Submitted:
22 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
The potyvirus helper component proteinase (HcPro) is a multifunctional protein, one of its most documented functions being host antiviral RNA silencing suppression. This study shows that the HcPro of potato virus Y (PVY), an important member of the potyvirus group, prevents replication of a related competing secondary virus. This phenomenon, referred to as superinfection exclusion (SIE), is common in bacterial, human, and plant virus infections. We also report that HcPro induction of SIE is strain specific and this specificity is provided by the first four amino acid residues of the protein. Consistent with the mechanism of SIE, the study found that HcPro does not exclude a resident virus. Additionally, HcPro induction of SIE was observed to function independently of its ability to suppress antiviral RNA silencing. HcPro induction of SIE is relevant given the prevalence of multiple PVY strains that routinely co-infect the same cell and that may lead to recombination and emergence of new and more virulent strains. Furthermore, cross-protection that is employed in plant virus disease management occurs when SIE moves from the cellular level and spreads systemically, emphasizing the importance of SIE.
Keywords:
HcPro
; potato virus Y
; superinfection exclusion
; suppression of RNA silencing
; systemic acquired resistance (SAR)
1. Introduction
Diseases caused by potato virus Y (PVY) are a major constraint to potato (Solanum tuberosum L.) production worldwide. PVY is disseminated in potatoes by the aphid vector and through potato seed tubers, which are the main mode of potato propagation. PVY infected potato plants display leaf mosaic, crinkling, deformation, and leaf and tuber necrosis, all of which contribute to decreases in tuber yield and quality. Based on the host hypersensitive response, genome sequence and/or serological properties, PVY has been grouped into three main clusters, PVYO, PVYN, and PVYC [1]. Recombination amongst these clusters has resulted in new variants, and to date, at least nine recombination patterns of PVYO and PVYN sequences have been identified in potato-infecting PVY isolates, the most important being PVYNTN, PVYN:O, and PVYN-Wi [2,3].
The PVY genome is a positive-sense single-stranded RNA [(+)ssRNA)] that encodes a large polyprotein of ~3062 amino acid residues. The coding region is flanked by 5` and 3` untranslated regions (UTRs), and a virus encoded VPg (viral protein genome-linked) is covalently attached at the 5’ UTR, while the 3’ UTR is polyadenylated [4]. Upon translation, the polyprotein undergoes proteolytic processing, producing 10 functional proteins: protein 1 (P1), helper component-protease (HC-Pro), protein 3 (P3), 6 kilodalton (kDa) peptide 1 (6K1), cylindrical inclusion protein (CI), 6 kDa peptide 2 (6K2), viral protein genome-linked (VPg), nuclear inclusion-a protease (NIa-Pro), nuclear inclusion-b protein (NIb), and capsid protein (CP) [5]. Proteolytic processing is carried out by the three P1, HcPro, and the NIa-Pro [6,7,8]. During replication, slippage of the RNA dependent RNA polymerase (RdRp) at the conserved motif (5′-GAAAAAA-3′) in the P3 cistron results in the expression of an additional protein named P3N-PIPO (N-terminal half of P3 fused to pretty interesting Potyvirus ORF) [9,10,11,12].
The potyvirus HcPro is a multifunctional protein (Figure 1), and one of the most studied potyviral proteins, its most documented functions being the ability to self-cleave from the viral polyprotein [13,14], and suppression of RNA silencing [15,16,17,18,19]. Other HcPro functions include: mediation of aphid transmission of PVY [20], assembly of PVY particles [21,22], facilitation of local and systemic spread of progeny virus [23,24,25].
This ever-expanding functions of the potyviral HcPro include the ability to elicit superinfection exclusion (SIE), as was reported for HcPro of potato virus A [27]. In SIE, a virus-infected cell is protected from subsequent infection (superinfection) by the same or a closely related virus [28,29,30]. Other potyviral proteins that have been reported to elicit SIE are NIa-Pro, CP, and P3 [31,32]. Virus encoded protein inducers of SIE have been reported in other plant virus groups, including Orf1a, p33, and p23 of Citrus tristeza virus [30,33], and matrix protein of sonchus yellow net virus [34]. Induction of SIE has also been reported in human, animal, and especially bacterial viruses, including influenza A virus neuraminidase (NA) [35], bovine viral diarrhea virus E2 protein [36], and bacteriophage Pf PA0721 protein [37].
The phenomenon of cross-protection or systemic acquired resistance (SAR), where plants pre-infected with a mild isolate of a virus becomes protected against secondary infections by more severe isolates of the same virus, is used to manage plant virus diseases [38,39,40,41,42,43]. Mechanistically, it has been shown that SAR occurs when SIE moves from the cellular level and spreads systemically to the rest of the plant [38,44,45]. Therefore, to characterize the establishment of SAR in PVY, we sort to characterize HcPro induction of PVY SIE. Results of the study show that HcPro of PVYO and of PVYNTN causes the exclusion of the superinfecting parent virus strain but not that of the other strain. The study further shows that this specific exclusion is provided mainly, but not exclusively, by the first four amino acid residues of the HcPro cistron. Using mutational analysis, we further show that HcPro induction of SIE functions independently of its ability to suppress antiviral RNA silencing. Furthermore, PVYNTN HcPro amino acid residue Asp-419 has been shown to be responsible for the ability of PVYNTN to induce potato tuber necrosis ringspot disease (PTNRD) [46]. Therefore, we investigated whether these amino acid residues also influence HcPro suppression of RNA silencing. Results showed that swapping Asp-419 and Glu-419 between the HcPro of PVYO and PVYNTN attenuates RNA silencing suppression properties of both proteins.
2. Results
2.1. HcPro Induces SIE in a Sequence Specific Manner
Here we investigated the ability of this protein to inhibit a superinfecting virus using the PVYO and PVYNTN strains and their encoded HcPro cistrons (designated here HcProO and HcProNTN, respectively). HcPro, expressed under the control of cauliflower mosaic virus 35S promoter, was agroinfiltrated to one side of the midrib of N. tabacum leaf blade and the other side was agroinfiltrated with an empty vector (negative control). After 2 days, PVYO or PVYNTN was mechanically inoculated to the whole leaf. At 7 days post virus inoculation, total RNA was extracted, and viral RNA quantified using reverse transcription quantitative PCR (RT-qPCR). Results from four independent experiments showed that tissues expressing HcProO displayed lower levels of PVYO RNA than control tissues (Figure 2A). In contrast, HcProNTN was observed to cause an increase in levels of PVYO RNA. Correspondingly, levels of PVYNTN were significantly lower in tissues expressing HcProNTN (Figure 2B) while levels of PVYO were higher, compared with control samples. Thus, HcPro inhibits the cognate virus strain while enhancing levels of the other PVY strain. These results suggest that HcPro restricts entry and/or replication of the cognate virus, i.e., it elicits exclusion of the superinfecting cognate virus.
PVYN-Wi and PVYN:O belong to the same PVYN cluster as PVYNTN, and their encoded HcPro cistrons are within the same PVYN recombinant fragment with only one amino acid substitution between HcProNTN and HcPro of PVYN-Wi (K272N), and that of PVYN:O (A48S) (Supplementary Figure S1). We therefore reasoned that HcProNTN, but not HcProO, would likely elicit superinfection of PVYN-Wi and PVYN:O, based on the specificity of SIE. To investigate, HcProO and HcProNTN were agroinfiltrated into N. tabacum, followed by inoculation of PVYN-Wi and of PVYN:O as described above. Viral RNA quantification showed that tissues expressing HcProNTN recorded lower levels of PVYN-Wi and PVYN:O RNA than control tissues (Figure 2C and 2D). Accordingly, tissues expressing HcProO displayed lower levels of PVYN:O RNA, suggesting that sequence elements with which HcPro interacts to induce SIE are conserved between PVYO and PVYN:O. In contrast, tissues expressing HcProO showed significantly higher levels of PVYN-Wi (Figure 2D). These results are consistent with the specificity of the SIE mechanism. We note that HcProNTN, HcProN-Wi, and HcProN:O, which all have the 1GVMD4 motif, show 36 amino acid differences with HcProO (Supplementary Figure S1). Together, these data show that PVY deploys HcPro to regulate levels of the homologous PVY strain genomes in a sequence specific manner.
2.2. The Four N-Terminal Amino Acid Residues Provide Specificity of HcPro Induction of SIE
2.2.1. Characterization of HcPro Induction of PVYO SIE
Given that HcProO and HcProNTN restrict the homologous strain while enhancing levels of the other strain, we reasoned that there are amino acid residues or motifs that cause this dichotomy in the behavior of HcPro towards the two PVY strains. Thus, we swapped amino acid residues or motifs, which are either highly conserved between the two PVY clusters (1GVMD4 (HcProNTN)/1RVLN4 (HcProO), show considerable biochemical differences [245NRL247 (HcProNTN)/245IRK247(HcProO)] or were previously reported to cause differences in PVY strain symptoms [E419 (HcProNTN)/D419 (HcProO)]. The constructs were expressed from pEarley100 and the SIE induction properties of these constructs were examined following procedures described above. Results from four independent experiments showed that similar to results recorded above, tissues expressing HcProO recorded lower levels of PVYO, while HcProNTN had the opposite effect (Figure 3A-i & 3A-ii). Like in HcProNTN, tissues expressing HcProO(GVMD) (HcProO containing 1GVMD4 from HcProNTN) were found to display enhanced levels of PVYO RNA compared with tissues containing the empty vector (Figure 3A-iii). This outcome suggests that 1RVLN4 is involved in HcProO induction of PVYO SIE, and 1GVMD4 from HcProNTN likely mediates HcProNTN enhancement of PVYO levels. Furthermore, no significant differences in PVYO RNA levels were observed between tissues expressing HcProNTN(RVLN) and the negative control (Figure 3A-iv), indicating that 1GVMD4 is involved in the ability of HcProNTN to enhance PVYO levels. Also, unlike HcProNTN, HcProNTN(IRK) did not significantly enhance PVYO (Figure 3A-vi), suggesting a role for 245NRL247 motif in the ability of HcProNTN to enhance PVYO levels. This indicates that in addition to the first amino acid residues, other amino acid residues are involved in an effective HcPro induction of SIE. Correspondingly, tissues expressing HcProO(D419E) and HcProNTN(E419D) did not display significantly different levels of PVYO (Figure 3A-vii & 3A-viii), indicating that these amino acids are required for HcPro induction of SIE.
2.2.2. HcProO Does not Induce Exclusion of PVYO
In the experiments described above, HcPro was expressed prior to inoculation of PVYO, which was therefore the superinfecting virus. To determine whether HcPro will have a similar influence on a resident or primary virus, HcPro cistrons, and HcPro mutants were each agroinfiltrated on one side of the leaf, and an empty vector on the other side of leaves systemically infected by PVYO at 21 days post infection (dpi). Interestingly, unlike the resident HcProO, the superinfecting HcProO did not inhibit PVYO, while HcProNTN and HcProO(GVMD) caused a decline in the level of PVYO (Figure 3B-i, 3B-ii & 3B-iii). These differential observations between a primary and a secondary infection are consistent with the mechanism of SIE, which occurs only on a superinfecting virus [28,29,30]. Further analyses showed that, unlike the resident HcProNTN(RVLN/IRK), the superinfecting HcProNTN(RVLN/IRK) caused an increase in the levels of PVYO (Figure 3B-v). Taken together, these data show that HcProO has evolved to regulate the superinfecting cognate PVYO, but not a resident virus.
2.2.3. HcProNTN Induction of PVYNTN SIE
We also characterized induction of PVYNTN SIE in leaf tissues expressing mutants of HcProNTN and HcProO. Here, similar to observations made above (Figure 2A), tissues expressing HcProNTN, but not those expressing HcProO, displayed lower levels of PVYNTN RNA compared with control tissues (Figure 4A-i & Figure 4A-ii). However, unlike HcProNTN, tissues expressing HcProNTN(RVLN) did not display lower levels of PVYNTN RNA, indicating that 1GVMD2 plays the same role in HcProNTN induction of PVYNTN SIE as does 1RVLN2 in HcProO induction of PVYO SIE. Strikingly, replacement of 245NRL247 with 245IRK247 from HcProO reconstituted the ability of HcProNTN(RVLN) to induce PVYNTN SIE as evidenced by the fact that HcProNTN(RVLN/IRK) caused substantial reduction in levels of a superinfecting PVYNTN RNA (Figure 4A-v). The role of 245IRK247 in reconstitution of SIE is confirmed by the fact that HcProNTN(IRK) also induced SIE of PVYNTN (4A-vi). Furthermore, mutants HcProNTN(D419E) and HcProNTN(E419D) showed no exclusion of PVYNTN, thus confirming a role for Asp-419 and Glu-419, respectively, in HcPro induction of SIE.
2.2.4. The Superinfecting HcPro Does not Induce Exclusion of a Resident PVYNTN
We also investigated whether HcProNTN inhibits a resident PVYNTN. Like for a resident PVYO above, N. tabacum leaves systemically infected by PVYNTN were infiltrated with HcPro constructs, and with an empty vector, as described above. Analysis of PVYNTN RNA levels showed that unlike observations made on a superinfecting PVYNTN, levels of the resident PVYNTN RNA were enhanced by a superinfecting HcProNTN (Figure 4B-i & Figure 4B-ii). Also, in contrast to the superinfecting PVYNTN experiment in Figure 4A, HcProNTN(IRK), as well as HcProNTN(RVLN/IRK) were found to enhance the levels of a resident PVYNTN RNA (Figure 4B-v & Figure 4B-vi). Together, these observations emphasize that HcPro is a typical inducer of SIE and may deploy its other function of suppressor of RNA silencing to a resident PVY.
2.2.5. Subcellular Localization Does not Influence Behavior of HcPro Mutants
Subcellular localization is critical for protein function, as it determines its access to interaction partners and provides the cellular environment needed for interactions. To determine whether differences in HcPro construct induction of SIE was due to subcellular localization, we investigated subcellular localization using confocal microscopy. These constructs were cloned into pEarleygate101, which contains a C-terminally fused yellow fluorescent protein (YFP), and transformed into Agrobacterium tumifaciens strain GV3101, cultures of which were infiltrated into N. benthamina plant leaves. Confocal microscope imaging at 48 hours post infiltration showed that HcPro localizes in the cytoplasm and nucleus. All mutants showed a similar pattern of distribution between the nucleus and cytoplasm as wild-type HcPro (Figure 5). These observations show that cellular localization did not influence SIE induction.
Together, these observations show that HcPro mediated superinfection exclusion is strain-specific, and that HcPro of a superinfecting PVY is unable to exclude a resident cognate PVY strain. In SIE, specificity is provided mainly, but not exclusively, by the first four amino acid residues of the HcPro cistron. The results also show that HcProO exclusion of PVYO is stronger than HcProNTN exclusion of PVYNTN.
2.3. HcPro Induction of SIE and RNA Silencing Suppression Are Regulated Through Distinct Mechanisms
2.3.1. HcPro Suppression of PTGS
From this study, it is clear that HcPro elicits exclusion of a superinfecting cognate PVY strain (cis-inhibition). It also mediates increased levels of a different viral strain, likely through its canonical RNA silencing suppression function. SIE and suppression of RNA silencing produce opposing outcomes since silencing suppressors cause increases in virus levels by countering the host antiviral defense. This contrasts with SIE, which limits levels of a superinfecting homologous virus. Thus, there is a dichotomy in both HcPro functions, which are likely mediated by different amino acid residues and/or motifs. To characterize RNA silencing suppression properties of HcPro mutants described above, we used the classic GFP silencing in N. benthamiana gfp-transgenic plants model [48]. First, we assessed suppression of post transcriptional gene silencing (PTGS) of RNA expressed from the 35S promoter. HcPro constructs described above, and GFP cloned into plasmid pEarley100 were transformed into Agrobacterium as described above. Each construct was then co-infiltrated with GFP into N. benthamiana line 16c plants as described previously [49,50,51]; control plants were co-infiltrated with an empty pEarley100 vector and GFP.
GFP fluorescence in infiltrated leaf patches was monitored with a handheld longwave UV lamp. Imaging results showed that all infiltrated N. benthamiana 16c leaf patches displayed bright green fluorescence within 2 days post infiltration (dpi), and by 5 dpi, there was a strong green fluorescence (Figure 6A). However, by 8 dpi, control leaves infiltrated with an empty vector and GFP had lost most of the green fluorescence, consistent with silence of gfp gene. In contrast, leaf patches expressing HcProO, and HcProNTN, as well as HcProO(GVMD) and HcProNTN(IRK) continued to display green fluorescence (Figure 6B), which persisted for up to 6 more days before being replaced by red chlorophyll fluorescence.
GFP levels were quantified using anti-GFP antibodies (α-GFP). Here, total plant protein was extracted at 5 and at 12 dpi from infiltrated leaves and GFP levels determined in an immunoblot analysis. This analysis showed that at 5 dpi, there were similar levels of GFP in all infiltrated leaves (Figure 7). However, at 12 dpi, there were lower levels of GFP in samples expressing HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProO(D419E), HcProNTN(E419D), and pEarley100 vector, while samples expressing HcProO(GVMD) and HcProNTN(IRK) showed similarly high levels of GFP as tissues expressing wild-type HcProO and HcProNTN. These observations indicate that swapping “1RVLN4” from HcProO to HcProNTN negatively affects the ability of HcProNTN to suppress RNA silencing; in contrast, “1GVMD4” appears not to affect HcProO suppression of silencing.
The pEarley101 plasmid to which HcPro constructs were cloned for subcellular localization contains a C-terminal HA tag fusion in addition to YFP. This allowed us to confirm that these HcPro constructs were expressed by carrying out an immunoblot analysis using anti-HA tag antibodies (α-HA). All HcPro constructs were detected (Figure 7), confirming expression. In these immunoblot assays, the heat shock complex 70 (HSC70) housekeeping protein was the loading control.
3. Discussion
This study has shown that one of the PVY HcPro cistron multiple functions in the virus infection cycle is induction of superinfection exclusion (SIE). SIE is a mechanism in which a resident or primary virus prevents homologous superinfecting viruses from entry into the cell [38,52]. Mounting evidence indicates that during co-infection by related viruses, including potyvirus strains, the co-infecting viruses largely mutually exclude and colonize separate cell clusters [53,54,55]. HcPro likely contributes to this segregation by excluding the superinfecting cognate virus. HcPro induction of SIE is particularly important because of the existence of multiple prevailing PVY strains, which routinely co-infect the same plant, a phenomenon that is common in viral infections [54,56,57]. It has been suggested that SIE is a winning strategy for viruses in the short-term because it protects a cell from being infected by competing viruses, giving the resident virus a greater chance of proliferating [58]. However, excluding related virus strains promotes homogeneity in virus populations, and this prevents recombination, and emergence of beneficial mutations that produce new adaptations leading to a more fit population in the long-term. Therefore, HcPro induction of SIE likely influences virus population structure and thus has an impact on virus evolution.
Unsurprisingly, inhibition of secondary infection by the primary virus has been suggested to be “collateral damage” of virus autoregulation [38] where the SIE mechanism is unable to distinguish between own progeny genomes and homologous superinfecting virus genomes. Naturally, viruses must limit exponential replication where progeny genomes replicate unchecked. To do this, the parent virus must develop a mechanism to prevent progeny genomes from replicating and overwhelming the system. Hence, these large progeny virus populations are the intended target of SIE, while the few homologous superinfectors are only an unintended target. This appears to be the mechanism exhibited by HcPro, given that in this study, HcProO and HcProNTN were observed to limit the levels of the cognate PVY strain and not those of the other strain. These observations partly explain why unrelated viruses generally interact in a synergistic manner, which has a facilitative effect on one or both viral partners, while related viruses antagonize [54,59]. It is not surprising that a superinfecting HcPro does not elicit SIE since the resident virus already autoregulates. Therefore, the process is stoichiometric and “external” HcPro does not add to the process.
Mechanistically, the phenomenon of SAR, which is common in DNA and RNA plant virus infections [60,61,62,63], has been shown to be a result of SIE becoming systemic (organismal) [38]. Thus, it is caused by the ability of the primary virus to use at least one of its proteins to prevent proliferation of its progeny and closely related viruses throughout the plant. SAR has been known since 1929 [64], and is used to control plant viruses by infecting plants with a mild strain as protection against future infection by a more severe strain [38,39,40,41]. In this case, the mild strain as the primary virus establishes throughout the plant and excludes subsequent entry and replication by a more virulent strain, thereby protecting the plant from a severe disease.
One of the key defenses plants deploy to counter virus infection is antiviral RNA silencing, where invading viral RNA is targeted and degraded by the host. To overcome this host defense, viruses typically encode at least one protein suppressor of RNA silencing, which enables the virus to replicate and spread. HcPro is one of three identified suppressors of RNA silencing encoded by potyviruses, the others being VPg [65], and NIb (Niraula et al., 2025; unpublished data). Thus, HcPro both induces SIE, and suppresses RNA silencing. SIE and suppression of RNA silencing produce opposing outcomes since SIE limits replication while silencing suppressors cause increases in virus levels. HcPro therefore appears to function as a master “master regulator” in the virus infection cycle.
To uncouple HcPro induction of SIE and RNA silencing suppression, we used mutational analysis and showed that the two mechanisms are mediated independently. Two of the mutants that illustrate this are HcProO(GVMD) and HcProNTN(IRK). Similar to wild-type HcPro, HcProO(GVMD) in which 1RVLN4 of PVYO HcPro was replaced with 1GVMD4 from PVYNTN HcPro, was found to suppress GFP in N. benthamiana line 16c plants. However, unlike HcProO, it significantly enhanced levels of a superinfecting PVYO. This shows that these mechanisms can function independently. Also of interest is HcProNTN(IRK), which like HcProO(GVMD) suppressed RNA silencing, however, it also elicited SIE of the cognate PVYNTN strain while HcProO(GVMD) did not.
Finally, we note that SIE provides new opportunities to manage plant virus diseases by taking advantage of recent advances in biotechnological strategies for crop improvement [66,67,68]. For example, mosquitoes that were engineered to express nsP2, a SIE-inducing viral protein, were found to be resistant to subsequent infection by Sindbis virus (SINV) and chikungunya virus (CHIKV) [69]. Our study identified specific HcPro sequence elements that induce SIE. Beyond expressing a whole protein, virus resistant plants or insect vectors can be engineered to express these SIE sequence elements using biotechnological strategies in plant virus management. Correspondingly, prime editing (PE), the novel CRISPR-based genome editing method, can be used for targeted and precise modification of the genome [70]. This provides an excellent opportunity to precisely insert sequences that elicit SIE into plant or insect vectors.
4. Materials and Methods
4.1. Plasmid Construction and Inoculations
HcPro cistrons investigated in this study were RT-PCR amplified from PVY strains PVYNTN and PVYO using primers shown in Table 1. Amplification products were introduced into the entry vector pDONR/Zeo, and then shuttled into the binary vector pEarleygate100 or pEarleygate101 [71] following the GatewayTM technology protocol (Life Technologies, USA). These plasmids were transformed into A. tumefaciens strain GV3101 for transient expression in tobacco. Mutations were introduced in pDONR/Zeo HcPro clones with Q5 Site-Directed Mutagenesis Kit (New England Biolabs, USA) using primers listed in Table 1. HcPro mutants were then shuttled from pDONR/Zeo entry clone into pEarleygate100 or pEarleygate101. These constructs were introduced into A. tumefaciens (strain GV3101) using the freeze-thaw method [72], and cultures were grown at 30 °C. Agrobacterium suspensions were pelleted and resuspended in an agroinfiltration buffer infiltration buffer (10 mM MES, pH 5.5; 10 mM MgSO4; 100 μM acetosyringone) at an optical density at 600 nm of 0.8.
PVY strains were mechanically inoculated to three- to four-week-old N. tabacum cv. Samsun plants. Virus inoculum was prepared by homogenizing systemically infected leaves of N. tabacum plants in a 0.01 M phosphate buffer, pH 7.0 at a dilution of 1/20 (w/v). Plants were placed in a room at 16 h light and 22 ± 3 °C temperature.
4.2. Isolation of RNA and Reverse Transcription Quantitative PCR (RT-qPCR)
Total RNA was isolated from leaf samples using the RNeasy Mini kit (Qiagen, USA) with modifications. Here, approximately 200 mg of ground leaf and tuber tissues, respectively, were aliquoted for total RNA extraction using a leaf extraction buffer (5 M guanidine thiocyanate, 50 mM Tris-HCl pH 8, 10 mM EDTA, 2% N-lauroylsarcosine). The quality and quantity of RNA were determined using a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, USA) and agarose gel electrophoresis. The Invitrogen SuperScript III Platinum™ SYBR™ green one-step RT-qPCR Kit (Thermo Fisher Scientific, USA) was used to quantify viral RNA in 50 ng of total RNA on a Light Cycler 480 system (Roche Diagnostics). The 10 µl reaction mix contained 5 µl SYBR Green Supermix, 0.25 µl SuperScript III reverse transcriptase (RT)/Platinum® Taq DNA polymerase enzyme mix, 0.4 µl 100 µM forward and reverse primers, and 2 µl of total RNA. The RT-qPCR conditions were as follows: cDNA synthesis at 50ºC for 15 min; qPCR cycling at 95ºC for 5 min and 45 three-step cycles, each at 95ºC for 15 sec, 60ºC for 30 sec, and 72ºC for 10 sec, followed by a final melting curve of 65-95ºC for 5 sec.
4.3. Determination of Relative Viral RNA Levels
Here, we used elongation factor 1 (EF1) as the housekeeping gene. To compensate for potential variation between runs and plates, EF1 Ct values were normalized by subtracting the median Ct value from individual EF1 Ct values in each replicate. Normalized EF1 Ct values were then subtracted from individual target Ct values to obtain normalized target Ct values, which were used to calculate target ΔCt and ΔΔCt as we described recently [73]. In the calculation of relative viral RNA accumulation, we removed variation from background signals unrelated to viral RNA target by subjecting healthy tissues to the same amplification conditions as for each treatment [74], followed by calculation of mean ΔCt and ΔΔCt of the background noise. These background ΔCt and ΔΔCt values were then subtracted from experimental ΔCt and ΔΔCt values, respectively, the results of which were used to calculate individual 2-ΔΔCt values [47] to obtain relative viral RNA accumulation. Error bars were computed to indicate the standard deviation of three different technical repeats from three biological replicates; significant difference between 2-ΔΔCt values were determined using a t-test, and differences were considered statistically significant when the p-value was less than 0.05.
4.4. Western Blot Analysis
At 5 and 12 dpi, leaves co-infiltrated with GFP and the respective HcPro constructs, were extracted in a protein extraction buffer [50] and the proteins separated in a 4–12% SDS-PAGE reducing gel (Genscript, USA). The proteins were electrophoretically blotted to PVDF membranes, then blocked in 5% milk in TBST for 1 h followed by incubation at 4 °C overnight with rocking in rabbit anti-GFP primary antibody (GenScript, USA) or rabbit anti-HA tag primary antibody (Proteintech, USA), each at a dilution of 1:5,000. Membranes were washed three times (5 min each) with TBST, then incubated with goat anti-rabbit IgG (H+L) secondary antibody, HRP (Invitrogen, USA) at a dilution of 1:1,000, followed by three 5-min washes with TBST. Chemiluminescent signals were generated using SuperSignal™ West Atto Ultimate Sensitivity Substrate (Thermo Fisher Scientific, USA), and emitted light captured using a G:Box and GeneSys software (Syngene, USA).
4.5. Confocal Microscopy
Leaf pieces were excised, gently vacuum infiltrated with water and placed on a slide and covered with a coverslip for confocal imaging. Images were acquired on an upright Zeiss LSM 780 laser-scanning microscope (Carl Zeiss, Inc., Oberkochen, Germany) using a Zeiss 40X C-Apochromat (NA 1.2) objective lens. Multichannel images of YFP and chloroplast autofluorescence were acquired using the 488 nm line of an Argon/Krypton laser with the 500 to 550 band pass and 650 long-pas 390/ emission filters, respectively. Images were captured as single optical sections or as a z-series of optical sections, and z-series data sets were displayed as single maximum intensity projection generated with Zeiss Zen Black software vSP2.
5. Conclusion
The phenomenon of superinfection exclusion (SIE) plays a key role in virus infection dynamics and influences viral fitness and population diversity, and evolution. This is because excluding a related competing secondary virus from the cell that has been successfully infected by the primary virus, eliminates competition for host resources, given that multiple infections allow for the exchange of genetic material between viruses through recombination and reassortment (for segmented genome virus); this increases diversity and improves the efficiency of selection. However, this may decrease fitness by promoting the presence of deleterious mutants at low frequencies [75]. Our study shows that HcPro, a suppressors of RNA silencing, induces SIE exclusion of closely related PVY strains. Thus, HcPro contributes in shaping the genetic structure of PVY and its evolution.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, V.N.F.; methodology, V.N.F.; investigation, P.M.N.; resources, V.N.F.; writing—original draft preparation, V.N.F.; writing—review and editing, V.N.Y., P.M.N.; project administration, V.N.F.; funding acquisition, V.N.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Institute of Food and Agriculture, grant number 2020-67018-31180.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Singh, R.P.; Valkonen, J.P.T.; Gray, S.M.; Boonham, N.; Jones, R. a. C.; Kerlan, C.; Schubert, J. Discussion Paper: The Naming of Potato Virus Y Strains Infecting Potato. Arch. Virol. 2008, 153, 1–13. [Google Scholar] [CrossRef]
- Hu, X.; Karasev, A.V.; Brown, C.J.; Lorenzen, J.H. Sequence Characteristics of Potato Virus Y Recombinants. J. Gen. Virol. 2009, 90, 3033–3041. [Google Scholar] [CrossRef] [PubMed]
- Rowley, J.S.; Gray, S.M.; Karasev, A. V. Screening Potato Cultivars for New Sources of Resistance to Potato Virus Y. Am. J. Potato Res. 2014 921 2015, 92, 38–48. [Google Scholar] [CrossRef]
- Quenouille, J.; Vassilakos, N.; Moury, B. Potato Virus Y: A Major Crop Pathogen That Has Provided Major Insights into the Evolution of Viral Pathogenicity. Mol. Plant Pathol. 2013, 14, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Merits, A.; Rajamäki, M.-L.; Lindholm, P.; Runeberg-Roos, P.; Kekarainen, T.; Puustinen, P.; Mäkeläinen, K.; Valkonen, J.P.T.; Saarma, M. Proteolytic Processing of Potyviral Proteins and Polyprotein Processing Intermediates in Insect and Plant Cells. J. Gen. Virol. 2002, 83, 1211–1221. [Google Scholar] [CrossRef]
- Chung, B.Y.-W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An Overlapping Essential Gene in the Potyviridae. Proc. Natl. Acad. Sci. 2008, 105, 5897–5902. [Google Scholar] [CrossRef]
- Revers, F.; García, J.A. Molecular Biology of Potyviruses. Adv. Virus Res. 2015, 92, 101–199. [Google Scholar] [CrossRef]
- Urcuqui-Inchima, S.; Haenni, A.-L.; Bernardi, F. Potyvirus Proteins: A Wealth of Functions. Virus Res. 2001, 74, 157–175. [Google Scholar] [CrossRef]
- Kozieł, E.; Surowiecki, P.; Przewodowska, A.; Bujarski, J.J.; Otulak-Kozieł, K. Modulation of Expression of PVYNTN RNA-Dependent RNA Polymerase (NIb) and Heat Shock Cognate Host Protein HSC70 in Susceptible and Hypersensitive Potato Cultivars. Vaccines 2021, 9, 1254. [Google Scholar] [CrossRef]
- Olspert, A.; Chung, B.Y.-W.; Atkins, J.F.; Carr, J.P.; Firth, A.E. Transcriptional Slippage in the Positive-Sense RNA Virus Family Potyviridae. EMBO Rep. 2015, 16, 995–1004. [Google Scholar] [CrossRef]
- Rodamilans, B.; Valli, A.; Mingot, A.; San León, D.; Baulcombe, D.; López-Moya, J.J.; García, J.A. RNA Polymerase Slippage as a Mechanism for the Production of Frameshift Gene Products in Plant Viruses of the Potyviridae Family. J. Virol. 2015, 89, 6965–6967. [Google Scholar] [CrossRef]
- Chai, M.; Wu, X.; Liu, J.; Fang, Y.; Luan, Y.; Cui, X.; Zhou, X.; Wang, A.; Cheng, X. P3N-PIPO Interacts with P3 via the Shared N-Terminal Domain To Recruit Viral Replication Vesicles for Cell-to-Cell Movement. J. Virol. 2020, 94, e01898-19. [Google Scholar] [CrossRef]
- Carrington, J.C.; Herndon, K.L. Characterization of the Potyviral HC-pro Autoproteolytic Cleavage Site. Virology 1992, 187, 308–315. [Google Scholar] [CrossRef]
- Verchot, J.; Herndon, K.L.; Carrington, J.C. Mutational Analysis of the Tobacco Etch Potyviral 35-kDa Proteinase: Identification of Essential Residues and Requirements for Autoproteolysis. Virology 1992, 190, 298–306. [Google Scholar] [CrossRef]
- Anandalakshmi, R.; Pruss, G.J.; Ge, X.; Marathe, R.; Mallory, A.C.; Smith, T.H.; Vance, V.B. A Viral Suppressor of Gene Silencing in Plants. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 13079–13084. [Google Scholar] [CrossRef]
- Kasschau, K.D.; Carrington, J.C. A Counterdefensive Strategy of Plant Viruses: Suppression of Posttranscriptional Gene Silencing. Cell 1998, 95, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Pruss, G.; Ge, X.; Shi, X.M.; Carrington, J.C.; Bowman Vance, V. Plant Viral Synergism: The Potyviral Genome Encodes a Broad-Range Pathogenicity Enhancer That Transactivates Replication of Heterologous Viruses. Plant Cell 1997, 9, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, K.I.; Eskelin, K.; Bašić, M.; De, S.; Lõhmus, A.; Varjosalo, M.; Mäkinen, K. Molecular Insights into the Function of the Viral RNA Silencing Suppressor HCPro. Plant J. Cell Mol. Biol. 2016, 85, 30–45. [Google Scholar] [CrossRef]
- Hu, S.-F.; Wei, W.-L.; Hong, S.-F.; Fang, R.-Y.; Wu, H.-Y.; Lin, P.-C.; Sanobar, N.; Wang, H.-P.; Sulistio, M.; Wu, C.-T.; et al. Investigation of the Effects of P1 on HC-pro-Mediated Gene Silencing Suppression through Genetics and Omics Approaches. Bot. Stud. 2020, 61, 22. [Google Scholar] [CrossRef] [PubMed]
- Govier, D.A.; Kassanis, B.; Pirone, T.P. Partial Purification and Characterization of the Potato Virus Y Helper Component. Virology 1977, 78, 306–314. [Google Scholar] [CrossRef]
- Gallo, A.; Valli, A.; Calvo, M.; García, J.A. A Functional Link between RNA Replication and Virion Assembly in the Potyvirus Plum Pox Virus. J. Virol. 2018, 92, e02179-17. [Google Scholar] [CrossRef]
- Valli, A.; Gallo, A.; Calvo, M.; Pérez, J. de J.; García, J.A. A Novel Role of the Potyviral Helper Component Proteinase Contributes To Enhance the Yield of Viral Particles. J. Virol. 2014, 88, 9808–9818. [Google Scholar] [CrossRef]
- Kd, K.; S, C.; Jc, C. Genome Amplification and Long-Distance Movement Functions Associated with the Central Domain of Tobacco Etch Potyvirus Helper Component-Proteinase. Virology 1997, 228. [Google Scholar] [CrossRef]
- Rojas, M.R.; Zerbini, F.M.; Allison, R.F.; Gilbertson, R.L.; Lucas, W.J. Capsid Protein and Helper Component-Proteinase Function as Potyvirus Cell-to-Cell Movement Proteins. Virology 1997, 237, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Sáenz, P.; Salvador, B.; Simón-Mateo, C.; Kasschau, K.D.; Carrington, J.C.; García, J.A. Host-Specific Involvement of the HC Protein in the Long-Distance Movement of Potyviruses. J. Virol. 2002, 76, 1922–1931. [Google Scholar] [CrossRef]
- Plisson, C.; Drucker, M.; Blanc, S.; German-Retana, S.; Le Gall, O.; Thomas, D.; Bron, P. Structural Characterization of HC-Pro, a Plant Virus Multifunctional Protein*. J. Biol. Chem. 2003, 278, 23753–23761. [Google Scholar] [CrossRef] [PubMed]
- Valkonen, J.P.T.; Rajamäki, M.-L.; Kekarainen, T. Mapping of Viral Genomic Regions Important in Cross-Protection between Strains of a Potyvirus. Mol. Plant-Microbe Interact. MPMI 2002, 15, 683–692. [Google Scholar] [CrossRef]
- Bergua, M.; Zwart, M.P.; El-Mohtar, C.; Shilts, T.; Elena, S.F.; Folimonova, S.Y. A Viral Protein Mediates Superinfection Exclusion at the Whole-Organism Level but Is Not Required for Exclusion at the Cellular Level. J. Virol. 2014, 88, 11327–11338. [Google Scholar] [CrossRef]
- Folimonova, S.Y. Superinfection Exclusion Is an Active Virus-Controlled Function That Requires a Specific Viral Protein. J. Virol. 2012, 86, 5554–5561. [Google Scholar] [CrossRef]
- Folimonova, S.Y.; Harper, S.J.; Leonard, M.T.; Triplett, E.W.; Shilts, T. Superinfection Exclusion by Citrus Tristeza Virus Does Not Correlate with the Production of Viral Small RNAs. Virology 2014, 468–470, 462–471. [Google Scholar] [CrossRef]
- Tatineni, S.; French, R. The Coat Protein and NIa Protease of Two Potyviridae Family Members Independently Confer Superinfection Exclusion. J. Virol. 2016, 90, 10886–10905. [Google Scholar] [CrossRef]
- Nunna, H.; Qu, F.; Tatineni, S. P3 and NIa-Pro of Turnip Mosaic Virus Are Independent Elicitors of Superinfection Exclusion. Viruses 2023, 15, 1459. [Google Scholar] [CrossRef] [PubMed]
- Licciardello, G.; Scuderi, G.; Russo, M.; Bazzano, M.; Bar-Joseph, M.; Catara, A.F. Minor Variants of Orf1a, P33, and P23 Genes of VT Strain Citrus Tristeza Virus Isolates Show Symptomless Reactions on Sour Orange and Prevent Superinfection of Severe VT Isolates. Viruses 2023, 15, 2037. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Sun, K.; Zhou, X.; Jackson, A.O.; Li, Z. The Matrix Protein of a Plant Rhabdovirus Mediates Superinfection Exclusion by Inhibiting Viral Transcription. J. Virol. 2019, 93, e00680-19. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.-C.; Li, W.; Sui, J.; Marasco, W.; Choe, H.; Farzan, M. Influenza A Virus Neuraminidase Limits Viral Superinfection. J. Virol. 2008, 82, 4834–4843. [Google Scholar] [CrossRef]
- Lee, Y.-M.; Tscherne, D.M.; Yun, S.-I.; Frolov, I.; Rice, C.M. Dual Mechanisms of Pestiviral Superinfection Exclusion at Entry and RNA Replication. J. Virol. 2005, 79, 3231–3242. [Google Scholar] [CrossRef]
- Schmidt, A.K.; Fitzpatrick, A.D.; Schwartzkopf, C.M.; Faith, D.R.; Jennings, L.K.; Coluccio, A.; Hunt, D.J.; Michaels, L.A.; Hargil, A.; Chen, Q.; et al. A Filamentous Bacteriophage Protein Inhibits Type IV Pili To Prevent Superinfection of Pseudomonas Aeruginosa. mBio 2022, e0244121. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Zhang, S.; Guo, Q.; Sun, R.; Wei, T.; Qu, F. A New Mechanistic Model for Viral Cross Protection and Superinfection Exclusion. Front. Plant Sci. 2018, 9, 40. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, W.; Gao, Y.; Meng, F.; Nie, X.; Bai, Y. Potato Virus Y Strain N-Wi Offers Cross-Protection in Potato Against Strain NTN-NW by Superior Competition. Plant Dis. 2022, 106, 1566–1572. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, S.; Wang, Z.; Yu, C.; Yuan, X. A New Strategy of Cross-Protection Based on Attenuated Vaccines: RNA Viruses Are Used as Vectors to Control DNA Viruses. Agronomy 2023, 13, 2334. [Google Scholar] [CrossRef]
- Hernando, Y.; Aranda, M.A. Cross-Protection against Pepino Mosaic Virus, More than a Decade of Efficient Disease Control. Ann. Appl. Biol. 2024, 184, 174–182. [Google Scholar] [CrossRef]
- Khatoon, H.; Chavan, D.D.; Meena, V.K.; Kashyap, A.S.; Sathiyaseelan, K.; Elangovan, M.; Patil, L.; Meena, B.R.; Gauns, A.; Bhattacharyya, U.K.; et al. Differentiation and Validation of Mild and Severe Strains of Citrus Tristeza Virus through Codon Usage Bias, Host Adaptation, and Biochemical Profiling. Front. Microbiol. 2025, 16. [Google Scholar] [CrossRef] [PubMed]
- Licciardello, G.; Scuderi, G.; Russo, M.; Bazzano, M.; Paradiso, G.; Bar-Joseph, M.; Catara, A.F. Progress in Our Understanding of the Cross-Protection Mechanism of CTV-VT No-SY Isolates Against Homologous SY Isolates. Pathogens 2025, 14, 701. [Google Scholar] [CrossRef] [PubMed]
- Gozzo, F.; Faoro, F. Systemic Acquired Resistance (50 Years after Discovery): Moving from the Lab to the Field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Alexandersson, E.; Mulugeta, T.; Lankinen, Å.; Liljeroth, E.; Andreasson, E. Plant Resistance Inducers against Pathogens in Solanaceae Species—From Molecular Mechanisms to Field Application. Int. J. Mol. Sci. 2016, 17, 1673. [Google Scholar] [CrossRef]
- Glais, L.; Faurez, F.; Tribodet, M.; Boulard, F.; Jacquot, E. The Amino Acid 419 in HC-Pro Is Involved in the Ability of PVY Isolate N605 to Induce Necrotic Symptoms on Potato Tubers. Virus Res. 2015, 208, 110–119. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Voinnet, O.; Vain, P.; Angell, S.; Baulcombe, D.C. Systemic Spread of Sequence-Specific Transgene RNA Degradation in Plants Is Initiated by Localized Introduction of Ectopic Promoterless DNA. Cell 1998, 95, 177–187. [Google Scholar] [CrossRef]
- Fondong, V.N.; Reddy, R.V.C.; Lu, C.; Hankoua, B.; Felton, C.; Czymmek, K.; Achenjang, F. The Consensus N-Myristoylation Motif of a Geminivirus AC4 Protein Is Required for Membrane Binding and Pathogenicity. Mol. Plant-Microbe Interact. MPMI 2007, 20, 380–391. [Google Scholar] [CrossRef]
- Chowda-Reddy, R.V.; Achenjang, F.; Felton, C.; Etarock, M.T.; Anangfac, M.-T.; Nugent, P.; Fondong, V.N. Role of a Geminivirus AV2 Protein Putative Protein Kinase C Motif on Subcellular Localization and Pathogenicity. Virus Res. 2008, 135, 115–124. [Google Scholar] [CrossRef]
- Chowda-Reddy, R.V.; Dong, W.; Felton, C.; Ryman, D.; Ballard, K.; Fondong, V.N. Characterization of the Cassava Geminivirus Transcription Activation Protein Putative Nuclear Localization Signal. Virus Res. 2009, 145, 270–278. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Sun, R.; Guo, Q.; Zhang, S.; Meulia, T.; Halfmann, R.; Li, D.; Qu, F. A Self-Perpetuating Repressive State of a Viral Replication Protein Blocks Superinfection by the Same Virus. PLOS Pathog. 2017, 13, e1006253. [Google Scholar] [CrossRef]
- Syller, J. Facilitative and Antagonistic Interactions between Plant Viruses in Mixed Infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef]
- Syller, J.; Grupa, A. Antagonistic Within-Host Interactions between Plant Viruses: Molecular Basis and Impact on Viral and Host Fitness. Mol. Plant Pathol. 2016, 17, 769–782. [Google Scholar] [CrossRef]
- Dietrich, C.; Maiss, E. Fluorescent Labelling Reveals Spatial Separation of Potyvirus Populations in Mixed Infected Nicotiana Benthamiana Plants. J. Gen. Virol. 2003, 84, 2871–2876. [Google Scholar] [CrossRef]
- Fondong, V.N.; Pita, J.S.; Rey, M.E.C.; De Kochko, A.; Beachy, R.N.; Fauquet, C.M. Evidence of Synergism between African Cassava Mosaic Virus and a New Double-Recombinant Geminivirus Infecting Cassava in Cameroon. J. Gen. Virol. 2000, 81, 287–297. [Google Scholar] [CrossRef]
- Mondal, S.; Lin, Y.-H.; Carroll, J.E.; Wenninger, E.J.; Bosque-Pérez, N.A.; Whitworth, J.L.; Hutchinson, P.; Eigenbrode, S.; Gray, S.M. Potato Virus Y Transmission Efficiency from Potato Infected with Single or Multiple Virus Strains. Phytopathology 2017, 107, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.; Fusco, D. Superinfection Exclusion: A Viral Strategy with Short-Term Benefits and Long-Term Drawbacks. PLoS Comput. Biol. 2022, 18, e1010125. [Google Scholar] [CrossRef] [PubMed]
- Tatineni, S.; Riethoven, J.-J.M.; Graybosch, R.A.; French, R.; Mitra, A. Dynamics of Small RNA Profiles of Virus and Host Origin in Wheat Cultivars Synergistically Infected by Wheat Streak Mosaic Virus and Triticum Mosaic Virus: Virus Infection Caused a Drastic Shift in the Endogenous Small RNA Profile. PloS One 2014, 9, e111577. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Rush, C.M. Evidence of Cross-Protection Between Beet Soilborne Mosaic Virus and Beet Necrotic Yellow Vein Virus in Sugar Beet. Plant Dis. 1999, 83, 521–526. [Google Scholar] [CrossRef]
- Nakazono-Nagaoka, E.; Takahashi, T.; Shimizu, T.; Kosaka, Y.; Natsuaki, T.; Omura, T.; Sasaya, T. Cross-Protection Against Bean Yellow Mosaic Virus (BYMV) and Clover Yellow Vein Virus by Attenuated BYMV Isolate M11. Phytopathology® 2009, 99, 251–257. [Google Scholar] [CrossRef]
- Reddy, R.V.C.; Dong, W.; Njock, T.; Rey, M.E.C.; Fondong, V.N. Molecular Interaction between Two Cassava Geminiviruses Exhibiting Cross-Protection. Virus Res. 2012, 163, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Ameyaw, G.A.; Domfeh, O.; Dzahini-Obiatey, H.; Ollennu, L. a. A.; Owusu, G.K. Appraisal of Cocoa Swollen Shoot Virus (CSSV) Mild Isolates for Cross Protection of Cocoa Against Severe Strains in Ghana. Plant Dis. 2016, 100, 810–815. [Google Scholar] [CrossRef] [PubMed]
- McKinney, H.H. Mosaic Diseases in the Canary Islands, West Africa, and Gibraltar. 1929, 557-578.
- Cheng, X.; Wang, A. The Potyvirus Silencing Suppressor Protein VPg Mediates Degradation of SGS3 via Ubiquitination and Autophagy Pathways. J. Virol. 2016, 91, e01478-16. [Google Scholar] [CrossRef] [PubMed]
- Fondong, V.N.; Rey, C.; Fondong, V.N.; Rey, C. Recent Biotechnological Advances in the Improvement of Cassava; IntechOpen, 2018; ISBN 978-953-51-3741-2.
- Giudice, G.; Moffa, L.; Varotto, S.; Cardone, M.F.; Bergamini, C.; De Lorenzis, G.; Velasco, R.; Nerva, L.; Chitarra, W. Novel and Emerging Biotechnological Crop Protection Approaches. Plant Biotechnol. J. 2021, 19, 1495–1510. [Google Scholar] [CrossRef]
- Niraula, P.M.; Fondong, V.N. Development and Adoption of Genetically Engineered Plants for Virus Resistance: Advances, Opportunities and Challenges. Plants Basel Switz. 2021, 10, 2339. [Google Scholar] [CrossRef]
- Reitmayer, C.M.; Levitt, E.; Basu, S.; Atkinson, B.; Fragkoudis, R.; Merits, A.; Lumley, S.; Larner, W.; Diaz, A.V.; Rooney, S.; et al. Mimicking Superinfection Exclusion Disrupts Alphavirus Infection and Transmission in the Yellow Fever Mosquito Aedes Aegypti. Proc. Natl. Acad. Sci. 2023, 120, e2303080120. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-Replace Genome Editing without Double-Strand Breaks or Donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Earley, K.W.; Haag, J.R.; Pontes, O.; Opper, K.; Juehne, T.; Song, K.; Pikaard, C.S. Gateway-Compatible Vectors for Plant Functional Genomics and Proteomics. Plant J. Cell Mol. Biol. 2006, 45, 616–629. [Google Scholar] [CrossRef]
- Weigel, D.; Glazebrook, J. Transformation of Agrobacterium Using the Freeze-Thaw Method. CSH Protoc. 2006, 2006, pdb.prot4666. [Google Scholar] [CrossRef]
- Niraula, P.M.; Baldrich, P.; Cheema, J.A.; Cheema, H.A.; Gaiter, D.S.; Meyers, B.C.; Fondong, V.N. Antagonism and Synergism Characterize the Interactions between Four North American Potato Virus Y Strains. Int. J. Plant Biol. 2024, 15, 412–428. [Google Scholar] [CrossRef]
- Wilhelm, J.; Pingoud, A.; Hahn, M. Validation of an Algorithm for Automatic Quantification of Nucleic Acid Copy Numbers by Real-Time Polymerase Chain Reaction. Anal. Biochem. 2003, 317, 218–225. [Google Scholar] [CrossRef]
- Hunter, M.; Fusco, D. Superinfection Exclusion: A Viral Strategy with Short-Term Benefits and Long-Term Drawbacks. PLoS Comput. Biol. 2022, 18, e1010125. [Google Scholar] [CrossRef]
Figure 1.
Functional domains of HcPro and mutations introduced in this study. The N-terminal region is involved in aphid vector binding and transmission of the virus particle while the central region participates in RNA silencing suppression, genome amplification, symptom severity, viral movement, and viral capsid-binding. The C-terminal domain displays proteolytic activity and self-cleaves from the viral polyprotein [26]. Open inverted triangles indicate amino acid residues that were mutated in this study.
Figure 1.
Functional domains of HcPro and mutations introduced in this study. The N-terminal region is involved in aphid vector binding and transmission of the virus particle while the central region participates in RNA silencing suppression, genome amplification, symptom severity, viral movement, and viral capsid-binding. The C-terminal domain displays proteolytic activity and self-cleaves from the viral polyprotein [26]. Open inverted triangles indicate amino acid residues that were mutated in this study.
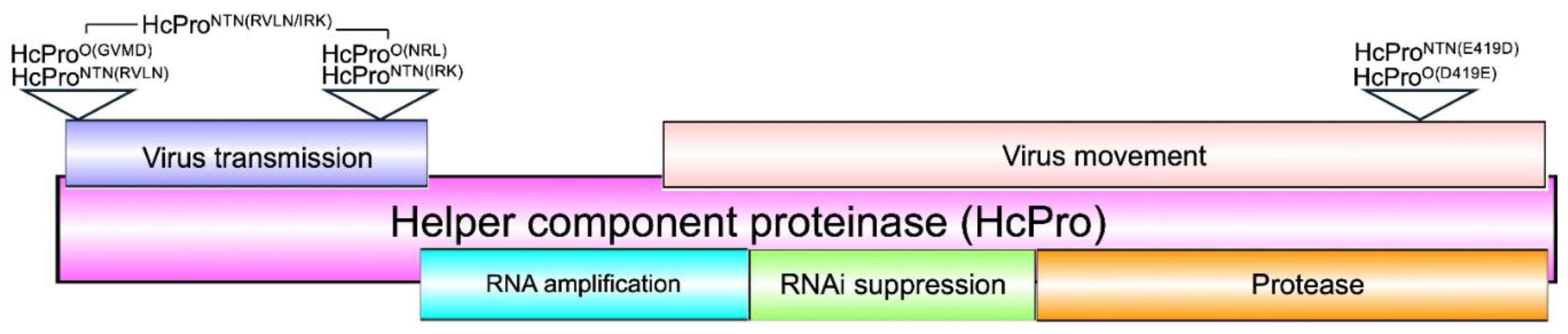
Figure 2.
HcPro induces exclusion of a superinfecting cognate PVY strain. HcPro of PVYO (HcProO) caused a decrease in PVYO RNA levels while enhancing RNA levels of PVYNTN and PVYN-Wi. In contrast, but consistent with the phenomenon of SIE, HcPro of PVYNTN (HcProNTN) induced a decrease in the RNA levels of members of the PVYN cluster, PVYNTN, PVYN-Wi, and PVYN:O and enhanced levels of PVYO. Relative RNA levels were determined using 2-ΔΔCT method [47]; 2-ΔΔCT values represent the mean, and the error bars show the standard error of the means (of four independent experiments). Pair-wise t-test was used to determine the significance between 22-ΔΔCT values; *, p <0.05; **, p <0.01; n.s., non-significant.
Figure 2.
HcPro induces exclusion of a superinfecting cognate PVY strain. HcPro of PVYO (HcProO) caused a decrease in PVYO RNA levels while enhancing RNA levels of PVYNTN and PVYN-Wi. In contrast, but consistent with the phenomenon of SIE, HcPro of PVYNTN (HcProNTN) induced a decrease in the RNA levels of members of the PVYN cluster, PVYNTN, PVYN-Wi, and PVYN:O and enhanced levels of PVYO. Relative RNA levels were determined using 2-ΔΔCT method [47]; 2-ΔΔCT values represent the mean, and the error bars show the standard error of the means (of four independent experiments). Pair-wise t-test was used to determine the significance between 22-ΔΔCT values; *, p <0.05; **, p <0.01; n.s., non-significant.
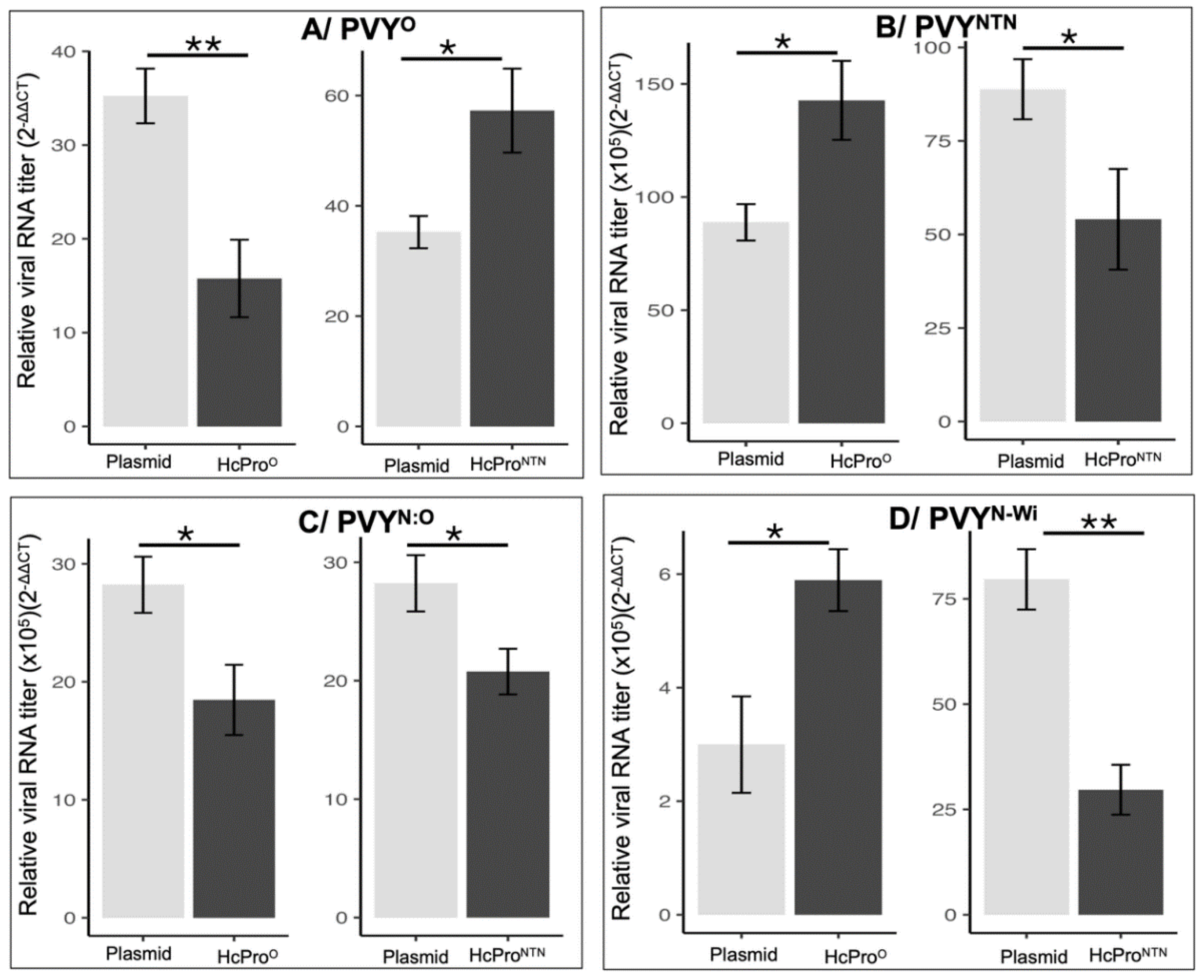
Figure 3.
The N-terminal first four amino acid residues are specificity determinants of HcPro induced exclusion of a superinfecting, but not a resident PVYO. A/ Unlike HcProO, HcProO(GVMD and HcProNTN caused an increase in levels of the superinfecting PVYO RNA, while other mutants did not significantly affect PVYO RNA levels. B/ Unlike the resident HcProO, the superinfecting HcProO did not cause a significant decrease in the level of a resident PVYO RNA, while HcProNTN and HcProO(GVMD) caused a decrease in the level of a resident PVYO RNA. Relative RNA levels were determined using 2-ΔΔCT as described above; 22-ΔΔCT values represent the mean, and the error bars show the standard error of the means (of four independent experiments). Pair-wise t-test was used to determine the significance between 22-ΔΔCT values; *, p <0.05; **, p <0.01; n.s., non-significant.
Figure 3.
The N-terminal first four amino acid residues are specificity determinants of HcPro induced exclusion of a superinfecting, but not a resident PVYO. A/ Unlike HcProO, HcProO(GVMD and HcProNTN caused an increase in levels of the superinfecting PVYO RNA, while other mutants did not significantly affect PVYO RNA levels. B/ Unlike the resident HcProO, the superinfecting HcProO did not cause a significant decrease in the level of a resident PVYO RNA, while HcProNTN and HcProO(GVMD) caused a decrease in the level of a resident PVYO RNA. Relative RNA levels were determined using 2-ΔΔCT as described above; 22-ΔΔCT values represent the mean, and the error bars show the standard error of the means (of four independent experiments). Pair-wise t-test was used to determine the significance between 22-ΔΔCT values; *, p <0.05; **, p <0.01; n.s., non-significant.
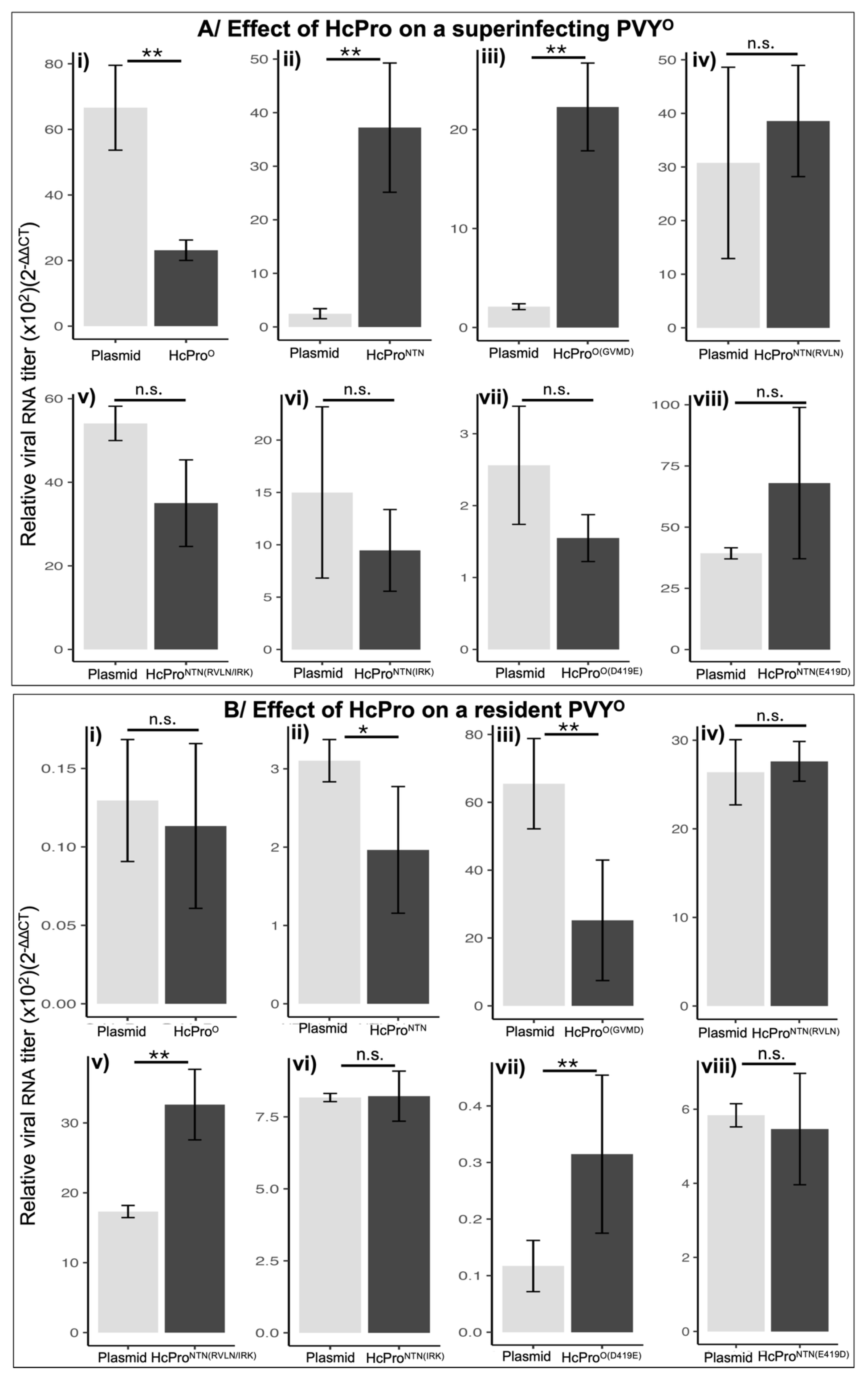
Figure 4.
The N-terminal first four amino acid residues are specificity determinants of HcPro induced exclusion of a superinfecting virus. To test the effect of mutants on PVYNTN, the experiment described in Figure 3 was replicated for a superinfecting (A/), as well as a resident (B/) PVYNTN. The 2-ΔΔCT values represent the mean, and the error bars show the standard error of the means (of four independent experiments). Pair-wise t-test was used to determine the significance between 22-ΔΔCT values; *, p <0.05; **, p <0.01; n.s., non-significant.
Figure 4.
The N-terminal first four amino acid residues are specificity determinants of HcPro induced exclusion of a superinfecting virus. To test the effect of mutants on PVYNTN, the experiment described in Figure 3 was replicated for a superinfecting (A/), as well as a resident (B/) PVYNTN. The 2-ΔΔCT values represent the mean, and the error bars show the standard error of the means (of four independent experiments). Pair-wise t-test was used to determine the significance between 22-ΔΔCT values; *, p <0.05; **, p <0.01; n.s., non-significant.
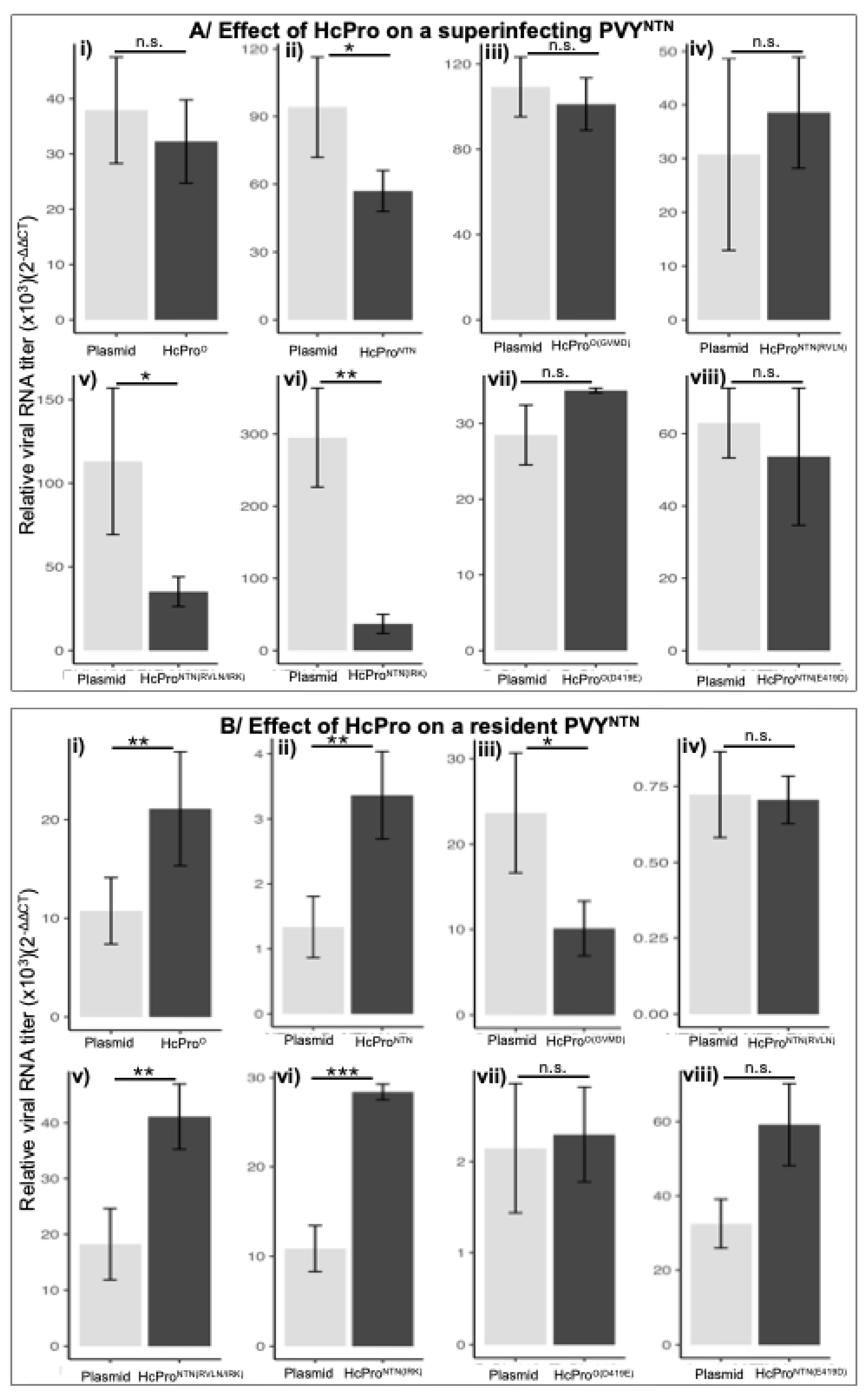
Figure 5.
Subcellular localization of HcPro and mutants using confocal microscopy. Wild-type PVYNTN and PVYO HcPro and mutants HcProO(GVMD, HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProNTN(IRK), HcProO(D419E), and HcProNTN(E419D) all localized to both the nucleus (indicated by arrows) and the cytoplasm. Images were acquired on an upright Zeiss LSM 780 laser-scanning microscope (using a Zeiss 40X C-Apochromat (NA 1.2) objective lens.
Figure 5.
Subcellular localization of HcPro and mutants using confocal microscopy. Wild-type PVYNTN and PVYO HcPro and mutants HcProO(GVMD, HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProNTN(IRK), HcProO(D419E), and HcProNTN(E419D) all localized to both the nucleus (indicated by arrows) and the cytoplasm. Images were acquired on an upright Zeiss LSM 780 laser-scanning microscope (using a Zeiss 40X C-Apochromat (NA 1.2) objective lens.
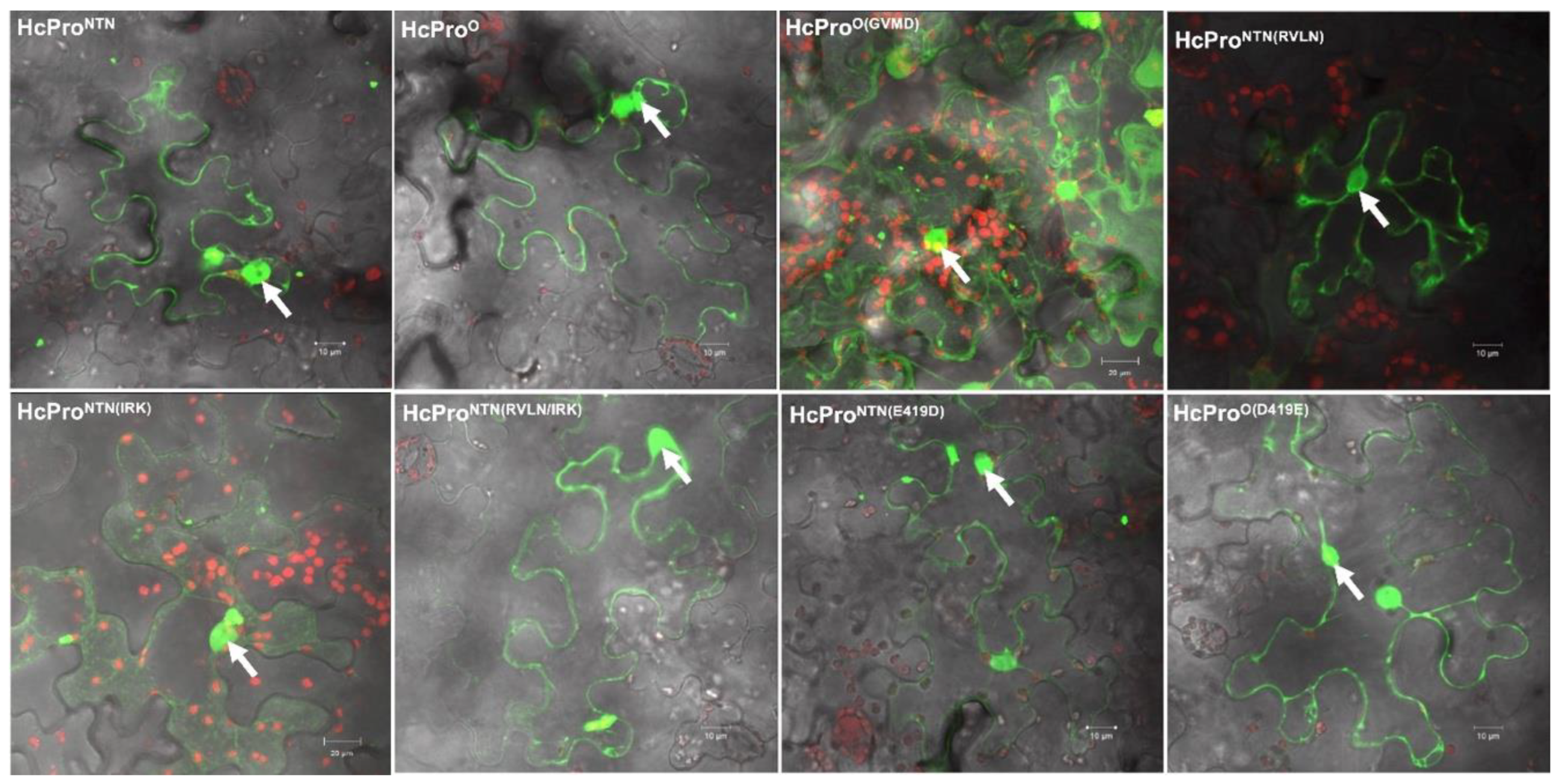
Figure 6.
PVY HcPro suppression of post transcriptional gene silencing (PTGS) in N. benthamiana line 16c GFP plants. gfp gene silencing suppression was assessed by co-infiltrating plants with GFP, and HcPro constructs. At 5 dpi, all constructs displayed bright green fluorescence. However, at 8 dpi, HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProO(D419E), HcProNTN(E419D, and GFP had lost much of the green fluorescence, while HcProNTN, HcProO, HcProO(GVMD), and HcProNTN(IRK) continued to display green fluorescence.
Figure 6.
PVY HcPro suppression of post transcriptional gene silencing (PTGS) in N. benthamiana line 16c GFP plants. gfp gene silencing suppression was assessed by co-infiltrating plants with GFP, and HcPro constructs. At 5 dpi, all constructs displayed bright green fluorescence. However, at 8 dpi, HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProO(D419E), HcProNTN(E419D, and GFP had lost much of the green fluorescence, while HcProNTN, HcProO, HcProO(GVMD), and HcProNTN(IRK) continued to display green fluorescence.
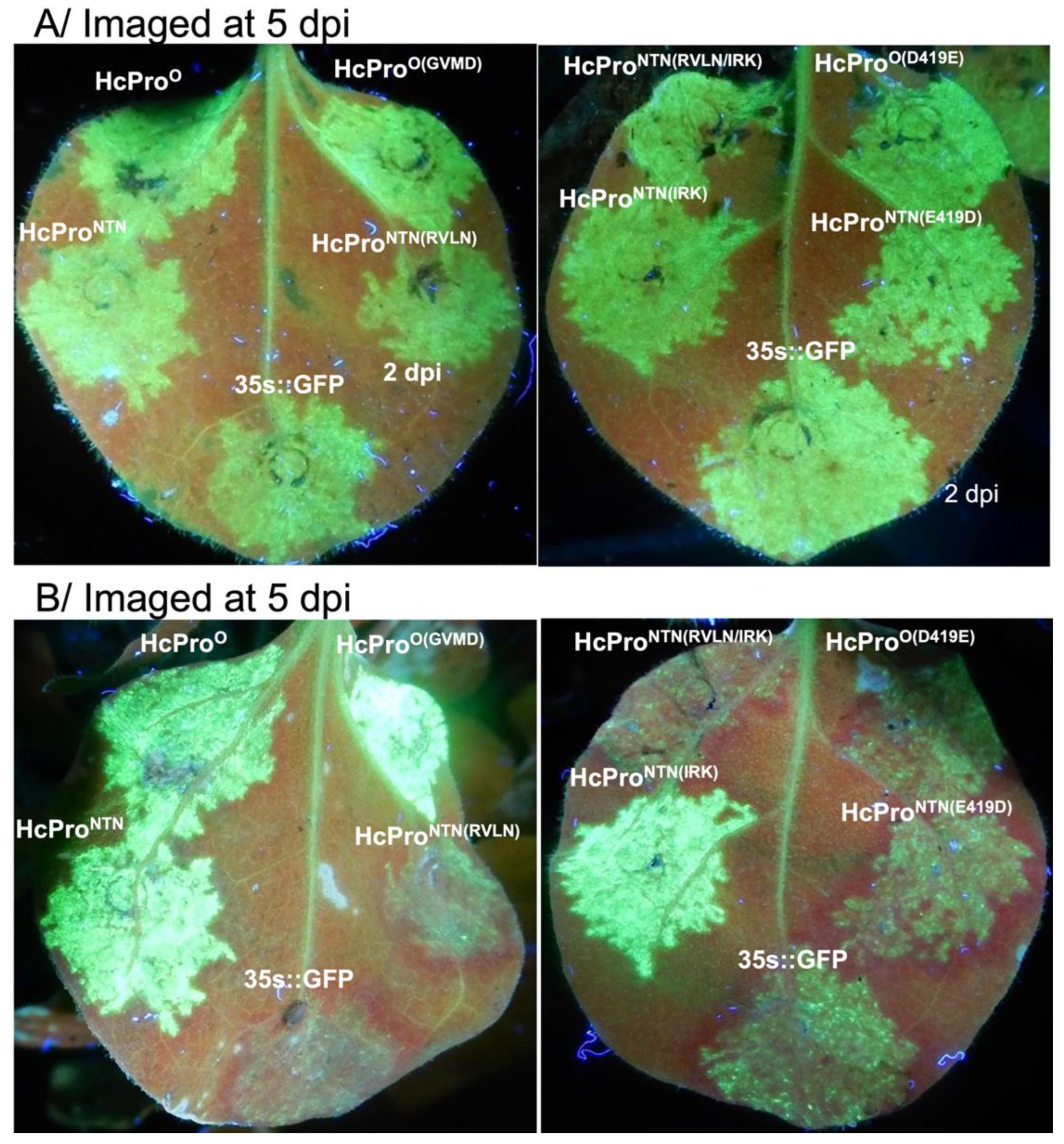
Figure 7.
Western blot analysis of GFP in infiltrated 16c leaves infiltrated with HCProNTN and HcProO, as well as constructs expressed from plasmid pEarleygate100. At 5 dpi, leaves co-infiltrated with GFP and HcProNTN, HcProO, HcProO(GVMD, HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProNTN(IRK), HcProO(D419E), HcProNTN(E419D, and pEarley100, respectively were analyzed using anti-GFP antibodies. By 12 dpi, GFP showed lower levels in HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProO(D419E), HcProNTN(E419D), and pEarley100. For the loading control, membranes were stripped of the GFP antibody and probed with heat shock complex 70 antibody (α-HSC70).
Figure 7.
Western blot analysis of GFP in infiltrated 16c leaves infiltrated with HCProNTN and HcProO, as well as constructs expressed from plasmid pEarleygate100. At 5 dpi, leaves co-infiltrated with GFP and HcProNTN, HcProO, HcProO(GVMD, HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProNTN(IRK), HcProO(D419E), HcProNTN(E419D, and pEarley100, respectively were analyzed using anti-GFP antibodies. By 12 dpi, GFP showed lower levels in HcProNTN(RVLN), HcProNTN(RVLN/IRK), HcProO(D419E), HcProNTN(E419D), and pEarley100. For the loading control, membranes were stripped of the GFP antibody and probed with heat shock complex 70 antibody (α-HSC70).
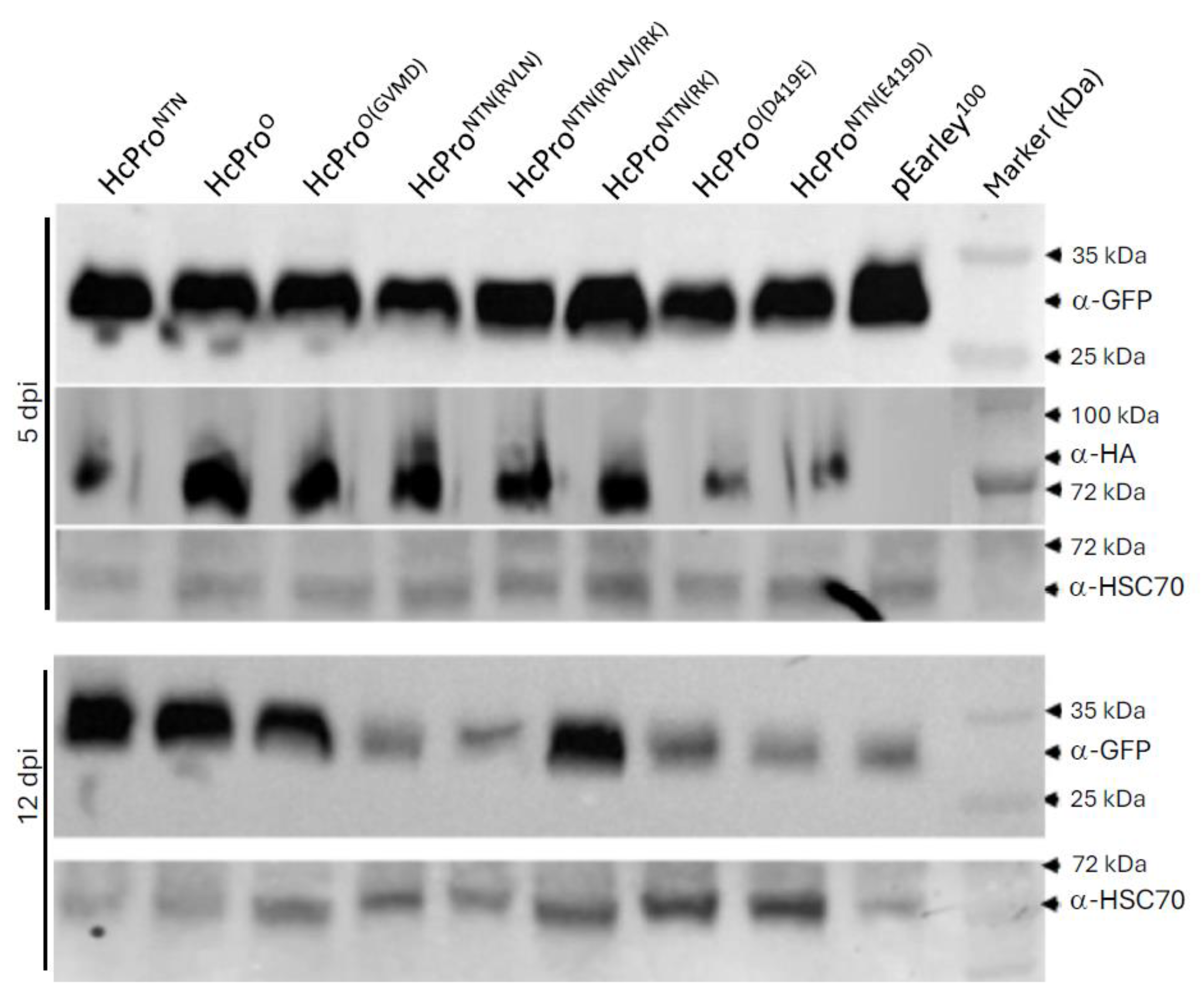

Table 1.
Primers used in this work.
| Designation | Sequence | Construct |
|---|---|---|
|
NTN.Hc-Pro.F NTN.Hc-Pro.R |
attB1+GGGGTTATGGATTCA | HcProNTN |
| attB2+ ACCAACTCTATAGTGCTTAATGTCAGACT | ||
|
O.HcPro.F O.HcPro-Pro.R |
attB1+CGTGTTTTGAACTCAA | HcProO |
| attB2+ACCAACTCTATAATGTTTTATATCAGATTCTAATTCATCATTTGC | ||
|
GVMD(1-4)RVLN-F GVMD(1-4)RVLN-R |
TTGAACTCAATGGTTCAGTTCTCAAGC | HcProNTN(RVLN) |
| AACACGGGCCATAAGGGCGAATTC | ||
|
RVLN(1-4)GVMD -F RVLN(1-4)GVMD-R |
ATGGATTCAATGATCCAGTTTTCGAATG | HcProO(GVMD |
| AACCCCGGCCATTAAGCCTGCTTT | ||
|
NRL(246-248)IRK-F NRL(246-248)IRK-R |
CAAGCATCCGAATGGAACAAGAAAAC | HcProNTN(IRK) |
| CGGATTTCATATGCTGAATAGCC | ||
| GVMD(1-4)RVLN-F | TTGAACTCAATGGTTCAGTTCTCAAGC | HcProNTN(RVLN/IRL)* |
| GVMD(1-4)RVLN-R | AACACGGGCCATAAGGGCGAATTC | |
|
OHcProGlu419-F OHcProGlu419-R |
TTGACCATGAAACTCAAACGTG | HcProO(D419E) |
| CCAATATTCTAGGCAGTTC | ||
|
NTNHcProAsp-F NTNHcProAsp-R |
TCGATCACGACACGCAGACAT | HcProNTN(E419D) |
| CTAGTATTCTAGGCAGTTCTG |
attB: site specific recombination; *HcProNTN(RVLN/IRL) was obtained by using HcProNTN(IRK) primers and HcProNTN(RVLN) template.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



