preclinical assessment of the efficacy and specificity of GD2-B7H3 SynNotch CAR-T in metastatic neuroblastoma. Nat Commun 2021, 12, 511. [Google Scholar] [CrossRef]
- Neal, J.T.; Li, X.; Zhu, J.; et al. Organoid Modeling of the Tumor Immune Microenvironment. Cell 2018, 175, 1972–1988. [Google Scholar] [CrossRef]
- Rossi, G.; Manfrin, A.; Lutolf, M.P. Progress and potential in organoid research. Nat Rev Genet 2018, 19, 671–687. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H.M.; Louzada, J.; Bardgett, R.D.; et al. Assessing the Importance of Intraspecific Variability in Dung Beetle Functional Traits. PLoS One 2016, 11, e145598. [Google Scholar] [CrossRef]
- Lavazza, A.; Massimini, M. Cerebral organoids and consciousness: how far are we willing to go? . J Med Ethics 2018, 44, 613–614. [Google Scholar] [CrossRef] [PubMed]
- Trepagnier, D.M. Human embryonic stem cell research: implications from an ethical and legal standpoint. J La State Med Soc 2000, 152, 616–624. [Google Scholar]
- Lensink, M.A.; Boers, S.N.; Jongsma, K.R.; et al. Organoids for personalized treatment of Cystic Fibrosis: Professional perspectives on the ethics and governance of organoid biobanking. J Cyst Fibros 2021, 20, 443–451. [Google Scholar] [CrossRef]
- Wright, C.B.; Becker, S.M.; Low, L.A.; et al. Improved Ocular Tissue Models and Eye-On-A-Chip Technologies Will Facilitate Ophthalmic Drug Development. J Ocul Pharmacol Ther 2020, 36, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhao, H.; Zhang, W.; et al. An Automated Organoid Platform with Inter-organoid Homogeneity and Inter-patient Heterogeneity. Cell Rep Med 2020, 1, 100161. [Google Scholar] [CrossRef]
- Metzger, J.J.; Pereda, C.; Adhikari, A.; et al. Deep-learning analysis of micropattern-based organoids enables high-throughput drug screening of Huntington’s disease models. Cell Rep Methods 2022, 2, 100297. [Google Scholar] [CrossRef]
- Song, E.; Kim, M.; Lee, S.; et al. VONet: A deep learning network for 3D reconstruction of organoid structures with a minimal number of confocal images. Patterns (N Y) 2024, 5, 101063. [Google Scholar] [CrossRef]
- Cao, Y.; Fu, L.; Wu, J.; et al. Integrated analysis of multimodal single-cell data with structural similarity. Nucleic Acids Res 2022, 50, e121. [Google Scholar] [CrossRef]
- Maramraju, S.; Kowalczewski, A.; Kaza, A.; et al. AI-organoid integrated systems for biomedical studies and applications. Bioeng Transl Med 2024, 9, e10641. [Google Scholar] [CrossRef]
- Bai, L.; Wu, Y.; Li, G.; et al. AI-enabled organoids: Construction, analysis, and application. Bioact Mater 2024, 31, 525–548. [Google Scholar] [CrossRef] [PubMed]
- Stuart, T.; Satija, R. Integrative single-cell analysis. Nat Rev Genet 2019, 20, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, B.; Wang, Y.; et al. Microfluidic Chip-Based Automatic System for Sequencing Patient-Derived Organoids at the Single-Cell Level. Anal Chem 2024, 96, 17027–17036. [Google Scholar] [CrossRef] [PubMed]
- Pagella, P.; Soderholm, S.; Nordin, A.; et al. The time-resolved genomic impact of Wnt/beta-catenin signaling. Cell Syst 2023, 14, 563–581. [Google Scholar] [CrossRef]
- Montorsi, L.; Pitcher, M.J.; Zhao, Y.; et al. Double-negative B cells and DNASE1L3 colocalise with microbiota in gut-associated lymphoid tissue. Nat Commun 2024, 15, 4051. [Google Scholar] [CrossRef]
- Bravo Gonzalez-Blas, C.; Matetovici, I.; Hillen, H.; et al. Single-cell spatial multi-omics and deep learning dissect enhancer-driven gene regulatory networks in liver zonation. Nat Cell Biol 2024, 26, 153–167. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Muto, Y.; Ledru, N.; et al. A single-cell multiomic analysis of kidney organoid differentiation. Proc Natl Acad Sci U S A 2023, 120, e2075268176. [Google Scholar] [CrossRef]
- Kim, S.; Kim, S.; Lim, S.; et al. Modeling Clinical Responses to Targeted Therapies by Patient-Derived Organoids of Advanced Lung Adenocarcinoma. Clin Cancer Res 2021, 27, 4397–4409. [Google Scholar] [CrossRef]
- Bulcaen, M.; Kortleven, P.; Liu, R.B.; et al. Prime editing functionally corrects cystic fibrosis-causing CFTR mutations in human organoids and airway epithelial cells. Cell Rep Med 2024, 5, 101544. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Fujimori, N.; Ichihara, K.; et al. Patient-derived organoids of pancreatic ductal adenocarcinoma for subtype determination and clinical outcome prediction. J Gastroenterol 2024, 59, 629–640. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Disease Modeling and Drug Development Platform Based on Organoids and Organ-on-a-Chip. This platform integrates multidisciplinary technologies to advance precision medicine. Its primary applications include: A-B: Disease Modeling: Constructing models for diseases such as cancer through gene profiling and single-cell long-read sequencing. C-D: Drug Testing: Utilizing organoid models for pre-clinical drug efficacy testing (e.g., response, no response, or mild response) and drug safety testing. E: Regenerative Medicine and Clinical Translation: Combining AI-predictive models and clinical diagnostics to ultimately apply research findings in regenerative medicine for patient care. F: Biobanking: Establishing organ-on-a-chip and organoid models from healthy donors or patients to create biobanks for academic studies.
Figure 1.
Disease Modeling and Drug Development Platform Based on Organoids and Organ-on-a-Chip. This platform integrates multidisciplinary technologies to advance precision medicine. Its primary applications include: A-B: Disease Modeling: Constructing models for diseases such as cancer through gene profiling and single-cell long-read sequencing. C-D: Drug Testing: Utilizing organoid models for pre-clinical drug efficacy testing (e.g., response, no response, or mild response) and drug safety testing. E: Regenerative Medicine and Clinical Translation: Combining AI-predictive models and clinical diagnostics to ultimately apply research findings in regenerative medicine for patient care. F: Biobanking: Establishing organ-on-a-chip and organoid models from healthy donors or patients to create biobanks for academic studies.
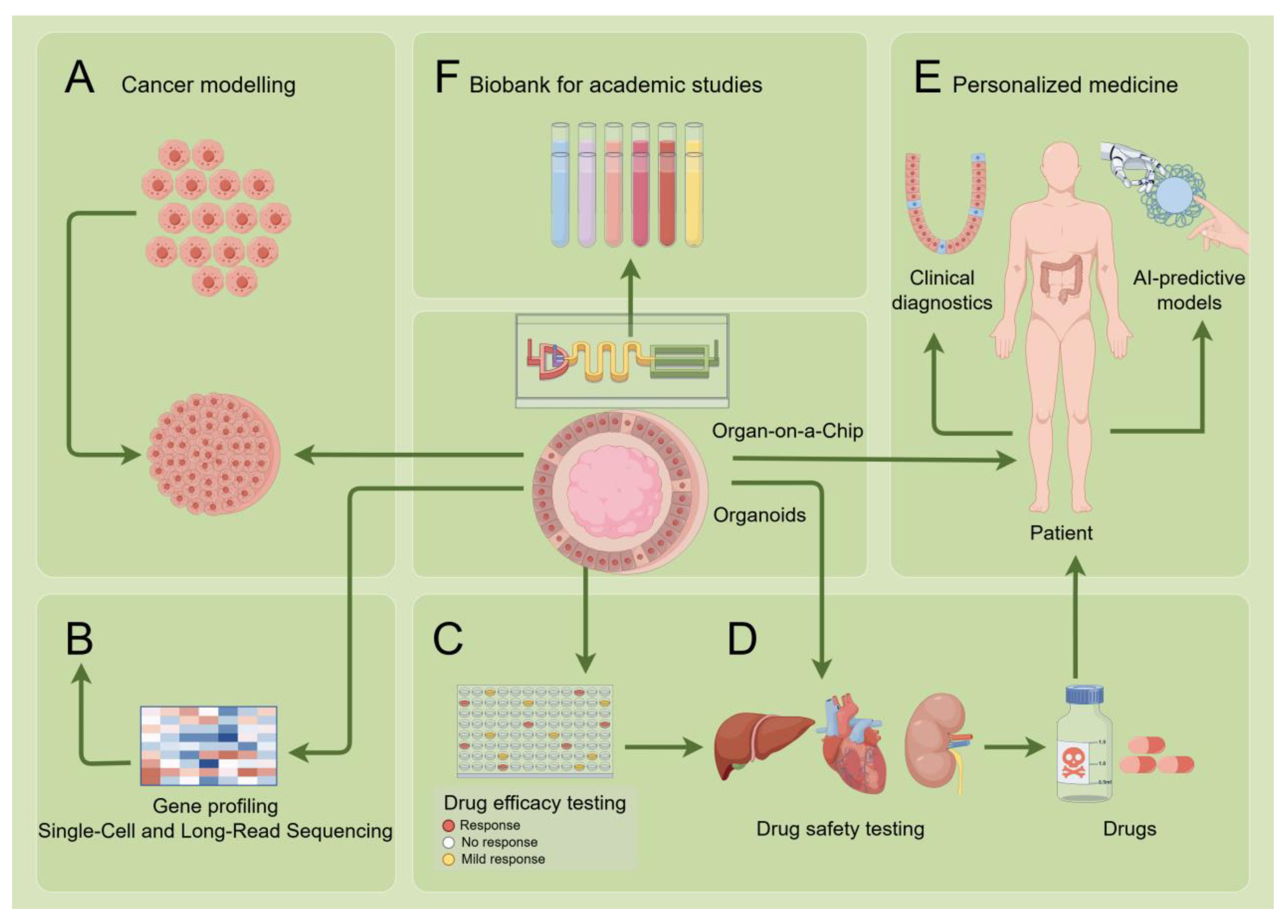
Figure 2.
This schematic illustrates an integrated research strategy for modeling and deriving complete cancer phenotypes in vitro. The pipeline begins with the establishment of stable cell lines, which are then used to generate patient-derived xenograft (PDX) models in mice for in vivo validation. Subsequently, organoids are derived from PDX models and cultured in a three-dimensional system using defined stromal components (e.g., POTX, Matingel). This system recapitulates the tumor progression sequence from normal epithelium to adenoma and adenocarcinoma, and incorporates critical tumor microenvironment elements, including vasculature, mesenchymal cells, immune cells, and the extracellular matrix. By synergistically utilizing cell lines, PDX models, and defined stroma, this platform enables the systematic generation and study of diverse, comprehensive cancer phenotypes.
Figure 2.
This schematic illustrates an integrated research strategy for modeling and deriving complete cancer phenotypes in vitro. The pipeline begins with the establishment of stable cell lines, which are then used to generate patient-derived xenograft (PDX) models in mice for in vivo validation. Subsequently, organoids are derived from PDX models and cultured in a three-dimensional system using defined stromal components (e.g., POTX, Matingel). This system recapitulates the tumor progression sequence from normal epithelium to adenoma and adenocarcinoma, and incorporates critical tumor microenvironment elements, including vasculature, mesenchymal cells, immune cells, and the extracellular matrix. By synergistically utilizing cell lines, PDX models, and defined stroma, this platform enables the systematic generation and study of diverse, comprehensive cancer phenotypes.
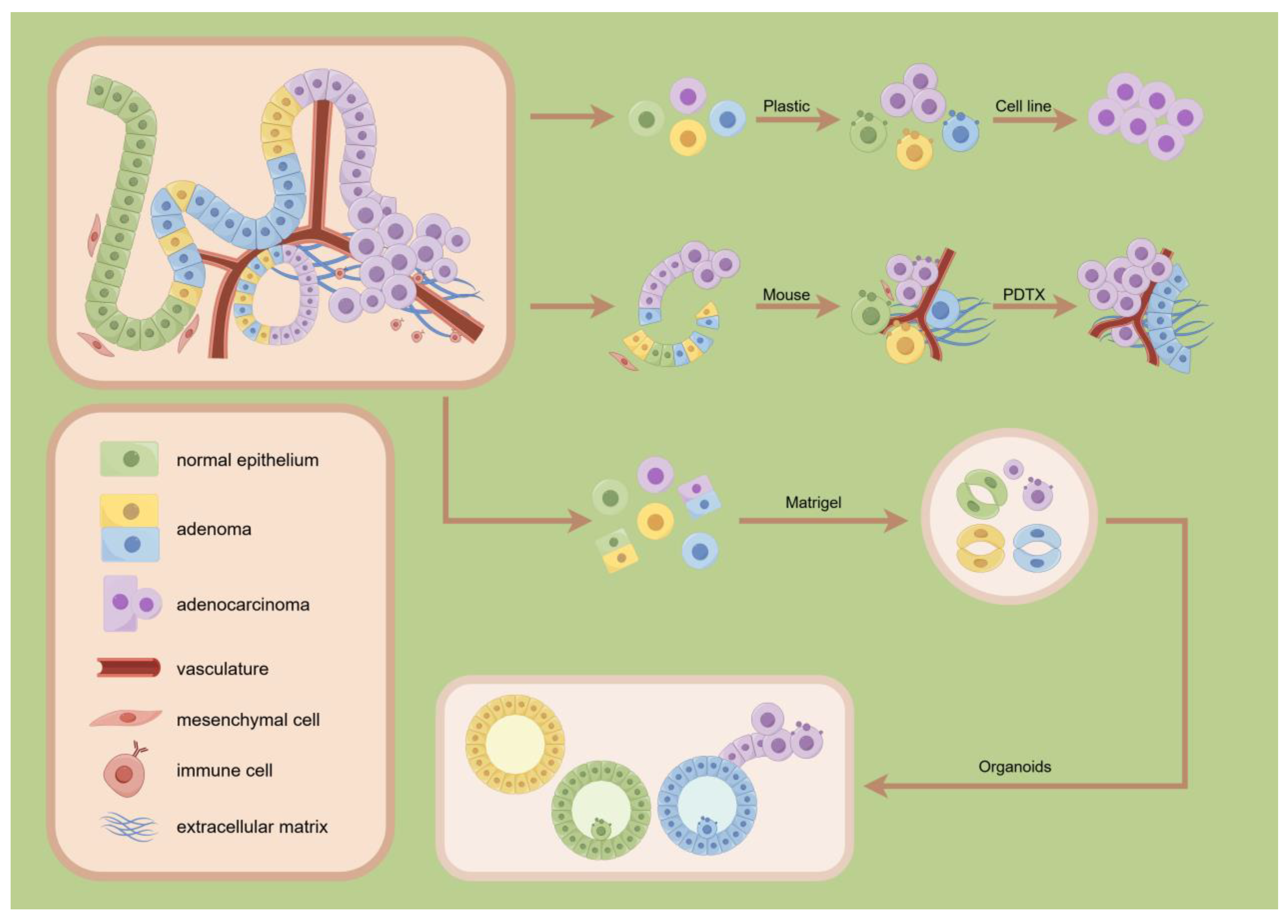
Figure 3.
Schematic illustration of the establishment of tumor organoids. Tumor organoids can be generated from three primary sources: (1) cancer tissues or cancer cells obtained from patients; (2) pluripotent stem cells, including embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs); and (3) normal tissues or somatic stem cells. These starting materials are cultured in three-dimensional (3D) matrices with specific culture media to form self-organizing structures. The resulting models encompass both normal organoids and cancer organoids, which recapitulate key functional units and can be utilized as floating spheroids for a wide range of biomedical applications.
Figure 3.
Schematic illustration of the establishment of tumor organoids. Tumor organoids can be generated from three primary sources: (1) cancer tissues or cancer cells obtained from patients; (2) pluripotent stem cells, including embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs); and (3) normal tissues or somatic stem cells. These starting materials are cultured in three-dimensional (3D) matrices with specific culture media to form self-organizing structures. The resulting models encompass both normal organoids and cancer organoids, which recapitulate key functional units and can be utilized as floating spheroids for a wide range of biomedical applications.
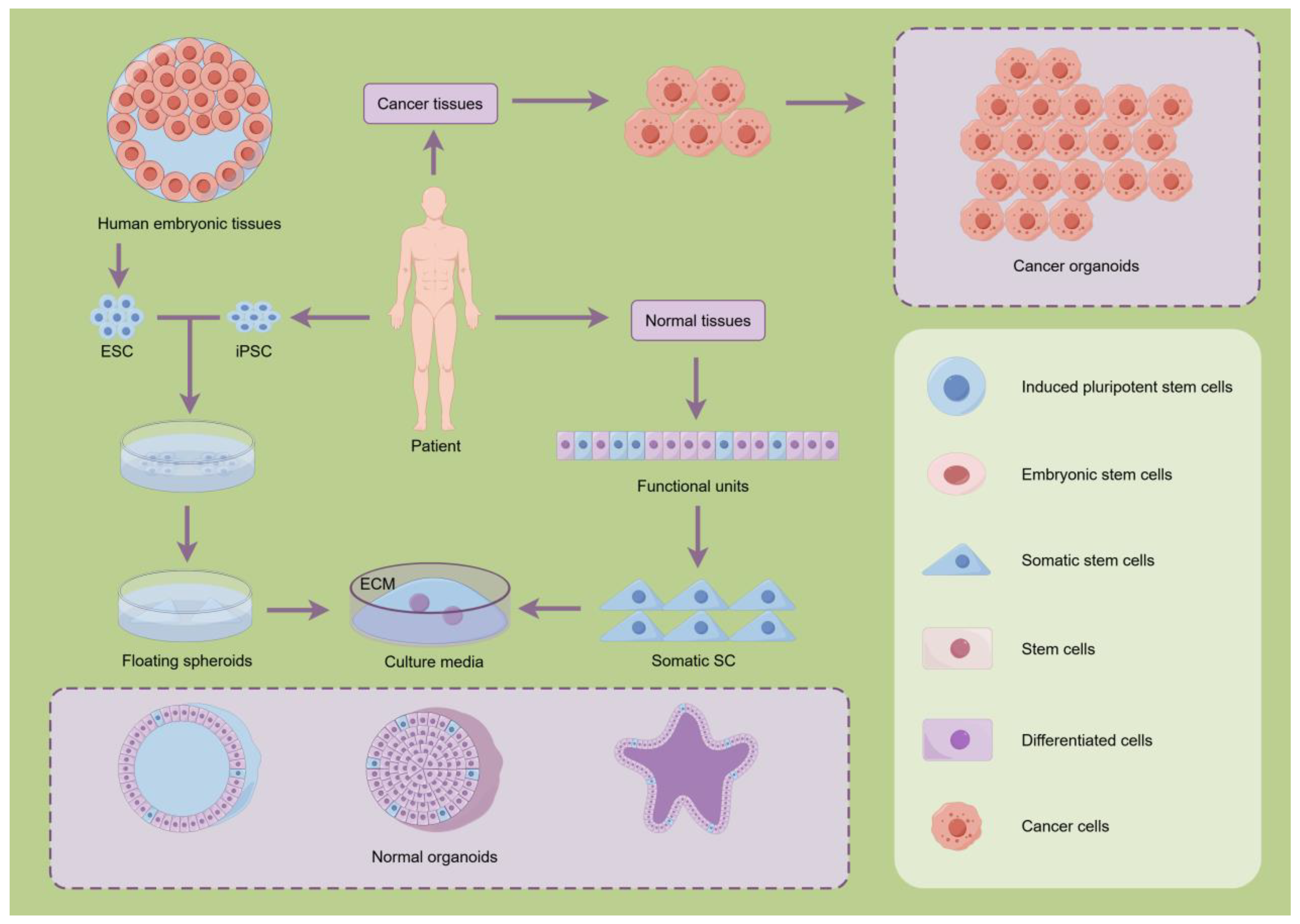
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



