
Submitted:
06 October 2025
Posted:
07 October 2025
You are already at the latest version
Abstract
Pulmonary hypertension (PH) causes morbidity and mortality in sickle cell disease (SCD). The release of heme during hemolysis triggers endothelial dysfunction and contributes to PH. Long non-coding RNAs (lncRNAs) may play a pivotal role in endo-thelial dysfunction and PH pathogenesis. This study assessed the regulatory role of the lncRNA heme oxygenase-1 (HMOX1) axis in SCD-PH pathogenesis. Total RNAs were isolated from 15-17 weeks sickle cell (SS) mice and littermate controls (AA) lungs and subjected to lncRNA expression profiling using the Arrystar™ lncRNA array. Raw signal intensities were normalized in quantile method by GeneSpring GX software. Volcano plot filtering was used to screen for differentially expressed (DE) lncRNAs and mRNAs with statistical significance (fold change >1.5, p< 0.05). A total of 2,302 lncRNAs were upregulated and a total of 2,546 lncRNAs were downregulated in lungs of SS mice compared to AA mice. To validate DE of lncRNAs and mRNAs, 6 lncRNAs and 6 mRNAs were selected for quantitative PCR. Metastasis-associated lung adenocarcino-ma transcript 1 (MALAT1) and HMOX1 levels were found to be increased in SS samples. Human pulmonary artery endothelial cells (HPAECs) were treated with hemin and analyzed for MALAT1 or HMOX1 levels, which were increased in hemin-treated HPAECs. Lastly, loss of MALAT1 function reduced HMOX1 levels and increased markers of endothelial dysfunction (ET-1 and VCAM1). These results suggest that SCD modulates the MALAT1-HMOX1 axis and further characterization of MALAT1 function may provide new insights into SCD-associated endothelial dysfunction, PH pathogen-esis, and identify novel therapeutic targets.
Keywords:
lncRNA
; MALAT1
; sickle cell disease
; pulmonary hypertension
; endothelial dysfunction
1. Introduction
Pulmonary hypertension (PH) is a recognized complication of sickle cell disease (SCD) associated with high morbidity and mortality, regardless of PH severity.[1,2,3] SCD is characterized by chronic intravascular hemolysis that generates excess heme in the blood and recurrent vaso-occlusive insults associated with painful crisis, acute chest syndrome, and chronic organ damage and failure.[4] Intravascular hemolysis activates pulmonary endothelium, which in turn upregulates hemin activating adhesion molecule expression, such as vascular endothelial cell adhesion molecule 1 (VCAM1), leading to endothelial dysfunction.[5] Endothelial dysfunction caused by hemolysis represents a fundamental derangement that is central to end-organ dysfunction in SCD-associated PH (SCD-PH). Current therapies for SCD-PH that target vascular dilation do not reverse the abnormal, proliferative, vascular cell phenotypes that underlie the pulmonary vasculopathy in this disorder. These considerations indicate that new therapeutic approaches are needed. For example, endothelin-1 (ET-1) is elevated in both SCD and PH patients.[6,7,8,9,10,11,12] ET-1 causes endothelial dysfunction, vasoconstriction, vascular cell migration, and proliferation in PH.[7,13] However, clinical trials in SCD-PH examining the ET-1 receptor antagonist, bosentan, produced insignificant decreases in pulmonary vascular resistance and conflicting results.[14] Therefore, the current study seeks to address the complex pathobiology of SCD-PH and this critical derangement by examining novel targetable molecular mechanisms of endothelial dysfunction in SCD-PH.
Emerging evidence suggests long non-coding RNAs (lncRNAs) play important roles in regulating gene expression in multiple critical cellular pathways including regulation of uncontrolled cell proliferation, cell growth, and apoptosis.[15,16,17,18] LncRNA can promote or inhibit gene expression at the level of mRNA transcription, translation, RNA stabilization, and through their ability to interact with microRNAs (miRNAs).[19] Moreover, mounting evidence demonstrates the role of lncRNAs in PH pathobiology and could serve as viable therapeutic molecular targets.
Heme oxygenase 1 (HMOX1) is an essential enzyme that degrades heme released during intravascular hemolysis and promotes endothelial health through antioxidant, cytoprotective, and heme catabolic pathways. HMOX1 also protects endothelial function in SCD.[20] In relation to PH, HMOX-1 expression is reduced in the lungs of patients with PH, while knockout of HMOX1 exacerbates hypoxia-induced PH and right ventricular hypertrophy (RVH) in mice.[22] Furthermore, HMOX1 overexpression in alveolar epithelial cells decreased hypoxia-induced PH.[23] Intriguingly, increased HMOX1 levels have been observed in neutrophils, mononuclear cells[24], and tissues of patients and transgenic mice with SCD.[25] However, the mechanisms and functional significance underlying these HMOX1 increases via lncRNAs in SCD have not been defined.
In this study, we report a novel link between lncRNA MALAT1 and HMOX1 in SCD-PH and that MALAT1 could serve as an effective therapeutic target in SCD-PH.
2. Materials and Methods
2.1. Sickle Mouse Lung Tissues
Lung tissues were isolated from wild-type (AA) mice and from the Townes humanized mouse model of SCD (SS) (JAX:013071).[26]
2.2. Human Pulmonary Artery Endothelial Cells (HPAECs)
HPAECs (passages 2-6, ScienCell, Carlsbad, CA) were incubated with 5 µM hemin (HEM) or dimethyl sulfoxide (DMSO, used as control [CON]) for up to 72 hours as previously reported.[27]
2.3. Assessment of PH in MALAT1-Overpressing Sickle Cell Mice
Selected AA or SS mice (15-17 weeks old) were treated with control fluorescent protein (GFP) or adenoviral MALAT1 (AdMALAT1) particles using intranasal injection as previously reported.[27] At day 7 following transfection, mice were subjected to measurements of right ventricular systolic pressure (RVSP) and right ventricular hypertrophy (RVH) as reported.[28] All experiments using mice were approved by the Institutional Animal Care and Use Committee of the Atlanta Veterans Affairs Medical Center and were conducted in accordance with institutional standards for the humane treatment of laboratory animals.
2.4. Reagents
HPAECs were obtained from ScienCell Research Laboratories (Carlsbad, CA). HMOX1 antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The following reagents were obtained from Sigma-Aldrich (St. Louis, MO): GAPDH antibody, hemin, fetal bovine serum (FBS), dimethyl sulfoxide (DMSO), and actinomycin D.
2.5. Cell-Based Enzyme-Linked Immunosorbent Assay (ELISA)
To measure HMOX1 activity in HPAECs, an In-Cell ELISA assay was performed and normalized to GAPDH levels within the same well using the Odyssey Imaging System (LI-COR Biosciences, Lincoln, NE), according to the manufacturer’s instructions.
2.6. MALAT1 Loss or Gain of Function
For MALAT1 loss-of-function experiments, HPAECs were transfected with MALAT1 antisense oligonucleotide or appropriate control (1 nM, Qiagen, Germantown MD) using Lipofectamine 3000 transfection reagent (Thermo Fisher Scientific, Waltham, MA), according to the manufacturer’s instructions. At 6 hours following transfection, the transfection medium was replaced with endothelial cells growth medium (EGM) containing 5% FBS, and cells were incubated at room temperature for 72 hours. Selected HPAECs were treated ± HEM (5 μM) or DMSO directly to the culture medium for up to 72 hours. HPAEC lysates were harvested and analyzed for MALAT1, ET-1, VCAM1, and HMOX1 levels by qRT-PCR.
To overexpress MALAT1, HPAECs were transfected with adenovirus-mediated MALAT1 (AdMALAT1, 5 multiplicity of infection, MOI) or green fluorescent protein (GFP)-tagged control adenovirus particles, as reported.[29] At 6 hours following transfection, media were replaced with fresh 5% FBS EGM, and HPAECs were treated with DMSO or HEM (5 µM) for 72 hours. HPAEC lysates were then harvested and examined for MALAT1, ET-1, VCAM1, and HMOX1 levels using qRT-PCR analysis.
2.7. HMOX1 siRNA
For HMOX1 loss-of-function, HPAECs were transfected with scrambled or HMOX1 RNAi duplexes (10 nM, Integrated DNA Technologies, Coralville, IA) using Lipofectamine 3000 transfection reagent (Invitrogen), according to the manufacturer’s instructions. After 6 hours of transfection, the transfection medium was replaced with EGM containing 5% FBS, and cells were incubated at room temperature for 72 hours. To determine whether depletion of HMOX1 affects HMOX1 activity, the In-Cell ELISA assay was performed. Further, HPAEC lysates were harvested and examined for ET-1 and VCAM1 levels using qRT-PCR analysis.
2.8. Hematoxylin and Eosin (H&E) Staining
To examine pulmonary vascular remodeling in mouse small arterioles, lung sections were stained with H&E staining according to the manufacturer’s instructions (Abcam, Cambridge, MA). For each lung, sections from 10 separate tissue blocks were analyzed. Images were captured using Keyence imaging system (Itasca, IL).
2.9. Messenger RNA Stability Assay
To inhibit de novo HMOX1 mRNA synthesis, HPAECs were transfected with ASO control or ASO MALAT1 and incubated for 72 hours, and then 5 μg/ml actinomycin D (Apexbio, Houston, TX) was treated time-dependently. Total RNAs were isolated using mirVana kit (Thermo Fisher Scientific), and HMOX1 mRNA levels were measured by qRT-PCR analysis. HMOX1 mRNA half-life was determined by comparing its level at each time point to the baseline level before actinomycin D treatment.
2.10. Messenger RNA Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
To measure MALAT1, ET-1, VCAM1, and HMOX1 levels, total RNAs were isolated from mouse lungs and HPAECs using the mirVana kit (Thermo Fisher Scientific). MALAT1, ET-1, VCAM1, and HMOX1 mRNA levels were quantified by qRT-PCR, as previously described[30]. GAPDH mRNA levels were used as a control.
2.11. Western Blot Analysis
Homogenates from mouse lungs or from hemin-treated HPAECs were subjected to Western blot analysis, as previously reported.[30] Primary HMOX1 mouse monoclonal antibody (1:500 dilution, Cat # SC-136960, 32 kDa) was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). GAPDH rabbit polyclonal antibody (1:10,000 dilution, Cat # G9545, 37 kDa) was purchased from Sigma-Aldrich (St. Louis, MO). Relative protein levels were visualized using Li-Cor proprietary software, quantified Image J software, and normalized to GAPDH levels within the same lane.
2.12. Statistical Analysis
All data are presented as mean ± standard error of the mean (SE). Shapiro-Wilk test was used to assess normality. For data that were not normally distributed, the Mann-Whitney U test was used to compare two independent groups, and the Kruskal-Wallis analysis of variance (ANOVA) was used to compare three or more independent groups. Post hoc analyses were performed using the Student Neuman Keuls test to detect differences between specific groups. In studies comparing only two experimental groups, data were analyzed with Student’s t-test to determine the significance of treatment effects. The level of statistical significance was taken as p<0.05. Statistical analyses were carried out using GraphPad Prism, Version 8.0 software (LaJolla, CA).
3. Results
3.1. MALAT1 Expression Is Increased in SS Mouse Lungs and in Hemin-Treated HPAECs
To define differentially expressed lncRNAs and molecular mechanisms of lncRNAs, we examined unbiased lncRNA microarray in SS mouse lungs (ArraystarTM). Microarray study revealed that 2,302 lncRNAs were upregulated (red) and 2,546 lncRNAs were downregulated (green) in lungs of SS mice compared to AA mice (Figure 1A). To validate the differentially expressed lncRNAs, we selected six upregulated and six downregulated conserved lncRNAs for quantitative analyses. Among 12 lncRNAs which were assessed, MALAT1 was consistently upregulated in both lncRNA microarray and qRT-PCR analysis in SS lungs (Figure 1B). In parallel, we found that MALAT1 is induced in SS mouse lungs (Figure 1C) and in hemin-treated HPAECs (Figure 1D) suggesting that hemolysis might play a role in inducing MALAT1 expression. Therefore, we further explored whether intravascular hemolysis in SCD stimulates MALAT1 expression to protect against PH and endothelial dysfunction.
3.2. MALAT1 Regulates Expression of Endothelial Dysfunction Markers ET-1 and VCAM1 In Vitro
To investigate the role of MALAT1 in the pathogenesis of SCD-PH, we examined its functional impact on endothelial behavior in HPAECs. We first examined whether loss of MALAT1 can lead to endothelial dysfunction in hemin-treated HPAECs to simulate a chronic hemolytic state in SS mice. We showed that antisense oligonucleotide (ASO)-mediated depletion of HPAEC MALAT1 increased expression of markers of EC dysfunction, including ET-1 (Figure 2A) and vascular cell adhesion molecule-1 (VCAM1) (Figure 2B). Further, depletion of MALAT1 augmented ET-1 and VCAM1 expression in hemin-treated HPAECs (Figure 2A-B). In parallel, we examined whether gain of MALAT1 could protect HPAECs against hemin-induced endothelial dysfunction using adenovirus-mediated MALAT1 (AdMALAT1) particles. We found that MALAT1 overexpression attenuated ET-1 (Figure 2C) and VCAM1 expression (Figure 2D). MALAT1 overexpression effectively protected HPAECs from hemin-induced ET-1 and VCAM1 expression (Figure 2C-D). Collectively, these results support that MALAT1 plays a protective role in endothelial function, and MALAT1 treatment may attenuate EC dysfunction in hemin-treated HPAECs.
3.3. MALAT1 Overexpression Reduces PH, RVH, and Vascular Remodeling with Downregulation of Endothelial Cells Dysfunction Markers, ET-1 and VCAM1
We previously reported that SCD-PH is associated with vascular remodeling and endothelial dysfunction.[27,31] To determine the functional significance of MALAT1 in SCD-PH, we examined if gain of MALAT1 function regulates SCD-PH, RV hypertrophy (RVH), vascular remodeling, and markers of endothelial dysfunction. As shown in Figure 3, AdMALAT1 in SS mouse lungs attenuated SCD-PH as seen by a reduction in RV systolic pressure (RVSP; Figure 3A), RVH (Figure 3B), vascular remodeling (Figure 3C), ET-1 (Figure 3D), and VCAM1 levels (Figure 3E). Collectively, the data presented in Figure 3 support our hypothesis that MALAT1 plays a protective role in vascular remodeling in SCD-PH, and MALAT1 treatment may attenuate EC dysfunction, vascular remodeling, and PH.
3.4. HMOX1 Protects Endothelial Function in SCD-PH
Because HMOX1 is an essential enzyme that degrades heme released during intravascular hemolysis and protects endothelial function in SCD, we measured HMOX1 levels and its activity in vivo and in vitro. As shown in Figure 4, HMOX1 levels are increased in SS mouse lungs (Figure 4A and 4B) and in hemin-treated HPAECs (Figure 4C and 4D). To determine if loss of HMOX1 modulates endothelial function, HPAECs were transfected with siHMOX1 and treated with hemin. Consistent with data shown in Figure 4A-D, treatment of HPAEC with hemin upregulated HMOX1 activity (Figure 4E) and markers for endothelial dysfunction (Figure 4F and 4G). Further, we demonstrated that knockdown of HMOX1 reduced not only HMOX1 activity in hemin-treated HPAECs (Figure 4E), but increased ET-1 (Figure 4F) and augmented hemin-induced VCAM1 expression (Figure 4G). These data suggest that hemolysis upregulates HMOX1 and that HMOX1 might have a protective role in hemolysis-induced endothelial dysfunction.
3.5. MALAT1 Regulates HMOX1 Expression
To assess whether MALAT1 can mediate HMOX1, we overexpressed MALAT1 in vivo and in vitro using adMALAT1. In Figure 5, we showed that MALAT1 overexpression upregulated HMOX1 in AA mice lungs and augmented HMOX1 expression in SS mice lung (Figure 5A). In parallel, we showed that MALAT1 overexpression upregulates HPAECs HMOX1 expression but did not further augment the HMOX1 expression in hemin-treated HPAECs (Figure 5B). In contrast, ASO-mediated depletion of HPAEC MALAT1 reduced hemin-induced HMOX1 levels in HPAECs and attenuated hemin-induced HMOX1 expression (Figure 5C). We further found that knockdown of MALAT1 reduced HMOX1 mRNA stability (Figure 5D) and HMOX1 activity (Figure 5E). These findings indicate that MALAT1 regulates the constitutive expression of HMOX1. Taken together, the findings in Figure 5 support the postulate that MALAT1 regulates endothelial function via HMOX1 activation, a previously undescribed pathobiological pathway in SCD-related endothelial dysfunction and PH (Figure 5F).
4. Discussion
In this study, we present data supporting a functional role of MALAT1 in SCD-PH pathogenesis. Using lncRNA microarray and confirmed by quantitative PCR, we showed that MALAT1 is upregulated in a chronic hemolysis environment including in SS mouse lungs and hemin-treated HPAECs. Further, depletion of MALAT1 promoted and exacerbated hemin-induced endothelial dysfunction as reflected by an upregulation of both ET-1 and VCAM1. Conversely, MALAT1 overexpression suppressed ET-1 and VCAM1 expression and atteuated hemin-induced ET-1 and VCAM1 expression in HPAECs. Moreover, in vivo MALAT1 overexpression decreased RVSP, RVH, endothelial proliferation, and markers of endothelial dysfunction. To understand the mechanism underlying the protective effect of MALAT1 in SCD, we identified HMOX1 as a candidate target for MALAT1. We show that HMOX1 levels increase in SS mouse lungs and hemin-treated HPAECs. Similar to MALAT1, suppression of HMOX1 is associated with an increase in endothelial cell dysfunction suggesting that HMOX1 and MALAT1 affect HPAECs in a similar fashion. In parallel, we show that MALAT1 directly mediates HMOX1 as MALAT1 overexpression upregulated HMOX1 while inhibition of MALAT1 reciprocally suppressed HMOX1. Finally, inhibition of MALAT1 is associated with HMOX1 RNA instability. Collectively, these data support our hypothesis that the MALAT1–HMOX1 axis plays an important role in hemolysis-induced endothelial dysfunction and, ultimately, hemolysis-associated pulmonary hypertension as seen in SCD lungs.
Long non-coding RNAs (lncRNA) are abundantly expressed in all tissues and have been shown to participate in cellular functions and disease pathogenesis.[32] lncRNAs can regulate gene and protein expression by acting as gene activators or repressors, mRNA stabilizers, miRNA precursors and inhibitors, and direct interaction with DNA or protein.[32] LncRNA MALAT1 consists of ~8,000 bp and is highly conserved and abundantly expressed in various cells and tissues including in PAECs.[33] MALAT1 polymorphisms either increase the risk of developing PH (i.e. rs619586A>G single nucleotide polymorphism) or are protective (i.e. G variant).[34] The protective effect of MALAT1 G variant is through its function as a competitive endogenous RNA for miR-214, which leads to an inhibition of vascular EC proliferation and migration.[34] Several lncRNAs have been implicated in PH pathogenesis including MALAT1. Specifically, MALAT1 is highly expressed in hypoxia-induced pulmonary artery smooth muscle cells (PASMCs) and PAECs.[35] In this study, we utilized a microarray for lncRNAs to identify possible novel diagnostic or therapeutic targets and identified MALAT1 as the only lncRNA that was consistently upregulated in SS lungs. As vascular EC dysfunction is one of first triggers leading to development of PH,[36] we assessed the function of MALAT1 on EC dysfunction in a chronic hemolysis model as seen in people with SCD. As presented in this report, modulation of MALAT1 has an inverse relationship with markers of EC dysfunction. Further, overexpression of MALAT1 protected EC against hemin-induced EC dysfunction. These findings support the functional significance of MALAT1 in hemin (chronic hemolysis)-induced EC dysfunction.
The pathobiology of SCD-PH involves abnormal proliferation of pulmonary vascular wall cells, vascular remodeling, and muscularization of small pulmonary vessels.[37] These structural and functional alterations in the pulmonary vasculature increase pulmonary vascular resistance and promote progressive right-sided heart failure and death. Sickle cell mice spontaneously develop PH as shown by elevated pulmonary artery pressure, decreased cardiac output, and right-heart failure.[38] Furthermore, our group and others showed that SS mice have evidence of intravascular hemolysis, which induces EC pro-inflammatory responses, including IL-8, IL-6, VCAM-1, and ET-1 levels.[31,39] Given that MALAT1 is protective against EC dysfunction, we assessed the function of MALAT1 in SS mouse lungs. Consistent with the in vitro findings, we show that overexpression of MALAT1 in SS mouse lungs attenuated RVSP, RV hypertrophy, vascular remodeling, and markers of EC dysfunction. These data suggest that therapeutic upregulation of MALAT1 could prevent or possibly reverse histological and physiological changes consistent with SCD-PH in SS mouse lungs.
A hallmark of SCD is intravascular hemolysis leading to a release of free heme (hemin) leading to disrupted EC function and exacerbated oxidative stress.[40,41] HMOX1 is an essential enzyme that degrades heme released during intravascular hemolysis, protects EC function in SCD, and is implicated in PH.[21,42,43] HMOX1 activity promotes antioxidant, cytoprotective, and heme catabolic pathways as well as protects endothelial function.[21] Reduced HMOX1 expression has been observed in the lungs of patients with PH[44] and hypoxia exacerbates right ventricular dilatation in Hmox1 KO mice.[22] Conversely, alveolar epithelial cell-targeted human HMOX1 overexpression decreased hypoxia-induced PH.[23] Intriguingly, increases in HMOX1 were observed in neutrophils, mononuclear cells,[24] and tissues in patients and transgenic mice with SCD.[25] In this study, we show that lungs from SS mice express higher levels of HMOX1 compared to littermate control lungs and hemin treatment induces HMOX-1 expression in HPAECs. Further, inhibition of HMOX1 expression exacerbates EC dysfunction in hemin-treated HPAECs. These findings are consistent with previous reports suggesting that HMOX1 is induced in the setting of hemolysis, and the lack of HMOX1 promotes EC dysfunction.
The regulatory networks involving various types of non-coding RNAs have been increasingly explored. These non-coding RNAs include microRNAs (miRNAs or miR) and lncRNAs, which can form a complex regulatory network. Several miRNAs have been shown to regulate HMOX1, including miR-377 in combination with miR-217,[45] let-7,[46] miR-155,[47] miR-135a, and miR-135b.[48] However, less is known about the role of lncRNA in HMOX1 regulation. Given the reciprocal changes between MALAT1 and HMOX1 in SS mouse lungs, we speculated that MALAT1 might play a regulatory role on HMOX1. Consistent with our speculation, we showed that overexpression of MALAT1 upregulated HMOX1, while inhibition of MALAT1 resulted in suppression of HMOX1 in mouse lungs and HPAECs.
In summary, the current study identifies novel functions of the MALAT1–HMOX1 axis in the pathogenesis of SCD-PH. The outcomes of the proposed studies will advance our understanding of lncRNA MALAT1 in SCD-PH and provide new therapeutic targets for treating people with SCD-PH and related diseases.
Author Contributions
Conception, hypothesis delineation, and design by B.Y.K.; acquisition of data, analysis, and interpretation by V.S., S.S.C., J.M., D.R.A., S.O.A., R.L.S., B.T.K., C.P., C.M.H., and B.Y.K.; writing of the article by V.S., D.R.A., S.O.A., B.T.K., C.P., C.M.H., and B.Y.K.
Funding
This study was supported by funding from VA BLR&D Merit Review Award (I01 BX004263 to C.M.H.), NIH National Heart, Lung, and Blood Institute R01 grants (HL102167 to C.M.H. and R.L.S., HL119291 to C.P. 1R01AA029690-01A1 to V.S, and B.Y.K., and HL133053 to B.Y.K.), NIH National Institute of Alcoholism and Abuse grants (AA027662 to V.S.), and the National Center for Advancing Translational Sciences of the National Institutes of Health UL1TR002378 (BYK).
Informed Consent Statement
The authors have no competing interests. The contents of this report represent the views of the authors and do not represent the views of the Department of Veterans Affairs or the United States Government.
Conflicts of Interest
All authors declare that they have no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| PH | Pulmonary hypertension |
| SCD | Sickle Cell Disease |
| MALAT1 | Metastasis-associated lung adenocarcinoma transcript 1 |
| SS | Sickle cell |
| HMOX1 | Heme oxygenase-1 |
| lncRNAs | Long non-coding RNAs |
| AA | Littermate controls |
| HPAECs | Human pulmonary artery endothelial cells |
| VCAM1 | Vascular endothelial cell adhesion molecule 1 |
| RVH | Right ventricular hypertrophy |
| ASO | Antisense oligonucleotide |
| ET-1 | Endothelin-1 |
| HEM | Hemin |
| RVSP | Right ventricular systolic pressure |
References
- Lilienfeld, D.E.; Rubin, L.J. Mortality from Primary Pulmonary Hypertension in the United States, 1979–1996. Chest 2000, 117, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Platt, O.S.; Brambilla, D.J.; Rosse, W.F.; Milner, P.F.; Castro, O.; Steinberg, M.H.; Klug, P.P. Mortality in Sickle Cell Disease—Life Expectancy and Risk Factors for Early Death. New Engl. J. Med. 1994, 330, 1639–1644. [Google Scholar]
- Powars, D.; A Weidman, J.; Odom-Maryon, T.; Niland, J.C.; Johnson, C. Sickle cell chronic lung disease: prior morbidity and the risk of pulmonary failure. Medicine (Baltimore) 1988, 67, 66–76. [Google Scholar] [PubMed]
- Rees, D.C.; Williams, T.N.; Gladwin, M.T. Sickle-Cell Disease. Lancet 2010, 376, 2018–2031. [Google Scholar] [CrossRef] [PubMed]
- Bensinger, T.A.; Gillette, P.N. Hemolysis in Sickle Cell Disease. Arch Intern Med 1974, 133, 624–31. [Google Scholar] [CrossRef]
- Giaid, A.; Yanagisawa, M.; Langleben, D.; Michel, R.P.; Levy, R.; Shennib, H.; Kimura, S.; Masaki, T.; Duguid, W.P.; Stewart, D.J. Expression of Endothelin-1 in the Lungs of Patients with Pulmonary Hypertension. New Engl. J. Med. 1993, 328, 1732–1739. [Google Scholar] [CrossRef]
- McLaughlin, V.V.; McGoon, M.D. Pulmonary Arterial Hypertension. Circulation 2006, 114, 1417–1431. [Google Scholar] [CrossRef]
- Stewart, D.J.; Levy, R.D.; Cernacek, P.; Langleben, D. Increased Plasma Endothelin-1 in Pulmonary Hypertension: Marker or Mediator of Disease? Ann. Intern. Med. 1991, 114, 464–469. [Google Scholar] [CrossRef]
- Yoshibayashi, M.; Nishioka, K.; Nakao, K.; Saito, Y.; Matsumura, M.; Ueda, T.; Temma, S.; Shirakami, G.; Imura, H.; Mikawa, H. Plasma endothelin concentrations in patients with pulmonary hypertension associated with congenital heart defects. Evidence for increased production of endothelin in pulmonary circulation. Circulation 1991, 84, 2280–2285. [Google Scholar] [CrossRef]
- Hammerman, S.I.; Kourembanas, S.; Conca, T.J.; Tucci, M.; Brauer, M.; Farber, H.W. Endothelin-1 Production during the Acute Chest Syndrome in Sickle Cell Disease. Am. J. Respir. Crit. Care Med. 1997, 156, 280–285. [Google Scholar] [CrossRef]
- Rybicki, A.C.; Benjamin, L.J. Increased Levels of Endothelin-1 in Plasma of Sickle Cell Anemia Patients. Blood 1998, 92, 2594–2596. [Google Scholar]
- Werdehoff, S.G.; Moore, R.B.; Hoff, C.J.; Fillingim, E.; Hackman, A.M. Elevated plasma endothelin-1 levels in sickle cell anemia: Relationships to oxygen saturation and left ventricular hypertrophy. Am. J. Hematol. 1998, 58, 195–199. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Minniti, C.P.; Machado, R.F.; Coles, W.A.; Sachdev, V.; Gladwin, M.T.; Kato, G.J. Endothelin receptor antagonists for pulmonary hypertension in adult patients with sickle cell disease. Br. J. Haematol. 2009, 147, 737–743. [Google Scholar] [CrossRef]
- Puthanveetil, P.; Gutschner, T.; Lorenzen, J. MALAT1: A Therapeutic candidate for a broad spectrum of vascular and cardiorenal complications. Hypertens. Res. 2020, 43, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ma, L. New Insights into Long Non-Coding RNA MALAT1 in Cancer and Metastasis. Cancers 2019, 11, 216. [Google Scholar] [CrossRef]
- Jaé, N.; Heumüller, A.W.; Fouani, Y.; Dimmeler, S. Long non-coding RNAs in vascular biology and disease. Vasc. Pharmacol. 2019, 114, 13–22. [Google Scholar]
- Moráňová, L.; Bartošík, M. Long Non-Coding RNAs – Current Methods of Detection and Clinical Applications. Klin. Onkol. 2019, 32, 65–71. [Google Scholar] [CrossRef]
- Bunch, H. Gene regulation of mammalian long non-coding RNA. Mol. Genet. Genom. 2018, 293, 1–15. [Google Scholar]
- Liu, Y.; Jing, F.; Yi, W.; Mendelson, A.; Shi, P.; Walsh, R.; Friedman, D.F.; Minniti, C.; Manwani, D.; Chou, S.T.; et al. Ho-1(Hi) Patrolling Monocytes Protect against Vaso-Occlusion in Sickle Cell Disease. Blood 2018, 131, 1600–1610. [Google Scholar] [CrossRef]
- Yachie, A. Heme Oxygenase-1 Deficiency and Oxidative Stress: A Review of 9 Independent Human Cases and Animal Models. Int. J. Mol. Sci. 2021, 22, 1514. [Google Scholar] [CrossRef]
- Yet, S.-F.; Perrella, M.A.; Layne, M.D.; Hsieh, C.-M.; Maemura, K.; Kobzik, L.; Wiesel, P.; Christou, H.; Kourembanas, S.; Lee, M.-E. Hypoxia induces severe right ventricular dilatation and infarction in heme oxygenase-1 null mice. J. Clin. Investig. 1999, 103, R23–R29. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Christou, H.; Hsieh, C.-M.; Liu, Y.; Dhawan, V.; Abraham, N.G.; Perrella, M.A.; Mitsialis, S.A.; Kourembanas, S. Targeted expression of heme oxygenase-1 prevents the pulmonary inflammatory and vascular responses to hypoxia. Proc. Natl. Acad. Sci. USA 2001, 98, 8798–8803. [Google Scholar]
- Lanaro, C.; Franco-Penteado, C.F.; Albuqueque, D.M.; O Saad, S.T.; Conran, N.; Costa, F.F. Altered levels of cytokines and inflammatory mediators in plasma and leukocytes of sickle cell anemia patients and effects of hydroxyurea therapy. J. Leukoc. Biol. 2009, 85, 235–242. [Google Scholar]
- Ghosh, S.; Tan, F.; Yu, T.; Li, Y.; Adisa, O.; Mosunjac, M.; Ofori-Acquah, S.F. Global Gene Expression Profiling of Endothelium Exposed to Heme Reveals an Organ-Specific Induction of Cytoprotective Enzymes in Sickle Cell Disease. PLOS ONE 2011, 6, e18399. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-C.; Sun, C.-W.; Ryan, T.M.; Pawlik, K.M.; Ren, J.; Townes, T.M. Correction of sickle cell disease by homologous recombination in embryonic stem cells. Blood 2006, 108, 1183–1188. [Google Scholar] [CrossRef]
- Kang, B.-Y.; Park, K.; Kleinhenz, J.M.; Murphy, T.C.; Sutliff, R.L.; Archer, D.; Hart, C.M. Peroxisome Proliferator-Activated Receptor Gamma Regulates the V-Ets Avian Erythroblastosis Virus E26 Oncogene Homolog 1/Microrna-27a Axis to Reduce Endothelin-1 and Endothelial Dysfunction in the Sickle Cell Mouse Lung. Am. J. Respir. Cell Mol. Biol. 2017, 56, 131–144. [Google Scholar]
- Nisbet, R.E.; Bland, J.M.; Kleinhenz, D.J.; Mitchell, P.O.; Walp, E.R.; Sutliff, R.L.; Hart, C.M. Rosiglitazone Attenuates Chronic Hypoxia–Induced Pulmonary Hypertension in a Mouse Model. Am. J. Respir. Cell Mol. Biol. 2010, 42, 482–490. [Google Scholar] [CrossRef]
- Kang, B.Y.; Park, K.K.; Kleinhenz, J.M.; Murphy, T.C.; Green, D.E.; Bijli, K.M.; Yeligar, S.M.; Carthan, K.A.; Searles, C.D.; Sutliff, R.L.; Hart, C.M. Ppargamma Activation Reduces Hypoxia-Induced Endothelin-1 Expression through Upregulation of Mir-98. Am J Respir Cell Mol Biol 2015. [Google Scholar]
- Kang, B.-Y.; Kleinhenz, J.M.; Murphy, T.C.; Hart, C.M.; Silpanisong, J.; Kim, D.; Williams, J.M.; Adeoye, O.O.; Thorpe, R.B.; Pearce, W.J.; et al. The Ppargamma Ligand Rosiglitazone Attenuates Hypoxia-Induced Endothelin Signaling in Vitro and in Vivo. Am. J. Physiol. Cell. Mol. Physiol. 2011, 301, L881–L891. [Google Scholar] [CrossRef] [PubMed]
- Jang, A.J.; Chang, S.S.; Park, C.; Lee, C.M.; Benza, R.L.; Passineau, M.J.; Ma, J.; Archer, D.R.; Sutliff, R.L.; Hart, C.M.; Kang, B.Y. Ppargamma Increases Huwe1 to Attenuate Nf-Kappab/P65 and Sickle Cell Disease with Pulmonary Hypertension. Blood Adv 2021, 5, 399–413. [Google Scholar]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Non-coding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin β4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Zeng, Q.; Zhang, P.; Li, G.; Xie, Q.; Cheng, Y. Functional Polymorphism of Lncrna Malat1 Contributes to Pulmonary Arterial Hypertension Susceptibility in Chinese People. Clin Chem Lab Med 2017, 55, 38–46. [Google Scholar]
- Brock, M.; Schuoler, C.; Leuenberger, C.; Bühlmann, C.; Haider, T.J.; Vogel, J.; Ulrich, S.; Gassmann, M.; Kohler, M.; Huber, L.C. Analysis of hypoxia-induced noncoding RNAs reveals metastasis-associated lung adenocarcinoma transcript 1 as an important regulator of vascular smooth muscle cell proliferation. Exp. Biol. Med. 2017, 242, 487–496. [Google Scholar]
- Kurakula, K.; Smolders, V.F.E.D.; Tura-Ceide, O.; Jukema, J.W.; Quax, P.H.A.; Goumans, M.-J. Endothelial Dysfunction in Pulmonary Hypertension: Cause or Consequence? Biomedicines 2021, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, M. Molecular pathogenesis of pulmonary arterial hypertension. J. Clin. Investig. 2008, 118, 2372–2379. [Google Scholar] [CrossRef]
- Hsu, L.L.; Champion, H.C.; Campbell-Lee, S.A.; Bivalacqua, T.J.; Manci, E.A.; Diwan, B.A.; Schimel, D.M.; Cochard, A.E.; Wang, X.; Schechter, A.N.; et al. Hemolysis in sickle cell mice causes pulmonary hypertension due to global impairment in nitric oxide bioavailability. Blood 2007, 109, 3088–3098. [Google Scholar] [CrossRef]
- Zhang, P.; Nguyen, J.; Abdulla, F.; Nelson, A.T.; Beckman, J.D.; Vercellotti, G.M.; Belcher, J.D. Soluble MD-2 and Heme in Sickle Cell Disease Plasma Promote Pro-Inflammatory Signaling in Endothelial Cells. Front. Immunol. 2021, 12, 632709. [Google Scholar] [CrossRef]
- Potoka, K.P.; Gladwin, M.T. Vasculopathy and pulmonary hypertension in sickle cell disease. Am J Physiol Lung Cell Mol Physiol 2015, 308, L314–L324. [Google Scholar]
- Reiter, C.D., X. Wang, J.E. Tanus-Santos, N. Hogg, R.O. Cannon, 3rd, A.N. Schechter, and M. T. Gladwin. Cell-Free Hemoglobin Limits Nitric Oxide Bioavailability in Sickle-Cell Disease. Nat Med 2002, 8, 1383-9. [Google Scholar]
- Abid, S.; Kebe, K.; Houssaïni, A.; Tomberli, F.; Marcos, E.M.; Bizard, E.M.; Breau, M.M.; Parpaleix, A.; Tissot, C.-M.; Maitre, B.; et al. New Nitric Oxide Donor NCX 1443: Therapeutic Effects on Pulmonary Hypertension in the SAD Mouse Model of Sickle Cell Disease. J. Cardiovasc. Pharmacol. 2018, 71, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Liang, O.D.; Mitsialis, S.A.; Chang, M.S.; Vergadi, E.; Lee, C.; Aslam, M.; Fernandez-Gonzalez, A.; Liu, X.; Baveja, R.; Kourembanas, S. Mesenchymal Stromal Cells Expressing Heme Oxygenase-1 Reverse Pulmonary Hypertension. Stem Cells 2011, 29, 99–107. [Google Scholar]
- Solari, V.; Piotrowska, A.P.; Puri, P. Expression of heme oxygenase-1 and endothelial nitric oxide synthase in the lung of newborns with congenital diaphragmatic hernia and persistent pulmonary hypertension. J. Pediatr. Surg. 2003, 38, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.D.; Chen, C.; Nguyen, J.; Thayanithy, V.; Subramanian, S.; Steer, C.J.; Vercellotti, G.M. Regulation of Heme Oxygenase-1 Protein Expression by miR-377 in Combination with miR-217. J. Biol. Chem. 2011, 286, 3194–3202. [Google Scholar] [CrossRef]
- Hou, W.; Tian, Q.; Steuerwald, N.M.; Schrum, L.W.; Bonkovsky, H.L. The let-7 microRNA enhances heme oxygenase-1 by suppressing Bach1 and attenuates oxidant injury in human hepatocytes. Biochim Biophys Acta 2012, 1819, 1113–1122. [Google Scholar]
- Pulkkinen, K.H.; Ylä-Herttuala, S.; Levonen, A.-L. Heme oxygenase 1 is induced by miR-155 via reduced BACH1 translation in endothelial cells. Free. Radic. Biol. Med. 2011, 51, 2124–2131. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shen, H.; Huang, Q.; Li, Q. The Circular Rna Cdr1as Regulates the Proliferation and Apoptosis of Human Cardiomyocytes through the Mir-135a/Hmox1 and Mir-135b/Hmox1 Axes. Genet. Test. Mol. Biomarkers 2020, 24, 537–548. [Google Scholar] [CrossRef]
Figure 1.
Hemolysis induces MALAT1 levels in vivo and in vitro. Whole lung homogenates were collected from littermate control (AA) and sickle cell (SS) mice at age 15-17 weeks. (A) Total RNAs were subjected to lncRNA expression profiling using the Arrystar™ lncRNA array. (B) Selected lncRNAs from SS lung were subjected to qRT-PCR for validation of microarray data. (C) Lung MALAT1 levels were measured with qRT-PCR and expressed relative to lung GAPDH. *p<0.05 vs AA, n=4. (D) HPAECs were treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for 72 hours. Mean HPAEC MALAT1 levels were measured with qRT-PCR. All bars represent the mean MALAT1 levels relative to GAPDH ± SE as fold-change vs. CON. *p<0.05 vs. CON, n=6.
Figure 1.
Hemolysis induces MALAT1 levels in vivo and in vitro. Whole lung homogenates were collected from littermate control (AA) and sickle cell (SS) mice at age 15-17 weeks. (A) Total RNAs were subjected to lncRNA expression profiling using the Arrystar™ lncRNA array. (B) Selected lncRNAs from SS lung were subjected to qRT-PCR for validation of microarray data. (C) Lung MALAT1 levels were measured with qRT-PCR and expressed relative to lung GAPDH. *p<0.05 vs AA, n=4. (D) HPAECs were treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for 72 hours. Mean HPAEC MALAT1 levels were measured with qRT-PCR. All bars represent the mean MALAT1 levels relative to GAPDH ± SE as fold-change vs. CON. *p<0.05 vs. CON, n=6.
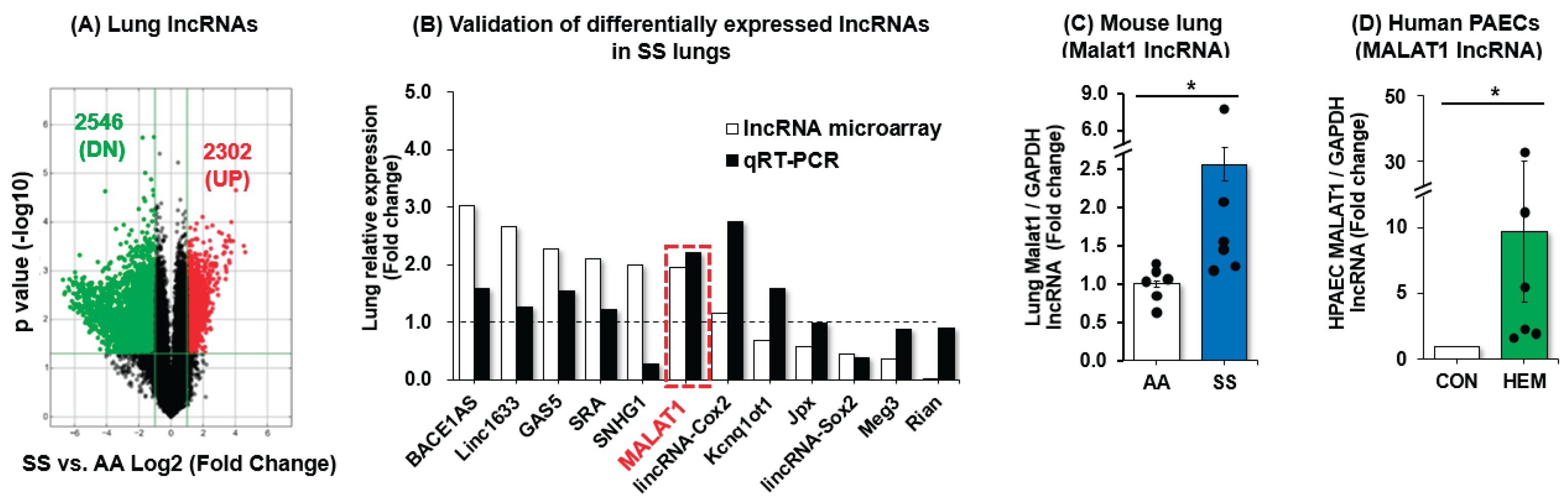
Figure 2.
MALAT1 inversely mediates ET-1 and VCAM1 expression. HPAECs were treated with scrambled ASO or ASO MALAT1 (25 nM) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC ET-1 (A) or VCAM1 (B) levels were measured with qRT-PCR. All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/ASO MALAT1(-). *p<0.05 vs. CON/ASO MALAT1(-), +p<0.05 vs. HEM/ASO MALAT1(-), n=4. In parallel, HPAECs were treated with scrambled AdGFP or AdMALAT1 particles (5 MOI) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC ET-1 (C) or VCAM1 (D) levels were measured with qRT-PCR. All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/AdMALAT1(-). *p<0.05 vs. CON/AdMALAT1(-), +p<0.05 vs. HEM/AdMALAT1(-), n=3-4.
Figure 2.
MALAT1 inversely mediates ET-1 and VCAM1 expression. HPAECs were treated with scrambled ASO or ASO MALAT1 (25 nM) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC ET-1 (A) or VCAM1 (B) levels were measured with qRT-PCR. All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/ASO MALAT1(-). *p<0.05 vs. CON/ASO MALAT1(-), +p<0.05 vs. HEM/ASO MALAT1(-), n=4. In parallel, HPAECs were treated with scrambled AdGFP or AdMALAT1 particles (5 MOI) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC ET-1 (C) or VCAM1 (D) levels were measured with qRT-PCR. All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/AdMALAT1(-). *p<0.05 vs. CON/AdMALAT1(-), +p<0.05 vs. HEM/AdMALAT1(-), n=3-4.
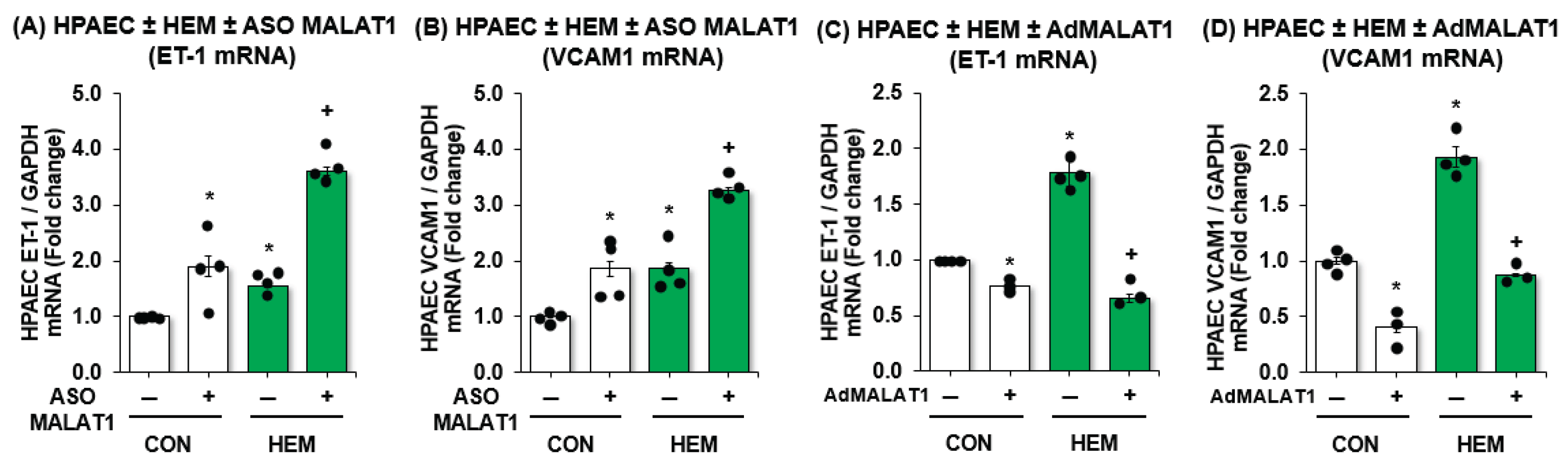
Figure 3.
MALAT1 overexpression attenuates RV systolic pressure, RVH, vascular remodeling, and endothelial dysfunction markers ET-1 and VCAM1 gene expressions. AA and SS mice were infected with scrambled AdGFP or AdMALAT1 particles and 2 weeks later, right ventricular systolic pressure (RVSP) (A) and RVH (B) were measured. Each bar represents mean RVSP or RVH ± SE in mmHg. n=3-5. The resulting lungs were imaged (C) and subjected to qRT-PCR analysis. Mean ET-1 (D) or VCAM1 (E) levels examined as fold-change vs. AA/AdMALAT1(-). *p<0.05 vs. AA/AdMALAT1(-), +p<0.05 vs. SS/AdMALAT1(-), n=3-5.
Figure 3.
MALAT1 overexpression attenuates RV systolic pressure, RVH, vascular remodeling, and endothelial dysfunction markers ET-1 and VCAM1 gene expressions. AA and SS mice were infected with scrambled AdGFP or AdMALAT1 particles and 2 weeks later, right ventricular systolic pressure (RVSP) (A) and RVH (B) were measured. Each bar represents mean RVSP or RVH ± SE in mmHg. n=3-5. The resulting lungs were imaged (C) and subjected to qRT-PCR analysis. Mean ET-1 (D) or VCAM1 (E) levels examined as fold-change vs. AA/AdMALAT1(-). *p<0.05 vs. AA/AdMALAT1(-), +p<0.05 vs. SS/AdMALAT1(-), n=3-5.
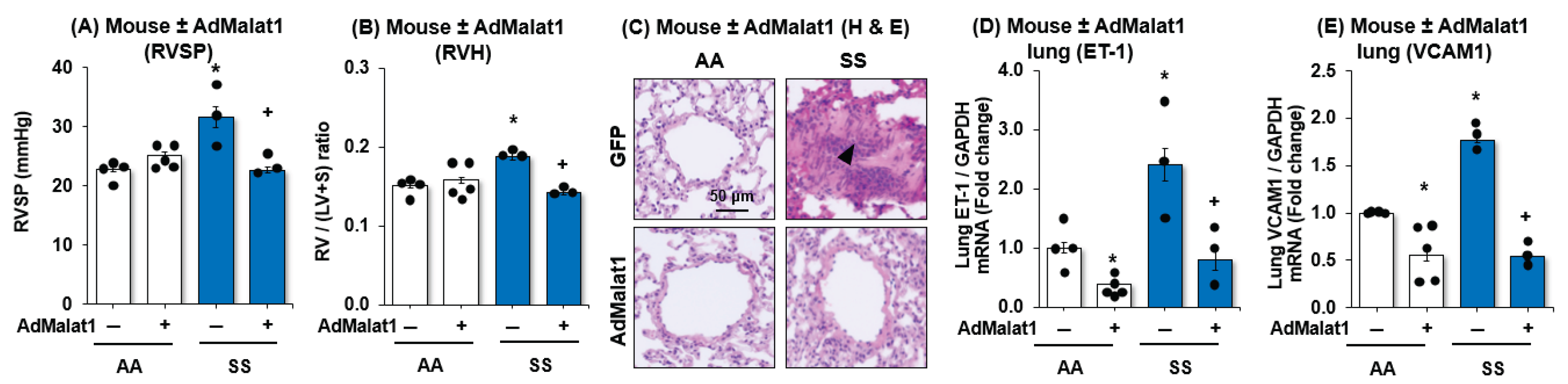
Figure 4.
HMOX1 protects endothelial function in SCD-PH. (A,B) Whole lung homogenates were collected from wildtype (AA) or sickle cell (SS) mice at age 15-17 weeks. Mean HPAEC HMOX1 protein (A) or mRNA (B) were measured with Western blots or qRT-PCR, respectively. Each bar represents the mean ± SE expressed as fold-change vs. AA. *p<0.05 vs. n=6. (C,D) HPAECs were treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for 72 hours. Mean HPAEC HMOX1 protein (C) or mRNA (D) were measured with Western blots or qRT-PCR, respectively. Each bar represents the mean ± SE expressed as fold-change vs. CON. *p<0.05 vs. n=3-6. (E-G) HPAECs were treated with scrambled siRNA or siHMOX1 for 6 hours. Selected cells were treated CON or HEM for 72 hours. Mean HPAEC HMOX1 activity was measured with In-Cell ELISA assay (E). Mean HPAEC ET-1 (F) or VCAM1 (G) levels were measured with qRT-PCR. All bars represent the mean levels relative to GAPDH ± SE expressed as fold-change vs. CON/siHMOX1(-), +p<0.05 vs. HEM/siHMOX1(-), n=3-4.
Figure 4.
HMOX1 protects endothelial function in SCD-PH. (A,B) Whole lung homogenates were collected from wildtype (AA) or sickle cell (SS) mice at age 15-17 weeks. Mean HPAEC HMOX1 protein (A) or mRNA (B) were measured with Western blots or qRT-PCR, respectively. Each bar represents the mean ± SE expressed as fold-change vs. AA. *p<0.05 vs. n=6. (C,D) HPAECs were treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for 72 hours. Mean HPAEC HMOX1 protein (C) or mRNA (D) were measured with Western blots or qRT-PCR, respectively. Each bar represents the mean ± SE expressed as fold-change vs. CON. *p<0.05 vs. n=3-6. (E-G) HPAECs were treated with scrambled siRNA or siHMOX1 for 6 hours. Selected cells were treated CON or HEM for 72 hours. Mean HPAEC HMOX1 activity was measured with In-Cell ELISA assay (E). Mean HPAEC ET-1 (F) or VCAM1 (G) levels were measured with qRT-PCR. All bars represent the mean levels relative to GAPDH ± SE expressed as fold-change vs. CON/siHMOX1(-), +p<0.05 vs. HEM/siHMOX1(-), n=3-4.
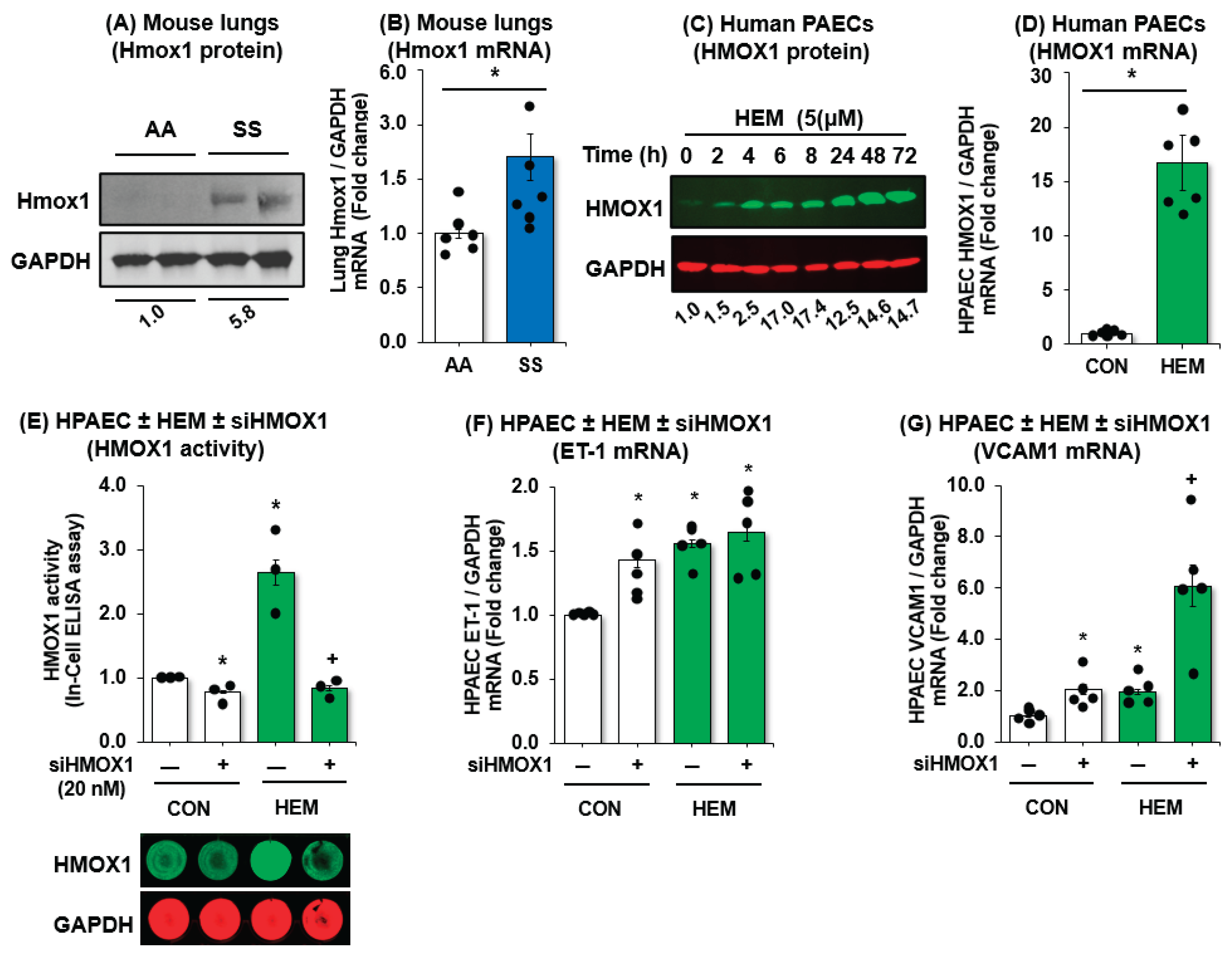
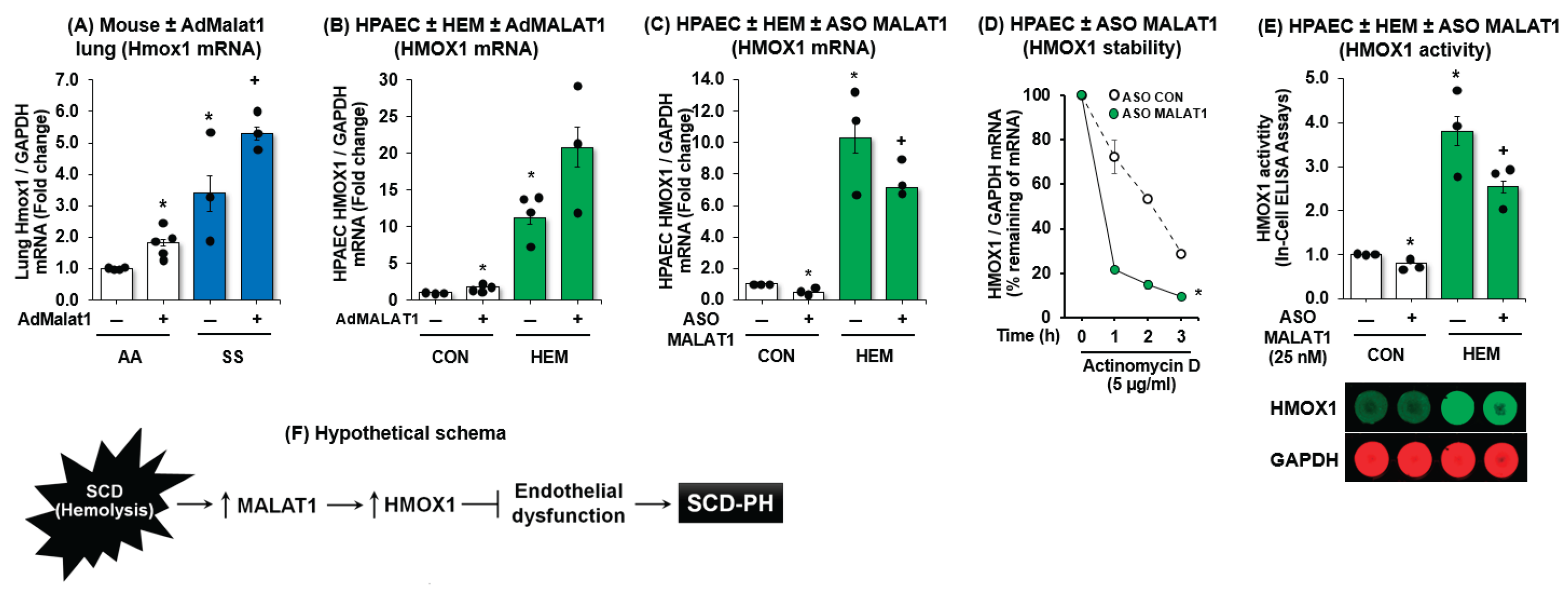
Figure 5.
MALAT1 regulates HMOX1 expression and activity. (A) AA and SS mice were infected with AdGFP or AdMALAT1 and 2 weeks later, whole lung homogenates were collected and subjected to qRT-PCR analysis. Mean HMOX1 levels examined as fold-change vs. AA/AdMALAT1(-). *p<0.05 vs. AA/AdMALAT1(-), +p<0.05 vs. SS/AdMALAT1(-), n=3-5. (B) HPAECs were treated with scrambled AdGFP or AdMALAT1 particles (5 MOI) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC HMOX1 levels were measured with qRT-PCR. All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/AdMALAT1(-). *p<0.05 vs. CON/AdMALAT1(-), +p<0.05 vs. HEM/AdMALAT1(-), n=3-4. (C,D) HPAECs were treated with scrambled anti-sense oligonucleotide (ASO CON) or ASO MALAT1 (25 nM) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC HMOX1 levels were measured with qRT-PCR (C) or HMOX1 activity using In-Cell ELISA assay (D). All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/AdMALAT1(-). *p<0.05 vs. CON/AdMALAT1(-), +p<0.05 vs. HEM/AdMALAT1(-), n=3-4. (E) To inhibit de novo HMOX1 mRNA synthesis, selected cells were treated with 5 μg/ml actinomycin D (in a time-dependent manner). HMOX1 mRNA half-life was determined by comparing to the mRNA level before adding actinomycin D. Error bars represent mean ± SE, n=3, *p<0.05 vs. scrambled controls (SCR). (F) A hypothetical schema defining the role of MALAT1 in SCD-PH pathogenesis.
Figure 5.
MALAT1 regulates HMOX1 expression and activity. (A) AA and SS mice were infected with AdGFP or AdMALAT1 and 2 weeks later, whole lung homogenates were collected and subjected to qRT-PCR analysis. Mean HMOX1 levels examined as fold-change vs. AA/AdMALAT1(-). *p<0.05 vs. AA/AdMALAT1(-), +p<0.05 vs. SS/AdMALAT1(-), n=3-5. (B) HPAECs were treated with scrambled AdGFP or AdMALAT1 particles (5 MOI) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC HMOX1 levels were measured with qRT-PCR. All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/AdMALAT1(-). *p<0.05 vs. CON/AdMALAT1(-), +p<0.05 vs. HEM/AdMALAT1(-), n=3-4. (C,D) HPAECs were treated with scrambled anti-sense oligonucleotide (ASO CON) or ASO MALAT1 (25 nM) for 6 h then treated with dimethyl sulfoxide vehicle (CON) or hemin (HEM, 5 µM) for an additional 72 h. Mean HPAEC HMOX1 levels were measured with qRT-PCR (C) or HMOX1 activity using In-Cell ELISA assay (D). All bars represent the mean mRNA levels relative to GAPDH ± SE expressed as fold-change vs. CON/AdMALAT1(-). *p<0.05 vs. CON/AdMALAT1(-), +p<0.05 vs. HEM/AdMALAT1(-), n=3-4. (E) To inhibit de novo HMOX1 mRNA synthesis, selected cells were treated with 5 μg/ml actinomycin D (in a time-dependent manner). HMOX1 mRNA half-life was determined by comparing to the mRNA level before adding actinomycin D. Error bars represent mean ± SE, n=3, *p<0.05 vs. scrambled controls (SCR). (F) A hypothetical schema defining the role of MALAT1 in SCD-PH pathogenesis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



