
Submitted:
10 November 2025
Posted:
12 November 2025
You are already at the latest version
Abstract
Diffuse large B-cell lymphoma (DLBCL) is the most common and clinically aggressive subtype of non-Hodgkin lymphoma (NHL). Although novel therapeutic agents, in-cluding rituximab and polatuzumab vedotin, have improved outcomes, almost one-third of patients ultimately develop relapsed or refractory disease. MicroRNAs (miRNAs), a class of endogenous single-stranded RNAs approximately 22 nucleotides in length, play a pivotal role in the regulation of gene expression at the post-transcriptional level through interactions with complementary target RNAs and contribute significantly to the de-velopment, progression, and treatment response of DLBCL. Oncogenic miRNAs, such as miR-155, miR-21, and the miR-17–92 cluster, promote proliferation, survival, immune evasion, and therapy resistance by modulating pathways including PI3K/AKT, NF-κB, and MYC. Conversely, tumor-suppressive miRNAs such as miR-34a, miR-144, miR-181a, and miR-124-3p inhibit oncogene activity and enhance apoptosis, with their loss often associated with adverse outcomes. Among these, miR-155 and miR-21 are particularly well studied, playing central roles in both tumor progression and remodeling of the tumor microenvironment. This review summarizes current evidence on the biological and clinical relevance of miRNAs in DLBCL, emphasizing their diagnostic and prognostic potential.
Keywords:
miRNAs
; DLBCL
; lymphoma
; miR-155
; miR-21
; miR-17-92
; non-Hodgkin
; non-coding RNA
; miR-34
1. Introduction
Diffuse large B-cell lymphoma (DLBCL) is a highly aggressive type of non-Hodgkin lymphoma (NHL), characterized by extensive molecular and pathological heterogeneity. This variation is reflected in its clinical behavior, resulting in diverse therapeutic responses and prognoses [1,2]. DLBCL is the most frequent form of NHL, with incidence rates over recent years. Rituximab (R), a monoclonal antibody targeting the CD20 antigen on B-cell surfaces, plays a central role in the treatment of DLBCL. Its addition to the CHOP regimen—comprising cyclophosphamide, doxorubicin, vincristine, and prednisolone—markedly improved 5-year overall survival (OS) rates (58% compared to 45%). As a result, the R-CHOP combination became the established first-line therapy for DLBCL patients for many years [3]. R-CHOP induces durable remission in 50–70% of patients; however, up to 30% relapse and 20% present with primary refractory disease [4,5,6]. In first-line treatment, substitution of vincristine with polatuzumab vedotin, an antibody–drug conjugate targeting CD79b, resulted in improved progression-free survival (PFS) but did not significantly affect OS [7]. DLBCL may arise through the transformation of a pre-existing indolent lymphoma, such as follicular lymphoma (FL). However, most DLBCL cases emerge de novo, without prior indolent lymphoma [8].
In terms of molecular characteristics, gene expression profiling has classified DLBCL into three distinct molecular subtypes: the germinal center B-cell–like (GCB) subtype, the activated B-cell–like (ABC) subtype, and a third category referred to as type 3 or unclassified cases [9]. More recently, advanced taxonomic systems have provided a more detailed classification of DLBCL based on comprehensive molecular and cytogenetic analyses [10]. A proposed molecular framework includes the following genetic subtypes: (a) MCD, characterized by co-mutations in MYD88 L265P and CD79B; (b) BN2, defined by BCL6 fusions or NOTCH2 mutations; (c) N1, associated with NOTCH1 mutations; and (d) EZB, involving EZH2 mutations or BCL2 translocations. Despite these advances, a substantial subset of patients still does not fit into any of these defined categories [11]. A genomic analysis of 304 DLBCL cases led to the proposal of an alternative molecular classification, comprising five distinct clusters (C):
C1, characterized by NOTCH2 mutations and linked to favorable prognosis;
C2, associated with aneuploidy and biallelic inactivation of TP53, correlating with poor clinical outcomes;
C3, defined by BCL2 mutations and translocations, along with mutations in genes involved in epigenetic regulation, is also linked to adverse prognosis;
C4, marked by alterations in signaling pathways such as JAK/STAT, and associated with better outcomes; and
C5, which includes cases characterized by 18q chromosomal gains and co-occurring MYD88 and CD79B mutations, is typically linked to poor prognosis.
In this study, the prognostic relevance of various genetic alterations was also examined. Notably, gain of 13q31.3—which includes the miR-17-92 cluster—was identified as an independent predictor of worse progression-free survival (PFS), highlighting the potential role of miRNA dysregulation in the pathogenesis and progression of DLBCL [12]. The recently developed classification systems for DLBCL, which define more precise subtypes based on genetic, molecular, and immunophenotypic profiles, are designed to better reflect the disease’s complexity and to support a more tailored and effective therapeutic approach. Nonetheless, these classifications rely on advanced molecular methodologies that are not yet routinely available in clinical settings or still require additional validation for widespread clinical use [13,14].
To better stratify DLBCL patients into defined risk groups and more precisely predict clinical outcomes, several prognostic scoring systems have been introduced. The most widely used among them is the International Prognostic Index (IPI), although it was developed prior to the introduction of rituximab. This index considers factors such as patient age, serum lactate dehydrogenase (LDH) levels, Ann Arbor stage, ECOG performance status, and the number of disease sites. Based on these parameters, patients are classified into four risk categories—low, low-intermediate, high-intermediate, and high—with corresponding 3-year OS estimates ranging from 91% in the low-risk group to 59% in the high-risk group [15]. Following the integration of rituximab into standard treatment protocols, the revised International Prognostic Index (R-IPI) was introduced. While it retained the original risk factors, it simplified patient stratification into three prognostic groups: very good (no risk factors), good (one to two risk factors), and poor (three to four risk factors) [16]. Additionally, the NCCN-IPI scoring system was developed specifically for newly diagnosed DLBCL patients intended to receive R-CHOP therapy [17]. The NCCN-IPI was developed using data from the National Comprehensive Cancer Network database and was largely based on the same adverse prognostic factors as earlier scoring systems. However, it introduced more refined risk stratification, categorizing patients once again into four risk groups: low, low-intermediate, high-intermediate, and high. Among the three scoring models, the NCCN-IPI appears to offer the greatest predictive accuracy for both PFS and OS. Nonetheless, none of these tools have been able to clearly identify a subgroup of patients with very poor prognosis, highlighting a continuing unmet need in the risk stratification of DLBCL [18,19]. A more recent prognostic tool for DLBCL patients, known as the International Metabolic Prognostic Index, has been introduced. This model incorporates patient age, disease stage, and baseline metabolic tumor volume (MTV), as assessed through positron emission tomography (PET) imaging [20].
There is a clear need to improve prognostic evaluation in patients with DLBCL, which may be addressed through the identification and validation of novel biomarkers by multiple research groups. This can be achieved by exploring the use of novel biomarkers from multiple research groups. Among the emerging biomarkers, circulating tumor DNA (ctDNA) has gained considerable attention, especially after early studies in patients with non-hematologic solid tumors. ctDNA consists of fragmented DNA—ranging in size from approximately 70 base pairs to 21 kilobases—that is released into the bloodstream by malignant cells through processes such as apoptosis, necrosis, or active secretion [21]. Multiple studies have highlighted the potential of ctDNA as a non-invasive biomarker for predicting prognosis, tracking treatment response, and enabling early detection of relapse in patients with DLBCL [22]. A systematic review examining the role of ctDNA as a "liquid biopsy" in DLBCL concluded that IGH gene rearrangements and somatic mutations identified in ctDNA currently represent the most promising biomarkers for evaluating treatment response. However, the prognostic significance of ctDNA concentration and its methylation status remains uncertain and warrants further investigation [23]. Remarkably, several clinical trials of novel treatment agents, such as polatuzumab vedotin or odronextamab, have used ctDNA as a biomarker of molecular response. Studies have shown that patients with undetectable ctDNA levels during treatment or upon its completion generally experience longer survival. Therefore, ctDNA could potentially support clinical decision-making by guiding treatment escalation or de-escalation at specific time points [24].
MiRNAs are part of a large family of naturally occurring, endogenous, single-stranded RNAs, approximately 22 nucleotides in length. They regulate gene expression at the post-transcriptional level by binding to target RNAs in a sequence-specific manner [25,26]. Victor Ambros and Gary Ruvkun were awarded the Nobel Prize in Physiology or Medicine in 2024 for their pioneering discovery and characterization of miRNAs, first identified in the roundworm Caenorhabditis elegans [27,28,29]. Since their discovery, it is estimated that miRNAs regulate nearly two-thirds of all human genes [30]. miRNAs participate in the regulation of numerous biological processes [31], such as cell proliferation, differentiation, development, and programmed cell death (apoptosis)[32]. Their dysregulation has been linked to various human diseases [33], including diabetes, cardiovascular conditions, and cancer [33,34]. They are considered promising biomarkers, as they can be detected in both tumor tissues and peripheral blood, and they remain stable during sample processing and storage [35]. This review provides an overview of miRNA biogenesis and focuses on the role of miRNAs in the pathogenesis of DLBCL. Additionally, it discusses their potential utility as biomarkers in patients with DLBCL.
2. Materials/Methods
This narrative review was conducted to synthesize current evidence regarding the role of miRNAs as biomarkers in DLBCL. A comprehensive literature search was performed using PubMed, Google Scholar, and Scopus, covering the period from January 1, 2010, to August 1, 2025. The search strategy incorporated keywords such as “DLBCL,” “lymphoma,” “miRNAs,” and “non-coding RNAs”, encompassing all types of relevant publications. The review included peer-reviewed primary studies, reviews, and meta-analyses; human studies focused on DLBCL; and articles addressing the diagnostic, prognostic, or therapeutic relevance of miRNAs. Only English-language articles were considered. Reference lists of selected papers were also manually screened to identify additional pertinent studies. Given the narrative nature of this review, a formal risk-of-bias assessment was not performed [36].
3. miRNAs Biogenesis
MiRNA biogenesis begins with the processing of transcripts generated by RNA polymerase II or III, processed either during transcription or shortly after it [37]. Approximately half of all identified miRNAs are located within genes (intragenic), primarily derived from introns and, to a lesser extent, from exons of protein-coding genes. The rest are located between genes (intergenic), transcribed independently from their own promoters and not associated with host genes [38,39]. The primary and most common route for miRNA production is the canonical biogenesis pathway. In this process, primary miRNA transcripts (pri-miRNAs) are first produced from miRNA genes and then cleaved into precursor miRNAs (pre-miRNAs) by the microprocessor complex. This complex is made up of the RNA-binding protein DGCR8 (DiGeorge Syndrome Critical Region 8) and the RNase III enzyme Drosha [40]. DGCR8 identifies specific sequence motifs within the pri-miRNA, such as the N6-methyladenylated GGAC motif [41]. Drosha, then, cuts the pri-miRNA at the base of its typical hairpin structure, generating a precursor miRNA (pre-miRNA) with a two-nucleotide 3′ overhang [42]. After pre-miRNAs are produced, they are transported from the nucleus to the cytoplasm by the exportin 5 (XPO5)/RanGTP complex. Once in the cytoplasm, the RNase III enzyme Dicer processes them further by removing the terminal loop, producing a mature miRNA duplex [40,43]. The strand direction defines the naming of the mature miRNA: the 5p strand comes from the 5′ end of the pre-miRNA hairpin, while the 3p strand comes from the 3′ end [44]. Both strands can be incorporated into Argonaute (AGO) proteins (AGO1–4 in humans) through an ATP-dependent mechanism [45]. The miRNA duplex is subsequently incorporated into an Argonaute protein, which facilitates the formation of the RNA-induced silencing complex (RISC), a ribonucleoprotein complex. RISC is mainly composed of the transactivation-responsive RNA-binding protein (TRBP) and Argonaute 2 (Ago2)[46].
The RISC complex helps locate target mRNAs by binding to sequences that are complementary, typically within the 3′ untranslated region (3′ UTR) of the mRNA. In humans, perfect base pairing is not required for this interaction to suppress gene expression, which may occur through mRNA degradation, destabilization, or inhibition of translation [47]. The biosynthesis of miRNAs is depicted in Figure 1. Because pri- or pre-miRNAs can be imprecisely cleaved at either the 5′ or 3′ end, a single miRNA gene may give rise to multiple isoforms, known as isomiRs. These different isomiRs can target various genes and signaling pathways, even when originating from the same miRNA. Moreover, certain isomiRs are specific to particular cancer types, highlighting their potential as promising biomarkers for future research and diagnostics [48].
Created in https://BioRender.com (Accessed on 8 July 2025). Modified from Seyhan, Attila 2023 [49].
4. miRNAs and Their Role in DLBCL Pathogenesis and Prognosis
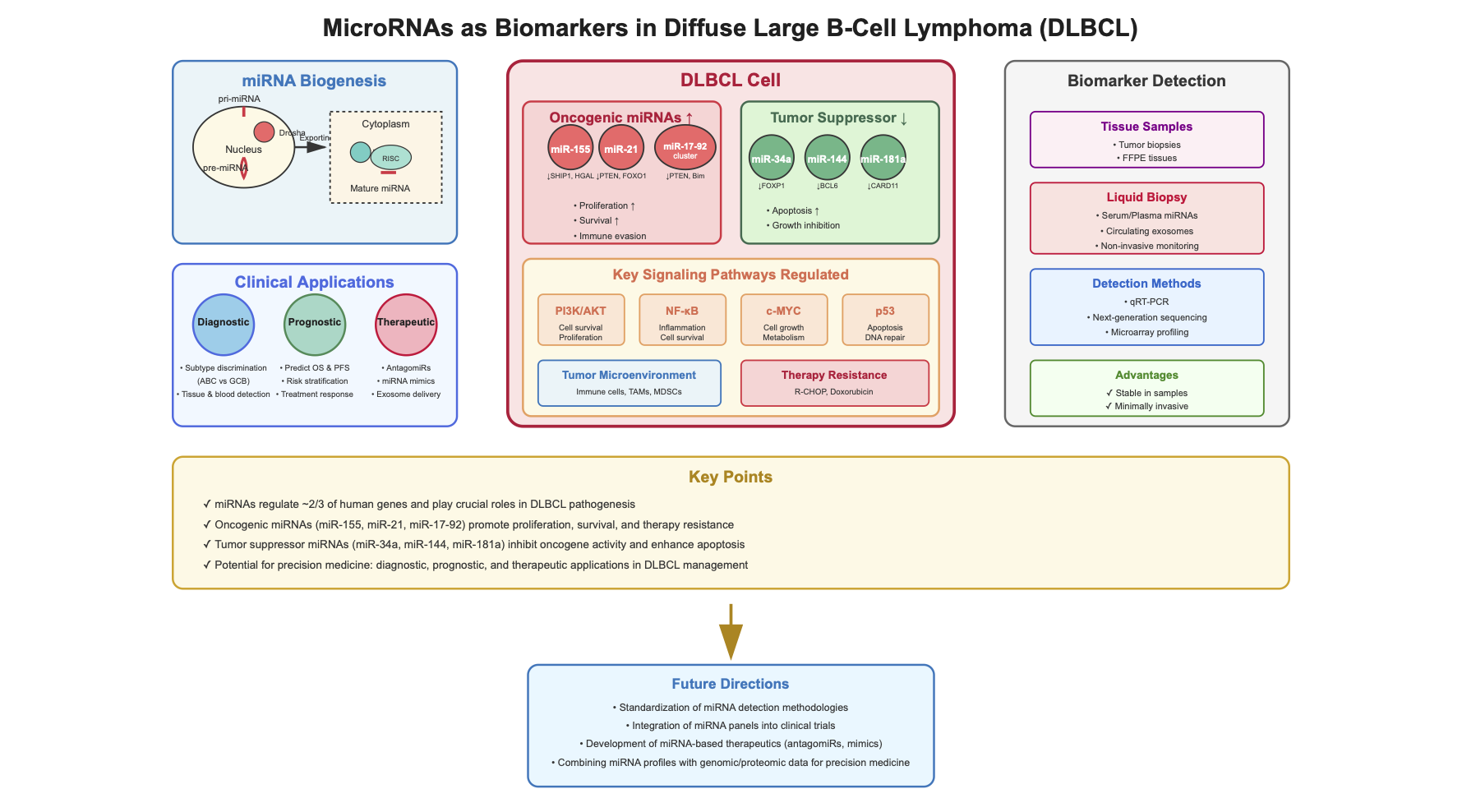
MiRNAs are key regulators in the development of DLBCL and influence how the disease responds to treatment [50]. Several miRNAs, such as miR-155, miR-21, and the miR-17-92 cluster, act as oncogenes by suppressing tumor-suppressor genes, thereby enhancing cell growth, survival, and the ability to evade the immune system. In contrast, miRNAs like miR-34a, miR-144, and miR-181a serve as tumor suppressors by downregulating oncogenes such as SIRT1, BCL6, and CARD11[50,51]. Presented below is an overview of each miRNA and its association with the pathogenesis of DLBCL.
4.1. miR-155
MiR-155 is a well-characterized oncomiR encoded by the B-cell integration cluster (BIC) gene located on chromosome 21 [52]. Initially discovered through its activation by avian leukosis virus insertion in B-cell lymphomas, miR-155 has since been recognized as a critical regulator of immune function, hematopoiesis, and inflammation [53]. MiR-155 has been shown to influence the differentiation of T helper cells and the germinal center (GC) response, helping to generate an effective antibody response dependent on T cells, partly by controlling the production of various cytokines [54]. MiR-155 regulates the development of myeloid cells and the production of inflammatory cytokines by targeting SOCS1 and SHIP1, both of which are negative regulators of the PI3K/Akt signaling pathway. Its abnormal expression has been linked to various hematologic and solid tumors, which is why it is classified as an oncomiR [55].
The cancer-promoting role of miR-155 was demonstrated in Eµ-miR-155 transgenic mice, which developed aggressive pre-B-cell tumors within 3 to 4 weeks. In this model, miR-155 was also shown to promote B-cell malignancies by reducing the expression of SHIP1 and C/EBPβ proteins [56]. SHIP1 and CEBPβ are key regulators within the IL-6 signaling pathway. When miR-155 is upregulated, it suppresses the expression of these genes, leading to impaired B-cell differentiation and enhanced cell survival. This survival advantage is linked to the activation of the PI3K/Akt and MAPK signaling pathways [56,57]. PI3K inhibitors have been tested in clinical trials for patients with relapsed or refractory DLBCL, and their efficacy may be greater in cases where miR-155 is highly expressed [58]. In addition to its other targets, miR-155 also downregulates the Human Germinal-center Associated Lymphoma (HGAL) gene. HGAL is specifically expressed in GC B-cells and plays a role in reducing lymphocyte and lymphoma cell movement by activating the RhoA signaling pathway and interacting with actin and myosin [59,60]. In patients with DLBCL, HGAL expression has been linked to improved OS, independent of the International Prognostic Index (IPI) score [59]. When miR-155 is overexpressed, it suppresses HGAL, which may contribute to increased lymphoma cell spread and tumor aggressiveness [60].
MiR-155 also functions as a regulator of immune responses [61]. Exosomes—small lipid-based extracellular vesicles—are believed to play a significant role in cell-to-cell communication [62]. These vesicles can carry miRNAs, such as miR-155, and facilitate their transfer between cells [62,63]. Dendritic cells (DCs) release exosomes containing miR-155, which can influence and modulate the inflammatory response [64]. The anti-tumor function of miR-155 in DC activity has been supported by both in vitro [64] and in vivo [65] studies. These studies show that exosomes enriched with miR-155 boost DC-driven cytokine production and promote greater tumor infiltration by cytotoxic and helper T cells, while simultaneously decreasing the presence of regulatory T cells [64,65]. The spread of colorectal cancer has been associated with miR-155-containing exosomes that facilitate communication between cancer cells and cancer-associated fibroblasts (CAFs) [66].
Cell-free RNAs, including miRNAs and circRNAs, have shown potential as biomarkers for precision medicine approaches in DLBCL [67]. Among them, miR-155 stands out as a promising candidate for diagnostic, prognostic, and predictive use in this lymphoma subtype [35]. As a candidate biomarker, the expression of miR-155 has been studied both in tissues and in peripheral blood.
Research investigating the role of miR-155 as a diagnostic biomarker in both tissue and blood samples has produced mixed findings [68,69,70]. Several studies have reported elevated miR-155 expression in tumor tissues and the serum of DLBCL patients compared to non-cancerous lymph nodes, normal peripheral B-cells, and the serum/plasma or exosomal vesicles from healthy individuals [71,72,73]. However, other studies have not found any significant differences in miR-155 levels between DLBCL patients and controls [74,75]. Notably, serum miR-155 levels have been shown to correlate positively with its expression in tumor tissue, suggesting its potential as a minimally invasive blood-based biomarker [76]. Interestingly, miR-155 levels in serum may vary depending on the disease phase—fluctuating at diagnosis, during treatment, or in the event of disease progression [77]. When comparing miRNA profiles between patients under treatment and those with progressive disease, a general decline in miRNA levels has been observed [77].
Consensus exists regarding the diagnostic utility of miR-155 in distinguishing between the ABC and GCB subtypes of DLBCL. Several studies have reported that miR-155 expression levels—whether in tissue or serum—are elevated in patients with the non-GCB/ABC subtype compared to those with the GCB subtype [70,71,72,74,76], likely due to miR-155 targeting the HGAL gene as previously mentioned [60]. Moreover, miR-155 expression appears to differ significantly between de novo DLBCL and transformed aggressive B-cell lymphoma [78], with higher levels observed in de novo cases. However, a separate study examining the transformation of FL to DLBCL identified changes in the expression of other miRNAs (including miR-223, 217, 222, 221, and let-7i/7b) [71]. Notably, miR-155 was not among the miRNAs found to be differentially expressed between de novo and transformed DLBCL in that study [71]. As a result, miR-155 does not appear to be a reliable marker for distinguishing FL from DLBCL or for predicting FL transformation into DLBCL [71].
The role of miR-155 as a prognostic biomarker in DLBCL remains a subject of ongoing debate. One study analyzing 90 tissue samples from patients with de novo DLBCL found that lower miR-155 expression was linked to improved 5-year PFS [61]. Similarly, two other studies reported that elevated miR-155 levels were associated with poorer OS [79]. In a cohort of 118 patients, real-time PCR analysis stratified cases by median miR-155 expression into high and low miR-155 expression, revealed significantly shorter survival in those with higher expression [80]. Contrarily, another study found that in patients with the GCB subtype of DLBCL treated with R-CHOP, high miR-155 levels correlated with more favorable outcomes, independent of IPI score [81]. miR-155 has also been investigated in peripheral blood as a potential prognostic indicator. While one study concluded that serum levels of miR-155 lacked prognostic value [71], later research suggested that elevated circulating miR-155 was independently associated with worse prognosis, regardless of IPI classification [75,82]. As with its diagnostic utility, further prospective, randomized studies are needed to clarify the prognostic significance of miR-155 in DLBCL. These discrepancies underscore the need for standardized quantification methods and large prospective validation studies [83]. Overall, miR-155 represents a promising, but currently inconclusive biomarker in DLBCL, with relevance in disease pathogenesis, subtype differentiation, and potentially treatment response prediction.
4.2. miR-21
MiR-21 is one of the most prevalent and evolutionarily conserved miRNAs. It is expressed in nearly all cell types and has key regulatory roles in both normal physiology and pathological conditions [84]. In DLBCL, miR-21 is consistently overexpressed in tumor tissues, serum, and cell lines [85,86,87,88]. It is considered a canonical oncomiR, implicated in promoting tumor progression through suppression of tumor suppressor genes.
Several studies have elucidated the oncogenic role of miR-21 in DLBCL via the PI3K/AKT pathway, primarily by targeting FOXO1 and PTEN. PTEN functions as a tumor suppressor by negatively regulating the PI3K/AKT signaling cascade and is a well-established direct target of miR-21 [89]. FOXO1, a transcription factor, modulates the expression of genes such as p27, p21, FasL, and Bim, all of which contribute to cell cycle arrest and apoptosis [90]. Upon activation, AKT phosphorylates FOXO1, leading to its cytoplasmic translocation and degradation by the proteasome [91]. To address this, Go et al. screened the expression levels of miR-21, FOXO1 and PTEN in a variety of DLBCL cell lines [92]. Immunohistochemical analysis of human DLBCL tissues revealed FOXO1 expression in 80.8% (126 out of 156) of the cases, displaying varying staining intensities and patterns that were associated with miR-21 levels. Specifically, low miR-21 expression was linked to increased nuclear localization of FOXO1, while high miR-21 levels were commonly associated with FOXO1 localization in the cytoplasm. Inhibition of miR-21 led to increased luciferase activity in reporter constructs containing FOXO1-binding sites from the Bim promoter region, indicating that FOXO1 enhances Bim transcription. Supporting this, the use of the transcriptional inhibitor actinomycin D blocked the upregulation of Bim even after miR-21 inhibition [92]. In human DLBCL tissues, Bim expression tended to show an inverse association with miR-21 levels. Specifically, Bim mRNA levels were lower in samples with high miR-21 expression compared to those with low miR-21. Immunohistochemical analysis categorized Bim expression as low (none to mild) or high (moderate to strong) based on staining intensity. Among patients with high miR-21, low Bim expression was more commonly observed. Conversely, high Bim expression appeared more frequently in the low miR-21 group, although this trend did not reach statistical significance [92].
Recent studies have established that miRNAs are critical regulators of tumorigenesis, representing an additional layer of post-transcriptional gene regulation. Their expression profiles are often strongly associated with specific clinical and pathological features of cancer, enabling their use as biomarkers for distinguishing malignant from non-malignant tissues and for predicting patient outcomes [93]. Among them, miR-21 has been implicated in tumor progression by downregulating tumor suppressor genes such as PDCD4, PTEN, and TPM1 [94]. Wang et al. reported that miR-21 expression was significantly elevated in hepatocellular carcinoma tissues compared to adjacent non-tumorous liver tissues [95], while markedly increased levels of miR-21 have also been detected in the cerebrospinal fluid of patients with primary central nervous system lymphoma compared to healthy controls [96]. In the context of DLBCL, miR-21 expression is upregulated both in serum and in tumor tissue [71]. Additionally, another study reported upregulated miR-21 expression in various DLBCL cell lines, including OCI-Ly1, OCI-Ly3, OCI-Ly4, OCI-Ly7, OCI-Ly8, OCI-Ly10, OCI-Ly18, OCI-Ly19, and HBL [88]. Patients with elevated miR-21 expression levels showed poorer OS compared to those with lower expression. Analysis using a Cox proportional hazards model, which accounted for other survival-related factors in DLBCL, identified miR-21 overexpression as an independent prognostic indicator. These findings suggest that miR-21 plays a significant role in the development and progression of DLBCL. Similarly, Mao et al. reported that serum miR-21 serves as a strong and independent predictor of OS in patients with primary central nervous system lymphoma [97]. Go et al. found that elevated miR-21 expression was significantly linked to shorter progression-free and overall survival in DLBCL patients receiving rituximab-based chemotherapy. miR-21 was also identified as an independent prognostic factor in this setting [92]. Similarly, MG Narducci et al. reported that miR-21 was upregulated in cutaneous T-cell lymphoma and could distinguish between patients with poor and favorable prognosis [98].
4.3. miR-34
The miR-34 family, which consists of miR-34a and the miR-34b/c cluster, targets the tumor suppressor gene p53. Additionally, the loss of miR-34 expression has been associated with resistance to apoptosis induced by p53-activating agents used in chemotherapy [99,100]. Moreover, knockdown of C-MYC was shown to elevate miR-34a expression levels, reduce forkhead box P1 (Foxp1) expression, and induce apoptosis in DLBCL cells [101]. miR-34 also modulates PI3K/AKT pathway which regulates cell survival and growth. In other words, MiR-34s can suppress cancer cell proliferation and induce cell death by inhibiting genes involved in the PI3K/AKT signaling pathway [102].
In chronic lymphocytic leukemia, altered miR-125a-5p and miR-34a-5p expression may predict Richter transformation. Especially, overexpression of miR-125a-5p or low expression of miR -34a-5p can predict almost 50% of Richter Syndrome 0.5 to 5 years before it occurs and their prediction value can guide clinical doctors to select the most appropriate therapeutic strategy considering the potential of transformation of the CLL into a more aggressive disease [103]. In a study in DLBCL cases, investigators found that the co-occurrence of TP53 mutations and miR-34a methylation was associated with a significantly poor prognosis, with a median survival of 9.4 months (P<0.0001), whereas single alterations in TP53 or miR-34a/b/c promoter methylation did not impact survival [104]. The expression level of miR-34a was significantly reduced in Mucosa-Associated Lymphoid Tissue (MALT) lymphomas and DLBCLs compared to normal gastric tissues and peripheral blood mononuclear cells. Furthermore, low levels of miR-34a along with increased FOXP1, p53, and BCL2 coexpression were indicators of poor prognosis [105]. On the other hand, overexpression of miR-34a was associated with better OS and greater sensitivity to doxorubicin in DLBCL [106].
The ability of miR-34s to regulate gene expression and control essential processes such as cell proliferation, apoptosis, and metastasis has also made them promising candidates for targeted cancer therapies, including lymphomas [102]. In conclusion, miR-34a is a key tumor suppressor in DLBCL with well-defined molecular targets and clinical associations. Its loss contributes to disease progression and poor prognosis, while its restoration shows promise in therapeutic strategies. Further studies are warranted to validate its prognostic utility and optimize delivery of miR-34a–based treatments in clinical settings.
4.4. miR-17-92
miR-17-92 is a polycistronic miRNA cluster (consisting of miR-17, miR-18a, miR-19a, miR-19b, miR-20a, and miR-92a) that often is overexpressed in particular solid and lymphoid malignancies. The miR-17-92 miRNA cluster targets the tumor suppressor PTEN and the proapoptotic protein Bim by inhibiting their expression [107]. The miR-17-92 cluster is also known as “oncomiR-1” and plays a crucial role in the cell cycle, proliferation, apoptosis, and other essential cellular processes [108]. The transcription factor MYC activates the expression of miR-17-92 by directly binding to its genomic locus [109]. Moreover, the miR-17-92 cluster modulates the expression of E2F1 or both E2F2 and E2F3 (transcription factors that play a crucial role in cell cycle) [108].
Among the molecular subtypes of DLBCL identified through gene-expression profiling, GC-DLBCL is characterized by elevated levels of miR-17-92 and frequent amplification of the 13q31.3 region, where the miR-17-92 locus resides [109,110]. Expression profiling of the miR-17-92 cluster may help distinguish GC-DLBCL from high-grade FL (overexpressed in DLBCL) [111]. The prognostic value of the miR17-92 cluster is under investigation. Overexpression of miR-18a was linked to shorter OS in DLBCL patients treated with R-CHOP [112]. In addition, investigators have found that overexpression of miR-18, miR-19a, and miR-92a was associated with a significant reduction in OS in patients with B-NHL, including patients with DLBCL , while elevated levels of miR-19a and miR-92a were linked to decreased event-free survival (EFS) [113].
4.5. Other miRNAs of Importance in DLBCL
Beyond these miRNAs, several others have been implicated in DLBCL. More specifically, miR-142 is the only human miRNA gene found to be mutated in about 20% of DLBCL cases. These mutations often occur in the seed sequences of both miR-142-3p and miR-142-5p, which are critical for target recognition and gene regulation [114]. Known and newly identified targets of miR-142 include CFL2, CLIC4, STAU1, TWF1, AKT1S1, CCNB1 (Cyclin B1), LIMA1, and TFRC. These targets are involved in cell cycle regulation, cytoskeletal organization, and iron metabolism, all of which can influence tumor growth and behavior [115]. Knockout of miR-142 in DLBCL cell lines leads to significant changes in the proteome, with upregulation of proteins that promote tumorigenesis and downregulation of proteins necessary for immune response, such as those involved in MHC-I presentation [115]. Both miR-142-3p and miR-142-5p are highly expressed in DLBCL, but mutations can lead to loss of function and altered regulation of cellular pathways [68,114,116].
Another miRNA of interest is miR-22. MiR-22 has emerged as a dynamic regulator of therapy response in DLBCL. Its expression and secretion are modulated in response to R-CHOP chemotherapy, with post-treatment upregulation potentially reflecting alterations in tumor biology or treatment efficacy [117]. Mechanistically, miR-22 regulates key signaling pathways, including p53 signaling, by targeting cell cycle regulators such as CDK6 and CDKN1A (p21), thereby influencing both cell cycle arrest and apoptosis [117,118]. In addition, miR-22 modulates the c-MYC/MYCBP axis and the PTEN-AKT pathway, highlighting its broader role in cell survival and proliferation control. These findings suggest that miR-22 may serve as both a biomarker of treatment response and a candidate for targeted therapy, particularly in molecular subtypes of DLBCL characterized by high miR-22 expression [118].
In DLBCL, miR-181a functions as a tumor suppressor, especially in the ABC subtype. Its expression is reduced in ABC-like cases compared to the GCB subtype, and reintroduction of miR-181a suppresses tumor cell growth and enhances cell death [119]. miR-181a suppresses several key elements of the NF-κB signaling pathway—such as CARD11, IKBα, p105/p50, and C-Rel—resulting in decreased NF-κB activity [120]. This reduction is critical, as NF-κB signaling supports the survival and proliferation of ABC-like DLBCL cells. Higher miR-181a expression is associated with improved survival and better progression-free survival in DLBCL patients treated with R-CHOP, making it a potential prognostic biomarker [121]. These findings highlight miR-181a as a master regulator of NF-κB signaling and a promising biomarker and therapeutic target in DLBCL, especially for the ABC-like subtype [122].
Also, miR-144 acts as a tumor suppressor in DLBCL. Its expression is significantly downregulated in DLBCL tissues and cell lines compared to normal controls, and this downregulation is associated with increased tumor proliferation and invasiveness. miR-144 targets the BCL6 gene, an essential transcriptional repressor that promotes DLBCL progression [123]. Through binding to the 3′ untranslated region of BCL6 mRNA, miR-144 reduces BCL6 expression, leading to decreased proliferation and invasiveness of DLBCL cells. Although miR-144 is downregulated in DLBCL, its plasma levels could contribute to a biomarker panel for monitoring lymphoma [122]. However, its standalone diagnostic utility in DLBCL remains to be confirmed through further studies. These data underscore miR-144 as a key regulator of BCL6 and a potential target for therapeutic intervention in DLBCL [122,123].
Finally, miR-124-3p acts as a tumor suppressor in DLBCL. Its expression is significantly downregulated in DLBCL cells, while its target, NFATc1, is upregulated [124]. In DLBCL cells, miR-124-3p overexpression suppresses cell proliferation and induces apoptosis both in vitro and in vivo. These effects occur via inhibition of the NFATc1/c-MYC pathway, a key driver of tumor growth and survival [124]. The ability of miR-124-3p to inhibit DLBCL progression via the NFATc1/cMYC pathway suggests it could be a promising therapeutic target or adjunct in DLBCL treatment strategies [125]. These results identify miR-124-3p as a strong tumor suppressor in DLBCL, with important mechanistic and therapeutic implications through its control of the NFATc1/c-MYC signaling pathway.
A summary of miRNAs associated with DLBCL is presented below.

Table 1.
Key DLBCL-related miRNAs and their primary functions summarized.
| miRNAs | Function | Reference |
|---|---|---|
| miR-155-5p | Oncogenic; regulates NF-κB pathway, promotes proliferation, useful for diagnosis and classification | [56,57,58] |
| miR-21-5p | Oncogenic; regulates PI3K/AKT pathway, promotes cell survival, useful for diagnosis | [44,90,95] |
| miR-17-92 | Oncogenic; alters MYC, SHIP expression, promotes tumorigenesis | [122] |
| miR-34a | Tumor suppressor; targets SIRT1, inhibits tumor growth | [51] |
| miR-144 | Tumor suppressor; targets BCL6, inhibits tumor growth | [51] |
| miR-181a | Tumor suppressor; targets CARD11, inhibits tumor growth | [51] |
| miR-124-3p | Tumor suppressor; inhibits NFATc1/cMYC pathway, suppresses proliferation, promotes apoptosis | [91,124] |
| miR-142-5p | Promotes immunosuppressive macrophage phenotype, associated with poor response to immunochemotherapy | [126] |
5. miRNAs and Their Role in Tumor Microenvironment in DLBCL
The tumor microenvironment (TME) is a critical determinant of DLBCL biology and clinical behavior, influencing immune escape, therapy response, and progression. It comprises a complex network of cellular and noncellular components that surround and interact with the tumor [127]. In summary, miRNAs are central to DLBCL pathogenesis through their regulation of oncogenic pathways and impact on tumor microenvironment. MiR-155 drives tumor progression and shapes the tumor microenvironment [61].
Moreover, in response to danger signals—such as pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs) like IFN-β—macrophages and dendritic cells (DCs) exhibit increased expression of miR-155. This upregulation also influences the behavior of myeloid-derived suppressor cells (MDSCs), a group of immature myeloid cells known for their potent immunosuppressive functions [61,128]. Importantly, the accumulation of MDSCs within the tumor microenvironment is commonly linked to poor clinical outcomes [129]. Furthermore, high miR-155 levels are linked to reduced peripheral CD8+ T cells and impaired T-cell receptor signaling. miR-155 also increases PD-L1 expression on lymphoma cells, promoting immune evasion by inhibiting CD8+ T cell function through the PD-1/PD-L1 pathway. Targeting this axis with PD-L1 blockade can restore CD8+ T cell activity and suppress tumor growth, especially in EBV-associated DLBCL [75]. miRNAs can also regulate cytokine production and cell migration within the TME, as seen with miR-155 targeting DEPTOR, affecting pro-inflammatory cytokine expression and DLBCL cell motility [130].
MiR-21 plays a central role in driving disease progression in B-cell lymphoma [131] with experimental studies demonstarting that its overexpression induces a pre-B-cell malignant lymphoid-like phenotype [132]. Clinically, elevated levels of circulating miR-21 in the serum of DLBCL patients correlate with tumor tissue expression, more advanced disease stages, and poorer OS outcomes [88,133]. Dysregulated tumor microenvironment determines cancer cell chemosensitivity [134]. Zheng et al. demonstrated that forced expression of miR-21 induced chemoresistance in B-lymphoma cells, an effect that was even more pronounced when these cells were co-cultured with immune and endothelial cells—key components of the tumor microenvironment. This aligns with earlier studies showing that miR-21 promotes adhesion of myeloma cells to bone marrow stromal cells and contributes to resistance against chemotherapy [135]. Additionally, miR-21 has been shown to activate inflammatory pathways in HER2-positive breast cancer, thereby diminishing the efficacy of neoadjuvant trastuzumab and chemotherapy [136].
Other miRNAs orchestrate complex interactions within the DLBCL TME, influencing immune cell function, tumor progression, and response to therapy. miR-142-5p is upregulated in immunosuppressive tumor-associated macrophages (TAMs) in DLBCL. Inhibiting miR-142-5p can repolarize these macrophages to a more anti-tumor phenotype, enhancing antibody-dependent cellular phagocytosis and improving immunotherapy outcomes. High miR-142-5p and CD206+ TAMs predict poor response to immunochemotherapy [126]. A prognostic model incorporating four circulating miRNAs (miR-21, miR-130b, miR-155, and miR-28) has been associated with elevated levels of myeloid-derived suppressor cells (MDSCs) and Th17 cells within the TME, promoting immunosuppression and leading to poorer clinical outcomes. These miRNAs are involved in modulating oncogenic pathways, such as Ras signaling, as well as shaping the immune cell landscape in DLBCL [137]. Overall, modulating specific miRNAs in the TME, such as inhibiting miR-155 or miR-142-5p, offers promising strategies to enhance immune responses and improve the efficacy of immunotherapies in DLBCL [75,126,138].
6. Conclusion
DLBCL remains a biologically and clinically heterogeneous disease, with outcomes varying widely despite advances in immunochemotherapy. Mounting evidence shows that miRNAs are key regulators of DLBCL initiation, progression, and treatment response. These small non-coding RNAs modulate apoptosis, proliferation, immune evasion, and drug resistance through post-transcriptional regulation of pivotal signaling pathways.
Among them, miR-21 and miR-155 are the most extensively characterized, while miR-181a, miR-124-3p, miR-144, miR-142, and miR-22 also exhibit strong mechanistic and prognostic relevance.
Diagnostic studies have shown that distinct miRNA expression profiles can discriminate between DLBCL subtypes, differentiate malignant from non-malignant lymphoid tissues, and even detect disease through peripheral blood samples. Nevertheless, inconsistencies—such as the variable diagnostic sensitivity and specificity of miR-155—underscore the need for methodological standardization in sample handling, detection platforms, and data interpretation.
From a prognostic and predictive perspective, miRNAs hold promise in refining risk stratification, guiding treatment selection, and monitoring therapeutic response. Elevated miR-21 and miR-155 levels have been associated with treatment resistance or differential sensitivity to agents such as vincristine, highlighting their potential as biomarkers for personalized medicine. Mechanistic studies further reveal that several miRNAs directly regulate core signaling cascades, including PI3K/AKT, NF-κB, c-MYC, and p53, thereby offering both biomarker and therapeutic opportunities.
The expanding understanding of miRNA biology in DLBCL paves the way for innovative therapeutic interventions. Strategies employing antagomiRs, miRNA mimics, and exosome-based delivery systems are under investigation to inhibit oncogenic miRNAs or restore tumor-suppressive ones. However, successful clinical translation will depend on overcoming challenges related to delivery specificity, off-target effects, and the integration of miRNA therapeutics with established treatment modalities.
In summary, miRNAs constitute a powerful class of regulatory molecules with multidimensional roles in DLBCL pathogenesis, diagnosis, prognosis, and therapy. Future research should prioritize the validation of robust miRNA panels, the harmonization of analytical methodologies, and the incorporation of miRNA profiling into prospective clinical trials. Such efforts could ultimately enable precision-guided treatment strategies and novel therapeutics that harness the regulatory potential of miRNAs to improve outcomes for patients with DLBCL. Integrating miRNA signatures with genomic and proteomic data may redefine DLBCL taxonomy, enabling truly personalized therapeutic algorithms.
Author Contributions
Conceptualization, E.H., A.P.-B., P.K. and E.K. (Evangelos Kolettas); writing—original draft preparation, E.P., E.K. (Epameinondas Koumpis), V.G. and G.P.B.; writing—review and editing, E.P., E.K. (Epameinondas Koumpis), V.G., G.P.B., A.P.-B., P.K., E.K. (Evangelos Kolettas) and E.H.; visualization, E.P., E.K. (Epameinondas Koumpis) and E.H.; supervision, E.K. (Evangelos Kolettas), A.P.-B., P.K. and E.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
No acknowledgements.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Teras LR, D.C., Cerhan JR, Morton LM, Jemal A, Flowers CR. US lymphoid malignancy statistics by World Health Organization subtypes. CA Cancer J Clin. 2016, 66, 443–459. [CrossRef]
- Chapuy B, S.C., Dunford AJ, Kim J, Kamburov A, Redd RA, Lawrence MS, Roemer MGM, Li AJ, Ziepert M, Staiger AM, Wala JA, Ducar MD, Leshchiner I, Rheinbay E, Taylor-Weiner A, Coughlin CA, Hess JM, Pedamallu CS, Livitz D, Rosebrock D, Rosenberg M, Tracy AA, Horn H, van Hummelen P, Feldman AL, Link BK, Novak AJ, Cerhan JR, Habermann TM, Siebert R, Rosenwald A, Thorner AR, Meyerson ML, Golub TR, Beroukhim R, Wulf GG, Ott G, Rodig SJ, Monti S, Neuberg DS, Loeffler M, Pfreundschuh M, Trümper L, Getz G, Shipp MA. Molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat. Med. 2018, 679–690. [CrossRef]
- Feugier P, V.H.A., Sebban C, Solal-Celigny P, Bouabdallah R, Fermé C, Christian B, Lepage E, Tilly H, Morschhauser F, Gaulard P, Salles G, Bosly A, Gisselbrecht C, Reyes F, Coiffier B. Long-term results of the R-CHOP study in the treatment of elderly patients with diffuse large B-cell lymphoma: A study by the Groupe d'Etude des Lymphomes de l'Adulte. J Clin Oncol. 2005, 23, 4117–4126. [CrossRef]
- Coiffier B, L.E., Briere J, Herbrecht R, Tilly H, Bouabdallah R, Morel P, Van Den Neste E, Salles G, Gaulard P, Reyes F, Lederlin P, Gisselbrecht C. CHOP chemotherapy plus rituximab compared with CHOP alone in elderly patients with diffuse large-B-cell lymphoma. N Engl J Med 2002, 346, 235–242. [CrossRef] [PubMed]
- Pfreundschuh M, K.E., Trümper L, Osterborg A, Trneny M, Shepherd L, Gill DS, Walewski J, Pettengell R, Jaeger U, Zinzani PL, Shpilberg O, Kvaloy S, de Nully Brown P, Stahel R, Milpied N, López-Guillermo A, Poeschel V, Grass S, Loeffler M, Murawski N; . MabThera International Trial (MInT) Group. CHOP-like chemotherapy with or without rituximab in young patients with good-prognosis diffuse large-B-cell lymphoma: 6-year results of an open-label randomised study of the MabThera International Trial (MInT) Group. Lancet Oncol 2011, 11, 1013–1022. [CrossRef]
- Coiffier B, S.C. Diffuse large B-cell lymphoma: R-CHOP failure-what to do? Hematology Am Soc Hematol Educ Program 2016, 1, 366–378. [CrossRef]
- Tilly, H.; Morschhauser, F.; Sehn, L.H.; Friedberg, J.W.; Trněný, M.; Sharman, J.P.; Herbaux, C.; Burke, J.M.; Matasar, M.; Rai, S.; et al. Polatuzumab Vedotin in Previously Untreated Diffuse Large B-Cell Lymphoma. N Engl J Med 2022, 386, 351–363. [CrossRef]
- Parry EM, R.S., Okosun J. DLBCL arising from indolent lymphomas: How are they different? . Semin Hematol. 2023, 60, 277–284. [CrossRef]
- Rosenwald A, W.G., Chan WC, Connors JM, Campo E, Fisher RI, Gascoyne RD, Muller-Hermelink HK, Smeland EB, Giltnane JM, Hurt EM, Zhao H, Averett L, Yang L, Wilson WH, Jaffe ES, Simon R, Klausner RD, Powell J, Duffey PL, Longo DL, Greiner TC, Weisenburger DD, Sanger WG, Dave BJ, Lynch JC, Vose J, Armitage JO, Montserrat E, López-Guillermo A, Grogan TM, Miller TP, LeBlanc M, Ott G, Kvaloy S, Delabie J, Holte H, Krajci P, Stokke T, Staudt LM;. Lymphoma/Leukemia Molecular Profiling Project. The use of molecular profiling to predict survival after chemotherapy for diffuse large-B-cell lymphoma. N Engl J Med 2002, 346, 1937–1947. [CrossRef]
- Wright GW, H.D., Phelan JD, Coulibaly ZA, Roulland S, Young RM, Wang JQ, Schmitz R, Morin RD, Tang J, Jiang A, Bagaev A, Plotnikova O, Kotlov N, Johnson CA, Wilson WH, Scott DW, Staudt LM. A Probabilistic Classification Tool for Genetic Subtypes of Diffuse Large B Cell Lymphoma with Therapeutic Implications. Cancer Cell 2020, 37, 551–568. [CrossRef]
- Schmitz R, W.G., Huang DW, Johnson CA, Phelan JD, Wang JQ, Roulland S, Kasbekar M, Young RM, Shaffer AL, Hodson DJ, Xiao W, Yu X, Yang Y, Zhao H, Xu W, Liu X, Zhou B, Du W, Chan WC, Jaffe ES, Gascoyne RD, Connors JM, Campo E, Lopez-Guillermo A, Rosenwald A, Ott G, Delabie J, Rimsza LM, Tay Kuang Wei K, Zelenetz AD, Leonard JP, Bartlett NL, Tran B, Shetty J, Zhao Y, Soppet DR, Pittaluga S, Wilson WH, Staudt LM. Genetics and Pathogenesis of Diffuse Large B-Cell Lymphoma. N Engl J Med 2018, 378, 1396–1407. [CrossRef]
- Sehn LH, S.G. Diffuse Large B-Cell Lymphoma. N Engl J Med 2021, 384, 842–858. [CrossRef]
- Alaggio R, A.C., Anagnostopoulos I, Attygalle AD, Araujo IBO, Berti E, Bhagat G, Borges AM, Boyer D, Calaminici M, Chadburn A, Chan JKC, Cheuk W, Chng WJ, Choi JK, Chuang SS, Coupland SE, Czader M, Dave SS, de Jong D, Du MQ, Elenitoba-Johnson KS, Ferry J, Geyer J, Gratzinger D, Guitart J, Gujral S, Harris M, Harrison CJ, Hartmann S, Hochhaus A, Jansen PM, Karube K, Kempf W, Khoury J, Kimura H, Klapper W, Kovach AE, Kumar S, Lazar AJ, Lazzi S, Leoncini L, Leung N, Leventaki V, Li XQ, Lim MS, Liu WP, Louissaint A Jr, Marcogliese A, Medeiros LJ, Michal M, Miranda RN, Mitteldorf C, Montes-Moreno S, Morice W, Nardi V, Naresh KN, Natkunam Y, Ng SB, Oschlies I, Ott G, Parrens M, Pulitzer M, Rajkumar SV, Rawstron AC, Rech K, Rosenwald A, Said J, Sarkozy C, Sayed S, Saygin C, Schuh A, Sewell W, Siebert R, Sohani AR, Tooze R, Traverse-Glehen A, Vega F, Vergier B, Wechalekar AD, Wood B, Xerri L, Xiao W. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Lymphoid Neoplasms. Leukemia 2022, 36, 1720–1748. [CrossRef]
- Campo E, J.E., Cook JR, Quintanilla-Martinez L, Swerdlow SH, Anderson KC, Brousset P, Cerroni L, de Leval L, Dirnhofer S, Dogan A, Feldman AL, Fend F, Friedberg JW, Gaulard P, Ghia P, Horwitz SM, King RL, Salles G, San-Miguel J, Seymour JF, Treon SP, Vose JM, Zucca E, Advani R, Ansell S, Au WY, Barrionuevo C, Bergsagel L, Chan WC, Cohen JI, d'Amore F, Davies A, Falini B, Ghobrial IM, Goodlad JR, Gribben JG, Hsi ED, Kahl BS, Kim WS, Kumar S, LaCasce AS, Laurent C, Lenz G, Leonard JP, Link MP, Lopez-Guillermo A, Mateos MV, Macintyre E, Melnick AM, Morschhauser F, Nakamura S, Narbaitz M, Pavlovsky A, Pileri SA, Piris M, Pro B, Rajkumar V, Rosen ST, Sander B, Sehn L, Shipp MA, Smith SM, Staudt LM, Thieblemont C, Tousseyn T, Wilson WH, Yoshino T, Zinzani PL, Dreyling M, Scott DW, Winter JN, Zelenetz AD. The International Consensus Classification of Mature Lymphoid Neoplasms: A report from the Clinical Advisory Committee. Blood 2022, 140, 1229–1253. [CrossRef]
- Project., I.N.-H.s.L.P.F. A predictive model for aggressive non-Hodgkin's lymphoma. N Engl J Med 1993, 329, 987–994. [CrossRef]
- Sehn LH, B.B., Chhanabhai M, Fitzgerald C, Gill K, Hoskins P, Klasa R, Savage KJ, Shenkier T, Sutherland J, Gascoyne RD, Connors JM. The revised International Prognostic Index (R-IPI) is a better predictor of outcome than the standard IPI for patients with diffuse large B-cell lymphoma treated with R-CHOP. Blood 2007, 109, 1857–1861. [CrossRef]
- Zhou Z, S.L., Rademaker AW, Gordon LI, Lacasce AS, Crosby-Thompson A, Vanderplas A, Zelenetz AD, Abel GA, Rodriguez MA, Nademanee A, Kaminski MS, Czuczman MS, Millenson M, Niland J, Gascoyne RD, Connors JM, Friedberg JW, Winter JN. An enhanced International Prognostic Index (NCCN-IPI) for patients with diffuse large B-cell lymphoma treated in the rituximab era. Blood 2014, 123, 837–842. [CrossRef]
- Jelicic J, J.-J.K., Bukumiric Z, Roost Clausen M, Ludvigsen Al-Mashhadi A, Pedersen RS, Poulsen CB, Brown P, El-Galaly TC, Stauffer Larsen T. Prognostic indices in diffuse large B-cell lymphoma: A population-based comparison and validation study of multiple models. Blood Cancer J. 2023, 13, 157. [CrossRef]
- Ruppert AS, D.J., Salles G, Wall A, Cunningham D, Poeschel V, Haioun C, Tilly H, Ghesquieres H, Ziepert M, Flament J, Flowers C, Shi Q, Schmitz N. International prognostic indices in diffuse large B-cell lymphoma: A comparison of IPI, R-IPI, and NCCN-IPI. Blood 2020, 135, 2041–2048. [CrossRef]
- Mikhaeel NG, H.M., Eertink JJ, de Vet HCW, Boellaard R, Dührsen U, Ceriani L, Schmitz C, Wiegers SE, Hüttmann A, Lugtenburg PJ, Zucca E, Zwezerijnen GJC, Hoekstra OS, Zijlstra JM, Barrington SF. Proposed New Dynamic Prognostic Index for Diffuse Large B-Cell Lymphoma: International Metabolic Prognostic Index. J Clin Oncol. 2022, 40, 2352–2360. [CrossRef]
- Kim H, P.K. Clinical Circulating Tumor DNA Testing for Precision Oncology. Cancer Res Treat 2023, 55, 351–366. [CrossRef]
- Wu FT, L.L., Xu W, Li JY. Circulating tumor DNA: Clinical roles in diffuse large B cell lymphoma. 9. Ann Hematol. 2019, 98, 255–269. [CrossRef]
- Arzuaga-Mendez J, P.-F.E., Lopez-Lopez E, Martin-Guerrero I, García-Ruiz JC, García-Orad A. Cell-free DNA as a biomarker in diffuse large B-cell lymphoma: A systematic review. Crit Rev Oncol Hematol. 2019, 139, 7–15. [CrossRef]
- Tavakkoli M, B.S. 2024 Update: Advances in the risk stratification and management of large B-cell lymphoma. Am J Hematol. 2023, 98, 1791–1805. [CrossRef]
- DP., B. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [CrossRef]
- DP., B. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [CrossRef]
- H., L. MicroRNAs won the Nobel - will they ever be useful as medicines? . Nature 2024. [CrossRef]
- Wightman B, H.I., Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [CrossRef]
- Lee RC, F.R., Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [CrossRef]
- Friedman, R.C.F., K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [CrossRef]
- Chen CZ, L.H. MicroRNAs as regulators of mammalian hematopoiesis. Semin Immunol. 2005, 17, 155–165. [CrossRef]
- Saliminejad K, K.K.H., Soleymani Fard S, Ghaffari SH. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J Cell Physiol. 2019, 234, 5451–5465. [CrossRef] [PubMed]
- Paul P, C.A., Sarkar D, Langthasa M, Rahman M, Bari M, Singha RS, Malakar AK, Chakraborty S. Interplay between miRNAs and human diseases. J Cell Physiol. 2018, 233, 2007–2018. [CrossRef]
- Markopoulos GS, R.E., Tokamani M, Chavdoula E, Hatziapostolou M, Polytarchou C, Marcu KB, Papavassiliou AG, Sandaltzopoulos R, Kolettas E. A step-by-step microRNA guide to cancer development and metastasis. Cell Oncol (Dordr). 2017, 40, 303–339. [CrossRef]
- Due H, S.P., Bødker JS, Schmitz A, Bøgsted M, Johnsen HE, El-Galaly TC, Roug AS, Dybkær K. miR-155 as a Biomarker in B-Cell Malignancies. Biomed Res Int. 2016. [CrossRef]
- Koumpis E, G.V., Papathanasiou K, Papoudou-Bai A, Kanavaros P, Kolettas E, Hatzimichael E. The Role of microRNA-155 as a Biomarker in Diffuse Large B-Cell Lymphoma. Biomedicines 2024, 12, 2658. [CrossRef]
- Ha M, K.V. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014, 15, 509–524. [CrossRef]
- de Rie D, A.I., Alam T, Arner E, Arner P, Ashoor H; et al. An integrated expression atlas of miRNAs and their promoters in human and mouse. Nat Biotechnol. 2017, 35, 872–878. [CrossRef]
- W., H. MicroRNAs: Biomarkers, diagnostics, and therapeutics. Methods Mol Biol. 1617, 57–67. [CrossRef]
- Denli AM, T.B., Plasterk RH, Ketting RF, Hannon GJ. Processing of primary microRNAs by the Microprocessor complex. Nature 2005, 432, 231–235. [CrossRef]
- Alarcon CR, L.H., Goodarzi H, Halberg N, Tavazoie SF. N6- methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [CrossRef]
- Han J, L.Y., Yeom KH, Kim YK, Jin H, Kim VN. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev 2004, 18, 3016–3027. [CrossRef]
- Okada C, Y.E., Lee SJ, Shibata S, Katahira J, Nakagawa A; et al. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [CrossRef]
- Zhang H, K.F., Jaskiewicz L, Westhof E, Filipowicz W. Single processing center models for human Dicer and bacterial RNase III. Cell 2004, 118, 57–68. [CrossRef]
- Yoda M, K.T., Paroo Z, Ye X, Iwasaki S, Liu Q; et al. ATP-dependent human RISC assembly pathways. Nat Struct Mol Biol. 2010, 17, 17–23. [CrossRef]
- Chendrimada TP, G.R., Kumaraswamy E, Norman J, Cooch N, Nishikura K, Shiekhattar R. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005, 436, 740–744. [CrossRef]
- Westholm JO, L.E.M.m.b.v.s.B.N.-d.j.b.E.J. Mirtrons: microRNA biogenesis via splicing. Biochimie 2011, 93, 1897–1904. [CrossRef]
- Telonis AG, M.R., Loher P, Chervoneva I, Londin E, Rigoutsos I. Knowledge about the presence or absence of miRNA isoforms (isomiRs) can successfully discriminate amongst 32 TCGA cancer types. Nucleic Acids Res. 2017, 45, 2973–2985. [CrossRef]
- Seyhan, A. Circulating microRNAs as Potential Biomarkers in Pancreatic Cancer—Advances and Challenges. International Journal of Molecular Sciences. 2023, 24, 13340. [CrossRef]
- Gartenhaus, K.M.-M.a.R.B. Role of microRNA deregulation in the pathogenesis of diffuse large B-cell lymphoma (DLBCL). Leuk Res. 2013, 37. [CrossRef]
- Mohammed Alsaadi, M.Y.K., Mahmood Hassan Dalhat, Salem Bahashwan,; Muhammad Uzair Khan, A.A., Hussein Almehdar and Ishtiaq Qadri Dysregulation of miRNAs in DLBCL: Causative Factor for Pathogenesis, Diagnosis and Prognosis. Diagnostics 2021, 11. [CrossRef]
- Faraoni I, A.F., Cardone J, Bonmassar E. miR-155 gene: A typical multifunctional microRNA. Biochim Biophys Acta 2009, 1792, 497–505. [CrossRef]
- Bondada MS, Y.Y., Nair V. Multifunctional miR-155 Pathway in Avian Oncogenic Virus-Induced Neoplastic Diseases. Non-Coding RNA 2019, 5, 24. [CrossRef]
- Thai TH, C.D., Casola S, Ansel KM, Xiao C, Xue Y, Murphy A, Frendewey D, Valenzuela D, Kutok JL, Schmidt-Supprian M, Rajewsky N, Yancopoulos G, Rao A, Rajewsky K. Regulation of the germinal center response by microRNA-155. Science 2007, 316, 604–608. [CrossRef]
- Mattiske S, S.R., Neilsen PM, Callen DF. The oncogenic role of miR-155 in breast cancer. Cancer Epidemiol Biomarkers Prev. 2012, 21, 1236–1243. [CrossRef]
- Costinean S, S.S., Pedersen IM, Tili E, Trotta R, Perrotti D, Ciarlariello D, Neviani P, Harb J, Kauffman LR, Shidham A, Croce CM. Src homology 2 domain-containing inositol-5-phosphatase and CCAAT enhancer-binding protein beta are targeted by miR-155 in B cells of Emicro-MiR-155 transgenic mice. Blood 2009, 114, 1374–1382. [CrossRef]
- O'Connell RM, C.A., Rao DS, Baltimore D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc NatI Acad Sci USA. 2009, 106, 7113–7118. [CrossRef]
- Lunning M, V.J., Nastoupil L, Fowler N, Burger JA, Wierda WG, Schreeder MT, Siddiqi T, Flowers CR, Cohen JB, Sportelli P, Miskin HP, Weiss MS, O'Brien S. Ublituximab and umbralisib in relapsed/refractory B-cell non-Hodgkin lymphoma and chronic lymphocytic leukemia. Blood 2019, 134, 1811–1820. [CrossRef]
- Lossos IS, A.A., Rajapaksa R, Tibshirani R, Levy R. HGAL is a novel interleukin-4-inducible gene that strongly predicts survival in diffuse large B-cell lymphoma. Blood 2003, 101, 433–440. [CrossRef]
- Dagan LN, J.X., Bhatt S, Cubedo E, Rajewsky K, Lossos IS. miR-155 regulates HGAL expression and increases lymphoma cell motility. Blood 2012, 119, 513–520. [CrossRef]
- Kalkusova K, T.P., Stakheev D, Smrz D. The Role of miR-155 in Antitumor Immunity. Cancers (Basel) 2022, 14, 5414. [CrossRef]
- Bauer, K.M.R., J.L.; O’Connell, R.M. No small matter: Emerging roles for exosomal miRNAs in the immune system. Febs J. 2022, 289, 4021–4037. [CrossRef]
- Alexander M, H.R., Runtsch MC, Kagele DA, Mosbruger TL, Tolmachova T, Seabra MC, Round JL, Ward DM, O'Connell RM. Exosome-delivered microRNAs modulate the inflammatory response to endotoxin. Nat Commun 2015, 6. [CrossRef]
- Hiramoto JS, T.K., Bedolli M, Norton JA, Hirose R. Antitumor immunity induced by dendritic cell-based vaccination is dependent on interferon-gamma and interleukin-12. J Surg Res. 2004, 116, 64–69. [CrossRef]
- Asadirad A, B.K., Hashemi SM; et al. Dendritic cell immunotherapy with miR-155 enriched tumor-derived exosome suppressed cancer growth and induced antitumor immune responses in murine model of colorectal cancer induced by CT26 cell line. International Immunopharmacology 2022, 104, 108493. [CrossRef] [PubMed]
- Wang, D., Wang, X., Song, Y. et al. Exosomal miR-146a-5p and miR-155-5p promote CXCL12/CXCR7-induced metastasis of colorectal cancer by crosstalk with cancer-associated fibroblasts. Cell Death Dis 2022, 13. [CrossRef]
- Decruyenaere P, O.F., Vandesompele J. Circulating RNA biomarkers in diffuse large B-cell lymphoma: A systematic review. Exp Hematol Oncol. 2021, 10, 13. [CrossRef]
- Lopez-Santillan M, L.-E.A., Arzuaga-Mendez J, Lopez-Lopez E, Garcia-Orad A. Circulating miRNAs as biomarkers in diffuse large B-cell lymphoma: A systematic review. Oncotarget 2018, 9, 22850–22861. [CrossRef]
- Huskova H, K.K., Karban J, Vargova J, Vargova K, Dusilkova N, Trneny M, Stopka T. Oncogenic microRNA-155 and its target PU.1: An integrative gene expression study in six of the most prevalent lymphomas. Int J Hematol. 2015, 102, 441–450. [CrossRef] [PubMed]
- Zhong H, X.L., Zhong JH, Xiao F, Liu Q, Huang HH, Chen FY. Clinical and prognostic significance of miR-155 and miR-146a expression levels in formalin-fixed/paraffin-embedded tissue of patients with diffuse large B-cell lymphoma. Exp Ther Med. 2012, 3, 763–770. [CrossRef]
- Lawrie CH, G.S., Dunlop HM, Pushkaran B, Liggins AP, Pulford K, Banham AH, Pezzella F, Boultwood J, Wainscoat JS, Hatton CS, Harris AL. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br J Haematol. 2008, 141, 672–675. [CrossRef]
- Fang C, Z.D., Dong HJ, Zhou ZJ, Wang YH, Liu L, Fan L, Miao KR, Liu P, Xu W, Li JY. Serum microRNAs are promising novel biomarkers for diffuse large B cell lymphoma. Ann Hematol. 2012, 91, 553–559. [CrossRef]
- Yuan, W.X.G., Y.X.; Na, W.N.; Chao, J.; Yang, X. Circulating microRNA-125b and microRNA-130a expression profiles predict chemoresistance to R-CHOP in diffuse large B-cell lymphoma patients. Oncol. Lett 2016, 11, 423–432. [CrossRef]
- Inada, K.O., Y.; Cho, Y.; Saito, H.; Iijima, T.; Hori, M.; Kojima, H. Availability of Circulating MicroRNAs as a Biomarker for Early Diagnosis of Diffuse Large B-Cell Lymphoma. Open J. Blood Dis. 2015, 48–58.
- Zheng Z, S.R., Zhao HJ, Fu D, Zhong HJ, Weng XQ, Qu B, Zhao Y, Wang L, Zhao WL. MiR155 sensitized B-lymphoma cells to anti-PD-L1 antibody via PD-1/PD-L1-mediated lymphoma cell interaction with CD8+T cells. Mol Cancer. 2019, 18, 54. [CrossRef]
- Yuan, W.X.G., Y.X.; Na, W.N.; Chao, J.; Yang, X. Circulating microRNA-125b and microRNA-130a expression profiles predict chemoresistance to R-CHOP in diffuse large B-cell lymphoma patients. Oncol. Lett 2016, 11, 423–432. [CrossRef]
- Beheshti A, S.K., Vanderburg C, Ravi D, McDonald JT, Christie AL, Shigemori K, Jester H, Weinstock DM, Evens AM. Identification of Circulating Serum Multi-MicroRNA Signatures in Human DLBCL Models. Sci Rep. 2019, 9, 17261. [CrossRef]
- Caramuta S, L.L., Ozata DM, Akçakaya P, Georgii-Hemming P, Xie H, Amini RM, Lawrie CH, Enblad G, Larsson C, Berglund M, Lui WO. Role of microRNAs and microRNA machinery in the pathogenesis of diffuse large B-cell lymphoma. Blood Cancer J. 2013, 3, 152. [CrossRef]
- Iqbal J, S.Y., Huang X, Liu Y, Wake L, Liu C, Deffenbacher K, Lachel CM, Wang C, Rohr J, Guo S, Smith LM, Wright G, Bhagavathi S, Dybkaer K, Fu K, Greiner TC, Vose JM, Jaffe E, Rimsza L, Rosenwald A, Ott G, Delabie J, Campo E, Braziel RM, Cook JR, Tubbs RR, Armitage JO, Weisenburger DD, Staudt LM, Gascoyne RD, McKeithan TW, Chan WC. Global microRNA expression profiling uncovers molecular markers for classification and prognosis in aggressive B-cell lymphoma. Blood 2015, 125, 1137–1145. [CrossRef]
- Zhu FQ, Z.L., Tang N, Tang YP, Zhou BP, Li FF, Wu WG, Zeng XB, Peng SS. MicroRNA-155 Downregulation Promotes Cell Cycle Arrest and Apoptosis in Diffuse Large B-Cell Lymphoma. Oncol Res. 2016, 24, 415–427. [CrossRef]
- Due H, S.P., Bødker JS, Schmitz A, Bøgsted M, Johnsen HE, El-Galaly TC, Roug AS, Dybkær K. miR-155 as a Biomarker in B-Cell Malignancies. Biomed Res Int. 2016;2016:9513037. [CrossRef] [PubMed] [PubMed Central]
- Ahmadvand M, E.M., Pashaiefar H, Yaghmaie M, Manoochehrabadi S, Khakpour G, Sheikhsaran F, Montazer Zohour M. Over expression of circulating miR-155 predicts prognosis in diffuse large B-cell lymphoma. Leuk. Res 2018, 70, 45–48. [CrossRef]
- Koumpis, E.; Georgoulis, V.; Papathanasiou, K.; Papoudou-Bai, A.; Kanavaros, P.; Kolettas, E.; Hatzimichael, E. The Role of microRNA-155 as a Biomarker in Diffuse Large B-Cell Lymphoma. Biomedicines 2024, 12. [CrossRef]
- Vigorito E, P.K., Abreu-Goodger C, Bunting S, Xiang Z, Kohlhaas S, Das PP, Miska EA, Rodriguez A, Bradley A, Smith KG, Rada C, Enright AJ, Toellner KM, Maclennan IC, Turner M. microRNA-155 regulates the generation of immunoglobulin class-switched plasma cells. Immunity 2007, 27, 847–859. [CrossRef]
- Chan JA, K.A., Kosik KS. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res. 2005, 65, 6029–6033. [CrossRef]
- Xu LF, W.Z., Chen Y, Zhu QS, Hamidi S, Navab R. MicroRNA-21 (miR-21) regulates cellular proliferation, invasion, migration, and apoptosis by targeting PTEN, RECK and Bcl-2 in lung squamous carcinoma, Gejiu City, China. PLoS ONE 2014, 9, 103698. [CrossRef]
- Chen L, Z.C., Wang T, You H, Yao R. Curcumin Inhibits the Proliferation, Migration, Invasion, and Apoptosis of Diffuse Large B-Cell Lymphoma Cell Line by Regulating MiR-21/VHL Axis. Yonsei Med J. 2020, 61, 20–29. [CrossRef]
- Chen W, W.H., Chen H, Liu S, Lu H, Kong D, Huang X, Kong Q, Lu Z. Clinical significance and detection of microRNA-21 in serum of patients with diffuse large B-cell lymphoma in Chinese population. Eur J Haematol 2014, 92, 407–412. [CrossRef]
- Wang Y, L.C. MicroRNA and cancer--focus on apoptosis. J Cell Mol Med. 2009, 13, 12–23. [CrossRef]
- Ho KK, M.S., Lam EW. Many forks in the path: Cycling with FoxO. Oncogene 2008, 27, 2300–2311. [CrossRef]
- Zhao X, G.L., Pan H, Kan D, Majeski M, Adam SA, Unterman TG. Biochem J. Multiple elements regulate nuclear/cytoplasmic shuttling of FOXO1: Characterization of phosphorylation- and 14-3-3-dependent and -independent mechanisms. 2004, 378, 839–849. [CrossRef]
- Go H, J.J., Kim PJ, Kim YG, Nam SJ, Paik JH, Kim TM, Heo DS, Kim CW, Jeon YK. MicroRNA-21 plays an oncogenic role by targeting FOXO1 and activating the PI3K/AKT pathway in diffuse large B-cell lymphoma. Oncotarget. 2015, 6, 15035–15049. [CrossRef]
- Huang Y, S.X., Zou Q, Wang SP, Tang SM, Zhang GZ. Biological functions of microRNAs: A review. J Physiol Biochem 2011, 67, 129–139. [CrossRef]
- Calin GA, C.C. MicroRNA signatures in human cancers. Nat Rev Cancer 2006, 6, 857–866. [CrossRef]
- Wang WY, Z.H., Wang L, Ma YP, Gao F, Zhang SJ, Wang LC. miR-21 expression predicts prognosis in hepatocellular carcinoma. Clin Res Hepatol Gastroenterol. 2014, 38, 715–719. [CrossRef]
- Baraniskin A, K.J., Schlegel U, Chan A, Deckert M, Gold R, Maghnouj A, Zöllner H, Reinacher-Schick A, Schmiegel W, Hahn SA, Schroers R. Identification of microRNAs in the cerebrospinal fluid as marker for primary diffuse large B-cell lymphoma of the central nervous system. Blood 2011, 117, 3140–3146. [CrossRef]
- Mao X, S.Y., Tang J. Serum miR-21 is a diagnostic and prognostic marker of primary central nervous system lymphoma. Neuro Sci. 2014, 35, 233–238. [CrossRef]
- Narducci MG, A.D., Picchio MC, Lazzeri C,; Pagani E, S.F., Scala E, Fadda P, Cristofoletti C, Facchiano A, Frontani M, Monopoli; A, F.M., Negrini M, Lombardo GA, Caprini; G., E.a.R. MicroRNA profiling reveals that miR-21, miR486 and miR-214 are upregulated and involved in cell survival in Sezary syndrome. Cell Death Dis 2011, 2, e151. [CrossRef]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ 2010, 17, 193–199. [CrossRef]
- Chim, C.S.; Wong, K.Y.; Qi, Y.; Loong, F.; Lam, W.L.; Wong, L.G.; Jin, D.Y.; Costello, J.F.; Liang, R. Epigenetic inactivation of the miR-34a in hematological malignancies. Carcinogenesis 2010, 31, 745–750. [CrossRef]
- Zhai, L.; Zhao, Y.; Liu, Z.; Wu, J.; Lin, L. mRNA expression profile analysis reveals a C-MYC/miR-34a pathway involved in the apoptosis of diffuse large B-cell lymphoma cells induced by Yiqichutan treatment. Exp Ther Med 2020, 20, 2157–2165. [CrossRef]
- Fu, J.; Imani, S.; Wu, M.Y.; Wu, R.C. MicroRNA-34 Family in Cancers: Role, Mechanism, and Therapeutic Potential. Cancers (Basel) 2023, 15. [CrossRef]
- Balatti, V.; Tomasello, L.; Rassenti, L.Z.; Veneziano, D.; Nigita, G.; Wang, H.Y.; Thorson, J.A.; Kipps, T.J.; Pekarsky, Y.; Croce, C.M. miR-125a and miR-34a expression predicts Richter syndrome in chronic lymphocytic leukemia patients. Blood 2018, 132, 2179–2182. [CrossRef]
- Asmar, F.; Hother, C.; Kulosman, G.; Treppendahl, M.B.; Nielsen, H.M.; Ralfkiaer, U.; Pedersen, A.; Møller, M.B.; Ralfkiaer, E.; de Nully Brown, P.; et al. Diffuse large B-cell lymphoma with combined TP53 mutation and MIR34A methylation: Another "double hit" lymphoma with very poor outcome? Oncotarget 2014, 5, 1912–1925. [CrossRef]
- He, M.; Gao, L.; Zhang, S.; Tao, L.; Wang, J.; Yang, J.; Zhu, M. Prognostic significance of miR-34a and its target proteins of FOXP1, p53, and BCL2 in gastric MALT lymphoma and DLBCL. Gastric Cancer 2014, 17, 431–441. [CrossRef]
- Marques, S.C.; Ranjbar, B.; Laursen, M.B.; Falgreen, S.; Bilgrau, A.E.; Bødker, J.S.; Jørgensen, L.K.; Primo, M.N.; Schmitz, A.; Ettrup, M.S.; et al. High miR-34a expression improves response to doxorubicin in diffuse large B-cell lymphoma. Exp Hematol 2016, 44, 238–246.e232. [CrossRef]
- Xiao, C.; Srinivasan, L.; Calado, D.P.; Patterson, H.C.; Zhang, B.; Wang, J.; Henderson, J.M.; Kutok, J.L.; Rajewsky, K. Lymphoproliferative disease and autoimmunity in mice with increased miR-17-92 expression in lymphocytes. Nat Immunol 2008, 9, 405–414. [CrossRef]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: A comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ 2013, 20, 1603–1614. [CrossRef]
- Dal Bo, M.; Bomben, R.; Hernández, L.; Gattei, V. The MYC/miR-17-92 axis in lymphoproliferative disorders: A common pathway with therapeutic potential. Oncotarget 2015, 6, 19381–19392. [CrossRef]
- Ota, A.; Tagawa, H.; Karnan, S.; Tsuzuki, S.; Karpas, A.; Kira, S.; Yoshida, Y.; Seto, M. Identification and characterization of a novel gene, C13orf25, as a target for 13q31-q32 amplification in malignant lymphoma. Cancer Res 2004, 64, 3087–3095. [CrossRef]
- Fassina, A.; Marino, F.; Siri, M.; Zambello, R.; Ventura, L.; Fassan, M.; Simonato, F.; Cappellesso, R. The miR-17-92 microRNA cluster: A novel diagnostic tool in large B-cell malignancies. Lab Invest 2012, 92, 1574–1582. [CrossRef]
- Alencar, A.J.; Malumbres, R.; Kozloski, G.A.; Advani, R.; Talreja, N.; Chinichian, S.; Briones, J.; Natkunam, Y.; Sehn, L.H.; Gascoyne, R.D.; et al. MicroRNAs are independent predictors of outcome in diffuse large B-cell lymphoma patients treated with R-CHOP. Clin Cancer Res 2011, 17, 4125–4135. [CrossRef]
- Yan, S.; Jia, C.; Quan, L.; Zhao, L.; Tian, Y.; Liu, A. Significance of the microRNA-17-92 gene cluster expressed in B-cell non-Hodgkin's lymphoma. Mol Med Rep 2019, 20, 2459–2467. [CrossRef]
- Kwanhian, W., Lenze, D., Alles, J., Motsch, N., Barth, S., Döll, C., Imig, J., Hummel, M., Tinguely, M., Trivedi, P., Lulitanond, V., Meister, G., Renner, C., & Grässer, F. MicroRNA-142 is mutated in about 20% of diffuse large B-cell lymphoma. Cancer Medicine 2012, 1, 141–155. [CrossRef]
- Menegatti, J., Nakel, J., Stepanov, Y., Caban, K., Ludwig, N., Nord, R., Pfitzner, T., Yazdani, M., Vilimova, M., Kehl, T., Lenhof, H., Philipp, S., Meese, E., Fröhlich, T., Grässer, F., & Hart, M. Changes of Protein Expression after CRISPR/Cas9 Knockout of miRNA-142 in Cell Lines Derived from Diffuse Large B-Cell Lymphoma. Cancers 2022, 14. [CrossRef]
- Larrabeiti-Etxebarria, A., Lopez-Santillan, M., Santos-Zorrozua, B., Lopez-Lopez, E., & García-Orad, A. Systematic Review of the Potential of MicroRNAs in Diffuse Large B Cell Lymphoma. Cancers 2019, 11. [CrossRef]
- Rinaldi, F., Marchesi, F., Palombi, F., Pelosi, A., Di Pace, A., Sacconi, A., Terrenato, I., Annibali, O., Tomarchio, V., Marino, M., Cantonetti, M., Vaccarini, S., Papa, E., Moretta, L., Bertoni, F., Mengarelli, A., Regazzo, G., & Rizzo, M. (2021). MiR-22, a serum predictor of poor outcome and therapy response in diffuse large B-cell lymphoma patients. British Journal of Haematology, 195. MiR-22, a serum predictor of poor outcome and therapy response in diffuse large B-cell lymphoma patients. British Journal of Haematology 2021, 195. [CrossRef]
- Marchesi, F., Regazzo, G., Palombi, F., Terrenato, I., Sacconi, A., Spagnuolo, M., Donzelli, S., Marino, M., Ercolani, C., Di Benedetto, A., Blandino, G., Ciliberto, G., Mengarelli, A., & Rizzo, M. Serum miR-22 as potential non-invasive predictor of poor clinical outcome in newly diagnosed, uniformly treated patients with diffuse large B-cell lymphoma: An explorative pilot study. Journal of Experimental & Clinical Cancer Research 2018, 37. [CrossRef]
- Kozloski, G., Jiang, X., Bhatt, S., Ruiz, J., Vega, F., Shaknovich, R., Melnick, A., & Lossos, I. miR-181a negatively regulates NF-κB signaling and affects activated B-cell-like diffuse large B-cell lymphoma pathogenesis. Blood 2016, 127, 2856–2866. [CrossRef]
- Zhu, D., Fang, C., He, W., Wu, C., Li, X., & Wu, J. (2019). MicroRNA-181a Inhibits Activated B-Cell-Like Diffuse Large B-Cell Lymphoma Progression by Repressing CARD11. Journal of Oncology 2019. [CrossRef]
- Kozloski, G., Jiang, X., Bunting, K., Melnick, A., & Lossos, I. MiR-181a Is a Master Regulator of the Nuclear Factor-κB Signaling Pathway in Diffuse Large B Cell Lymphoma. Blood 2012, 120, 417. [CrossRef]
- Alsaadi, M., Khan, M., Dalhat, M., Bahashwan, S., Khan, M., Albar, A., Almehdar, H., & Qadri, I. Dysregulation of miRNAs in DLBCL: Causative Factor for Pathogenesis, Diagnosis and Prognosis. Diagnostics 2021, 11. [CrossRef]
- Wang, H., Wang, A., Hu, Z., Xu, X., Liu, Z., & Wang, Z. (2016). A Critical Role of miR-144 in Diffuse Large B-cell Lymphoma Proliferation and Invasion. Cancer Immunology Research 2016, 4, 337–344. [CrossRef]
- Zhao, X., Xu, M., Hu, X., Ding, X., Zhang, X., Xu, L., Li, L., Sun, X., & Song, J. Human bone marrow-derived mesenchymal stem overexpressing microRNA-124-3p inhibit DLBCL progression by downregulating the NFATc1/cMYC pathway. Stem Cell Research & Therapy 2023, 14. [CrossRef]
- Ding, X., Xu, L., Sun, X., Zhao, X., Gao, B., Cheng, Y., Liu, D., Zhao, J., Zhang, X., Xu, L., & Song, J. Human Bone Marrow-Derived Mesenchymal Stem Cell-Secreted Exosomes Overexpressing Microrna-124-3p Inhibit DLBCL Progression By Downregulating NFATc1. 2020. [CrossRef]
- Longley, J., Foxall, R., Thirdborough, S., Beers, S., & Cragg, M. MicroRNA manipulation of macrophage polarization in DLBCL to augment antibody immunotherapy. Cancer Reasearch 2024. [CrossRef]
- Koumpis, E.; Papoudou-Bai, A.; Papathanasiou, K.; Kolettas, E.; Kanavaros, P.; Hatzimichael, E. Unraveling the Immune Microenvironment in Diffuse Large B-Cell Lymphoma: Prognostic and Potential Therapeutic Implications. Curr Issues Mol Biol 2024, 46, 7048–7064. [CrossRef]
- Veglia F, S.E., Gabrilovich DI. Myeloid-derived suppressor cells in the era of increasing myeloid cell diversity. Nat Rev Immunol. 2021, 21, 485–498. [CrossRef]
- Ai L, M.S., Wang Y, Wang H, Cai L, Li W, Hu Y. Prognostic role of myeloid-derived suppressor cells in cancers: A systematic review and meta-analysis. Prognostic role of myeloid-derived suppressor cells in cancers: A systematic review and meta-analysis. BMC Cancer 2018, 18, 1220. [CrossRef]
- Jabłońska, E., Białopiotrowicz, E., Szydłowski, M., Prochorec-Sobieszek, M., Juszczyński, P., & Szumera-Ciećkiewicz, A. DEPTOR is a microRNA-155 target regulating migration and cytokine production in diffuse large B-cell lymphoma cells. Experimental Hematology 2020. [CrossRef]
- CH., L. MicroRNAs and lymphomagenesis: A functional review. Br J Haematol. 2013, 160, 571–581. [CrossRef]
- Medina PP, N.M., Slack FJ. OncomiR addiction in an in vivo model of microRNA-21-induced pre-B-cell lymphoma. Nature 2010, 467, 86–90. [CrossRef]
- Li J, F.R., Yang L, Tu W. miR-21 expression predicts prognosis in diffuse large B-cell lymphoma. 2015;8(11):15019–24. Int J Clin Exp Pathol. 2015, 8, 15019–15024.
- An G, A.C., Feng X, Wen K, Zhong M, Zhang L, Munshi NC, Qiu L, Tai YT, Anderson KC. Osteoclasts promote immune suppressive microenvironment in multiple myeloma: Therapeutic implication. Blood 2016, 128, 1590–1603. [CrossRef]
- Wang X, L.C., Ju S, Wang Y, Wang H, Zhong R. Myeloma cell adhesion to bone marrow stromal cells confers drug resistance by microRNA-21 up-regulation. Leuk Lymphoma 2011, 52, 1991–1998. [CrossRef]
- De Mattos-Arruda L, B.G., Nuciforo PG, Di Tommaso L, Giovannetti E, Peg V, Losurdo A, Pérez-Garcia J, Masci G, Corsi F, Cortés J, Seoane J, Calin GA, Santarpia L. MicroRNA-21 links epithelial-to-mesenchymal transition and inflammatory signals to confer resistance to neoadjuvant trastuzumab and chemotherapy in HER2-positive breast cancer patients. Oncotarget 2015, 6, 37269–37280. [CrossRef]
- Sun, R., Zheng, Z., Wang, L., Cheng, S., Shi, Q., Qu, B., Fu, D., Leboeuf, C., Zhao, Y., Ye, J., Janin, A., & Zhao, W. A novel prognostic model based on four circulating miRNA in diffuse large B-cell lymphoma: Implications for the roles of MDSC and Th17 cells in lymphoma progression. Molecular Oncology 2020, 15. [CrossRef]
- Zheng Z, X.P., Wang L, Zhao HJ, Weng XQ, Zhong HJ, Qu B, Xiong J, Zhao Y, Wang XF, Janin A, Zhao WL. MiR21 sensitized B-lymphoma cells to ABT-199 via ICOS/ICOSL-mediated interaction of Treg cells with endothelial cells. J Exp Clin Cancer Res. 2017, 36, 82. [CrossRef] [PubMed] [PubMed Central]
Figure 1.
miRNAs are transcribed by RNA polymerase II as primary transcripts (pri-miRNAs), which are then processed in the nucleus by the enzyme Drosha along with its cofactor DGCR8 into precursor miRNAs (pre-miRNAs). These pre-miRNAs are transported to the cytoplasm via exportin 5, where they associate with the Dicer/TRBP complex and are cleaved into short double-stranded RNA molecules. One strand of this miRNA duplex is then incorporated into the Argonaute protein to form the RNA-induced silencing complex (RISC). RISC then binds to specific target mRNAs, leading to their degradation, destabilization, or inhibition of translation. here are also non-canonical miRNA biogenesis pathways, such as Drosha-independent routes (where the pri-miRNA is processed by the spliceosome) and Dicer-independent mechanisms. Abbreviations: Ago2: Argonaute 2, DGCR8: DiGeorge syndrome critical region 8, RISC: RNA-induced silencing complex, TRBP: Transactivation response element RNA-binding protein
Figure 1.
miRNAs are transcribed by RNA polymerase II as primary transcripts (pri-miRNAs), which are then processed in the nucleus by the enzyme Drosha along with its cofactor DGCR8 into precursor miRNAs (pre-miRNAs). These pre-miRNAs are transported to the cytoplasm via exportin 5, where they associate with the Dicer/TRBP complex and are cleaved into short double-stranded RNA molecules. One strand of this miRNA duplex is then incorporated into the Argonaute protein to form the RNA-induced silencing complex (RISC). RISC then binds to specific target mRNAs, leading to their degradation, destabilization, or inhibition of translation. here are also non-canonical miRNA biogenesis pathways, such as Drosha-independent routes (where the pri-miRNA is processed by the spliceosome) and Dicer-independent mechanisms. Abbreviations: Ago2: Argonaute 2, DGCR8: DiGeorge syndrome critical region 8, RISC: RNA-induced silencing complex, TRBP: Transactivation response element RNA-binding protein
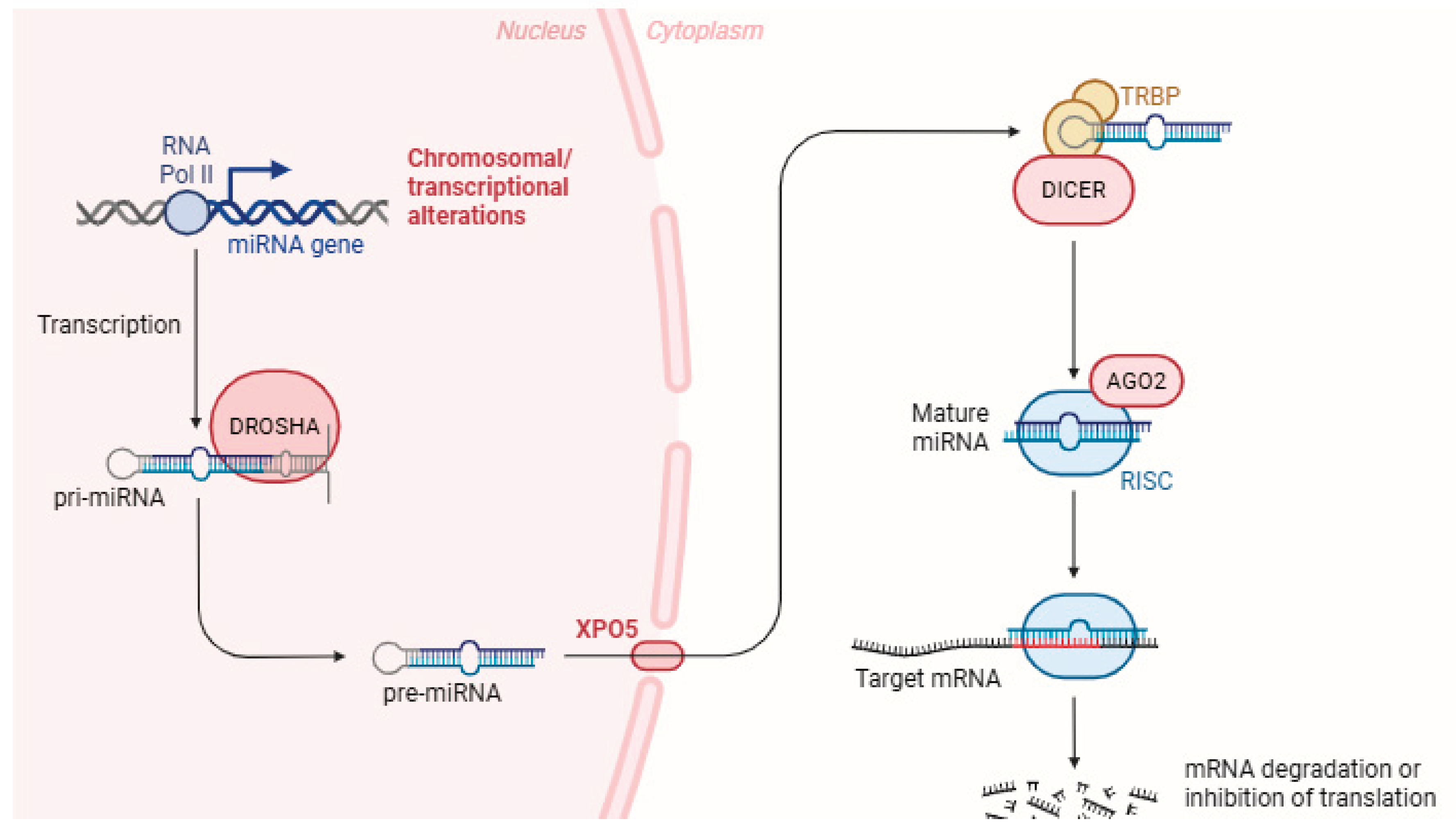
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



