Submitted:
20 November 2025
Posted:
21 November 2025
You are already at the latest version
Abstract
The Strait of Messina (Central Mediterranean Sea) has always been known for the stranding of marine organisms, especially during the spring. We came across an extraordinary event of mass stranding in April 2025, with 453 specimens of Macroramphosus sp. found through a single night. A total of 571 post-larvae and juvenile individuals stranded between February and May 2025 were examined for this study. Clear morphological differences related to the size, especially in post-larvae were highlighted. The relationship between Body Length (BL) and other morphometric measurements, such as Dorsal Spine Length (DSL), Snout Length (SNL), and Body Height (BH) were studied with the aim of identifying without any doubt the species Macroramphosus gracilis. A slightly negative allometric relationship between length and weight shows that it grows faster in length and slower in weight. This study aims to improve the state of knowledge on slender snipefish M. gracilis, and particularly on the somatic features of different post-larval development stages. All these morphological changes could give us a hint at the ecological adaptation to the habit shift related to the development. Larvae and post-larvae of M. gracilis under 36 mm SL appear to be more linked to the epipelagic/hyponeustonic habitat, while adult individuals seem to prefer benthopelagic habits. Our results support the hypothesis that the Strait of Messina represents an exclusive and crucial zone for the reproduction and growth of M. gracilis.
Keywords:
Macroramphosus gracilis
; slender snipefish
; post-larvae/juveniles
; morphometrics
; development changes
; ecology
; Strait of Messina
; Mediterranean Sea
1. Introduction
Both sympatric species, Macroramphosus gracilis (Lowe, 1839) (Figure 1), commonly known as slender snipefish, and Macroramphosus scolopax (Linnaeus, 1758), known as long-spined snipefish, are broadly distributed throughout all the world’s oceans [1]. The two species differ mainly in body shape [2,3,4,5], growth patterns [2,6], feeding behaviours [1,2,3,5,7,8], and on the basis of morphometric data [1,5,6,9,10,11]. Much of the available information on presence, geographical distribution, and depth primarily refers to Macroramphosus scolopax, as reported in fish checklists for most Mediterranean areas [12,13,14,15,16,17,18]. A study on the biometry and distribution of M. scolopax was done in the Adriatic Sea [19] while the presence of larvae between the surface and 60 m was reported in the northern Ionian Sea and southern Tyrrhenian Sea [20,21]. Both species have no commercial value, since M. scolopax is frequently captured as bycatch in commercial fishing in the Tyrrhenian waters, ending up discarded [22]. No nutritional importance as human food is known for M. gracilis in Italian waters either.
Macroramphosus gracilis was for a long time mistaken for the juvenile stage of M. scolopax. So, its occurrence data are very scarce and refer mostly to the Eastern Mediterranean Sea, as Cyprus, Israel and Aegean Sea [23,24,25,26]. More recently, [27] confirms its presence in the Eastern Mediterranean Sea, reporting data on the biometry, taxonomy, distribution, biology and ecology.
Due to its hydrodynamic and biological characteristics, the Strait of Messina has been considered a peculiar upwelling ecosystem in the Central Mediterranean Sea. For this reason, August David Krohn (1803-1891), a zoologist of German origin, was the first to define the Strait of Messina as “the paradise of zoologists”, referring to its high biodiversity and the ease of finding meso- and bathypelagic fish stranded in excellent conditions, often still alive. The stranding phenomenon was studied for the first time by Anastasio Cocco (1829) from Messina, who also gave the name to many new species [28]. The first systematic list of stranded meso- and bathypelagic species is due to [29], followed by [30], who updated both the biodiversity, the places and periods of stranding, relating this phenomenon to the synergy between currents and winds speed-direction. Up to more recent times, updates and in-depth studies on the role of mesopelagic fish in the deep pelagic food web followed, both on larval and juvenile stages and adults of many species, also in relation to the lunar phases e.g. [31,32,33].
Available data on Macroramphosus in our study area are quite confusing, as is not listed by [29,30] and [34] among the deep-sea fishes stranded in the Strait of Messina. Then, for the first time, [34,35,36] reported massive strandings of Macroramphosus gracilis along the Messina coast of Faro and Ganzirri sites, even if the same authors considered the individuals as juvenile stages of M. scolopax. As mentioned earlier, this misidentification has been quite common in less recent papers, as [37]. More recent works, as [38], and in particular [39], reported both species stranded in the Strait of Messina between 2015 and 2016, with the macroscopic difference that only one specimen of M. scolopax was collected in October 2015 while a total of 375 specimens of M. gracilis stranded in 2016 during the months of February (3 specimens), March (30 specimens), April (137 specimens) and May (205 specimens).
This study aims to improve the state of knowledge on slender snipefish M. gracilis, and particularly on the somatic features of different post-larval development stages, contributing to the knowledge about this long-neglected fish.
2. Materials and Methods
2.1. Study Area
2.1.1. Physical Features
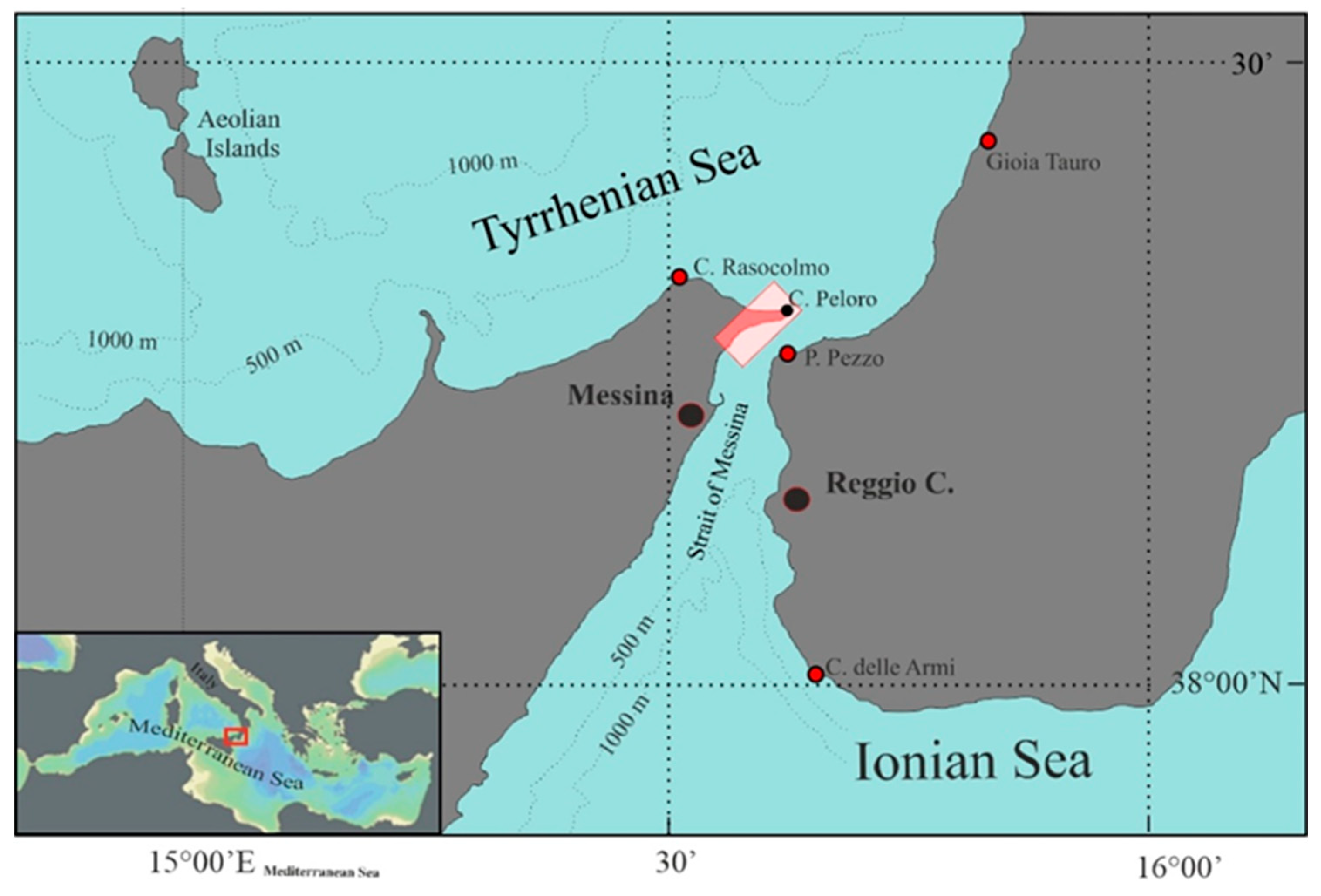
The Strait of Messina represents one of the most productive pelagic ecosystems in the Mediterranean Sea. Its unique characteristics are determined by geomorphological conformation of the seabed, the opposing tidal phases of the two neighboring seas, the northern Ionian and the southern Tyrrhenian, and the different physical and chemical characteristics of their water masses. Its funnel shape, with the widest and deepest part facing south towards the Ionian Sea (2000 m in front of Capo d'Armi) and the narrowest part (about 3 km) and less deep towards the northern Tyrrhenian entrance (about 75 m on the threshold that rises between Punta Pezzo-Calabria and Ganzirri-Sicily, Figure 2 and 3), determines complex and turbulent hydrodynamic processes [40,41,42,43,44], characterised by the formation of advective vortices and strong horizontal current cuts, generally located close to the most prominent capes (Capo Peloro, Sicily and Punta Pezzo, Calabria). The lunar phases influence the tidal currents, which can reach speeds of 300 cm s-1 during the new and full moon, speeds that can increase up to 500 cm s-1 in conjunction with other factors such as wind drift, meteorological waves, atmospheric pressure and turbulence [45]. Two stationary tidal currents alternate regularly every 6 hours (semi-diurnal), called "montante” (ascending current, Figure 2) which flows from the Ionian Sea to the Tyrrhenian Sea causing an upwelling of colder and saltier Ionian deep waters especially from the mesopelagic zone (200-1000 m) and "scendente” (descending current) which flows superficially from the Tyrrhenian Sea towards the Ionian Sea dragged by the surface cyclonic circulation [40,41,42,43,44,46]. The first is the most important from a biological point of view, as on the one hand it mixes the nutrient-rich Levantine Intermediate Waters (LIW) with the surface waters of the Atlantic, enriching the euphotic layer for the benefit of the entire food chain (Figure 2). On the other hand, depending on the prevailing wind, it determines the stranding of meso- and bathipelagic organisms both along the Sicilian coast of Messina (southern sirocco winds) and along the Calabrian coast (northern mistral winds).
Figure 3.
Details of the area where the stranded Macroramphosus gracilis specimens were collected.
2.1.2. Biological Features
The continuous supply of nutrients due to upwelling currents (Figure 2) makes the Strait of Messina a highly productive ecosystem, resulting in high biodiversity at all levels of the pelagic food chain, from micro-phytoplankton to zooplankton, up to micronekton (such as euphausiids, pelagic decapods and mesopelagic fish [47,48]. Maximum fluorescence values have been recorded during the spring season in the southern area of the Strait [49,50]. It has also been known for a long time [48] that in some particular areas of the Strait there are “accumulations” of zooplankton organisms, many of which, considered rare in the Mediterranean Sea, reach significant concentrations of individuals [47,52,53,54]. Upwelling currents also cause the regular stranding of mesopelagic and bathypelagic organisms [30,32,55]. Consequently, this upwelling system attracts predators from different trophic levels [47,56] constituting feeding areas for both top predators such as tuna (Thunnus thynnus), swordfish (Xiphias gladius), albacore (Thunnus alalunga), and for many marine mammals that cross the Strait of Messina during their migrations [57].
2.2. Samples Collection
A total of 571 Macroramphosus gracilis specimens stranded from the first half of February 2025 to the first half of May 2025 were collected from the Sicilian coast of the Strait of Messina (Figure 3). The samplings were always made before dawn on the beach of Capo Peloro to avoid the competition of seabirds, ants and wasps and the sun's drying effect. In Figure 4 it is shown the individual number of M. gracilis collected stranded in the four months from February to May, in relation to some variables such as the lunar phases and the direction and speed of the rising current (S-N). A progressive increase in stranded organisms is evident from February to May, with a maximum of 477 individuals collected in April with a current of 3.0-3.4 kn. In particular, we came across an extraordinary event of mass stranding when 453 specimens of Macroramphosus gracilis found through a single night, on April 3 2025, during waxing crescent lunar phase.
In Table 1, zooplankton, micronekton and other deep fish stranded together with M. gracilis were listed. A total of 110 teleosts were collected, 63 in March and 47 in April. Of these, 11 species belong to mesopelagic fishes, including juveniles and adults (4 species), juveniles (3 species), and adults (4 species). Adult Hygophum benoiti was the species with the highest number of stranded individuals in both March and April, followed by Argyropelecus hemigymnus, Cyclothone braueri, and Vinciguerria attenuata (only in March). In order of species abundance, amphipods follow with 4 species and 1genus (24 individuals) in March and 1 species and 1 genus (2 individuals) in April. 45 individuals of the euphausiids Thysanoessa gregaria and 1 of Euphausia krohni (only in March) were found. No other organisms were found stranded together to M. gracilis in May.
2.3. Data Analysis
According to [1,3,5,10], a primary laboratory sorting of the specimens based on five characters as colour and body shape was made (Table 2). This, coupled with an assessment of morphometric ratios (see below), allowed us to assign all collected specimens to M. gracilis. Each specimen was measured to the nearest 1 mm (Standard Length, SL) using a ruler for larger individuals and graph paper for smaller ones and weighed to the nearest 0.01 g (Wet Weight, WW). These data were then used for statistical analyses. In addition, further morphometric measurements in accordance with the methods outlined by [1,3,5,10] were taken. These measurements included SNL (Snout Length, also referred to as beak length, measured from the tip of the snout to the front edge of the eye orbit), DSL (Dorsal Spine Length, commonly known as Spine Length), BH (Body Height, the maximum depth of the body), and DSFD (Dorsal Spine Fin Distance, the distance between dorsal spine and second dorsal fin origin; see Figure 5 and Table 2).
Since the length of the beak changes with size, the Body Length (BL; Figure 6), obtained by subtracting the length of the beak (Snout Length, SNL) from the Standard Length (BL=SL-SNL), is a useful morphometric measure to study the effective growth of Macroramphosus spp. [3,5]. Therefore, BL was used in place of SL in the assessment of Length-Weight relationships. The latter were calculated by the function W=aLb [58] where W is the WW measured in g, L is BL measured in mm, and a and b constants. Instead, Length-Length relationships were calculated using the function y = ax + b, where x = BL, y = concerned, a = slope, and b = intercept. Using PAST 4.03 [59] software, obtained curve was used to interpolate weight values as a function of length.
In addition, percentage ratios between BL and other morphometric measures were used to determine without doubt the species Macroramphosus gracilis, according to the statement of [5] (see Table 2). The following ratios between the measurements were assessed:
SNL/BL = Snout Length/Body Length. Snout length should be indicative of the organism's overall growth, considering that it is almost absent during the larval stage and longer in the adult stage;
DSL/BL = Dorsal Spine Length (longer ray of the first dorsal fin)/ Body Length. This measurement was already taken by [3], although in the paper it is indicated for Macroramphosus sp. For the present study it was used this ratio on only M. gracilis;
BL/BH = Body Length /Body Height (maximum body depth). A standard measurement that was already taken by [10].
Furthermore, some morphological differences were observed between our specimens. On the smallest individuals, we observed the presence of a great number of spinules, which covered almost completely the upper part of the body from the cephalic region to the first dorsal fin and beyond (Figure 7).
3. Results
From visual observation of the 571 collected specimens, it was possible to notice a slender, hydrodynamic shape, a dorsally dark colouration with silver sides, and a straight ventral body profile (Figure 1 and Figure 8). In seven individuals, we observed soft shades of light red on the silver sides, slightly more evident in one of these (Figure 9). The BH/BL ratio was found in most cases to be between 20 and 30%, in accordance with what was stated by [5] for M. gracilis (Table 2), while in some of the specimens (65 out of 571) it was found to be higher, and closer to the congener M. scolopax. In 7 specimens, it was found to be lower than 20% (Table 3). The remaining morphometric ratios were found in the large majority of cases to correspond to M. gracilis [5], with very few exceptions, numerically rather small, for example, 4 individuals with the spine slightly longer in relation to the body length (DSL/BL ratio=33.3%, while the maximum for M. gracilis is 32.6% [5]).
We named the spinules sets as it follows: dorso-cephalic spinules (Figure 10a), supraorbital spinules (Figure 10b), supra-opercular spinules (Figure 10c), and lateral spinules (Figure 10d). Then we can establish 4 size classes: A, early post-larvae 9-27 mm SL (n=432); B, mid post-larvae 28-34 mm SL (n=31); C, late post-larvae 35-45 mm SL (n=75); D, early juvenile stage > 46 mm SL (n=33). We observed all the spinules sets on individuals up to 27 mm SL and could be intended as an early transition from larval to juvenile stage. The dorso-cephalic ones, well evident in the first size class had almost completely disappeared in the range 28-34 mm, and only a supraorbital serrated pair of spinules, two supra-opercular serrated pairs of spinules, three straight pairs of spinules on the lateral line remained. From each pair of spinules on the lateral line, a row of smaller spinules originates, and goes up on the dorsal side. On specimens from 35-45 mm SL, we observed the absence of the supraorbital set, but the other two sets were still present, with the only difference being their smaller size. Finally, for individuals bigger than 46 mm SL we observed no spinules at all, feature that recalls the adult form.
Figure 11 shows a slightly negative allometric relationship between length and weight, in which weight increases less rapidly than length. Data are fairly well distributed around the curve, indicating a excellent quality of model fit (R² = 0.9727). Some points out of trend (outliers), visible especially around BL ≈ 20 mm and BL ≈ 40 mm, which means that the remaining 2.73% of the variation can be due to other factors (biological variability, environmental conditions, etc.). The b value = 2.8536 indicates negative allometric growth, with 95% confidence intervals ranging from 2.786 to 2.905.
The relationship between BL and Snout Length (SNL) (Figure 12) showed an angular coefficient of 0.4703, with 95% confidence interval ranging from 0.46017 to 0.48012, suggesting a negative allometric growth, which means that the Snout Length grows slower that the Body Length. The good value of R² = 0.9466 and p-value < 0.0001 confirm the statistical robustness of the model, and suggests that 94.66% of variability in SNL is explained by BL.
The relationship between BL and Dorsal Spine Length (DSL) (Figure 13) showed an angular coefficient of 0.2267, with 95% confidence interval ranging from 0.21859 to 0.23528, suggesting a negative allometric growth that translates in Dorsal Spine Length growing slower than BL. The good value of R² = 0.9076 and p-value < 0.0001 suggests that 90.76% of variability in DSL is explained by BL.
The relationship between BL and Body Height (BH) (Figure 14) showed an angular coefficient of 0.318, with 95% confidence interval ranging from 0.30981 to 0.32581, which suggests a negative allometric growth where BH grows slower than BL. The good value of R² = 0.9258 and p-value < 0.0001 confirm the statistical robustness of the model, suggesting that 92.58% of variability in BH is explained by BL.
Since all these intervals showed a negative allometric growth it can be concluded that, proportionally, all the values reported as dependent variable (y axis) grow slower than the independent variable (BL). In other words, all the other morphological features remain smaller in proportion to the body length throughout all the individuals examined, even though some of the considered body parts seem to grow faster than the others, like SNL.
4. Discussion
The reported results allowed us to unequivocally assign all investigated specimens to the species Macroramphosus gracilis. Indeed, both visual observation and morphometric measurements and ratios match those reported by [1,5,8,9,10] for M. gracilis. In our opinion, some slight discrepancies may be due to possible intraspecific variability in the local population, which in turn could be due to the unique ecological characteristics of the study area [40,41,42,43,44,45,46,47,48,49,50,51,52,53,54]. Furthermore, a certain intraspecific morphological variability is also found in the congener M. scolopax [1,3,4,9,10], but this does not prevent us from clearly and unequivocally distinguishing between the two species [1,5,8,9,10].
All the plots showed in the result section display a high goodness of fit in the regression models. The length-weight relationship shows that this fish grows faster in length and slower in weight. This means that when they finally reach the adult stage, they should obtain a slenderer shape; this feature is totally reasonable given that the specimen examined in this paper were collected stranded by the upwelling strong current, typical of the Strait of Messina. In fact, this kind of shape is expectable from species that live in turbulent environments where they need to be agile to swim through strong currents. According to [8], M. gracilis may be a better swimmer than M. scolopax due to its slender and fusiform body. This feature may increase capture success in off-bottom waters, thereby reducing hydrodynamic drag [60]. This underlines that the major body shape difference between M. gracilis and M. scolopax could be due to the different environment for which they adapted: is in fact known that M. scolopax lives mostly as a demersal species, feeding on soft seabeds using the long beak-like mouth to strike the prey with high speed to catch it [61], while M. gracilis is a benthopelagic species and feeds mostly on prey like pelagic plankton [1,5,8]. This is probably what [1] and [11] stated in their papers even though they only identified the fishes from the genera Macroramphosus as type-b (benthic) and type-p (pelagic) feeding, with a low frequent intermediate feeding type. This difference is observed also in [5], where they give the name of B-type and D-type to fishes that feed in a similar way respectively to type-b and type-p from [1] and [11], and then finally [8] links the feeding type to the two species, confirming that the p-type corresponds to M. gracilis.
Moreover, [5] stated that larvae and post-larvae measuring 5.6-36.5 mm SL were still present at the surface in the month of April, meaning that they were probably still feeding on pelagic plankton before growing and descending to the continental slope. This could be a quite similar situation to the one presented hereby; in fact, all the specimens examined in this paper were under 52 mm SL and probably not even a year in age following the length-age table from [6]. The fact that only post-larvae and juveniles were stranded on the beach could mean that no adults were present in the Strait of Messina, as they live near soft seafloors even though capable of pelagic migrations, as seemingly confirmed by [5] in which paper they assert that some sand was found in some D-type stomach. Given the fact that on the Ionian side of the Strait the soft seafloor is scarcely present due to hydrodynamic conditions [62], the absence of adult individuals could be due to the lack of habitat, and therefore this does not affect the juvenile population as they only need the epipelagic zone in order to sustain themselves. So, we can hypothesize that the Strait of Messina could represent a nursery area and/or a feeding area for M. gracilis larvae and juveniles, given its high productivity [49,50], resulting in important trophic effects and a rapid short food chain [63] and, consequently, high concentrations of prey, making the Strait of Messina a zone of food accumulation and “insemination” of neighbouring areas [47,56,64]. Once adults, they may migrate southeastward to the eastern Mediterranean basin where M. gracilis has been repeatedly found [23,24,25,26,27], while its presence in the western Mediterranean basin is still uncertain. The high food availability in the Strait of Messina may cause the population of M. gracilis to present higher rates of growth in comparison to the one found in the Atlantic Ocean. Borges [6] stated that M. gracilis presents an isometric growth, while we found a slightly negative allometric growth meaning that, in the Strait, M. gracilis specimens are longer in relation to the weight. This could be a similar situation to the one described in [65], where individuals of the jellyfish Pelagia noctiluca that were feeding in this area grew at a much faster rate than the normal. For M. gracilis, this could translate into reaching the first maturity size faster than in other areas of the world. A similar scenario could be happening in the coastal waters of Portugal, where fishes of the genera Macroramphosus grow faster than the same species found in Marocco Atlantic waters because of them living in an environment subject to cold upwelling events that rise the productivity of the area [6]. Therefore, it is possible that the Strait of Messina represents an exclusive and crucial zone for the reproduction and the growth for M. gracilis in the eastern basin of the Mediterranean Sea due to its peculiar characteristics like the colder and highly productive waters [50]. Indeed, [6] stated that reproduction of Macroramphosus sp. occurs over a time span from late winter to early spring (January to March in the northeastern Atlantic Ocean) [2,6,66], which is consistent with our sampling of early post-larvae (n=432) from February to May. Furthermore, the seawater temperature in the Strait of Messina is significantly lower than in the rest of the Mediterranean Sea due to the upwelling dynamics [53], which is perfectly in line with the statement [2], who considers the temperature range of 16-24°C optimal for the survival of this species and consequently for its reproduction. This could be confirmed by our findings, plotting Body Length in function of the Dorsal Spine Length, the Snout Length and the Body Height; all the ratios presented a negative allometric growth, which means that the three lengths grew slower in relation to the BL, that in turn grows faster in relation to the weight. As stated earlier, all these morphological changes may be an ecological adaptation to benthopelagic habits in adult individuals of M. gracilis. A shorter dorsal spine, coupled with a shorter and hydrodynamic-shaped snout and a lower body height, reduces hydrodynamic drag. This may align with a better swimming ability useful for broader displacement through the water column, granting the adults access to both pelagic and benthic food resources [1,5,8,11,60], unlike what is stated for M. scolopax, which showed a significant preference for a benthic diet [1,5,8,22,67] and therefore needs a higher manoeuvrability near the seabed [68], and a longer snout for catching benthic prey [61]. Moreover, M. scolopax presents a bigger dorsal spike [5], which could highlight a predator avoiding strategy [69,70] less based on a fast escape, in contrast to M. gracilis [71]. This latter, in particular, could use schooling behaviour to protect themselves from predation [72,73], taking advantage by both their good swimming ability [1,5,60] and their colouration, dark dorsally with silver sides [5], which we were able to observe in this paper. This pigmentation in fact helps pelagic fish in the euphotic zone through countershading, camouflaging themselves when seen from below thanks to the pale belly. Conversely, when seen from above, thanks to the dark upper part, they appear less visible thanks to the darker background of deeper waters. Furthermore, they exploit the reflectivity to confuse the sight of predators through the already mentioned schooling behaviours [72,73]. On the contrary, the red-orange pigmentation of M. scolopax helps it to camouflage itself in the deeper water layers, close to the bottom, where it lives, since unlike M. gracilis, it inhabits the epipelagic/hyponeustonic zone only at the larval stage (below 12 mm SL) [5]. In fact, the red colour is one of those that fades first with increasing depth [74], as seen in red crustaceans [73]. However, we couldn’t determine the exact depth at which the stranded specimens collected could have been living, and we can only hypothesize that they were living in the upper euphotic zone and possibly in deeper areas for the larger individuals. The fact that this fish was found stranded more often with non-migrant mesopelagic species as Argyropelecus hemigymnus, Cyclothone braueri, Maurolicus muelleri, some amphipods species, and strong migrant species as Hygophum benoiti and Vinciguerria attenuata [31,75] could indicate that our postlarvae specimens of M. gracilis could occupy the upper epipelagic euphotic zone and instead the juveniles the mid euphotic zone, respectively. The lack of stranded adult Macroramphosus specimens together to mesopelagic fish, confirms that they are not present in this bathymetric range. To confirm this, more studies on the trophic ecology of this fish are required, with a particular focus on the juveniles and on the specimen stranded in the Strait of Messina. In fact, identifying prey items could reveal their feeding habits and therefore trace their movements.
From a fisheries perspective, as already mentioned, neither M. gracilis nor M. scolopax have any commercial or food importance for humans, at least in Italian waters [22]. However, M. scolopax is known to be part of the diet of several demersal fishes, as Zeus faber [76], Pagellus bogaraveo and P. acarne [77], Scyliorhinus canicula [78], and finally of the very commercially important European hake Merluccius merluccius [79]. Nevertheless, we think the lack of knowledge about this matter, as soon as the whole ecology of Macroramphosus gracilis could be due to that, in the past, this species was often confused with M. scolopax, as just stated in our study. This may be even more true in the case of dietary studies, considering that digestive processes can make prey recognition more complicated, increasing the probability of misidentification. Moreover, otoliths of both species of Macroramphosus are very similar to each other [80], and the identification could be uncertain. Further studies will be useful to shed light on this and other aspects of the ecology of M. gracilis, including potential assessment of threats and pollution sensitivity and its role in the trophic chain, also as a potential prey of commercial fishes.
Author Contributions
Conceptualization, A.G. (Andrea Geraci), A.S., R.M., D.D.P., L.G. and A.G. (Antonia Granata); Methodology, A.G. (Andrea Geraci), A.S. and Y.G.; Software, D.D.P. and P.C.; Validation, A.G. (Andrea Geraci), A.S., R.M., S.G. and A.G. (Antonia Granata); Formal Analysis, A.G. (Andrea Geraci), A.S. and D.D.P.; Investigation, A.G. (Andrea Geraci), A.S., Y.G., R.M., D.D.P., P.C., L.G., S.G., R.F. and A.G. (Antonia Granata); Data Curation, A.G. (Andrea Geraci), A.S. and L.G.; Writing – Original Draft Preparation, A.G. (Andrea Geraci), A.S., Y.G., R.M., D.D.P., P.C., L.G., S.G., R.F. and A.G. (Antonia Granata); Writing – Review & Editing, A.G. (Andrea Geraci), A.S., R.M., L.G. and A.G. (Antonia Granata); Supervision, A.G. (Antonia Granata).
Funding
No funds are currently available for this research.
Data Availability Statement
Data are contained within the article. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Matthiessen, B.; Fock, H.O.; von Westernhagen, H. Evidence for two sympatric species of snipefishes Macroramphosus spp. (Syngnathiformes, Centriscidae) on Great Meteor Seamount. Helgoland Marine Research 2003, 57, 63–72. [Google Scholar] [CrossRef]
- Brêthes, J.-C. Contribution à l'étude des populations de Macrorhamphosus scolopax (L., 1758) et Macrorhamphosus gracilis (Lowe, 1839) des côtes atlantiques marocaines. Bulletin de l'institut des pêches maritimes du Maroc 1979, 24, 1-69.
- Clarke, T.A. Diet and morphological variation in snipefishes, presently recognized as Macrorhamphosus scolopax, from southeast Australia: evidence for two sexually dimorphic species. Copeia 1984, 595–608. [Google Scholar] [CrossRef]
- Assis, C.A. On the systematics of Macrorhamphosus scolopax (Linnaeus, 1758) and Macrorhamphosus gracilis (Lowe, 1839). II - Multivariate morphometric analysis. Arquivos do Museu Bocage (new series) 1993, 22, 383–402. [Google Scholar]
- Miyazaki, E.; Sasaki, K.; Mitani, T.; Ishida, M.; Uehara, S. The occurrence of two species of Macroramphosus (Gasterosteiformes: Macroramphosidae) in Japan: morphological and ecological observations on larvae, juveniles, and adults. Ichthyological Research 2004, 51, 256–262. [Google Scholar] [CrossRef]
- Borges, L. Age and growth of the snipefish, Macrorhamphosus spp., in the Portuguese continental waters. Journal of the Marine Biological Association of the United Kingdom 2000, 80, 147–153. [Google Scholar] [CrossRef]
- Farah, I.H.; Kamal, M.; Mahfoud, B.; Najib, C. Snipefish (Macroramphosus spp.) abundance and trophic dynamics in response to upwelling regime in the atlantic region from cape blanc to cape boujdor (20°50 N to 26°00 N). International Journal of Professional Business Review 2024, 9(11):e05136. [CrossRef]
- Matthiessen, B.; Fock, H.O. A null model for the analysis of dietary overlap in Macroramphosus spp. at the Great Meteor Seamount (subtropical North-east Atlantic). Archive of Fishery and Marine Research 2004, 51, 294–304. [Google Scholar]
- Assis, C. On the systematics of Macrorhamphosus scolopax (Linnaeus, 1758) and Macrorhamphosus gracilis (Lowe, 1839). I. A preliminary biometrical approach. Boletim da Sociedade Portuguesa de Ciências Naturais (2nd Sér.) 1992, 25, 5–19. [Google Scholar]
- Lopes, M.; Murta, A.G.; Cabral, H.N. Discrimination of snipefish Macroramphosus species and boarfish Capros aper morphotypes through multivariate analysis of body shape. Helgoland Marine Research 2006, 60, 18–24. [Google Scholar] [CrossRef]
- Christiansen, B.; Martin, B.; Hirch, S. The benthopelagic fish fauna on the summit of Seine Seamount, NE Atlantic: Composition, population structure and diets. Deep Sea Research Part II: Topical Studies in Oceanography 2009, 56, 2705–2712. [Google Scholar] [CrossRef]
- D'onghia, G.; Tursi, A.; Maiorano, P.; Matarrese, A.; Panza, M. Demersal fish assemblages from the bathyal grounds of the Ionian Sea (middle-eastern Mediterranean). Italian Journal of Zoology 1998, 65, 287–292. [Google Scholar] [CrossRef]
- Gaertner, J.-C.; Bertrand, J.A.; Relini, G.; Papaconstantinou, C.; Mazouni, N.; De Sola, L.G; Durbec, J.-P.; Jukic-Peladic, S.; Souplet, A. Spatial pattern in species richness of demersal fish assemblages on the continental shelf of the northern Mediterranean Sea: a multiscale analysis. Marine Ecology Progress Series 2007, 341, 191–203. [Google Scholar] [CrossRef]
- Bilecenoğlu, M.; Kaya, M.; Cihangir, B.; Çiçek, E. An updated checklist of the marine fishes of Turkey. Turkish Journal of Zoology 2014, 38, 901–929. [Google Scholar] [CrossRef]
- Carpentieri, P.; Colloca, F.; Ardizzone, G. Day-night variations in the demersal nekton assemblage on the Mediterranean shelf-break. Estuarine, Coastal and Shelf Science 2005, 63, 577–588. [Google Scholar] [CrossRef]
- Guy-Haim, T.; Stern, N.; Sisma-Ventura, G. Trophic ecology of deep-sea megafauna in the ultra-oligotrophic Southeastern Mediterranean Sea. Frontiers in Marine Science 2022, 9, 857179. [Google Scholar] [CrossRef]
- Lauria, V.; Gristina, M.; Fiorentino, F.; Attrill, M.J.; Garofalo, G. Spatial management units as an ecosystem-based approach for managing bottom-towed fisheries in the Central Mediterranean Sea. Frontiers in Marine Science 2020, 7, 233. [Google Scholar] [CrossRef]
- Kovačić, M.; Lipej, L.; Dulčić, J.; Iglesias, S.P.; Goren, M. Evidence-based checklist of the Mediterranean Sea fishes. Zootaxa 2021, 4998, 1–115. [Google Scholar] [CrossRef] [PubMed]
- Zorica, B.; Vrgoč, N. Biometry and distribution of snipefish, Macroramphosus scolopax (Linnaeus, 1758), in the Adriatic Sea. Acta adriatica 2005, 46, 99–106. [Google Scholar]
- Granata, A.; Cubeta, A.; Minutoli, R.; Bergamasco, A.; Guglielmo, L. Distribution and abundance of fish larvae in the northern Ionian Sea (Eastern Mediterranean). Helgoland Marine Research 2011, 65, 381–398. [Google Scholar] [CrossRef]
- Giordano, D.; Profeta, A.; Busalacchi, B.; Minutoli, R.; Guglielmo, L.; Bergamasco, A.; Granata, A. Summer larval fish assemblages in the Southern Tyrrhenian Sea (Western Mediterranean Sea). Marine Ecology 2015, 36, 104–117. [Google Scholar] [CrossRef]
- Carpentieri, P.; Serpetti. N.; Colloca. F.; Criscoli. A.; Ardizzone. G. Food preferences and rhythms of feeding activity of two co-existing demersal fish, the longspine snipefish, Macroramphosus scolopax (Linnaeus, 1758), and the boarfish Capros aper (Linnaeus, 1758), on the Mediterranean deep shelf. Marine Ecology 2016, 37, 106-118.
- Ben-Tuvia, A. Revised list of the Mediterranean fishes of Israel. Israel Journal of Zoology 1971, 20, 1–39. [Google Scholar]
- Papaconstantinou, C.; Tsimenidis, N. Some uncommon fishes from the Aegean Sea. Cybium 1979, 3, 3–14. [Google Scholar]
- Demetropoulos, A.; Neocleous, D. The fishes and crustaceans of Cyprus. Fisheries Bulletin of the Ministry of Agriculture and Natural Resources of Cyprus 1969, 1, 3–21. [Google Scholar]
- Ben-Tuvia, A. Collection of fishes from Cyprus. The Bulletin of the Research Council of Israel 1962, 11, 132–145. [Google Scholar]
- Bilecenoglu, M. Status of the genus Macroramphosus (Syngnathiformes: Centriscidae) in the eastern Mediterranean Sea. Zootaxa 2006, 1273, 55–64. [Google Scholar] [CrossRef]
- Cocco, A. Su di alcuni nuovi pesci de’mari di Messina. Giornale di Scienze Lettere e Arti per La Sicilia, 1829, 7, 138–147. [Google Scholar]
- Mazzarelli, G. Gli animali abissali e le correnti sottomarine dello Stretto di Messina. Rivista Mensile di Pesca e Idrobiologia 1909, 11, 177–218. [Google Scholar]
- Genovese, S.; Guglielmo, L. Spiaggiamenti di fauna abissale nello Stretto di Messina. Atti della Società Peloritana di Scienze Fisiche Matematiche e Naturali 1971, 17, 331–370. [Google Scholar]
- Di Carlo, B.S.; Costanzo, G.; Fresi, E.; Guglielmo, L. Feeding ecology and stranding mechanisms in two lanternfishes, Hygophum benoiti and Myctophum punctatum. Marine Ecology Progress Series 1982, 9, 13–24. [Google Scholar] [CrossRef]
- Battaglia, P.; Ammendolia, G.; Cavallaro, M.; Consoli, P.; Esposito, V.; Malara, D.; Rao, I.; Romeo, T.; Andaloro, F. Influence of lunar phases, winds and seasonality on the stranding of mesopelagic fish in the Strait of Messina (Central Mediterranean Sea). Marine Ecology 2017, 38, e12459. [Google Scholar] [CrossRef]
- Battaglia, P.; Pedà, C.; Malara, D.; Milisenda, G.; MacKenzie, B.R.; Esposito, V.; Consoli, P.; Vicchio, T.M.; Stipa, M.G.; Pagano, L. Importance of the lunar cycle on mesopelagic foraging by Atlantic Bluefin Tuna in the upwelling area of the Strait of Messina (Central Mediterranean Sea). Animals 2022, 12, 2261. [Google Scholar] [CrossRef]
- Berdar, A.; Cavaliere, A.; Cavallaro, G.; Giuffre, G.; Potoschi, A. Lo studio degli organismi marini spiaggiati nello Stretto di Messina negli ultimi due secoli. Naturalista Siciliano 1983, 7, 3–17. [Google Scholar]
- Berdar, A.; Cavallaro, G.; Giuffre, G.; Potoschi, A. Contributo alla conoscenza dei pesci spiaggiati lungo il litorale siciliano dello Stretto di Messina. Memorie di Biologia Marina e di Oceanografia 1981, 7, 77–87. [Google Scholar]
- Berdar, A.; Berdar, N.; Costa, F. Diminuzione di ittiofauna meso e batipelagica spiaggiata nello Stretto di Messina. Memorie di Biologia Marina e Oceanografia 1988, 17, 43–60. [Google Scholar]
- Costa, F. I pesci del Mediterraneo: stadi larvali e giovanili; Grafo editor: Brescia, Italia, 1999; p. 285. [Google Scholar]
- Cavallaro, M.; Villari, A.; Ammendolia, G.; Spadola, F.; Bonfiglio, L.; Mangano, G.; Panzera, M. Le collezioni di faune ittiche mesopelagiche e malacologica “A. Villari” del Museo della Fauna di Messina. In Atti XXV Congresso Associazione Nazionale Musei Scientifici, Torino, Italia, 11-13 November 2015; Università degli Studi di Torino, 2015; p. 48.
- Cavallaro, M.; Ammendolia, G.; Rao, I.; Villari, A.; Battaglia, P. Variazioni pluriennali del fenomeno dello spiaggiamento di specie ittiche nello Stretto di Messina, con particolare attenzione alle specie mesopelagiche. Annales, Series Historia Naturalis 2021, 31, 69-78.
- Bignami, F.; Salusti, E. Tidal currents and transient phenomena in the Strait of Messina: A review. In The Physical Oceanography of Sea Straits; Pratt, L.J., Ed.; NATO ASI Series: Springer, Dordrecht, 1990; Volume 318, pp. 95–124. [Google Scholar]
- Mosetti, R. Optimal policies in a Bioeconomic model of eutrophication. Applied Mathematics and Computation 1988, 26, 257–265. [Google Scholar] [CrossRef]
- Hopkins, T.S.; Salusti, E.; Settimi, D. Tidal forcing of the water mass interface in the Strait of Messina. Journal of Geophysical Research: Oceans 1984, 89, 2013–2024. [Google Scholar] [CrossRef]
- Vercelli, F. Crociere per lo studio dei fenomeni nello Stretto di Messina; Office Grafiche C. Ferrari: Venezia, Italia, 1926.
- Vercelli, F. Il regime delle correnti e delle maree nello Stretto di Messina. Office Grafiche Ferrari: Venezia, Italia, 1925.
- Mosetti, F. Tidal and other currents in the Straits of Messina. In The Straits of Messina ecosystem, Proceedings of the Symposium, Messina, Italia, 4-16 Aprile 1991; Guglielmo, L., Manganaro, A., De Domenico, E., Eds.; Università degli Studi di Messina, Italia, 1995; pp. 15-29.
- Povero, P.; Hopkins, T.; Fabiano, M. Oxygen and nutrient observations in the southern Tyrrhenian Sea. Oceanologica acta 1990, 13, 299–305. [Google Scholar]
- Guglielmo, L.; Crescenti, N.; Costanzo, G.; Zagami, G. Zooplankton and micronekton communities in the Straits of Messina. In The Straits of Messina ecosystem, Proceedings of the Symposium, Messina, Italia, 4-16 Aprile 1991; Guglielmo, L., Manganaro, A., De Domenico, E., Eds.; Università degli Studi di Messina, Italia, 1995; pp. 247-270.
- Guglielmo, L.; Marabello, F.; Vanucci, S. The role of the mesopelagic fishes in the pelagic food web of the Straits of Messina. In The Straits of Messina ecosystem, Proceedings of the Symposium, Messina, Italia, 4-16 Aprile 1991; Guglielmo, L., Manganaro, A., De Domenico, E., Eds.; Università degli Studi di Messina, Italia, 1995; pp. 223-246.
- Decembrini, F.; Hopkins, T.; Azzaro, F. Variability and sustenance of the deep-chlorophyll maximum over a narrow shelf, Augusta Gulf (Sicily). Chemistry and Ecology 2004, 20, 231–247. [Google Scholar] [CrossRef]
- Azzaro, F.; Decembrini, F.; Raffa, F.; Crisafi, E. Seasonal variability of phytoplankton fluorescence in relation to the Straits of Messina (Sicily) tidal upwelling. Ocean Science 2007, 3, 451–460. [Google Scholar] [CrossRef]
- Currieri, G. Sulle cause meccanico biologiche della formazione degli accumuli di plancton. Bollettino della Società zoologica italiana I 1900, 4.
- Zagami, G.; Badalamenti, F.; Guglielmo, L.; Manganaro, A. Short-term variations of the zooplankton community near the Straits of Messina (North-eastern Sicily): relationships with the hydrodynamic regime. Estuarine, Coastal and Shelf Science 1996, 42, 667-681.
- Guglielmo, L. Distribuzione dei chetognati nell'area idrografica dello Stretto di Messina. Pubblicazioni della Stazione Zoologica di Napoli 1976, 40, 34–72. [Google Scholar]
- Sparla, M.; Guglielmo, L. Distribuzione del microzooplancton nello Stretto di Messina (estate 1990). In Atti X Congresso AIOL, Alassio, Italia, 4-6 Novembre 1992;1994, pp. 307-325.
- Battaglia, P.; Andaloro, F.; Esposito, V.; Granata, A.; Guglielmo, L.; Guglielmo, R.; Musolino, S.; Romeo, T.; Zagami, G. Diet and trophic ecology of the lanternfish Electrona risso (Cocco 1829) in the Strait of Messina (central Mediterranean Sea) and potential resource utilization from the Deep Scattering Layer (DSL). Journal of Marine Systems 2016, 159, 100–108. [Google Scholar] [CrossRef]
- Battaglia, P.; Andaloro, F.; Consoli, P.; Esposito, V.; Malara, D.; Musolino, S.; Pedà, C.; Romeo, T. Feeding habits of the Atlantic bluefin tuna, Thunnus thynnus (L. 1758), in the central Mediterranean Sea (Strait of Messina). Helgoland Marine Research 2013, 67, 97–107. [Google Scholar] [CrossRef]
- Granata, A.; Tringali, L.; Bruno, R.; Guglielmo, L.; Guglielmo, R.; Minutoli, R. Lo Stretto di Messina: via di migrazione per pesci e mammiferi marini. In Sviluppo sostenibile dei trasporti marittimi nel Mediterraneo; Pellegrino, F., Ed.; Edizioni Scientifiche Italiane (ESI): Napoli, Italia, 2013; pp. 691–755. [Google Scholar]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. Fisheries Research Board of Canada Bulletin 1975, 191, 1–382. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T. , Ryan P.D. Past: paleontological statistics software package for educaton and data anlysis. Palaeontologia electronica 2001, 4, 1. http://palaeo-electronica.org/2001_1/past/issue1_01.htm.
- Schluter, D. Adaptive radiation in sticklebacks: size, shape, and habitat use efficiency. Ecology 1993, 74, 699–709. [Google Scholar] [CrossRef]
- Longo, S.J.; Goodearly, T.; Wainwright, P.C. Extremely fast feeding strikes are powered by elastic recoil in a seahorse relative, the snipefish, Macroramphosus scolopax. Proceedings of the Royal Society B: Biological Sciences 2018, 285, 20181078. [Google Scholar] [CrossRef] [PubMed]
- Santoro, V.C.; Amore, E.; Cavallaro, L.; Cozzo, G.; Foti, E. Sand waves in the Messina strait, Italy. Journal of Coastal Research 2002, 36, 640–653. [Google Scholar] [CrossRef]
- Fortier, L. Export production and the production of fish larvae and their prey at hydrodynamic singularities. In The Straits of Messina ecosystem, Proceedings of the Symposium, Messina, Italia, 4-16 Aprile 1991; Guglielmo, L., Manganaro, A., De Domenico, E., Eds.; Università degli Studi di Messina, Italia, 1995; pp. 213-225.
- Brancato, G.; Minutoli, R.; Granata, A.; Sidoti, O.; Guglielmo, L. Diversity and vertical migration of euphausiids across the Straits of Messina area. In Mediterranean ecosystems: structures and processes; Faranda, F.M., Guglielmo, L., Spezie, G., Eds.; Springer: Milano, Italia, 2001; pp. 131–141. [Google Scholar]
- Bergamasco, A.; Cucco, A.; Guglielmo, L.; Minutoli, R.; Quattrocchi, G.; Guglielmo, R.; Palumbo, F.; Pansera, M.; Zagami, G.; Vodopivec, M. Observing and modeling long-term persistence of P. noctiluca in coupled complementary marine systems (Southern Tyrrhenian Sea and Messina Strait). Scientific reports 2022, 12, 14905. [Google Scholar] [CrossRef]
- Arruda, L.; Andrade, J.; Cunha, M. Abundance, diversity, and community structure of the fish population in the Ria de Aveiro (Portugal). Oceanologica Acta 1988, 11, 235–240. [Google Scholar]
- Matallanas, J. Aspectos generales del regimen alimentario de Macroramphosus scolopax (Linnaeus 1758) (Pisces, Macroramphosidae) en las costas catalanas (Mediterrâneo occidental). Cahiers De Biologie Marine 1982, 23, 243–252. [Google Scholar]
- Motta, P.J.; Clifton, K.B.; Hernandez, P.; Eggold, B.T. Ecomorphological correlates in ten species of subtropical seagrass fishes: diet and microhabitat utilization. Environmental biology of fishes 1995, 44, 37–60. [Google Scholar] [CrossRef]
- Hoogland, R.; Morris, D.; Tinbergen, N. The spines of sticklebacks (Gasterosteus and Pygosteus) as means of defence against predators (Perca and Esox). Behaviour 1956, 10, 205–236. [Google Scholar] [CrossRef]
- Bosher, B.T.; Newton, S.H.; Fine, M.L. The spines of the channel catfish, Ictalurus punctatus, as an anti-predator adaptation: an experimental study. Ethology 2006, 112, 188–195. [Google Scholar] [CrossRef]
- Domenici, P. Escape responses in fish: kinematics, performance and behavior. In Fish locomotion: An eco-ethological perspective; Domenici, P., Ed.; CRC Press: Boca Raton, USA, 2010; pp. 123–170. [Google Scholar]
- Magurran, A.E. The adaptive significance of schooling as an anti-predator defence in fish. Annales Zoologici Fennici, 1990, 27, 51–66. [Google Scholar]
- McFall-Ngai, M.J. Crypsis in the pelagic environment. American Zoologist 1990, 30, 175–188. [Google Scholar] [CrossRef]
- Marshall, J. Vision and lack of vision in the ocean. Current biology 2017, 27, R494–R502. [Google Scholar] [CrossRef] [PubMed]
- Granata, A.; Bergamasco, A.; Battaglia, P.; Milisenda, G.; Pansera, M.; Bonanzinga, V.; Arena, G.; Andaloro, F.; Giacobbe, S.; Greco, S. Vertical distribution and diel migration of zooplankton and micronekton in Polcevera submarine canyon of the Ligurian mesopelagic zone (NW Mediterranean Sea). Progress in Oceanography 2020, 183, 102298. [Google Scholar] [CrossRef]
- Silva, A. Feeding habits of John Dory, Zeus faber, off the Portuguese continental coast. Journal of the Marine Biological Association of the United Kingdom 1999, 79, 333–340. [Google Scholar] [CrossRef]
- Morato, T.; Solà, E.; Grós, M.P.; Menezes, G. Feeding habits of two congener species of seabreams, Pagellus bogaraveo and Pagellus acarne, off the Azores (northeastern Atlantic) during spring of 1996 and 1997. Bulletin of marine science 2001, 69, 1073–1087. [Google Scholar]
- Gravino, F.; Dimech, M.; Schembri, P.J. Feeding habits of the small-spotted catshark Scyliorhinus canicula (L., 1758) in the central Mediterranean. Rapport Commission International de la Mer Mediterranee 2010, 39, 538. [Google Scholar]
- D’Iglio, C.; Porcino, N.; Savoca, S.; Profeta, A.; Perdichizzi, A.; Armeli Minicante, E.; Salvati, D.; Soraci, F.; Rinelli, P.; Giordano, D. Ontogenetic shift and feeding habits of the European hake (Merluccius merluccius L., 1758) in Central and Southern Tyrrhenian Sea (Western Mediterranean Sea): A comparison between past and present data. Ecology and Evolution 2022, 12, e8634. [Google Scholar] [CrossRef] [PubMed]
- Tuset, V.M.; Lombarte, A.; Assis, C.A. Otolith atlas for the western Mediterranean, north and central eastern Atlantic. Scientia Marina 2008, 72, 7–198. [Google Scholar] [CrossRef]
Figure 1.
Seven specimens of Macroramphosus gracilis (Lowe, 1839) from our collection (photo by Andrea Geraci).
Figure 1.
Seven specimens of Macroramphosus gracilis (Lowe, 1839) from our collection (photo by Andrea Geraci).

Figure 2.
Section of the Strait of Messina with upwelling current (S-N).
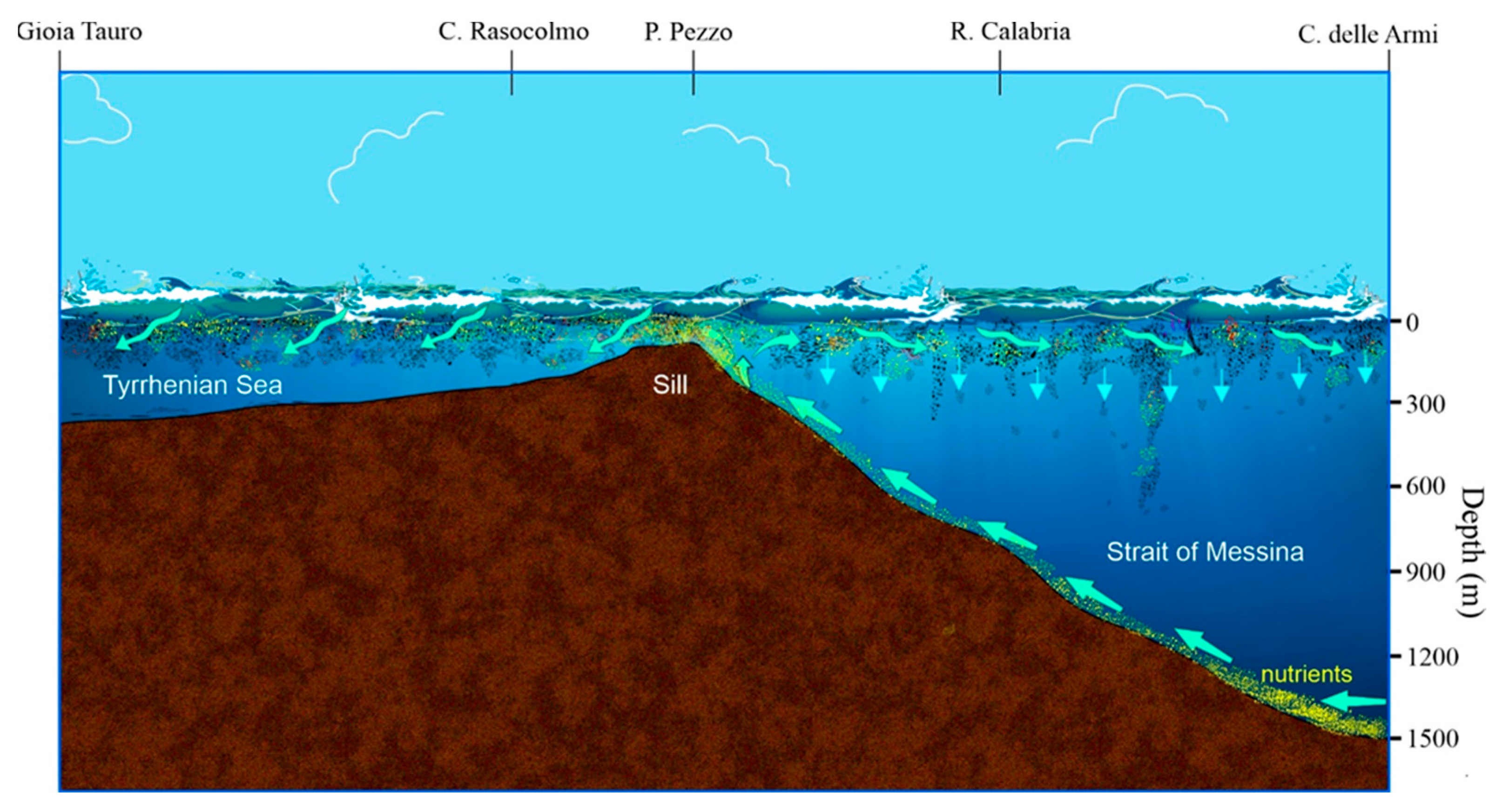
Figure 4.
Number of Macroramphosus gracilis stranded in relation to the months. For each month, upwelling current speed (S-N) and linked lunar phase (under x axis) for each sampling event are shown.
Figure 4.
Number of Macroramphosus gracilis stranded in relation to the months. For each month, upwelling current speed (S-N) and linked lunar phase (under x axis) for each sampling event are shown.
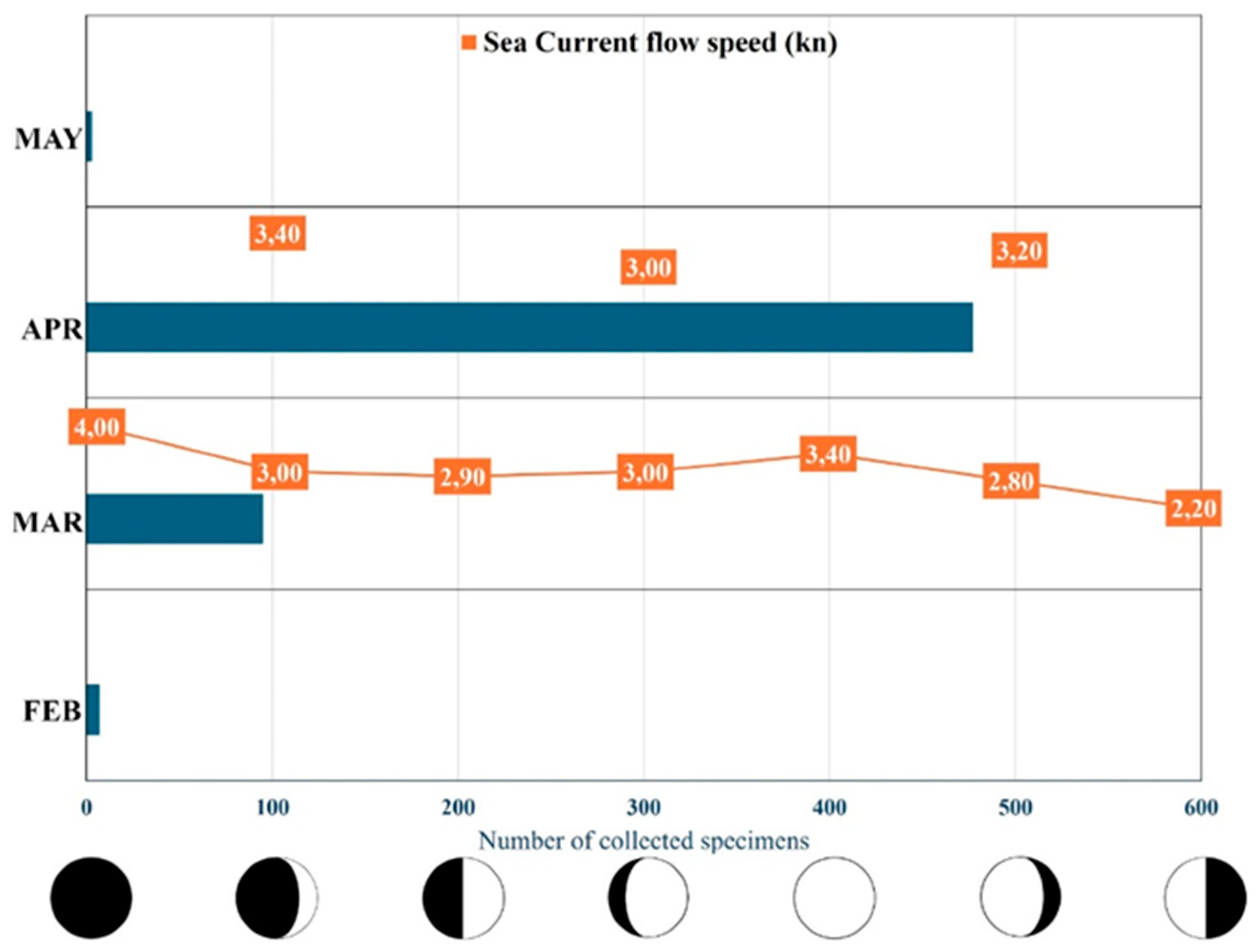
Figure 5.
Schematic drawing of Macroramphosus gracilis (modified from [10]) with highlighted points of morphometric measurements, as pointed out by [3,5] (see also following table).
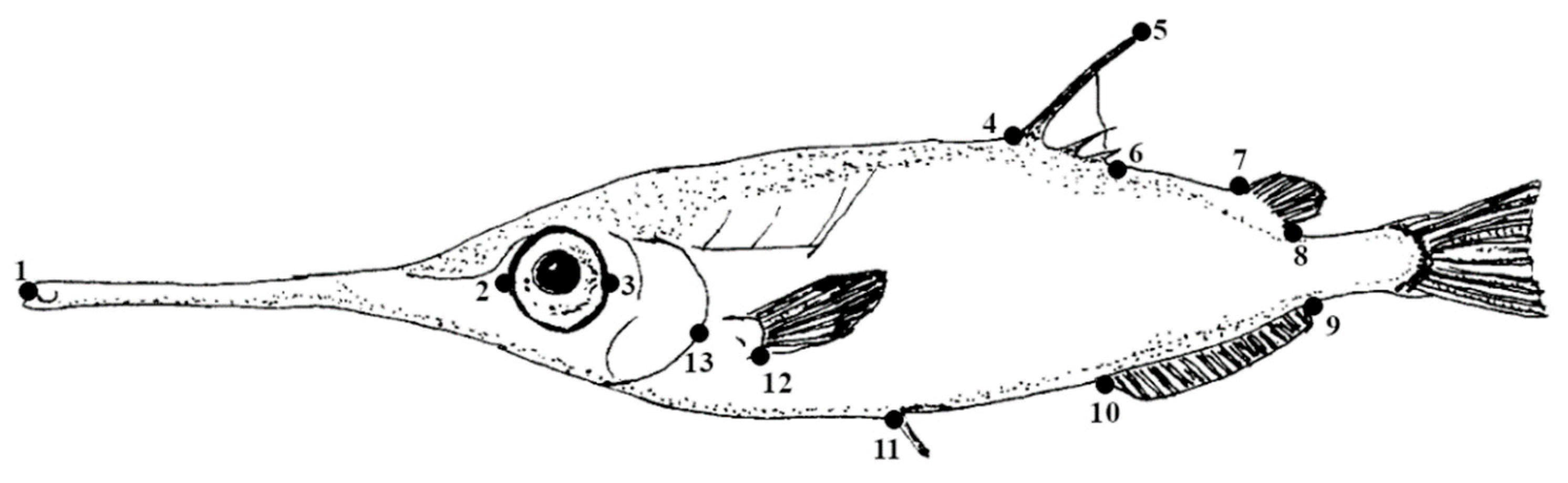
Figure 6.
Picture of one specimen of Macroramphosus gracilis from our collection, with highlighted SNL and BL (photo by Andrea Geraci).
Figure 6.
Picture of one specimen of Macroramphosus gracilis from our collection, with highlighted SNL and BL (photo by Andrea Geraci).

Figure 7.
Schematic drawing of the position of the spinule series on Macroramphosus gracilis. In yellow, the dorso-cephalic series; in red, the supra-orbital series; in blue, the supra-opercular series; in turquoise, the lateral series (modified from [10]).
Figure 7.
Schematic drawing of the position of the spinule series on Macroramphosus gracilis. In yellow, the dorso-cephalic series; in red, the supra-orbital series; in blue, the supra-opercular series; in turquoise, the lateral series (modified from [10]).
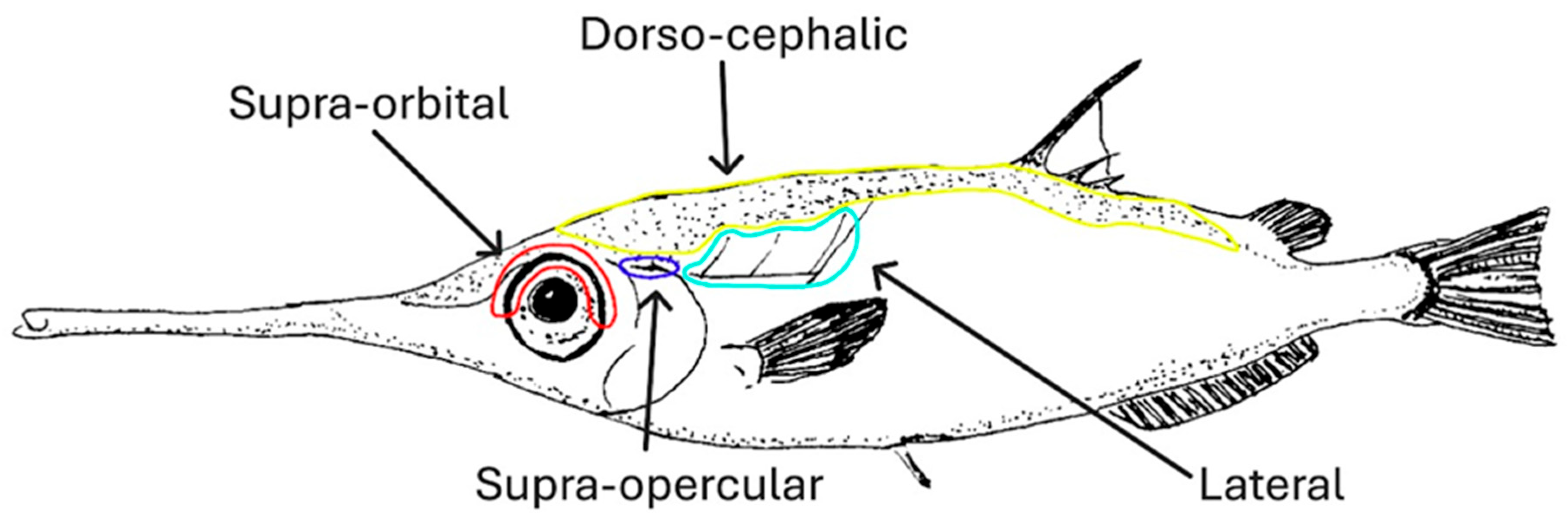
Figure 8.
Some collected specimens of Macroramphosus gracilis of various sizes (photo by Andrea Geraci).
Figure 8.
Some collected specimens of Macroramphosus gracilis of various sizes (photo by Andrea Geraci).
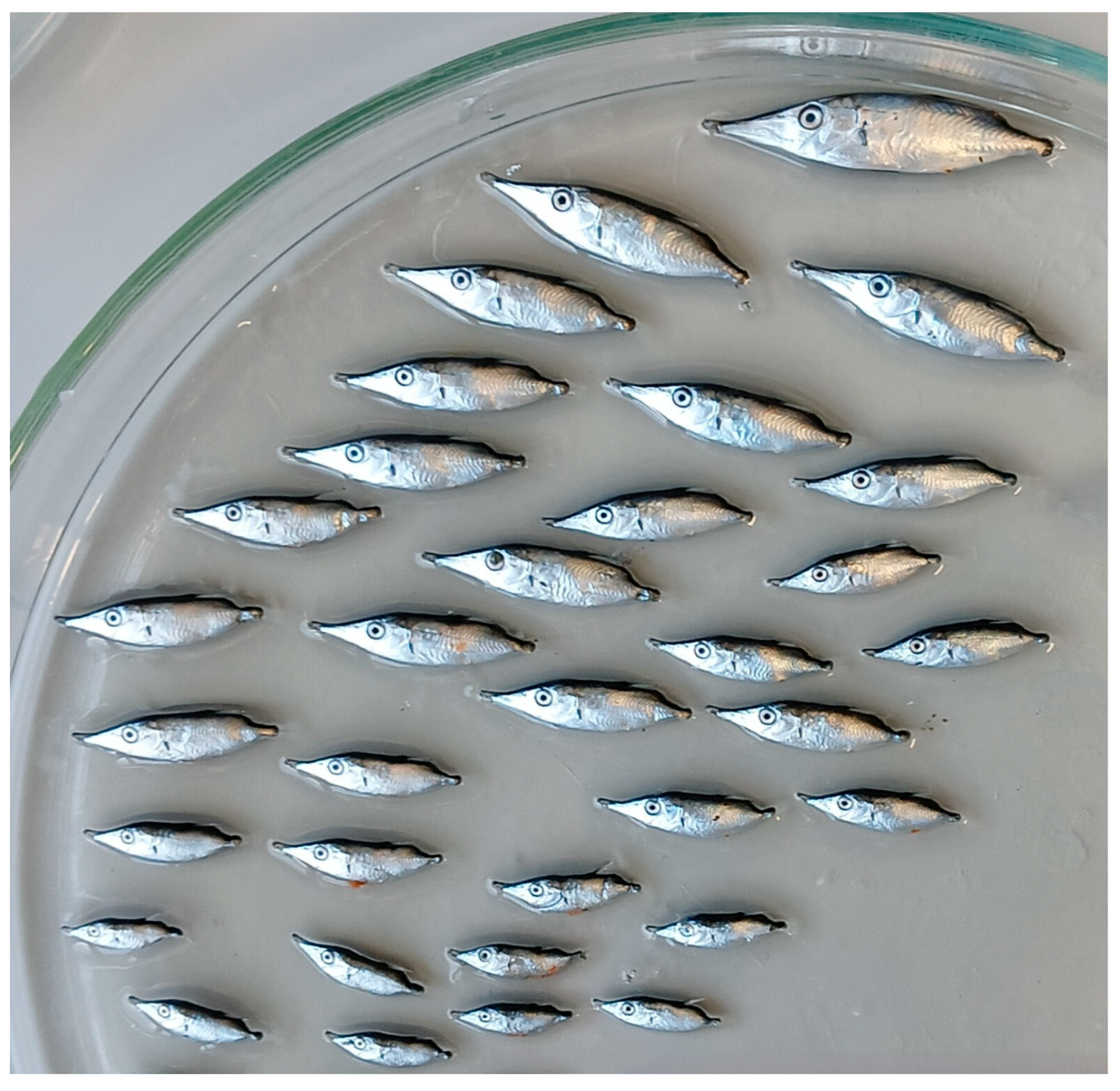
Figure 9.
One of the seven specimens that showed light red shades (above), compared to a standard-colored individual, dark dorsally with silver sides (below) (photo by Andrea Geraci).
Figure 9.
One of the seven specimens that showed light red shades (above), compared to a standard-colored individual, dark dorsally with silver sides (below) (photo by Andrea Geraci).

Figure 10.
a. Pictures of the dorso-cephalic spinules series. The pictures go from the first size class A to the fourth size class D (photo by Davide Di Paola). b. Pictures of the supraorbital spinules series (photo by Davide Di Paola). c. Pictures of the supra-opercular spinules series (photo by Davide Di Paola). d. Pictures of the lateral spinules series. Note the small rows of spinules departing from the bigger ones on the side (photo by Davide Di Paola).
Figure 10.
a. Pictures of the dorso-cephalic spinules series. The pictures go from the first size class A to the fourth size class D (photo by Davide Di Paola). b. Pictures of the supraorbital spinules series (photo by Davide Di Paola). c. Pictures of the supra-opercular spinules series (photo by Davide Di Paola). d. Pictures of the lateral spinules series. Note the small rows of spinules departing from the bigger ones on the side (photo by Davide Di Paola).
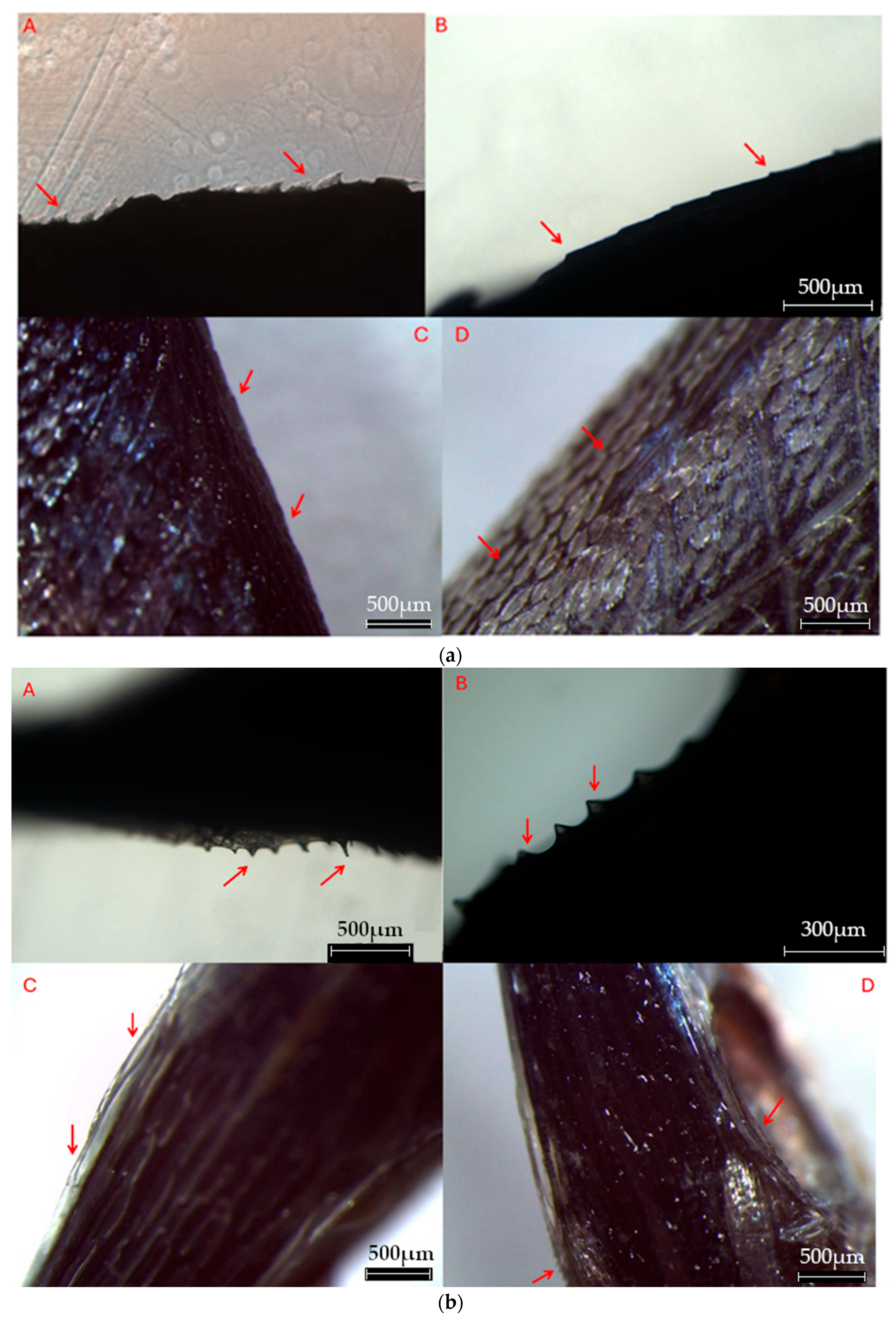

Figure 11.
Plots of length-weight relationships for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based non-linear regression analysis (p-value <0.0001).
Figure 11.
Plots of length-weight relationships for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based non-linear regression analysis (p-value <0.0001).
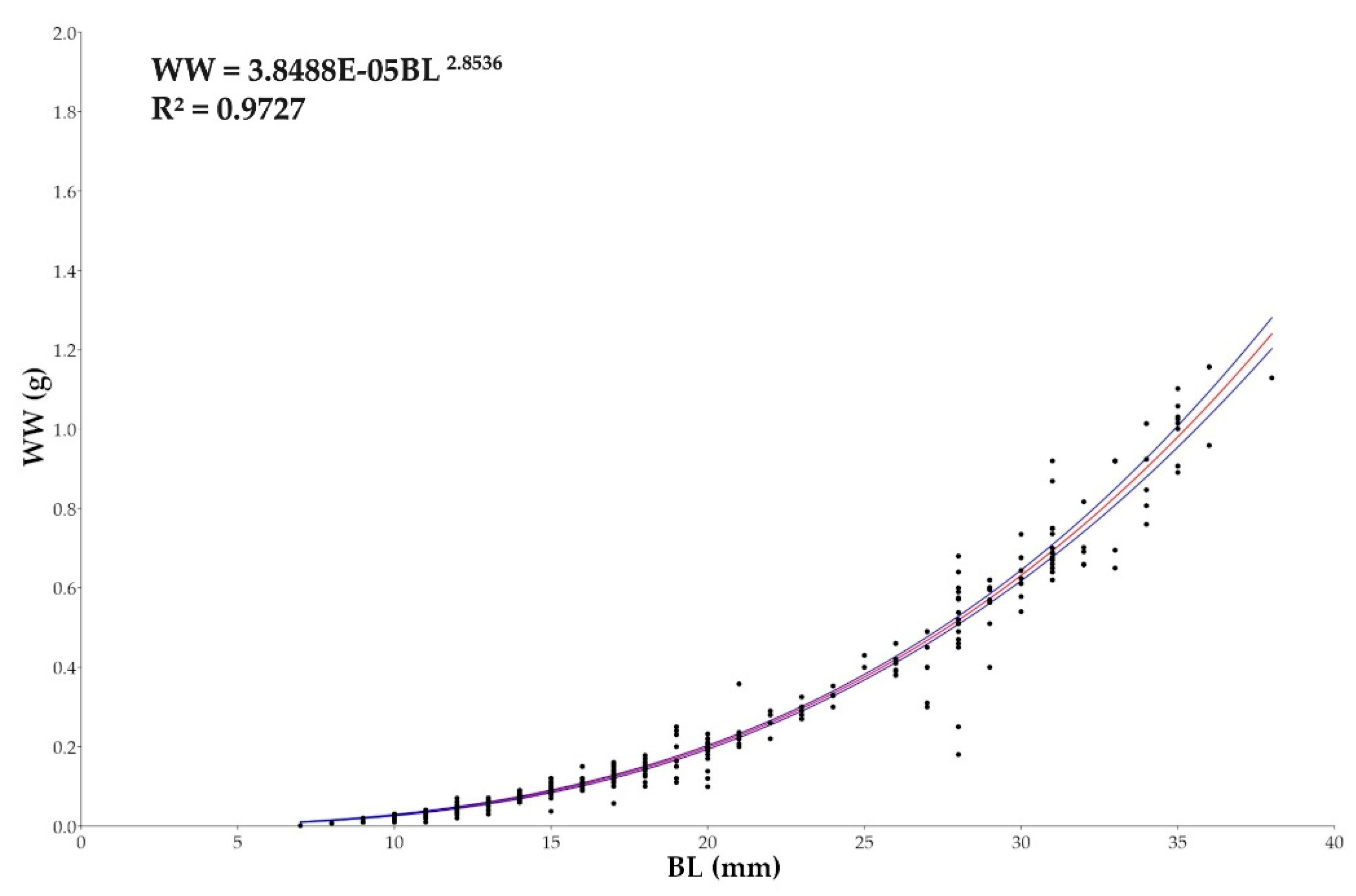
Figure 12.
Plots of Length (BL, in mm) and Snout Length (SNL, in mm) relationship for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based linear regression analysis (p-value <0.0001).
Figure 12.
Plots of Length (BL, in mm) and Snout Length (SNL, in mm) relationship for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based linear regression analysis (p-value <0.0001).
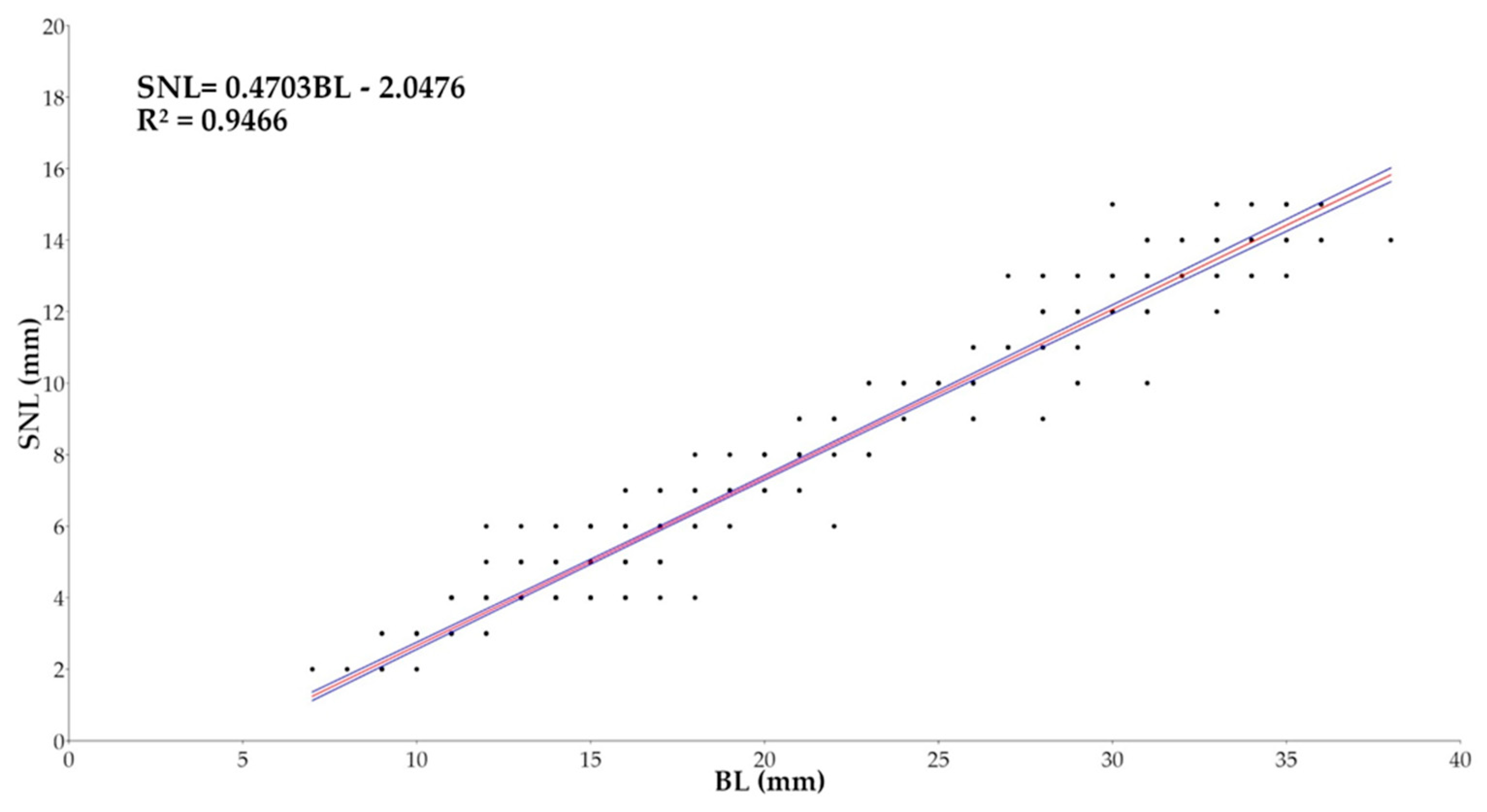
Figure 13.
Plots of Length (BL, in mm) and Dorsal Spine Length (DSL, in mm) relationship for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based linear regression analysis (p-value <0.0001).
Figure 13.
Plots of Length (BL, in mm) and Dorsal Spine Length (DSL, in mm) relationship for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based linear regression analysis (p-value <0.0001).
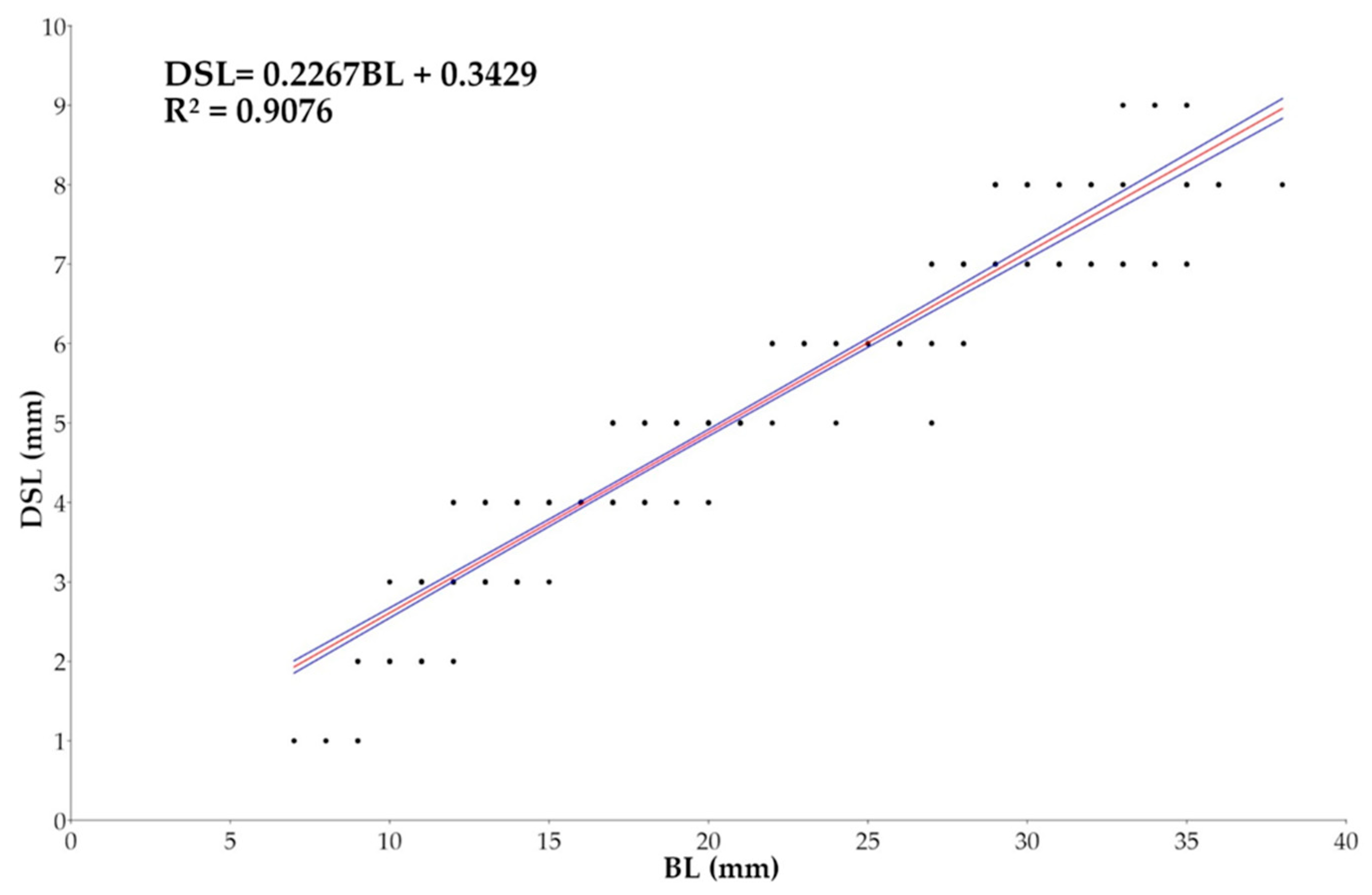
Figure 14.
Plots of Length (BL, in mm) and Body Height (BH, in mm) relationship for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based linear regression analysis (p-value <0.0001).
Figure 14.
Plots of Length (BL, in mm) and Body Height (BH, in mm) relationship for Macroramphosus gracilis post-larvae and juvenile (n= 571) as part of a standard curve-based linear regression analysis (p-value <0.0001).
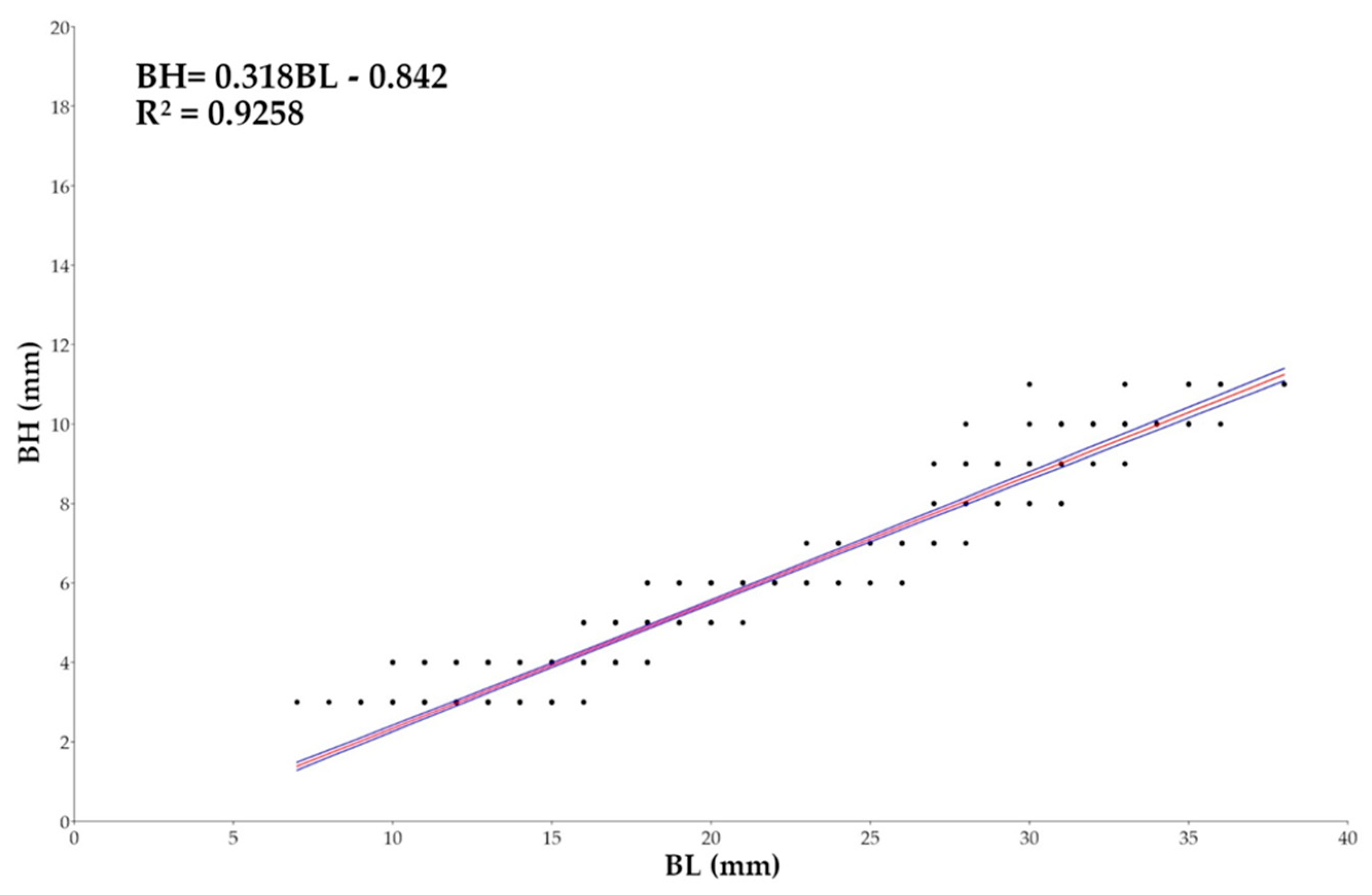

Table 1.
Number of individuals of the species collected stranded together to Macroramphosus gracilis.
Table 1.
Number of individuals of the species collected stranded together to Macroramphosus gracilis.
| Species | February | March | April | |
|---|---|---|---|---|
| Teleosts | Argyropelecus hemigymnus | -- | 11 | -- |
| Argyropelecus hemigymnus juv. | -- | 8 | -- | |
| Conger sp. juv. | -- | 1 | -- | |
| Cyclothone braueri | -- | 6 | -- | |
| Diaphus metopoclampus juv. | -- | 1 | -- | |
| Electrona risso juv. | -- | 2 | -- | |
| Engraulis encrasicolus | -- | 1 | -- | |
| Hygophum benoiti | -- | 25 | 36 | |
| Hygophum hygomii juv. | -- | -- | 2 | |
| Maurolicus muelleri | -- | -- | 1 | |
| Myctophum punctatum | -- | -- | 1 | |
| Microstoma microstoma | -- | 1 | 1 | |
| Microstoma microstoma juv. | -- | -- | 4 | |
| Nansenia oblita | -- | -- | 1 | |
| Nansenia oblita juv. | -- | 1 | -- | |
| Vinciguerria attenuata | -- | 6 | -- | |
| Vinciguerria attenuata juv. | -- | -- | 1 | |
| Amphipods | Lestrigonus schizogeneios | -- | 6 | -- |
| Phronima atlantica | -- | 4 | 1 | |
| Phronima sp. | -- | 11 | 1 | |
| Phrosina semilunata | -- | 2 | -- | |
| Scina crassicornis | -- | 1 | -- | |
| Euphausiids | Euphausia krohni | -- | 1 | -- |
| Thysanoessa gregaria | -- | 45 | -- | |
| Mysids | Siriella sp. | -- | -- | 1 |
| Pteropods | Cymbulia peronii | 1 | -- | 3 |
Table 2.
Morphometric measurement of Macroramphosus spp. [5] (see previous figure).
Table 2.
Morphometric measurement of Macroramphosus spp. [5] (see previous figure).
| M. gracilis | M. scolopax | |
|---|---|---|
|
Ventral body profile in larval and juvenile |
Straight | Notched |
| Body colour | Dark dorsally, with silver sides |
Red-orange with few melanophores dorsally |
| Posterior margin of dorsal fin spine (spike) in specimens larger than 50 mm SL | Smooth | Serrated with spinules |
| Lenght of dorsal fin spine (DSL, Dorsal Spine Length; 4-5) | Relatively short: 17.9-32.6% BL. 62.4-138% distance between dorsal spine-second dorsal fin origin (DSFD, Dorsal Spine Fin Distance; 4-7). |
Long: 23.7-46.2% BL. 98.9-231% distance between dorsal spine-second dorsal fin origin (DSFD, Dorsal Spine Fin Distance; 4-7). |
| Body Height (BH; 4-10) | Relatively slender: 20.9-30.8% BL. | Relatively deep: 23.4-36.7% BL. |
Table 3.
Morphological features and morphometric ratios observed in the examined specimens, compared with the statements of [5]. The "x" symbol indicates the presence of the feature, while the "/" symbol indicates its absence.
Table 3.
Morphological features and morphometric ratios observed in the examined specimens, compared with the statements of [5]. The "x" symbol indicates the presence of the feature, while the "/" symbol indicates its absence.
| Macroramphosus gracilis features | Macroramphosus scolopax features | Outlier features | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N. | Straight ventral body profile | Silver body, dor-sally dark | DSL/ BL ratio 17.9-32.6% |
DSL/ DSFD ratio 62.4-138% |
BH/BL ratio 20.9- 30.8% |
Notched ventral body profile | Red- orange body | DSL/ BL ratio 23.7-46.2% |
DSL/ DSFD ratio 98.9-231% |
BH/BL ratio 23.4- 36.7% |
Silver body, light red shaded, dorsally dark | DSL/ DSFD ratio <62.4% |
BH/ BL ratio >36.7% |
|
| 490 | x | x | x | x | x | / | / | / | / | / | / | / | / | |
| 61 | x | x | x | x | / | / | / | / | / | x | / | / | / | |
| 5 | x | x | x | x | / | / | / | / | / | / | / | / | x | |
| 2 | x | x | x | / | / | / | / | / | / | / | / | x | x | |
| 4 | x | / | x | x | x | / | / | / | / | / | x | / | / | |
| 3 | x | / | x | x | / | / | / | / | / | x | x | / | / | |
| 2 | x | x | x | / | x | / | / | / | / | / | / | x | / | |
| 1 | x | x | / | x | / | / | / | x | / | x | / | / | / | |
| 3 | x | x | / | x | x | / | / | x | / | / | / | / | / | |
| Tot | 571 | 571 | 564 | 567 | 567 | 499 | 0 | 0 | 4 | 0 | 65 | 7 | 4 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



