Submitted:
17 October 2025
Posted:
20 October 2025
You are already at the latest version
Abstract
Pancreatic cancer often goes undetected in its early stages due to minimal or no symptoms, leading to late diagnosis with limited treatment options. Challenges include late detection, drug resistance, and the tumor's complexity, but advances are being made in targeted therapies, immunotherapy, metabolism-based strategies, and early detection techniques. Current treatments focus on boosting immune responses, improving survival rates, and enhancing patients' quality of life. Immunotherapy has sparked interest in cell-based approaches, with studies showing that natural killer (NK) cells can eliminate pancreatic tumor stem-like cells and promote tumor differentiation through cytokines. NK cells play a key role in tumor destruction, though reduced cytotoxic activity has been observed in NK cells from pancreatic cancer patients’ blood. Probiotics are being investigated as potential aids in pancreatic cancer therapy to restore immune function, inhibit tumor growth, and improve treatment outcomes. By activating immune cells like PBMCs, NK cells, T cells, and antigen-presenting cells, probiotics can help combat pancreatic tumors. They also enhance NK cell-based immunotherapies and improve the effectiveness of immunotherapy, radiation, and chemotherapy. Combining probiotics with NK cell therapy has shown promise in reducing tumor burden, restoring immune function, and reversing tumor-induced bone damage in pancreatic cancer models, offering hope for future treatments.
Keywords:
pancreatic cancer
; NK cells
; probiotics
; immunotherapy
; immune cells
; cytotoxicity
; IFN-γ
1. Introduction and Background
Pancreatic cancer is an aggressive disease often diagnosed late due to its vague early symptoms, diverse tumor types, and poor prognosis, with a 5-year survival rate of less than 10% [1,2]. Early detection is tough because symptoms usually arise only after the tumor grows significantly or spreads. It’s been one of the top causes of cancer-related deaths worldwide, with little improvement in survival rates over the years [1]. Common symptoms include fatigue, weakness, unexplained weight loss, abdominal pain radiating to the back, jaundice, digestive problems like nausea or bloating, new-onset diabetes, and blood clots. Tumors in the pancreas head often cause early jaundice from bile duct obstruction, while tumors in the body or tail typically result in pain and weight loss later, often after metastasis [3]. The causes are not fully understood but are linked to genetic mutations, hereditary factors, smoking, obesity, heavy alcohol use, chronic pancreatitis, aging (65-74 years), family history, chemical exposure, and diabetes [3,4]. Diagnosis usually involves imaging tests like computed tomography (CT) scans, magnetic resonance imaging (MRI), positron emission tomography (PET) scans, and endoscopic ultrasound. Blood tests for markers like cancer antigen 19-9 (CA 19-9) can help monitor the disease, but aren’t definitive, and biopsies are often guided by imaging [1,5].
Most pancreatic tumors (90-95%) are exocrine, primarily pancreatic adenocarcinomas (PDAC), which arise from enzyme-producing cells near the ducts, grow rapidly, and spread early [6]. The remaining 5-10% are pancreatic neuroendocrine tumors (PNETs), originating from hormone-producing cells, growing more slowly, and offering a better prognosis [7]. PDAC progresses due to a mix of genetic mutations, disrupted signaling pathways, tumor microenvironment interactions, and cellular adaptability [6,8]. Key genetic drivers include KRAS mutations, found in about 95% of PDAC cases, which activate growth and survival pathways, along with mutations in tumor suppressor genes like TP53, CDKN2A (p16), and SMAD4, causing uncontrolled cell growth, avoidance of cell death, and tumor development [8,9]. Early detection significantly improves outcomes, and PNETs generally have better survival rates than adenocarcinomas [9]. Pancreatic cancer patients often face severe nutritional deficiencies, muscle wasting, and bone issues, with the first case of bone metastasis documented in Russian literature in 1963 [10]. Pancreatic cancer often spreads to the liver, peritoneal cavity, lungs, bones, adrenal glands, and brain [10,11,12,13,14,15,16]. The liver (50-70%) is the primary site for metastasis due to portal circulation, worsening the prognosis [11,13]. The peritoneum (around 50% in advanced cases) often develops carcinomatosis, causing symptoms and limiting treatment options [12]. Pulmonary metastases (15-30%) affect survival as tumor cells become trapped in the lung’s capillaries. Bone metastases (10-20%) are less common but cause pain and fractures [10]. Adrenal gland involvement (5-10%) is rare but can disrupt hormonal balance [14]. Brain metastases, occurring in less than 1% of cases, are a rare complication with poor outcomes [15,16].
The pancreatic tumor microenvironment (TME) is characterized by a dense fibrotic stroma, cancer-associated fibroblasts (CAFs), immune-suppressive cells like regulatory T cells and myeloid-derived suppressor cells, extracellular matrix (ECM), and cytokines [17,18,19]. This environment facilitates tumor growth, hinders drug delivery, enables immune evasion, and promotes metastasis [20]. Secreted factors and cytokines, such as TGF-β, further modify the microenvironment to foster tumor growth, fibrosis, angiogenesis, metastasis, and immune suppression [9,21]. Treatment for pancreatic cancer depends on the stage, location, and the patient’s overall health [22]. Surgery, including the Whipple procedure, distal pancreatectomy, or total pancreatectomy, offers the only potential cure but is limited to localized cases [22,23]. Chemotherapy (e.g.; FOLFIRINOX, gemcitabine) and radiation therapy are used to control the disease and shrink tumors [22]. Other strategies involve MAPK pathway inhibitors like Trametinib and Binimetinib, KRAS-specific inhibitors such as Sotorasib and Adagrasib, and immunotherapy, though immune evasion remains a challenge [22,24]. Targeted therapies and immunotherapies hold promise for specific genetic subtypes or advanced cases, while palliative care focuses on relieving symptoms, particularly pain [22]. Strategies to modify TME, using a combination of CAR-T with immune checkpoint inhibitors and cytokine modulations, are also under investigation to boost pancreatic cancer therapeutics [25]. The effectiveness of natural killer (NK) cell-based therapies has been demonstrated in various solid tumors [26,27,28,29,30,31].
NK cells play a key role in inhibiting pancreatic cancer through their effector functions, such as direct cytotoxicity and antibody-dependent cellular cytotoxicity (ADCC). They also regulate or activate the anti-cancer activity of other immune effectors through the cytokines and chemokines they secrete [32,33,34]. As vital components of innate immunity, NK cells identify and destroy tumor and virus-infected cells without prior sensitization [35,36,37]. The proper functioning of NK cells or NK cell-based therapies is crucial for improving the prognosis of pancreatic cancer patients and reducing tumor relapse [38,39,40,41,42,43,44,45,46,47]. Maintaining balanced numbers and anti-cancer activity of peripheral blood-derived NK cells, along with increased NK cell infiltration in tumor tissues, significantly enhances patient outcomes [48,49,50,51,52]. However, studies have shown reduced numbers and diminished anti-cancer activity of NK cells in cancer patients, both in tumor tissues and peripheral blood-derived immune subsets [48,49,50,51,52]. To address this, several technologies have been developed to expand and activate NK cells, enabling large-scale production for therapeutic use [53,54,55,56,57]. This review focuses on combining probiotics with feeder cells to promote significant NK cell expansion and enhance their anti-cancer functionality [51,58,59,60].
Probiotics, discovered by Elie Metchnikoff in the early 20th century, are crucial for maintaining gut balance [61,62]. They may influence pancreatic cancer by impacting gut microbiota, immune responses, and tumor pathways, though the evidence is mixed and context-dependent, with risks like potentially promoting tumor growth [63,64,65]. In vivo studies show probiotic treatments can enhance tumor immune cell recruitment, boost anti-cancer activity, improve PBMC cytotoxicity and cytokine secretion, and reduce tumor burden [59,66]. Clinical trials reveal better immune markers and therapy outcomes in cancer patients using probiotics [63]. Some probiotic regimens, combined with standard therapy, have been shown to improve survival, reduce complications, and enhance quality of life [63]. Host factors like baseline microbiome, genetics, diet, and immune status also play a significant role in these effects [63].
This review discusses how activating immune cells through probiotics can directly or indirectly contribute to the breakdown and inhibition of pancreatic cancer growth. Combining probiotic treatments with feeder cells has been shown to promote NK cell expansion and functional activity. Probiotic-based NK cell therapies have demonstrated enhanced lifespan, cell proliferation, cytotoxic effects, and cytokine secretion, improving the ability to target and destroy pancreatic cancer both in vivo and in vitro. Administering probiotics orally, alongside surgery, radiotherapy, chemotherapy, and NK cell therapy, has shown additive benefits in reducing tumor burden, restoring immune function, and preventing bone loss in pancreatic cancer models in mice or humans.
2. Probiotics Boost the Ability of Immune Cells to Fight Cancer
Probiotics help balance the gut microbiome, activate peripheral blood mononuclear cells (PBMCs), reduce pathogen-induced inflammation, and lower exposure to systemic carcinogens [67]. They support the intestinal barrier and microbial balance by boosting proteins and mucus that block pathogens and prevent excessive immune reactions [68]. Through direct interaction and signaling with immune cells, probiotics enhance immunity, strengthen anti-cancer properties, and maintain gut tolerance [68,69]. Their benefits are strain-specific, as outlined in Table 1, with beneficial formulations including Lactobacillus and Bifidobacterium [67,69].
PBMCs or PBMCs-derived lymphocytes, such as T cells and NK cells, are vital for anti-tumor immunity [99,100]. Probiotics, often Lactobacillus and Bifidobacterium, interact directly or indirectly with PBMCs to enhance their anti-tumor effects [98] (Figure 1). These bacteria produce short-chain fatty acids (SCFAs) like butyrate, acetate, and propionate, which promote PBMCs or PBMCs-derived NK cells and T cells-mediated apoptosis of pancreatic cancer via Fas/FasL pathways and support T-cell differentiation and activation [101,102,103]. Probiotics also create bioactive compounds like bacteriocins and exopolysaccharides (EPS) that boost PBMC activation and selectively target tumor cells [104]. Enhanced PBMC cytotoxicity can halt tumor cell cycles at G0/G1 or G2/M phases, inhibiting tumor growth and metastasis [105]. Probiotic treatments increase anti-angiogenic cytokines, particularly IFN-γ and TNF-α in PBMCs, NK cells, and T cells (CD8+. CD4+ and γδ T cells); these cytokines can inhibit tumor growth by promoting differentiation [70]. Differentiated tumors proliferate more slowly and express more surface molecules like major histocompatibility complex-class I (MHC-I), CD54, and PD-L1, improving immune recognition and reducing tumor-supportive microenvironments [70,106].
Probiotics, especially lactic acid bacteria like Lactobacillus and Bifidobacterium, provide significant immunomodulatory benefits by enhancing NK cell-driven anti-cancer responses [70]. Probiotic-treated NK cells exhibit increased cytolytic activity against cancer cells, producing more perforins and granzyme B for precise cancer cell targeting [70]. They activate NK cells through cytokine stimulation, promoting tumor differentiation, growth inhibition, and improved immune recognition via upregulated expression of MHC-class I, CD54, and PD-L1 [70]. Strains like Lactobacillus paracasei support Th1- and Th17-mediated anti-tumor immunity, indirectly activating NK cells [70]. Probiotics also stimulate dendritic cells (DCs) and other antigen-presenting cells to present antigens more effectively, bolstering NK cells and T cells’ anti-cancer functions [107]. They enhance DC maturation by upregulating CD80 and CD86, improving antigen presentation to naive T cells. Activated DCs release cytokines like IL-10, IL-12, and TGF-β, guiding naive T cells to differentiate into regulatory T cells (Tregs) to suppress excessive inflammation [108,109]. Effector T cells, including Th1 and Th17, play a role in anti-tumor immunity and work in synergy with chemotherapy drugs like cyclophosphamide and gemcitabine [110].
Probiotic treatment in PBMCs shows increased CD8+ T cell/Treg cell ratios, higher IFN-γ production by NK, CD4+ T, and γδ T cells, improved tumor recognition, and enhanced cytotoxicity and tumor growth inhibition via differentiation. These effects may boost cancer therapy by restoring immune surveillance, suppressing tumor progression, and improving outcomes. Additionally, these mechanisms amplify the effectiveness of conventional therapies, including immunotherapy [111,112,113] (Figure 1).
3. Probiotics, When Combined with Feeder Cells, Contribute to Improving the Development of NK Cell-Based Immunotherapies
Natural killer (NK) cells, known for their strong anti-tumor activity, can effectively target cells that lack or have altered major histocompatibility complex (MHC) class I molecules [114,115]. While NK cell-based therapies are considered safe, their effectiveness is limited due to challenges in enhancing their expansion and anti-tumor capabilities [116,117]. Autologous NK cells from pancreatic cancer patients often face impairments caused by prior immunosuppression, making their expansion more difficult compared to those from healthy donors [118,119,120]. Studies show that NK cells expanded from pancreatic cancer patients demonstrate lower cytotoxicity and reduced IFN-γ secretion [121]. These cells also exhibit decreased levels of activating receptors like CD16, CD56, Nkp30, Nkp44, Nkp46, NKG2D, and CD54 [121]. To address these issues, healthy allogeneic NK cells are being explored for pancreatic cancer therapies and are under clinical investigation [122]. These allogeneic cells, sourced from peripheral blood, cord blood, hematopoietic stem cells, or induced pluripotent stem cells, can be expanded and cryopreserved for convenient use [123,124,125]. Several methods have been developed to expand NK cells ex vivo, often using feeder cells with or without cytokines and other activation signals [53,55,56,126,127,128,129,130,131,132]. Feeder cells play a vital role in activating and proliferating NK cells by providing receptor-ligand interactions and cytokine support, enabling large-scale therapeutic applications [132,133,134,135]. Cytokines like IL-2, IL-15, and IL-21, along with membrane-bound cytokines and antibodies, are commonly used to supplement feeder cell cultures [136,137]. When there’s a donor-recipient mismatch, donor NK cells with inhibitory receptors like KIR don’t recognize HLA class I on recipient cells. This lack of recognition activates the donor NK cells, allowing them to attack cancer cells that lack the MHC ligands needed for KIR inhibition. As a result, alloreactive NK cells destroy these cancer cells [122,138,139]. Engineered or feeder-expanded NK cells, with enhanced KIR expression, increased cytokine secretion, improved ADCC through elevated levels of CD16 and NKG2D, reduced inhibitory receptors like NKG2A, and the ability to kill tumor cells regardless of MHC-class I expression, show great promise as allogeneic cancer therapies [132,140,141,142,143]. Additionally, the absence of graft-versus-host disease (GVHD) in NK cells ensures their safety in adoptive cell-based therapies [144,145].
Combining probiotics with feeder cells has been shown to enhance the effectiveness of NK cell therapy compared to using feeder cells alone (Figure 2) [132]. Selected probiotic strains like Streptococcus thermophilus, Bifidobacterium longum, Bifidobacterium breve, Bifidobacterium infantis, Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus paracasei, and Lactobacillus bulgaricus boosted NK cell activation and cytokine secretion [70]. NK cells expanded with probiotics and feeder cells displayed improved expansion, longer lifespan, increased cytotoxicity, and higher cytokine secretion compared to those expanded with feeder cells alone [132,146]. These expanded NK cells also showed a greater ability to induce pancreatic tumor killing and differentiation in both in vivo and in vitro studies [59,66]. They exhibited enhanced survival in the pancreatic tumor microenvironment due to higher anti-apoptotic protein levels like BCL2 and reduced pro-apoptotic proteins, allowing them to resist tumor-induced cell death. Elevated cytotoxic granules and Trail expression further improved their cytotoxic function [66,140]. Additionally, these expanded NK cells showed better regulatory functions, with most cells in an active cycling phase, increased expression of proliferation- and memory-associated genes, and enhanced activating receptors (e.g., CD16, CD56, Nkp30, Nkp44, Nkp46, NKG2D) while reducing inhibitory receptors like NKG2A, PD-1, and TIGIT [132,140]. Compared to NK cells expanded without probiotics, those expanded with probiotics demonstrated remarkable anti-cancer properties [132] (Figure 2).
The effectiveness of probiotic combined with feeder cells expanded NK cells therapies has been shown in vivo using humanized mouse models, highlighting their potential for clinical use [59,66,146]. Infusing these expanded NK cells into healthy and pancreatic cancer-bearing humanized mice over eight weeks resulted in no toxicity, pain, distress, or adverse events such as cytokine-release syndrome (CRS) or immune effector cell-associated neurotoxicity syndrome (ICANS) [66]. Combining this NK cell-based therapy with chemotherapy or checkpoint inhibitors in pancreatic tumor-bearing humanized mice enhanced the effectiveness of both treatments [146]. Advances in NK cell expansion techniques have opened up exciting new therapeutic possibilities for pancreatic cancer [55,147].
4. Benefits of Incorporating Probiotics as an Additional Therapy in the Treatment of Pancreatic Cancer
Several probiotics are being studied in preclinical and clinical settings for their potential benefits in cancer patients. A diet rich in probiotics has been linked to reduced rates of pancreatitis and pancreatic cancer [148,149]. Probiotics like Lactobacillus and Bifidobacterium spp. help maintain gut microbial balance, strengthen intestinal barriers, and decrease pro-inflammatory microbial products reaching the pancreas [91,150]. Animal studies indicate they may reduce pancreatic inflammation, suppress enzyme activity, and enhance proteins like occludin and claudins to prevent tumors [151]. Probiotics can also boost immune responses and promote cancer cell death, with compounds like heptelidic acid from Aspergillus oryzae and ferrichrome from Lactobacillus casei showing tumor-inhibiting properties [152]. Ferrichrome from Lactobacillus casei, an iron chelate derivative, has demonstrated antitumor effects in refractory and 5-fluorouracil-resistant pancreatic cancer by regulating the tumor cell cycle through p53 activation [152,153]. They may also inhibit angiogenesis, EMT, and metastasis, potentially slowing tumor growth. Using Lactobacillus plantarum to ferment stevia extract generated bioactive metabolite clonogenic acid methyl ester (CAME) was found to arrest pancreatic cancer cells (PANC-1) in the G0/G1 phase and induce apoptosis suggesting its role as pancreatic cancer therapeutic[88]. The effects on pancreatic cancer depend on specific strains and the overall context, making strain selection crucial for achieving therapeutic goals (Table 1).
Short-chain fatty acids (SCFAs) derived from probiotics, like butyrate, acetate, and propionate, can activate tumor suppressor genes epigenetically and inhibit pancreatic cancer cell invasion and metastasis by reducing integrin β4 expression or inhibiting histone deacetylase (HDAC) activity [149,154,155]. Probiotic treatments also decrease VEGF expression in tumors, limiting angiogenesis, promoting differentiation, and inducing tumor-antigen surface expression in pancreatic tumors [156]. Strains such as Lactobacillus casei and Lactobacillus reuteri show promise in suppressing pancreatic tumors by downregulating TLR4, encouraging macrophage M1 polarization, and supporting gut microbial balance [85,105]. Probiotics may also reduce cancer cell growth, prevent PanIN progression, and curb metastasis through mechanisms possibly involving the TGF-β signaling pathway [157]. Probiotic exposure, including strains like Lactobacillus rhamnosus, Lactobacillus helveticus, Lactobacillus casei, and Saccharomyces boulardii, has been associated with improved survival in pancreatic cancer patients under palliative care [158]. Additionally, patients consuming Streptococcus thermophiles, Bifidobacterium strains, Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus paracasei, and Lactobacillus bulgaris (125 billion CFU per capsule, three times daily for four weeks) demonstrated enhanced IFN-γ levels and increased cytotoxic activity in PBMCs and NK cells [159].
Probiotics as an additional treatment for pancreatic conditions are still in the early stages of research, with limited data available. Common strains like Bifidobacteria and Lactobacillus are often used in multistrain formulations, with optimal doses generally ranging from 108 to over 1010 CFU daily [160,161,162,163,164]. Recent studies suggest potential benefits in pancreatic cancer for both mouse models and humans, especially when combined with surgery, chemotherapy, radiotherapy, or NK cell-based therapies [66,105,160,165,166]. In mice, probiotics have been shown to enhance drug effectiveness, lessen chemotherapy side effects, and slow tumor progression [152]. For human patients on palliative chemotherapy, probiotics are associated with improved survival outcomes. They also help reduce gastrointestinal side effects from aggressive therapies, boosting treatment adherence and quality of life [105,160,165,166]. Additionally, probiotics have been linked to increased tumor-infiltrating CD8+ T cells, higher IFN-γ expression, reduced inflammatory cytokines, and fewer postoperative complications like anastomotic leakage and bacteremia[59,66,167].
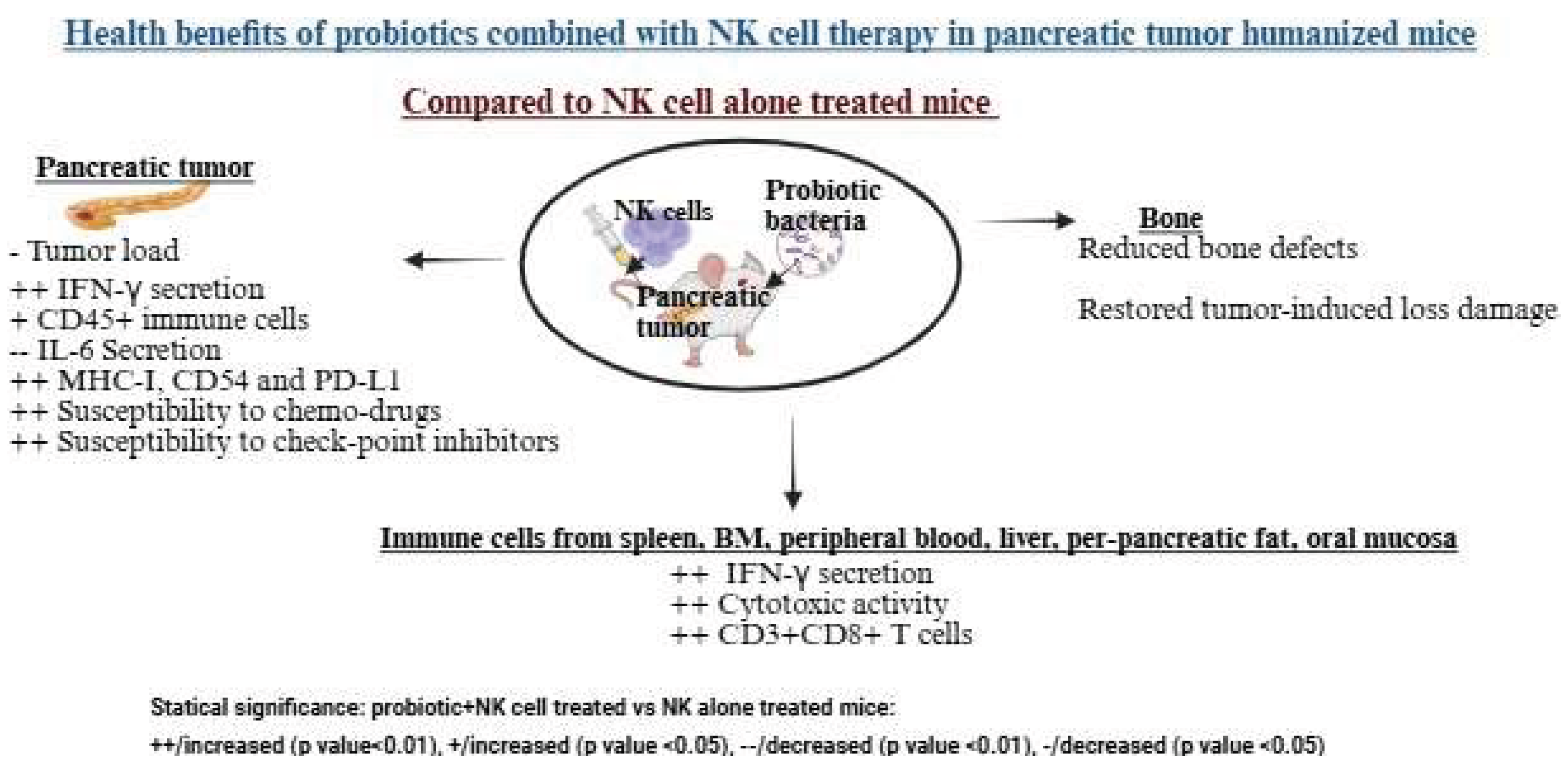
Oral administration of Streptococcus thermophiles, Bifidobacterium longum, Bifidobacterium breve, Bifidobacterium infantis, Lactobacillus acidophilus, Lactobacillus plantarum, and Lactobacillus paracasei, combined with NK cell therapy, significantly reduced tumor growth in pancreatic tumor-bearing humanized mice [59,66]. This combination enhanced NK cell activation in vivo, leading to tumors with increased PD-L1, CD54, and MHC-class I expression, slower growth, and greater sensitivity to checkpoint inhibitors and chemotherapy [59]. These changes likely improved CD8+ T cell-induced killing due to higher MHC-class I expression. Treated mice showed increased infiltration of human CD45+ immune cells, elevated IFN-γ secretion, and reduced IL-6 secretion in the tumor microenvironment, emphasizing the therapeutic benefits of probiotics and NK cells [59]. Probiotics also boosted tumor-infiltrating lymphocyte activity, particularly CD8+ T cell recruitment and cytotoxicity, enhancing cancer-specific immunity. Immune cells from peripheral blood, bone marrow, spleen, pancreas, oral mucosa, liver, and peri-pancreatic fatty tissue in probiotic and NK cell-treated mice exhibited higher cytotoxicity and cytokine secretion compared to those treated with NK cells alone [59]. Additionally, combining probiotics with NK cell therapy not only inhibited tumors but also reduced pancreatic tumor-induced bone defects and restored bone integrity [66].
Probiotics could potentially enhance the effectiveness of standard therapy by reducing immunosuppression and inflammation in the tumor microenvironment. However, clinical validation is necessary to identify the best strains, dosage, duration, and combinations with therapy. While adverse events are uncommon, they can occur in immunocompromised individuals, so their use requires careful consideration, especially for those who are immunocompromised or have metastatic conditions.
Figure 3.
The combination of probiotics and NK cells improved immune function and reduced tumor load in a pancreatic tumor-bearing mouse model. These mice were orthotopically injected with human pancreatic tumors into the pancreas. One to two weeks after tumor implantation, they received NK cells via tail-vein injection and were given 5 billion CFU probiotics orally every 48 hours, starting a week before tumor implantation. At the end of the experiment, hu-BLT mice were sacrificed, and their tissues and tumors were collected and analyzed. Tumors from the probiotic and NK cell-treated group showed enhanced immune function, immune cell infiltration, and differentiation compared to the NK cell-only group. Tissues from the probiotic and NK cell-treated group had higher immune function and a greater percentage of CD3+CD8+ T cells compared to the NK cell-only group. Bones from the probiotic and NK cell-treated group displayed fewer defects and showed recovery from tumor-induced bone loss compared to the NK cell-only group. Illustration created with https://BioRender.com on September 26, 2025. https://app.biorender.com/illustrations/68cedfb1c9e8ad7e0f109b30?slideId=b6075359-d4a7-4a6b-8d8d-1b825340710f.
Figure 3.
The combination of probiotics and NK cells improved immune function and reduced tumor load in a pancreatic tumor-bearing mouse model. These mice were orthotopically injected with human pancreatic tumors into the pancreas. One to two weeks after tumor implantation, they received NK cells via tail-vein injection and were given 5 billion CFU probiotics orally every 48 hours, starting a week before tumor implantation. At the end of the experiment, hu-BLT mice were sacrificed, and their tissues and tumors were collected and analyzed. Tumors from the probiotic and NK cell-treated group showed enhanced immune function, immune cell infiltration, and differentiation compared to the NK cell-only group. Tissues from the probiotic and NK cell-treated group had higher immune function and a greater percentage of CD3+CD8+ T cells compared to the NK cell-only group. Bones from the probiotic and NK cell-treated group displayed fewer defects and showed recovery from tumor-induced bone loss compared to the NK cell-only group. Illustration created with https://BioRender.com on September 26, 2025. https://app.biorender.com/illustrations/68cedfb1c9e8ad7e0f109b30?slideId=b6075359-d4a7-4a6b-8d8d-1b825340710f.
5. Conclusion
This review highlights the significance of probiotics, particularly the strains listed in Table 1, in activating immune cells or signals to fight pancreatic cancer, their role in advancing cancer therapies, and enhancing the effectiveness of treatments. It emphasizes the benefits of probiotic bacteria and combination therapies, especially NK cell-based treatments, in reducing pancreatic tumor growth and spread. Probiotics offer advantages as a supplementary therapy, including gut health support, improved immune response, and fewer side effects from conventional treatments. Additionally, oral probiotics, alone or with NK cell infusions, help prevent tumor-related bone damage. These findings suggest that probiotics, whether used alone or alongside surgery, radiation, chemotherapy, and immunotherapies, hold promise for treating pancreatic cancer.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare that the work reviewed in the article was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Gheorghe, G.; et al. Early Diagnosis of Pancreatic Cancer: The Key for Survival. Diagnostics (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Eissa, M.A.L.; et al. Promoter methylation of ADAMTS1 and BNC1 as potential biomarkers for early detection of pancreatic cancer in blood. Clin Epigenetics 2019, 11, 59. [Google Scholar] [CrossRef]
- Singh, S.; Sawal, A. Comprehensive Review on Pancreatic Head Cancer: Pathogenesis, Diagnosis, and Treatment Challenges in the Quest for Improved Survival. Cureus 2024, 16, e54290. [Google Scholar] [CrossRef]
- Grigorescu, R.R.; Husar-Sburlan, I.A.; Gheorghe, C. Pancreatic Cancer: A Review of Risk Factors. Life (Basel) 2024, 14. [Google Scholar] [CrossRef]
- Lee, E.S.; Lee, J.M. Imaging diagnosis of pancreatic cancer: a state-of-the-art review. World J Gastroenterol 2014, 20, 7864–77. [Google Scholar] [CrossRef] [PubMed]
- Luchini, C.; et al. Malignant epithelial/exocrine tumors of the pancreas. Pathologica 2020, 112, 210–226. [Google Scholar] [CrossRef]
- Halfdanarson, T.R.; et al. Pancreatic neuroendocrine tumors (PNETs): incidence, prognosis and recent trend toward improved survival. Ann Oncol 2008, 19, 1727–33. [Google Scholar] [CrossRef]
- Javadrashid, D.; et al. Pancreatic Cancer Signaling Pathways, Genetic Alterations, and Tumor Microenvironment: The Barriers Affecting the Method of Treatment. Biomedicines 2021, 9. [Google Scholar] [CrossRef]
- Truong, L.H.; Pauklin, S. Pancreatic Cancer Microenvironment and Cellular Composition: Current Understandings and Therapeutic Approaches. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Borad, M.J.; et al. Skeletal metastases in pancreatic cancer: a retrospective study and review of the literature. Yale J Biol Med 2009, 82, 1–6. [Google Scholar]
- Liu, Z.; Gou, A.; Wu, X. Liver metastasis of pancreatic cancer: the new choice at the crossroads. Hepatobiliary Surg Nutr 2023, 12, 88–91. [Google Scholar] [CrossRef]
- Avula, L.R.; Hagerty, B.; Alewine, C. Molecular mediators of peritoneal metastasis in pancreatic cancer. Cancer Metastasis Rev 2020, 39, 1223–1243. [Google Scholar] [CrossRef]
- Bellon, E.; et al. Pancreatic cancer and liver metastases: state of the art. Updates Surg 2016, 68, 247–251. [Google Scholar] [CrossRef]
- Yachida, S.; Iacobuzio-Donahue, C.A. The pathology and genetics of metastatic pancreatic cancer. Arch Pathol Lab Med 2009, 133, 413–22. [Google Scholar] [CrossRef]
- Ma, Y.; et al. Characterization and treatment of brain metastases from pancreatic cancer: a systematic review. Discov Oncol 2025, 16, 1274. [Google Scholar] [CrossRef]
- Friedant, A.; Khurana, S.; Dissin, J. Rare Cerebral Manifestation of Pancreatic Adenocarcinoma: Insights from a Calcified Brain Metastasis (P1-6.009). Neurology 2025. 104(7_Supplement_1): p. 4549.
- Wang, K.; He, H. Pancreatic Tumor Microenvironment. Adv Exp Med Biol 2020. 1296, 243–257. [Google Scholar]
- Ho, W.J.; Jaffee, E.M.; Zheng, L. The tumour microenvironment in pancreatic cancer — clinical challenges and opportunities. Nature Reviews Clinical Oncology 2020, 17, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Carlomagno, S.; et al. Pancreatic ductal adenocarcinoma microenvironment: Soluble factors and cancer associated fibroblasts as modulators of NK cell functions. Immunology Letters 2024. 269, 106898. [CrossRef] [PubMed]
- Pan, D.; et al. Immunosuppressive tumor microenvironment in pancreatic cancer: mechanisms and therapeutic targets. Front Immunol 2025. 16, 1582305. [CrossRef]
- Ahmed, S.; et al. The TGF-β/Smad4 Signaling Pathway in Pancreatic Carcinogenesis and Its Clinical Significance. J Clin Med 2017, 6. [Google Scholar] [CrossRef]
- Wang, J.; et al. Consensus, debate, and prospective on pancreatic cancer treatments. J Hematol Oncol 2024, 17, 92. [Google Scholar] [CrossRef]
- Aoyama, T.; et al. Adjuvant Treatment for Resectable Pancreatic Cancer. Anticancer Res 2025, 45, 1329–1341. [Google Scholar] [CrossRef]
- Pergolizzi, R.G.; Brower, S.T. Molecular Targets for the Diagnosis and Treatment of Pancreatic Cancer. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Baghel, K.; Mehrotra, S.; Prajapati, V.K. Revolutionizing pancreatic cancer treatment with CAR-T therapy. Adv Protein Chem Struct Biol 2025. 144, 331–353.
- Ruggeri, L.; et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science 2002, 295, 2097–100. [Google Scholar] [CrossRef] [PubMed]
- Venstrom, J.M.; et al. HLA-C-dependent prevention of leukemia relapse by donor activating KIR2DS1. N Engl J Med 2012, 367, 805–16. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulou, E.G.; et al. A phase I trial of adoptive transfer of allogeneic natural killer cells in patients with advanced non-small cell lung cancer. Cancer Immunol Immunother 2010, 59, 1781–9. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S.; et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood 2005, 105, 3051–7. [Google Scholar] [CrossRef]
- Re, F.; et al. Killer cell Ig-like receptors ligand-mismatched, alloreactive natural killer cells lyse primary solid tumors. Cancer 2006, 107, 640–8. [Google Scholar] [CrossRef]
- Geller, M.A.; et al. A phase II study of allogeneic natural killer cell therapy to treat patients with recurrent ovarian and breast cancer. Cytotherapy 2011, 13, 98–107. [Google Scholar] [CrossRef]
- Palmer, J.M.; et al. Clinical relevance of natural killer cells following hematopoietic stem cell transplantation. J Cancer 2013, 4, 25–35. [Google Scholar] [CrossRef]
- Fildes, J.E.; Yonan, N.; Leonard, C.T. Natural killer cells and lung transplantation, roles in rejection, infection, and tolerance. Transpl Immunol 2008, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Farag, S.S.; Caligiuri, M.A. Human natural killer cell development and biology. Blood Rev 2006, 20, 123–37. [Google Scholar] [CrossRef]
- Moretta, A.; et al. NK cells at the interface between innate and adaptive immunity. Cell Death Differ 2008, 15, 226–33. [Google Scholar] [CrossRef]
- Vivier, E.; et al. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–9. [Google Scholar] [CrossRef]
- Jurišić, V. Investigation of NK cell function against two target hematological cell line using radioactive chromium assay. Appl Radiat Isot 2024. 206, 111251. [CrossRef]
- Burke, S.; et al. New views on natural killer cell-based immunotherapy for melanoma treatment. Trends Immunol 2010, 31, 339–45. [Google Scholar] [CrossRef] [PubMed]
- Jurisic, V.; et al. Clinical stage-depending decrease of NK cell activity in multiple myeloma patients. Med Oncol 2007, 24, 312–7. [Google Scholar] [CrossRef] [PubMed]
- Larsen, S.K.; Y. Gao.; Basse, P.H. NK cells in the tumor microenvironment. Crit Rev Oncog 2014. 19(1-2): p. 91-105.
- Imai, K.; et al. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general population. Lancet 2000, 356, 1795–9. [Google Scholar] [CrossRef]
- Visvader, J.E.; Lindeman, G.J. Cancer stem cells: current status and evolving complexities. Cell Stem Cell 2012, 10, 717–28. [Google Scholar] [CrossRef]
- Yu, C.C.; et al. miR145 targets the SOX9/ADAM17 axis to inhibit tumor-initiating cells and IL-6-mediated paracrine effects in head and neck cancer. Cancer Res 2013, 73, 3425–40. [Google Scholar] [CrossRef]
- Chiou, S.H.; et al. Positive correlations of Oct-4 and Nanog in oral cancer stem-like cells and high-grade oral squamous cell carcinoma. Clin Cancer Res 2008, 14, 4085–95. [Google Scholar] [CrossRef] [PubMed]
- Grandis, J.R.; et al. Human leukocyte antigen class I allelic and haplotype loss in squamous cell carcinoma of the head and neck: clinical and immunogenetic consequences. Clin Cancer Res 2000, 6, 2794–802. [Google Scholar]
- Uppaluri, R.; Dunn, G.P.; Lewis, J.S., Jr. Focus on TILs: prognostic significance of tumor infiltrating lymphocytes in head and neck cancers. Cancer Immun 2008. 8, 16.
- Staveley-O’Carroll, K.; et al. Induction of antigen-specific T cell anergy: An early event in the course of tumor progression. Proc Natl Acad Sci U S A 1998, 95, 1178–83. [Google Scholar] [CrossRef]
- Türkseven, M.R.; Oygür, T. Evaluation of natural killer cell defense in oral squamous cell carcinoma. Oral Oncol 2010, 46, e34–7. [Google Scholar] [CrossRef]
- Accomando, W.P.; et al. Decreased NK cells in patients with head and neck cancer determined in archival DNA. Clin Cancer Res 2012, 18, 6147–54. [Google Scholar] [CrossRef] [PubMed]
- Mickel, R.A.; et al. Natural killer cell cytotoxicity in the peripheral blood, cervical lymph nodes, and tumor of head and neck cancer patients. Cancer Res 1988, 48, 5017–22. [Google Scholar]
- Kaur, K.; et al. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Front Immunol 2017. 8, 297. [CrossRef] [PubMed]
- Turkseven, M.R.; Oygur, T. Evaluation of natural killer cell defense in oral squamous cell carcinoma. Oral Oncol 2010, 46, e34–7. [Google Scholar] [CrossRef]
- Igarashi, T.; et al. Enhanced cytotoxicity of allogeneic NK cells with killer immunoglobulin-like receptor ligand incompatibility against melanoma and renal cell carcinoma cells. Blood 2004, 104, 170–7. [Google Scholar] [CrossRef]
- Alici, E.; et al. Autologous antitumor activity by NK cells expanded from myeloma patients using GMP-compliant components. Blood 2008, 111, 3155–62. [Google Scholar] [CrossRef]
- Fujisaki, H.; et al. Expansion of highly cytotoxic human natural killer cells for cancer cell therapy. Cancer Res 2009, 69, 4010–7. [Google Scholar] [CrossRef]
- Berg, M.; et al. Clinical-grade ex vivo-expanded human natural killer cells up-regulate activating receptors and death receptor ligands and have enhanced cytolytic activity against tumor cells. Cytotherapy 2009, 11, 341–55. [Google Scholar] [CrossRef]
- Shah, N.; et al. Antigen presenting cell-mediated expansion of human umbilical cord blood yields log-scale expansion of natural killer cells with anti-myeloma activity. PLoS One 2013, 8, e76781. [Google Scholar] [CrossRef]
- Kaur, K.; et al. Osteoclast-expanded super-charged NK-cells preferentially select and expand CD8+ T cells. Scientific Reports 2020, 10, 20363. [Google Scholar] [CrossRef]
- Kaur, K.; et al. Probiotic-Treated Super-Charged NK Cells Efficiently Clear Poorly Differentiated Pancreatic Tumors in Hu-BLT Mice. Cancers 2020, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; et al. Sequential therapy with supercharged NK cells with either chemotherapy drug cisplatin or anti-PD-1 antibody decreases the tumor size and significantly enhances the NK function in Hu-BLT mice. Front Immunol 2023. 14, 1132807. [CrossRef] [PubMed]
- Mackowiak, P.A. ; Recycling metchnikoff: probiotics, the intestinal microbiome and the quest for long life. Front Public Health 2013. 1, 52. [CrossRef]
- Mowat, A.M. ; Historical Perspective: Metchnikoff and the intestinal microbiome. J Leukoc Biol 2021, 109, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; et al. The dual role of gut microbiota in pancreatic cancer: new insights into onset and treatment. Ther Adv Med Oncol 2025. 17, 17588359251324882. [CrossRef]
- Han, Z.-Y.; et al. Probiotics functionalized with a gallium-polyphenol network modulate the intratumor microbiota and promote anti-tumor immune responses in pancreatic cancer. Nature Communications 2024, 15, 7096. [Google Scholar] [CrossRef]
- Li, P.; Zhang, H.; Dai, M. Current status and prospect of gut and oral microbiome in pancreatic cancer: Clinical and translational perspectives. Cancer Letters 2024. 604, 217274. [CrossRef]
- Kaur, K.; et al. Natural Killer Cell Therapy Combined with Probiotic Bacteria Supplementation Restores Bone Integrity in Cancer by Promoting IFN-γ Production. Cells 2025, 14, 1347. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, P.; Weiskirchen, S.; Weiskirchen, R. Effects of Probiotics on Gut Microbiota: An Overview. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat Rev Immunol 2016, 16, 341–52. [Google Scholar] [CrossRef]
- Kerry, R.G.; et al. Benefaction of probiotics for human health: A review. J Food Drug Anal 2018, 26, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Bui, V.T.; et al. Augmented IFN-γ and TNF-α Induced by Probiotic Bacteria in NK Cells Mediate Differentiation of Stem-Like Tumors Leading to Inhibition of Tumor Growth and Reduction in Inflammatory Cytokine Release; Regulation by IL-10. Frontiers in Immunology 2015, 6, 2015. [Google Scholar] [CrossRef] [PubMed]
- Latvala, S.; et al. Lactobacillus rhamnosus GG and Streptococcus thermophilus induce suppressor of cytokine signalling 3 (SOCS3) gene expression directly and indirectly via interleukin-10 in human primary macrophages. Clinical and Experimental Immunology 2011, 165, 94–103. [Google Scholar] [CrossRef]
- Dargahi, N.; Johnson, J.C.; Apostolopoulos, V. Immune Modulatory Effects of Probiotic Streptococcus thermophilus on Human Monocytes. Biologics 2021, 1, 396–415. [Google Scholar] [CrossRef]
- Mandelbaum, N.; et al. Extracellular vesicles of the Gram-positive gut symbiont Bifidobacterium longum induce immune-modulatory, anti-inflammatory effects. NPJ Biofilms Microbiomes 2023, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Pei, B.; et al. Bifidobacterium modulation of tumor immunotherapy and its mechanism. Cancer Immunology, Immunotherapy 2024, 73, 94.
- Li, Z.; et al. Critical role of the gut microbiota in immune responses and cancer immunotherapy. Journal of Hematology & Oncology 2024, 17, 33.
- Gavzy, S.J.; et al. Bifidobacterium mechanisms of immune modulation and tolerance. Gut Microbes 2023, 15, 2291164. [Google Scholar] [CrossRef] [PubMed]
- Fanning, S.; et al. Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc Natl Acad Sci U S A 2012, 109, 2108–13. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Mercado, A.I.; et al. Bifidobacterium longum subsp. infantis CECT 7210 Reduces Inflammatory Cytokine Secretion in Caco-2 Cells Cultured in the Presence of Escherichia coli CECT 515. International Journal of Molecular Sciences 2022, 23, 10813.
- You, J.; Yaqoob, P. Evidence of immunomodulatory effects of a novel probiotic, Bifidobacterium longum bv. infantis CCUG 52486. FEMS Immunology & Medical Microbiology 2012, 66, 353-362.
- Kim, D.S.; et al. Lactobacillus acidophilus Supplementation Exerts a Synergistic Effect on Tacrolimus Efficacy by Modulating Th17/Treg Balance in Lupus-Prone Mice via the SIGNR3 Pathway. Frontiers in Immunology 2021, 12, 2021. [Google Scholar] [CrossRef]
- Vissers, Y.M.; et al. Lactobacillus strains differentially modulate cytokine production by hPBMC from pollen-allergic patients. FEMS Immunology & Medical Microbiology 2011, 61, 28-40.
- Yue, Y.; et al. Effects of Lactobacillus acidophilus KLDS1.0901 on Proliferation and Apoptosis of Colon Cancer Cells. Frontiers in Microbiology 2022. Volume 12 - 2021.
- Arasu, K.A.; Rajasekar, T. Immunomodulatory Activity of Postbiotics from Lactobacillus, in Postbiotics, D. Dharumadurai, Editor. 2024, Springer US: New York, NY. p. 181-186.
- Takagi, A.; et al. Relationship between the in vitro response of dendritic cells to Lactobacillus and prevention of tumorigenesis in the mouse. Journal of Gastroenterology 2008, 43, 661–669. [Google Scholar] [CrossRef]
- Zhu, Z.; et al. Lactobacillus casei combined with Lactobacillus reuteri alleviate pancreatic cancer by inhibiting TLR4 to promote macrophage M1 polarization and regulate gut microbial homeostasis. BMC Cancer 2023, 23, 1044. [Google Scholar] [CrossRef]
- Hao, R.; et al. Anti-inflammatory effect of <i>Lactiplantibacillus plantarum</i> T1 cell-free supernatants through suppression of oxidative stress and NF-κB- and MAPK-signaling pathways. Applied and Environmental Microbiology 2023, 89, e00608–23. [Google Scholar]
- Chiu, Y.-H.; et al. Lactobacillus plantarum MYL26 induces endotoxin tolerance phenotype in Caco-2 cells. BMC Microbiology 2013, 13, 190. [Google Scholar] [CrossRef]
- Zhang, R.; et al. Stevia Leaf Extract Fermented with Plant-Derived Lactobacillus plantarum SN13T Displays Anticancer Activity to Pancreatic Cancer PANC-1 Cell Line. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef]
- Zhang, S.L.; et al. Lacticaseibacillus paracasei sh2020 induced antitumor immunity and synergized with anti-programmed cell death 1 to reduce tumor burden in mice. Gut Microbes 2022, 14, 2046246. [Google Scholar] [CrossRef]
- Qin, D.; et al. Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei. Life (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Shah, A.B.; et al. Probiotic significance of Lactobacillus strains: a comprehensive review on health impacts, research gaps, and future prospects. Gut Microbes 2024, 16, 2431643. [Google Scholar] [CrossRef]
- Si, W.; et al. Lactobacillus rhamnosus GG induces STING-dependent IL-10 in intestinal monocytes and alleviates inflammatory colitis in mice. J Clin Invest 2025, 135. [Google Scholar] [CrossRef]
- Fong, F.L.Y.; et al. Immunomodulatory effects of Lactobacillus rhamnosus GG on dendritic cells, macrophages and monocytes from healthy donors. Journal of Functional Foods 2015. 13, 71–79. [CrossRef]
- Wang, L.; Liang, H.L.; Weichselbaum, R. Lactobacillus rhamnosus GG re-shapes gut microbiota and triggers STING-type I IFN-dependent antitumor immunity. The Journal of Immunology, 2022; 208, Supplement_1, 120.04. [Google Scholar]
- Ludwig, I.S.; et al. Lactobacillus rhamnosus GG-Derived Soluble Mediators Modulate Adaptive Immune Cells. Frontiers in Immunology 2018, 9, 2018. [Google Scholar] [CrossRef] [PubMed]
- Daillère, R.; et al. Enterococcus hirae and Barnesiella intestinihominis Facilitate Cyclophosphamide-Induced Therapeutic Immunomodulatory Effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Goubet, A.-G.; et al. Multifaceted modes of action of the anticancer probiotic Enterococcus hirae. Cell Death & Differentiation 2021, 28, 2276-2295.
- Hua, M.C.; et al. Probiotic Bio-Three induces Th1 and anti-inflammatory effects in PBMC and dendritic cells. World J Gastroenterol 2010, 16, 3529–40. [Google Scholar] [CrossRef]
- Alexovič, M.; et al. Human peripheral blood mononuclear cells: A review of recent proteomic applications. Proteomics 2022. 22(15-16): p. e2200026.
- Su, L.; et al. HMGN2, a new anti-tumor effector molecule of CD8+ T cells. Mol Cancer 2014. 13, 178. [CrossRef] [PubMed]
- Flint, H.J.; et al. Links between diet, gut microbiota composition and gut metabolism. Proc Nutr Soc 2015, 74, 13–22. [Google Scholar] [CrossRef]
- Al-Qadami, G.H.; et al. Gut Microbiota-Derived Short-Chain Fatty Acids: Impact on Cancer Treatment Response and Toxicities. Microorganisms 2022, 10, 2048. [Google Scholar] [CrossRef]
- Rangan, P.; Mondino, A. Microbial short-chain fatty acids: a strategy to tune adoptive T cell therapy. Journal for ImmunoTherapy of Cancer 2022, 10, e004147. [Google Scholar] [CrossRef]
- Thoda, C.; Touraki, M. Immunomodulatory Properties of Probiotics and Their Derived Bioactive Compounds. Applied Sciences 2023, 13, 4726. [Google Scholar] [CrossRef]
- Ahmad, M.F.; et al. Probiotics and Cancer: Mechanistic Insights and Organ-Specific Impact. Biomolecules 2025, 15, 879. [Google Scholar] [CrossRef]
- Shen, J.; et al. Anti-cancer therapy with TNFα and IFNγ: A comprehensive review. Cell Prolif 2018, 51, e12441. [Google Scholar] [CrossRef]
- Fink, L.N.; et al. Distinct gut-derived lactic acid bacteria elicit divergent dendritic cell-mediated NK cell responses. Int Immunol 2007, 19, 1319–27. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–73. [Google Scholar] [CrossRef] [PubMed]
- Mohamadzadeh, M.; et al. Lactobacilli activate human dendritic cells that skew T cells toward T helper 1 polarization. Proc Natl Acad Sci U S A 2005, 102, 2880–5. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; et al. The Multifaceted Role of Th1, Th9, and Th17 Cells in Immune Checkpoint Inhibition Therapy. Frontiers in Immunology 2021. Volume 12 - 2021.
- Mao, J.; et al. Probiotics Can Boost the Antitumor Immunity of CD8(+)T Cells in BALB/c Mice and Patients with Colorectal Carcinoma. J Immunol Res 2020. 2020, 4092472. [Google Scholar] [CrossRef]
- Fan, S.; et al. Microbiota-Derived L-SeMet Potentiates CD8+ T Cell Effector Functions and Facilitates Anti-Tumor Responses. International Journal of Molecular Sciences 2025, 26, 2511. [Google Scholar] [CrossRef]
- Römer, P.S.; et al. Preculture of PBMCs at high cell density increases sensitivity of T-cell responses, revealing cytokine release by CD28 superagonist TGN1412. Blood 2011, 118, 6772–6782. [Google Scholar] [CrossRef]
- Coënon, L.; et al. Natural Killer cells at the frontline in the fight against cancer. Cell Death & Disease 2024, 15, 614.
- Luo, N.; et al. Natural Killer Cell-Mediated Antitumor Immunity: Molecular Mechanisms and Clinical Applications. MedComm (2020) 2025, 6, e70387. [Google Scholar] [CrossRef]
- Morcillo-Martín-Romo, P.; et al. The Role of NK Cells in Cancer Immunotherapy: Mechanisms, Evasion Strategies, and Therapeutic Advances. Biomedicines 2025, 13. [Google Scholar] [CrossRef]
- Tao, J.-H.; et al. Nature killer cell for solid tumors: Current obstacles and prospective remedies in NK cell therapy and beyond. Critical Reviews in Oncology/Hematology 2025. 205, 104553.
- Fincham, R.E.A.; et al. Natural killer cells in pancreatic cancer stroma. World J Gastroenterol 2021, 27, 3483–3501. [Google Scholar] [CrossRef]
- Kumar, V.; Mahato, R.I. Natural killer cells for pancreatic cancer immunotherapy: Role of nanoparticles. Cancer Letters 2023. 579, 216462. [CrossRef]
- Fanijavadi, S.; Thomassen, M.; Jensen, L.H. Targeting Triple NK Cell Suppression Mechanisms: A Comprehensive Review of Biomarkers in Pancreatic Cancer Therapy. International Journal of Molecular Sciences 2025, 26, 515. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; et al. Defective NK cell expansion, cytotoxicity, and lack of ability to differentiate tumors from a pancreatic cancer patient in a long term follow-up: implication in the progression of cancer. Cancer Immunology, Immunotherapy 2022, 71, 1033-1047.
- Berrien-Elliott, M.M.; Jacobs, M.T.; Fehniger, T.A. Allogeneic natural killer cell therapy. Blood 2023, 141, 856–868. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; et al. Technical advances in NK cell-based cellular immunotherapy. Cancer Biol Med 2019, 16, 647–654. [Google Scholar] [CrossRef]
- Liu, S.; et al. NK cell-based cancer immunotherapy: from basic biology to clinical development. J Hematol Oncol 2021, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Maia, A.; et al. Building a Better Defense: Expanding and Improving Natural Killer Cells for Adoptive Cell Therapy. Cells 2024, 13. [Google Scholar] [CrossRef]
- Perussia, B.; et al. Preferential proliferation of natural killer cells among peripheral blood mononuclear cells cocultured with B lymphoblastoid cell lines. Nat Immun Cell Growth Regul 1987, 6, 171–88. [Google Scholar]
- Rabinowich, H.; et al. Increased proliferation, lytic activity, and purity of human natural killer cells cocultured with mitogen-activated feeder cells. Cell Immunol 1991, 135, 454–70. [Google Scholar] [CrossRef]
- Srivastava, S.; Lundqvist, A.; Childs, R.W. Natural killer cell immunotherapy for cancer: a new hope. Cytotherapy 2008, 10, 775–83. [Google Scholar] [CrossRef] [PubMed]
- Gras Navarro, A.; Björklund, A.; Chekenya, M. Therapeutic potential and challenges of Natural killer cells in treatment of solid tumors. Frontiers in Immunology 2015. 6.
- Alici, E.; et al. Autologous antitumor activity by NK cells expanded from myeloma patients using GMP-compliant components. Blood 2008, 111, 3155–62. [Google Scholar] [CrossRef] [PubMed]
- Garg, T.K.; et al. Highly activated and expanded natural killer cells for multiple myeloma immunotherapy. Haematologica 2012, 97, 1348–56. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; et al. Novel Strategy to Expand Super-Charged NK Cells with Significant Potential to Lyse and Differentiate Cancer Stem Cells: Differences in NK Expansion and Function between Healthy and Cancer Patients. Frontiers in Immunology 2017, 8, 2017. [Google Scholar] [CrossRef]
- Gurney, M.; et al. Feeder Cells at the Interface of Natural Killer Cell Activation, Expansion and Gene Editing. Front Immunol 2022. 13, 802906. [CrossRef]
- Michen, S.; et al. Artificial feeder cells expressing ligands for killer cell immunoglobulin-like receptors and CD94/NKG2A for expansion of functional primary natural killer cells with tolerance to self. Cytotherapy 2020, 22, 354–368. [Google Scholar] [CrossRef]
- Tran, T.B.T.; et al. In Vitro Expansion and Transduction of Primary NK Cells Using Feeder Cells Expressing Costimulatory Molecules and IL-21. Cancer Science 2025, 116, 1847–1860. [Google Scholar] [CrossRef]
- Marr, B.; et al. Cytokines in Focus: IL-2 and IL-15 in NK Adoptive Cell Cancer Immunotherapy. Immune Netw 2025, 25, e17. [Google Scholar] [CrossRef]
- Denman, C.J.; et al. Membrane-bound IL-21 promotes sustained ex vivo proliferation of human natural killer cells. PLoS One 2012, 7, e30264. [Google Scholar] [CrossRef]
- Gao, F.; et al. Influence of KIR and NK Cell Reconstitution in the Outcomes of Hematopoietic Stem Cell Transplantation. Front Immunol 2020. 11, 2022. [CrossRef] [PubMed]
- Mehta, R.S.; Rezvani, K. Can we make a better match or mismatch with KIR genotyping? Hematology Am Soc Hematol Educ Program 2016, 2016, 2016, 106–118. [Google Scholar] [CrossRef]
- Ko, M.W.; et al. Osteoclast-expanded supercharged NK cells perform superior antitumour effector functions. BMJ Oncol 2025, 4, e000676. [Google Scholar] [CrossRef]
- Kaur, K.; et al. ADCC against MICA/B Is Mediated against Differentiated Oral and Pancreatic and Not Stem-Like/Poorly Differentiated Tumors by the NK Cells; Loss in Cancer Patients due to Down-Modulation of CD16 Receptor. Cancers 2021, 13, 239. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Chen, P.-C.; Jewett, A. Supercharged NK cells: a unique population of NK cells capable of differentiating stem cells and lysis of MHC class I high differentiated tumors. Cell Death & Disease 2025, 16, 665.
- Huerta-Yepez, S.; et al. Supercharged NK cells, unlike primary activated NK cells, effectively target ovarian cancer cells irrespective of MHC-class I expression. BMJ Oncol 2025, 4, e000618. [Google Scholar] [CrossRef]
- Liu, E.; et al. Use of CAR-Transduced Natural Killer Cells in CD19-Positive Lymphoid Tumors. N Engl J Med 2020, 382, 545–553. [Google Scholar] [CrossRef]
- Becker, P.S.; et al. Selection and expansion of natural killer cells for NK cell-based immunotherapy. Cancer Immunol Immunother 2016, 65, 477–84. [Google Scholar] [CrossRef]
- Kaur, K.; et al. Sequential therapy with supercharged NK cells with either chemotherapy drug cisplatin or anti-PD-1 antibody decreases the tumor size and significantly enhances the NK function in Hu-BLT mice. Frontiers in Immunology 2023, 14, 2023. [Google Scholar] [CrossRef]
- Leivas, A.; et al. Novel treatment strategy with autologous activated and expanded natural killer cells plus anti-myeloma drugs for multiple myeloma. Oncoimmunology 2016, 5, e1250051. [Google Scholar] [CrossRef] [PubMed]
- Sexton, R.E.; et al. Connecting the Human Microbiome and Pancreatic Cancer. Cancer Metastasis Rev 2022, 41, 317–331. [Google Scholar] [CrossRef]
- Mohamed Elfadil, O.; et al. Butyrate: More Than a Short Chain Fatty Acid. Curr Nutr Rep 2023, 12, 255–262. [Google Scholar] [CrossRef]
- Qin, W.; et al. The role of probiotic foods in acute pancreatitis: current status and future directions. Current Opinion in Food Science 2024. 60, 101231. [CrossRef]
- Werawatganon, D.; et al. Effects of probiotics on pancreatic inflammation and intestinal integrity in mice with acute pancreatitis. BMC Complement Med Ther 2023, 23, 166. [Google Scholar] [CrossRef]
- Chen, S.M.; et al. The synergistic tumor growth-inhibitory effect of probiotic Lactobacillus on transgenic mouse model of pancreatic cancer treated with gemcitabine. Sci Rep 2020, 10, 20319. [Google Scholar] [CrossRef] [PubMed]
- Kita, A.; et al. Probiotic-derived ferrichrome inhibits the growth of refractory pancreatic cancer cells. Int J Oncol 2020, 57, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Farrow, B.; et al. Butyrate inhibits pancreatic cancer invasion. J Gastrointest Surg 2003, 7, 864–70. [Google Scholar] [CrossRef]
- Davie, J.R. ; Inhibition of histone deacetylase activity by butyrate. J Nutr 2003, 133(7 Suppl), 2485s–2493s. [Google Scholar] [CrossRef]
- Sun, J.; et al. Butyrate as a promising therapeutic target in cancer: From pathogenesis to clinic (Review). Int J Oncol 2024, 64. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; et al. Lactobacillus Attenuate the Progression of Pancreatic Cancer Promoted by Porphyromonas Gingivalis in K-ras(G12D) Transgenic Mice. Cancers (Basel) 2020, 12. [Google Scholar]
- Lee, T.S. ; Are Probiotics Beneficial or Harmful for Pancreatic Cancer Outcomes? Probiotics and Antimicrobial Proteins 2025, 17, 2293–2300. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.-W.; et al. Defective Patient NK Function Is Reversed by AJ2 Probiotic Bacteria or Addition of Allogeneic Healthy Monocytes. Cells 2022, 11, 697. [Google Scholar] [CrossRef]
- Yang, Z.; et al. The effect of probiotics supplementation on cancer-treatment complications: a critical umbrella review of interventional meta-analyses. Crit Rev Food Sci Nutr 2025, 65, 3702–3727. [Google Scholar] [CrossRef] [PubMed]
- Dikeocha, I.J.; et al. Probiotics supplementation in patients with colorectal cancer: a systematic review of randomized controlled trials. Nutrition Reviews 2021, 80, 22–49. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.L.; et al. Effect of Probiotics on Metabolic Outcomes in Pregnant Women with Gestational Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Zarezadeh, M.; et al. Probiotic therapy, a novel and efficient adjuvant approach to improve glycemic status: An umbrella meta-analysis. Pharmacological Research 2022. 183, 106397. [CrossRef]
- Musazadeh, V.; et al. Probiotics as an effective therapeutic approach in alleviating depression symptoms: an umbrella meta-analysis. Critical Reviews in Food Science and Nutrition 2023, 63, 8292–8300. [Google Scholar] [CrossRef]
- Chen, S.-M.; et al. The synergistic tumor growth-inhibitory effect of probiotic Lactobacillus on transgenic mouse model of pancreatic cancer treated with gemcitabine. Scientific Reports 2020, 10, 20319. [Google Scholar] [CrossRef]
- Moreira, M.M.; et al. The impact of pre-, pro- and synbiotics supplementation in colorectal cancer treatment: a systematic review. Front Oncol 2024. 14, 1395966.
- Maher, S.; et al. Synergistic immunomodulatory effect of synbiotics pre- and postoperative resection of pancreatic ductal adenocarcinoma: a randomized controlled study. Cancer Immunology, Immunotherapy 2024, 73, 109.
Figure 1.
Probiotic treatments enhance cytotoxic activity, increase anti-inflammatory cytokines, decrease pro-inflammatory cytokines in PBMCs and PBMCs-derived immune cells, ultimately leading to lysis and growth inhibition of pancreatic cancer. They boost the anti-cancer effects of PBMCs, as well as PBMC-derived NK and T cells, either directly or indirectly by modulating dendritic cells or other antigen-presenting cells. The heightened cytotoxic activity of PBMCs, NK, and T cells supports tumor apoptosis, while increased cytokine secretion aids in tumor differentiation. Upon differentiation, tumors proliferate and metastasize at a minimal rate. Antigen-presenting cells drive T-cell polarization and enhance T-cell cytotoxicity, working in synergy with radiotherapy, chemotherapy, and immune checkpoint inhibitors. These processes highlight the potential of probiotics to encourage apoptosis, cell cycle arrest, and tumor growth inhibition. Illustration created with https://BioRender.com on September 24, 2025. https://app.biorender.com/illustrations/68cdb701d629a499caac78aa?slideId=9ad3a503-198a-4fc5-8316-c2e0f35b40ec.
Figure 1.
Probiotic treatments enhance cytotoxic activity, increase anti-inflammatory cytokines, decrease pro-inflammatory cytokines in PBMCs and PBMCs-derived immune cells, ultimately leading to lysis and growth inhibition of pancreatic cancer. They boost the anti-cancer effects of PBMCs, as well as PBMC-derived NK and T cells, either directly or indirectly by modulating dendritic cells or other antigen-presenting cells. The heightened cytotoxic activity of PBMCs, NK, and T cells supports tumor apoptosis, while increased cytokine secretion aids in tumor differentiation. Upon differentiation, tumors proliferate and metastasize at a minimal rate. Antigen-presenting cells drive T-cell polarization and enhance T-cell cytotoxicity, working in synergy with radiotherapy, chemotherapy, and immune checkpoint inhibitors. These processes highlight the potential of probiotics to encourage apoptosis, cell cycle arrest, and tumor growth inhibition. Illustration created with https://BioRender.com on September 24, 2025. https://app.biorender.com/illustrations/68cdb701d629a499caac78aa?slideId=9ad3a503-198a-4fc5-8316-c2e0f35b40ec.
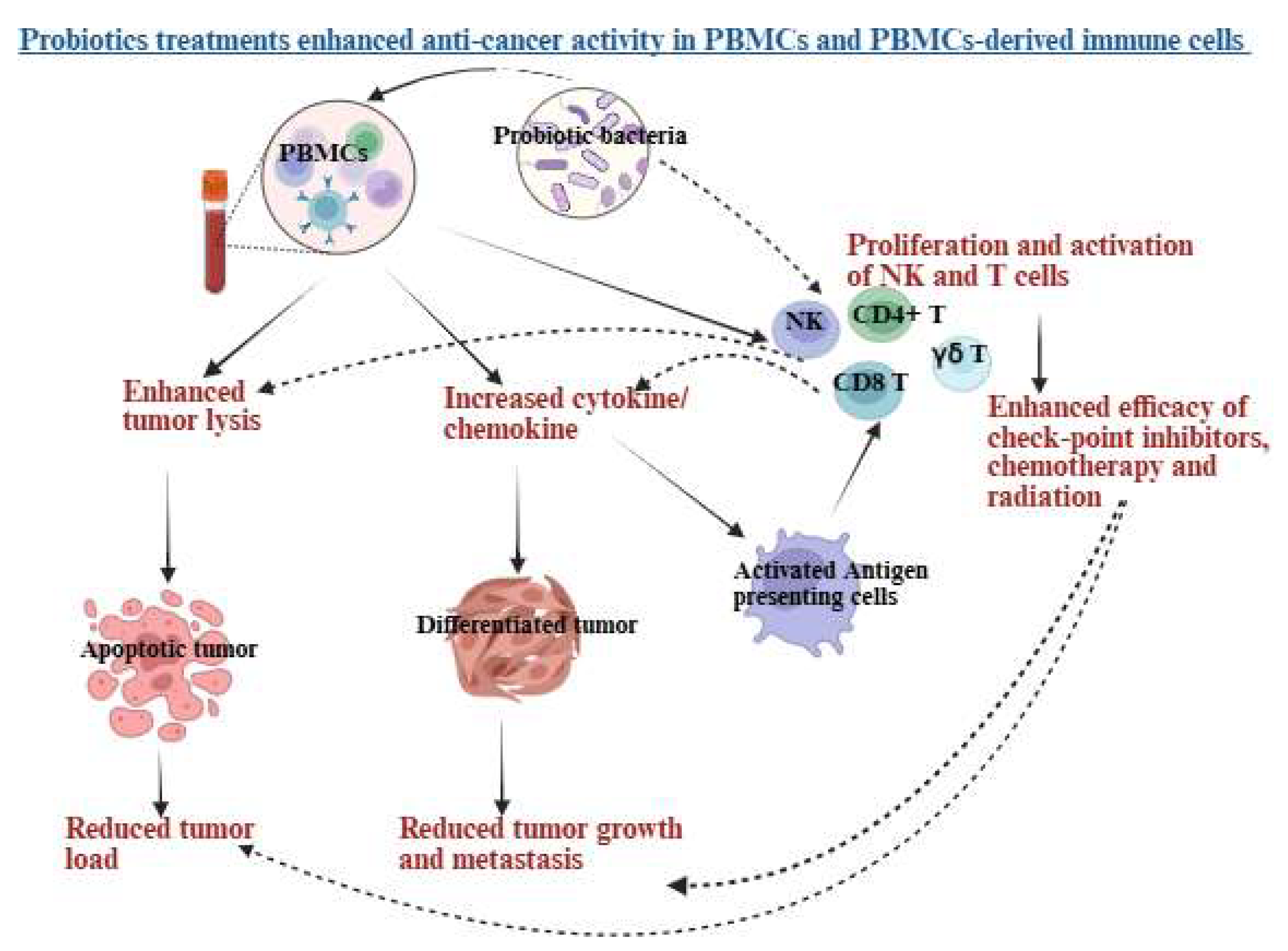
Figure 2.
A schematic overview of the generation and characteristics of expanded NK cells using probiotics and feeder cells. Human peripheral blood-derived cells are treated with cytokines and antibodies and co-cultured with feeder cells alongside probiotic bacteria. NK cells expanded with probiotics and feeder cells showed enhanced expansion and functional activation compared to those expanded with feeder cells alone. Illustration created with https://BioRender.com on September 26, 2025. https://app.biorender.com/illustrations/68cdea1c0412ec45e81a342d?slideId=9ad3a503-198a-4fc5-8316-c2e0f35b40ec.
Figure 2.
A schematic overview of the generation and characteristics of expanded NK cells using probiotics and feeder cells. Human peripheral blood-derived cells are treated with cytokines and antibodies and co-cultured with feeder cells alongside probiotic bacteria. NK cells expanded with probiotics and feeder cells showed enhanced expansion and functional activation compared to those expanded with feeder cells alone. Illustration created with https://BioRender.com on September 26, 2025. https://app.biorender.com/illustrations/68cdea1c0412ec45e81a342d?slideId=9ad3a503-198a-4fc5-8316-c2e0f35b40ec.
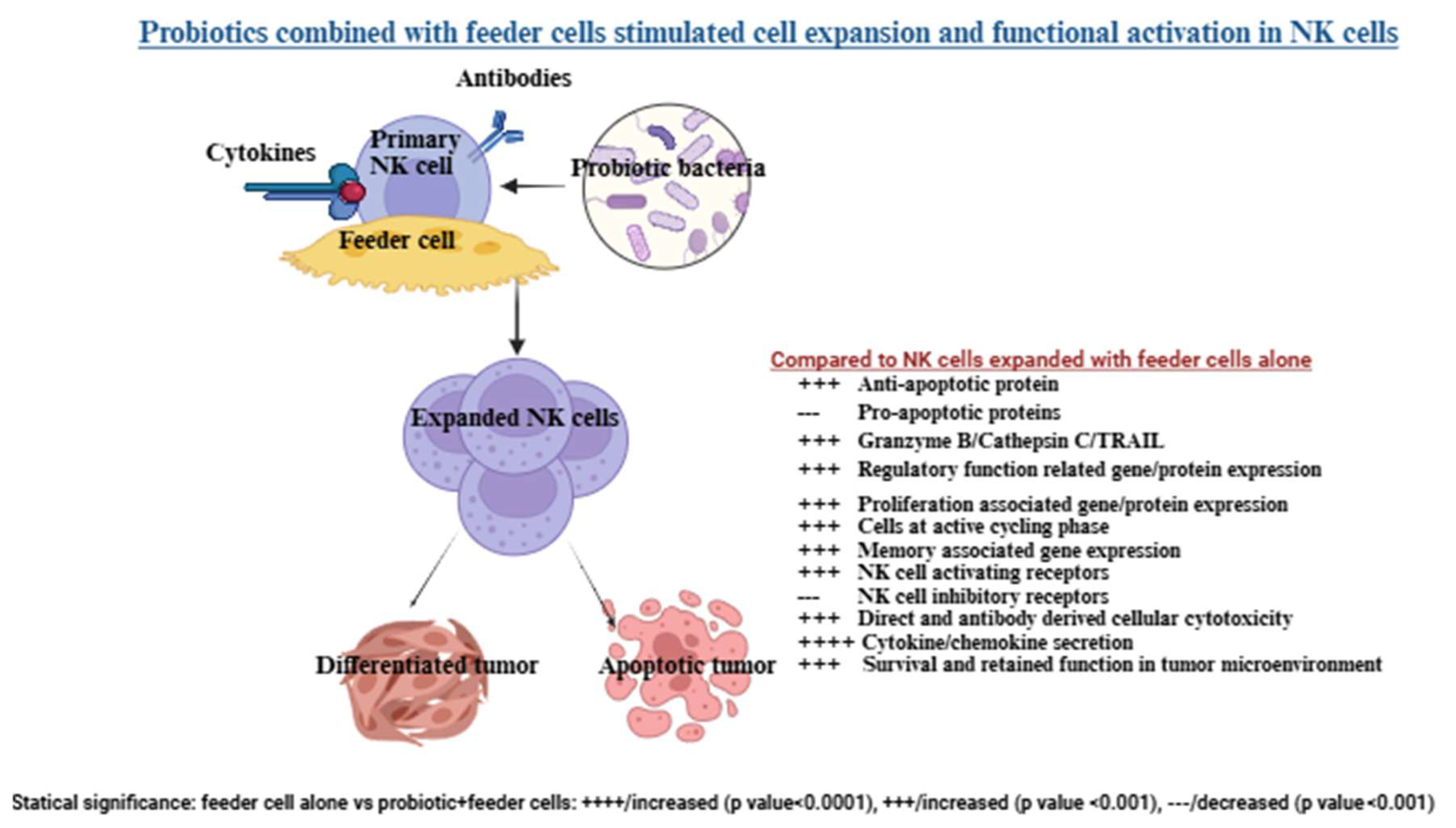

Table 1.
Probiotic strains that may help inhibit pancreatic cancer and improve its prognosis.
| Probiotic strains | Anti-cancer effects | References |
|---|---|---|
| Streptococcus thermophilus | - Promote Th1-type cytokine profile, increasing IL-12 and IFN-γ in PBMCs, NK and T cells - Boost cytotoxic activity in PBMCs, NK and T cells |
[70,71,72] |
| Bifidobacterium longum | - Regulate cytokine secretion in PBMCs and NK cells - Boost cytotoxic activity in PBMCs, NK and T cells - Increase the number of total T cells, NK cells, and increase the CD8+/CD4+ T ratio - Enhance CD8+T cells priming and accumulation in TME in mice - Increase anti-cancer gene expressions on dendritic cells - Increase efficacy of PD-1 therapy in pancreatic cancer - Protection against chemo- and radiotherapy-induced fever and diarrhea in pancreatic cancer patients -Inhibit pancreatic tumor proliferation |
[70,73,74,75] |
|
Bifidobacterium breve |
- Boosts IL-10 production and cytotoxic activity in PBMCs - Encourage CD4+ and CD8+ T cell proliferation - Boost IFN-γ production in PBMCs, NK and T cells |
[70,76,77] |
| Bifidobacterium infantis | -Regulated cytokine secretion in human PBMCs and NK cells - Support Th2 profile with higher IL-10 and IL-6 compared to IL-12 and IFN-γ |
[70,78,79] |
| Lactobacillus acidophilus | - Regulate cytokine secretion in human PBMCs and NK cells - Induce proliferation of CD4+ and CD8+ T cells -Protection against chemo- and radiotherapy-induced fever and diarrhea in pancreatic cancer patients -Sensitized tumor cells to chemotherapy by faster activation of caspase-3 and downregulation of p21 protein -Inactivated the NF-kB inflammatory pathway |
[70,80,81,82,83] |
| Lactobacillus Lactis | -Inhibit tumor proliferation -Reduce pro-inflammatory cytokines -Enhance Th17 immune response against cancer - Boost antigen presentation by dendritic cells, enhancing cytotoxic T cell responses |
[70,84] |
| Lactobacillus reutri | -Inhibit p53-p21-Cyclin B1/Cdk1 signaling pathway resulting in growth arrest at G2 growth phase of tumors -Inhibit pancreatic cancer proliferation, migration and invasion |
[85] |
| Lactobacillus plantarum | - Regulate cytokine secretion in PBMCs, NK and T cells -Increase tumor infiltration of CD4+ and CD8+ T cells -Promoted Th1-type CD4+ T cell differentiation - Reduces NF-κB and Wnt/β-catenin in tumors through PBMC signaling - Induce cytotoxicity in pancreatic cancer |
[70,86,87,88] |
| Lactobacillus paracasei/casei | - Regulate cytokine secretion in PBMCs and NK cells - Promote tumor-specific T cell infiltration and reduce pro-tumoral IL-6 levels, enhancing cancer-specific immunity - Stimulate Th1 and Th17 responses, which synergize with chemotherapy drugs like cyclophosphamide and gemcitabine -Inhibit tumor growth and proliferation -Promoted apoptotic cell death in tumors -Upregulated the epression if apoptosis-inducing ligand TRAIL and dowregulates transcription expressions of cyclin D1 and BIRC5a in cancer cells -Increase NK cell-mediated cytotoxicity -Increase the numbers of total T cells, NK cells, and increased CD8+/CD4+ T ratio -Sensitize tumor cells to chemotherapy by faster activation of caspase-3 and dowregulation of p21 protein -Activated c-jun N-terminal kinase (JNK) mediated apoptosis of tumors -Inhibit tumor growth by decreasing matrix metalloproteinase-9 (MMP-9) activity - Enhance efficacy of anti-PD1 |
[70,85,89,90,91] |
| Lactobacillus bulgaricus |
- Regulate cytokine secretion in PBMCs and NK cells | [70] |
| Lactobacillus rhamnosus | - Boost IL-10 production and cytotoxic activity in PBMCs - Encourage CD4+ and CD8+ T cell proliferation - Boost IFN-γ production in PBMCs, NK and T cells - Stimulate Th1 and Th17 responses, which synergize with chemotherapy drugs like cyclophosphamide and gemcitabine -Alleviate effects of pro-inflammatory cytokines on epithelial barrier and inflammation through inhibition of NF-kB signalling -Inhibits tumor growth by decreasing matrix metalloproteinase-9 (MMP-9) activity |
[92,93,94,95] |
| Enterococcus hirae | - Improve CD8+/Treg ratio - Stimulate Th1 and Th17 responses, which synergize with chemotherapy drugs like cyclophosphamide and gemcitabine |
[96,97] |
| Bacillus mesentericus | - Stimulate Th1 immune response, downregulate pro-inflammatory cytokines, and upregulate anti-inflammatory cytokines in PBMCs - Increase surface expression of CD11b, HLA-DR, CD4, CD45Ram CD25, CD44 and CD69 in PBMCs - Increase secretion of IL-10 and IL-12 in dendritic cells |
[98] |
| Clostridium butyricum | - Stimulate Th1 immune response, downregulate pro-inflammatory cytokines, and upregulate anti-inflammatory cytokines in PBMCs - Increase surface expression of CD11b, HLA-DR, CD4, CD45Ram CD25, CD44 and CD69 in PBMCs - Increase secretion of IL-10 and IL-12 in dendritic cells |
[98] |
| Enterococcus faecalis | - Stimulate Th1 immune response, downregulate pro-inflammatory cytokines, and upregulate anti-inflammatory cytokines in PBMCs - Increase surface expression of CD11b, HLA-DR, CD4, CD45Ram CD25, CD44 and CD69 in PBMCs - Increase secretion of IL-10 and IL-12 in dendritic cells |
[98] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



