
Submitted:
11 November 2025
Posted:
12 November 2025
You are already at the latest version
Abstract
The chromatin remodelling proteins DAXX and ATRX are key regulators of ge-nome stability and epigenetic processes. Alterations in their expression have been as-sociated with tumour stage and prognostic outcomes in various humane cancer types, whereas their role in veterinary oncology has received little investigation to date. We analysed canine prostate and urinary bladder samples, including 18 prostate carcino-mas (12 adenocarcinomas, 6 prostatic urothelial carcinomas), 10 non-malignant pros-tate tissues, 22 carcinomas of the urinary bladder and 6 non-malignant bladder tissues. Nuclear expression of DAXX and ATRX was assessed using fully quantitative digital immunohistochemistry. Overall, DAXX exhibited consistently higher expression than ATRX across benign and malignant samples from the prostate and bladder. DAXX and ATRX expression demonstrated a positive correlation across all prostate (ρ = 0.692, p < 0.05) and bladder (ρ = 0.488, p < 0.05) samples, suggesting coordinated regulation. They displayed organ-specific expression patterns: prostate carcinomas showed increased expression of both proteins compared with non-malignant prostate tissues. In contrast, in the bladder, expression declined with increasing tumour aggressiveness (p < 0.05).
Our findings provide new insights into the potential of DAXX and ATRX as bi-omarkers, offering new insights into their clinical relevance in dogs.
Keywords:
dog
; molecular profiling
; DAXX
; ATRX
; immunohistochemistry
; digital
1. Introduction
As neoplastic diseases represent the leading cause of death in dogs [1], a thorough understanding of the underlying pathomechanisms is essential to make informed clinical decisions. In oncology, tumour biomarkers hold significant value for diagnosis, prognosis and the development of targeted therapies [2]. Biomarkers can be defined as characteristics of neoplastic processes that can be objectively measured and evaluated as indicators of pathogenic biological activity [3]. Among the numerous molecular biomarkers under investigation, the chromatin remodelling proteins DAXX (death-domain associated protein) and ATRX (alpha-thalassemia/mental retardation syndrome X-linked) have gained increasing attention [4,5] due to their role in epigenetic regulation, genomic stability, and telomere maintenance [6,7]. Among their various functions [8,9], DAXX and ATRX form a complex which is responsible for the deposition of the histone variant H3.3 at repetitive regions of the genome, especially telomeric and pericentromeric DNA [6]. In healthy cells, the proper attachment of the ATRX/DAXX/Histone3.3 complex to these regions is essential for genome stability and heterochromatic architecture, as it prevents aberrant transcription through restricting transcriptional processes [10].
Alterations in the expression of DAXX and/or ATRX can influence tumour biology and have been correlated with tumour stage and prognostic outcomes across different human cancer types, including meningiomas, melanomas, various sarcomas and neuroendocrine neoplasms [8,11]. Among these, pancreatic neuroendocrine tumours (PanNENs) are the best investigated neoplasms in this respect to date [12]. Because loss of DAXX and ATRX expression is associated with shorter patient survival and more aggressive tumour behaviour in PanNENs [13], immunohistochemistry for these markers is included in routine molecular diagnostic panels to support risk stratification and clinical decision making in human medicine [14]. In contrast, Vries et al. have already shown that canine PanNENs differ from their human counterparts regarding their ATRX gene expression, although the majority of canine PanNENs (~ 90%) are aggressive carcinomas in dogs [15]. No loss of ATRX expression was observed in canine PanNENs [16], in contrast to the frequent loss (in ~ 36% of cases) reported in human PanNENs [17].
Furthermore, in veterinary medicine, Kreilmeier et al. investigated DAXX and ATRX expression in the context of alternative lengthening of telomeres (ALT) in various canine sarcomas [18]. Loss of ATRX expression was detected in hemangiosarcoma (1 of 8), histiocytic sarcoma (2 of 5), osteosarcoma (1 of 4), multilobular osteochondrosarcoma (1 of 1), sticker sarcoma (1 of 2), fibrosarcoma (1 of 15), and one undifferentiated sarcoma (1 of 3), whereas loss of DAXX expression was not observed. In addition, Wong et al. reported ATRX mutations in 4 of 15 canine visceral hemangiosarcomas [19]. Mutations of DAXX and ATRX appear to correlate with loss of nuclear immunolabelling, as seen in PanNENs [13,20], although exceptions may occur depending on the tumour type [21].
To date, no veterinary studies have immunohistochemically investigated DAXX or ATRX expression in canine neoplasms other than PanNENs [16] and sarcomas [18]. Nevertheless, their potential value as molecular tumour markers may also be relevant in canine prostatic and urothelial carcinomas. In men, alterations in DAXX and/or ATRX expression have been reported for both prostate and urinary bladder carcinomas. In human prostate carcinomas, strong DAXX protein expression has been shown to correlate with higher malignancy (e.g., higher Gleason grades) and increased tumour cell proliferation [22]. Immunohistochemical studies assessing nuclear ATRX expression in the prostate remain to be investigated, however ATRX mRNA was found to be significantly downregulated in human prostate carcinomas [23]. In human urinary bladder carcinomas, DAXX expression was characterised by reduced nuclear staining intensity compared with normal urothelium [24] and loss of expression was associated with higher tumour stage [25]. Studies specifically assessing ATRX protein expression in urinary bladder carcinomas are currently lacking.
Although DAXX and ATRX immunohistochemistry has not yet been implemented in routine molecular diagnostic panels for prostate or urinary bladder carcinomas in human oncology, these findings support a potential diagnostic and prognostic role for altered expression, and may additionally provide therapeutic relevance in the future as targeted approaches evolve.
In pathological practice, formalin-fixed paraffin-embedded (FFPE) tissue samples represent the most commonly available material for diagnostic and research purposes. Therefore, immunohistochemistry (IHC) represents an ideal approach to estimate protein expression and identify potential new biomarkers, such as DAXX and ATRX. Both antibodies used in this study have been established for canine tissues, and were previously examined in canine sarcomas [18] and pancreatic neuroendocrine neoplasms [16]. As DAXX and ATRX primarily exert their biological functions within the nucleus, nuclear expression is considered the most relevant parameter for immunohistochemical evaluation. ATRX expression is consistently described as nuclear, whereas cytoplasmic localisation of DAXX has been observed in certain human tumour types, for example in gastric cancer [26], and may carry additional biological significance [27]. Nevertheless, nuclear staining remains the established standard for assessing the functional activity of both proteins.
The evaluation of IHC-stained tissue slides traditionally relies on visual assessment of staining intensity. However, this approach can be subject to a certain degree of variability. Even among experienced pathologists, both interobserver [28,29] and intraobsever variability [30] have been reported. To address these limitations, digital pathology is becoming increasingly important for the evaluation and quantification of tissue biomarkers. Automated analysis of scanned tissue slides, frequently aided by artificial intelligence (AI) algorithms, can help pathologists produce results that are more objective, quantitative, and reproducible [31].
This study aims to investigate alterations in the epithelial nuclear expression levels of DAXX and ATRX in canine carcinomas of the prostate and bladder using immunohistochemical methods combined with AI-assisted digital image analysis.
2. Materials and Methods
2.1. Samples
Formalin-fixed, paraffin-embedded (FFPE) samples of benign and malignant prostate and urinary bladder tissues were available from 56 dogs. Included were 18 prostate carcinomas (PC; 12 adenocarcinomas, 6 prostatic urothelial carcinomas), 10 non-malignant prostate samples, 22 urothelial carcinomas of the bladder (UC) and 6 non-malignant bladder samples. An overview of included cases is provided in Supplementary Table S1.
Samples originated from Laboklin GmbH and Co.KG and the Institute of Animal Pathology at the University of Bern, and were retrieved from archival diagnostic material collected between 2000 and 2022. Submission of samples included consent for research use, as specified on the respective submission forms.
2.2. Histology and Tissue Microarray (TMA)
Haematoxylin and eosin (HE)-stained slides were available from each sample. General inclusion criteria required an unambiguous histological diagnosis. Based on HE-stained whole tissue sections, all examined UC originated of the transitional epithelium of the urinary bladder, whereas PC were classified as either adenocarcinomas or prostatic urothelial carcinomas, depending histology and the reported sampling site, following the system proposed by Palmieri et al. [32]. Furthermore, PC were histologically graded using the modified Gleason score [33], ranging from 2 to 10, with score 10 representing the most aggressive and poorly differentiated PC. The Gleason score was applied only to cases in which architectural Gleason patterns could be reliably identified; cases of prostatic urothelial carcinomas remained ungraded. UC were graded as Grade 1, 2, or 3 based on the system described by Valli et al. [34], with Grade 1 representing well-differentiated and Grade 3 highly anaplastic UC. For information on the histological grading of each case, please refer to Supplementary Table S1.
Two tissue microarray (TMA) blocks were created; one for prostate and one for urinary bladder samples, with core diameters of 2 mm and 1 to 6 cores from each case. TMA blocks were manufactured using the TMA Grand Master (3DHistech, Budapest, Hungary) at the Comparative Experimental Pathology department of the Institute of Pathology at the Technical University of Munich in Germany. For each sample, regions of interest were identified on the HE-stained whole tissue sections and subsequently manually transferred to the donor blocks. Where applicable, tissue cores were selected to capture different tumour growth patterns and adjacent non-neoplastic areas. TMA sections were HE-stained before immunohistochemical analyses.
2.3. Immunohistochemistry (IHC)
DAXX and ATRX immunohistochemistry were performed at the Technical University of Munich on SuperFrost slides (Langenbrinck, Emmendingen, Germany) from the TMA blocks. Both antibodies were anti-rabbit polyclonal IgG antibodies from Sigma Aldrich® (DAXX: HPA008736; ATRX: HPA001906). For antigen retrieval, slides were prepared using Tris-EDTA buffer for 30 minutes for DAXX and citrate buffer for 20 minutes for ATRX. Staining for both antibodies was performed using a Bond RXm system (Leica Microsystems, Wetzlar, Germany) with the Bond Polymer Refine Detection Kit. The staining protocol began with a 5-minute peroxide block (Refine Detection Kit Peroxide Block) to suppress endogenous peroxidase activity, followed by incubation with the primary antibody for 15 minutes. Next, an 8-minute incubation with a polymer-based secondary detection system (Refine Detection Kit Polymer) was performed, followed by chromogen development with Diaminobenzidine (Refine Detection Kit Mixed DAB Refine) for 10 minutes. Finally, slides were counterstained with hematoxylin for 5 minutes (Refine Detection Kit Hematoxylin).
Human tonsil tissue was used as positive control for DAXX, and human glioblastoma tissue as positive control for ATRX. Prostate and bladder TMA slides were stained in the same run for both antibodies. Simultaneously with the immunohistochemical staining, slides from both TMAs were prepared without the primary antibody to serve as a negative reagent control (NRC).
2.4. Digital IHC Quantification
Following HE and IHC staining, the glass slides were scanned at 40x resolution using the S360 Nanozoomer (Hamamatsu Photonics, Shizuoka, Japan). Digital assessment of DAXX and ATRX IHC staining was performed by two board-certified veterinary pathologists (S.d.B., L.A.) and a pathology resident (A.S.) using Visiopharm software (version 2025.02 x64, Hørsholm, Denmark). The following workflow was applied: 1) Tissue detection and segmentation into epithelial and stromal (deep learning (DL) classification, U-Net). Epithelium (benign or malignant) was defined as regions of interest (ROI); 2) Revision of ROIs with manual corrections where needed; 3) Detection and labelling of individual epithelial cell nuclei (DL classification, U-Net); 4) Generation of output: total nuclear HDAB-DAB pixel values (range 0 (dark brown; strong immunolabeling) to 255 (white; no immunolabeling) per TMA core (mean, median, standard deviation).
2.5. Statistical Analysis
For all calculations, the mean HDAB-DAB pixel value of all epithelial nuclei within each TMA core was used as the raw data. For each case, aggregated mean values were then calculated as the unweighted average of the means from all TMA cores belonging to that specific case, since all cores were considered equally representative for the respective case.
All statistics were generated in R (Version 4.5.1). HDAB-DAB values (i.e., brown IHC expression levels) were compared between groups (e.g., malignant, non-malignant, Grade 2, Grade 3) using the Mann–Whitney U test. Continuity correction was not applied, as the sample sizes did not allow reliable assumptions of normality, as assessed by the Shapiro-Wilk test. Spearman’s rank correlation was used to evaluate the relationship between DAXX and ATRX expression. Statistical significance was set at p < 0.05. All statistical analyses were performed using the aggregated mean values.
For visualisation purposes, values were linearly inverted (x′ = 255 – x) so that higher values corresponded to stronger immunohistochemical staining intensity. This monotonic transformation preserved relative differences and statistical relationships while enhancing interpretability. The inverted mean values per case are summarised in Supplementary Table S1.
3. Results
3.1. Histomorphological Evaluation in HE-Stained Sections
Among the prostate samples (n = 28), eighteen were classified as prostate carcinoma (PC). Of all PC, twelve were identified as adenocarcinomas originating from the glandular epithelium of the prostate and six were classified as prostatic urothelial carcinomas arising from the prostatic urethra or the prostatic ducts near the urethra. All adenocarcinomas displayed mixed solid and tubular growth patterns, except for one case (case 9), which had a predominantly tubular pattern. Intravascular tumour cells were detected in three adenocarcinomas (cases 1, 2 and 5). Using the veterinary Gleason Scoring system [33], all adenocarcinomas exhibited aggressive histological patterns, ranging from fused glands and vacuolated cytoplasm to solid sheets, cords or single cells without glandular differentiation and comedonecrosis. Consequently, four adenocarcinomas were assigned a Gleason score of 9 (cases 1 – 4), and eight samples were assigned a Gleason score of 10 (cases 5 -12). The number of Gleason score 9 cases in our cohort was relatively small (n = 4). Therefore, differences between adenocarcinomas with Gleason scores 9 and 10 were not further evaluated in this study. Similarly, in human medicine, Gleason scores 9 and 10 are both categorised as Grade Group 5 and are considered to have comparable prognostic behaviour [35].
All six prostatic urethral carcinomas exhibited typical urothelial morphology, without any further subdivision possible. Non-malignant prostate samples included two cases of benign prostatic hyperplasia (cases 26 and 27), one atrophic prostate (case 28), four prostates from juvenile dogs (cases 22 – 25), and three samples of normal prostate tissue (cases 19 – 21).
Among the urinary bladder samples (n = 28), twenty-two were classified as urothelial carcinomas of the bladder (UC), showing clear signs of malignancy, such as disorganised growth, variation in cell and nuclear size with often irregular nuclear crowding and molding, as well as prominent nucleoli. In the most aggressive cases (cases 46 – 50), muscular invasion and numerous, partly abnormal, mitotic figures could be observed.
The predominant growth pattern was papillary in thirteen cases, solid in eight cases, and mixed solid and papillary in one case (case 38). Intravascular tumour cells were detected in two papillary UC (cases 32 and 47) and in the single mixed solid-papillary UC (case 38). According to the grading system from Valli et al. [34], seventeen UC were assigned Grade 2 (cases 29 – 45), and five were UC Grade 3 (cases 46 – 50), while no cases met the criteria for Grade 1. Non-malignant bladder samples included three papillomas (cases 54 – 56), one hyperplasia (case 53), and two samples of healthy urothelium (cases 51 and 52).
Detailed information for each case, including the dog’s breed, sex and age, is provided in Supplementary Table S1.
3.2. Overall DAXX and ATRX Expression in Prostate and Bladder Tissues
The DAXX and ATRX antibodies showed valid and interpretable immunolabeling. Expression of both proteins was indicated by a variably intense brown staining, predominantly localised to the nuclei of positive cells. In cases with very strong expression, immunolabeling extended into the cytoplasm, which was frequently seen in cases with a strong DAXX expression. Even in such cases, however, nuclear expression remained clearly distinguishable from cytoplasmic staining, since the nuclei showed markedly greater staining intensity. For both markers and both anatomical sites, intratumoral heterogeneous expression was frequently observed. In general, DAXX showed stronger staining compared to ATRX, with a mean epithelial nuclear expression level of 165 and 54, respectively.
For DAXX, staining intensity in the interstitial connective tissue varied depending on the presence and extent of inflammation. Stromal cells showed little to no staining, whereas leukocytes were DAXX-positive with variable staining intensity. For ATRX, the staining of stromal cells and leukocytes was generally negative or only weakly positive. Since epithelial nuclear expression was considered more relevant for assessing alterations of DAXX and ATRX in carcinomas, variations in stromal expression were not further analysed.
3.3. Epithelial Nuclear DAXX and ATRX Expression in the Prostate
PC (n = 18) showed a significant increase in epithelial nuclear expression of both DAXX (p = 0.002) and ATRX (p = 0.03) compared with non-malignant prostate tissues (n = 10), as indicated by a notably stronger nuclear staining intensity (Figure 1 and Figure 2). This was particularly evident seen for the DAXX expression. However, one individual case of prostatic adenocarcinoma (case 8) deviated from this pattern, showing clear malignancy despite relatively weak expression of both markers.
No differences in expression levels could be determined between different dog breeds or histological types (adenocarcinoma vs. prostatic urothelial carcinoma; p > 0.05).
All prostate samples demonstrated a strong positive correlation between DAXX and ATRX expression levels (ρ = 0.692, p < 0.001) in both non-malignant tissues and PC.
3.4. Epithelial Nuclear DAXX and ATRX Expression in the Bladder
Comparison of all UC samples (n = 22) with non-malignant bladder tissue (n = 6) revealed no statistically significant difference in epithelial nuclear expression of DAXX or ATRX (p > 0.05). When examining UC of histological Grade 2 (n = 17), both markers showed a tendency towards stronger expression compared to non-malignant tissues, although this did not reach statistical significance (p = 0.071 for DAXX; p = 0.080 for ATRX).
In contrast, UC of histological Grade 3 (n = 5) exhibited lower DAXX expression compared with non-malignant samples (p = 0.046), while ATRX did not differ significantly (p > 0.05).
Importantly, direct comparison between Grade 2 UC and Grade 3 UC revealed significantly lower expression of DAXX (p = 0.002) and ATRX (p = 0.011) in the more aggressive Grade 3 tumours (Figure 3). On histological examination of the HE-stained slides, UC with reduced expression were characterised by high cellular and nuclear pleomorphism and an invasive growth pattern. In contrast, well-differentiated UC with preserved organoid architecture (corresponding to Grade 2) exhibited noticeably stronger expression.
In summary, Grade 2 UC predominantly displayed strong nuclear DAXX and ATRX expression, whereas more invasive and pleomorphic Grade 3 UC showed reduced expression levels (Figure 4).
No significant difference in either DAXX or ATRX expression was observed between different dog breeds.
Similar to the prostate samples, DAXX and ATRX expression levels correlated in both non-malignant tissues and UC, although to a lesser extent (ρ = 0.488, p = 0.0098).
3.5. Main Findings
DAXX and ATRX expression levels showed a positive correlation in both non-malignant and tumour tissues of the prostate and bladder.
The patterns of DAXX and ATRX expression varied between non-malignant and tumour tissues for prostate and bladder. In the prostate, carcinomas showed a significant increase in expression of both proteins compared with non-malignant samples. In the bladder, Grade 2 UC tended to show stronger expression than non-malignant tissues, although this did not reach statistical significance. With increasing malignancy, however, the more aggressive Grade 3 UC displayed a clear reduction in expression of both markers compared with the less malignant Grade 2 UC.
4. Discussion
4.1. Methodological Considerations
Several methodological aspects of this study are important to consider when interpreting the results. The use of AI-based digital IHC quantification enabled the objective evaluation of nuclear expression levels, minimising interobserver or intraobserver variability. This was especially helpful for evaluating ATRX expression, where the generally weaker staining could have made it difficult to recognize subtle differences by eye alone. The method also allowed a precise and reproducible quantification of expression intensity, capturing variations beyond the limits of visual assessment. Nevertheless, the use of AI in immunohistochemical evaluations is not without limitations. Variability in slide preparation, staining quality, and scanner settings can affect algorithmic performance, which is why quality control by an experienced pathologist remains essential to ensure reliable results [36]. In addition, the use of TMAs is advantageous in this regard, as it enables uniform processing conditions across large sample cohorts and thereby reduces technical variability.
Both antibodies used in this study demonstrated reliable immunolabelling, confirming their suitability for use in canine tissues, underlining the results from previous studies [16,18]. The heterogeneity of the tissue collection in our study can be viewed as both a strength and a limitation. While it allowed us to observe DAXX and ATRX expression across different groups of samples (benign vs. malignant; various growth patterns), it also presented challenges for the interpretation of the immunohistochemical results. However, the consistently correlated expression patterns of DAXX and ATRX in both prostate and bladder strongly suggest that our findings reflect a true biological effect rather than an artefact of sample variability.
4.2. Organ-Specific Expression Patterns in Canine Prostate and Urinary Bladder Tissue
Our results indicate that DAXX and ATRX undergo coordinated expression changes during malignant transformation in both prostate and bladder tissues, consistent with their known role as functional partners in chromatin regulation and telomere maintenance in human cancers [8].
In our cohort, DAXX and ATRX expression showed different expression patterns in prostate and urinary bladder cancers. This variation was not completely unexpected, as their expression levels are known to differ markedly across tumour entities, for example ranging from reduced levels in human pancreatic neuroendocrine tumours [13,37] to increased expression in human high-grade meningiomas [11]. Together with our results, these findings highlight that the role of DAXX and ATRX in tumour biology must always be interpreted within the specific context of each tumour type, both in human and veterinary oncology.
In our study, prostate carcinomas demonstrated elevated nuclear expression relative to non-malignant tissues, without significant differences between adenocarcinomas and prostatic urothelial carcinomas. Our observation of DAXX overexpression in prostate cancer aligns with reports from human oncology, where strong DAXX expression has been associated with higher Gleason scores and increased tumour cell proliferation [22]. The observed increase in ATRX expression in canine prostate carcinomas appears to differ from findings in human prostate cancer. While studies assessing ATRX nuclear protein expression in human prostate carcinomas are lacking, Coutinho-Camillo et al. reported significant ATRX mRNA downregulation in most cases [23]. The precise roles of ATRX, and to a lesser extent DAXX, in prostate carcinogenesis remain incompletely understood, and further research is needed to clarify their potential clinical relevance. While overexpression, as observed in the PC of our cohort, may contribute to tumour progression through mechanisms such as autophagy inhibition, as proposed by Puto et al. for primary prostatic malignancies [38], loss of expression could likewise promote genomic instability and thus facilitate tumour initiation [8].
In the urinary bladder, the expression levels of DAXX and ATRX did not differ significantly between non-malignant samples and UC. When comparing UC of histological Grade 3 with non-malignant samples, only DAXX showed a statistically significant decrease in expression. Notably, however, expression levels for both markers could clearly differentiate Grade 2 from Grade 3 UC, with the more malignant Grade 3 UC showing a loss of expression. The reduced DAXX and ATRX expression in more malignant cases is consistent with findings in human oncology, particularly the study from Segersten et al., which reported that loss of DAXX expression was significantly associated with higher tumour stage (T2 – 4 compared to Ta and T1) in human urothelial bladder carcinomas [25].
4.3. Clinical Relevance in Human Medicine and Implications for Veterinary Medicine
In human medicine, DAXX and ATRX have gained increasing clinical relevance as molecular biomarkers. Diagnostically, assessment of their nuclear expression is used routinely to help distinguish well-differentiated neuroendocrine tumours (NETs) and poorly differentiated neuroendocrine carcinomas (NECs), as loss of DAXX or ATRX expression strongly supports the diagnosis of NET, whereas retained nuclear expression is more typical for NEC [14]. For veterinary oncology, our findings indicate a potential diagnostic applicability in canine prostate cancer, as increased expression of both markers could help distinguish carcinomas from non-malignant prostate tissues such as benign prostatic hyperplasia. This may be useful in challenging cases in which conventional histopathological assessment is limited, for example when only small biopsies or fine-needle aspiration (FNA) material is available, or artefacts impede accurate interpretation.
The prognostic value of both markers appears to vary considerably depending on the tumour type. While loss of expression for DAXX and ATRX in human medicine is associated with a worse prognosis in PanNENs [13,39] and urothelial bladder carcinomas [25], loss of ATRX may conversely contribute to a better prognosis in patients with nasopharyngeal carcinoma [40]. Furthermore, high-grade serous ovarian carcinomas seem to show poorer overall survival when exhibiting elevated nuclear DAXX expression [41]. Our findings suggest a potential prognostic value of DAXX and ATRX in canine UC. Although expression levels did not distinguish non-malignant from malignant bladder tissues, UC showed a clear reduction of DAXX and ATRX expression with increasing tumour aggressiveness. Thus, both markers may help identify biologically more aggressive UC and could serve as indicators of malignant progression.
From a therapeutic perspective, loss of DAXX and/or ATRX expression has been associated with activation of alternative lengthening of telomeres (ALT), a mechanism that enables cancer cells to maintain telomere length and sustain proliferation [42]. Immunohistochemically detectable loss of these proteins serve as a practical surrogate marker for ALT. Therapeutic approaches targeting ALT-positive tumours, such as ATR-kinase inhibitors like Ceralasertib, are currently under investigation in human oncology [43,44]. As already mentioned, the occurrence of ALT activity has also been examined in canine sarcomas by Kreilmeier et al. [18]. They detected ALT in 9.4% of analysed cases, with all ALT-positive sarcomas belonging to non-soft tissue entities. Notably, all ALT-positive tumours in their study demonstrated a loss of ATRX expression. Similarly, Bicanova et al. reported ALT activity in 20% (10 of 50) of canine appendicular osteosarcoma [45]. To our knowledge no further studies in veterinary medicine have yet examined for ALT mechanisms in canine tumour types other than sarcomas. Furthermore, the mutation or loss of expression of ATRX may increase sensitivity to poly(ADP-ribose)-polymerase (PARP) inhibitors [46]. Notably, the PARP inhibitor Olaparib has already shown promising effects in canine cancer cell lines and may serve as a potential oral therapy alternative to classic chemotherapy in dogs with hematopoietic cancer [47]. Our findings suggest that therapeutic approaches targeting tumours with loss of expression for DAXX and/or ATRX may be of particular interest for highly aggressive UC in veterinary oncology. Future studies should investigate whether UC with reduced DAXX and ATRX expression also show activation of the ALT pathway.
Experimental strategies focusing on inhibiting of DAXX and ATRX overexpression are currently being explored in human medicine, although no clinically established inhibitors are available to date [48,49]. This may will become relevant for canine tumour types with elevated expression of these proteins, such as the prostate carcinomas in our study.
While personalised tumour therapy, which is precisely adapted to the individual patient and genetic profile of the tumour, is already well established in human medicine [50], veterinary oncology is still in the early stages of applying such approaches [51]. Therefore, it is particularly important to deepen our understanding of the underlying mechanisms driving tumour development and progression and to further explore potential therapeutic options in veterinary medicine.
4.4. Conclusions
In conclusion, our study demonstrated that epithelial DAXX and ATRX expression levels are positively correlated and display organ-specific patterns in prostate and bladder tissues. Prostate carcinomas showed increased expression of both proteins compared with non-malignant tissues, whereas in the bladder, expression declined with increasing aggressiveness of the tumour. These findings offer novel insights into the potential role of DAXX and ATRX as biomarkers for these cancer types, providing a foundation for future studies to explore their clinical significance in dogs. AI-assisted digital IHC evaluation enables a fully quantitative, automated, and reproducible analysis, which is particularly important for validating new biomarkers, such as DAXX and ATRX.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Overview of included cases examined for DAXX and ATRX expression.
Author Contributions
Conceptualization, A.S., S.d.B. and H.A.-L; methodology, A.S., S.d.B. and H.A.-L.; software, S.d.B.; data curation, S.d.B., A.S. and M.S.; writing—original draft preparation, A.S.; writing—review and editing, H.A.-L., S.d.B., S.W., L.A., M.S., R.K. and A.S.; visualization, A.S. and M.S.; supervision, S.d.B. and H.A.-L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Albert Heim Foundation (grant number 148) and by internal funding from the Bern Center for Precision Medicine.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to thank the technical staff at the Institute of Animal Pathology at the University of Bern, the Institute of Pathology at the Technical University of Munich, and LABOKLIN GmbH and Co. KG for their assistance with slide preparation and scanning.
Conflicts of Interest
The authors Annika Spitzer, Martin Spitzer and Heike Aupperle-Lellbach are employees of Company LABOKLIN GmbH and Co.KG. The remaining authors have no conflicts of interest to declare.
References
- Adams, V.J.; Evans, K.M.; Sampson, J.; Wood, J.L.N. Methods and mortality results of a health survey of purebred dogs in the UK. J. Small Anim. Pract. 2010, 51, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tao, L.; Qiu, J.; Xu, J.; Yang, X.; Zhang, Y.; Tian, X.; Guan, X.; Cen, X.; Zhao, Y. Tumor biomarkers for diagnosis, prognosis and targeted therapy. Signal Transduct. Target. Ther. 2024, 9, 132. [Google Scholar] [CrossRef]
- Biomarkers Definitions working Group. Biomarkers and surrogate endpoints: preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar] [CrossRef]
- Clatterbuck Soper, S.F.; Walker, R.L.; Pineda, M.A.; Zhu, Y.J.; Dalgleish, J.L.T.; Wang, J.; Meltzer, P.S. Cancer-associated DAXX mutations reveal a critical role for ATRX localization in ALT suppression. bioRxiv 2024. [Google Scholar] [CrossRef]
- Pang, Y.; Chen, X.; Ji, T.; Cheng, M.; Wang, R.; Zhang, C.; Liu, M.; Zhang, J.; Zhong, C. The Chromatin Remodeler ATRX: Role and Mechanism in Biology and Cancer. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.D.; Banaszynski, L.A.; Noh, K.-M.; Lewis, P.W.; Elsaesser, S.J.; Stadler, S.; Dewell, S.; Law, M.; Guo, X.; Li, X.; et al. Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell 2010, 140, 678–691. [Google Scholar] [CrossRef] [PubMed]
- Campos, E.I.; Reinberg, D. New chaps in the histone chaperone arena. Genes Dev. 2010, 24, 1334–1338. [Google Scholar] [CrossRef]
- Dyer, M.A.; Qadeer, Z.A.; Valle-Garcia, D.; Bernstein, E. ATRX and DAXX: Mechanisms and Mutations. Cold Spring Harb. Perspect. Med. 2017, 7. [Google Scholar] [CrossRef]
- Aguilera, P.; López-Contreras, A.J. ATRX, a guardian of chromatin. Trends Genet. 2023, 39, 505–519. [Google Scholar] [CrossRef]
- Groth, A.; Rocha, W.; Verreault, A.; Almouzni, G. Chromatin challenges during DNA replication and repair. Cell 2007, 128, 721–733. [Google Scholar] [CrossRef]
- Cavalcante, S.G.; Pereira, B.J.A.; Lerario, A.M.; Sola, P.R.; Oba-Shinjo, S.M.; Marie, S.K.N. The chromatin remodeler complex ATRX-DAXX-H3.3 and telomere length in meningiomas. Clin. Neurol. Neurosurg. 2021, 210, 106962. [Google Scholar] [CrossRef]
- Wilde, R.F. de; Heaphy, C.M.; Maitra, A.; Meeker, A.K.; Edil, B.H.; Wolfgang, C.L.; Ellison, T.A.; Schulick, R.D.; Molenaar, I.Q.; Valk, G.D.; et al. Loss of ATRX or DAXX expression and concomitant acquisition of the alternative lengthening of telomeres phenotype are late events in a small subset of MEN-1 syndrome pancreatic neuroendocrine tumors. Mod. Pathol. 2012, 25, 1033–1039. [Google Scholar] [CrossRef]
- Marinoni, I.; Kurrer, A.S.; Vassella, E.; Dettmer, M.; Rudolph, T.; Banz, V.; Hunger, F.; Pasquinelli, S.; Speel, E.-J.; Perren, A. Loss of DAXX and ATRX are associated with chromosome instability and reduced survival of patients with pancreatic neuroendocrine tumors. Gastroenterology 2014, 146, 453–60.e5. [Google Scholar] [CrossRef] [PubMed]
- Rindi, G.; Mete, O.; Uccella, S.; Basturk, O.; La Rosa, S.; Brosens, L.A.A.; Ezzat, S.; Herder, W.W. de; Klimstra, D.S.; Papotti, M.; et al. Overview of the 2022 WHO Classification of Neuroendocrine Neoplasms. Endocr. Pathol. 2022, 33, 115–154. [Google Scholar] [CrossRef] [PubMed]
- Tumors in domestic animals; Meuten, D. J., Ed., 5th ed.; John Wiley & Sons: Ames, 2017; ISBN 9780813821795. [Google Scholar]
- Vries, C. de; Konukiewitz, B.; Weichert, W.; Klöppel, G.; Aupperle-Lellbach, H.; Steiger, K. Do Canine Pancreatic Neuroendocrine Neoplasms Resemble Human Pancreatic Neuroendocrine Tumours? A Comparative Morphological and Immunohistochemical Investigation. J. Comp. Pathol. 2020, 181, 73–85. [Google Scholar] [CrossRef]
- Yachida, S.; Vakiani, E.; White, C.M.; Zhong, Y.; Saunders, T.; Morgan, R.; Wilde, R.F. de; Maitra, A.; Hicks, J.; Demarzo, A.M.; et al. Small cell and large cell neuroendocrine carcinomas of the pancreas are genetically similar and distinct from well-differentiated pancreatic neuroendocrine tumors. Am. J. Surg. Pathol. 2012, 36, 173–184. [Google Scholar] [CrossRef]
- Kreilmeier, T.; Sampl, S.; Deloria, A.J.; Walter, I.; Reifinger, M.; Hauck, M.; Borst, L.B.; Holzmann, K.; Kleiter, M. Alternative lengthening of telomeres does exist in various canine sarcomas. Mol. Carcinog. 2017, 56, 923–935. [Google Scholar] [CrossRef]
- Wong, K.; Ludwig, L.; Krijgsman, O.; Adams, D.J.; Wood, G.A.; van der Weyden, L. Comparison of the oncogenomic landscape of canine and feline hemangiosarcoma shows novel parallels with human angiosarcoma. Dis. Model. Mech. 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Shi, C.; Edil, B.H.; Wilde, R.F. de; Klimstra, D.S.; Maitra, A.; Schulick, R.D.; Tang, L.H.; Wolfgang, C.L.; Choti, M.A.; et al. DAXX/ATRX, MEN1, and mTOR pathway genes are frequently altered in pancreatic neuroendocrine tumors. Science 2011, 331, 1199–1203. [Google Scholar] [CrossRef]
- Chami, R.; Marrano, P.; Teerapakpinyo, C.; Arnoldo, A.; Shago, M.; Shuangshoti, S.; Thorner, P.S. Immunohistochemistry for ATRX Can Miss ATRX Mutations: Lessons From Neuroblastoma. Am. J. Surg. Pathol. 2019, 43, 1203–1211. [Google Scholar] [CrossRef]
- Tsourlakis, M.C.; Schoop, M.; Plass, C.; Huland, H.; Graefen, M.; Steuber, T.; Schlomm, T.; Simon, R.; Sauter, G.; Sirma, H.; et al. Overexpression of the chromatin remodeler death-domain-associated protein in prostate cancer is an independent predictor of early prostate-specific antigen recurrence. Hum. Pathol. 2013, 44, 1789–1796. [Google Scholar] [CrossRef]
- Coutinho-Camillo, C.M.; Miracca, E.C.; dos Santos, M.L.; Salaorni, S.; Sarkis, A.S.; Nagai, M.A. Identification of differentially expressed genes in prostatic epithelium in relation to androgen receptor CAG repeat length. Int. J. Biol. Markers 2006, 21, 96–105. [Google Scholar] [CrossRef]
- Zizzi, A.; Montironi, M.A.; Mazzucchelli, R.; Scarpelli, M.; Lopez-Beltran, A.; Cheng, L.; Paone, N.; Castellini, P.; Montironi, R. Immunohistochemical analysis of chromatin remodeler DAXX in high grade urothelial carcinoma. Diagn. Pathol. 2013, 8, 111. [Google Scholar] [CrossRef]
- Segersten, M.U.; Edlund, E.K.; Micke, P.; La Torre, M. de; Hamberg, H.; Edvinsson, A.E.L.; Andersson, S.E.C.; Malmström, P.-U.; Wester, H.K. A novel strategy based on histological protein profiling in-silico for identifying potential biomarkers in urinary bladder cancer. BJU international 2009, 104, 1780–1785. [Google Scholar] [CrossRef]
- Chen, C.; Sun, X.; Xie, W.; Chen, S.; Hu, Y.; Xing, D.; Xu, J.; Chen, X.; Zhao, Z.; Han, Z.; et al. Opposing biological functions of the cytoplasm and nucleus DAXX modified by SUMO-2/3 in gastric cancer. Cell Death Dis. 2020, 11, 514. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, I.; Liao, D. DAXX in cancer: phenomena, processes, mechanisms and regulation. Nucleic Acids Res. 2019, 47, 7734–7752. [Google Scholar] [CrossRef] [PubMed]
- Klarskov, L.; Ladelund, S.; Holck, S.; Roenlund, K.; Lindebjerg, J.; Elebro, J.; Halvarsson, B.; Salomé, J. von; Bernstein, I.; Nilbert, M. Interobserver variability in the evaluation of mismatch repair protein immunostaining. Hum. Pathol. 2010, 41, 1387–1396. [Google Scholar] [CrossRef]
- Butter, R.; Hondelink, L.M.; van Elswijk, L.; Blaauwgeers, J.L.G.; Bloemena, E.; Britstra, R.; Bulkmans, N.; van Gulik, A.L.; Monkhorst, K.; Rooij, M.J. de; et al. The impact of a pathologist’s personality on the interobserver variability and diagnostic accuracy of predictive PD-L1 immunohistochemistry in lung cancer. Lung Cancer 2022, 166, 143–149. [Google Scholar] [CrossRef]
- Gavrielides, M.A.; Gallas, B.D.; Lenz, P.; Badano, A.; Hewitt, S.M. Observer variability in the interpretation of HER2/neu immunohistochemical expression with unaided and computer-aided digital microscopy. Arch. Pathol. Lab. Med. 2011, 135, 233–242. [Google Scholar] [CrossRef]
- Zuraw, A.; Aeffner, F. Whole-slide imaging, tissue image analysis, and artificial intelligence in veterinary pathology: An updated introduction and review. Vet. Pathol. 2022, 59, 6–25. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, C.; Foster, R.A.; Grieco, V.; Fonseca-Alves, C.E.; Wood, G.A.; Culp, W.T.N.; Murua Escobar, H.; Marzo, A.M. de; Laufer-Amorim, R. Histopathological Terminology Standards for the Reporting of Prostatic Epithelial Lesions in Dogs. J. Comp. Pathol. 2019, 171, 30–37. [Google Scholar] [CrossRef]
- Palmieri, C.; Grieco, V. Proposal of Gleason-like grading system of canine prostate carcinoma in veterinary pathology practice. Res. Vet. Sci. 2015, 103, 11–15. [Google Scholar] [CrossRef]
- Valli, V.E.; Norris, A.; Jacobs, R.M.; Laing, E.; Withrow, S.; Macy, D.; Tomlinson, J.; McCaw, D.; Ogilvie, G.K.; Pidgeon, G. Pathology of canine bladder and urethral cancer and correlation with tumour progression and survival. J. Comp. Pathol. 1995, 113, 113–130. [Google Scholar] [CrossRef]
- Pierorazio, P.M.; Walsh, P.C.; Partin, A.W.; Epstein, J.I. Prognostic Gleason grade grouping: data based on the modified Gleason scoring system. BJU international 2013, 111, 753–760. [Google Scholar] [CrossRef]
- Poalelungi, D.G.; Neagu, A.I.; Fulga, A.; Neagu, M.; Tutunaru, D.; Nechita, A.; Fulga, I. Revolutionizing Pathology with Artificial Intelligence: Innovations in Immunohistochemistry. J. Pers. Med. 2024, 14. [Google Scholar] [CrossRef]
- Cives, M.; Partelli, S.; Palmirotta, R.; Lovero, D.; Mandriani, B.; Quaresmini, D.; Pelle’, E.; Andreasi, V.; Castelli, P.; Strosberg, J.; et al. DAXX mutations as potential genomic markers of malignant evolution in small nonfunctioning pancreatic neuroendocrine tumors. Sci. Rep. 2019, 9, 18614. [Google Scholar] [CrossRef] [PubMed]
- Puto, L.A.; Brognard, J.; Hunter, T. Transcriptional Repressor DAXX Promotes Prostate Cancer Tumorigenicity via Suppression of Autophagy. J. Biol. Chem. 2015, 290, 15406–15420. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, I. Prognostic value of DAXX/ATRX loss of expression and ALT activation in PanNETs: is it time for clinical implementation? Gut 2022, 71, 847–848. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, H.; Wang, S.; Feng, Y.; Feng, Y.; Fan, S.; Hu, C.; Liu, X.; Hou, T. Clinicopathological Significance of ATRX Expression in Nasopharyngeal Carcinoma Patients: A Retrospective Study. J. Cancer 2021, 12, 6931–6936. [Google Scholar] [CrossRef]
- Davidson, B.; McFadden, E.; Holth, A.; Brunetti, M.; Flørenes, V.A. Death domain-associated protein (DAXX) expression is associated with poor survival in metastatic high-grade serous carcinoma. Virchows Arch. 2020, 477, 857–864. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.; Watters, A.K.; To, J.T.; Young, M.W.; Muratori, J.; Wilkoff, M.H.; Abraham, R.G.; Plummer, M.M.; Zhang, D. ALT Positivity in Human Cancers: Prevalence and Clinical Insights. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Carson, L.M.; Flynn, R.L. Highlighting vulnerabilities in the alternative lengthening of telomeres pathway. Curr. Opin. Pharmacol. 2023, 70, 102380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; Zou, L. Alternative lengthening of telomeres: from molecular mechanisms to therapeutic outlooks. Cell Biosci. 2020, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Bicanova, L.; Kreilmeier-Berger, T.; Reifinger, M.; Holzmann, K.; Kleiter, M. Prevalence and potentially prognostic value of C-circles associated with alternative lengthening of telomeres in canine appendicular osteosarcoma. Vet. Comp. Oncol. 2021, 19, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Garbarino, J.; Eckroate, J.; Sundaram, R.K.; Jensen, R.B.; Bindra, R.S. Loss of ATRX confers DNA repair defects and PARP inhibitor sensitivity. Translational oncology 2021, 14, 101147. [Google Scholar] [CrossRef]
- Pasaol, J.C.; Dejnaka, E.; Mucignat, G.; Bajzert, J.; Henklewska, M.; Obmińska-Mrukowicz, B.; Giantin, M.; Pauletto, M.; Zdyrski, C.; Dacasto, M.; et al. PARP inhibitor olaparib induces DNA damage and acts as a drug sensitizer in an in vitro model of canine hematopoietic cancer. BMC Vet. Res. 2025, 21, 439. [Google Scholar] [CrossRef]
- Scbt. Daxx Inhibitoren | SCBT - Santa Cruz Biotechnology. Available online: https://www.scbt.com/browse/daxx-inhibitors?pageNum=2 (accessed on 9 October 2025).
- Scbt. ATRX Inhibitors | SCBT - Santa Cruz Biotechnology. Available online: https://www.scbt.com/browse/atrx-inhibitors?srsltid=AfmBOooT6ZhjNw6OP1KpJZL0COGLr7SItvehYQ1DsYUlz4bwpt7DyrOk (accessed on 29 October 2025).
- Singh, D.; Dhiman, V.K.; Pandey, M.; Dhiman, V.K.; Sharma, A.; Pandey, H.; Verma, S.K.; Pandey, R. Personalized medicine: An alternative for cancer treatment. Cancer Treat. Res. Commun. 2024, 42, 100860. [Google Scholar] [CrossRef]
- Chibuk, J.; Flory, A.; Kruglyak, K.M.; Leibman, N.; Nahama, A.; Dharajiya, N.; van den Boom, D.; Jensen, T.J.; Friedman, J.S.; Shen, M.R.; et al. Horizons in Veterinary Precision Oncology: Fundamentals of Cancer Genomics and Applications of Liquid Biopsy for the Detection, Characterization, and Management of Cancer in Dogs. Front. Vet. Sci. 2021, 8, 664718. [Google Scholar] [CrossRef]
Figure 1.
Nuclear expression levels of DAXX (left) and ATRX (right) in prostate tissues. Prostate carcinomas exhibited overall stronger expression compared with non-malignant samples. This was particularly pronounced for DAXX (p = 0.002) but also evident for ATRX (p = 0.030). Across all samples, DAXX expression was generally stronger than ATRX expression. No statistically significant differences were observed between prostatic urothelial carcinomas (PUC) and adenocarcinomas (PAC). Black bars represent the median expression values for each subgroup. Abbreviations: PUC, prostatic urothelial carcinomas; PAC, prostatic adenocarcinomas.
Figure 1.
Nuclear expression levels of DAXX (left) and ATRX (right) in prostate tissues. Prostate carcinomas exhibited overall stronger expression compared with non-malignant samples. This was particularly pronounced for DAXX (p = 0.002) but also evident for ATRX (p = 0.030). Across all samples, DAXX expression was generally stronger than ATRX expression. No statistically significant differences were observed between prostatic urothelial carcinomas (PUC) and adenocarcinomas (PAC). Black bars represent the median expression values for each subgroup. Abbreviations: PUC, prostatic urothelial carcinomas; PAC, prostatic adenocarcinomas.
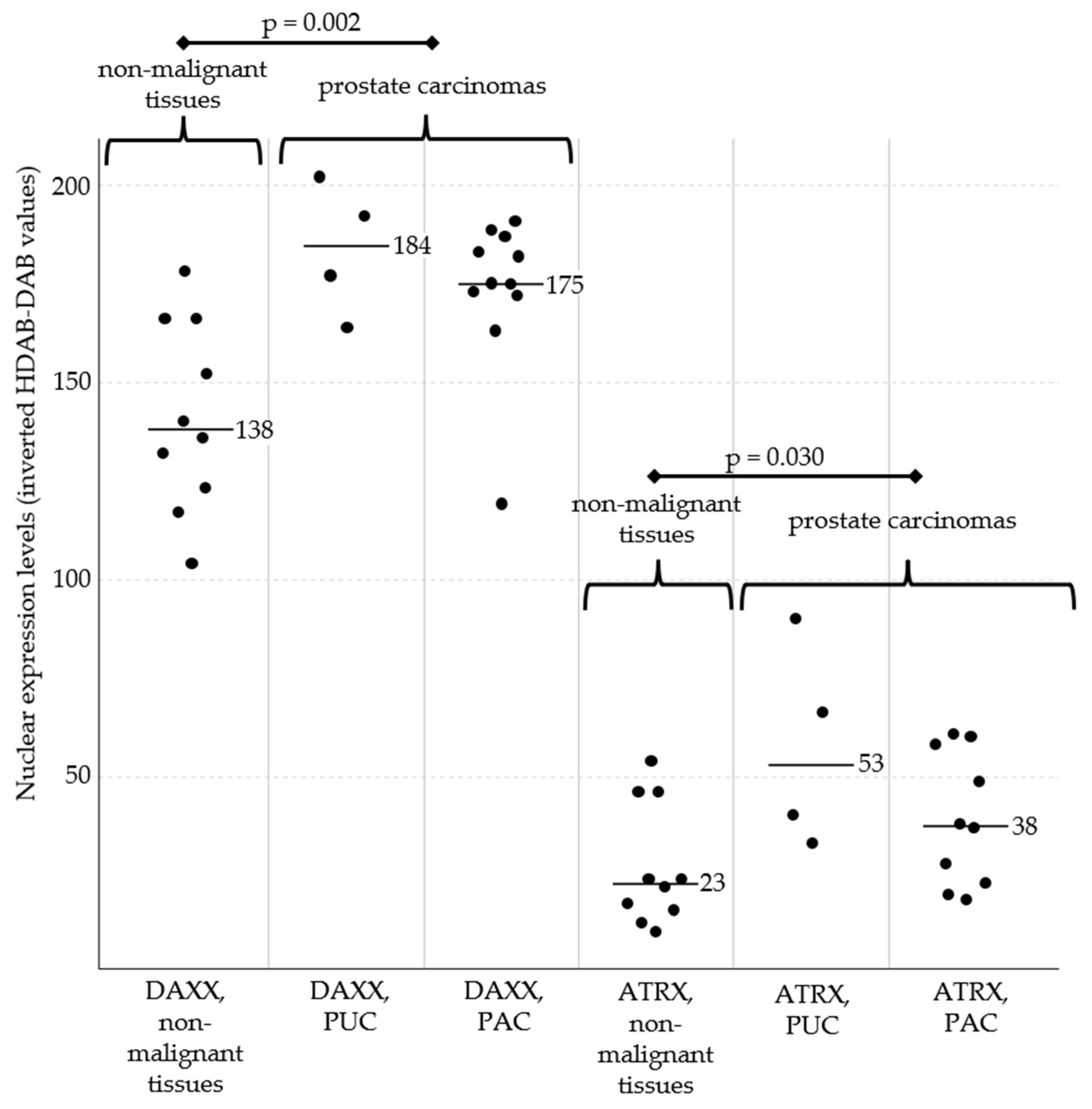
Figure 2.
DAXX and ATRX immunohistochemistry of canine prostate tissue; bar indicates 100 µm. Immunostaining with both antibodies showed predominant nuclear expression of DAXX and ATRX in epithelial cells. DAXX (a, c, e) showed overall stronger expression compared to ATRX (b, d, f) across all prostate samples. PC (c - f) showed stronger DAXX and ATRX expression compared to benign prostatic conditions such as hyperplasia (a, b; case 20). This increase was observed in both, prostatic urothelial carcinoma (c, d; case 15) and adenocarcinoma (e, f; case 7), and was particularly pronounced for DAXX. Intratumoral heterogeneity in the expression of epithelial DAXX and ATRX could be observed, represented by variable strong immunolabelling of individual tumour cell nuclei.
Figure 2.
DAXX and ATRX immunohistochemistry of canine prostate tissue; bar indicates 100 µm. Immunostaining with both antibodies showed predominant nuclear expression of DAXX and ATRX in epithelial cells. DAXX (a, c, e) showed overall stronger expression compared to ATRX (b, d, f) across all prostate samples. PC (c - f) showed stronger DAXX and ATRX expression compared to benign prostatic conditions such as hyperplasia (a, b; case 20). This increase was observed in both, prostatic urothelial carcinoma (c, d; case 15) and adenocarcinoma (e, f; case 7), and was particularly pronounced for DAXX. Intratumoral heterogeneity in the expression of epithelial DAXX and ATRX could be observed, represented by variable strong immunolabelling of individual tumour cell nuclei.
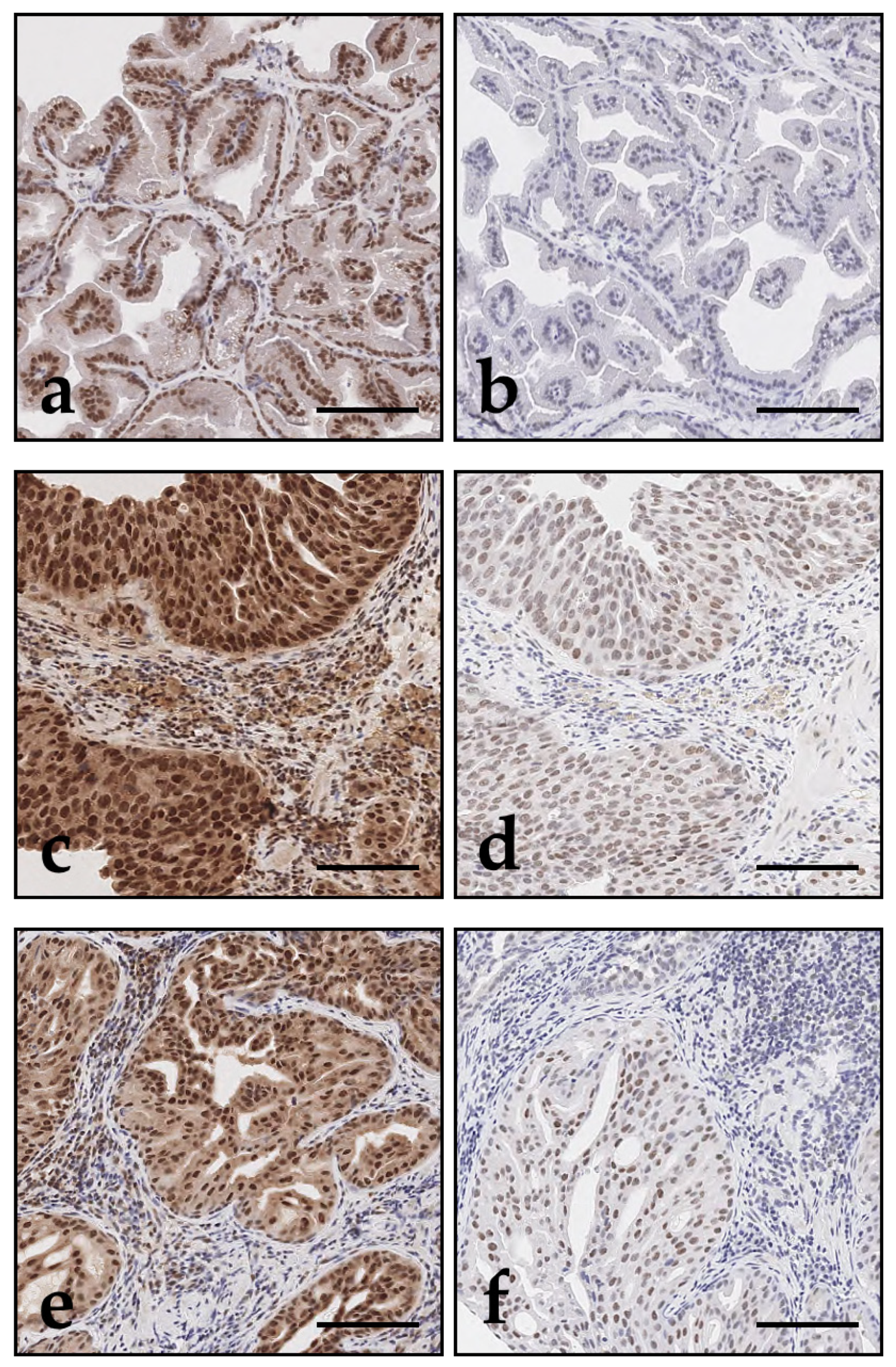
Figure 3.
Nuclear expression levels of DAXX (left) and ATRX (right) in urinary bladder tissues. DAXX and ATRX expression did not significantly differ between all urothelial carcinomas (UC) and non-malignant bladder tissues. UC of histological Grade 2 showed a tendency towards stronger expression of both markers compared with non-malignant samples, although this did not reach statistical significance (p = 0.071 for DAXX; p = 0.080 for ATRX). Importantly, the more aggressive Grade 3 UC showed significantly lower expression of DAXX and ATRX compared to Grade 2 UC. Across all samples, DAXX expression was generally stronger than ATRX expression. Black bars represent the median expression values for each subgroup. Abbreviations: UC, urothelial carcinomas of the urinary bladder.
Figure 3.
Nuclear expression levels of DAXX (left) and ATRX (right) in urinary bladder tissues. DAXX and ATRX expression did not significantly differ between all urothelial carcinomas (UC) and non-malignant bladder tissues. UC of histological Grade 2 showed a tendency towards stronger expression of both markers compared with non-malignant samples, although this did not reach statistical significance (p = 0.071 for DAXX; p = 0.080 for ATRX). Importantly, the more aggressive Grade 3 UC showed significantly lower expression of DAXX and ATRX compared to Grade 2 UC. Across all samples, DAXX expression was generally stronger than ATRX expression. Black bars represent the median expression values for each subgroup. Abbreviations: UC, urothelial carcinomas of the urinary bladder.
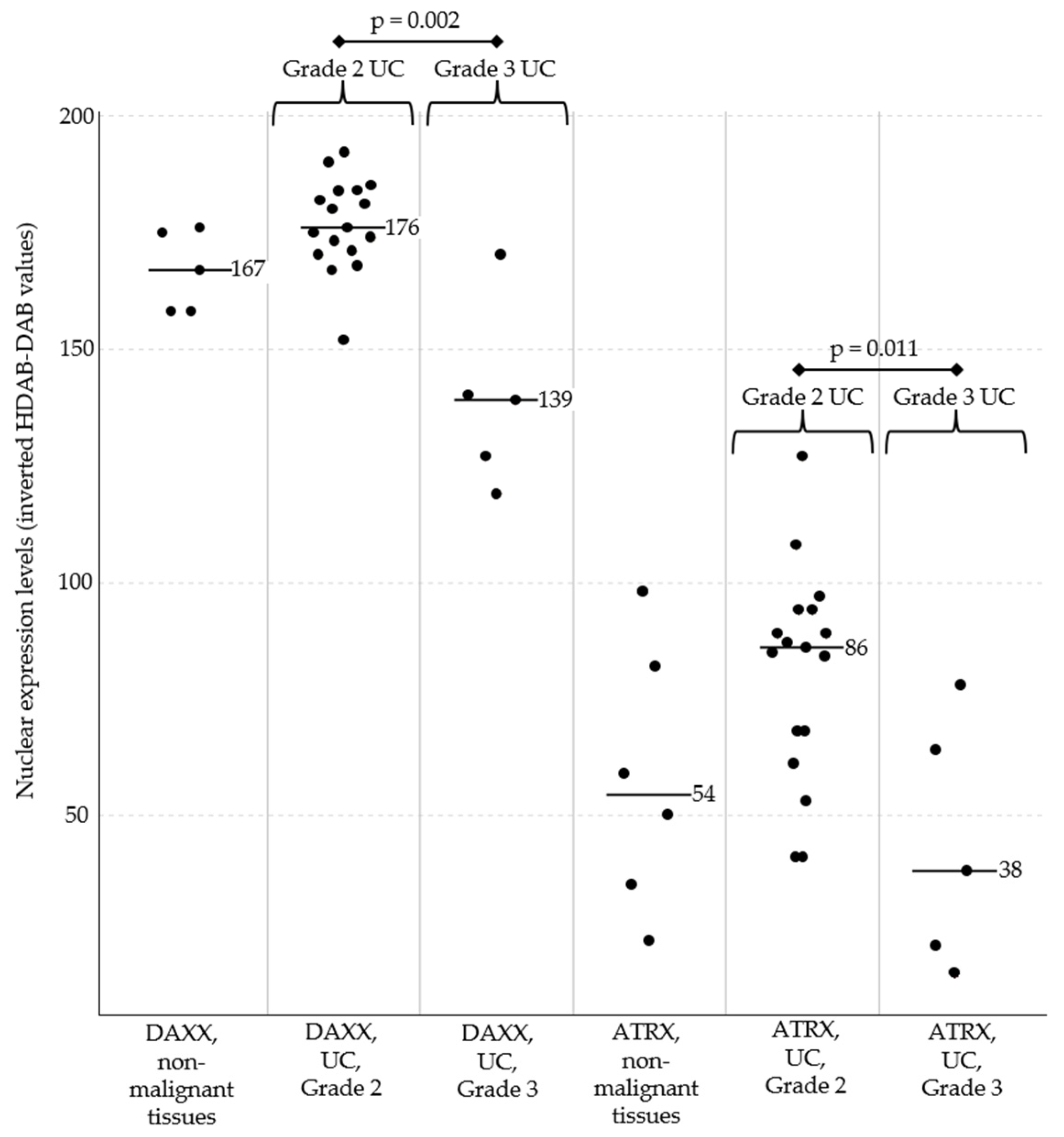
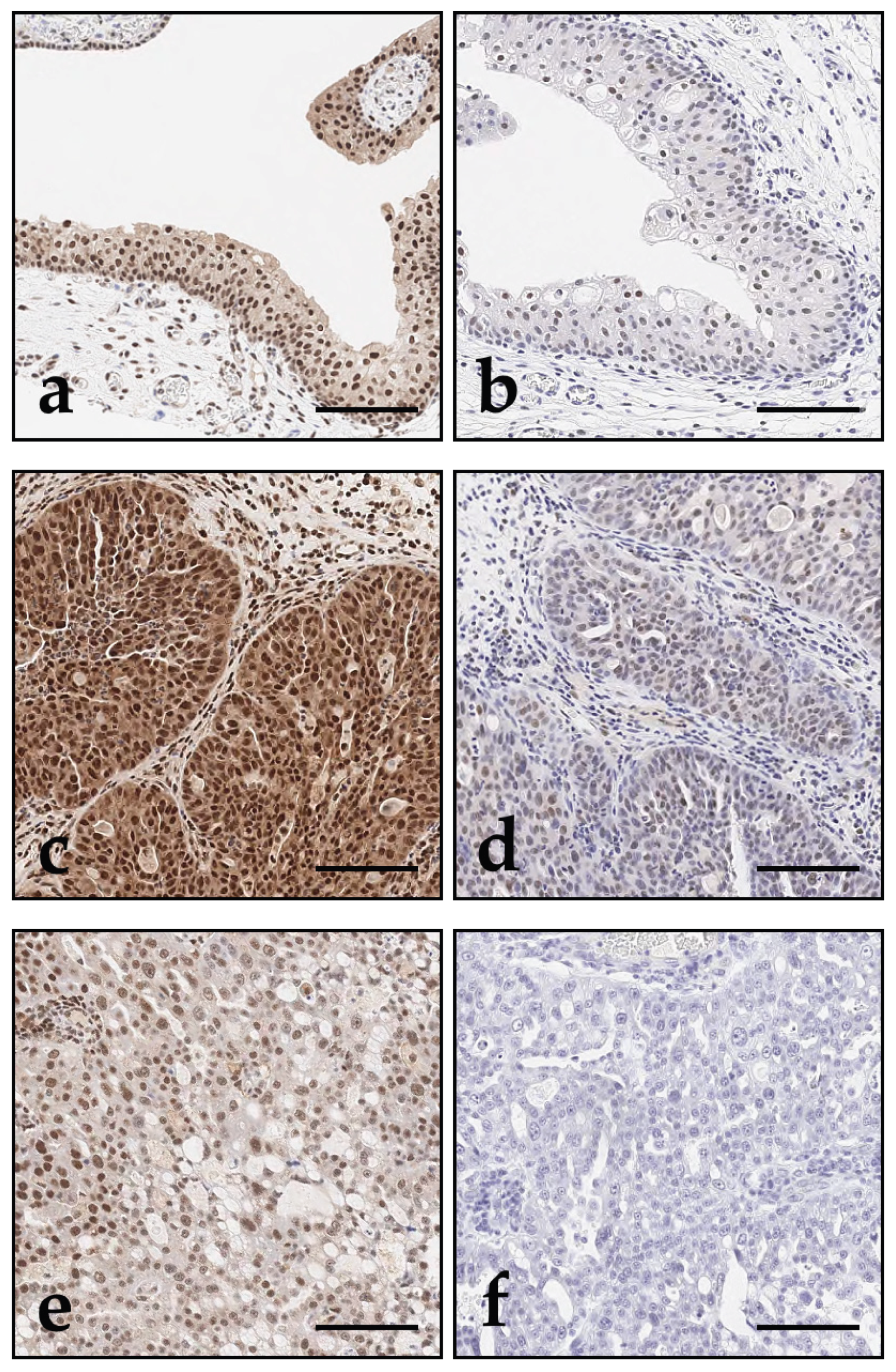
Figure 4.
DAXX and ATRX immunohistochemistry of canine bladder tissue; bar indicates 100 µm. Across all bladder samples, DAXX (a, c, e) showed overall stronger expression compared to ATRX (b, d, f). While Grade 2 UC (c, d; case 45) tended to show stronger expression compared with non-malignant urothelium (e, f; case 52), expression of both DAXX and ATRX decreased in the more aggressive Grade 3 UC (e, f; case 49). Within tumours, expression heterogeneity was observed, with loss of nuclear immunolabelling for DAXX and ATRX in regions with increased tumour cell pleomorphism (e, f).
Figure 4.
DAXX and ATRX immunohistochemistry of canine bladder tissue; bar indicates 100 µm. Across all bladder samples, DAXX (a, c, e) showed overall stronger expression compared to ATRX (b, d, f). While Grade 2 UC (c, d; case 45) tended to show stronger expression compared with non-malignant urothelium (e, f; case 52), expression of both DAXX and ATRX decreased in the more aggressive Grade 3 UC (e, f; case 49). Within tumours, expression heterogeneity was observed, with loss of nuclear immunolabelling for DAXX and ATRX in regions with increased tumour cell pleomorphism (e, f).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



