
Submitted:
24 October 2025
Posted:
27 October 2025
You are already at the latest version
Abstract
Protein gelation is a key mechanism for structuring food systems, as it determines texture, water retention, and overall product stability. Therefore, understanding how processing factors influence gelation is essential for designing functional protein-based matrices. In this work, the effect of ionic strength on the gelation and techno-functional properties of Acheta domesticus (house cricket) protein extract is investigated. Protein gels were prepared with increasing NaCl concentrations (0 – 0.5 M) and evaluated using rheological analysis. Additionally, techno-functional properties of the cricket protein extract, including solubility, emulsifying properties and water/oil holding capacity, were determined. Small-amplitude oscillatory shear tests revealed that moderate salt levels enhance gel elasticity, while excessive ionic strength produces brittle structures with reduced deformability. Frequency sweeps revealed decreasing tan δ and n values, confirming a transition from weakly entangled to strong, solid-like networks. These molecular changes were mirrored by functional responses: water-holding capacity increased at high salt concentration, whereas solubility and emulsifying activity declined. Collectively, the results highlight ionic strength as a tunable parameter linking protein interactions to macroscopic texture. By adjusting salt concentration, elasticity, hydration, and interfacial stability can be strategically modulated, enabling the rational design of insect-based food gels for different applications, from emulsified matrices to fibrous meat analogues.
Keywords:
gels
; protein gelation
; ionic strength
; rheology
; techno-functional properties
; Acheta domesticus
; insect protein
; sustainable protein
1. Introduction
The global shift toward sustainable protein sources is reshaping food production systems, driven by the need to reduce environmental impacts while meeting the nutritional demands of a growing population [1,2]. Among emerging alternatives, insect proteins have gained attention for their favorable amino acid profile, digestibility, and resource efficiency compared to conventional livestock [3,4,5]. Among edible insects, Acheta domesticus (house cricket) is considered one of the most suitable species for food applications [6]. However, beyond their nutritional potential, the integration of insect proteins into structured foods depends largely on their functional and gelation properties, which are essential for delivering the texture, stability, taste and, ultimately, consumer acceptance.
Protein gelation is a fundamental process in food structuring. Upon heat treatment or environmental changes, proteins unfold and associate into three-dimensional networks that govern water retention, elasticity, and rheological performance [7]. Such properties are particularly relevant in the development of meat analogues, where gels provide the cohesive texture and firmness needed to mimic muscle tissue [8]. In plant and dairy proteins, extensive work has shown that gelation and related functional attributes can be modulated by pH, thermal treatment, and ionic strength [9,10,11].
Particularly, ionic strength plays a pivotal role in controlling protein–protein interactions. By altering electrostatic forces and hydration layers, salts influence solubility, aggregation, and the balance between elasticity and brittleness in gels. Studies on soy, whey, and pea proteins have demonstrated that moderate salt concentrations enhance gel strength and water-holding capacity, while excessive ionic strength promotes aggregation and destabilization [12,13,14]. These findings underscore the importance of ionic environments in tuning protein gel performance.
In contrast, the gelation behavior of insect proteins under varying ionic conditions remains largely unexplored. Previous studies have focused on solubility, emulsification, and foaming of insect proteins [15,16], but systematic evaluation of gelation and its modulation by ionic strength is scarce. This gap limits both the understanding of protein gel science and the ability to develop insect-based structured foods.
The present study aims to address this gap by investigating the effect of ionic strength on the gelation and techno-functional properties of cricket (Acheta domesticus) protein extracts. Rheological characterization of the cricket protein extracts-based gels at different concentration of NaCl was combined with measurements of water-holding capacity, solubility, and emulsifying properties of the protein extracts. The results reveal how ionic conditions can be strategically adjusted to control gel elasticity, hydration, and interfacial stability. This work contributes to a broader understanding of protein gelation and identifies ionic strength as a tunable parameter for designing sustainable protein gels with potential applications in food structuring.
2. Results and Discussion
2.1. Techno-Functional Properties of the Protein Extract
2.1.1. Protein Solubility
The solubility of cricket protein extracts (Figure 1) showed a clear dependence on ionic strength. Based on a total protein content of 85.65% (w/w dry extract) [17], at 0 M NaCl protein solubility was 57.97%, reflecting a good dispersion capacity of the extract in aqueous media. A statistically significant increase (p < 0.05) was observed at 0.1 M NaCl, where solubility peaked, while, beyond 0.2 M, solubility decreased progressively, reaching 64.42 ± 2.01% at 0.5 M (p < 0.05 compared to values at 0.1 M). This observed trend is consistent with the classical “salting-in/salting-out” effect. At low ionic strength, NaCl ions shield electrostatic charges, reducing repulsion and allowing proteins to remain solvated. However, at higher concentrations, competition for water molecules leads to dehydration of protein surfaces, promoting hydrophobic interactions and aggregation [12,18]. This mechanism has been well documented in plant protein systems: soy protein isolates typically show sharp decline above 0.3 M NaCl [19], while pea protein isolates exhibit 40 – 50% reduction under similar conditions [13].
Our results confirm that insect protein extracts follow comparable physicochemical principles. From a functional standpoint, solubility is a prerequisite for effective participation in network formation and interfacial activity. Proteins must be sufficiently dispersed to unfold, interact, and form structured gels upon heating or other treatments [9]. However, partial loss of solubility at higher ionic strength does not necessarily hinder gelation; instead, it can promote protein–protein interactions that lead to denser and more elastic networks. This dual effect highlights a functional trade-off between network strengthening and interfacial performance. Similar behavior has been reported for soy and whey protein isolates, where reduced solubility at high salt concentrations is associated with stronger gels but poorer emulsifying stability [20,21]. These results underscore the complex role of solubility in the functional behavior of insect protein extracts, influencing gel elasticity and interfacial properties in opposite ways depending on ionic strength
Interestingly, although the trend mirrors that of soy and pea proteins, the absolute solubility values of cricket extracts remained relatively high at the uppermost value of investigated ionic strength (≥ 64% at 0.5 M NaCl), suggesting a certain stability of solubility under ionic stress. This may be linked to the heterogeneous composition of the insect extract, which contains which contains both readily soluble globular proteins (e.g., enzymatic and storage proteins) and less soluble structural or cuticle-associated proteins. The coexistence of these fractions may broaden the functional window of the extract compared with purified isolates, maintaining dispersion even under elevated salt conditions [4,16]. The stability of solubility of the protein extracts may represent an advantage in food processing, where protein systems are often subjected to fluctuating ionic conditions.
2.1.2. Water-Holding Capacity and Oil-Holding Capacity
The Water-Holding Capacity (WHC) of cricket protein extracts was not affected by ionic strength, as shown in Figure 2. At 0 M NaCl, WHC is 3.71 ± 0.61% (w/w dry extract), reflecting the hydration ability of the extract. Across the investigated ionic strength range, WHC values showed no significant differences (p > 0.05), varying from 3.94% to 4.88%, with the highest mean value observed at 0.4 M NaCl. Although the differences were not significant, the upward trend suggests that moderate salt concentrations may enhance protein–protein interactions through electrostatic screening, promoting water entrapment within the protein matrix without causing extensive aggregation [10,11,12,13]. This observation is particularly relevant when considered alongside solubility behaviors: although solubility decreased at high ionic strength, WHC increased, suggesting that proteins excluded from the soluble fraction contributed effectively to matrix formation and water retention.
From a food structuring perspective, higher WHC at elevated salt levels suggests the potential to produce gels with improved juiciness and reduced syneresis, properties essential in applications such as meat analogues.
The Oil-Holding Capacity (OHC) of the dialyzed protein extract was 3.98 ± 0.03 g % (w/w dry extract). This high value indicates a strong lipophilic interaction capacity, likely related to the exposure of hydrophobic amino acid residues during extraction. From a functional perspective, high OHC suggests that the extract can bind non-polar molecules such as lipids and volatile flavor compounds, which is advantageous for the development of stable and palatable food systems, including emulsions, dressings, and fat-rich matrices. Although OHC was not measured across varying ionic strengths, it remains an important complementary property to WHC. Whereas WHC governs gel hydration and elasticity, OHC represents the capacity of the extract to bind lipids, thereby contributing to improved sensory perception and formulation stability.
2.1.3. Emulsifying Properties
The Emulsifying Activity Index (EAI, Figure 3a) and Emulsion Stability Index (ESI, Figure 3b) of cricket protein extracts showed different non-monotonic responses to the NaCl concentration.
The EAI progressively increased from 0 to 0.3 M NaCl, where the highest activity was recorded, and then declined at higher concentrations, reaching values comparable to the salt-free extract. In contrast the ESI reached its highest value at 0.1 M NaCl (p < 0.05) and then declined sharply beyond 0.2 M, showing the lowest stability at 0.5 M. The increase in EAI up to 0.3 M may be attributed to partial charge screening that facilitates protein adsorption and interfacial packing, as widely reported for globular protein systems [21,22]. At moderate ionic strength, salt ions reduce electrostatic repulsion between protein molecules, enabling closer packing at the oil–water interface and improving interfacial coverage.
However, the simultaneous drop in ESI at high salt concentrations suggests that, despite faster interfacial coverage, the interfacial films formed are more susceptible to flocculation/coalescence due to strong screening of electrostatic repulsions between droplets—an effect observed in protein-stabilized emulsions at elevated ionic strength [21]. This behavior can be explained by excessive charge screening, which suppresses electrostatic stabilization between emulsion droplets, which is a commonly observed effect in protein-stabilized emulsions at elevated ionic strength [21]. At 0.5 M NaCl, both EAI and ESI were low, consistent with bulk aggregation and reduced solubility, which limit the number of mobile, interfacially active molecules [19]. These trends are consistent with the solubility behavior, where higher ionic strength promoted aggregation and reduced protein dispersion. Lower solubility consequently diminishes the number of surface-active molecules available to migrate and stabilize oil droplets. Conversely, at low to moderate salt concentrations, where solubility remains high, proteins are sufficiently unfolded and flexible to efficiently adsorb at the interface, explaining the higher EAI and ESI values observed in this range. The relatively high OHC of the extract further supports its intrinsic emulsifying potential, since exposed hydrophobic residues favor anchoring at the oil–water interface. Together, these results suggest that ionic strength modulates emulsifying behavior through its simultaneous influence on solubility and hydrophobic interactions, balancing dispersion and aggregation within the system.
ESI and ESI results are compatible with reports on soy proteins under NaCl, where moderate salt can improve interfacial activity, but excessive ionic strength induces aggregation and destabilization [19], and align with observations on insect protein extracts, whose emulsifying performance is sensitive to ionic environment and dispersion state [4]
2.2. Rheological Properties of Protein Gels
To evaluate the effect of ionic strength on the viscoelastic behavior of the gels, amplitude sweep tests were carried out. The results (Figure 4) revealed an elastic-dominant profile (G′ > G′′), confirming the establishment of three-dimensional protein networks, capable of storing deformation energy rather than dissipating it through flow [24].
The Linear Viscoelastic Region (LVR) decreased with increasing salt concentration, indicating that the gels became less tolerant to deformation before structural breakdown. Simultaneously, the storage modulus (G′) raised consistently with NaCl, from less than 200 Pa at 0 M NaCl to more than 1300 Pa at 0.5 M, reflecting denser and more cross-linked protein assemblies.
These changes denote that moderate to high ionic strength enhances protein–protein interactions by screening repulsive charges and favoring hydrophobic associations. However, yield strain shifted downward, reflecting a trade-off: stronger gels formed, but they became more brittle and prone to fracture under stress. Similar outcomes have been observed in pea and soy gels [25,26], where the addition of salts increases cross-linking and rigidity at the expense of plasticity.
Interestingly, a distinctive and somewhat atypical observation derived from the amplitude sweep tests is the initial ascending trend in both moduli within what is conventionally defined as the LVR. In ideal gel systems, the LVR is rigorously defined by a plateau region where G′ and G′′ maintain constant values, indicating that the applied strain is insufficient to induce disruption of the network structural integrity [27]. In the present case, a perfectly linear region could not be clearly identified, nevertheless, the strain interval preceding the decrease in G′ can be reasonably considered an apparent LVR, since the variation of G′ remained within approximately 10% of its initial value, which is an operational criterion commonly applied in non-ideal gel systems [27,28]. Overall, we attribute the observed increase in the moduli to either slow gel maturation during testing or to minor strain-induced microstructural rearrangements that transiently enhance network connectivity. Similar low-strain stiffening or alignment phenomena have been reported in whey protein systems, where a mild strain-hardening response within the nominally linear region was attributed to microstructural rearrangements and progressive formation of junction zones that temporarily increase network connectivity and stiffness [28]. Additional analyses would help discriminate between time-dependent aging and genuine strain-induced stiffening.
Frequency sweeps (Figure 5) further underline the solid-like nature of the gels across the 0.1 – 100 Hz range: the relatively limited dependence of both G′ and G′′ on angular frequency, in fact, reinforces the conclusion that stable, predominantly elastic, cross-linked networks were successfully formed [29]. This behavior is indeed characteristic of gel-like structure, wherein the intrinsic relaxation times of the network significantly exceed the inverse of the applied oscillation frequency.
Increasing NaCl concentration causes G′ to rise quickly, whereas G′′ increases only slightly, resulting in lower tan δ values at higher salt levels. The lowest value of tan δ, registered 0.3 M NaCl (Table 1), corresponds to the most elastic and energy-storing gels. Such behavior suggests that the presence of salt promotes a more interconnected and less dissipative network, consistent with the “salting-in” effect on partially aggregated proteins that reorganize into continuous structures during heating. At 0.5 M NaCl, tan δ remained statistically comparable to that at 0.3 M (p > 0.05), suggesting that gel elasticity reached a plateau. This suggests that further ionic screening no longer strengthens the network measurably; instead, excessive salt may begin to promote slight phase separation or weaker junction zones at the microstructural level [12,14,22]. Nevertheless, the overall elastic dominance of the gels was maintained, as tan δ values remained significantly lower than the control (0 M).
To further quantify these rheological trends and better interpret the structural evolution of the gels, each sweep was fitted to a power-law model, according to equations (6) and (7) within the linear, monotonic region of the log–log plots (Table 2).
This approach is widely adopted for characterizing protein and biopolymer gels under small-amplitude oscillatory shear, proximate scaling accurately captures the viscoelastic behavior [11,25,30,31,32]. This approach provides complementary insight into the mechanical strength and organization of the protein network beyond the qualitative trends of G′ and G′′. The consistency indices k′ and k′′, related to the magnitude of G′ and G′′, respectively, increase markedly with salt concentration, confirming the progressive reinforcement of the gel matrix. The largest rise of both indices is observed between 0.1 and 0.3 M NaCl, indicating the formation of a stable, compact network capable of storing mechanical energy. The frequency-dependence exponents n′ and n″ quantify the sensitivity of the moduli to oscillation frequency: as NaCl increases, both n′ and n″ decreases, implying that both G′ and G′′ become progressively less frequency-dependent. The strong increase in k′ and the corresponding decrease in n′ with NaCl concentration highlight the transition from soft, weakly connected aggregates to densely cross-linked, elastic structures. This trend is characteristic of well-developed gels in which structural relaxation is limited—a pattern also reported for plant proteins under similar ionic conditions [25,26]. Moreover, the combined reduction in n values and tan δ thus reflects a transition from weak, entangled networks toward elastic, solid-like matrices, reinforcing the mechanistic role of ionic screening in promoting junction zone formation and stabilizing the gel network. Hence, the power law modeling framework validates the observed experimental evolution of G′ and tan δ, confirming that ionic strength drives a shift from viscous-dominated to elasticity-dominated behavior.
To connect the power-law model with the compositional variable, the fitted parameters were empirically correlated with NaCl concentration. Linear regressions were applied to the consistency indices (k′, k″), while exponential functions were used for the frequency-dependence exponents (n′, n″), as described by Equations (8) and (9). The fitting coefficients derived from these models are summarized in Table 3, and the resulting trends of the power-law parameters as a function of NaCl concentration are illustrated in Figure 6.
Although purely descriptive, these relationships provide a functional map linking the rheological parameters to ionic conditions, allowing identification of the salt range associated with maximal stiffness (high k′) and minimal frequency dependence (low n′). Such empirical correlations are particularly useful in translating experimental rheological data into formulation strategies. By expressing k′ and n′ as functions of NaCl concentration, it becomes possible to predict the mechanical response of cricket protein gels under given ionic environments without performing a full frequency sweep. This approach is consistent with prior modeling of biopolymer networks, where power-law parameters have been shown to capture the degree of cross-linking, relaxation behavior, and energy dissipation in protein and polysaccharide gels [31,32].
In this context, the increase in k′ and decrease in n′ with ionic strength reflect the formation of a progressively stronger, less frequency-dependent structure, characteristic of networks with reduced segmental mobility and enhanced cooperative junction density. Such structural evolution underlies the observed transition from soft, weakly connected aggregates to compact, elastic networks as salt concentration rises. In contrast, the correlations for k″ and particularly for n″ were weaker (R² < 0.83), suggesting indicating greater data variability and the absence of a consistent dependence of the viscous component on salt concentration. Taken together, these empirical correlations can be viewed as a useful descriptive framework for understanding how ionic strength modulates gel stiffness and relaxation dynamics. However, the limited number of experimental points constrains their statistical robustness and prevents rigorous extrapolation. Extending the analysis to a broader ionic strength range and performing independent validation would be necessary to confirm the quantitative and predictive value of these relationships for protein gel systems.
From a molecular standpoint, this evolution, as captured by the fitted power-law parameters, can be attributed to electrostatic screening that facilitates close packing and hydrophobic clustering of partially unfolded protein molecules, leading to a greater number of effective junction zones. According to previous results, the increase in G′ and k’ parallels the trends in WHC, indicating that tighter matrices retain more water, while the decline in solubility confirms that aggregation rather than dispersion dominates the behavior at elevated ionic strength. Conversely, the same compactness that strengthens elasticity reduces protein availability at the oil–water interface, impairing emulsifying properties. Thus, rheology bridges solubility, WHC, and emulsification: salt promotes aggregation, which enhances gel elasticity and hydration but diminishes interfacial stability. Such a balance is crucial in food applications where different textures and functionalities are targeted.
3. Conclusions
This work demonstrated that ionic strength plays a pivotal role in tuning both the techno-functional properties and the gelation behavior of Acheta domesticus protein extracts. The observed trends revealed a dynamic balance in which solubility governs the availability of reactive protein species, while water retention and viscoelasticity depend on the degree of aggregation and network consolidation promoted by ionic screening. At moderate salt concentrations, these interactions led to the formation of elastic, water-rich gels, whereas excessive ionic strength promoted extensive aggregation resulting in stronger but less functional matrices
From a technological perspective, this duality represents a powerful tool for product design. Ionic strength adjustment enables the tailoring of protein-based gels to achieve specific textures and hydration levels suited to different applications—soft, emulsifying matrices at low salt for spreads or patties; firm, cohesive gels at higher salt for fibrous analogues or structured meat substitutes. Such tunability can be further exploited in other food systems, such as sauces, dressings, or protein-rich snacks, where texture and water retention are decisive for product quality. The ability to control gel elasticity through salt concentration also offers potential to reduce or eliminate added hydrocolloids, supporting clean-label formulation trends.
Moreover, the rheological modeling performed in this study provided a quantitative link between ionic strength and the resulting mechanical behavior. The integration of power-law analysis with empirical correlations of the parameters (k′, k″, n′, n″) against NaCl concentration offered a simplified framework to describe how ionic environments modulate gel stiffness and frequency-dependent relaxation. Such modeling, with further validation through expanded datasets, cross-validation, and mechanistic interpretation, could evolve into a robust predictive tool capable of identifying optimal salt ranges for desired textural properties, thereby reinforcing the concept of controllable, salt-responsive gelation.
Finally, from a broader formulation perspective, Acheta domesticus protein extracts emerged as versatile structuring ingredient, able to complement or replace conventional proteins in sustainable food systems. Their salt-sensitive gelation behavior provides a practical means to modulate elasticity, hydration, and stability directly through formulation parameters, without the need for external hydrocolloids. This controllable balance between firmness and functionality positions insect proteins as a promising ingredient for the development of clean-label, high-protein foods where texture, water retention, and structural integrity are critical to product quality.
4. Materials and Methods
4.1. Materials
The Acheta domesticus flour was purchased from Small Giants S.r.l. (Milan, Italy). The nutritional composition of the flour, as reported by the manufacturer, is reported in Table 4. The Pierce™ Bradford Protein Assay reagent was purchased from Thermo Fisher Scientific Inc. (Waltham, Massachusetts, USA) The Slide – A – Lyzer dialysis flasks were purchased from Thermo Fisher Scientific Inc. (Waltham, Massachusetts, USA). All other chemicals were of analytical grade and purchased from Merck (Darmstadt, Germania). The palm oil was purchased from Euronut s.p.a (Avellino, Italy).
4.2. Protein Extraction
Cricket flour was first defatted [33] by solvent extraction with n-hexane (1:5, w/v) under continuous stirring for 3 h at room temperature, followed by centrifugation (SL 8 Small Benchtop, Thermo Fisher Scientific, Waltham, MA, USA) at 3100g for 10 min and overnight air-drying of the pellet under a fume hood to remove residual solvent.
Subsequently, protein extraction was carried out by alkaline solubilization coupled with isoelectric precipitation [34]: the defatted flour was suspended in 0.25 M NaOH (1:15, w/v), stirred for 2 h at 40 °C, and centrifuged at 3500g for 20 min; a second extraction was repeated on the pellet under the same condition and pooled supernatants were adjusted to pH 4.4 with 1 M HCl to induce precipitation. The protein extract was collected by centrifugation (2500g, 20 min), neutralized to pH 7.0 and freeze dried. According to our previous study the protein content of the extract was close to 86% [17]. To remove salts and low-molecular-weight compounds, protein extract suspensions (12% w/v in distilled water) were dialyzed in 3.5 kDa MWCO cassettes (Slide – A – Lyzer dyalisis, Thermofisher, USA) against distilled water under gentle stirring for 24 h [35]. Dialyzed suspensions were recovered, freeze-dried, and used for subsequent analyses.
4.3. Protein Solubility
Protein solubility was determined according to a colorimetric method based on the Bradford assay [36]. Cricket protein extract (0.1% w/v) was dispersed in Phosphate Buffer Solutions (PBS, 0.1M, pH 7) with NaCl concentration ranging from 0.1 to 0.5 M. A dispersion of the extract in PBS without added salt was used as control. Samples were stirred for 60 min at room temperature and centrifuged at 3200g for 20 min. Supernatants (50 μL) were mixed with 1.5 mL of Pierce™ Bradford Protein Assay reagent, incubated for 10 min, and absorbance was measured at 595 nm using a UV-6300PC spectrophotometer (VWR International, Leuven, Belgium). Blanks were prepared with the corresponding buffer and assay reagent. Protein concentrations were quantified against a calibration curve prepared with bovine serum albumin as standard.
Solubility was calculated as the percentage of soluble protein (PSoluble) relative to the total protein content (PTotal), according to Equation (1):
Total protein content was determined by the Kjeldahl method [37], using 6.25 as a nitrogen-to-protein conversion factor.
4.4. Water-Holding Capacity and Oil-Holding Capacity
The Water-Holding Capacity (WHC) and Oil-Holding Capacity (OHC) of the samples were evaluated following gravimetric method [38], with minor modifications. Briefly, 0.1 g of sample was suspended in PBS at different concentration of NaCl (0 to 0.5 M). PBS/palm oil was slowly added to the centrifugal tube until the sample was covered and then, tubes were vortexed for 1 min (REAX 2000, Heidolph GmbH, Germany), left to stand at room temperature for 30 min, and centrifuged at 3200g for 15 min at 20 °C. The supernatant was carefully decanted, and the retained PBS/oil was quantified gravimetrically.
WHC and OHC were expressed as the percentage of water or oil retained by the extract relative to the total mass of the sample on dry basis, according to the following equations:
where m0 is the mass (g) of the protein extract (dry basis), m₁ is the initial weight of the centrifuge tube with the sample, and m₂ is the final weight of the tube after supernatant removal.
4.5. Emulsifying Properties
The evaluation of the emulsifying activity index (EAI) and emulsion stability index (ESI) was conducted through turbidimetric method [39]. Briefly, a 0.1% w/v protein extract solution was prepared in PBS (0.1 M, pH 7.0) and different NaCl concentrations, ranging from 0 to 0.5 M. Emulsions were prepared by homogenizing the protein solution and palm oil at a 1:4 (v/v) ratio using an Ultra-Turrax T25 homogenizer (IKA, Staufen, Germany) at 10,000 rpm for 1 min. Aliquots (50 μL) of each emulsion were collected immediately after homogenization and after 10 min, diluted in 5 mL of 0.1% w/v SDS solution, and absorbance were measured at 500 nm (UV–Visible spectrophotometer, VWR International, USA), with 0.1% SDS solution as blank control. EAI (m²/g) and ESI (min) were calculated using the following equations:
where A0 and A10 are the absorbances of the diluted emulsions at 0 min and 10 min, respectively, DF is the dilution factor (100), C is the concentration of protein (g/mL) before emulsification, φ is the proportion of the oil phase (0.25), and θ is the optical path (1 cm) and 104 is a unit conversion factor to express EAI as m²/g.
4.6. Gel Preparation
Protein gels were prepared from cricket protein extract (15% w/v) in PBS containing different NaCl concentrations (0.1, 0.3 and 0.5 M). Solutions were heated in a water bath (WB-M5, Falc, Treviglio, Italy) to 90 °C to induce gelation, held at this temperature for 15 min, cooled at room temperature for 10 min, and subsequently stored at 4 °C for 20 min to complete gel formation. A control gel was prepared using PBS without NaCl addition under the same conditions.
4.7. Rheological Characterization
The viscoelastic properties of the gels were evaluated using a rotational rheometer (Anton Paar MCR 102e) equipped with a Peltier temperature device (P-PTD200/56) and plate – plate geometry (25 mm diameter, PP25 system). Immediately after preparation, gel samples were carefully loaded onto the rheometer, and the upper plate was adjusted to a final gap of 2 mm [25]. All rheological measurements were conducted at 20 °C
4.7.1. Amplitude Sweep
Amplitude sweep tests were conducted at a fixed frequency of 0.1 Hz to determine the linear viscoelastic region (LVR), with strain ranging from 0.01% to 100%. The RheoCompass® software (Anton Paar, Austria) provided the storage modulus (G′, Pa) and loss modulus (G′′, Pa), which reflect the elastic and viscous behavior of the gels, respectively. The yield strain was defined as the strain point where the storage modulus (G′) began to decline by more than 5%, indicating the onset of structural breakdown [26].
4.7.1. Frequency Sweep
Frequency sweep tests were performed within the LVR over a frequency range of 0.1 – 100 Hz. The storage modulus (G′, Pa) and loss modulus (G′′, Pa) were recorded and the loss factor (tan δ = G”/ G′) was also provided by the software as indication of the balance between viscous and elastic contributions to the gel network. The frequency sweep data were fitted using a power law model in order to describe the dependence of the storage modulus (G′) and the loss modulus (G′′) on the angular frequency (ω), as expressed in Equations (6) and (7) [40]:
where k′ and k′′ are the consistency indices associated with G′ and G′′, respectively, and n′ and n′′ represent the frequency-dependence exponents.
To integrate the power-law modeling with the compositional variable, the parameters were empirically correlated with NaCl concentration using linear and exponential fits for the consistency indices and the exponents respectively:
where a, b, c and d are fitting coefficients, and x represents the NaCl concentration.
4.8. Statistical Analysis
All measurements were performed in triplicate, and results are expressed as mean ± standard deviation (SD). One-way analysis of variance (ANOVA) was applied to evaluate overall differences among treatments. When ANOVA indicated significance (p < 0.05), Tukey’s honest significant difference (HSD) post hoc test was used to perform pairwise comparisons between groups. Statistical analyses were conducted using JMP statistical software (SAS Institute. Inc. Cary, NC, USA).
Author Contributions
Conceptualization, D.A. and R.F.L.; methodology, R.F.L.; validation, R.F.L., M.D. and F.Mal.; formal analysis, R.F.L. and M.D.; investigation, R.F.L., M.D., D.A. and F.Mal.; data curation, R.F.L. and M.D.; writing—original draft preparation, R.F.L.; writing—review and editing, R.F.L., E.M., D.A., F.Mal., F.M. and O.H.C.; visualization, F.M. and O.H.C.; supervision, D.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Payne, C.L.R.; Scarborough, P.; Rayner, M.; Nonaka, K. A systematic review of nutrient composition data available for twelve commercially available edible insects, and comparison with reference values. Trends Food Sci. Technol. 2016, 47, 69–77. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.M.; Sagis, L.M.; Eisner-Schadler, V.; van Huis, A.; van Boekel, M.A. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef] [PubMed]
- Mishyna, M.; Chen, J.; Benjamin, O. Sensory attributes of edible insects and insect-based foods—Future outlooks for enhancing consumer appeal. Trends Food Sci. Technol. 2020, 95, 141–148. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Jakubczyk, A. Antioxidant activity of predigested protein obtained from a range of farmed edible insects. Int. J. Food Sci. Technol. 2017, 52, 306–312. [Google Scholar] [CrossRef]
- Clark, A.H.; Ross-Murphy, S.B. Structural and mechanical properties of biopolymer gels. Adv. Polym. Sci. 1983, 83, 57–192. [Google Scholar] [CrossRef]
- Dekkers, B.L.; Boom, R.M.; van der Goot, A.J. Structuring processes for meat analogues. Trends Food Sci. Technol. 2018, 81, 25–36. [Google Scholar] [CrossRef]
- Foegeding, E.A.; Davis, J.P. Food protein functionality: A comprehensive approach. Food Hydrocoll. 2011, 25, 1853–1864. [Google Scholar] [CrossRef]
- Hermansson, A.M. Water- and fat-holding. In Functional Properties of Food Macromolecules; 1st ed. Elsevier Applied Science; London, UK, 1986; pp. 273–314.
- Bryant, C.M.; McClements, D.J. Molecular basis of protein functionality with special consideration of cold-set gels derived from heat-denatured whey. Trends Food Sci. Technol. 1998, 9, 143–151. [Google Scholar] [CrossRef]
- Renkema, J.M.S.; Gruppen, H.; van Vliet, T. Influence of pH and ionic strength on heat-induced formation and rheological properties of soy protein gels in relation to denaturation and their protein compositions. J. Agric. Food Chem. 2002, 50, 6064–6071. [Google Scholar] [CrossRef] [PubMed]
- Tanger, C.; Müller, M.; Andlinger, D.; Kulozik, U. Influence of pH and ionic strength on the thermal gelation behaviour of pea protein. Food Hydrocoll. 2022, 123, 106903. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Kilara, A. Gelation of pH-aggregated whey protein isolate solution induced by heat, protease, calcium salt, and acidulant. J. Agric. Food Chem. 1998, 46, 1830–1835. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Mishyna, M.; Martinez, J.J.I.; Chen, J.; Benjamin, O. Extraction, characterization and functional properties of soluble proteins from edible grasshopper (Schistocerca gregaria) and honey bee (Apis mellifera). Food Res. Int. 2019, 116, 697–706. [Google Scholar] [CrossRef]
- Lanza, R.F.; Muccio, E.; Malvano, F.; Marra, F.; Albanese, D. Exploring the Potential of Acheta domesticus Protein Extracts for the Future of Meat Analogues. Chem. Eng. Trans. 2025, 118, 79–84. [Google Scholar] [CrossRef]
- Stone, A.K.; Karalash, A.; Tyler, R.T.; Warkentin, T.D.; Nickerson, M.T. Functional attributes of pea protein isolates prepared using different extraction methods and cultivars. Food Res. Int. 2015, 76, 31–38. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, J.; Xiong, Y.L. Structural and emulsifying properties of soy protein isolate subjected to acid and alkaline pH-shifting processes. J. Agric. Food Chem. 2009, 57, 7576–7583. [Google Scholar] [CrossRef]
- Sager, V.F.; de la Hera, E.; Gomez, M. Formulation of Heat-Induced Whey Protein Gels for Extrusion-Based 3D Printing. Foods 2021, 10, 8. [Google Scholar] [CrossRef]
- Dickinson, E. Flocculation of protein-stabilized oil-in-water emulsions. Colloids Surf. B Biointerfaces 2010, 81, 130–140. [Google Scholar] [CrossRef]
- Langton, M.; Hermansson, A.M. Fine-stranded and particulate gels of β-lactoglobulin and whey protein at varying ionic strength. Food Hydrocoll. 1992, 5, 523–539. [Google Scholar] [CrossRef]
- McClements, D.J. Protein-stabilized emulsions. Curr. Opin. Colloid Interface Sci. 2004, 9, 305–313. [Google Scholar] [CrossRef]
- Sun, C.; Purohit, P.K. Rheology of fibrous gels under compression. Extreme Mech. Lett. 2022, 54, 101757. [Google Scholar] [CrossRef]
- Klost, M.; Drusch, S. Structure formation and rheological properties of pea protein-based gels. Food Hydrocoll. 2019, 94, 622–630. [Google Scholar] [CrossRef]
- Kaboorani, A.; Blanchet, P. Determining the linear viscoelastic region of sugar maple wood by dynamic mechanical analysis. BioResources 2014, 9, 4392–4409. [Google Scholar] [CrossRef]
- Rivera-Hernández, L.; Chavarría-Hernández, N.; Tecante, A.; López-Ortega, M.A.; López Cuellar, M.R.; Rodríguez-Hernández, A.I. Mixed gels based on low acyl gellan and citrus pectin: A linear viscoelastic analysis. Food Hydrocoll. 2022, 137, 108353. [Google Scholar] [CrossRef]
- Ewoldt, R.H.; Hosoi, A.E.; McKinley, G.H. New measures for characterizing nonlinear viscoelasticity in large amplitude oscillatory shear. J. Rheol. 2008, 52, 1427–1458. [Google Scholar] [CrossRef]
- Ghosh, K.; Shu, X.Z.; Mou, R.; Lombardi, J.; Prestwich, G.D.; Rafailovich, M.H.; Clark, R.A.F. Rheological characterization of in situ cross-linkable hyaluronan hydrogels. Biomacromolecules 2005, 6, 2857–2865. [Google Scholar] [CrossRef]
- Winter, H.H.; Chambon, F. Analysis of linear viscoelasticity of a crosslinking polymer at the gel point. J. Rheol. 1986, 30, 367–382. [Google Scholar] [CrossRef]
- Mezger, T.G. The Rheology Handbook, 4th ed.; Vincentz Network: Hannover, 2014. [Google Scholar]
- Ross-Murphy, S.B. Rheological characterization of gels. J. Texture Stud. 1995, 26, 391–400. [Google Scholar] [CrossRef]
- Choi, B.D.; Wong, N.A.K.; Auh, J.-H. Defatting and sonication enhances protein extraction from edible insects. Korean J. Food Sci. Anim. Resour. 2017, 37, 955–961. [Google Scholar] [CrossRef]
- Zhao, X.; Vázquez-Gutiérrez, J.L.; Johansson, D.P.; Landberg, R.; Langton, M. Yellow mealworm protein for food purposes—Extraction and functional properties. PLoS ONE 2016, 11, e0147791. [Google Scholar] [CrossRef]
- Thermo Fisher Scientific. Slide-A-Lyzer™ Dialysis Cassettes and Flasks. 2019. Available online: https://www.thermofisher.com/it/en/home/life-science/protein-biology/protein-purification-isolation/protein-dialysis-desalting-concentration/dialysis-products/slide-a-lyzer-dialysis-cassettes.html (accessed on 2 June 2025).
- Zhang, Z.; Holden, G.; Wang, B.; Adhikari, B. Maillard reaction-based conjugation of Spirulina protein with maltodextrin using wet-heating route and characterisation of conjugates. Food Chem. 2023, 406, 134931. [Google Scholar] [CrossRef]
- AOAC International. AOAC International. AOAC Official Method 2001.11 Protein (Crude) in Animal Feed, Forage (Plant Tissue), Grain, and Oilseeds. In Official Methods of Analysis of AOAC International; Oxford University Press: Oxford, UK, 2023. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, D.; Wang, J.; Yang, Y.; Zhang, L.; Li, J.; Wang, S. Functional properties and structural characteristics of phosphorylated pea protein isolate. Int. J. Food Sci. Technol. 2020, 55, 2002–2010. [Google Scholar] [CrossRef]
- Purdi, T.S.; Setiowati, A.D.; Ningrum, A. Ultrasound-assisted extraction of Spirulina platensis protein: Physicochemical characteristic and techno-functional properties. J. Food Meas. Charact. 2023, 17, 5474–5486. [Google Scholar] [CrossRef]
- Alvarez, M.D.; Canet, W. Dynamic Viscoelastic Behavior of Vegetable-Based Infant Purees. J. Texture Stud. 2013, 44, 205–224. [Google Scholar] [CrossRef]
Figure 1.
Protein solubility of cricket protein extract as a function of NaCl concentration. Data points followed by different letters (a - d) are significantly different (p < 0.05).
Figure 1.
Protein solubility of cricket protein extract as a function of NaCl concentration. Data points followed by different letters (a - d) are significantly different (p < 0.05).
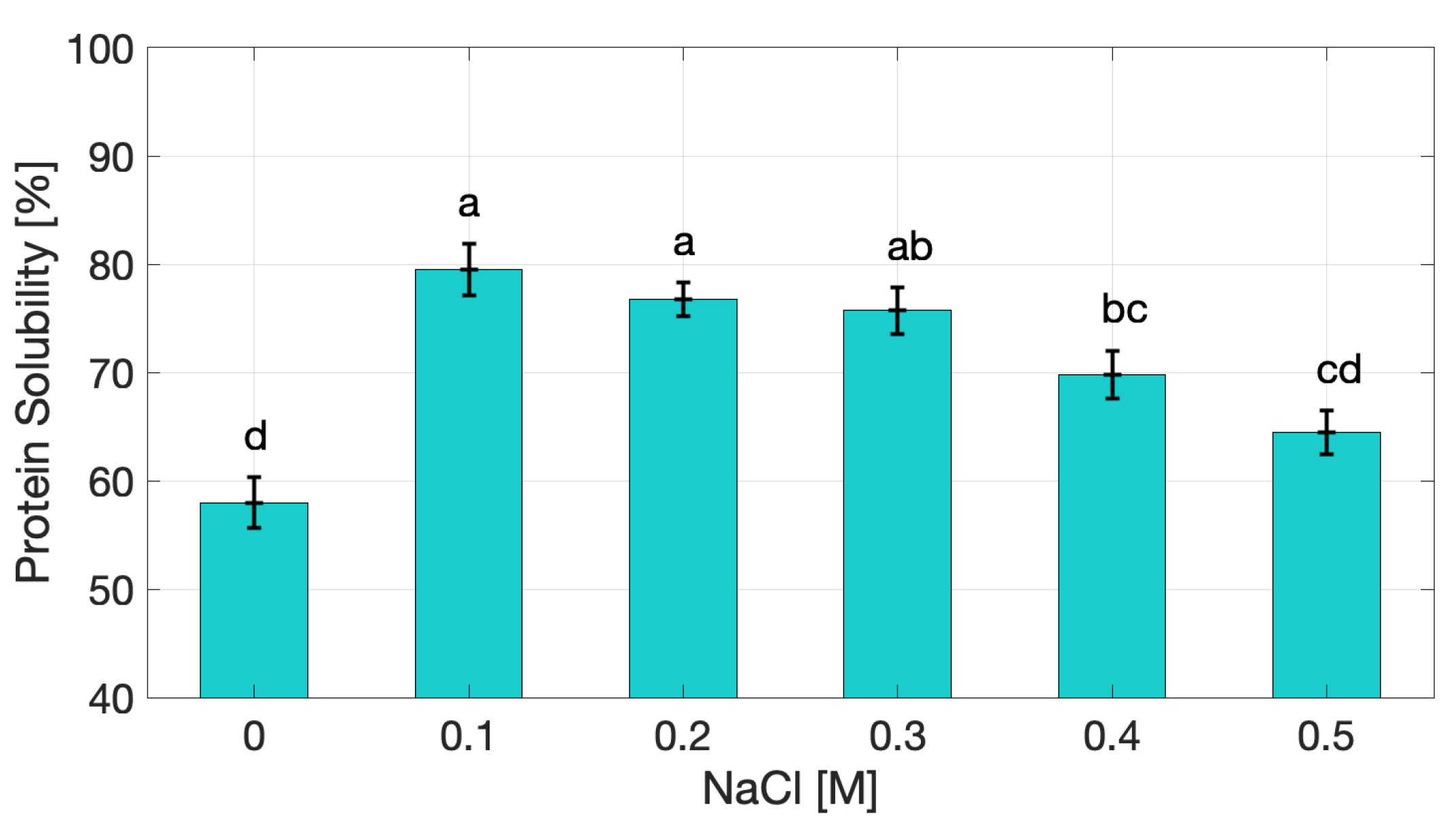
Figure 2.
Water Holding Capacity of cricket protein extract as a function of NaCl concentration.
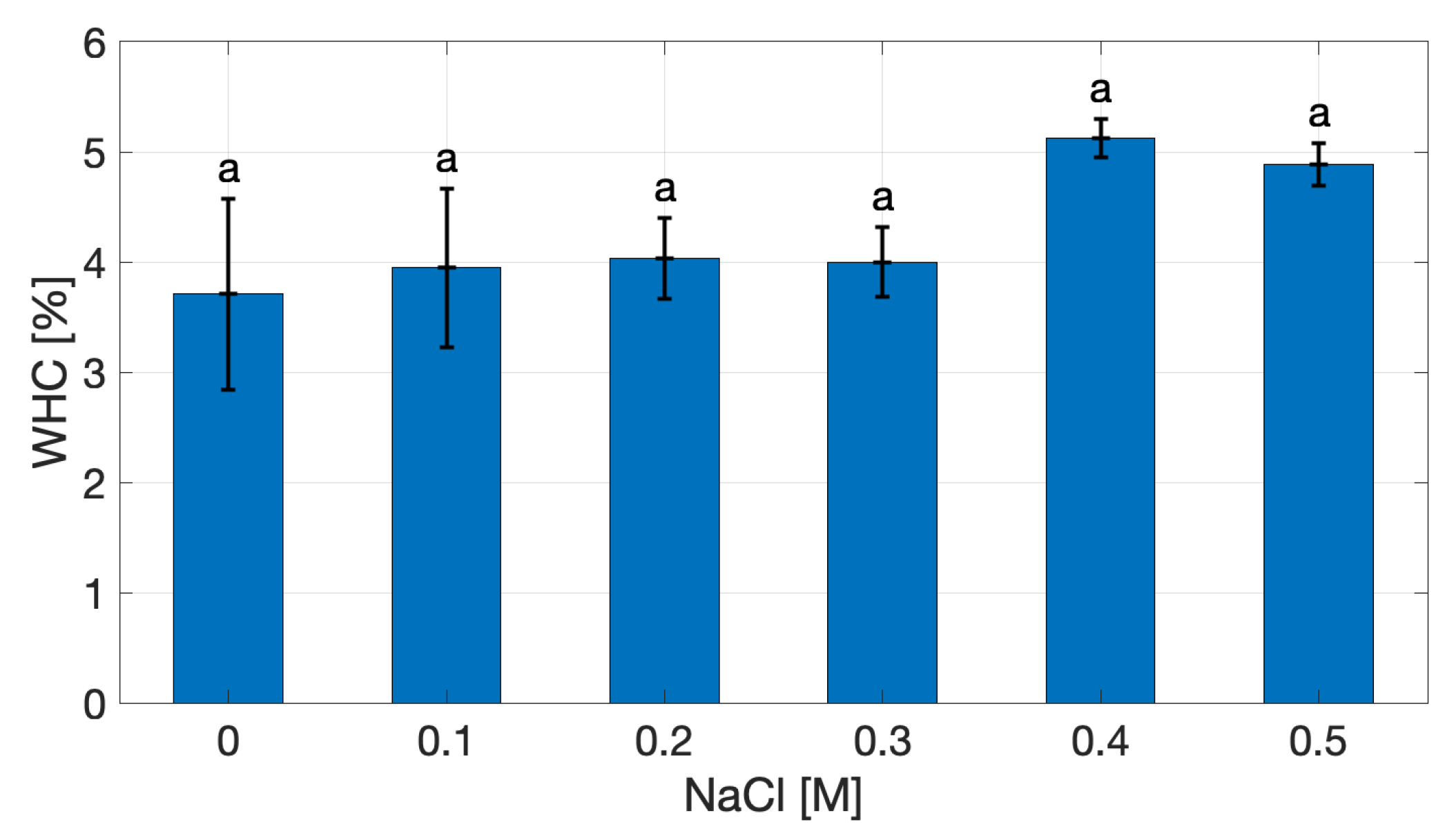
Figure 3.
Emulsion Activity Index (a) and Emulsion Stability Index (b) of cricket protein extract under varying NaCl concentrations. Values marked with different letters (a – d and A – B) indicate statistically significant differences (p < 0.05).
Figure 3.
Emulsion Activity Index (a) and Emulsion Stability Index (b) of cricket protein extract under varying NaCl concentrations. Values marked with different letters (a – d and A – B) indicate statistically significant differences (p < 0.05).
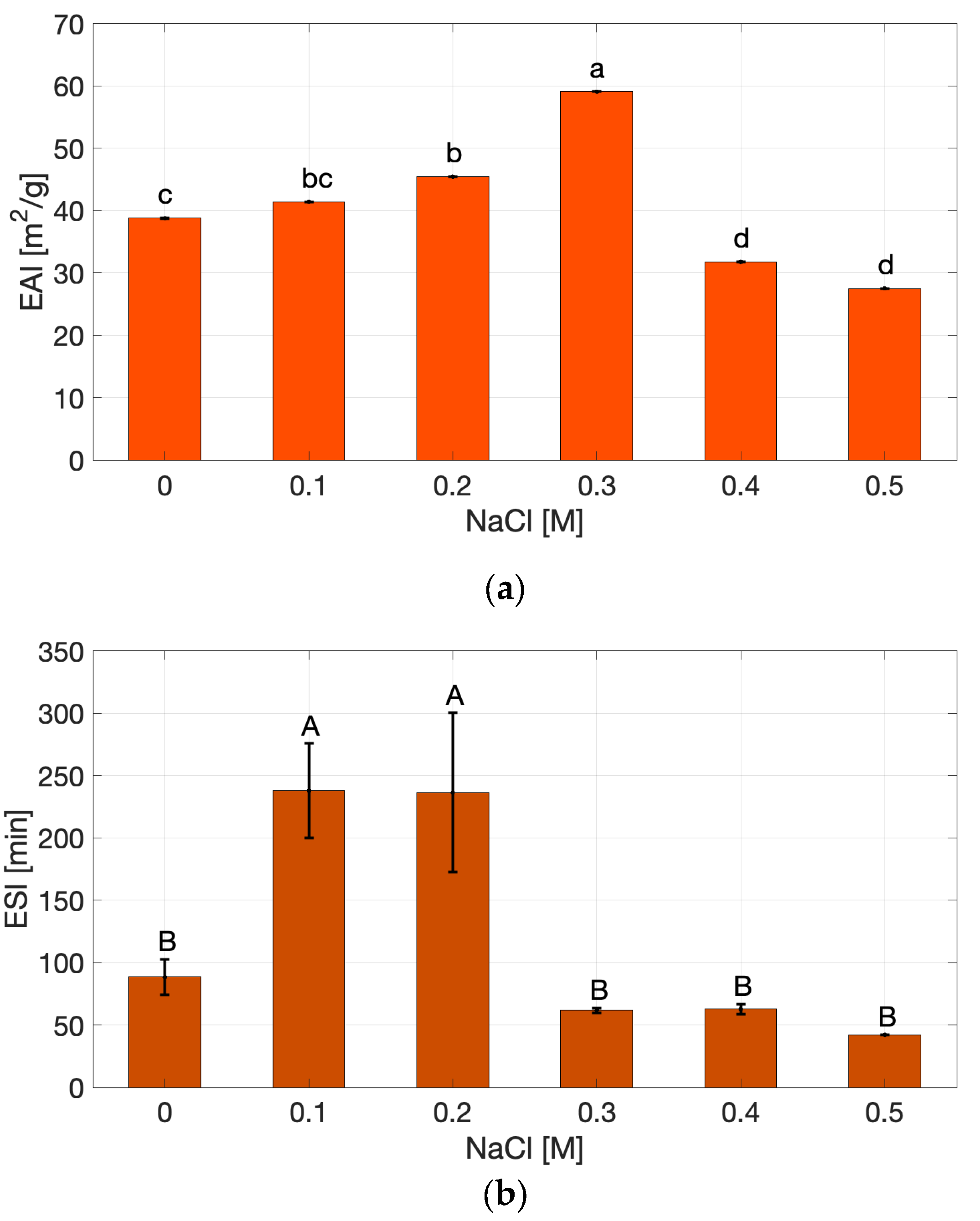
Figure 4.
Evolution of the storage modulus (G′) (a) and of the loss modulus (G”) (b) of cricket protein gels at varying NaCl concentrations during amplitude sweep tests.
Figure 4.
Evolution of the storage modulus (G′) (a) and of the loss modulus (G”) (b) of cricket protein gels at varying NaCl concentrations during amplitude sweep tests.
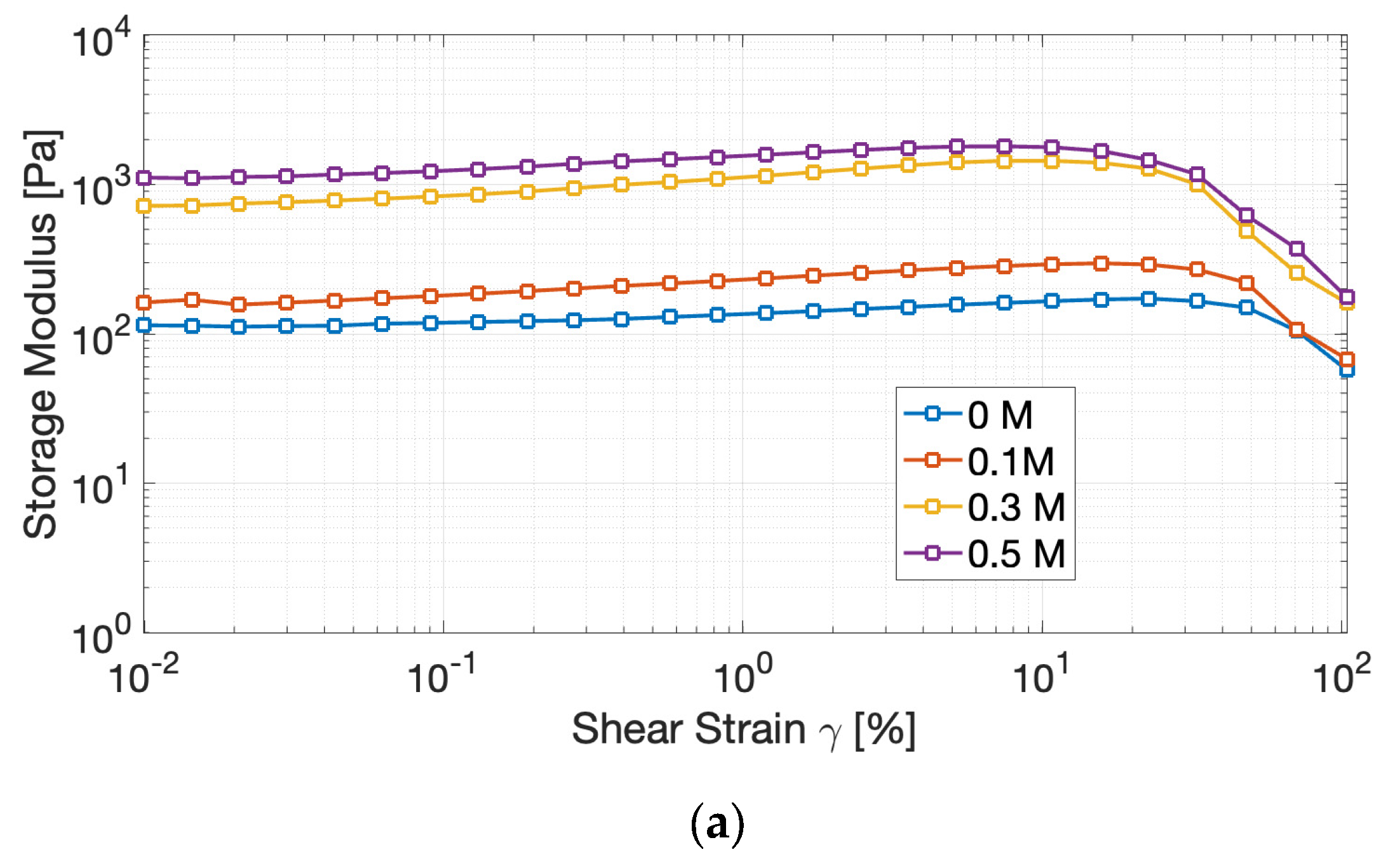
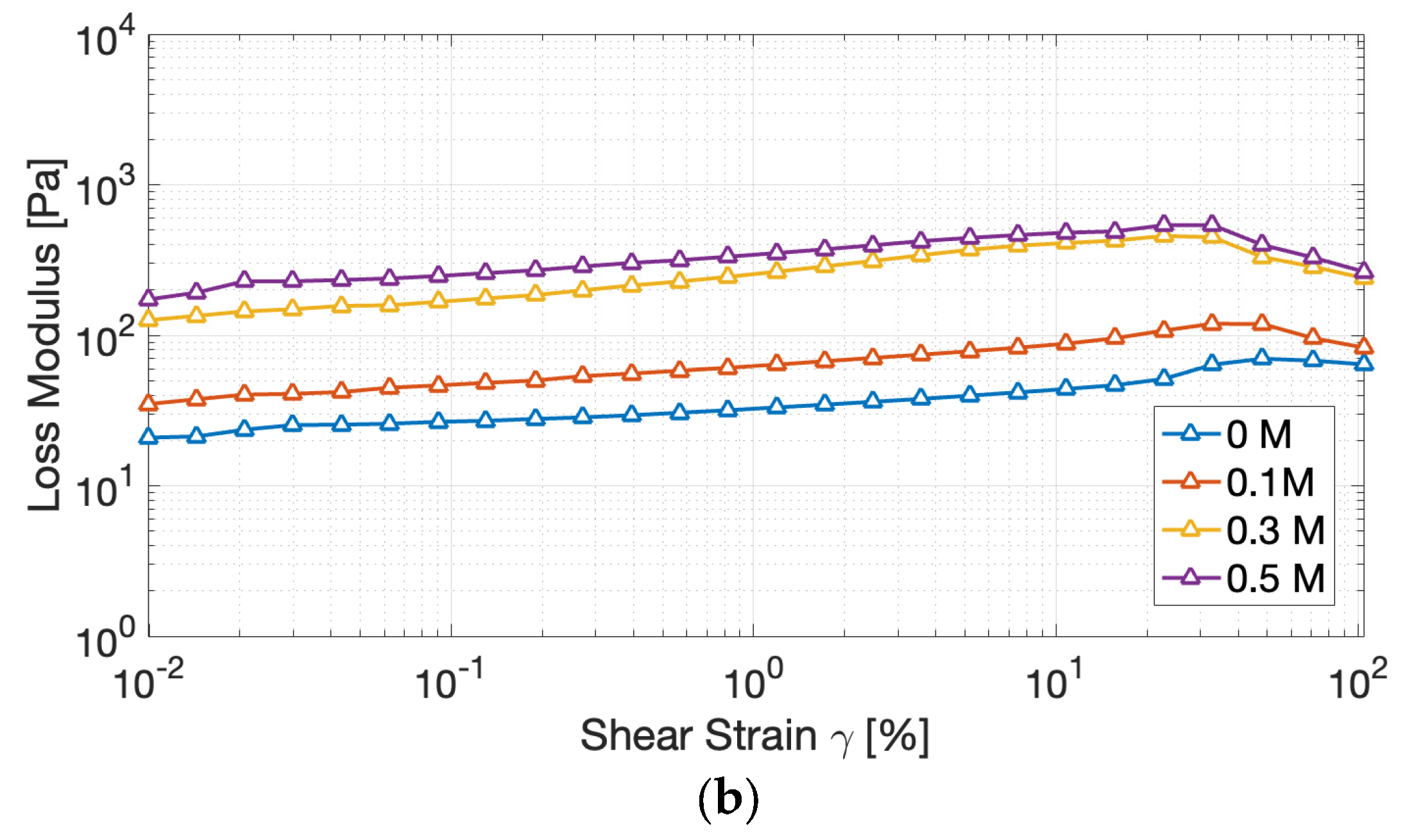
Figure 5.
Evolution of the storage modulus (G′) (a) and of the loss modulus (G”) (b) of cricket protein gels at varying NaCl concentrations during frequency sweep tests.
Figure 5.
Evolution of the storage modulus (G′) (a) and of the loss modulus (G”) (b) of cricket protein gels at varying NaCl concentrations during frequency sweep tests.
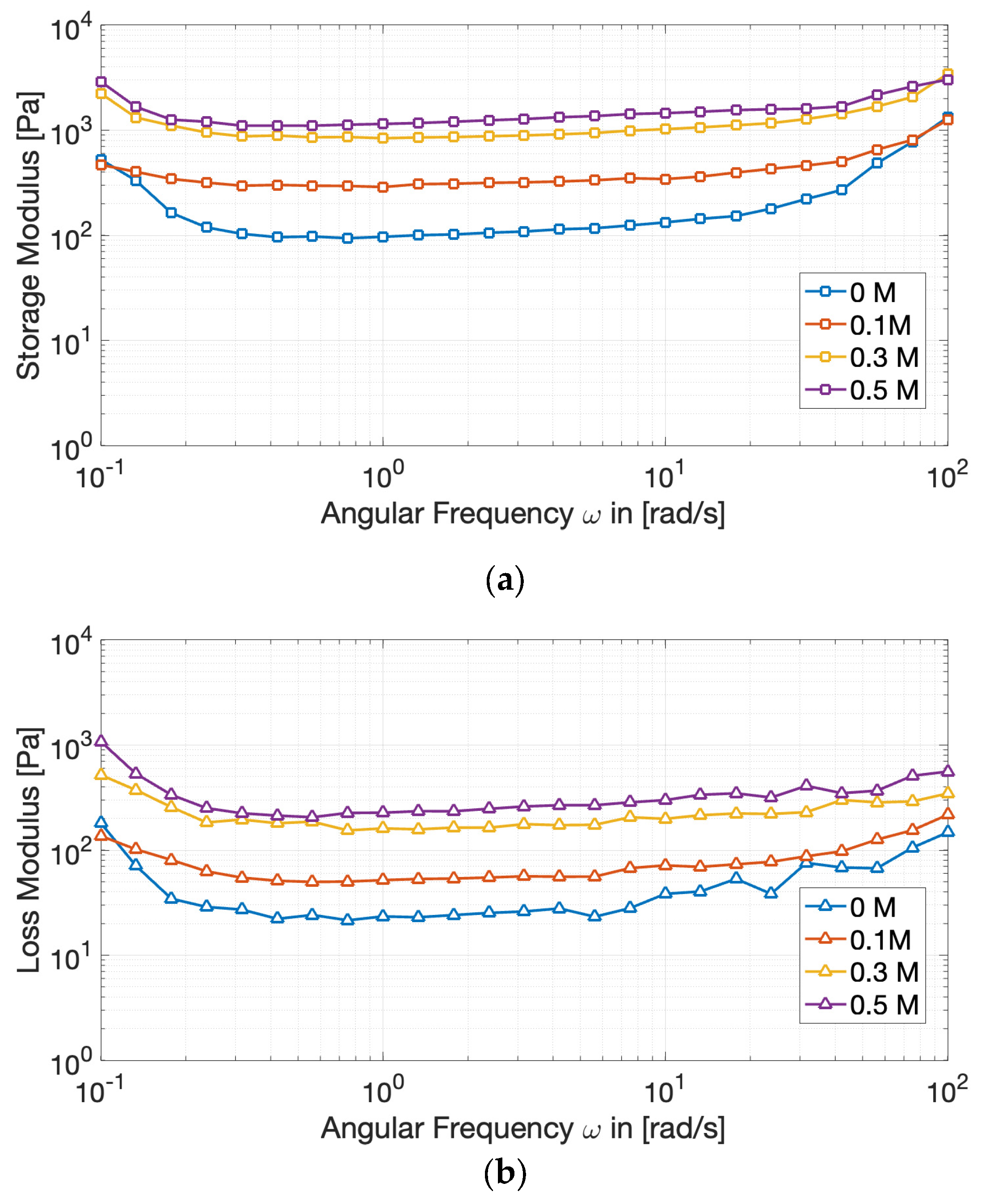
Figure 6.
Variation of the power-law parameters with NaCl concentration: (a) consistency index k′, (b) frequency-dependence exponent n′, (c) consistency index k″, and (d) frequency-dependence exponent n″. Symbols represent experimental data, and solid lines correspond to the fitted correlations.
Figure 6.
Variation of the power-law parameters with NaCl concentration: (a) consistency index k′, (b) frequency-dependence exponent n′, (c) consistency index k″, and (d) frequency-dependence exponent n″. Symbols represent experimental data, and solid lines correspond to the fitted correlations.
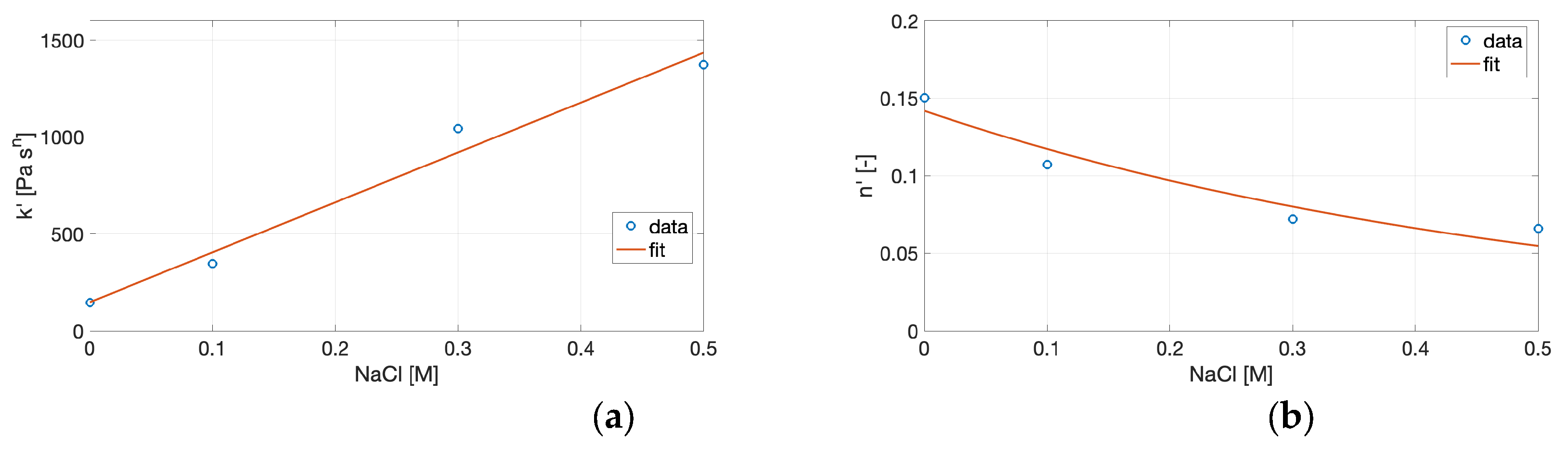
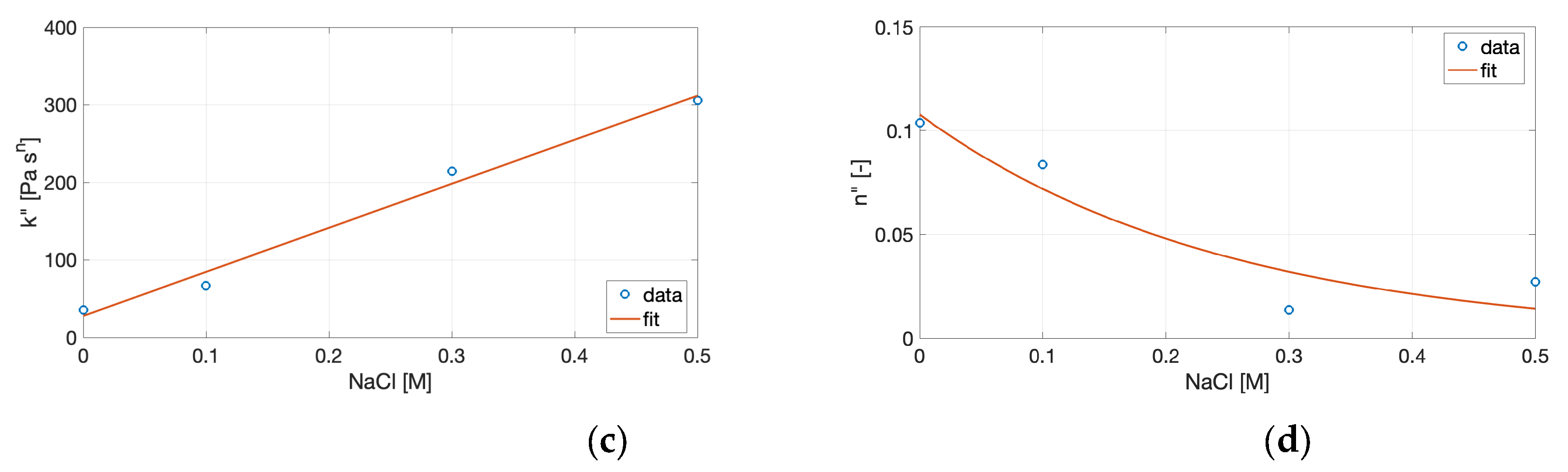

Table 1.
Loss factor values for cricket protein extract gels at different NaCl concentrations.
| NaCl [M] |
tan δ [-] |
| 0 | 0.31 ± 0.14A |
| 0.1 | 0.28 ± 0.06AB |
| 0.3 | 0.19 ± 0.04C |
| 0.5 | 0.24 ± 0.08BC |
Values followed by different letters (A-C) are significantly different (p < 0.05).
Table 2.
Parameters of the power law model for cricket protein extract gels at different NaCl concentrations.
Table 2.
Parameters of the power law model for cricket protein extract gels at different NaCl concentrations.
| NaCl [M] |
k’ [Pa⋅sn] |
k” [Pa⋅sn] |
n’ [-] |
n” [-] |
| 0 | 145.54 | 35.43 | 0.15 | 0.1 |
| 0.1 | 343.43 | 66.91 | 0.11 | 0.08 |
| 0.3 | 1042.90 | 214.50 | 0.07 | 0.01 |
| 0.5 | 1372.10 | 305.61 | 0.06 | 0.03 |
Table 3.
Fitting parameters of the empirical correlations between the power-law coefficients (k′, k″, n′, n″) and NaCl concentration.
Table 3.
Fitting parameters of the empirical correlations between the power-law coefficients (k′, k″, n′, n″) and NaCl concentration.
| Parameter | a | b | R2 | c | d | R2 |
| k’ | 2577.60 | 146.64 | 0.977 | - | - | - |
| k” | 568.12 | 27.78 | 0.986 | - | - | - |
| n’ | - | - | - | 0.14 | -1.91 | 0.826 |
| n” | - | - | - | 0.11 | -4.05 | 0.775 |
Table 4.
Nutritional composition of the Acheta domesticus flour as stated by the manufacturer.
| Nutrient (per 100 g) |
Value [g] |
| Fat | 11.60 |
| Carbohydrates | 4.60 |
| Fiber | 8.80 |
| Protein | 74.60 |
| Salt | 0.395 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



