
Submitted:
17 October 2025
Posted:
20 October 2025
You are already at the latest version
Abstract
Deuterium, a stable isotope of hydrogen present in natural water at ~150 ppm, has been implicated in modulating cellular metabolism and tumor progression. While deuterium-depleted water (DDW) has shown anticancer effects in preclinical and clinical studies, the underlying transcriptional mechanisms remain incompletely defined. Here, we profiled gene expression in A549 lung adenocarcinoma cells cultured for 72 hours in media containing four graded deuterium concentrations (40, 80, 150, and 300 ppm) using a targeted NanoString panel of 236 cancer-related genes. After stringent quality filtering, 87 genes were retained and classified into nine distinct expression patterns based on fold-change trends relative to the 150 ppm control. High deuterium (300 ppm) induced strong upregulation of oncogenic and survival-related genes (e.g., EGFR, CTNNB1, STAT3, CD44), while DDW (40–80 ppm) led to selective downregulation of oncogenes (e.g., MYCN, ETS2, IRF1) and drug-resistance genes (e.g., ABCB1). Several genes involved in DNA repair, apoptosis, and extracellular matrix remodeling exhibited dose-dependent responses, suggesting coordinated regulation by deuterium abundance. These findings demonstrate that deuterium concentration functions as a biologically active variable capable of modulating cancer-relevant gene networks. This exploratory dataset refines mechanistic models of DDW action and provides a foundation for future studies incorporating biological replication, functional assays, and in vivo validation. Significance: Deuterium concentration modulation alters oncogenic, apoptotic, and drug-resistance gene networks in lung adenocarcinoma cells, refining prior models of deuterium-depleted water effects. These findings identify deuterium concentration as a biologically active variable warranting further mechanistic and translational investigation.
Keywords:
deuterium concentration modulation
; deuterium depleted water
; in vitro
; lung adenocarcinoma
; A549 cells
; gene expression profiling
; NanoString technology
; oncogenic signaling
; apoptosis regulation
; drug resistance
1. Introduction
Cancer progression is driven by complex interactions between oncogenes, tumor suppressors, and environmental factors [1]. While the role of oncogenes and tumor suppressors is well established, the contribution of stable isotopes such as deuterium to tumor biology has received comparatively little attention. Naturally occurring at ~150 ppm in drinking water [2], deuterium is incorporated into biomolecules through metabolic processes, subtly altering reaction kinetics and hydrogen-bonding networks [3,4,5]. Deuterium depleted water (DDW) has been shown to inhibit proliferation in multiple cancer models and, in some clinical contexts, to prolong survival. Proposed mechanisms include modulation of mitochondrial function, redox balance, and signal transduction pathways [6]. By modulating fundamental molecular interactions, shifts in the intracellular and extracellular D/H ratio may contribute to altered gene expression programs, changes in cell-cycle progression, and the acquisition of drug-resistant phenotypes — hallmarks of malignant transformation and tumor progression. Given these mechanistic links, modulation of deuterium abundance represents a potential metabolic and signaling checkpoint in tumor biology. Previous studies have suggested that lowering the D/H ratio can influence mitochondrial function, oxidative stress responses, and apoptosis induction in malignant cells, while sparing normal tissues [7,8,9,10,11,12,13]. Two studies have investigated the effect of DDW on a limited set of cancer-related genes in carcinogen-induced mice. The findings showed that DDW inhibited the upregulation of Kras, H-ras, Bcl2, Myc, and p53 gene expression, further supporting the role of an elevated D/H ratio in regulating gene expression [14,15]. However, the transcriptomic landscape underlying these effects remains to be better defined.
To address this gap, we employed NanoString technology to profile [16] gene expression [17,18] in lung adenocarcinoma cells exposed to variable deuterium concentrations. To reveal the impact of D concentration on gene expression a more accurate and gene specific analysis was carried out compared to a former analysis [9]. This approach enabled the identification of coordinated transcriptional responses and the delineation of dose-dependent expression patterns. By integrating pathway analysis with functional categorization, we aimed to uncover molecular signatures that could lead to the therapeutic application of deuterium depletion in oncology.
2. Results
2.1. Gene Expression Classification Results
We selected 87 genes (vide infra in Section 4) and classified them based on their expression ratios relative to the 150 ppm control. The summary of expression ratios in each classification category (upregulated, stable and downregulated) for the different deuterium levels is presented in Table 1.
The results in Table 1 show that exposure to 300 ppm deuterium did not cause downregulation in any of the genes examined. Instead, 72 genes were upregulated under these conditions. This contrasts sharply with the results from the deuterium-depleted medium, which led to the downregulation of 7 to 19 genes and only a moderate 23% upregulation in two exposed to 40 ppm deuterium. None of the investigated genes were upregulated above 20% in 80 ppm deuterium content medium. The average downregulation is 0.91 and 0.95 in 40 and 80 ppm medium, respectively. We noticed that 8 of 66 stable expression ratio genes have an expression ratio below 0.85 thus these genes are close to falling into the moderately downregulated category in 40 ppm medium. The average upregulation is 1.47 in 300 ppm medium.
2.2. Pattern Analysis and Gene Functions
To gain deeper insights into the biological implications, the 87 genes were grouped into distinct expression patterns based on their classification across the 40 ppm, 80 ppm, and 300 ppm deuterium levels. The ratios of the average copy numbers vs the reference 150 ppm deuterium content media, the copy numbers, the averages, CV and Z values can be found in the Supplementary Data.
Pattern 1: Stable at 40 ppm and 80 ppm, Strongly Upregulated at 300 ppm (29 genes)
This pattern represents genes that maintain stable expression at lower deuterium concentrations (40 ppm and 80 ppm) but exhibit a significant increase in expression at high deuterium (300 ppm).
Genes in Pattern 1: BRAF, BRCA1, CD44, CTNNB1, DAPK1, DEK, EGFR, ERBB3, ERCC4, FAT1, FGFR1, IFNGR1, ITGB1, JUN, LAMB1, LIF, MSH6, MYC, PLAUR, PTGS2, RB1, SPP1, STAT3, TFRC, TGFA, TGFBI, TGFBR2, XRCC5, YES1.
Figure 1.
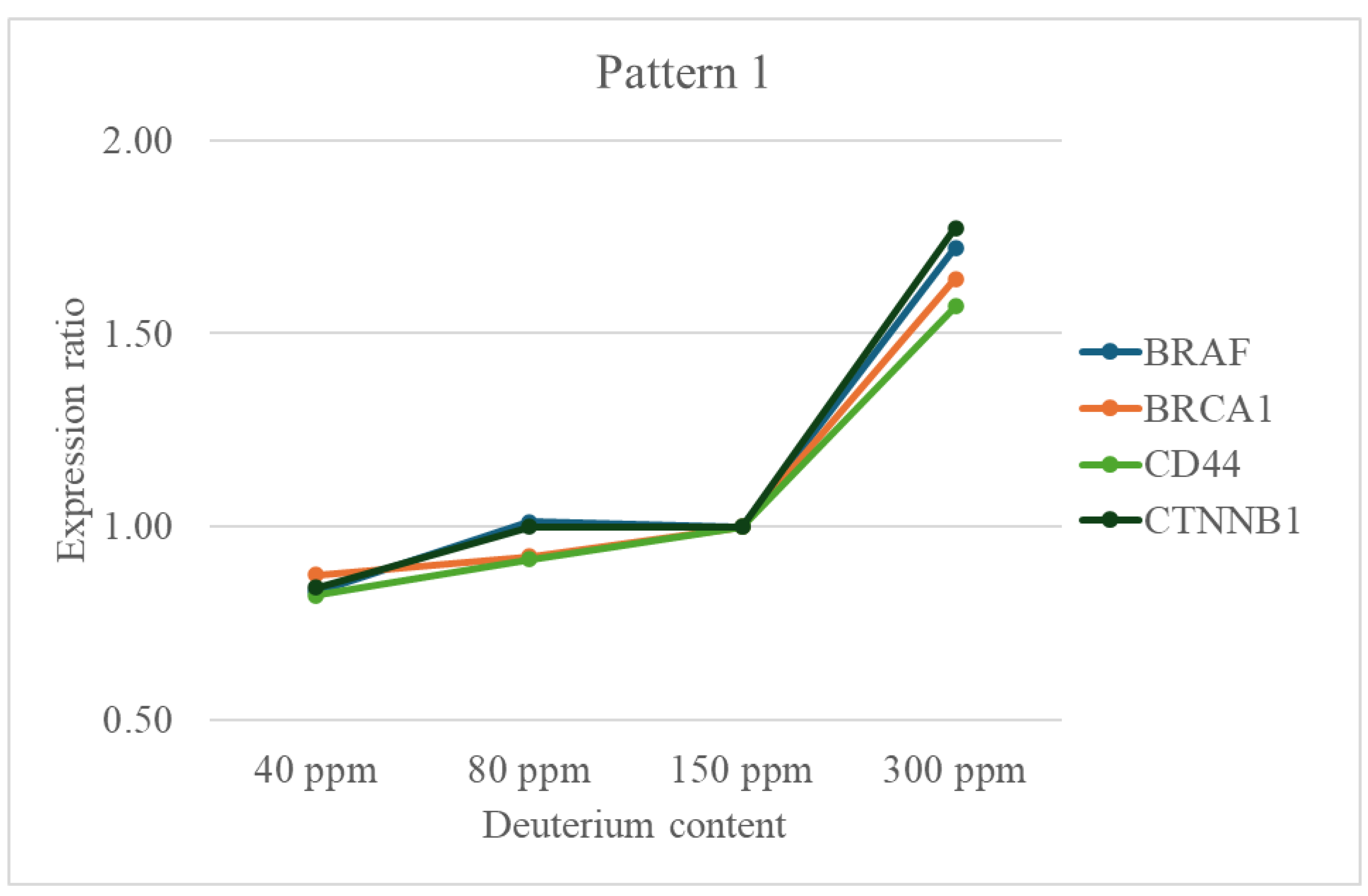
Representative Pattern 1 gene expression profiles in A549 lung adenocarcinoma cells exposed to varying deuterium concentrations. Line graphs show the relative mRNA expression (y-axis) of four representative Pattern 1 genes (BRAF. BRCA1, CD44 and CTNNB1) in cells cultured for 72 h in media containing 40 ppm, 80 ppm, 150 ppm (natural abundance control), or 300 ppm deuterium (x-axis). Expression values are presented as the ratio of the mean NanoString copy number at each deuterium level to the mean value in the 150 ppm control. Pattern 1 genes are defined by stable expression at 40 ppm and 80 ppm (ratios within 0.83–1.20) and strong upregulation (>1.5-fold) at 300 ppm. These genes are predominantly involved in oncogenic signaling (e.g., MAPK/ERK, PI3K/AKT, Wnt, JAK/STAT), cell adhesion, and extracellular matrix remodeling. The coordinated induction at high deuterium suggests activation of pro-proliferative and pro-survival pathways, potentially enhancing tumor aggressiveness. Horizontal axis spacing is not to scale; connecting lines are provided for visual guidance only.
Common Characteristics: These genes are predominantly involved in core oncogenic processes.
Representative Pattern 1 gene expression profiles in A549 lung adenocarcinoma cells exposed to varying deuterium concentrations. Line graphs show the relative mRNA expression (y-axis) of four representative Pattern 1 genes (BRAF. BRCA1, CD44 and CTNNB1) in cells cultured for 72 h in media containing 40 ppm, 80 ppm, 150 ppm (natural abundance control), or 300 ppm deuterium (x-axis). Expression values are presented as the ratio of the mean NanoString copy number at each deuterium level to the mean value in the 150 ppm control. Pattern 1 genes are defined by stable expression at 40 ppm and 80 ppm (ratios within 0.83–1.20) and strong upregulation (>1.5-fold) at 300 ppm. These genes are predominantly involved in oncogenic signaling (e.g., MAPK/ERK, PI3K/AKT, Wnt, JAK/STAT), cell adhesion, and extracellular matrix remodeling. The coordinated induction at high deuterium suggests activation of pro-proliferative and pro-survival pathways, potentially enhancing tumor aggressiveness. Horizontal axis spacing is not to scale; connecting lines are provided for visual guidance only.
Common Characteristics: These genes are predominantly involved in core oncogenic processes.
- Cell Signaling Pathways: Many are components of key growth-promoting pathways such as MAPK/ERK (BRAF, EGFR, JUN, YES1), PI3K/AKT (EGFR, ERBB3, LIF, MYC), Wnt (CTNNB1), and JAK/STAT (LIF, STAT3). Their upregulation at 300 ppm suggests an amplification of these pro-survival and pro-proliferative signals under high deuterium conditions.
- Proliferation and Survival: Oncogenes like MYC, EGFR, and ERBB3 directly promote cell cycle progression and inhibit apoptosis. SPP1 and TGFA further enhance proliferation and tumor growth.
- Stress Response and DNA Repair: Genes like BRCA1, ERCC4, MSH6, and XRCC5 are crucial for maintaining genomic stability through DNA repair mechanisms. Their upregulation might indicate a cellular response to deuterium-induced stress, where increased repair capacity is needed to cope with potential DNA damage. DAPK1, a pro-apoptotic gene, may also be activated in response to stress.
- Cell Adhesion and Migration: CD44, ITGB1, LAMB1, FAT1, PLAUR, and TGFBI are integral to cell-matrix interactions and extracellular matrix (ECM) remodeling, processes critical for tumor invasion and metastasis. Their strong upregulation at 300 ppm suggests an enhanced metastatic potential.
- Immune and Inflammatory Modulation: IFNGR1 and PTGS2 (COX-2) play roles in immune responses and inflammation, which can contribute to a tumor-supportive microenvironment.
Connections: The 300 ppm condition triggered coordinated upregulation across multiple genes, suggesting activation of shared regulatory networks, including PI3K/AKT, p53, and NF-κB. D-induced changes in H-bonding may alter chromatin accessibility, influencing transcription rates. This pattern aligns with literature suggesting that higher deuterium concentrations can enhance oncogenic pathways (9,12).
Figure 2 shows Pattern 1 for the two most upregulated genes, with DAPK1 showing the highest increase (113% in the 300 ppm medium), followed by TGFBR2 with a 105% increase. TGFBR2 functions as a receptor for external signals — most notably TGF-β — while DAPK1 serves as an intracellular effector involved in apoptosis and autophagy. Their potential intersection lies in TGF-β-induced cell death or cell cycle arrest: TGFBR2 initiates the signaling cascade, and DAPK1 may act downstream to execute the death signal, particularly under stress conditions such as elevated reactive oxygen species (ROS) levels (19,20,21). Notably, both genes were among the most strongly induced at 300 ppm deuterium concentration, suggesting that they may play a highly specific and coordinated role in the regulatory network responding to shifts in the deuterium-to-hydrogen (D/H) ratio. Note the mild (8-10%) upregulation of DAPK1 at 40 and 80 ppm; however, determining whether this change has a biological function requires further investigation.
Relative mRNA expression (y-axis) of DAPK1 and TGFBR2 in A549 lung adenocarcinoma cells cultured for 72 h in media containing 40 ppm, 80 ppm, 150 ppm (control), or 300 ppm deuterium (x-axis). Values are expressed as fold change versus the 150 ppm control. Both genes remain stable at low deuterium, but these expressions are the most upregulated at 300 ppm: DAPK1 by 113% and TGFBR2 by 105%. DAPK1 encodes a pro-apoptotic serine/threonine kinase, while TGFBR2 encodes a receptor mediating TGF-β signaling. Their parallel induction suggests a coordinated stress-response pathway, potentially linking extracellular TGF-β cues to intracellular apoptotic execution under high-deuterium stress. Mild (8–10%) increases at 40–80 ppm may reflect early or subthreshold activation.
Pattern 2: Stable at 40 ppm and 80 ppm, Moderately Upregulated at 300 ppm (32 genes)
This pattern includes genes that are stable at lower deuterium levels but show a moderate increase in expression at 300 ppm.
Genes in Pattern 2: AKT2, BCL2, BCL2L1, CAV1, CCND1, CCND3, CDK6, CLTC, CSK, E2F3, ERBB2, FAS, FOSL2, GAPDH, GRB7, HRAS, IGFBP2, MET, MST1R, NOTCH1, NUMA1, OGG1, PCNA, PDGFA, PIM1, RARA, STAT1, TGFB1, TOP2A, TUBB, TYMS, TYRO3.
Common Characteristics:
- Cell Survival and Apoptosis Regulation: Genes like BCL2, BCL2L1, AKT2, and PIM1 are anti-apoptotic, promoting cancer cell survival. FAS, a pro-apoptotic receptor, also falls into this category, suggesting a fine-tuned balance in cell death pathways.
- Cell Cycle Regulation: CCND1, CCND3, CDK6, E2F3, PCNA, and TOP2A are directly involved in cell cycle progression, particularly the G1/S transition and DNA replication. Their moderate upregulation at 300 ppm supports increased proliferative activity.
- Cell Proliferation and Growth: ERBB2 (HER2), HRAS, MET, MST1R, and PDGFA are components of various growth factors signaling pathways, driving cell proliferation.
- Oncogenic Potential: Many genes in this pattern (e.g., AKT2, BCL2, CCND1, ERBB2, HRAS) are well-known oncogenes, frequently dysregulated in cancer. Their moderate upregulation at 300 ppm further supports the pro-cancer effects of high deuterium.
- DNA Repair and Maintenance: OGG1, PCNA, TOP2A, and TYMS are crucial for DNA repair and synthesis, ensuring genomic stability necessary for rapid cancer cell proliferation.
Connections: These genes are integral to major oncogenic pathways (PI3K/AKT, RAS/MAPK, TGF-beta, NOTCH). Their stability at 40 and 80 ppm and moderate upregulation at 300 ppm suggest a sensitivity to deuterium levels, possibly due to subtle effects on enzyme kinetics or membrane fluidity, which can be altered by deuterium's heavier mass (22).
Pattern 3: Moderately Downregulated at 40 ppm, Stable at 80 ppm, Upregulated at 300 ppm (15 genes)
This pattern characterizes genes that show moderate suppression at 40 ppm, stability at 80 ppm. Of the 15 genes in Pattern 5, 3 strongly upregulated and 12 moderately at 300 ppm.
Genes in Pattern 3: ETS2, HIF1A, HSP90AB1, IRF1, LYN, MLL (KMT2A), MMP9, NPM1, PTPRG, RAF1, RET, RRM1, SERPINE1 (PAI-1), TPR, XPC.
Six of the Pattern 1 genes: BRAF, CD44, CTNNB1, DEK, EGFR, ITGB1 show downregulation between 0.83 and 0.85 in 40 ppm medium thus these genes are close to the border between pattern 1 and 3. Two of the Pattern 2 genes TOP2A and TYMS show similar downregulation.
Common Characteristics:
- Cancer Progression and Survival: Most genes in this pattern are associated with hallmarks of cancer, including cell invasion (MMP9), proliferation (RAF1, RET), angiogenesis (HIF1A, SERPINE1), and cell survival (HSP90AB1, LYN).
- Deuterium Sensitivity: The moderate downregulation observed at 40 ppm and the upregulation at 300 ppm suggest that the expression of these genes is bidirectionally responsive to shifts in deuterium concentration. While these genes may not be directly involved in classical signaling pathways, their functions are nonetheless fundamental, indicating that they play a critical role in maintaining cellular homeostasis under varying D/H ratios.
- Diverse Cellular Processes: These genes span a wide range of cellular functions, including cell adhesion, transcription, signaling, matrix remodeling, DNA repair, and protein stability. This diversity indicates a broad network of cancer-related functions that are collectively modulated by deuterium's effects on cellular redox state.
Connections: These genes form a highly interconnected network involving MAPK, hypoxia, and invasion pathways.
Pattern 4: Strongly Downregulated at 40 ppm, Stable at 80 ppm, Upregulated at 300 ppm (2 genes)
This pattern describes genes that are significantly suppressed at very low deuterium (40 ppm), return to stable levels at 80 ppm, and become moderately or strongly upregulated at high deuterium (300 ppm). This is an interesting patten as the variable deuterium concentration has a large, steady effect on the copy numbers, the expression of the gene.
Genes in Pattern 4: BCL2A1, TGFBR3.
Figure 3 shows the pattern for BCL2A1, TGFBR3 genes. The BCL2A1 gene expression is rapidly increasing in the range of 40-80 ppm deuterium concentrations, in the next concentration range it is stable and in the 300 ppm concentration medium it abruptly increases to be very strongly upregulated. TGFBR3 expression shows a steady increase from strong down regulation to moderate upregulation.
Relative mRNA expression ratio (y-axis) of BCL2A1 and TGFBR3 is shown for A549 cells cultured at 40, 80, 150, and 300 ppm deuterium (x-axis), normalized to the 150 ppm control. BCL2A1, an anti-apoptotic mitochondrial membrane protein, is strongly downregulated at 40 ppm, recovers to stability at 80 ppm, and is sharply induced at 300 ppm. TGFBR3, a co-receptor modulating TGF-β signaling, shows a steady increase from strong downregulation at 40 ppm to moderate upregulation at 300 ppm. These bidirectional responses suggest that extreme deuterium depletion suppresses pro-survival signaling, while enrichment promotes it; a dynamic potentially exploitable in therapeutic modulation of apoptosis and metastasis.
Common Characteristics:
- Cell Survival and Stress Response: BCL2A1 is a potent anti-apoptotic protein, promoting cancer cell survival, especially under stress conditions like oxidative stress. TGFBR3 modulates TGF-β signaling, influencing cell survival and stress responses.
- Cancer Relevance with Context-Dependent Roles: BCL2A1 is oncogenic, supporting cancer cell survival. TGFBR3 can act as a tumor suppressor in early stages but may promote metastasis in advanced cancers.
- Deuterium Sensitivity: BCL2A1 is primarily localized to the mitochondrial outer membrane. Its location on the mitochondrial membrane is essential for its role in cell survival, particularly in conditions of stress or disease, such as in certain cancers where its overexpression contributes to chemotherapy resistance. The strong downregulation at 40 ppm suggests that very low deuterium levels suppress their expression which may contribute to the advantageous effect of DDW. The upregulation at 300 ppm aligns with the general trend of cancer gene activation under high deuterium.
Connections: The distinct response of these genes to varying deuterium levels suggests a complex interplay with cellular stress pathways. The suppression at 40 ppm could be beneficial for anti-cancer strategies, while their upregulation at 300 ppm indicates a pro-survival or pro-metastatic response.
Pattern 5: Stable at All Concentrations (4 genes)
This pattern identifies genes whose expression remains stable across all tested deuterium concentrations (40 ppm, 80 ppm, 150 ppm, and 300 ppm). In the 40 ppm medium, expression levels for these genes decrease to 84–89%, positioning them just above the cutoff for moderate downregulation.
Genes in Pattern 5: FANCG, MUC1, PCTK1, TFE3.
Common Characteristics:
- Cellular Protection and Maintenance: FANCG is involved in DNA repair, ensuring genomic integrity. MUC1 provides mucosal protection and can play roles in cell survival signaling. TFE3 regulates lysosomal biogenesis and metabolic homeostasis. PCTK1 supports cell cycle progression and neuronal differentiation.
- Limited Oncogenic Roles: While MUC1 and TFE3 have known associations with cancer, FANCG and PCTK1 are not primarily considered oncogenic drivers in the same vein as genes in Patterns 1 and 2.
- Stability Across DDW Levels: The consistent stability of these genes across all deuterium concentrations suggests that their functions are essential and tightly regulated, making them robust against variations in deuterium levels. This may indicate that these processes are fundamental for cell survival and are not significantly modulated by deuterium, or that their regulatory mechanisms are insensitive to these specific deuterium shifts.
Connections: These genes primarily function in cellular protection, maintenance, and basic cellular processes. Their stability contrasts with the dynamic regulation observed in other patterns, highlighting a subset of genes whose expression show small dependence on deuterium concentration within the tested range.
Pattern 6: Moderately Downregulated at 40 ppm and 80 ppm, Stable at 300 ppm (1 gene)
This pattern identifies a gene that is moderately suppressed at both 40 ppm and 80 ppm deuterium but shows stable expression at 300 ppm.
Gene in Pattern 6: FGFR4.
Characteristics:
- Oncogenic Receptor Tyrosine Kinase: FGFR4 is a receptor tyrosine kinase involved in cell proliferation, differentiation, and survival, often overexpressed in various cancers (e.g., hepatocellular carcinoma).
- Deuterium Sensitivity: The moderate downregulation at 40 ppm and 80 ppm suggests that lower deuterium levels may suppress FGFR4 expression. Its stability at 300 ppm, unlike the upregulation seen in many other oncogenes at high deuterium, might indicate tight regulatory control or a saturation of its signaling at normal/high deuterium levels.
The suppression of FGFR4 at low deuterium supports the potential anti-cancer effects of DDW by inhibiting oncogenic signaling.
Pattern 7: Stable at 40 ppm, Strongly Downregulated at 80 ppm, Moderately Upregulated at 300 ppm (1 gene)
This unique pattern describes a gene that is stable, even 10% upregulated at very low deuterium, suppressed considerably at 80 ppm deuterium depletion, and then moderately upregulated at high deuterium as shown in Figure 4.
Gene in Pattern 7: ABCB1.
Expression ratio of ABCB1 mRNA (y-axis) is plotted for A549 cells cultured at 40, 80, 150, and 300 ppm deuterium (x-axis), relative to the 150 ppm control. ABCB1, encoding the P-glycoprotein multidrug efflux pump, shows a slight increase (~12%) at 40 ppm, marked downregulation at 80 ppm, and moderate upregulation at 300 ppm. This pattern indicates that moderate deuterium depletion may suppress drug-resistance mechanisms, potentially enhancing chemotherapy efficacy, whereas high deuterium could promote efflux-mediated resistance. The sensitivity of ABCB1 to intermediate depletion levels highlights a narrow window for isotopic modulation to overcome multidrug resistance.
Characteristics:
- ABCB1is a member of the (ATP-Binding Cassette) transporter superfamily.
- Pathway: Transporter Superfamily/Drug Metabolism and Transport.
- Function: It acts as an ATP-powered efflux pump on the cell membrane, actively transporting a wide variety of structurally unrelated hydrophobic molecules (including many chemotherapy drugs like Vinca alkaloids, Taxanes, and Anthracyclines) out of the cell.
- Multidrug Resistance (MDR) Efflux Pump: ABCB1 (P-glycoprotein) is a key transporter conferring multidrug resistance in cancer by actively expelling chemotherapeutic drugs from cells. Overexpression of ABCB1 is a major mechanism by which cancer cells develop resistance to a wide range of chemotherapeutic agents, making treatments ineffective (23).
- Deuterium Sensitivity: The 12% increase of its expression at 40 ppm suggests that very low deuterium concentration may not significantly perturb its expression, but it might have adverse effects. However, the downregulation at 80 ppm indicates that moderate deuterium depletion can suppress ABCB1, potentially enhancing chemotherapy sensitivity by reducing drug efflux. The moderate upregulation at 300 ppm aligns with increased ROS at high deuterium, which can induce ABCB1 expression and promote drug resistance.
This pattern highlights a complex, dose-dependent response of the ABCB1 gene to deuterium, underscoring the extreme sensitivity of this membrane transporter to variations in D concentration. Further investigation is warranted to better characterize the regulation of ABCB1 in response to deuterium shifts, which may be crucial for optimizing the therapeutic use of DDW. Notably, lower deuterium levels—particularly around 80 ppm—may help overcome drug resistance, offering potential benefits for improving chemotherapy efficacy as it was published earlier (9)
Pattern 8: Moderately Upregulated at 40 ppm, Stable at 80 ppm, Upregulated at 300 ppm (2 genes)
This unique pattern contains genes that are moderately upregulated at 40 ppm deuterium by 23% no other gene shows such a considerable upregulation. BCL3 is moderately (almost strongly) upregulated by 49% while PTK7 is strongly upregulated by 66% at 300 ppm deuterium (cf. Supporting Material).
Gene in Pattern 8: BCL3 and PTK7.
Common Characteristics:
- Oncogenic and Pro-Metastatic Roles: BCL3 is an oncogenic transcription co-regulator promoting cell proliferation and survival. PTK7 is a pseudokinase involved in Wnt signaling, cell polarity, and migration, often promoting invasion and metastasis.
- Unique Deuterium Sensitivity: The moderate upregulation at 40 ppm is distinct from most other patterns, suggesting a unique sensitivity to very low deuterium. Their upregulation at 300 ppm aligns with general cancer gene activation at high deuterium.
Connections: This pattern suggests that BCL3 and PTK7 may enhance cancer progression under varying deuterium levels, potentially complicating the anti-cancer effects of DDW. Their regulation appears to be sensitive to subtle shifts in cellular environment induced by deuterium.
Pattern 9: Stable at 40 ppm, Moderately Downregulated at 80 ppm, Stable at 300 ppm (1 gene)
This pattern describes a gene that is stable at very low and high deuterium but suppressed to 76% at 80 ppm deuterium depletion.
Gene in Pattern 9: MYCN.
Characteristics:
- Potent Proto-Oncogene: MYCN is a transcription factor critical for neural development but is frequently amplified or overexpressed in aggressive cancers like neuroblastoma, driving rapid tumor growth.
- Complex Deuterium Response: Its stability at 40 ppm and 300 ppm suggests tight regulatory control or context-specific roles that prevent its activation by extreme deuterium levels. However, the moderate downregulation at 80 ppm indicates that moderate deuterium depletion can suppress MYCN, potentially inhibiting tumor growth, which aligns with DDW's anti-cancer effects in neuroblastoma models.
This unique pattern highlights a nuanced response of MYCN to deuterium, suggesting that optimal deuterium depletion may be required to achieve its suppression, while extreme depletion or enrichment might not have the same inhibitory effect.
3. Discussion
The comprehensive analysis of gene expression in A549 lung cancer cells under varying deuterium concentrations reveals profound and diverse impacts on cancer-related pathways. The initial data cleaning process was crucial, demonstrating that a significant proportion of raw gene expression measurements are unreliable due to low copy numbers or high variability. The retention of 87 high-quality gene expressions provides a robust foundation for drawing biologically meaningful conclusions. While the dataset provides compelling insights into dose-dependent transcriptional responses, it is important to note that the study was conducted using technical duplicates. As such, the findings should be interpreted as exploratory and hypothesis-generating. Future studies incorporating biological replicates will be essential to confirm the reproducibility and generalizability of these observations across cell lines and experimental conditions.
The most general observation is the strong and widespread upregulation of oncogenic and metastatic genes at 300 ppm deuterium. Genes like EGFR, CTNNB1, RAF1, STAT3, and ITGB1, which are central to cell proliferation, survival, and invasion, showed significant increases in expression. This aligns with existing literature suggesting that higher deuterium concentrations are consistent with enhanced tumor aggressiveness (7Error! Bookmark not defined.). The coordinated upregulation of genes involved in MAPK/ERK, PI3K/AKT, Wnt, and JAK/STAT signaling pathways indicates that high deuterium may broadly stimulate pro-cancer signaling networks, potentially by influencing metabolic processes or inducing oxidative stress (24).
Conversely, lower deuterium levels (40 ppm and 80 ppm) generally led to a more stable expression profile for many essential cellular genes (Pattern 5: FANCG, MUC1, PCTK1, TFE3), suggesting that fundamental cellular processes like DNA repair and metabolic homeostasis are robust against deuterium variations. However, specific genes, particularly those involved in cell survival and drug resistance, exhibited interesting patterns of downregulation at lower deuterium levels. For instance, ABCB1 (Pattern 7), a key multidrug resistance transporter, was downregulated at 80 ppm, implying that moderate deuterium depletion could potentially sensitize cancer cells to chemotherapy. Similarly, MYCN (Pattern 9), a potent proto-oncogene, was moderately downregulated at 80 ppm, suggesting an inhibitory effect of DDW on its expression.
The observed discrepancies and higher variability at 40 ppm deuterium warrant particular attention. While some of this variability could be attributed to technical factors, it is also plausible that extreme deuterium depletion induces a significant biological stress response in A549 cells. Genes like AREG (not retained in the cleaned set due to large CV and Z), which showed high CVs and Z-scores at 40 ppm, might represent genuine, albeit highly variable, biological responses to this extreme condition. Deuterium depletion can disrupt mitochondrial function and alter cellular redox states, potentially leading to oxidative stress or changes in enzyme kinetics that disproportionately affect certain sensitive genes (13). The unique upregulation of BCL3 and PTK7 (Pattern 8) at 40 ppm further supports the idea of complex, gene-specific responses to very low deuterium, possibly through transient activation of NF-κB or other stress-responsive pathways. Due to the limited replication, formal statistical testing (e.g., p-values, ANOVA) was not applied. Instead, we relied on fold-change thresholds, coefficient of variation (CV), and Z-score filtering to ensure robustness in gene selection and pattern classification. These descriptive metrics provide a transparent framework for interpreting gene-specific responses and identifying biologically meaningful trends despite the absence of inferential statistics.
Our novel classification system, dividing gene expression responses into nine distinct patterns (Pattern 1-9), elucidates the nuanced impact of deuterium concentration. Genes exhibiting upregulation at 300 ppm (Patterns 1, 2 and 3) were identified as key drivers of cancer progression. Consequently, targeted inhibition of these genes (e.g., EGFR with specific inhibitors) represents a plausible therapeutic strategy to counteract the pro-oncogenic effects associated with elevated deuterium levels. The pronounced, dose-dependent regulation of ABCB1 and other ABC transporters is particularly noteworthy. These membrane proteins influence drug efflux and chemoresistance, and their modulation by deuterium content raises the possibility that DDW could sensitize tumors to chemotherapy by altering transporter activity. This aligns with emerging evidence that metabolic and isotopic environments can influence drug response.
These findings corroborate previous observations that DDW inhibits cell proliferation, whereas DEW stimulates it. This supports the hypothesis that an elevated intracellular D/H ratio is critical for initiating the G1–S phase transition. A gene-specific analysis demonstrates that central cell cycle regulatory genes are activated under higher D/H conditions.
The observation that relatively few genes changed expression in response to DDW suggests the primary regulatory mechanism of cell division may involve preventing an increase in the D/H ratio, rather than reducing it outright. This molecular mechanism underpins the observed effects of DDW in human applications: a decrease in systemic deuterium concentration impedes the activation of genes essential for cell proliferation. Furthermore, DDW elicits a systemic metabolic effect, inducing cellular stress that culminates in a significant increase in ROS levels and subsequent apoptosis.
The intracellular D/H ratio is predominantly influenced by the cell membrane's Na⁺/H⁺ transport system and mitochondrial activity. The latter's influence on deuterium dynamics is modulated by dietary composition: a higher proportion of fat in caloric intake correlates with a lower deuterium concentration in the metabolically produced water. Fat metabolism inherently yields metabolic DDW.
In healthy cells, these two processes are in balance, maintaining D/H homeostasis. However, in cancer cells, the lack of a functional TCA cycle disrupts this equilibrium (24). As a result, cancer cells lack the metabolic counterbalance to the increase in D/H ratio caused by membrane transport activity, allowing deuterium accumulation and potentially contributing to uncontrolled cell proliferation. A recent analysis confirms that an increased D/H ratio is translated into molecular changes by activating genes essential for cell growth (7).
In this experiment, the D concentration was adjusted to levels 70 and 110 ppm lower, and 150 ppm higher than the natural D concentration. This study demonstrated that variations in D concentration can affect the expression of cancer-related genes. This concept builds on an earlier study that examined the effects of D concentration on blood glucose levels in diabetic rats (25). That initial study found that a D concentration of 25 ppm significantly reduced blood sugar levels. In a follow-up study testing seven D concentrations ranging from 75 to 150 ppm, researchers discovered that the optimal range for activating the insulin signaling pathway was between 125 and 140 ppm. These findings suggest that altering the D/H ratio—by increasing or decreasing the Dconcentration by approximately 20–30 ppm relative to natural levels—may be critical for modulating biological pathways supporting by a long-term observational study involving 2649 DDW-consuming patients (26).
Our transcriptomic survey of A549 lung adenocarcinoma cells across controlled deuterium concentrations reveals coordinated, dose-dependent modulation of cancer-relevant gene networks. Although the study is limited to technical replicates and a single cell line, the observed expression patterns refine earlier hypotheses regarding deuterium’s influence on oncogenic signaling, apoptosis regulation, and drug resistance pathways. These findings establish deuterium concentration as a biologically active variable with potential relevance for cancer biology. Future investigations should incorporate biological replication, functional validation, and in vivo models to determine the translational significance of these molecular responses and their therapeutic implications.
4. Materials and Methods
4.1. Experimental Setup and Cell Culture
A549 lung cancer cells were cultured in water media with four different deuterium concentrations: 40 ppm, 80 ppm, 150 ppm (control, representing natural deuterium abundance), and 300 ppm. The specific culture conditions (e.g., media composition, incubation temperature, duration of exposure to varying deuterium levels, see later) were maintained consistently across all groups as published earlier (9).
Production of Deuterium-Depleted Water
DDW with a final deuterium concentration of 25 ppm was produced by HYD LLC for Cancer Research and Drug Development (Budapest) by fractional distillation of purified ordinary tap water, which initially contained 150 ppm of deuterium (equivalent to 0.0158 mol/l HDO). Production followed Good Manufacturing Practice (GMP) standards for quality assurance. The final deuterium concentration was precisely verified using a Liquid-Water Isotope Analyser-24d (manufactured by Los Gatos Research Inc, USA) with a precision of ± 1 ppm. For gene expression studies, DDW at 40 ppm and 80 ppm deuterium concentrations were used, alongside a 150 ppm deuterium concentration as a control. Additionally, deuterium enhanced water (DEW) at 300 ppm was prepared by adding 99.9% heavy water (obtained from Sigma Aldrich, Budapest) to achieve the desired concentration. The reaction H2O + D2O ⇌ 2 HDO, occurs on a picosecond timescale, thus the equilibrium is reached immediately, leading to a mixture of 99.4% H2O and 0.06% HDO and negligible D2O after thorough mixing.
Cell Culture and Deuterium Exposure
A549 cells were seeded at a density of 400,000 cells per well in serum-free medium and incubated for 16 hours prior to experimental manipulation. Following this initial incubation, the serum-free medium was aspirated and replaced with experimental media containing varying concentrations of deuterium (40 ppm, 80 ppm, 150 ppm, and 300 ppm), supplemented with fetal calf serum. Cells were cultured in these media for 72 hours. All experimental groups, including a control group, were maintained under identical conditions. Cells were harvested when they reached approximately 60% confluence.
4.2. Gene Expression Quantification
NanoString technology was employed to quantify the expression of 236 cancer-related genes. NanoString's nCounter Analysis System allows for direct, digital quantification of RNA molecules without the need for reverse transcription or amplification, minimizing potential biases. The technology utilizes unique color-coded molecular barcodes that hybridize to specific RNA targets, which are then counted by an nCounter Analysis System from NanoString digital analyzer, providing precise measurements of gene expression levels.
4.3. Dataset Description
The raw dataset comprised expression measurements for 236 cancer-related genes, with each gene measured in duplicate at each of the four deuterium concentrations. The correlation coefficient between the duplicate measurements is 0.9996, proving that the results are reproducible.
4.4. Data Cleaning Criteria
We used the Coefficient of Variation (CV) and the robust Z-score to find statistically relevant data. The robust Z-score applied here helps identify outliers in gene expression differences while being less sensitive to extreme values than a traditional Z-score (27). For pairwise comparisons of gene expression ratios, we quantified variability using a dimensionless coefficient of variation defined as CV = (x₁ − x₂)/ (x₁ + x₂). This approach avoids inflated variance estimates from degrees-of-freedom corrections and provides a biologically interpretable measure of dispersion, especially under low-replicate conditions.
To ensure the reliability of downstream analyses, stringent data cleaning criteria were applied:
- Low Average Copy Number Exclusion: Genes were excluded if their average copy number was less than 20 at the 150 ppm deuterium level. This filter targets low-expression genes, which are inherently more susceptible to technical noise and may lack significant biological relevance in this context.
- High Coefficient of Variation Exclusion: Genes were excluded if their CV exceeded 20% and |Z| > 3 at the 150 ppm deuterium level. A high CV and Z indicate excessive variability between replicate measurements, potentially stemming from experimental errors or significant biological heterogeneity that could confound interpretation.
4.5. Gene Expression Classification
After data cleaning, 87 genes were retained and categorized based on their expression ratios relative to a 150 ppm control. This reduction highlights the presence of significant noise and variability in the raw measurements.
Gene expression changes were classified using the following thresholds:
- Strongly Upregulated: Ratio > 1.5 (expression is more than 50% higher than in the control medium).
- Moderately Upregulated: 1.2 < Ratio ≤ 1.5 (expression is 20% to 50% higher).
- Stable: 0.83 ≤ Ratio ≤ 1.2 (expression is within the range of -17% to +20% of the control).
- Moderately Downregulated: 0.67 ≤ Ratio < 0.83 (expression is 17% to 33% lower).
- Strongly Downregulated: Ratio < 0.67 (expression is less than two-thirds of the control).
These thresholds combine a standard 1.5-fold cutoff for significant changes with a smaller 1.2-fold cutoff to capture more subtle, yet potentially biologically relevant, shifts in expression.
4.6. Data Cleaning Outcomes
- Retained Genes: Genes such as BRCA1, TP53, and EGFR, known for their critical roles as tumor suppressors or oncogenes, were consistently retained. These genes exhibited stable expression (CV <10% across deuterium levels), indicating reliable measurements suitable for studying deuterium effects. The results are summarized in Table S1.
- Excluded Genes: A significant number of genes were excluded due to either low expression (e.g., AKT1, with an average copy number of 4 at 40 ppm) or high variability (e.g., AREG, with a CV of 33.33% at 40 ppm and a robust Z-score of 7.83). While the exclusion of low-expression genes helps reduce technical noise, the removal of high-variability genes, even those with high Z-scores, warrants careful consideration as they might represent genuine biological responses to deuterium-induced stress rather than mere experimental artifacts. Notably, the 40 ppm deuterium condition exhibited the largest discrepancies and highest average CV (~18% compared to ~10-14% for other levels), would have contributed disproportionately to gene exclusions. This suggests that extreme deuterium depletion may induce greater biological variability or be more susceptible to technical inconsistencies. These problems will be investigated in future studies.
The NanoString experiment was repeated twice for four retained cancer genes BRAF, CLTC, LYN and RAF1 in an independent experiment. For BRAF the copy numbers in the new experiment show a systematic increase of 100-105, but the pattern does not change (see 3.3, pattern 1). The two experiments for CLTC, LYN show excellent agreement, while for LYN a negligible systematic decrease of 2-12 can be observed which again does not change the outcome of the detailed analysis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org: The dataset was structured as a 22-column Excel file, with the following organization:.Column A: Gene Code (e.g., ABL1, BRCA1). Column B: Accession number. Columns C–G: Measurements for 40 ppm deuterium, including replicate counts ((40/1), (40/2)), calculated average, coefficient of variation (CV), and robust Z-score. Columns H–L, M–Q, R–V: Equivalent metrics for 80 ppm, 150 ppm, and 300 ppm deuterium, respectively.
Author Contributions
Conceptualization, G.I.C. and G.S.; methodology, G.I.C.; software, G.I.C.; validation, G.I.C., G.S., I.S. and A.P.; formal analysis, G.I.C.; investigation, G.I.C.; resources, G.I.C.; data curation, G.I.C.; writing—original draft preparation, G.I.C.; writing—review and editing, G.I.C., G.S.; visualization, G.I.C.; supervision, G.S., I.S.; project administration, G.S.; funding acquisition, G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.:.
Acknowledgments
We: wish to thank Lajos I. Nagy and László G Puskas for the gene expression data.
References
- Heron M, "Deaths: Leading Causes for 2019," Natl Vital Stat Rep, vol. 70(9), pp. 1-114, 2021.
- Španěl P, Shestivska V, Chippendale TWE, Smith D. Determination of the deuterium abundances in water from 156 to 10,000 ppm by SIFT-MS. J Am Soc Mass Spectrom. 2011;22(2):179–186.
- Maréchal, Y. IR spectra and dynamics of H₂O (D₂O, HDO) molecules in a still poorly known liquid: water. NATO ASI Ser C Math Phys Sci. 1993;435:149–168.
- Jancsó, G. Isotope Effects in Handbook of Nuclear Chemistry (eds. Vértes, A., Nagy, S. & Klencsár, Z.) 85-116 (Kluwer Academic Publishers: Dordrecht, Netherland, 2013).
- Sobczyk L, Obrzud M, Filarowski A. H/D isotope effects in hydrogen bonded systems. Molecules. 2013 Apr 16;18(4):4467-76. [CrossRef] [PubMed]
- Somlyai, G.; et al. Naturally occurring deuterium is essential for the normal growth rate of cells. FEBS Lett 1993, 317, 1–4. [Google Scholar] [CrossRef] [PubMed]
- G. Somlyai, B. Z. Kovács, I. Somlyai, A. Papp, L. I. Nagy and L. G. Puskás, "Deuterium depletion inhibits lung cancer cell growth and migration in vitro and results in severalfold increase of median survival time of non-small cell lung cancer patients receiving conventional therapy," Cancer Res The, vol. 9(2), pp. 12-19, 2021.
- Lu, Y. and Chen, H. Deuterium-Depleted Water in Cancer Therapy: A Systematic Review of Clinical and Experimental Trials, Nutrients, 2024 16(9), p. 1397.
- Kovács BZ, Puskás LG, Nagy LI, Papp A, Gyöngyi Z, Fórizs I, et al. Blocking the Increase of Intracellular Deuterium Concentration Prevents the Expression of Cancer-Related Genes, Tumor Development, and Tumor Recurrence in Cancer Patients. Cancer Control. 2022;10732748211068963.
- Lei, Z., Su, N., Li, M., Pan Z., Liu K., Zhang Y. Unraveling the role of deuterium in cancer: mechanisms, detection techniques, and therapeutic potential. Mol Divers (2025). [CrossRef]
- Cong F.S., Zhang Y.R., Sheng H.C., Ao Z.H., Zhang S.Y., Wang J.Y. Deuterium-Depleted Water Inhibits Human Lung Carcinoma Cell Growth by Apoptosis. Exp. Ther. Med. 1(2), 277-283.
- Boros LG, Somlyai I, Kovács BZ, Puskás LG, Nagy LI, Dux L, Farkas G, Somlyai G. Deuterium Depletion Inhibits Cell Proliferation, RNA and Nuclear Membrane Turnover to Enhance Survival in Pancreatic Cancer. Cancer Control. 2021 Jan-Dec;28:1073274821999655. [CrossRef]
- Zhang X, Gaetani M, Chernobrovkin A, Zubarev RA. Anticancer Effect of Deuterium Depleted Water – Redox Disbalance Leads to Oxidative Stress. Cancer Control. 2019;18:2373–87.
- Z. Gyöngyi, F. Budán, I. Szabó, I. Ember, K. Kiss, K. Krempels, I. Somlyai and G. Somlyai, "Deuterium depleted water effects on survival of lung cancer patients and expression of Kras, Bcl2, and Myc genes in mouse lung," Nutr Cancer, vol. 65(2), pp. 240-246, 2012.
- Gyöngyi, Z. & Somlyai, G. Deuterium depletion can decrease the expression of c-myc, Ha-ras and p53 gene in carcinogen-treated mice. In Vivo 14, 437–439; PMID.10904878 (2000).
- Fortina P, Surrey S. Digital mRNA profiling. Nat Biotechnol. 2008 Mar;26(3):293-4. [CrossRef]
- Kontomanolis EN, Koutras A, Syllaios A, Schizas D, Mastoraki A, Garmpis N, et al. Role of Oncogenes and Tumor-suppressor Genes in Carcinogenesis: A Review. Anticancer Res. 2020 Nov;40(11):6009-6015. [CrossRef]
- Carlo M. Croce, MD, Oncogenes and Cancer, 2008, N Engl J Med 2008;358:502-511. [CrossRef]
- Jang, C. W., Chen, C. H., Chen, C. C., Chen, J. Y., Su, Y. H., & Chen, R. H. (2002). TGF-β induces apoptosis through Smad-mediated expression of DAP-kinase. Nature Cell Biology, 4(1), 51–58. [CrossRef]
- Kim, N., Chen, D., Zhou, X. Z., & Lee, T. H. (2019). Death-associated protein kinase 1 phosphorylation in neuronal cell death and neurodegenerative disease. In International Journal of Molecular Sciences (Vol. 20, Issue 13). MDPI AG. [CrossRef]
- Liu, R. M., & Desai, L. P. (2015). Reciprocal regulation of TGF-β and reactive oxygen species: A perverse cycle for fibrosis. In Redox Biology (Vol. 6, pp. 565–577). Elsevier B.V. [CrossRef]
- Brown, M. F., Thurmond, R. L., Dodd, S. W., Otten, D., & Beyer, K. (2002). Elastic deformation of membrane bilayers probed by deuterium NMR relaxation. Journal of the American Chemical Society, 124(28), 8471–8484. [CrossRef]
- Xiao H, Zheng Y, Ma L, Tian L, Sun Q. Clinically-Relevant ABC Transporter for Anti-Cancer Drug Resistance. Front Pharmacol. 2021;12:648407. [CrossRef]
- Somlyai, G. Deuterium Depletion – A New Way in Curing Cancer and Preserving Health. Redwood City, CA: PublishDrive; 2021.
- Molnár M, Horváth K, Dankó T, Somlyai I, Kovács BZ, Somlyai G. Deuterium-depleted water stimulates GLUT4 translocation in the presence of insulin, which leads to decreased blood glucose concentration. Mol Cell Biochem. 2021;476(12):4507–16. [CrossRef] [PubMed]
- Somlyai G, Papp A, Somlyai I, Kovács BZ, Debrődi M. Real-World Data Confirm That the Integration of Deuterium Depletion into Conventional Cancer Therapy Multiplies the Survival Probability of Patients. Biomedicines. 2025;13:876. [CrossRef]
- Maronna RA, Martin RD, Yohai VJ. Robust Statistics: Theory and Methods (with R). 2nd ed. Chichester, UK: Wiley; 2019.
Figure 2.
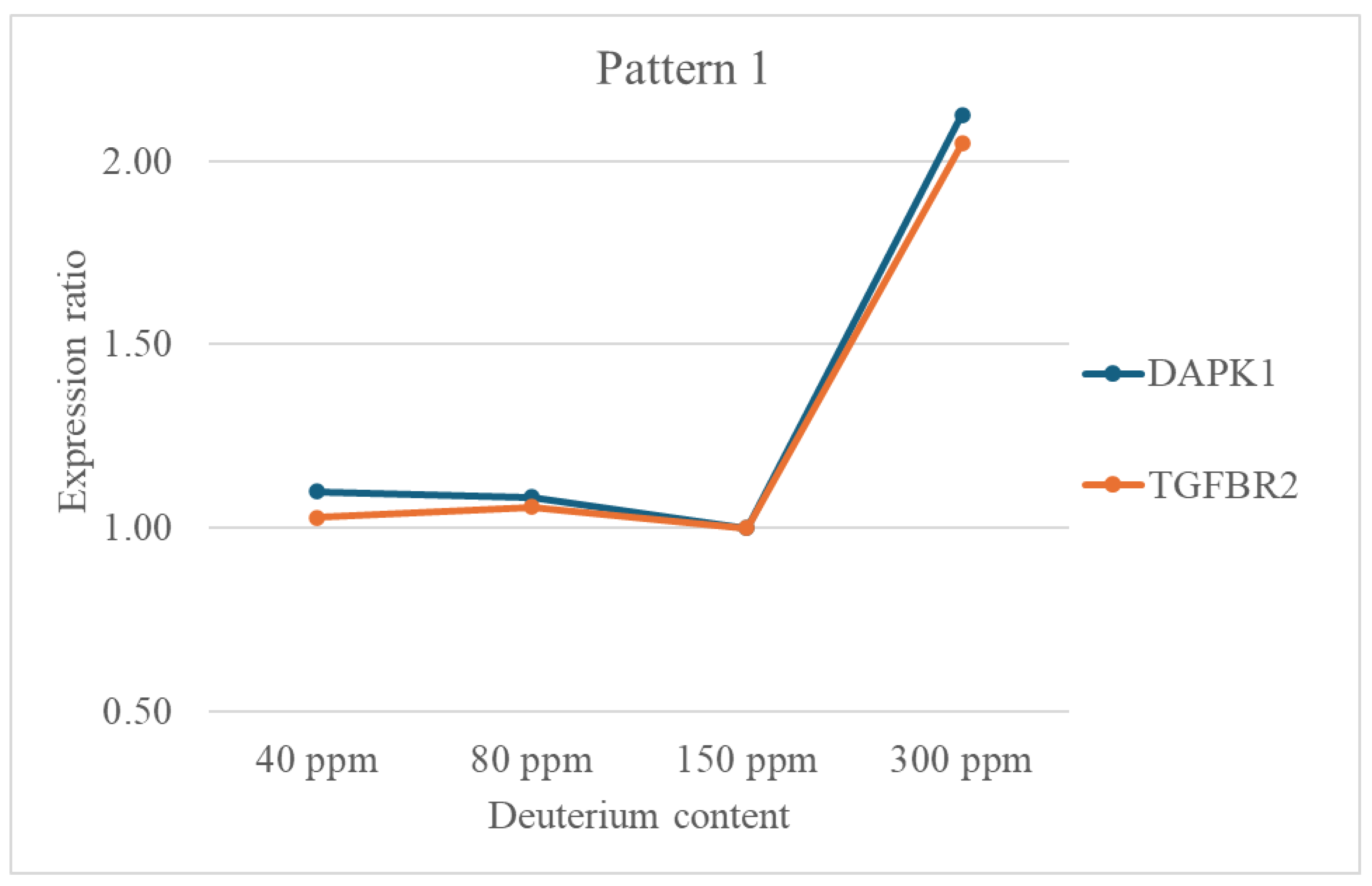
Figure 3.
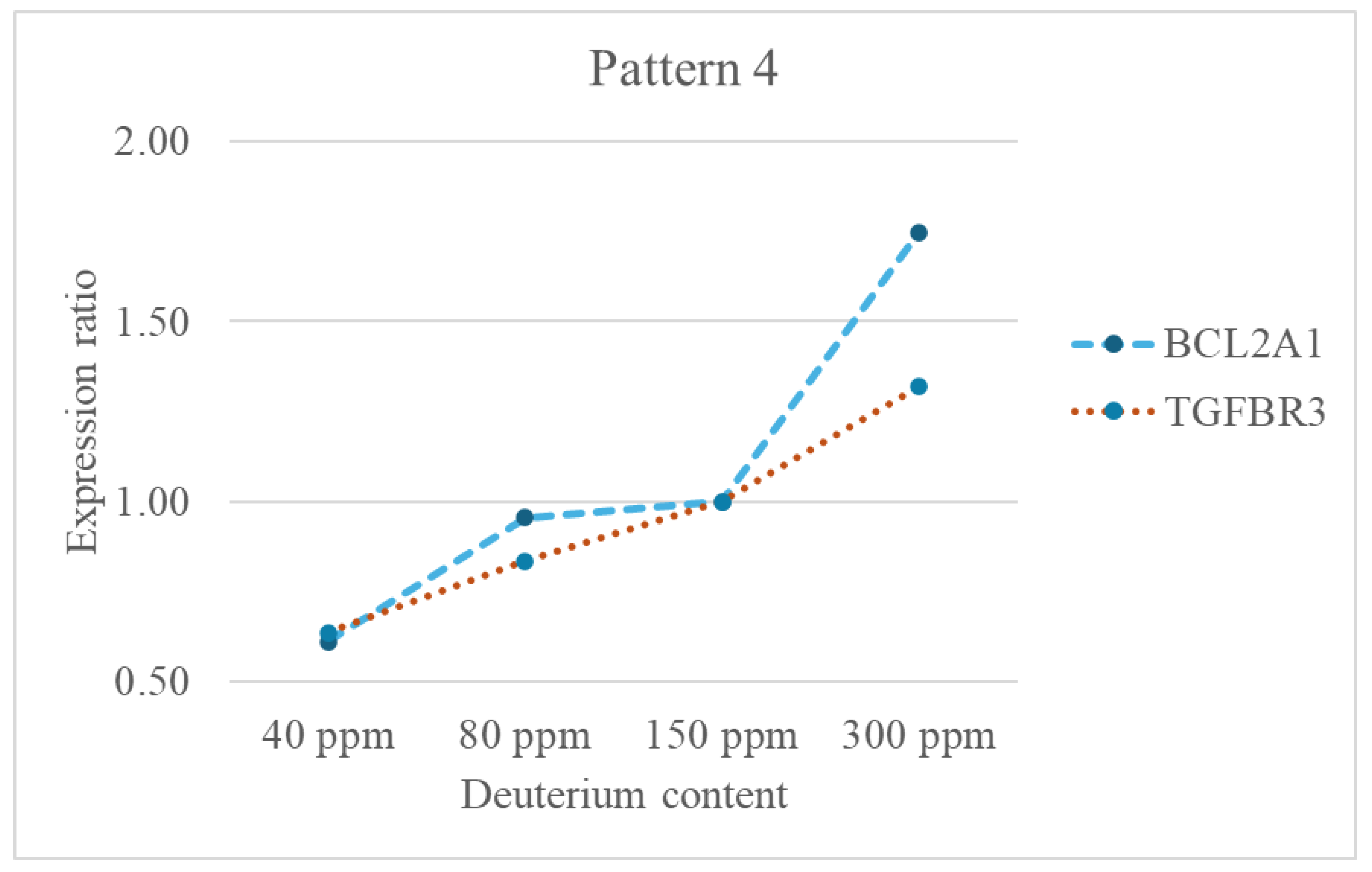
Figure 4.
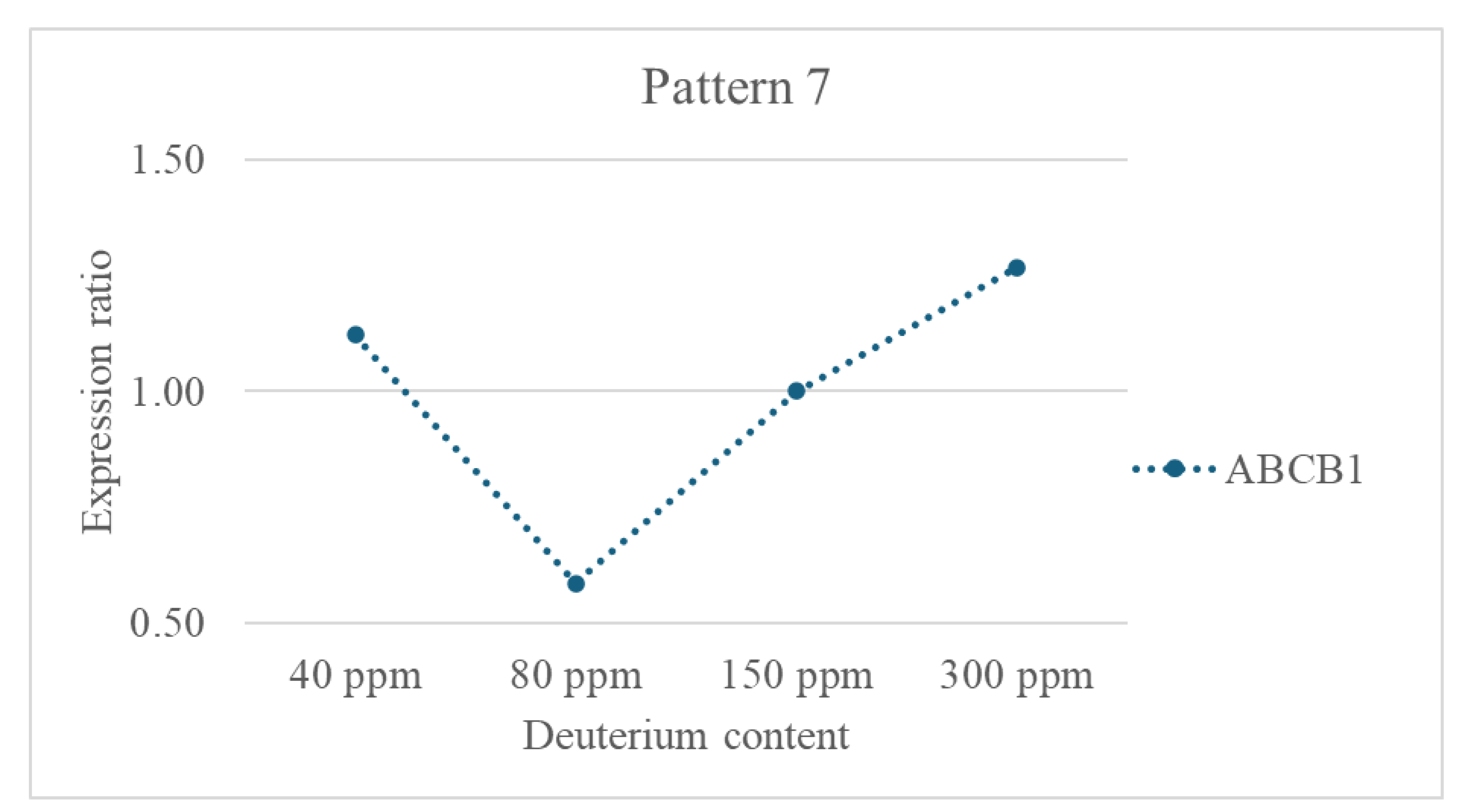

Table 1.
Summary of Expression Ratios of the 87 retained genes Relative to 150 ppm Control in Each Classification Category by Deuterium Content.
Table 1.
Summary of Expression Ratios of the 87 retained genes Relative to 150 ppm Control in Each Classification Category by Deuterium Content.
| Deuterium Content | Strong Up (>1.5) | Moderate Up (1.2–1.5) |
Stable (0.83–1.2) | Moderate Down (0.67–0.83) | Strong Down (<0.67) |
|---|---|---|---|---|---|
| 40 ppm | 0 | 2 | 66 | 17 | 2 |
| 80 ppm | 0 | 0 | 80 | 6 | 1 |
| 300 ppm | 44 | 28 | 15 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



