
Submitted:
03 December 2025
Posted:
04 December 2025
You are already at the latest version
Abstract
Major depressive disorder (MDD) remains a leading cause of disability worldwide, with conventional antidepressants offering incomplete and often transient relief. Mounting ev-idence highlights disturbances in tryptophan (Trp) metabolism as a key biological axis linking inflammation, neuroplasticity, and mood regulation. Plant-derived compounds that modulate this pathway, including 5-hydroxytryptophan, isoflavones, berberine, and polyphenols, have emerged as promising candidates for integrative treatment strategies. Yet, despite encouraging preclinical and clinical findings, knowledge gaps persist re-garding long-term efficacy, mechanistic specificity, and standardized therapeutic proto-cols. This narrative review explores how plant-derived Trp modulators influence central and peripheral mechanisms relevant to depression, from serotonergic synthesis and kynurenine shunting to gut–brain–immune interactions. Evidence from animal models and randomized clinical trials is critically synthesized, with particular attention to out-comes on mood stabilization, anxiety reduction, cognitive function, and sleep regulation. Special emphasis is placed on translational potential, methodological limitations, and the need for harmonized research frameworks. Here we highlight that phytochemical inter-ventions represent a mechanistically informed and biocompatible strategy for advancing depression management. By bridging neurobiology and clinical psychiatry, these insights may pave the way for next-generation therapeutics that integrate dietary, microbio-ta-targeted, and anti-inflammatory approaches. Broader application of this research could ultimately refine personalized psychiatry, expand therapeutic horizons, and contribute to global mental health resilience.
Keywords:
major depressive disorder
; tryptophan metabolism
; kynurenine pathway
; phytochemicals
; dietary supplements
; 5-hydroxytryptophan
; gut–brain axis
; neuroinflammation
; plant extracts
; precision nutrition
1. Introduction
MDD surpasses most chronic diseases in human toll, ranking consistently among the top causes of years lived with disability [1,2]. More than 300 million people currently experience the condition, and its relapsing course relentlessly erodes well-being, productivity, and social cohesion [2,3]. Annual indirect and direct costs already exceed one trillion United States dollars, weakening national economies while stigma quietly erodes family networks, impairs educational attainment, and narrows life opportunities [4,5]. Conventional monoamine-reuptake-based antidepressants, though invaluable, leave a sizeable fraction of patients with partial or transient relief, highlighting an unmet therapeutic horizon [2]. Recent neurobiological work has illuminated Trp metabolism as a convergent node linking serotonergic tone, KYN-driven neuroinflammation, and gut-brain communication [6,7]. Consequently, nutritional psychiatry is moving center stage, exploring dietary enrichment, phytochemical extracts, and complex plant polysaccharides that can steer Trp flux toward antidepressant pathways [3,7]. These multitarget, biocompatible candidates warrant rapid, translational exploration to transform MDD management [3,8].
5-HT occupies the fulcrum of affective neurocircuitry, yet its synthesis depends on a single dietary precursor, Trp [9,10]. When circulating 5-HT dwindles—whether through genetic polymorphisms, persistent inflammation, or inadequate intake—clinical and pre-clinical data reveal cascades of anhedonia, low mood, and suicidality [9,10]. Supplementation with Trp and, more recently, nanozyme-accelerated biosynthesis swiftly restores synaptic stores, underscoring causality [11,12]. Because no alternative substrate can replenish this indoleamine, strategies that enrich Trp availability or steer its fate toward the serotonergic branch merit priority; they simultaneously quell microglial inflammation and normalize behavior in rodents and humans [10,12]. The problem deepens when proinflammatory cytokines activate IDOs, shunting Trp down the KYN track, exhausting serotonin, and generating neurotoxic QA [10,13]. Meta-analytic biomarker work links this detour to melancholic features, treatment resistance, and suicidal intent, framing KYN dysregulation as a mechanistic bottleneck [9,10]. Encouragingly, plant-derived agents—ginsenoside Rk3, Pulsatilla saponins, Hypericum extracts, diverse polysaccharides, and indole alkaloids—re-calibrate this junction [12,14,15]. By activating tryptophan hydroxylase, repressing IDO1, re-shaping gut microbiota, and enriching anti-inflammatory indoles, they restore serotonergic dominance and offer a multimodal, biocompatible blueprint for next-generation antidepressant therapy [10,14].
Selective 5-HT and 5-HT-norepinephrine reuptake inhibitors still headline pharmacological guidelines for MDD, yet their benefit remains modest [16,17]. Clinical relief often arrives only after a fortnight—a critical delay for individuals experiencing suicidal ideation or severe impairment [16,18]. Adverse events accumulate in the interim: sexual dysfunction that may persist beyond discontinuation; progressive weight gain and hyponatremia threaten metabolic and electrolyte balance; emotional blunting, apathy, and cognitive fog sap motivation and adherence, especially in adolescents and older adults vulnerable to withdrawal syndromes [16,18,19,20]. Some patients become treatment-resistant after adequate trials, and relapse still strikes a part of the apparent responders within a year, each episode deepening psychosocial impairment and economic burden [18,19,21]. These intertwined shortcomings have energized the search for nutraceutical, phytochemical, and mind-body interventions [18,22,23,24]. Omega-3 fatty acids, flavonoid-rich extracts, acupuncture, mindfulness practice, and guided psilocybin sessions already display encouraging efficacy with fewer liabilities, earning cautious endorsement in emerging guidelines and stimulating expansive translational research [22,23,24,25]. Together, these trends justify systematic appraisal of plant-derived Trp as the next therapeutic frontier [18,22,26]. Figure 1 illustrates an overview of MDD burden and plant-derived modulation of Trp pathways.
Plant-derived modulators of Trp metabolism have moved from folklore to the laboratory, carrying a mechanistic sophistication once reserved for synthetics [27,28,29]. Oligosaccharides from Morinda officinalis boost gut 5-hydroxy-L-tryptophan, elevating brain 5-HT and reversing stress-induced anhedonia [14,27,28]. Ginsenoside Rk3 and Pulsatilla saponins suppress IDOs, redirect KYN flux, and calm neuroinflammation, while Hypericum fractions, probiotic indoles, and alkaloid derivatives further polish gut-brain signaling with striking behavioral gains [28,29,30]. These findings echo an ethnopharmacological legacy: lavender, turmeric, bacopa, the Chinese formula Xiaoyao-san, and Ayurvedic rasayanas have eased melancholy for centuries, with numerous mood-modulating plants catalogued across ancient pharmacopoeias [27,28,30]. Their renaissance coincides with nutritional psychiatry, a field treating diet as neuronal software by integrating microbiome science, polyphenol chemistry, and amino-acid neuroscience [14,27,28]. Some global guidelines now acknowledge nutraceuticals and phytoceuticals, and bibliometric analyses chart exponential growth in phytotherapeutic trials [27,28,29]. Moreover, xylopic acid, sclareol-quercetin blends, capsaicin, and Chaihu-Shugan-San enhance SSRI efficacy while tempering metabolic and cognitive liabilities, underscoring the promise of safer, synergistic, multicomponent antidepressant strategies [27,28,29].
The primary objective of this review is to systematically examine the efficacy and neurobiological mechanisms of plant-derived Trp in the treatment of depression. Because plant-derived Trp compounds act across multiple interacting pathways, a comprehensive synthesis requires revisiting core mechanisms such as TPH activity, KYN metabolism, cytokine signaling, and gut microbiota function, since each reappears in distinct clinical and translational contexts. By addressing both experimental and clinical evidence, the review seeks to clarify how interventions targeting Trp metabolism may influence mood regulation and therapeutic outcomes [31,32,33]. A central aim is to bridge the gap between fundamental neurobiology and clinical psychiatry, offering an integrated perspective that connects molecular mechanisms with patient-centered applications. Specifically, the review focuses on delineating the pathways through which Trp and its metabolites modulate the serotonergic and KYN systems, evaluating preclinical and clinical trial data, and identifying limitations that must be overcome to enhance translational relevance. Attention is given to unresolved issues in bioavailability, pharmacokinetics, and long-term safety. Ultimately, this review highlights future therapeutic prospects and emphasizes the importance of integrating mechanistic insights with clinical practice to guide precision-based strategies for depression management.
This review carries significant implications for psychiatry, nutritional sciences, and integrative medicine by advancing understanding of plant-derived Trp as a therapeutic option for depression. Its contributions extend beyond cataloguing current evidence, offering a critical synthesis that highlights both mechanistic insights and clinical applications. By evaluating how dietary and phytochemical interventions influence Trp metabolism and related neurobiological pathways, the review provides an innovative framework with direct translational relevance. The discussion emphasizes how such approaches may complement existing pharmacological treatments, reduce the burden of side effects, and support individualized, nutrition-based strategies in mental health care. Notably, the review underscores the potential of integrating dietary science with neuropsychiatric research to broaden treatment paradigms and improve patient outcomes. Here, we highlight the necessity of sustained interdisciplinary collaboration, uniting psychiatry, molecular biology, nutrition, and clinical practice. Only through such efforts can the full therapeutic potential of Trp-related interventions be realized in depression management.
2. Neurobiology of Depression: Beyond Serotonin (5-HT)
For much of the twentieth century, textbooks portrayed depression as a straightforward consequence of diminished synaptic 5-HT, a view seemingly vindicated by the success of selective 5-HT reuptake inhibitors and reinforced by simplified biochemical schematics [34,35,36]. Accumulating evidence now unpicks that narrative. An umbrella analysis fails to locate a reliable serotonergic deficit signature in MDD. At the same time, parallel studies reveal stress-evoked glutamatergic drift at AMPA receptors, a shift that disrupts excitatory balance and long-term potentiation [34,37,38,39]. Cytokine surges further interweave immune and neural realms, diverting Trp toward KYN metabolites with neurotoxic or neuroprotective potential, and tuning mitochondrial resilience in vulnerable circuits [40,41,42]. Animal paradigms of chronic unpredictable stress echo these molecular fingerprints, exposing dendritic retraction and synaptic pruning that compromise mood-regulating networks [34,41,42]. Against this backdrop, neuroplasticity models integrate receptor interactions, neurotrophic signaling, and gliovascular dynamics, offering a more comprehensive representation of the disorder’s heterogeneity [34,35,40]. Such multi-layered models do more than widen mechanistic horizons; they steer translational efforts toward modulators of plasticity, inflammation, and metabolism, promising interventions that transcend the monoaminergic lens and address the complex biology underlying depressive illness [40,43,44,45].
2.1. Microglia Activation and Neuroinflammation
Microglia constantly monitor the brain. They prune synapses and release trophic factors that help maintain metabolic and neural networks [46,47,48]. Stress shifts them into a reactive mode. In this state, they proliferate more rapidly, alter their metabolism, and release inflammatory signals that weaken excitatory synapses [46,47,49]. Glucocorticoids and catecholamines coursing through HPA and sympathetic pathways reinforce this shift, entrenching microglia in maladaptive, inflammatory phenotypes [46,50,51,52]. A spectrum of reactive states emerges: some microglia secrete cytokines and reactive metabolites that fracture neuronal integrity, while others attempt repair through anti-inflammatory mediators [48,53,54]. Early-life immune challenges further dysregulate microglial surveillance, triggering excessive dendritic spine engulfment and predisposing adolescents to mood disturbances [55,56,57]. In adulthood, sustained IL-6 release drives hippocampal astrocyte apoptosis, and prostaglandin signaling in striatal regions amplifies negative affect [49,58,59]. Crucially, pharmacological or genetic dampening of microglial activation reduces cytokine burden and restores synaptic function, alleviating depressive-like behaviors in animal models [48,49,58]. Such findings underscore microglia’s dual capacity for neuroprotection and neurotoxicity, and highlight inflammatory modulation as a promising axis for next-generation antidepressant development [46,48,49].
A consistent pattern emerges across studies. IL-1β, IL-6, TNF-α, and IFN-γ tend to rise with symptom severity and often predict relapse. Stress increases these cytokines via NLRP3 activation, while epigenetic regulators such as EZH2 further amplify them. Together, they build an inflammatory loop that shapes mood and cognition [60,61,62]. Meta-analytic syntheses, large observational cohorts, and translational models consistently show that concentrations of these cytokines rise in parallel with symptom load, forecast relapse, and differentiate treatment-resistant cases from treatment-responsive patients [63,64,65]. Stress priming activates NLRP3 inflammasomes, unleashing bursts of IL-1β that propagate glial crosstalk, while epigenetic regulators such as EZH2 intensify IL-6 and TNF-α transcription, accelerating synaptic attrition and anhedonia [66,67,68]. Longitudinal work in adolescents demonstrates that elevated baseline TNF-α or IL-6 anticipates chronic anhedonia and white-matter dysconnectivity years later, underscoring a trajectory of immune-driven neuroprogression [69,70,71]. Complementary primate and lipopolysaccharide paradigms replicate the cytokine surge and its behavioral sequelae, bolstering causality [72,73,74]. Crucially, immunomodulatory interventions can reverse this profile: ketamine lowers IL-6 and TNF-α in treatment-resistant depression in tandem with rapid affective recovery, and targeted anti-cytokine therapies normalize peripheral markers while lifting mood in case studies and pilot trials [75,76,77]. Together, these findings portray a feed-forward inflammatory circuit that not only marks depressive severity and chronicity but also delineates mechanistic targets for next-generation precision antidepressants, with potential for biomarker-guided stratification in clinical practice worldwide [72,73,78]. These inflammatory shifts directly reshape the neurogenic niche, setting the stage for changes in hippocampal plasticity.
2.2. Neurogenesis Impairment and Hippocampal Atrophy in Depressive Pathology
The hippocampus continually produces new granule cells. These cells integrate into memory and emotion circuits, helping maintain cognitive flexibility and emotional stability. Rodent, primate, and human proxy experiments reveal that boosting this renewal before stress inoculates against anhedonia and cognitive drift, whereas blocking it magnifies vulnerability and depressive phenotypes [79,80,81]. Mitochondrial fitness, autophagic clearance, and neuroimmune balance calibrate neurogenic yield, linking cellular housekeeping to psychological health and nominating the neurogenic niche as a tractable antidepressant target [82,83,84]. Chronic stress destabilizes this balance. Elevated corticosterone primes microglia and activates NF-κB. As a result, IL-1β, IL-6, TNF-α, and IFN-γ accumulate in the hippocampus. Because microglia are the primary source of many of these cytokines, their activation acts as a bottleneck for neurogenesis, constraining proliferation and survival of newborn neurons. These cytokines stop progenitors from dividing. They also divert BDNF signaling through CDK5-phosphorylated huntingtin. At the same time, astrocytes provide less VEGF, and neural precursors are pushed toward apoptosis [85,86,87]. Glial activation amplifies oxidative damage, ADAM17/TNF-α signaling, and aberrant pruning, cementing a neurogenic standstill that melatonin or ginsenoside Rg1 can dismantle [88,89,90]. Structural imaging corroborates the cellular narrative: meta-analyses and cross-diagnostic cohorts consistently report bilateral, often left-dominant, hippocampal atrophy in depression [87,90,91]. Seven-Tesla datasets pinpoint shrinkage to dentate gyrus and tail subfields, especially in treatment-resistant illness [89,91,92]. Interventions that rekindle plasticity, including electroconvulsive therapy and aerobic exercise, partially restore volume, implying that suppressed neurogenesis and dendritic retraction are reversible contributors to mood recovery [89,93,94]. Such structural reversibility converges with behavioral improvements, reinforcing adult neurogenesis as both a biomarker and a mediator of durable resilience [95,96].
Chronic neuroinflammation progressively dismantles the hippocampal neurogenic niche. Activated microglia, astrocytes, and infiltrating immune cells inundate progenitor zones with IL-1β, caspases, reactive oxygen species, and other cytotoxic mediators, stalling cell-cycle progression and truncating dendritic maturation [97,98,99]. Sustained IL-10 exposure or traumatic injury reprograms microglia toward a phagocytic, synapse-stripping phenotype that disrupts neuron–glia crosstalk and shortens the lifespan of newborn neurons [100,101,102]. Intrinsic genetic vulnerabilities intensify the problem: loss of the transcription factor Tcf4 unleashes latent inflammatory pathways inside neural stem cells, extinguishing their regenerative capacity [97,98]. Systemic infections or chronic gut inflammation widen this suppression, propagating cytokine waves that reverberate into cognitive decline and depressive affect [103,104]. Compounding these injuries, elevations in IL-6 and TNF-α draw down BDNF, a molecule essential for synaptic refinement and mood stability, thereby tightening the grip of inflammation on neuroplasticity [105,106,107]. Clinical imaging and behavioral studies mirror these molecular events: higher cytokine loads predict smaller hippocampi, poorer memory, and deeper anhedonia, while agents that either quiet neuroimmune signaling or re-energize neurogenesis—such as Akebia saponin D, liraglutide, naringenin, or KYN-pathway modulators—restore plasticity and relieve symptoms [105,108,109,110,111].
2.3. The Link Between Inflammation, Tryptophan (Trp) Metabolism, and Depressive Symptoms
Innate and adaptive immune signals divert Trp away from serotonergic fate, engaging IDOs and TDO that accelerate the KYN cascade of neuroactive compounds [112,113]. IFN-γ, IL-6, and TNF-α boost these enzymes across the gut and brain, shrinking 5-HT stores while raising KYN and its downstream quinolinate products [113,114]. Chronic stress or intestinal inflammation magnifies the shift, elevating KYN-to-5-HT ratios in rodents and paralleling anhedonia in patients [113,115]. This metabolic rerouting, therefore, links immune activation to depressed mood and cognitive decline [113,115,116,117] (Figure 2). Yet the pathway’s end stage proves most damaging. QA overstimulates NMDA receptors, drives calcium overload, and sparks oxidative stress [113,114,118,119,120]. Traumatic brain injury or systemic inflammation upregulates KYN monooxygenase, intensifying quinolinate build-up, halting progenitor proliferation, and eroding hippocampal circuits [112,113,118]. Microplastic exposure yields similar DNA fragmentation and microglial activation [112,113,121]. Therapeutic blockade with ketamine or selective enzyme inhibitors reduces quinolinate levels, restores neurogenesis, and rescues behavior [112,113,114]. Reviews across neurodegenerative contexts confirm that balancing toxic and protective KYNs is critical for hippocampal integrity and mood regulation [112,113,115,122]. Collectively, these findings reposition the KYN axis as both a biomarker and a mechanistic driver, inviting precision anti-inflammatory or metabolic interventions that could recalibrate Trp flux, preserve plasticity, and alleviate depressive illness.
Chronic immune activation diverts Trp metabolism by up-regulating IDOs and TDO, drawing substrate from 5-HT synthesis and inflating production of neuroactive KYNs such as QA [113,123,124]. Quinolinic excess overstimulates NMDA receptors, kindles oxidative stress, and arrests hippocampal neurogenesis, thereby reducing BDNF, pruning synapses, and disturbing gut–brain communication essential for mood and memory [123,125]. These pathophysiological threads recast depression as a metabolic-immune-plasticity disorder that demands therapies surpassing monoaminergic repair [113,123,124]. Evidence now endorses blended strategies that curb inflammation, recalibrate microbiota signaling, temper glutamatergic drive, and reignite neurogenesis [126,127]. Phytochemicals illustrate this integrative ethos: polyphenols, soy isoflavones, berberine, albiflorin, Pulsatilla saponins, and naringenin jointly silence microglial cytokine release, inhibit IDO1, activate tryptophan hydroxylase, and foster survival of newborn neurons, collectively improving affective and cognitive outcomes in stress-resistant models [128,129,130]. Such multi-layered modulation repairs molecular circuitry while offering a blueprint for precision nutrition and pharmacology that mirrors depression’s multifactorial biology, promising durable relief in clinical practice [131,132,133].
3. Tryptophan (Trp) Metabolism: Central Pathways and Peripheral Influences
This review uses the term plant-derived Trp compounds to refer to plant Trp sources, purified plant Trp-based supplements, and phytochemicals that modulate Trp metabolism. Shortened forms such as 'Trp' or 'phytochemicals' are used only when the context is specific.
Trp metabolism lies at the heart of neuropsychiatric research, serving as a biochemical crossroad between neurotransmission, immune regulation, and circadian control [134,135]. Clinical depression emerges from disruptions in these same pathways because even subtle shifts in serotonin or kynurenine flux can alter neuroplasticity, inflammatory tone, and stress responsiveness. Among its metabolic fates, the 5-HT and KYN pathways represent fundamental routes with profound implications for mood and cognition [136,137]. The 5-HT branch underpins neurotransmitter synthesis and neuroplasticity, while the KYN branch generates a spectrum of neuroactive metabolites that shape neuroinflammation and excitotoxicity [138,139]. Dysregulation of this delicate balance has been repeatedly implicated in depression, schizophrenia, and neurodegenerative disorders [13,140]. This section introduces these dual metabolic routes, emphasizing their molecular complexity, clinical relevance, and potential as therapeutic targets for precision psychiatry.
3.1. Serotonin (5HT) and Kynurenine (KYN) Metabolic Pathways: Fundamental Metabolic Routes
The 5-HT pathway begins when Trp is hydroxylated by TPH1 and TPH2, producing 5-HTP, which is then decarboxylated to form 5-HT [141,142]. This neurotransmitter engages multiple receptor families, including 5-HT1A, 5-HT2A, 5-HT4, and 5-HT6, each of which exerts distinct influences on mood regulation, neurogenesis, and synaptic plasticity [134,141]. Antidepressants such as SSRIs not only increase synaptic 5-HT but also enhance structural remodeling and circuit flexibility, shifting the focus from a deficit model to a neuroplasticity framework [142,143]. Receptor-receptor interactions, like 5-HT2A–TrkB heterodimerization, and receptor-specific mechanisms, such as biased 5-HT1A signaling or 5-HT4-mediated memory modulation, highlight the pathway’s molecular sophistication [13,143]. Psychedelic compounds further reveal how 5-HT2A activation may promote cortical plasticity, though 5-HT itself may not be the primary ligand driving such effects [134,142]. Yet, pressing gaps persist. The precise endogenous mechanisms underlying receptor-driven plasticity remain unclear, while phase-specific roles of 5-HT in declarative memory are insufficiently mapped. Translational barriers hinder the movement of receptor-targeted and psychedelic-based therapies into clinical use, particularly regarding long-term safety. Moreover, the interplay of diet, stress, and genetic variability complicates predictions of therapeutic response [134,143]. Clarifying these complexities is critical for developing precise, individualized treatments for mood and cognitive disorders. Figure 3 illustrates the multiple biological effects associated with melatonin.
The KYN metabolic pathway represents the primary route of Trp degradation, initiated by the rate-limiting enzymes IDOs and TDO, leading to the production of KYN [144,145]. This intermediate can be further metabolized into neuroactive derivatives with contrasting effects: KYNA, an NMDA receptor antagonist with neuroprotective properties, and QA, an NMDA receptor agonist with excitotoxic and pro-inflammatory actions [146,147]. Additional metabolites such as 3-HK contribute to oxidative stress, linking the pathway to neurodegenerative and psychiatric disorders [144,146]. Meta-analyses and experimental studies consistently show a pathological shift toward neurotoxic metabolites in depression, schizophrenia, Alzheimer’s disease, and neurodegeneration [13,148]. Yet significant challenges persist. The dichotomy of KYNA as purely protective and QA as strictly harmful is now recognized as overly simplistic, since their effects vary with concentration, brain region, and disease context [149,150]. Furthermore, the spatial distribution and regulation of key enzymes such as KMO, KATs, and QPRT remain insufficiently characterized in both central and peripheral systems [151,152]. Peripheral metabolite measures often fail to accurately represent brain activity, limiting their use as biomarkers [152,153]. Finally, although enzyme inhibitors and analogues are under clinical investigation, their long-term efficacy and safety are far from established [151,154].
3.2. Oxidative Stress and Inflammation: Drivers of Kynurenine (KYN) Pathway Activation
Trp is an essential amino acid indispensable for neurochemical homeostasis, mood regulation, and cognitive integrity [13,123,135]. While a minor fraction enters the serotonin pathway to support neurotransmission and neuroplasticity, the vast majority is shunted into the KYN pathway, generating metabolites with neuroactive and immunomodulatory properties [10,113,135]. Cytokine-driven diversion of Trp toward the KYN branch, described earlier, amplifies oxidative stress and metabolic imbalance, a shift consistently observed in depression, schizophrenia, and neurodegenerative disorders [13,113,155]. Accumulation of neurotoxic metabolites, such as QA, exacerbates excitotoxicity and oxidative stress, whereas KYNA may provide neuroprotection in a context-dependent manner [156,157,158]. Despite clear links between immune activation, oxidative stress, and altered Trp metabolism, significant questions remain unresolved [13,113,156]. The molecular crosstalk among cytokine signaling, mitochondrial dysfunction, and metabolic reprogramming remains unclear [13,156,159]. Similarly, the role of gut microbiota and platelets as peripheral modulators of Trp fate is incompletely defined [109,115,136,160]. Translational gaps persist in developing reliable biomarkers and effective therapeutic strategies, particularly antioxidants, pathway modulators, and microbiota-targeted interventions [13,156,159]. Addressing these challenges will be crucial for precision approaches to neuropsychiatric and neurodegenerative diseases.
IDOs and TDO catalyze the first, rate-limiting step of the KYN pathway, thereby functioning as pivotal gatekeepers of Trp metabolism [161,162,163]. Under conditions of inflammation and oxidative stress, these enzymes are strongly upregulated: TDO, primarily expressed in the liver and regulated by systemic Trp levels and glucocorticoids, and IDO, widely induced in immune cells by cytokines such as IFN-γ and TNF-α [162,164,165]. Their activation diverts Trp away from 5-HT synthesis and channels it toward KYN, which, in turn, generates metabolites such as QA, with excitotoxic and pro-inflammatory effects, and KYNA, with context-dependent neuroprotection [162,163,164]. Beyond enzymatic activity, KYN itself acts as an AhR ligand, modulating immune tolerance and tumor progression [161,166,167]. Despite these advances, substantial gaps remain. The tissue-specific regulation of IDO and TDO under stress conditions is not fully mapped, and their non-enzymatic roles in shaping the immune microenvironment remain underexplored [162,163,164]. Clinical trials with IDO inhibitors have yielded disappointing results, highlighting the need for dual IDO/TDO inhibition or combined targeting of downstream receptors [163,166,168]. Moreover, reliable biomarkers of enzyme activity and patient stratification strategies are lacking, while the dynamic regulation of heme incorporation and post-translational modifications introduces further complexity [161,162,163].
Elevated activity of the KYN pathway, particularly the accumulation of QA, has been strongly associated with depressive symptoms, impaired neurogenesis, and deficits in cognitive domains such as memory and executive function [169,170,171]. Neuroimaging and postmortem studies reveal that increased KYN metabolite ratios correlate with structural and functional brain abnormalities in depression. At the same time, animal models show that IDO-mediated pathway activation drives both neuroinflammation and cognitive decline [170,171,172]. Evidence also suggests that reduced kynurenic acid, a putative neuroprotective metabolite, exacerbates cognitive impairment, especially in mood disorders [148,173,174]. These findings have stimulated interest in KYN-targeted therapies, ranging from small molecules to microbiota-based interventions [115,136,150,175,176]. Yet, significant questions remain unresolved. Central and peripheral measures of pathway activity often diverge, complicating biomarker development [148,174,177,178]. The causal links between specific metabolites and clinical symptoms require more mechanistic studies [124,169,172]. Translational efforts remain limited, with few robust clinical trials validating efficacy [148,175,177]. Furthermore, the role of gut microbiota in modulating KYN metabolism and cognitive outcomes is only beginning to be understood [136,179,180]. Longitudinal studies are essential for mapping disease trajectories and clarifying whether metabolite alterations differ across depression subtypes and cognitive phenotypes [148,172,177].
3.3. Gut-Brain Axis and Intestinal Microbiota: Peripheral Modulators of Tryptophan (Trp) Metabolism
The gut-brain axis functions as a bidirectional regulator of Trp metabolism linking the gastrointestinal tract with the central nervous system through neural, immune, endocrine, and metabolic pathways [181,182,183]. This system orchestrates critical processes that regulate neuroinflammation, energy metabolism, and psychiatric outcomes, with mounting evidence implicating gut microbiota and their metabolites as central mediators [160,184]. Dysbiosis has been consistently associated with depression, anxiety, schizophrenia, and neurodegenerative disorders, primarily through altered signaling across the vagus nerve, HPA axis, and immune-inflammatory cascades [185,186]. At the same time, microbial metabolites such as short-chain fatty acids and Trp derivatives modulate neurotransmission and glial function, further linking gut health to brain plasticity and cognition [181,187]. Yet, despite compelling evidence, substantial gaps remain. The precise molecular and cellular mechanisms underlying gut-brain interactions are incompletely defined, particularly regarding causal links [188,189]. Translation of probiotics, prebiotics, symbiotics, and FMT into reliable therapeutic strategies is hindered by inconsistent clinical results [183,188,190]. Biomarker development for diagnosis and treatment monitoring is still in its infancy, while most studies are restricted to narrow cohorts, limiting generalizability [183,188,191]. Longitudinal and mechanistic trials are urgently needed to clarify causality and optimize intervention strategies in psychiatric and neurodegenerative disorders [181,188,191]. Figure 4 summarizes the dietary Trp metabolism by gut microbiota and its systemic effects.
As detailed earlier, microbial regulation of Trp availability shapes central signaling and mood through multiple converging pathways [136,160,192]. By modulating local inflammation, they alter the activity of enzymes that govern the diversion of Trp into the 5-HT and KYN branches, thereby influencing neurotransmission, neuroplasticity, and immune signaling [136,193,194]. Microbial metabolites also act directly: some species enhance 5-HT synthesis in the gut, while others promote KYN production or generate indole derivatives that engage the aryl hydrocarbon receptor to regulate neuroinflammation [195,196]. These interactions extend beyond the periphery, with microbial-driven metabolic shifts linked to depression, anxiety, and cognitive dysfunction in both preclinical models and clinical cohorts [136,160,192,194]. Yet, despite compelling evidence, essential uncertainties remain. The identity of key microbial species responsible for shaping Trp metabolism is incompletely defined, and the molecular mechanisms that orchestrate the balance between the 5-HT, KYN, and indole pathways remain unclear [194,197,198]. Translational progress is limited, as most findings derive from animal studies rather than large-scale human trials [194,199,200]. Therapeutic strategies such as probiotics, symbiotics, and dietary modulation hold promise, but their efficacy, safety, and mechanistic specificity need systematic evaluation [200,201,202]. Finally, reliable biomarkers to monitor microbiota-driven Trp metabolism and predict psychiatric outcomes are lacking, hindering the development of personalized interventions [194,197,203]. Figure 5 summarizes the Trp–indole metabolism and its pathological consequences.
Peripheral metabolism does not operate in isolation. The gut microbiota strongly shapes the amount of Trp available for serotonin synthesis or diversion into kynurenine metabolites, and these shifts can amplify or mitigate depressive risk. Emerging preclinical and clinical evidence suggests that targeting gut microbiota through probiotics, prebiotics, dietary phytochemicals, or even FMT can restore Trp balance, reduce neuroinflammation, and alleviate depressive symptoms [191,194,204]. Probiotics such as Lactobacillus and Bifidobacterium strains have been shown to enhance 5-HT availability, while prebiotics and phytochemicals promote beneficial microbial shifts that dampen KYN-driven neurotoxicity [205,206,207]. In animal models, microbiota modulation improves stress-induced depressive behaviors by redirecting Trp metabolism toward 5-HT rather than neurotoxic KYN derivatives, findings now echoed in early human trials reporting improved mood outcomes [205,206,208]. Nevertheless, significant gaps remain. Mechanistic clarity is limited, particularly regarding the molecular pathways by which specific microbial taxa regulate 5-HT and KYN branches [191,192,194]. Most clinical studies are small, short-term, and heterogeneous in design, making premature conclusions about efficacy and safety [208,209,210]. Inter-individual variability in microbiome composition complicates the development of standardized therapies, while long-term durability of benefits remains untested [191,194,209]. Finally, reliable microbial and metabolic biomarkers to guide patient stratification and therapeutic monitoring are lacking [191,194,209]. Addressing these gaps is essential for translating microbiota-based strategies into personalized, evidence-driven interventions for depression and related neuropsychiatric disorders. Figure 6 summarizes dual pathways of Trp metabolism and their neuropsychiatric implications.
4. Integrative Therapeutic Approaches as Modulators of Tryptophan (Trp) Metabolism with a Focus on Plant-Derived Dietary Strategies
Plant-derived compounds offer a versatile toolkit for modulating Trp metabolism at multiple biological levels, from enzyme activity to gut–brain signaling [31,134,211]. This section introduces how phytochemicals can rebalance flux between 5-HT and KYN pathways, thereby influencing mood, sleep, cognition, and neuroinflammation [31,134,212]. We integrate mechanistic insights with clinical observations, highlighting candidates such as 5-HTP, isoflavones, berberine, polyphenols, and Trp-rich dietary proteins [31,212,213]. Particular attention is given to bioavailability, pharmacokinetics, and microbiome interactions that shape therapeutic response [31,134,214]. By linking molecular targets to patient outcomes, this section frames an evidence-informed path toward translational strategies. It sets the stage for a critical appraisal of clinical trials in the subsections that follow.
4.1. Clinical Evidence for Plant-Derived Modulators of Tryptophan (Trp) Metabolism
Most available findings remain at an early stage. Many trials are small, short, or narrow in scope, so any mood or sleep improvements should be viewed as exploratory signals rather than evidence of clinical readiness [31,138,212]. Although several RCTs report changes in mood-related outcomes, these effects are usually modest, vary across studies, and remain unconfirmed in larger clinical samples [31,215,216]. Other interventions, such as Trp and magnesium-enriched Mediterranean diets tested in women with fibromyalgia, have explored the combined impact on sleep quality and anxiety. However, results remain mixed due to small cohorts and gender-restricted samples [217,218,219]. Broader inclusion criteria have encompassed patients with MDD or bulimia, while preclinical successes with compounds like albiflorin highlight the translational potential awaiting human validation [219,220,221]. Standard outcome measures include psychiatric rating scales, sleep quality indices, and neurochemical assays, allowing assessment of both symptomatic relief and mechanistic engagement [220,222,223].
The following table (Table 1) highlights relevant papers on dietary and non-dietary modulation of Trp in depression and related disorders, summarizing their focus, inclusion criteria, outcomes, and patient populations.
Randomized controlled trials investigating phytochemicals with modulatory effects on Trp metabolism have yielded encouraging, though heterogeneous, findings across domains of mood, anxiety, sleep, and cognition. Rosemary extract demonstrated significant reductions in anxiety and depressive symptoms among university students, along with measurable improvements in sleep quality and memory, suggesting a combined effect on mood regulation and cognitive processing [242,243,244]. Lavender oil (Silexan) consistently reduced generalized anxiety and restlessness while simultaneously enhancing sleep continuity, with benefits comparable to first-line anxiolytics but without sedative liabilities [245,246,247]. Trials with lemon balm further highlighted synergistic effects, showing improved psychological well-being, stress resilience, and sleep indices in adults with moderate emotional distress [245,248,249]. Beyond these botanicals, isoflavones and polyphenolic compounds have repeatedly been associated with improved affective stability and enhanced cognitive performance, likely mediated by their antioxidative and serotonergic interactions [250,251,252]. L-Trp supplementation and dietary enrichment strategies produced measurable anxiolytic and mood-stabilizing effects, although outcomes on sleep were inconsistent [251,252,253]. Berberine, while less frequently studied in formal RCTs, emerges from systematic analyses as a promising candidate due to its dual action on serotonergic pathways and neuroinflammation [15,252,254]. Taken together, the findings point to possible benefits across several symptom domains, yet the evidence remains preliminary because outcomes vary widely and most trials are too small to establish reliable clinical effects.
Clinical investigations of phytochemicals that influence Trp metabolism face several persistent limitations that limit their translational potential [255,256]. Variability in dosage and duration across trials complicates direct comparisons and limits the ability to establish optimal therapeutic regimens [255,257]. Intervention periods are often short, failing to capture long-term efficacy or safety profiles, particularly for compounds intended for chronic use in mood and anxiety disorders [255,257]. Patient populations are frequently homogeneous, with many studies confined to healthy adults or narrowly defined subgroups, reducing generalizability to clinically diverse cohorts [256,257]. Outcome measures further compound these issues, as trials employ a wide array of scales for mood, anxiety, and sleep, hindering reproducibility and meta-analytic synthesis [256,257]. Future research must prioritize methodological rigor by standardizing dosing protocols, harmonizing validated psychiatric and sleep measures, and extending trial durations [212,258]. Incorporating mechanistic endpoints through metabolomics, microbiome analysis, and neuroimaging would strengthen causal inference and bridge the translational gap [258,259].
4.2. Specific Tryptophan (Trp)-Rich Phytocompounds and Associated Clinical Outcomes
Griffonia simplicifolia seeds, a natural source of 5-HTP, have attracted considerable attention for their potential therapeutic effects on mood and anxiety disorders [260,261]. Small studies report improvements in mood, anxiety, and sleep, yet these observations remain preliminary because sample sizes are limited and replication is scarce. [261,262]. These early signals are promising, but they cannot be interpreted as evidence of clinical readiness at this stage. These outcomes are biologically plausible, given that 5-HTP serves as the immediate precursor of 5-HT, bypassing the tightly regulated hydroxylation of Trp and directly fueling central serotonergic pathways [262,263]. Several narrative reviews underscore its promise in alleviating depression and insomnia, while preliminary trials report measurable gains in affective stability and reduced sleep latency [261,262]. Importantly, these findings align with mechanistic insights showing that oral 5-HTP elevates central 5-HT availability, thereby modulating emotional processing, stress resilience, and circadian regulation [262,264]. Although current evidence remains constrained by methodological limitations and the absence of large randomized controlled trials, the accumulating data suggest a clinically relevant role worth further exploration [238,265].
Soy isoflavones, particularly genistein and S-equol, have emerged as promising candidates for mood regulation through their intertwined effects on neuroinflammation and Trp metabolism [266,267]. Clinical and preclinical findings suggest that these phytochemicals suppress pro-inflammatory signaling cascades, including TLR4/NF-κB activation, while simultaneously shifting Trp utilization away from the KYN pathway and toward 5-HT synthesis [250,267]. This dual action is highly relevant for psychiatric disorders, as chronic inflammation and altered Trp metabolism often converge to lower 5-HT availability and exacerbate depressive and anxiety symptoms [130,268]. Trials in animal models demonstrate that soy isoflavones normalize 5-HT levels, reduce KYN accumulation, and enhance synaptic plasticity, culminating in reduced depressive-like and anxiety-related behaviors [250,267]. Although direct human clinical trials remain scarce, pilot studies in related contexts indicate improvements in affective stability, sleep quality, and cognitive performance [269,270]. Together, these findings position soy isoflavones as a biologically grounded strategy with significant translational potential awaiting rigorous clinical validation.
Berberine and albiflorin have recently emerged as two of the most compelling phytochemicals targeting the gut–brain axis, with converging evidence suggesting they can influence mood and cognition through microbiota-driven mechanisms [271,272]. Preclinical and early translational studies show that berberine exerts profound regulatory effects on gut microbial composition, enhancing the production of short-chain fatty acids and restoring gut barrier integrity, while simultaneously reducing peripheral and central inflammatory markers [271]. These changes translate into improved hippocampal 5-HT, dopamine, and BDNF signaling, resulting in alleviation of depressive-like behaviors and measurable gains in memory and learning in animal models of both depression and Alzheimer’s disease [273,274]. Albiflorin, a bioactive monoterpene glycoside, demonstrates a complementary mode of action: gut microbes metabolize it into benzoic acid, which penetrates the blood–brain barrier and inhibits D-amino acid oxidase, ultimately fostering neurogenesis and reducing depressive symptoms [275,276]. Although direct human clinical trials remain sparse, small-scale investigations and mechanistic reviews highlight reductions in depression severity, relief from anxiety, and preliminary cognitive benefits consistent with these gut-brain interactions [277,278]. Together, the data suggest that berberine and albiflorin represent promising candidates for modulating Trp metabolism, suppressing neuroinflammation, and enhancing cognitive resilience, warranting rigorous clinical validation to confirm their therapeutic potential [279,280].
Polyphenols and flavonoids have gained significant attention as multi-targeted modulators of brain health, with a growing body of evidence supporting their roles in mood stabilization, neuroprotection, and anti-inflammatory activity [281,282,283]. Clinical studies involving polyphenol-rich extracts demonstrate improvements in depressive and anxiety symptoms, often accompanied by reductions in circulating inflammatory cytokines such as TNF-α and IL-6, and improved systemic redox balance [253,284,285]. Flavonoids like naringenin and quercetin show particular promise, as they not only suppress pro-inflammatory pathways including NF-κB and MAPK but also promote hippocampal neurogenesis and synaptic plasticity, thereby enhancing resilience against stress-related pathology [252,283,286]. A notable mechanistic insight involves their potential modulation of the KYN pathway. By restraining the diversion of Trp toward neurotoxic KYN metabolites, polyphenols may preserve 5-HT synthesis while fostering neuroprotective KYNA balance [252,287,288]. Preclinical studies reinforce these effects, highlighting synergistic actions between flavonoids and gut microbiota in maintaining metabolic and neurotransmitter homeostasis [287,288,289]. While large-scale psychiatric trials remain limited, smaller investigations and indirect clinical evidence suggest tangible benefits in mood stabilization, cognitive function, and anxiety reduction [252,288,290]. Collectively, these findings underscore the translational potential of polyphenols and flavonoids as pleiotropic agents that target both immune and neurotransmitter pathways. Yet, they demand well-designed trials to establish their clinical efficacy in psychiatric populations.
A visual figure illustrates the mechanisms by which key phytochemicals, such as 5-HTP, isoflavones, berberine, and polyphenols, influence depression biology (Figure 7). The diagram highlights their capacity to enhance 5-HT synthesis, regulate KYN metabolism, reduce neuroinflammation, and promote neurogenesis. Arrows connect peripheral processes, such as gut microbial modulation and immune signaling, to central effects in the brain, emphasizing the bidirectional nature of the gut–brain axis. By mapping these pathways together, the figure clarifies how dietary and phytotherapeutic interventions can complement pharmacological strategies. This visual summary strengthens understanding of mechanistic complexity while guiding translational applications in depression management.
5. Gaps and Controversies in Current Research
Despite growing enthusiasm for plant-derived Trp interventions, substantial uncertainties continue to challenge their clinical translation [291,292]. Research to date is marked by heterogeneity in trial design, inconsistency in dosing regimens, and insufficient mechanistic clarity, making it difficult to draw definitive conclusions about efficacy or safety [293,294]. Moreover, debates over bioavailability and the relative value of dietary versus supplemental sources remain unresolved [295]. This section critically examines the gaps and controversies that persist in the field, identifying key obstacles that must be addressed to advance evidence-based applications in psychiatry [292,295].
5.1. Identified Gaps in Current Research
Limited availability of long-term clinical trials on plant-derived Trp compounds represents a critical bottleneck in advancing depression therapeutics [296,297,298]. Most clinical investigations are confined to short durations, providing valuable insights into initial symptom reduction but leaving long-term efficacy and safety largely unexplored [32,298,299]. Historical work, such as Herrington’s comparative trial of L-Trp and electroconvulsive therapy, included a six-month follow-up but involved a limited sample, offering only fragmentary evidence regarding the durability of response [297]. This pattern highlights a broader issue: without longitudinal monitoring, it is impossible to determine whether early benefits represent genuine disease modification or transient relief [296,298,299]. Moreover, potential long-term risks, including serotonergic overstimulation, metabolic alterations, or shifts in gut–brain interactions, remain insufficiently characterized [12,298,300]. The absence of such data also obscures whether chronic administration leads to tolerance or adaptive physiological changes that diminish therapeutic value [297,298,300]. To honestly assess the role of plant-derived Trp compounds in the management of chronic depression, carefully designed, multi-year randomized controlled trials with repeated follow-up assessments are urgently required.
Inconsistency in dosing regimens, variability in formulation purity, and absence of standardized protocols represent significant obstacles to interpreting the therapeutic potential of plant-derived Trp compounds for depression treatment. Across clinical and preclinical studies, dosing practices vary widely, ranging from fixed low-dose supplementation to high-dose concentrated extracts, often administered at different frequencies. This lack of uniformity makes it nearly impossible to establish reliable dose–response relationships or determine optimal therapeutic windows. Differences in extraction methods, including solvent choice, temperature, and processing time, further affect yield, bioavailability, and pharmacological activity. Clinical trials using saffron or chamomile extracts, while reporting short-term improvements in depressive symptoms, often employ heterogeneous extraction techniques and undefined purity standards, preventing direct comparison or meta-analytic synthesis. Without harmonized dosing protocols and rigorous phytochemical standardization, conclusions regarding the safety and efficacy of these promising compounds remain provisional, limiting translation into clinical guidelines.
Although the Trp–KYN pathway has been extensively linked to depression and neuroinflammation, mechanistic clarity regarding how plant-derived phytochemicals act upon this axis is strikingly underdeveloped. Most studies remain descriptive, emphasizing correlations between altered Trp metabolism and psychiatric outcomes, while offering little insight into whether phytochemicals exert direct enzymatic modulation or act indirectly through immune regulation and microbiome interactions. For instance, trials investigating saffron and chamomile supplementation demonstrate short-term clinical benefits but fail to examine whether these effects are mediated by IDO inhibition, KYN–5-HT balance, or shifts in neuroinflammatory markers. Similarly, preclinical studies using Trp-rich diets in stress models suggest involvement of the gut–brain axis, but lack detailed mapping of metabolite flux or cell-type–specific enzyme regulation. Without such mechanistic endpoints, it remains speculative whether phytochemical interventions primarily restore serotonergic tone, rebalance KYN derivatives, or attenuate inflammatory cascades. The paucity of molecular and biochemical investigations severely limits translation into targeted therapies. To overcome this, future research must integrate omics-driven profiling, metabolite quantification, and longitudinal immune assays within both animal and human trials. Only through such rigor can we disentangle direct versus indirect pathways and fully clarify how plant-derived Trp interventions modulate the biochemical underpinnings of chronic depression. Such uncertainties make it difficult to identify responders, to define therapeutic windows, or to design trials that align mechanistic action with clinical endpoints.
5.2. Controversies Regarding Clinical Efficacy and Bioavailability
The debate surrounding dietary versus supplemental Trp remains unresolved, reflecting both biochemical complexity and methodological gaps in current research [138,301,302]. Opinions diverge sharply because supplements provide precision while dietary sources engage wider systemic pathways. Evidence supporting either scenario remains incomplete. Isolated supplementation offers clear pharmacological advantages, including predictable dosing, rapid absorption, and more consistent increases in serum Trp and downstream metabolites [301,303,304]. Systematic reviews demonstrate modest but reproducible improvements in mood and sleep quality with supplementation, yet optimal dosing strategies are far from established [301,303,304]. The evidence is encouraging but limited by small sample sizes, brief interventions, and inconsistent measurement panels. These constraints reduce confidence in effect size estimates and complicate comparisons across studies. In contrast, whole-food Trp intake introduces a range of variables: competition with other large neutral amino acids for transport across the blood–brain barrier, food matrix effects on absorption, and significant modulation by the gut microbiota [138,301,302]. Animal studies suggest that dietary Trp enhances intestinal barrier function and alters immune signaling pathways, implying potential systemic benefits distinct from supplementation, though mechanistic specificity remains elusive [305,306]. Although compelling, these findings primarily come from controlled laboratory settings and rarely translate directly into human physiology. Until head-to-head human trials are completed, conclusions remain tentative. For example, interventions in piglet and rodent models show that dietary Trp influences gut–brain and immune interactions more strongly than supplementation alone, but human trials directly comparing the two approaches are absent [307,308,309]. The resulting uncertainty complicates translation into clinical practice [195,308,309]. While supplements provide pharmacological precision, dietary sources may confer broader metabolic resilience, albeit with unpredictable bioavailability [309,310]. Head-to-head, long-term trials are urgently required to determine whether whole-food Trp and supplementation exert complementary or divergent therapeutic effects in depression [200,310,311,312].
Bioavailability remains one of the most contentious issues in evaluating dietary Trp and plant-based supplements, as absorption and metabolic fate are strongly shaped by gut microbiota composition, digestive efficiency, and individual genetic differences. While isolated supplementation produces predictable increases in plasma Trp, dietary sources are subject to competition with other large neutral amino acids and to modulation by the food matrix, resulting in less consistent outcomes. Compounding these pharmacokinetic challenges is the unresolved question of whether enhanced bioavailability directly translates into clinically meaningful improvements in depression, since mood regulation involves downstream serotonergic, KYN, and immune pathways. Finally, safety concerns linked to supplement impurities, as highlighted by the eosinophilia–myalgia syndrome outbreak, emphasize the critical need for rigorous pharmacokinetic and pharmacodynamic studies. Only with such evidence can bioavailability be meaningfully tied to therapeutic outcomes.
6. Clinical Translation: From Bench to Bedside
Research into plant-derived Trp compounds has expanded rapidly, yet critical uncertainties continue to limit their clinical translation [13,31,143]. Evidence from preclinical and early clinical studies highlights promising effects on mood, sleep, and stress resilience, but methodological gaps and unresolved controversies persist [31,240,301]. Questions surrounding long-term efficacy, optimal dosing, and mechanistic pathways remain central obstacles [138,143,303]. Likewise, debates over dietary versus supplemental sources and challenges of bioavailability further complicate interpretation [134,240,301]. This section examines these gaps and controversies, outlining unresolved issues that must be addressed before phytochemical modulation of Trp metabolism can be reliably integrated into psychiatric practice.
6.1. Practical Considerations for Clinical Application
Determining the effective and safe dosage of plant-derived Trp remains a central challenge in translating experimental findings into clinical practice [31,313,314]. Current evidence suggests a relatively wide therapeutic window, yet precise dosing guidelines are lacking, particularly for chronic depression [31,298,313]. Human studies indicate that daily intakes of 1–5 g are generally well tolerated, with a proposed NOAEL of 4.5 g/day in young adults [303,314,315]. These findings are consistent with short-term supplementation trials in healthy women, in which no safety concerns emerged after 3 weeks of use [298,304,315]. Animal studies further support this margin, showing no toxic effects at doses up to 2000 mg/kg/day over 90 days, underscoring a substantial buffer between practical and harmful exposure [31,240,316,317]. However, dosage requirements appear to vary depending on formulation: free L-Trp, fermented products, and protein-bound dietary forms display distinct bioavailability profiles that may influence clinical outcomes [31,304,316]. Patient-specific factors, such as age, sex, metabolic status, and comorbid conditions, are rarely addressed in current trials, leaving critical gaps in personalized dosing [31,304,316]. Significantly, historical episodes of eosinophilia-myalgia syndrome highlight the need for strict quality control, as impurities rather than Trp itself caused adverse events [240,298,318]. Collectively, available evidence supports cautious short-term use within established limits, while long-term efficacy and safety require rigorous randomized controlled trials across diverse populations.
Plant-derived Trp supplements are widely regarded as safe when used within recommended limits, yet their safety profile warrants careful consideration given both historical precedents and documented adverse reactions [314,319,320]. The most serious concern remains eosinophilia–myalgia syndrome, a rare but debilitating condition that emerged in the late 1980s due to contaminated L-Trp supplements [320,321,322]. Although later investigations confirmed that impurities rather than the amino acid itself were responsible, this episode underscores the importance of rigorous quality control and sourcing from reliable manufacturers [319,320,321]. This distinction is essential for current practice because modern formulations vary in quality, and safety remains dependent on manufacturing standards rather than the compound itself. Beyond this, clinical evidence indicates that most individuals tolerate doses up to 5 g per day for short durations without metabolic or systemic disturbances [314,318,323]. Nevertheless, higher intakes or prolonged use have occasionally been associated with nausea, tremor, dizziness, or excessive drowsiness. Interactions with serotonergic medications raise the additional risk of 5-HT syndrome, marked by neuromuscular rigidity, confusion, and fever, highlighting the need for medical oversight in polypharmacy contexts. Related compounds, such as 5-hydroxytryptophan, show similarly favorable safety profiles, though isolated case reports raise unresolved concerns that warrant continued vigilance [318,323,324]. Practical guidance emphasizes regular monitoring for muscle pain, skin changes, and gastrointestinal symptoms, patient education on recognizing side effects, and prompt discontinuation or dose reduction if adverse signs emerge, ensuring therapeutic benefit without undue risk [314,325,326].
Plant-derived Trp supplements raise important considerations regarding potential interactions with commonly prescribed medications, particularly antidepressants and anxiolytics that influence serotonergic signaling [31,134,327]. The most clinically significant risk is 5-HT syndrome. This potentially life-threatening condition can occur when Trp is combined with selective 5-HT reuptake inhibitors, 5-HT-norepinephrine reuptake inhibitors, monoamine oxidase inhibitors, or certain anxiolytics [31,134,327]. Characterized by agitation, hyperreflexia, autonomic instability, and, in severe cases, hyperthermia or coma, 5-HT syndrome underscores the need for clinicians to carefully review all concurrent serotonergic therapies before recommending Trp [142,328,329]. Beyond this, pharmacological overlap within the 5-HT and KYN pathways suggests that plant-derived Trp compounds could interfere with drugs targeting immune function or neuroinflammation, although robust clinical evidence remains sparse [31,134,330]. Pharmacokinetic interactions are less well defined but may involve competition for metabolic enzymes, potentially altering drug clearance and therapeutic effectiveness [142,329,331]. Contraindications include patients with a history of 5-HT syndrome, those taking multiple serotonergic agents, and individuals with unstable psychiatric or metabolic disorders [142,328,329]. Practical recommendations emphasize individualized assessment, use of the lowest effective dose, and structured patient education regarding early symptoms of adverse interactions [329,331,332]. Close monitoring and early discontinuation in the presence of concerning signs are essential to balance potential therapeutic benefits with safety in clinical application.
6.2. Personalized Medicine Approaches
Gut microbiota profiling is rapidly gaining attention as a tool for personalized depression management, with compelling evidence that microbial composition influences both Trp metabolism and therapeutic response to phytochemicals [192,194,333]. Altered microbiota profiles can shift the balance between 5-HT and KYN pathways, thereby modulating neuroinflammation and mood regulation [136,194,334]. Clinical studies now suggest that baseline microbiota signatures may predict antidepressant response: patients enriched in beneficial taxa, such as Faecalibacterium prausnitzii or Bifidobacterium, demonstrate higher remission rates, while reduced diversity often correlates with treatment resistance [333,335,336]. Early trials of Trp-rich dietary interventions and probiotics also reveal that therapeutic efficacy may hinge on an individual’s microbial ecosystem, with some patients showing robust mood improvement and others little benefit despite identical regimens [333,335,337]. This variability highlights the potential of stool-based profiling to guide personalized strategies, matching patients to dietary, probiotic, or phytotherapeutic interventions most likely to be effective [208,335,338]. Emerging data also point to machine learning approaches that integrate microbial, metabolic, and cytokine biomarkers for predictive modeling of treatment outcomes [338,339,340]. Yet translation into practice is still nascent: large-scale, longitudinal trials and functional multi-omics studies are essential to confirm which microbial profiles drive therapeutic benefit [115,194,336]. Ultimately, microbiota-informed stratification could transform the management of depression into a more precise and individualized discipline.
Genetic polymorphisms in key enzymes regulating Trp metabolism, such as IDOs, TDO, and TPH, play a decisive role in shaping vulnerability to depression and responsiveness to therapeutic interventions [192,194,333]. Variants in these genes influence the metabolic fate of Trp, determining whether it is directed toward 5-HT synthesis or shunted into the KYN pathway, with downstream consequences for neuroinflammation and mood regulation [115,136,334]. Clinical data suggest that polymorphisms in TPH2 and 5-HT transporter-linked regions correlate with depression severity and treatment resistance. At the same time, IDO and IFN-γ gene variants are associated with enhanced KYN production, amplifying immune-driven depressive symptoms [113,341,342]. Postpartum depression studies also reveal IDO-related polymorphisms as predictors of higher risk, underscoring the clinical relevance of pathway-specific genetic variation [341,343,344]. These insights highlight the potential of gene profiling to guide individualized treatment, in which patients with high-risk alleles may benefit more from interventions targeting the KYN pathway or from plant-derived Trp formulations designed to enhance serotonergic flux [10,113,341]. Although current evidence remains fragmented, integrating genotyping into personalized medicine frameworks could enable patient stratification, prediction of treatment outcomes, and rational selection of phytotherapeutic strategies [341,344,345]. Expanding multi-ethnic genetic studies and intervention trials is essential to validate these applications and move toward precision psychiatry [346].
7. Future Perspectives and Research Directions
Clinical translation represents the critical step of transforming mechanistic insights and preclinical discoveries into therapeutic strategies that can be safely and effectively applied in patient care [150,347,348]. For plant-derived Trp and related compounds, this bridge from bench to bedside demands careful attention to dosing, safety, interactions, and long-term monitoring [143,349,350]. Equally important is the integration of personalized medicine approaches, in which genetic, metabolic, and microbiome profiles guide therapeutic decisions [351,352,353]. By addressing these considerations, clinical application moves beyond experimental promise to evidence-based strategies capable of shaping future psychiatric treatments.
7.1. Advancing Clinical Evidence through Robust Study Designs
Large-scale, multi-center clinical trials represent the critical next step for validating plant-derived Trp therapies in depression [10,30,354]. While early-phase studies and small-scale trials suggest therapeutic promise, their limited sample sizes, short durations, and restricted demographic scope constrain clinical interpretation [115,354,355]. Multi-center designs allow for the inclusion of diverse populations across ethnic, geographic, and socioeconomic backgrounds, thereby addressing genetic polymorphisms in enzymes such as IDO, TDO, and TPH that influence Trp metabolism and treatment responsiveness [10,30,355]. Such diversity is essential to move beyond narrow, context-specific findings and toward results with broad clinical applicability. Equally important, multi-center collaborations enhance statistical power, enabling the detection of subtle treatment effects and the evaluation of subgroup responses [356,357]. Coordinated protocols across research networks also promote methodological consistency, ensuring that differences in formulation, dosing, and biomarker assessments do not confound outcomes [358,359,360]. Promising applications include testing whether specific genotypic or microbiota-defined subgroups derive greater benefit from phytotherapeutic interventions, a question that can be adequately addressed only through large, well-controlled trials [361,362,363]. Beyond efficacy, safety monitoring on a global scale provides crucial insight into rare adverse events and long-term tolerability [359,360,363]. Ultimately, multi-center trials create the foundation for standardized guidelines, ensuring that plant-derived Trp interventions are evaluated with the rigor required for integration into precision psychiatry [364,365,366,367,368].
Standardization of methodologies and outcome measures is indispensable for advancing plant-derived Trp therapies from promising pilot studies to clinically actionable interventions [369,370,371]. At present, heterogeneity in dosing protocols, extraction techniques, biomarker panels, and outcome assessments undermines comparability across trials and limits the ability to generate robust meta-analyses [372,373]. For instance, differences in extraction solvents and processing conditions can yield markedly divergent metabolite profiles, complicating interpretation of therapeutic efficacy [242,372]. Similarly, clinical studies often apply variable dosing schedules—ranging from milligram-scale supplementation to multi-gram intakes—without harmonized reference points, making it difficult to establish optimal therapeutic windows [374,375]. Biomarker selection presents another challenge: while some studies emphasize 5-HT and KYN metabolites, others focus on inflammatory cytokines or gut microbiota-derived indoles, creating fragmented datasets that resist synthesis [376,377,378]. Outcome assessment tools are equally inconsistent, with trials variously employing clinician-rated depression scales, patient self-reports, or biochemical endpoints [375,378]. A shift toward standardized protocols—including consensus dosing frameworks, validated extraction procedures, and a unified “indolome” biomarker panel—would significantly enhance reproducibility and clinical interpretability [379,380,381]. Such consistency is particularly critical for large-scale, multi-center trials, where methodological alignment ensures that findings are both statistically reliable and generalizable [358,379,382]. Ultimately, methodological standardization is not a technical detail but the foundation for transforming plant-derived Trp research into credible, guideline-ready therapies.
Long-term follow-up studies are indispensable for establishing the actual clinical value of plant-derived Trp therapies in depression management [10,31,383]. While short-term trials consistently report symptomatic improvements, they cannot determine whether these benefits persist over months or years, nor can they capture delayed adverse effects that may only emerge with prolonged use [297,298,383]. Longitudinal research is therefore essential to assess the durability of mood stabilization, sustained neurobiological changes, and overall psychiatric outcomes [10,32,384]. Equally important is evaluating patient adherence, as real-world compliance often declines over time and directly influences therapeutic success [32,298,385]. Early evidence from nutritional and probiotic interventions suggests that plant-derived Trp compounds can modulate 5-HT and KYN pathways in ways that might yield lasting benefit, yet without systematic long-term monitoring, such effects remain speculative [12,299,386]. Furthermore, the safety profile of chronic use is not fully characterized, particularly in populations with comorbidities or concurrent pharmacotherapies [241,383,384]. Long-term follow-up would also clarify whether plant-derived formulations maintain efficacy or whether tolerance, metabolic adaptation, or microbiota shifts reduce their impact over time [31,383,387]. Only through rigorous, multi-year studies with repeated biochemical, clinical, and functional assessments can the field move from preliminary promise to evidence-based recommendations that ensure both efficacy and safety in chronic depression care.
7.2. Combined Pharmacological and Dietary Intervention Strategies
Integrating dietary interventions with standard pharmacological treatments offers a compelling strategy for enhancing depression management [388,389,390]. Evidence from randomized controlled trials, such as the SMILES study, demonstrates that improving diet alongside usual care results in significantly greater reductions in depressive symptoms and higher remission rates than control conditions [390,391,392]. Meta-analyses further support these findings, showing that dietary modifications consistently improve mood outcomes, even when participants are already receiving antidepressants [393,394,395,396]. Nutrient-focused approaches, including supplementation with omega-3 fatty acids, vitamin D, and probiotics, have shown additive benefits, with effects on neurotransmitter synthesis, immune regulation, and gut–brain communication that complement pharmacological mechanisms [388,396,397]. Together, these findings highlight the rationale for combined strategies that address both biological and lifestyle determinants of mental health.
The integration of diet with pharmacotherapy also has practical advantages. Nutritional interventions may mitigate side effects commonly associated with antidepressant use, such as weight gain, fatigue, and gastrointestinal disturbances, thereby improving tolerability and adherence [396,397,398]. Patients often perceive dietary changes as more holistic and less stigmatizing than medication alone, thereby enhancing engagement and long-term treatment compliance [399,400,401]. From a mechanistic perspective, dietary modulation of inflammation, oxidative stress, and Trp metabolism provides synergistic support to monoamine-based drug action [389,394,402]. Thus, combining dietary and pharmacological approaches is not merely additive but can produce synergistic effects, optimizing both symptom relief and overall health outcomes [389,403]. This integrated model represents one of the most promising directions for personalized, sustainable depression care.
Future research on combined dietary Trp and antidepressant interventions requires rigorously designed clinical trials that address both short- and long-term outcomes. The most promising model is a randomized, double-blind, placebo-controlled design with multiple parallel arms: antidepressant plus Trp, antidepressant plus placebo, Trp alone, and full placebo [296,385,404]. Such a structure allows investigators to disentangle additive and synergistic effects while also clarifying the standalone impact of dietary supplementation [32,385,404]. Control conditions should be carefully balanced to avoid expectancy bias, and stratification by baseline nutritional status or genetic polymorphisms in Trp metabolism (e.g., IDO, TDO, TPH variants) would enhance precision in interpreting outcomes [32,210,405].
Biomarker monitoring should be a central feature of these trials. Serial assessments of plasma Trp, KYN metabolites, 5-HT levels, inflammatory cytokines, and sleep parameters would help map mechanistic pathways linking supplementation to clinical response [406,407,408]. Alongside these biological endpoints, standardized psychiatric outcome measures—such as HDRS and BDI scores—should be collected, complemented by cognitive and emotional processing tasks to capture subtle shifts in mood regulation [409,410,411]. Adherence can be monitored with digital tracking tools and serum metabolite checks [412,413,414]. A duration of at least 8–12 weeks is needed to capture short-term efficacy, but follow-up at 6 to 12 months will be critical to assess durability, safety, and adherence in real-world contexts [415,416]. These models would not only clarify the therapeutic potential of combined interventions but also provide insight into patient subgroups most likely to benefit. By integrating dietary, pharmacological, and mechanistic perspectives, such studies could build the evidence base needed to translate plant-derived Trp strategies into precision-guided clinical practice.
7.3. Leveraging Biotechnological Innovations for Personalized Depression Management
Precision nutrition is redefining how dietary strategies are applied in depression care by emphasizing individualized interventions informed by genetic and metabolic profiling [417,418,419]. Nutritional genomics has revealed that polymorphisms in genes such as SLC6A4 and BDNF can influence how individuals respond to nutrient-based therapies, including plant-derived Trp [417,420,421]. These differences suggest that personalized dosing algorithms may eventually replace one-size-fits-all approaches, although such models remain experimental at present. For example, specific 5-HT transporter variants may determine whether Trp is preferentially metabolized into 5-HT or diverted into the KYN pathway, directly affecting mood outcomes [332,422,423]. Integrating this genetic information into clinical planning could enable targeted dietary prescriptions tailored to a patient’s unique metabolic capacity, thereby improving the precision and efficacy of treatment.
Beyond genomics, advanced biomarker analyses provide powerful tools to individualize dietary interventions [424,425,426]. Gut microbiota profiling typically involves sequencing approaches that characterize intestinal microbial composition and functional metabolic capacity. Profiling gut microbiota, serum metabolites, and inflammatory markers allows clinicians to predict and monitor how patients metabolize Trp-rich foods or supplements [427,428,429]. Emerging evidence shows that personalized diets can reduce depressive symptoms, alter microbial composition, and improve quality of life in both older adults and adolescents [393,399,427]. These findings suggest that combining biomarker-driven monitoring with dietary tailoring may optimize long-term outcomes [425,426,430]. Importantly, precision nutrition also considers comorbidities: in patients with obesity or diabetes, individualized nutritional plans have demonstrated benefits not only for metabolic regulation but also for mood stabilization [265,428,430]. Together, these insights support a move away from generic dietary recommendations toward integrative, personalized protocols. When fully developed, such strategies could complement pharmacological treatments and establish plant-based Trp interventions as a cornerstone of personalized psychiatry.
Microbiome-targeted therapeutic approaches are emerging as innovative strategies for addressing depression by modulating gut–brain communication and regulating Trp metabolism [383,431,432]. Probiotics and prebiotics are the most extensively studied, with randomized controlled trials showing that probiotic supplementation can alleviate depressive symptoms, maintain microbial diversity, and enhance serotonergic signaling [386,433,434]. Specific strains, such as Lactobacillus and Bifidobacterium, have been linked to improved mood outcomes and altered brain activation patterns, suggesting that targeted microbial interventions may complement conventional pharmacotherapy [337,435,436]. Prebiotics, by selectively stimulating beneficial gut bacteria, also influence the production of neuroactive metabolites, offering an indirect yet consequential means of modulating the Trp–KYN pathway [31,431,437].
FMT represents a more radical but promising intervention [204,438,439]. Preclinical studies in rodent models demonstrate that transplantation from healthy donors reduces depressive behaviors, restores neurotransmitter balance, and dampens neuroinflammatory processes [440,441,442]. Early clinical findings echo these benefits, although long-term safety and efficacy remain insufficiently studied [443,444,445]. Beyond these approaches, engineered microbial therapies are on the horizon, with genetically modified strains designed to produce specific neuroactive compounds or regulate metabolic flux through Trp-related pathways [194,200,333]. When combined with multi-omics profiling, these advanced strategies could allow for precision-guided microbial interventions tailored to an individual’s baseline microbiome and metabolic profile [194,204,333]. Collectively, probiotics, prebiotics, FMT, and engineered microbes hold strong potential to enhance therapeutic outcomes, particularly when integrated into holistic models of depression care that also consider diet, pharmacology, and genetics.
8. Conclusions
This review synthesizes current knowledge on plant-derived Trp interventions and outlines how these mechanisms intersect with psychiatric research. The evidence highlights potential applications while clarifying where further work is needed. By weaving together evidence from neuroinflammation, KYN metabolism, gut–brain interactions, and phytochemical modulation, the article provides a comprehensive framework that highlights both theoretical insights and translational promise. Notably, the findings stress that phytocompounds can recalibrate serotonergic balance, temper neurotoxic cascades, and enhance resilience through multimodal pathways, supporting their integration alongside conventional antidepressants. At the same time, methodological limitations—heterogeneous dosing protocols, brief intervention periods, and insufficient long-term safety data—demand carefully designed clinical trials. Future research should prioritize precision nutrition strategies, biomarker-guided interventions, and longitudinal monitoring to elucidate the sustained impact of these therapies fully. Advancing this field will require interdisciplinary collaboration and coordinated multi-center trials that reflect real-world diversity and consistent methodological standards. The promise is real, yet current data remain preliminary because most trials are underpowered or methodologically heterogeneous. Larger, more mechanistically grounded studies are needed to confirm clinical relevance and advance the field toward safer, more effective, and biologically informed treatment paradigms. A significant gap remains in the absence of coordinated multi-center RCTs that apply harmonized dosing frameworks, standardized extraction procedures, and unified biomarker panels. Without such designs, it is difficult to determine whether early signals of efficacy translate into durable clinical benefit.
Author Contributions
Conceptualization, S.M.B., A.C., M.T. and L.F.L.; methodology, C.R.P.D.; investigation, A.C.C., E.P.L., V.E.V., C.R.P.D. and V.M.C.S.C., and L.F.L.; writing—original draft preparation, S.M.B., L.F.L, R.S.A.H. and M.T.; writing—review and editing, S.M.B., L.F.L., V.E.V., C.M.G., E.d.S.B.M.P. and R.S.A.H.; visualization, S.M.B., M.T. and C.M.G.; supervision, S.M.B. and M.T.; project administration, S.M.B.; funding acquisition, S.M.B. and M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the HUN-REN Hungarian Research Network to M.T.
Data Availability Statement
Not applicable.
Acknowledgments
Figure 1 was created using Mind the Graph (https://mindthegraph.com/, last accessed on November 2025). Its use is allowed under proper subscription. Figures 2-7 were built using resources from www.freepik.com, www.canva.com, and www.pixabay.com websites. Only free, royalty-free, or public-domain images were utilized from these latter sources. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 3-HK | 3-hydroxykynurenine |
| 5-HT | serotonin |
| 5-HTP | 5-hydroxytryptophan |
| AhR | aryl hydrocarbon receptor |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| BDI | Beck Depression Inventory |
| BDNF | brain-derived neurotrophic factor |
| CDK5 | cyclin-dependent kinase 5 |
| FMT | fecal microbiota transplantation |
| HDRS | Hamilton Depression Rating Scale |
| HPA | hypothalamic–pituitary–adrenal |
| IDO | indoleamine 2,3-dioxygenase |
| IFN | interferon |
| IL | interleukin |
| KAT | kynurenine aminotransferases |
| KMO | kynurenine 3-monooxydase |
| KYN | kynurenine |
| KYNA | kynurenic acid |
| MAPK | mitogen-activated protein kinase |
| MDD | major depressive disorder |
| NF-κB | nuclear factor kappa B |
| NLRP3 | NLR family pyrin domain-containing protein 3 |
| NMDA | N-Methyl-D-Aspartate |
| QA | quinolinic acid |
| QPRT | quinolinate phosphoribosyltransferase |
| RCT | randomized controlled trial |
| SSRI | selective serotonin reuptake inhibitor |
| TDO | tryptophan 2,3-dioxygenase |
| TLR4 | toll-like receptor 4 |
| TNF-α | tumor necrosis factor-alpha |
| Trp | tryptophan |
| TPH | tryptophan hydroxylase |
| VEGF | vascular endothelial growth factor |
References
- Filatova, E.V.; Shadrina, M.I.; Slominsky, P.A. Major Depression: One Brain, One Disease, One Set of Intertwined Processes. Cells 2021, 10. [CrossRef]
- Fries, G.R.; Saldana, V.A.; Finnstein, J.; Rein, T. Molecular pathways of major depressive disorder converge on the synapse. Mol Psychiatry 2023, 28, 284-297. [CrossRef]
- Cui, L.; Li, S.; Wang, S.; Wu, X.; Liu, Y.; Yu, W.; Wang, Y.; Tang, Y.; Xia, M.; Li, B. Major depressive disorder: hypothesis, mechanism, prevention and treatment. Signal Transduct Target Ther 2024, 9, 30. [CrossRef]
- Proudman, D.; Greenberg, P.; Nellesen, D. The Growing Burden of Major Depressive Disorders (MDD): Implications for Researchers and Policy Makers. Pharmacoeconomics 2021, 39, 619-625. [CrossRef]
- Greenberg, P.E.; Fournier, A.A.; Sisitsky, T.; Simes, M.; Berman, R.; Koenigsberg, S.H.; Kessler, R.C. The Economic Burden of Adults with Major Depressive Disorder in the United States (2010 and 2018). Pharmacoeconomics 2021, 39, 653-665. [CrossRef]
- Pu, J.; Liu, Y.; Zhang, H.; Tian, L.; Gui, S.; Yu, Y.; Chen, X.; Chen, Y.; Yang, L.; Ran, Y.; et al. An integrated meta-analysis of peripheral blood metabolites and biological functions in major depressive disorder. Mol Psychiatry 2021, 26, 4265-4276. [CrossRef]
- Kouba, B.R.; de Araujo Borba, L.; Borges de Souza, P.; Gil-Mohapel, J.; Rodrigues, A.L.S. Role of Inflammatory Mechanisms in Major Depressive Disorder: From Etiology to Potential Pharmacological Targets. Cells 2024, 13. [CrossRef]
- Athira, K.V.; Bandopadhyay, S.; Samudrala, P.K.; Naidu, V.G.M.; Lahkar, M.; Chakravarty, S. An Overview of the Heterogeneity of Major Depressive Disorder: Current Knowledge and Future Prospective. Curr Neuropharmacol 2020, 18, 168-187. [CrossRef]
- Ilavská, L.; Morvová, M., Jr.; Paduchová, Z.; Muchová, J.; Garaiova, I.; Ďuračková, Z.; Šikurová, L.; Trebatická, J. The kynurenine and serotonin pathway, neopterin and biopterin in depressed children and adolescents: an impact of omega-3 fatty acids, and association with markers related to depressive disorder. A randomized, blinded, prospective study. Front Psychiatry 2024, 15, 1347178. [CrossRef]
- Correia, A.S.; Vale, N. Tryptophan Metabolism in Depression: A Narrative Review with a Focus on Serotonin and Kynurenine Pathways. Int J Mol Sci 2022, 23. [CrossRef]
- Fang, X.; Wu, Y.; Dai, Y.; Xiao, H.; Li, S.; Chen, X.; Yuan, M.; Guo, Y.; Ma, L.; Lin, D.; et al. In Situ Recovery of Serotonin Synthesis by a Tryptophan Hydroxylase-Like Nanozyme for the Treatment of Depression. J Am Chem Soc 2025, 147, 9111-9121. [CrossRef]
- Wang, D.; Wu, J.; Zhu, P.; Xie, H.; Lu, L.; Bai, W.; Pan, W.; Shi, R.; Ye, J.; Xia, B.; et al. Tryptophan-rich diet ameliorates chronic unpredictable mild stress induced depression- and anxiety-like behavior in mice: The potential involvement of gut-brain axis. Food Res Int 2022, 157, 111289. [CrossRef]
- Huang, Y.; Zhao, M.; Chen, X.; Zhang, R.; Le, A.; Hong, M.; Zhang, Y.; Jia, L.; Zang, W.; Jiang, C.; et al. Tryptophan Metabolism in Central Nervous System Diseases: Pathophysiology and Potential Therapeutic Strategies. Aging Dis 2023, 14, 858-878. [CrossRef]
- Zhang, Z.W.; Gao, C.S.; Zhang, H.; Yang, J.; Wang, Y.P.; Pan, L.B.; Yu, H.; He, C.Y.; Luo, H.B.; Zhao, Z.X.; et al. Morinda officinalis oligosaccharides increase serotonin in the brain and ameliorate depression via promoting 5-hydroxytryptophan production in the gut microbiota. Acta Pharm Sin B 2022, 12, 3298-3312. [CrossRef]
- Figueiredo Godoy, A.C.; Frota, F.F.; Araújo, L.P.; Valenti, V.E.; Pereira, E.; Detregiachi, C.R.P.; Galhardi, C.M.; Caracio, F.C.; Haber, R.S.A.; Fornari Laurindo, L.; et al. Neuroinflammation and Natural Antidepressants: Balancing Fire with Flora. Biomedicines 2025, 13. [CrossRef]
- Cowen, P.J. SSRIs in the Treatment of Depression: A Pharmacological CUL-DE-SAC? Curr Top Behav Neurosci 2024, 66, 1-19. [CrossRef]
- Gabriel, F.C.; de Melo, D.O.; Fráguas, R.; Leite-Santos, N.C.; Mantovani da Silva, R.A.; Ribeiro, E. Pharmacological treatment of depression: A systematic review comparing clinical practice guideline recommendations. PLoS One 2020, 15, e0231700. [CrossRef]
- Stachowicz, K.; Sowa-Kućma, M. The treatment of depression - searching for new ideas. Front Pharmacol 2022, 13, 988648. [CrossRef]
- Dudek, D.; Chrobak, A.A.; Krupa, A.J.; Gorostowicz, A.; Gerlich, A.; Juryk, A.; Siwek, M. TED-trazodone effectiveness in depression: a naturalistic study of the effeciveness of trazodone in extended release formulation compared to SSRIs in patients with a major depressive disorder. Front Pharmacol 2023, 14, 1296639. [CrossRef]
- Srifuengfung, M.; Pennington, B.R.T.; Lenze, E.J. Optimizing treatment for older adults with depression. Ther Adv Psychopharmacol 2023, 13, 20451253231212327. [CrossRef]
- Zhao, Y.; Xu, D.; Wang, J.; Zhou, D.; Liu, A.; Sun, Y.; Yuan, Y.; Li, J.; Guo, W. The pharmacological mechanism of chaihu-jia-longgu-muli-tang for treating depression: integrated meta-analysis and network pharmacology analysis. Front Pharmacol 2023, 14, 1257617. [CrossRef]
- Zhao, X.; Zhang, H.; Wu, Y.; Yu, C. The efficacy and safety of St. John's wort extract in depression therapy compared to SSRIs in adults: A meta-analysis of randomized clinical trials. Adv Clin Exp Med 2023, 32, 151-161. [CrossRef]
- Zhang, Z.; Cai, X.; Liang, Y.; Zhang, R.; Liu, X.; Lu, L.; Huang, Y. Electroacupuncture as a rapid-onset and safer complementary therapy for depression: A systematic review and meta-analysis. Front Psychiatry 2022, 13, 1012606. [CrossRef]
- Shafiee, A.; Jafarabady, K.; Seighali, N.; Mohammadi, I.; Rajai Firouz Abadi, S.; Abhari, F.S.; Bakhtiyari, M. Effect of Saffron Versus Selective Serotonin Reuptake Inhibitors (SSRIs) in Treatment of Depression and Anxiety: A Meta-analysis of Randomized Controlled Trials. Nutr Rev 2025, 83, e751-e761. [CrossRef]
- Zhichao, H.; Ching, L.W.; Huijuan, L.; Liang, Y.; Zhiyu, W.; Weiyang, H.; Zhaoxiang, B.; Linda, Z.L.D. A network meta-analysis on the effectiveness and safety of acupuncture in treating patients with major depressive disorder. Sci Rep 2021, 11, 10384. [CrossRef]
- Nitzan, K.; David, D.; Franko, M.; Toledano, R.; Fidelman, S.; Tenenbaum, Y.S.; Blonder, M.; Armoza-Eilat, S.; Shamir, A.; Rehavi, M.; et al. Anxiolytic and antidepressants' effect of Crataegus pinnatifida (Shan Zha): biochemical mechanisms. Transl Psychiatry 2022, 12, 208. [CrossRef]
- Di Sotto, A.; Vitalone, A.; Di Giacomo, S. Plant-Derived Nutraceuticals and Immune System Modulation: An Evidence-Based Overview. Vaccines (Basel) 2020, 8. [CrossRef]
- Vrânceanu, M.; Galimberti, D.; Banc, R.; Dragoş, O.; Cozma-Petruţ, A.; Hegheş, S.C.; Voştinaru, O.; Cuciureanu, M.; Stroia, C.M.; Miere, D.; et al. The Anticancer Potential of Plant-Derived Nutraceuticals via the Modulation of Gene Expression. Plants (Basel) 2022, 11. [CrossRef]
- Xiong, B.; Wang, H.; Song, Y.X.; Lan, W.Y.; Li, J.; Wang, F. Natural saponins and macrophage polarization: Mechanistic insights and therapeutic perspectives in disease management. Front Pharmacol 2025, 16, 1584035. [CrossRef]
- Wang, T.; Song, Y.; Ai, Z.; Liu, Y.; Li, H.; Xu, W.; Chen, L.; Zhu, G.; Yang, M.; Su, D. Pulsatilla chinensis saponins ameliorated murine depression by inhibiting intestinal inflammation mediated IDO1 overexpression and rebalancing tryptophan metabolism. Phytomedicine 2023, 116, 154852. [CrossRef]
- Liaqat, H.; Parveen, A.; Kim, S.Y. Neuroprotective Natural Products' Regulatory Effects on Depression via Gut-Brain Axis Targeting Tryptophan. Nutrients 2022, 14. [CrossRef]
- Ahmad, S.; Azhar, A.; Tikmani, P.; Rafique, H.; Khan, A.; Mesiya, H.; Saeed, H. A randomized clinical trial to test efficacy of chamomile and saffron for neuroprotective and anti-inflammatory responses in depressive patients. Heliyon 2022, 8, e10774. [CrossRef]
- Yang, Y.H.; Li, C.X.; Zhang, R.B.; Shen, Y.; Xu, X.J.; Yu, Q.M. A review of the pharmacological action and mechanism of natural plant polysaccharides in depression. Front Pharmacol 2024, 15, 1348019. [CrossRef]
- Tartt, A.N.; Mariani, M.B.; Hen, R.; Mann, J.J.; Boldrini, M. Dysregulation of adult hippocampal neuroplasticity in major depression: pathogenesis and therapeutic implications. Mol Psychiatry 2022, 27, 2689-2699. [CrossRef]
- Colucci-D'Amato, L.; Speranza, L.; Volpicelli, F. Neurotrophic Factor BDNF, Physiological Functions and Therapeutic Potential in Depression, Neurodegeneration and Brain Cancer. Int J Mol Sci 2020, 21. [CrossRef]
- Sun, X.; Zu, Y.; Li, X.; Zhao, S.; Sun, X.; Li, L.; Zhang, X.; Wang, W.; Liang, Y.; Wang, W.; et al. Corticosterone-induced Hippocampal 5-HT Responses were Muted in Depressive-like State. ACS Chem Neurosci 2021, 12, 845-856. [CrossRef]
- Lee, M.T.; Peng, W.H.; Kan, H.W.; Wu, C.C.; Wang, D.W.; Ho, Y.C. Neurobiology of Depression: Chronic Stress Alters the Glutamatergic System in the Brain-Focusing on AMPA Receptor. Biomedicines 2022, 10. [CrossRef]
- He, J.G.; Zhou, H.Y.; Wang, F.; Chen, J.G. Dysfunction of Glutamatergic Synaptic Transmission in Depression: Focus on AMPA Receptor Trafficking. Biol Psychiatry Glob Open Sci 2023, 3, 187-196. [CrossRef]
- Moncrieff, J.; Cooper, R.E.; Stockmann, T.; Amendola, S.; Hengartner, M.P.; Horowitz, M.A. The serotonin theory of depression: a systematic umbrella review of the evidence. Molecular Psychiatry 2023, 28, 3243-3256. [CrossRef]
- Correia, A.S.; Cardoso, A.; Vale, N. Oxidative Stress in Depression: The Link with the Stress Response, Neuroinflammation, Serotonin, Neurogenesis and Synaptic Plasticity. Antioxidants (Basel) 2023, 12. [CrossRef]
- Cui, Y.; Cao, K.; Lin, H.; Cui, S.; Shen, C.; Wen, W.; Mo, H.; Dong, Z.; Bai, S.; Yang, L.; et al. Early-Life Stress Induces Depression-Like Behavior and Synaptic-Plasticity Changes in a Maternal Separation Rat Model: Gender Difference and Metabolomics Study. Front Pharmacol 2020, 11, 102. [CrossRef]
- Yan, Y.; Xu, X.; Chen, R.; Wu, S.; Yang, Z.; Wang, H.; Zhang, T. Down-regulation of MST1 in hippocampus protects against stress-induced depression-like behaviours and synaptic plasticity impairments. Brain Behav Immun 2021, 94, 196-209. [CrossRef]
- Aleksandrova, L.R.; Phillips, A.G. Neuroplasticity as a convergent mechanism of ketamine and classical psychedelics. Trends Pharmacol Sci 2021, 42, 929-942. [CrossRef]
- Brown, K.A.; Gould, T.D. Targeting metaplasticity mechanisms to promote sustained antidepressant actions. Mol Psychiatry 2024, 29, 1114-1127. [CrossRef]
- Tanaka, M.; Battaglia, S. Dualistic Dynamics in Neuropsychiatry: From Monoaminergic Modulators to Multiscale Biomarker Maps. Biomedicines 2025, 13. [CrossRef]
- Li, B.; Yang, W.; Ge, T.; Wang, Y.; Cui, R. Stress induced microglial activation contributes to depression. Pharmacol Res 2022, 179, 106145. [CrossRef]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. The semantics of microglia activation: neuroinflammation, homeostasis, and stress. J Neuroinflammation 2021, 18, 258. [CrossRef]
- Wang, H.; He, Y.; Sun, Z.; Ren, S.; Liu, M.; Wang, G.; Yang, J. Microglia in depression: an overview of microglia in the pathogenesis and treatment of depression. J Neuroinflammation 2022, 19, 132. [CrossRef]
- Kokkosis, A.G.; Madeira, M.M.; Hage, Z.; Valais, K.; Koliatsis, D.; Resutov, E.; Tsirka, S.E. Chronic psychosocial stress triggers microglial-/macrophage-induced inflammatory responses leading to neuronal dysfunction and depressive-related behavior. Glia 2024, 72, 111-132. [CrossRef]
- Sugama, S.; Kakinuma, Y. Stress and brain immunity: Microglial homeostasis through hypothalamus-pituitary-adrenal gland axis and sympathetic nervous system. Brain Behav Immun Health 2020, 7, 100111. [CrossRef]
- Picard, K.; Bisht, K.; Poggini, S.; Garofalo, S.; Golia, M.T.; Basilico, B.; Abdallah, F.; Ciano Albanese, N.; Amrein, I.; Vernoux, N.; et al. Microglial-glucocorticoid receptor depletion alters the response of hippocampal microglia and neurons in a chronic unpredictable mild stress paradigm in female mice. Brain Behav Immun 2021, 97, 423-439. [CrossRef]
- Fornari Laurindo, L.; Aparecido Dias, J.; Cressoni Araújo, A.; Torres Pomini, K.; Machado Galhardi, C.; Rucco Penteado Detregiachi, C.; Santos de Argollo Haber, L.; Donizeti Roque, D.; Dib Bechara, M.; Vialogo Marques de Castro, M.; et al. Immunological dimensions of neuroinflammation and microglial activation: exploring innovative immunomodulatory approaches to mitigate neuroinflammatory progression. Front Immunol 2023, 14, 1305933. [CrossRef]
- Asveda, T.; Talwar, P.; Ravanan, P. Exploring microglia and their phenomenal concatenation of stress responses in neurodegenerative disorders. Life Sci 2023, 328, 121920. [CrossRef]
- Biswas, K. Microglia mediated neuroinflammation in neurodegenerative diseases: A review on the cell signaling pathways involved in microglial activation. J Neuroimmunol 2023, 383, 578180. [CrossRef]
- Bolton, J.L.; Short, A.K.; Othy, S.; Kooiker, C.L.; Shao, M.; Gunn, B.G.; Beck, J.; Bai, X.; Law, S.M.; Savage, J.C.; et al. Early stress-induced impaired microglial pruning of excitatory synapses on immature CRH-expressing neurons provokes aberrant adult stress responses. Cell Rep 2022, 38, 110600. [CrossRef]
- Reemst, K.; Kracht, L.; Kotah, J.M.; Rahimian, R.; van Irsen, A.A.; Congrains Sotomayor, G.; Verboon, L.N.; Brouwer, N.; Simard, S.; Turecki, G. Early-life stress lastingly impacts microglial transcriptome and function under basal and immune-challenged conditions. Translational psychiatry 2022, 12, 507.
- Smail, M.A.; Lenz, K.M. Developmental functions of microglia: Impact of psychosocial and physiological early life stress. Neuropharmacology 2024, 258, 110084. [CrossRef]
- Shen, S.Y.; Liang, L.F.; Shi, T.L.; Shen, Z.Q.; Yin, S.Y.; Zhang, J.R.; Li, W.; Mi, W.L.; Wang, Y.Q.; Zhang, Y.Q.; et al. Microglia-Derived Interleukin-6 Triggers Astrocyte Apoptosis in the Hippocampus and Mediates Depression-Like Behavior. Adv Sci (Weinh) 2025, 12, e2412556. [CrossRef]
- Klawonn, A.M.; Fritz, M.; Castany, S.; Pignatelli, M.; Canal, C.; Similä, F.; Tejeda, H.A.; Levinsson, J.; Jaarola, M.; Jakobsson, J.; et al. Microglial activation elicits a negative affective state through prostaglandin-mediated modulation of striatal neurons. Immunity 2021, 54, 225-234.e226. [CrossRef]
- Wang, Y.L.; Wu, H.R.; Zhang, S.S.; Xiao, H.L.; Yu, J.; Ma, Y.Y.; Zhang, Y.D.; Liu, Q. Catalpol ameliorates depressive-like behaviors in CUMS mice via oxidative stress-mediated NLRP3 inflammasome and neuroinflammation. Transl Psychiatry 2021, 11, 353. [CrossRef]
- Xia, C.Y.; Guo, Y.X.; Lian, W.W.; Yan, Y.; Ma, B.Z.; Cheng, Y.C.; Xu, J.K.; He, J.; Zhang, W.K. The NLRP3 inflammasome in depression: Potential mechanisms and therapies. Pharmacol Res 2023, 187, 106625. [CrossRef]
- Wang, L.; Hauenstein, A.V. The NLRP3 inflammasome: Mechanism of action, role in disease and therapies. Mol Aspects Med 2020, 76, 100889. [CrossRef]
- Cattaneo, A.; Ferrari, C.; Turner, L.; Mariani, N.; Enache, D.; Hastings, C.; Kose, M.; Lombardo, G.; McLaughlin, A.P.; Nettis, M.A.; et al. Whole-blood expression of inflammasome- and glucocorticoid-related mRNAs correctly separates treatment-resistant depressed patients from drug-free and responsive patients in the BIODEP study. Transl Psychiatry 2020, 10, 232. [CrossRef]
- Xu, Y.; Liang, J.; Sun, Y.; Zhang, Y.; Shan, F.; Ge, J.; Xia, Q. Serum cytokines-based biomarkers in the diagnosis and monitoring of therapeutic response in patients with major depressive disorder. Int Immunopharmacol 2023, 118, 110108. [CrossRef]
- Bodnár, K.; Hermán, L.; Zsigmond, R.; Réthelyi, J. Investigation of cytokine imbalance in schizophrenia, assessment of the possible role of serum cytokine levels in predicting treatment response, prognosis and psychotic relapses. European Psychiatry 2024, 67, S685-S685.
- Zhao, Z.; Wang, Y.; Zhou, R.; Li, Y.; Gao, Y.; Tu, D.; Wilson, B.; Song, S.; Feng, J.; Hong, J.S.; et al. A novel role of NLRP3-generated IL-1β in the acute-chronic transition of peripheral lipopolysaccharide-elicited neuroinflammation: implications for sepsis-associated neurodegeneration. J Neuroinflammation 2020, 17, 64. [CrossRef]
- Moraes, C.A.; Hottz, E.D.; Dos Santos Ornellas, D.; Adesse, D.; de Azevedo, C.T.; d'Avila, J.C.; Zaverucha-do-Valle, C.; Maron-Gutierrez, T.; Barbosa, H.S.; Bozza, P.T.; et al. Microglial NLRP3 Inflammasome Induces Excitatory Synaptic Loss Through IL-1β-Enriched Microvesicle Release: Implications for Sepsis-Associated Encephalopathy. Mol Neurobiol 2023, 60, 481-494. [CrossRef]
- Xu, W.; Huang, Y.; Zhou, R. NLRP3 inflammasome in neuroinflammation and central nervous system diseases. Cell Mol Immunol 2025, 22, 341-355. [CrossRef]
- Lu, S.; Wu, C.; Jia, L.; Fang, Z.; Lu, J.; Mou, T.; Hu, S.; He, H.; Huang, M.; Xu, Y. Increased plasma levels of IL-6 are associated with striatal structural atrophy in major depressive disorder patients with anhedonia. Front Psychiatry 2022, 13, 1016735. [CrossRef]
- Sanders, A.F.P.; Tirado, B.; Seider, N.A.; Triplett, R.L.; Lean, R.E.; Neil, J.J.; Miller, J.P.; Tillman, R.; Smyser, T.A.; Barch, D.M.; et al. Prenatal exposure to maternal disadvantage-related inflammatory biomarkers: associations with neonatal white matter microstructure. Transl Psychiatry 2024, 14, 72. [CrossRef]
- Lim, J.; Sohn, H.; Kwon, M.S.; Kim, B. White Matter Alterations Associated with Pro-inflammatory Cytokines in Patients with Major Depressive Disorder. Clin Psychopharmacol Neurosci 2021, 19, 449-458. [CrossRef]
- Yin, R.; Zhang, K.; Li, Y.; Tang, Z.; Zheng, R.; Ma, Y.; Chen, Z.; Lei, N.; Xiong, L.; Guo, P.; et al. Lipopolysaccharide-induced depression-like model in mice: meta-analysis and systematic evaluation. Front Immunol 2023, 14, 1181973. [CrossRef]
- Lasselin, J.; Schedlowski, M.; Karshikoff, B.; Engler, H.; Lekander, M.; Konsman, J.P. Comparison of bacterial lipopolysaccharide-induced sickness behavior in rodents and humans: Relevance for symptoms of anxiety and depression. Neurosci Biobehav Rev 2020, 115, 15-24. [CrossRef]
- Su, J.; Pan, Y.W.; Wang, S.Q.; Li, X.Z.; Huang, F.; Ma, S.P. Saikosaponin-d attenuated lipopolysaccharide-induced depressive-like behaviors via inhibiting microglia activation and neuroinflammation. Int Immunopharmacol 2020, 80, 106181. [CrossRef]
- Tomaz, V.S.; Chaves Filho, A.J.M.; Cordeiro, R.C.; Jucá, P.M.; Soares, M.V.R.; Barroso, P.N.; Cristino, L.M.F.; Jiang, W.; Teixeira, A.L.; de Lucena, D.F.; et al. Antidepressants of different classes cause distinct behavioral and brain pro- and anti-inflammatory changes in mice submitted to an inflammatory model of depression. J Affect Disord 2020, 268, 188-200. [CrossRef]
- Yi, S.S.; Zou, J.J.; Meng, L.; Chen, H.M.; Hong, Z.Q.; Liu, X.F.; Farooq, U.; Chen, M.X.; Lin, Z.R.; Zhou, W.; et al. Ultrasound Stimulation of Prefrontal Cortex Improves Lipopolysaccharide-Induced Depressive-Like Behaviors in Mice. Front Psychiatry 2022, 13, 864481. [CrossRef]
- Kappelmann, N.; Lewis, G.; Dantzer, R.; Jones, P.B.; Khandaker, G.M. Antidepressant activity of anti-cytokine treatment: a systematic review and meta-analysis of clinical trials of chronic inflammatory conditions. Mol Psychiatry 2018, 23, 335-343. [CrossRef]
- Zhang, M.; Wang, X.L.; Shi, H.; Meng, L.Q.; Quan, H.F.; Yan, L.; Yang, H.F.; Peng, X.D. Betaine Inhibits NLRP3 Inflammasome Hyperactivation and Regulates Microglial M1/M2 Phenotypic Differentiation, Thereby Attenuating Lipopolysaccharide-Induced Depression-Like Behavior. J Immunol Res 2022, 2022, 9313436. [CrossRef]
- Planchez, B.; Lagunas, N.; Le Guisquet, A.M.; Legrand, M.; Surget, A.; Hen, R.; Belzung, C. Increasing Adult Hippocampal Neurogenesis Promotes Resilience in a Mouse Model of Depression. Cells 2021, 10. [CrossRef]
- Jones, K.L.; Zhou, M.; Jhaveri, D.J. Dissecting the role of adult hippocampal neurogenesis towards resilience versus susceptibility to stress-related mood disorders. NPJ Sci Learn 2022, 7, 16. [CrossRef]
- Fang, S.; Wu, Z.; Guo, Y.; Zhu, W.; Wan, C.; Yuan, N.; Chen, J.; Hao, W.; Mo, X.; Guo, X.; et al. Roles of microglia in adult hippocampal neurogenesis in depression and their therapeutics. Front Immunol 2023, 14, 1193053. [CrossRef]
- Tang, C.; Wang, Q.; Shen, J.; Wang, C.; Ding, H.; Wen, S.; Yang, F.; Jiao, R.; Wu, X.; Li, J.; et al. Neuron stem cell NLRP6 sustains hippocampal neurogenesis to resist stress-induced depression. Acta Pharm Sin B 2023, 13, 2017-2038. [CrossRef]
- Zhang, S.Q.; Deng, Q.; Zhu, Q.; Hu, Z.L.; Long, L.H.; Wu, P.F.; He, J.G.; Chen, H.S.; Yue, Z.; Lu, J.H.; et al. Cell type-specific NRBF2 orchestrates autophagic flux and adult hippocampal neurogenesis in chronic stress-induced depression. Cell Discov 2023, 9, 90. [CrossRef]
- Zhang, K.; Wang, F.; Zhai, M.; He, M.; Hu, Y.; Feng, L.; Li, Y.; Yang, J.; Wu, C. Hyperactive neuronal autophagy depletes BDNF and impairs adult hippocampal neurogenesis in a corticosterone-induced mouse model of depression. Theranostics 2023, 13, 1059.
- Du Preez, A.; Onorato, D.; Eiben, I.; Musaelyan, K.; Egeland, M.; Zunszain, P.A.; Fernandes, C.; Thuret, S.; Pariante, C.M. Chronic stress followed by social isolation promotes depressive-like behaviour, alters microglial and astrocyte biology and reduces hippocampal neurogenesis in male mice. Brain Behav Immun 2021, 91, 24-47. [CrossRef]
- Parul; Mishra, A.; Singh, S.; Singh, S.; Tiwari, V.; Chaturvedi, S.; Wahajuddin, M.; Palit, G.; Shukla, S. Chronic unpredictable stress negatively regulates hippocampal neurogenesis and promote anxious depression-like behavior via upregulating apoptosis and inflammatory signals in adult rats. Brain Res Bull 2021, 172, 164-179. [CrossRef]
- Zhang, J.; Rong, P.; Zhang, L.; He, H.; Zhou, T.; Fan, Y.; Mo, L.; Zhao, Q.; Han, Y.; Li, S.; et al. IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis. Sci Adv 2021, 7. [CrossRef]
- Zhang, H.; Wei, M.; Sun, N.; Wang, H.; Fan, H. Melatonin attenuates chronic stress-induced hippocampal inflammatory response and apoptosis by inhibiting ADAM17/TNF-α axis. Food Chem Toxicol 2022, 169, 113441. [CrossRef]
- Li, Y.; Wang, L.; Wang, P.; Fan, C.; Zhang, P.; Shen, J.; Yu, S.Y. Ginsenoside-Rg1 Rescues Stress-Induced Depression-Like Behaviors via Suppression of Oxidative Stress and Neural Inflammation in Rats. Oxid Med Cell Longev 2020, 2020, 2325391. [CrossRef]
- Li, J.; Gao, W.; Zhao, Z.; Li, Y.; Yang, L.; Wei, W.; Ren, F.; Li, Y.; Yu, Y.; Duan, W.; et al. Ginsenoside Rg1 Reduced Microglial Activation and Mitochondrial Dysfunction to Alleviate Depression-Like Behaviour Via the GAS5/EZH2/SOCS3/NRF2 Axis. Mol Neurobiol 2022, 59, 2855-2873. [CrossRef]
- Amanollahi, M.; Jameie, M.; Heidari, A.; Rezaei, N. The Dialogue Between Neuroinflammation and Adult Neurogenesis: Mechanisms Involved and Alterations in Neurological Diseases. Mol Neurobiol 2023, 60, 923-959. [CrossRef]
- Zhang, J.; He, H.; Qiao, Y.; Zhou, T.; He, H.; Yi, S.; Zhang, L.; Mo, L.; Li, Y.; Jiang, W.; et al. Priming of microglia with IFN-γ impairs adult hippocampal neurogenesis and leads to depression-like behaviors and cognitive defects. Glia 2020, 68, 2674-2692. [CrossRef]
- Zhao, R.; Tian, X.; Xu, H.; Wang, Y.; Lin, J.; Wang, B. Aerobic Exercise Restores Hippocampal Neurogenesis and Cognitive Function by Decreasing Microglia Inflammasome Formation Through Irisin/NLRP3 Pathway. Aging Cell 2025, 24, e70061. [CrossRef]
- Wang, W.; Wang, L.; Wang, L.; Li, Y.; Lan, T.; Wang, C.; Chen, X.; Chen, S.; Yu, S. Ginsenoside-Rg1 synergized with voluntary running exercise protects against glial activation and dysregulation of neuronal plasticity in depression. Food Funct 2023, 14, 7222-7239. [CrossRef]
- de Lima, E.P.; Tanaka, M.; Lamas, C.B.; Quesada, K.; Detregiachi, C.R.P.; Araújo, A.C.; Guiguer, E.L.; Catharin, V.; de Castro, M.V.M.; Junior, E.B.; et al. Vascular Impairment, Muscle Atrophy, and Cognitive Decline: Critical Age-Related Conditions. Biomedicines 2024, 12. [CrossRef]
- Liloia, D.; Zamfira, D.A.; Tanaka, M.; Manuello, J.; Crocetta, A.; Keller, R.; Cozzolino, M.; Duca, S.; Cauda, F.; Costa, T. Disentangling the role of gray matter volume and concentration in autism spectrum disorder: A meta-analytic investigation of 25 years of voxel-based morphometry research. Neurosci Biobehav Rev 2024, 164, 105791. [CrossRef]
- Müller, L.; Di Benedetto, S. Neuroimmune crosstalk in chronic neuroinflammation: microglial interactions and immune modulation. Front Cell Neurosci 2025, 19, 1575022. [CrossRef]
- Navabi, S.P.; Badreh, F.; Khombi Shooshtari, M.; Hajipour, S.; Moradi Vastegani, S.; Khoshnam, S.E. Microglia-induced neuroinflammation in hippocampal neurogenesis following traumatic brain injury. Heliyon 2024, 10, e35869. [CrossRef]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: the roles of microglia and astrocytes. Transl Neurodegener 2020, 9, 42. [CrossRef]
- Sanchez-Molina, P.; Almolda, B.; Giménez-Llort, L.; González, B.; Castellano, B. Chronic IL-10 overproduction disrupts microglia-neuron dialogue similar to aging, resulting in impaired hippocampal neurogenesis and spatial memory. Brain Behav Immun 2022, 101, 231-245. [CrossRef]
- Izzy, S.; Yahya, T.; Albastaki, O.; Abou-El-Hassan, H.; Aronchik, M.; Cao, T.; De Oliveira, M.G.; Lu, K.J.; Moreira, T.G.; da Silva, P.; et al. Nasal anti-CD3 monoclonal antibody ameliorates traumatic brain injury, enhances microglial phagocytosis and reduces neuroinflammation via IL-10-dependent T(reg)-microglia crosstalk. Nat Neurosci 2025, 28, 499-516. [CrossRef]
- Witcher, K.G.; Bray, C.E.; Chunchai, T.; Zhao, F.; O'Neil, S.M.; Gordillo, A.J.; Campbell, W.A.; McKim, D.B.; Liu, X.; Dziabis, J.E.; et al. Traumatic Brain Injury Causes Chronic Cortical Inflammation and Neuronal Dysfunction Mediated by Microglia. J Neurosci 2021, 41, 1597-1616. [CrossRef]
- Gao, Y.; Liu, X.; Gu, Y. Evaluation of serum brain-derived neurotrophic factor, IL-6, IL-10, and TNF-a cognitive function, and sleep quality in elderly patients with major depressive disorder and somatic symptoms. J Med Biochem 2025, 44, 568-575. [CrossRef]
- Pan, X.; Guo, A.; Guan, K.; Chen, C.; Xu, S.; Tang, Y.; Li, X.; Huang, Z. Lactobacillus rhamnosus GG attenuates depression-like behaviour and cognitive deficits in chronic ethanol exposure mice by down-regulating systemic inflammatory factors. Addict Biol 2024, 29, e13445. [CrossRef]
- Zhang, J.C.; Yao, W.; Hashimoto, K. Brain-derived Neurotrophic Factor (BDNF)-TrkB Signaling in Inflammation-related Depression and Potential Therapeutic Targets. Curr Neuropharmacol 2016, 14, 721-731. [CrossRef]
- Calabrese, F.; Rossetti, A.C.; Racagni, G.; Gass, P.; Riva, M.A.; Molteni, R. Brain-derived neurotrophic factor: a bridge between inflammation and neuroplasticity. Front Cell Neurosci 2014, 8, 430. [CrossRef]
- Carniel, B.P.; da Rocha, N.S. Brain-derived neurotrophic factor (BDNF) and inflammatory markers: Perspectives for the management of depression. Prog Neuropsychopharmacol Biol Psychiatry 2021, 108, 110151. [CrossRef]
- Liu, Q.; Zhang, J.; Xiao, C.; Su, D.; Li, L.; Yang, C.; Zhao, Z.; Jiang, W.; You, Z.; Zhou, T. Akebia saponin D protects hippocampal neurogenesis from microglia-mediated inflammation and ameliorates depressive-like behaviors and cognitive impairment in mice through the PI3K-Akt pathway. Front Pharmacol 2022, 13, 927419. [CrossRef]
- Zhang, Q.; Zhao, W.; Yun, Y.; Ma, T.; An, H.; Fan, N.; Wang, J.; Wang, Z.; Yang, F. Multiomics analysis reveals aberrant tryptophan-kynurenine metabolism and immunity linked gut microbiota with cognitive impairment in major depressive disorder. J Affect Disord 2025, 373, 273-283. [CrossRef]
- de Lima, E.P.; Laurindo, L.F.; Catharin, V.C.S.; Direito, R.; Tanaka, M.; Jasmin Santos German, I.; Lamas, C.B.; Guiguer, E.L.; Araújo, A.C.; Fiorini, A.M.R.; et al. Polyphenols, Alkaloids, and Terpenoids Against Neurodegeneration: Evaluating the Neuroprotective Effects of Phytocompounds Through a Comprehensive Review of the Current Evidence. Metabolites 2025, 15. [CrossRef]
- Barbalho, S.M.; Laurindo, L.F.; de Oliveira Zanuso, B.; da Silva, R.M.S.; Gallerani Caglioni, L.; Nunes Junqueira de Moraes, V.B.F.; Fornari Laurindo, L.; Dogani Rodrigues, V.; da Silva Camarinha Oliveira, J.; Beluce, M.E.; et al. AdipoRon's Impact on Alzheimer's Disease-A Systematic Review and Meta-Analysis. Int J Mol Sci 2025, 26. [CrossRef]
- Salminen, A. Role of indoleamine 2,3-dioxygenase 1 (IDO1) and kynurenine pathway in the regulation of the aging process. Ageing Res Rev 2022, 75, 101573. [CrossRef]
- Tanaka, M.; Tóth, F.; Polyák, H.; Szabó, Á.; Mándi, Y.; Vécsei, L. Immune Influencers in Action: Metabolites and Enzymes of the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9. [CrossRef]
- Dawood, S.; Bano, S.; Badawy, A.A. Inflammation and serotonin deficiency in major depressive disorder: molecular docking of antidepressant and anti-inflammatory drugs to tryptophan and indoleamine 2,3-dioxygenases. Biosci Rep 2022, 42. [CrossRef]
- Tiwari, S.; Paramanik, V. Role of Probiotics in Depression: Connecting Dots of Gut-Brain-Axis Through Hypothalamic-Pituitary Adrenal Axis and Tryptophan/Kynurenic Pathway involving Indoleamine-2,3-dioxygenase. Mol Neurobiol 2025, 62, 7230-7241. [CrossRef]
- Sublette, M.E.; Postolache, T.T. Neuroinflammation and depression: the role of indoleamine 2,3-dioxygenase (IDO) as a molecular pathway. Psychosom Med 2012, 74, 668-672. [CrossRef]
- Tanaka, M.; Battaglia, S. From Biomarkers to Behavior: Mapping the Neuroimmune Web of Pain, Mood, and Memory. 2025, 13, 2226.
- Stone, T.W.; Williams, R.O. Tryptophan metabolism as a 'reflex' feature of neuroimmune communication: Sensor and effector functions for the indoleamine-2, 3-dioxygenase kynurenine pathway. J Neurochem 2024, 168, 3333-3357. [CrossRef]
- Juhász, L.; Spisák, K.; Szolnoki, B.Z.; Nászai, A.; Szabó, Á.; Rutai, A.; Tallósy, S.P.; Szabó, A.; Toldi, J.; Tanaka, M.; et al. The Power Struggle: Kynurenine Pathway Enzyme Knockouts and Brain Mitochondrial Respiration. J Neurochem 2025, 169, e70075. [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Szűcs, M.; Martos, D.; Takeda, K.; Ozaki, K.; Inoue, H.; Yamamoto, S.; Toldi, J.; et al. Oxidative and Excitatory Neurotoxic Stresses in CRISPR/Cas9-Induced Kynurenine Aminotransferase Knockout Mice: A Novel Model for Despair-Based Depression and Post-Traumatic Stress Disorder. Front Biosci (Landmark Ed) 2025, 30, 25706. [CrossRef]
- Fiore, A.; Murray, P.J. Tryptophan and indole metabolism in immune regulation. Curr Opin Immunol 2021, 70, 7-14. [CrossRef]
- Szabó, Á.; Galla, Z.; Spekker, E.; Martos, D.; Szűcs, M.; Fejes-Szabó, A.; Fehér, Á.; Takeda, K.; Ozaki, K.; Inoue, H.; et al. Behavioral Balance in Tryptophan Turmoil: Regional Metabolic Rewiring in Kynurenine Aminotransferase II Knockout Mice. Cells 2025, 14. [CrossRef]
- Alfaro-Rodríguez, A.; Reyes-Long, S.; Roldan-Valadez, E.; González-Torres, M.; Bonilla-Jaime, H.; Bandala, C.; Avila-Luna, A.; Bueno-Nava, A.; Cabrera-Ruiz, E.; Sanchez-Aparicio, P.; et al. Association of the Serotonin and Kynurenine Pathways as Possible Therapeutic Targets to Modulate Pain in Patients with Fibromyalgia. Pharmaceuticals (Basel) 2024, 17. [CrossRef]
- Hunt, C.; Macedo, E.C.T.; Suchting, R.; de Dios, C.; Cuellar Leal, V.A.; Soares, J.C.; Dantzer, R.; Teixeira, A.L.; Selvaraj, S. Effect of immune activation on the kynurenine pathway and depression symptoms - A systematic review and meta-analysis. Neurosci Biobehav Rev 2020, 118, 514-523. [CrossRef]
- Almulla, A.F.; Supasitthumrong, T.; Tunvirachaisakul, C.; Algon, A.A.A.; Al-Hakeim, H.K.; Maes, M. The tryptophan catabolite or kynurenine pathway in COVID-19 and critical COVID-19: a systematic review and meta-analysis. BMC Infect Dis 2022, 22, 615. [CrossRef]
- Li, Y.; Peng, Y.; Shen, Y.; Zhang, Y.; Liu, L.; Yang, X. Dietary polyphenols: regulate the advanced glycation end products-RAGE axis and the microbiota-gut-brain axis to prevent neurodegenerative diseases. Crit Rev Food Sci Nutr 2023, 63, 9816-9842. [CrossRef]
- Ghimire, S.; Cady, N.M.; Lehman, P.; Peterson, S.R.; Shahi, S.K.; Rashid, F.; Giri, S.; Mangalam, A.K. Dietary Isoflavones Alter Gut Microbiota and Lipopolysaccharide Biosynthesis to Reduce Inflammation. Gut Microbes 2022, 14, 2127446. [CrossRef]
- Winiarska-Mieczan, A.; Kwiecień, M.; Jachimowicz-Rogowska, K.; Donaldson, J.; Tomaszewska, E.; Baranowska-Wójcik, E. Anti-Inflammatory, Antioxidant, and Neuroprotective Effects of Polyphenols-Polyphenols as an Element of Diet Therapy in Depressive Disorders. Int J Mol Sci 2023, 24. [CrossRef]
- Tian, E.; Sharma, G.; Dai, C. Neuroprotective Properties of Berberine: Molecular Mechanisms and Clinical Implications. Antioxidants (Basel) 2023, 12. [CrossRef]
- Wang, L.; Wu, X.; Ma, Y.; Li, X.; Zhang, J.; Zhao, L. Supplementation with soy isoflavones alleviates depression-like behaviour via reshaping the gut microbiota structure. Food Funct 2021, 12, 4995-5006. [CrossRef]
- Chatterjee, A.; Kumar, S.; Roy Sarkar, S.; Halder, R.; Kumari, R.; Banerjee, S.; Sarkar, B. Dietary polyphenols represent a phytotherapeutic alternative for gut dysbiosis associated neurodegeneration: A systematic review. J Nutr Biochem 2024, 129, 109622. [CrossRef]
- Tayab, M.A.; Islam, M.N.; Chowdhury, K.A.A.; Tasnim, F.M. Targeting neuroinflammation by polyphenols: A promising therapeutic approach against inflammation-associated depression. Biomed Pharmacother 2022, 147, 112668. [CrossRef]
- Abbaszadeh, F.; Fakhri, S.; Khan, H. Targeting apoptosis and autophagy following spinal cord injury: Therapeutic approaches to polyphenols and candidate phytochemicals. Pharmacol Res 2020, 160, 105069. [CrossRef]
- Grifka-Walk, H.M.; Jenkins, B.R.; Kominsky, D.J. Amino Acid Trp: The Far Out Impacts of Host and Commensal Tryptophan Metabolism. Front Immunol 2021, 12, 653208. [CrossRef]
- Comai, S.; Bertazzo, A.; Brughera, M.; Crotti, S. Tryptophan in health and disease. Adv Clin Chem 2020, 95, 165-218. [CrossRef]
- Deng, Y.; Zhou, M.; Wang, J.; Yao, J.; Yu, J.; Liu, W.; Wu, L.; Wang, J.; Gao, R. Involvement of the microbiota-gut-brain axis in chronic restraint stress: disturbances of the kynurenine metabolic pathway in both the gut and brain. Gut Microbes 2021, 13, 1-16. [CrossRef]
- Tutakhail, A.; Boulet, L.; Khabil, S.; Nazari, Q.A.; Hamid, H.; Coudoré, F. Neuropathology of kynurenine pathway of tryptophan metabolism. Current pharmacology reports 2020, 6, 8-23.
- Höglund, E.; Øverli, Ø.; Winberg, S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front Endocrinol (Lausanne) 2019, 10, 158. [CrossRef]
- Fujigaki, H.; Yamamoto, Y.; Saito, K. L-Tryptophan-kynurenine pathway enzymes are therapeutic target for neuropsychiatric diseases: Focus on cell type differences. Neuropharmacology 2017, 112, 264-274. [CrossRef]
- Messaoud, A.; Mensi, R.; Douki, W.; Neffati, F.; Najjar, M.F.; Gobbi, G.; Valtorta, F.; Gaha, L.; Comai, S. Reduced peripheral availability of tryptophan and increased activation of the kynurenine pathway and cortisol correlate with major depression and suicide. World J Biol Psychiatry 2019, 20, 703-711. [CrossRef]
- Chivite, M.; Leal, E.; Míguez, J.M.; Cerdá-Reverter, J.M. Distribution of two isoforms of tryptophan hydroxylase in the brain of rainbow trout (Oncorhynchus mykiss). An in situ hybridization study. Brain Struct Funct 2021, 226, 2265-2278. [CrossRef]
- Maffei, M.E. 5-Hydroxytryptophan (5-HTP): Natural Occurrence, Analysis, Biosynthesis, Biotechnology, Physiology and Toxicology. Int J Mol Sci 2020, 22. [CrossRef]
- Xue, C.; Li, G.; Zheng, Q.; Gu, X.; Shi, Q.; Su, Y.; Chu, Q.; Yuan, X.; Bao, Z.; Lu, J.; et al. Tryptophan metabolism in health and disease. Cell Metab 2023, 35, 1304-1326. [CrossRef]
- Zakrocka, I.; Urbańska, E.M.; Załuska, W.; Kronbichler, A. Kynurenine Pathway after Kidney Transplantation: Friend or Foe? International Journal of Molecular Sciences 2024, 25, 9940.
- Perez-Castro, L.; Garcia, R.; Venkateswaran, N.; Barnes, S.; Conacci-Sorrell, M. Tryptophan and its metabolites in normal physiology and cancer etiology. The FEBS journal 2023, 290, 7-27.
- Tanaka, M.; Bohár, Z.; Vécsei, L. Are Kynurenines Accomplices or Principal Villains in Dementia? Maintenance of Kynurenine Metabolism. Molecules 2020, 25. [CrossRef]
- Fathi, M.; Vakili, K.; Yaghoobpoor, S.; Tavasol, A.; Jazi, K.; Mohamadkhani, A.; Klegeris, A.; McElhinney, A.; Mafi, Z.; Hajiesmaeili, M.; et al. Dynamic changes in kynurenine pathway metabolites in multiple sclerosis: A systematic review. Front Immunol 2022, 13, 1013784. [CrossRef]
- Ou, W.; Chen, Y.; Ju, Y.; Ma, M.; Qin, Y.; Bi, Y.; Liao, M.; Liu, B.; Liu, J.; Zhang, Y.; et al. The kynurenine pathway in major depressive disorder under different disease states: A systematic review and meta-analysis. J Affect Disord 2023, 339, 624-632. [CrossRef]
- Kozieł, K.; Urbanska, E.M. Kynurenine Pathway in Diabetes Mellitus-Novel Pharmacological Target? Cells 2023, 12. [CrossRef]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Redefining Roles: A Paradigm Shift in Tryptophan-Kynurenine Metabolism for Innovative Clinical Applications. Int J Mol Sci 2024, 25. [CrossRef]
- Skorobogatov, K.; De Picker, L.; Verkerk, R.; Coppens, V.; Leboyer, M.; Müller, N.; Morrens, M. Brain Versus Blood: A Systematic Review on the Concordance Between Peripheral and Central Kynurenine Pathway Measures in Psychiatric Disorders. Front Immunol 2021, 12, 716980. [CrossRef]
- Pocivavsek, A.; Schwarcz, R.; Erhardt, S. Neuroactive Kynurenines as Pharmacological Targets: New Experimental Tools and Exciting Therapeutic Opportunities. Pharmacol Rev 2024, 76, 978-1008. [CrossRef]
- Zhang, S.; Sakuma, M.; Deora, G.S.; Levy, C.W.; Klausing, A.; Breda, C.; Read, K.D.; Edlin, C.D.; Ross, B.P.; Wright Muelas, M.; et al. A brain-permeable inhibitor of the neurodegenerative disease target kynurenine 3-monooxygenase prevents accumulation of neurotoxic metabolites. Commun Biol 2019, 2, 271. [CrossRef]
- Jones, S.P.; Franco, N.F.; Varney, B.; Sundaram, G.; Brown, D.A.; de Bie, J.; Lim, C.K.; Guillemin, G.J.; Brew, B.J. Expression of the Kynurenine Pathway in Human Peripheral Blood Mononuclear Cells: Implications for Inflammatory and Neurodegenerative Disease. PLoS One 2015, 10, e0131389. [CrossRef]
- Sathyasaikumar, K.V.; Notarangelo, F.M.; Kelly, D.L.; Rowland, L.M.; Hare, S.M.; Chen, S.; Mo, C.; Buchanan, R.W.; Schwarcz, R. Tryptophan Challenge in Healthy Controls and People with Schizophrenia: Acute Effects on Plasma Levels of Kynurenine, Kynurenic Acid and 5-Hydroxyindoleacetic Acid. Pharmaceuticals (Basel) 2022, 15. [CrossRef]
- Hestad, K.; Alexander, J.; Rootwelt, H.; Aaseth, J.O. The Role of Tryptophan Dysmetabolism and Quinolinic Acid in Depressive and Neurodegenerative Diseases. Biomolecules 2022, 12. [CrossRef]
- Martos, D.; Tuka, B.; Tanaka, M.; Vécsei, L.; Telegdy, G. Memory Enhancement with Kynurenic Acid and Its Mechanisms in Neurotransmission. Biomedicines 2022, 10. [CrossRef]
- Tanaka, M.; Szatmári, I.; Vécsei, L. Quinoline Quest: Kynurenic Acid Strategies for Next-Generation Therapeutics via Rational Drug Design. Pharmaceuticals (Basel) 2025, 18. [CrossRef]
- Mor, A.; Tankiewicz-Kwedlo, A.; Krupa, A.; Pawlak, D. Role of Kynurenine Pathway in Oxidative Stress during Neurodegenerative Disorders. Cells 2021, 10. [CrossRef]
- Tanaka, M.; Vécsei, L. From Microbial Switches to Metabolic Sensors: Rewiring the Gut-Brain Kynurenine Circuit. Biomedicines 2025, 13. [CrossRef]
- Cheong, J.E.; Sun, L. Targeting the IDO1/TDO2-KYN-AhR Pathway for Cancer Immunotherapy - Challenges and Opportunities. Trends Pharmacol Sci 2018, 39, 307-325. [CrossRef]
- Ye, Z.; Yue, L.; Shi, J.; Shao, M.; Wu, T. Role of IDO and TDO in Cancers and Related Diseases and the Therapeutic Implications. J Cancer 2019, 10, 2771-2782. [CrossRef]
- Lu, Z.; Zhang, C.; Zhang, J.; Su, W.; Wang, G.; Wang, Z. The Kynurenine Pathway and Indole Pathway in Tryptophan Metabolism Influence Tumor Progression. Cancer Med 2025, 14, e70703. [CrossRef]
- Stone, T.W.; Williams, R.O. Interactions of IDO and the Kynurenine Pathway with Cell Transduction Systems and Metabolism at the Inflammation-Cancer Interface. Cancers (Basel) 2023, 15. [CrossRef]
- Lashgari, N.A.; Roudsari, N.M.; Shayan, M.; Niazi Shahraki, F.; Hosseini, Y.; Momtaz, S.; Abdolghaffari, A.H. IDO/Kynurenine; novel insight for treatment of inflammatory diseases. Cytokine 2023, 166, 156206. [CrossRef]
- Labadie, B.W.; Bao, R.; Luke, J.J. Reimagining IDO Pathway Inhibition in Cancer Immunotherapy via Downstream Focus on the Tryptophan-Kynurenine-Aryl Hydrocarbon Axis. Clin Cancer Res 2019, 25, 1462-1471. [CrossRef]
- Campesato, L.F.; Budhu, S.; Tchaicha, J.; Weng, C.H.; Gigoux, M.; Cohen, I.J.; Redmond, D.; Mangarin, L.; Pourpe, S.; Liu, C.; et al. Blockade of the AHR restricts a Treg-macrophage suppressive axis induced by L-Kynurenine. Nat Commun 2020, 11, 4011. [CrossRef]
- Fujiwara, Y.; Kato, S.; Nesline, M.K.; Conroy, J.M.; DePietro, P.; Pabla, S.; Kurzrock, R. Indoleamine 2,3-dioxygenase (IDO) inhibitors and cancer immunotherapy. Cancer Treat Rev 2022, 110, 102461. [CrossRef]
- Bertollo, A.G.; Mingoti, M.E.D.; Ignácio, Z.M. Neurobiological mechanisms in the kynurenine pathway and major depressive disorder. Rev Neurosci 2025, 36, 169-187. [CrossRef]
- Gong, X.; Chang, R.; Zou, J.; Tan, S.; Huang, Z. The role and mechanism of tryptophan - kynurenine metabolic pathway in depression. Rev Neurosci 2023, 34, 313-324. [CrossRef]
- Anderson, E.W.; Fishbein, J.; Hong, J.; Roeser, J.; Furie, R.A.; Aranow, C.; Volpe, B.T.; Diamond, B.; Mackay, M. Quinolinic acid, a kynurenine/tryptophan pathway metabolite, associates with impaired cognitive test performance in systemic lupus erythematosus. Lupus Sci Med 2021, 8. [CrossRef]
- Wang, L.; Feng, Z.; Zheng, T.; Dai, G.; Wang, M.; Zhou, L.; Zheng, Y.; Chen, G. Associations between the kynurenine pathway and the brain in patients with major depressive disorder-A systematic review of neuroimaging studies. Prog Neuropsychopharmacol Biol Psychiatry 2023, 121, 110675. [CrossRef]
- Ogyu, K.; Kubo, K.; Noda, Y.; Iwata, Y.; Tsugawa, S.; Omura, Y.; Wada, M.; Tarumi, R.; Plitman, E.; Moriguchi, S.; et al. Kynurenine pathway in depression: A systematic review and meta-analysis. Neurosci Biobehav Rev 2018, 90, 16-25. [CrossRef]
- Marx, W.; McGuinness, A.J.; Rocks, T.; Ruusunen, A.; Cleminson, J.; Walker, A.J.; Gomes-da-Costa, S.; Lane, M.; Sanches, M.; Diaz, A.P.; et al. The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: a meta-analysis of 101 studies. Mol Psychiatry 2021, 26, 4158-4178. [CrossRef]
- Joisten, N.; Ruas, J.L.; Braidy, N.; Guillemin, G.J.; Zimmer, P. The kynurenine pathway in chronic diseases: a compensatory mechanism or a driving force? Trends Mol Med 2021, 27, 946-954. [CrossRef]
- Martos, D.; Lőrinczi, B.; Szatmári, I.; Vécsei, L.; Tanaka, M. Decoupling Behavioral Domains via Kynurenic Acid Analog Optimization: Implications for Schizophrenia and Parkinson's Disease Therapeutics. Cells 2025, 14. [CrossRef]
- Inam, M.E.; Fernandes, B.S.; Salagre, E.; Grande, I.; Vieta, E.; Quevedo, J.; Zhao, Z. The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: a systematic review and meta-analysis of cerebrospinal fluid studies. Braz J Psychiatry 2023, 45, 343-355. [CrossRef]
- Tanaka, M.; Battaglia, S.; Liloia, D. Navigating Neurodegeneration: Integrating Biomarkers, Neuroinflammation, and Imaging in Parkinson's, Alzheimer's, and Motor Neuron Disorders. Biomedicines 2025, 13. [CrossRef]
- Więdłocha, M.; Marcinowicz, P.; Janoska-Jaździk, M.; Szulc, A. Gut microbiota, kynurenine pathway and mental disorders - Review. Prog Neuropsychopharmacol Biol Psychiatry 2021, 106, 110145. [CrossRef]
- Chen, C.Y.; Wang, Y.F.; Lei, L.; Zhang, Y. Impacts of microbiota and its metabolites through gut-brain axis on pathophysiology of major depressive disorder. Life Sci 2024, 351, 122815. [CrossRef]
- Cryan, J.F.; O'Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol Rev 2019, 99, 1877-2013. [CrossRef]
- Ohara, T.E.; Hsiao, E.Y. Microbiota-neuroepithelial signalling across the gut-brain axis. Nat Rev Microbiol 2025, 23, 371-384. [CrossRef]
- Zheng, Y.; Bonfili, L.; Wei, T.; Eleuteri, A.M. Understanding the Gut-Brain Axis and Its Therapeutic Implications for Neurodegenerative Disorders. Nutrients 2023, 15. [CrossRef]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.F. Impact of microbiota on central nervous system and neurological diseases: the gut-brain axis. J Neuroinflammation 2019, 16, 53. [CrossRef]
- Góralczyk-Bińkowska, A.; Szmajda-Krygier, D.; Kozłowska, E. The Microbiota-Gut-Brain Axis in Psychiatric Disorders. Int J Mol Sci 2022, 23. [CrossRef]
- Han, Y.; Wang, B.; Gao, H.; He, C.; Hua, R.; Liang, C.; Zhang, S.; Wang, Y.; Xin, S.; Xu, J. Vagus Nerve and Underlying Impact on the Gut Microbiota-Brain Axis in Behavior and Neurodegenerative Diseases. J Inflamm Res 2022, 15, 6213-6230. [CrossRef]
- Naufel, M.F.; Truzzi, G.M.; Ferreira, C.M.; Coelho, F.M.S. The brain-gut-microbiota axis in the treatment of neurologic and psychiatric disorders. Arq Neuropsiquiatr 2023, 81, 670-684. [CrossRef]
- Loh, J.S.; Mak, W.Q.; Tan, L.K.S.; Ng, C.X.; Chan, H.H.; Yeow, S.H.; Foo, J.B.; Ong, Y.S.; How, C.W.; Khaw, K.Y. Microbiota-gut-brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct Target Ther 2024, 9, 37. [CrossRef]
- Rutsch, A.; Kantsjö, J.B.; Ronchi, F. The Gut-Brain Axis: How Microbiota and Host Inflammasome Influence Brain Physiology and Pathology. Front Immunol 2020, 11, 604179. [CrossRef]
- Long-Smith, C.; O'Riordan, K.J.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota-Gut-Brain Axis: New Therapeutic Opportunities. Annu Rev Pharmacol Toxicol 2020, 60, 477-502. [CrossRef]
- Lee, S.H.; Han, C.; Shin, C. IUPHAR review: Microbiota-gut-brain axis and its role in neuropsychiatric disorders. Pharmacol Res 2025, 216, 107749. [CrossRef]
- Zhou, M.; Fan, Y.; Xu, L.; Yu, Z.; Wang, S.; Xu, H.; Zhang, J.; Zhang, L.; Liu, W.; Wu, L.; et al. Microbiome and tryptophan metabolomics analysis in adolescent depression: roles of the gut microbiota in the regulation of tryptophan-derived neurotransmitters and behaviors in human and mice. Microbiome 2023, 11, 145. [CrossRef]
- Zhao, H.; Qiu, X.; Wang, S.; Wang, Y.; Xie, L.; Xia, X.; Li, W. Multiple pathways through which the gut microbiota regulates neuronal mitochondria constitute another possible direction for depression. Front Microbiol 2025, 16, 1578155. [CrossRef]
- Liu, L.; Wang, H.; Chen, X.; Zhang, Y.; Zhang, H.; Xie, P. Gut microbiota and its metabolites in depression: from pathogenesis to treatment. EBioMedicine 2023, 90, 104527. [CrossRef]
- Ma, N.; He, T.; Johnston, L.J.; Ma, X. Host-microbiome interactions: the aryl hydrocarbon receptor as a critical node in tryptophan metabolites to brain signaling. Gut Microbes 2020, 11, 1203-1219. [CrossRef]
- Salminen, A. Activation of aryl hydrocarbon receptor (AhR) in Alzheimer's disease: role of tryptophan metabolites generated by gut host-microbiota. J Mol Med (Berl) 2023, 101, 201-222. [CrossRef]
- Zhu, S.; Jiang, Y.; Xu, K.; Cui, M.; Ye, W.; Zhao, G.; Jin, L.; Chen, X. The progress of gut microbiome research related to brain disorders. J Neuroinflammation 2020, 17, 25. [CrossRef]
- You, M.; Chen, N.; Yang, Y.; Cheng, L.; He, H.; Cai, Y.; Liu, Y.; Liu, H.; Hong, G. The gut microbiota-brain axis in neurological disorders. MedComm (2020) 2024, 5, e656. [CrossRef]
- Zatorska, O. Impact of gut microbiota on the central nervous system relevance in neurodegenerative and psychiatric diseases. Current Problems of Psychiatry 2024, 25, 239-247.
- Varesi, A.; Campagnoli, L.I.M.; Chirumbolo, S.; Candiano, B.; Carrara, A.; Ricevuti, G.; Esposito, C.; Pascale, A. The brain-gut-microbiota interplay in depression: A key to design innovative therapeutic approaches. Pharmacol Res 2023, 192, 106799. [CrossRef]
- Palepu, M.S.K.; Dandekar, M.P. Remodeling of microbiota gut-brain axis using psychobiotics in depression. Eur J Pharmacol 2022, 931, 175171. [CrossRef]
- Bhatt, S.; Kanoujia, J.; Mohana Lakshmi, S.; Patil, C.R.; Gupta, G.; Chellappan, D.K.; Dua, K. Role of Brain-Gut-Microbiota Axis in Depression: Emerging Therapeutic Avenues. CNS Neurol Disord Drug Targets 2023, 22, 276-288. [CrossRef]
- Generoso, J.S.; Giridharan, V.V.; Lee, J.; Macedo, D.; Barichello, T. The role of the microbiota-gut-brain axis in neuropsychiatric disorders. Braz J Psychiatry 2021, 43, 293-305. [CrossRef]
- Zhang, Q.; Bi, Y.; Zhang, B.; Jiang, Q.; Mou, C.K.; Lei, L.; Deng, Y.; Li, Y.; Yu, J.; Liu, W.; et al. Current landscape of fecal microbiota transplantation in treating depression. Front Immunol 2024, 15, 1416961. [CrossRef]
- Qiao, Y.; Yu, J.; Zhang, Z.; Hou, Q.; Guo, Z.; Wang, Y. Regulatory effects of Lactobacillus zhachilii HBUAS52074(T) on depression-like behavior induced by chronic social defeat stress in mice: modulation of the gut microbiota. Food Funct 2025, 16, 691-706. [CrossRef]
- Li, W.; Xu, M.; Liu, Y.; Zhang, S.; Wang, J.; Zhang, Z.; Xiao, G.; Wang, R.; Zhang, J.; Xue, H. Lactiplantibacillus plantarum GOLDGUT-HNU082 Alleviates CUMS-Induced Depressive-like Behaviors in Mice by Modulating the Gut Microbiota and Neurotransmitter Levels. Foods 2025, 14. [CrossRef]
- Xiong, R.G.; Li, J.; Cheng, J.; Zhou, D.D.; Wu, S.X.; Huang, S.Y.; Saimaiti, A.; Yang, Z.J.; Gan, R.Y.; Li, H.B. The Role of Gut Microbiota in Anxiety, Depression, and Other Mental Disorders as Well as the Protective Effects of Dietary Components. Nutrients 2023, 15. [CrossRef]
- Sanada, K.; Nakajima, S.; Kurokawa, S.; Barceló-Soler, A.; Ikuse, D.; Hirata, A.; Yoshizawa, A.; Tomizawa, Y.; Salas-Valero, M.; Noda, Y.; et al. Gut microbiota and major depressive disorder: A systematic review and meta-analysis. J Affect Disord 2020, 266, 1-13. [CrossRef]
- Ng, Q.X.; Lim, Y.L.; Yaow, C.Y.L.; Ng, W.K.; Thumboo, J.; Liew, T.M. Effect of Probiotic Supplementation on Gut Microbiota in Patients with Major Depressive Disorders: A Systematic Review. Nutrients 2023, 15. [CrossRef]
- Hofmeister, M.; Clement, F.; Patten, S.; Li, J.; Dowsett, L.E.; Farkas, B.; Mastikhina, L.; Egunsola, O.; Diaz, R.; Cooke, N.C.A.; et al. The effect of interventions targeting gut microbiota on depressive symptoms: a systematic review and meta-analysis. CMAJ Open 2021, 9, E1195-e1204. [CrossRef]
- Esmeeta, A.; Adhikary, S.; Dharshnaa, V.; Swarnamughi, P.; Ummul Maqsummiya, Z.; Banerjee, A.; Pathak, S.; Duttaroy, A.K. Plant-derived bioactive compounds in colon cancer treatment: An updated review. Biomed Pharmacother 2022, 153, 113384. [CrossRef]
- Naoi, M.; Wu, Y.; Maruyama, W.; Shamoto-Nagai, M. Phytochemicals Modulate Biosynthesis and Function of Serotonin, Dopamine, and Norepinephrine for Treatment of Monoamine Neurotransmission-Related Psychiatric Diseases. Int J Mol Sci 2025, 26. [CrossRef]
- Rahman, M.A.; Hannan, M.A.; Dash, R.; Rahman, M.H.; Islam, R.; Uddin, M.J.; Sohag, A.A.M.; Rahman, M.H.; Rhim, H. Phytochemicals as a Complement to Cancer Chemotherapy: Pharmacological Modulation of the Autophagy-Apoptosis Pathway. Front Pharmacol 2021, 12, 639628. [CrossRef]
- Phan, M.A.T.; Paterson, J.; Bucknall, M.; Arcot, J. Interactions between phytochemicals from fruits and vegetables: Effects on bioactivities and bioavailability. Crit Rev Food Sci Nutr 2018, 58, 1310-1329. [CrossRef]
- Zhao, Q.; Liu, J.; Chen, L.; Gao, Z.; Lin, M.; Wang, Y.; Xiao, Z.; Chen, Y.; Huang, X. Phytomedicine Fructus Aurantii-derived two absorbed compounds unlock antidepressant and prokinetic multi-functions via modulating 5-HT(3)/GHSR. J Ethnopharmacol 2024, 323, 117703. [CrossRef]
- Cheng, S.; Zhu, Z.; Li, H.; Wang, W.; Jiang, Z.; Pan, F.; Liu, D.; Ho, R.C.M.; Ho, C.S.H. Rifaximin ameliorates depression-like behaviour in chronic unpredictable mild stress rats by regulating intestinal microbiota and hippocampal tryptophan metabolism. J Affect Disord 2023, 329, 30-41. [CrossRef]
- Martínez-Rodríguez, A.; Rubio-Arias, J.; Ramos-Campo, D.J.; Reche-García, C.; Leyva-Vela, B.; Nadal-Nicolás, Y. Psychological and Sleep Effects of Tryptophan and Magnesium-Enriched Mediterranean Diet in Women with Fibromyalgia. Int J Environ Res Public Health 2020, 17. [CrossRef]
- Patel, A.; Cheung, J. The effect of mediterranean diet and chrononutrition on sleep quality: a scoping review. Nutr J 2025, 24, 31. [CrossRef]
- Scoditti, E.; Tumolo, M.R.; Garbarino, S. Mediterranean Diet on Sleep: A Health Alliance. Nutrients 2022, 14. [CrossRef]
- Godos, J.; Ferri, R.; Lanza, G.; Caraci, F.; Vistorte, A.O.R.; Yelamos Torres, V.; Grosso, G.; Castellano, S. Mediterranean Diet and Sleep Features: A Systematic Review of Current Evidence. Nutrients 2024, 16. [CrossRef]
- Gantenbein, K.V.; Kanaka-Gantenbein, C. Mediterranean Diet as an Antioxidant: The Impact on Metabolic Health and Overall Wellbeing. Nutrients 2021, 13. [CrossRef]
- Godos, J.; Ferri, R.; Caraci, F.; Cosentino, F.I.I.; Castellano, S.; Galvano, F.; Grosso, G. Adherence to the Mediterranean Diet is Associated with Better Sleep Quality in Italian Adults. Nutrients 2019, 11. [CrossRef]
- Casas, I.; Nakaki, A.; Pascal, R.; Castro-Barquero, S.; Youssef, L.; Genero, M.; Benitez, L.; Larroya, M.; Boutet, M.L.; Casu, G.; et al. Effects of a Mediterranean Diet Intervention on Maternal Stress, Well-Being, and Sleep Quality throughout Gestation-The IMPACT-BCN Trial. Nutrients 2023, 15. [CrossRef]
- Booij, L.; Merens, W.; Markus, C.R.; Van der Does, A.J. Diet rich in alpha-lactalbumin improves memory in unmedicated recovered depressed patients and matched controls. J Psychopharmacol 2006, 20, 526-535. [CrossRef]
- Porter, R.J.; Phipps, A.J.; Gallagher, P.; Scott, A.; Stevenson, P.S.; O'Brien, J.T. Effects of acute tryptophan depletion on mood and cognitive functioning in older recovered depressed subjects. The American journal of geriatric psychiatry 2005, 13, 607-615.
- Badrasawi, M.M.; Shahar, S.; Abd Manaf, Z.; Haron, H. Effect of Talbinah food consumption on depressive symptoms among elderly individuals in long term care facilities, randomized clinical trial. Clin Interv Aging 2013, 8, 279-285. [CrossRef]
- Jangid, P.; Malik, P.; Singh, P.; Sharma, M.; Gulia, A.K. Comparative study of efficacy of l-5-hydroxytryptophan and fluoxetine in patients presenting with first depressive episode. Asian J Psychiatr 2013, 6, 29-34. [CrossRef]
- Meloni, M.; Puligheddu, M.; Carta, M.; Cannas, A.; Figorilli, M.; Defazio, G. Efficacy and safety of 5-hydroxytryptophan on depression and apathy in Parkinson's disease: a preliminary finding. Eur J Neurol 2020, 27, 779-786. [CrossRef]
- Markus, C.R.; Olivier, B.; Panhuysen, G.E.; Van Der Gugten, J.; Alles, M.S.; Tuiten, A.; Westenberg, H.G.; Fekkes, D.; Koppeschaar, H.F.; de Haan, E.E. The bovine protein alpha-lactalbumin increases the plasma ratio of tryptophan to the other large neutral amino acids, and in vulnerable subjects raises brain serotonin activity, reduces cortisol concentration, and improves mood under stress. Am J Clin Nutr 2000, 71, 1536-1544. [CrossRef]
- Firk, C.; Markus, C.R. Mood and cortisol responses following tryptophan-rich hydrolyzed protein and acute stress in healthy subjects with high and low cognitive reactivity to depression. Clin Nutr 2009, 28, 266-271. [CrossRef]
- Benkelfat, C.; Ellenbogen, M.A.; Dean, P.; Palmour, R.M.; Young, S.N. Mood-lowering effect of tryptophan depletion. Enhanced susceptibility in young men at genetic risk for major affective disorders. Arch Gen Psychiatry 1994, 51, 687-697. [CrossRef]
- Bruce, K.R.; Steiger, H.; Young, S.N.; Kin, N.M.; Israël, M.; Lévesque, M. Impact of acute tryptophan depletion on mood and eating-related urges in bulimic and nonbulimic women. J Psychiatry Neurosci 2009, 34, 376-382.
- Murphy, S.E.; Longhitano, C.; Ayres, R.E.; Cowen, P.J.; Harmer, C.J. Tryptophan supplementation induces a positive bias in the processing of emotional material in healthy female volunteers. Psychopharmacology (Berl) 2006, 187, 121-130. [CrossRef]
- Rueda, G.H.; Causada-Calo, N.; Borojevic, R.; Nardelli, A.; Pinto-Sanchez, M.I.; Constante, M.; Libertucci, J.; Mohan, V.; Langella, P.; Loonen, L.M.P.; et al. Oral tryptophan activates duodenal aryl hydrocarbon receptor in healthy subjects: a crossover randomized controlled trial. Am J Physiol Gastrointest Liver Physiol 2024, 326, G687-g696. [CrossRef]
- Cerit, H.; Schuur, R.J.; de Bruijn, E.R.; Van der Does, W. Tryptophan supplementation and the response to unfairness in healthy volunteers. Front Psychol 2015, 6, 1012. [CrossRef]
- Markus, C.R.; Firk, C.; Gerhardt, C.; Kloek, J.; Smolders, G.F. Effect of different tryptophan sources on amino acids availability to the brain and mood in healthy volunteers. Psychopharmacology (Berl) 2008, 201, 107-114. [CrossRef]
- Bravo, R.; Matito, S.; Cubero, J.; Paredes, S.D.; Franco, L.; Rivero, M.; Rodríguez, A.B.; Barriga, C. Tryptophan-enriched cereal intake improves nocturnal sleep, melatonin, serotonin, and total antioxidant capacity levels and mood in elderly humans. Age (Dordr) 2013, 35, 1277-1285. [CrossRef]
- Rondanelli, M.; Opizzi, A.; Faliva, M.; Mozzoni, M.; Antoniello, N.; Cazzola, R.; Savarè, R.; Cerutti, R.; Grossi, E.; Cestaro, B. Effects of a diet integration with an oily emulsion of DHA-phospholipids containing melatonin and tryptophan in elderly patients suffering from mild cognitive impairment. Nutr Neurosci 2012, 15, 46-54. [CrossRef]
- Connell, N.J.; Grevendonk, L.; Fealy, C.E.; Moonen-Kornips, E.; Bruls, Y.M.H.; Schrauwen-Hinderling, V.B.; de Vogel, J.; Hageman, R.; Geurts, J.; Zapata-Perez, R.; et al. NAD+-Precursor Supplementation With L-Tryptophan, Nicotinic Acid, and Nicotinamide Does Not Affect Mitochondrial Function or Skeletal Muscle Function in Physically Compromised Older Adults. J Nutr 2021, 151, 2917-2931. [CrossRef]
- Chojnacki, C.; Gąsiorowska, A.; Popławski, T.; Konrad, P.; Chojnacki, M.; Fila, M.; Blasiak, J. Beneficial Effect of Increased Tryptophan Intake on Its Metabolism and Mental State of the Elderly. Nutrients 2023, 15. [CrossRef]
- Lieberman, H.R.; Agarwal, S.; Fulgoni, V.L., 3rd. Tryptophan Intake in the US Adult Population Is Not Related to Liver or Kidney Function but Is Associated with Depression and Sleep Outcomes. J Nutr 2016, 146, 2609s-2615s. [CrossRef]
- Nematolahi, P.; Mehrabani, M.; Karami-Mohajeri, S.; Dabaghzadeh, F. Effects of Rosmarinus officinalis L. on memory performance, anxiety, depression, and sleep quality in university students: A randomized clinical trial. Complement Ther Clin Pract 2018, 30, 24-28. [CrossRef]
- Ghasemzadeh Rahbardar, M.; Hosseinzadeh, H. Therapeutic effects of rosemary (Rosmarinus officinalis L.) and its active constituents on nervous system disorders. Iran J Basic Med Sci 2020, 23, 1100-1112. [CrossRef]
- Li, T.; Wang, W.; Guo, Q.; Li, J.; Tang, T.; Wang, Y.; Liu, D.; Yang, K.; Li, J.; Deng, K.; et al. Rosemary (Rosmarinus officinalis L.) hydrosol based on serotonergic synapse for insomnia. J Ethnopharmacol 2024, 318, 116984. [CrossRef]
- Kenda, M.; Kočevar Glavač, N.; Nagy, M.; Sollner Dolenc, M. Medicinal Plants Used for Anxiety, Depression, or Stress Treatment: An Update. Molecules 2022, 27. [CrossRef]
- Can, S.; Yildirim Usta, Y.; Yildiz, S.; Tayfun, K. The effect of lavender and rosemary aromatherapy application on cognitive functions, anxiety, and sleep quality in the elderly with diabetes. Explore (NY) 2024, 20, 103033. [CrossRef]
- Ghasemi, A.; Sharafabadi, F.M.; Marani, E. The Effect of Inhalation Aromatherapy with Essential Oils of Various Plants on Anxiety and Sleep Quality in Diabetic Patients: A Review Study. Iranian journal of diabetes and obesity 2025.
- Caballero-Gallardo, K.; Quintero-Rincón, P.; Olivero-Verbel, J. Aromatherapy and Essential Oils: Holistic Strategies in Complementary and Alternative Medicine for Integral Wellbeing. Plants (Basel) 2025, 14. [CrossRef]
- Mathews, I.M.; Eastwood, J.; Lamport, D.J.; Cozannet, R.L.; Fanca-Berthon, P.; Williams, C.M. Clinical Efficacy and Tolerability of Lemon Balm (Melissa officinalis L.) in Psychological Well-Being: A Review. Nutrients 2024, 16. [CrossRef]
- Lu, C.; Wei, Z.; Jiang, N.; Chen, Y.; Wang, Y.; Li, S.; Wang, Q.; Fan, B.; Liu, X.; Wang, F. Soy isoflavones protects against cognitive deficits induced by chronic sleep deprivation via alleviating oxidative stress and suppressing neuroinflammation. Phytother Res 2022, 36, 2072-2080. [CrossRef]
- Mendonça, C.R.; Manhães-de-Castro, R.; de Santana, B.; Olegário da Silva, L.; Toscano, A.E.; Guzmán-Quevedo, O.; Galindo, L.C.M. Effects of flavonols on emotional behavior and compounds of the serotonergic system: A preclinical systematic review. Eur J Pharmacol 2022, 916, 174697. [CrossRef]
- Hamsalakshmi; Alex, A.M.; Arehally Marappa, M.; Joghee, S.; Chidambaram, S.B. Therapeutic benefits of flavonoids against neuroinflammation: a systematic review. Inflammopharmacology 2022, 30, 111-136. [CrossRef]
- Scuto, M.; Majzúnová, M.; Torcitto, G.; Antonuzzo, S.; Rampulla, F.; Di Fatta, E.; Trovato Salinaro, A. Functional Food Nutrients, Redox Resilience Signaling and Neurosteroids for Brain Health. Int J Mol Sci 2024, 25. [CrossRef]
- Zhang, Z.; Li, X.; Li, F.; An, L. Berberine alleviates postoperative cognitive dysfunction by suppressing neuroinflammation in aged mice. Int Immunopharmacol 2016, 38, 426-433. [CrossRef]
- Premkumar, L.S. Transient receptor potential channels as targets for phytochemicals. ACS Chem Neurosci 2014, 5, 1117-1130. [CrossRef]
- Pham, D.C.; Shibu, M.A.; Mahalakshmi, B.; Velmurugan, B.K. Effects of phytochemicals on cellular signaling: reviewing their recent usage approaches. Crit Rev Food Sci Nutr 2020, 60, 3522-3546. [CrossRef]
- Rao, P.P. Phytochemicals in Obesity Management: Mechanisms and Clinical Perspectives. Curr Nutr Rep 2025, 14, 17. [CrossRef]
- Sarris, J.; Ravindran, A.; Yatham, L.N.; Marx, W.; Rucklidge, J.J.; McIntyre, R.S.; Akhondzadeh, S.; Benedetti, F.; Caneo, C.; Cramer, H.; et al. Clinician guidelines for the treatment of psychiatric disorders with nutraceuticals and phytoceuticals: The World Federation of Societies of Biological Psychiatry (WFSBP) and Canadian Network for Mood and Anxiety Treatments (CANMAT) Taskforce. World J Biol Psychiatry 2022, 23, 424-455. [CrossRef]
- Hao, W.Z.; Ma, Q.Y.; Tao, G.; Huang, J.Q.; Chen, J.X. Oral coniferyl ferulate attenuated depression symptoms in mice via reshaping gut microbiota and microbial metabolism. Food Funct 2021, 12, 12550-12564. [CrossRef]
- Mannino, G.; Serio, G.; Gaglio, R.; Maffei, M.E.; Settanni, L.; Di Stefano, V.; Gentile, C. Biological Activity and Metabolomics of Griffonia simplicifolia Seeds Extracted with Different Methodologies. Antioxidants (Basel) 2023, 12. [CrossRef]
- Ribas, R.; Oliveira, D.; Silva, P. The different roles of Griffonia simplicifolia in the treatment of depression: a narrative review. Int J Complement Alt Med 2021, 14, 167-172.
- Moharir, Y. 5-HTP for Improved Sleep, Mood and Weight Loss.
- Fellows, L.E.; Bell, E.A. 5-Hydroxy-L-tryptophan, 5-hydroxytryptamine and L-tryptophan-5-hydroxylase in Griffonia simplicifolia. Phytochemistry 1970, 9, 2389-2396.
- Vigliante, I.; Mannino, G.; Maffei, M.E. Chemical Characterization and DNA Fingerprinting of Griffonia simplicifolia Baill. Molecules 2019, 24. [CrossRef]
- Fijałkowska, A.; Jędrejko, K.; Sułkowska-Ziaja, K.; Ziaja, M.; Kała, K.; Muszyńska, B. Edible Mushrooms as a Potential Component of Dietary Interventions for Major Depressive Disorder. Foods 2022, 11. [CrossRef]
- Lu, C.; Wei, Z.; Wang, Y.; Li, S.; Tong, L.; Liu, X.; Fan, B.; Wang, F. Soy isoflavones alleviate lipopolysaccharide-induced depressive-like behavior by suppressing neuroinflammation, mediating tryptophan metabolism and promoting synaptic plasticity. Food Funct 2022, 13, 9513-9522. [CrossRef]
- Lu, C.; Gao, R.; Zhang, Y.; Jiang, N.; Chen, Y.; Sun, J.; Wang, Q.; Fan, B.; Liu, X.; Wang, F. S-equol, a metabolite of dietary soy isoflavones, alleviates lipopolysaccharide-induced depressive-like behavior in mice by inhibiting neuroinflammation and enhancing synaptic plasticity. Food Funct 2021, 12, 5770-5778. [CrossRef]
- McLaren, S.; Seidler, K.; Neil, J. Investigating the Role of 17β-Estradiol on the Serotonergic System, Targeting Soy Isoflavones as a Strategy to Reduce Menopausal Depression: A Mechanistic Review. J Am Nutr Assoc 2024, 43, 221-235. [CrossRef]
- Sekikawa, A.; Wharton, W.; Butts, B.; Veliky, C.V.; Garfein, J.; Li, J.; Goon, S.; Fort, A.; Li, M.; Hughes, T.M. Potential Protective Mechanisms of S-equol, a Metabolite of Soy Isoflavone by the Gut Microbiome, on Cognitive Decline and Dementia. Int J Mol Sci 2022, 23. [CrossRef]
- Cui, C.; Birru, R.; Snitz, B.E.; Ihara, M.; Lopresti, B.J.; Aizenstein, H.J.; Lopez, O.L.; Mathis, C.; Miyamoto, Y.; Kuller, L.H. P3-615: EFFECTS OF SOY ISOFLAVONES ON COGNITIVE FUNCTION: A SYSTEMATIC REVIEW AND META-ANALYSIS OF RANDOMIZED CONTROLLED TRIALS. Alzheimer's & Dementia 2018, 14, P1365-P1366.
- Huang, M.; He, Y.; Tian, L.; Yu, L.; Cheng, Q.; Li, Z.; Gao, L.; Gao, S.; Yu, C. Gut microbiota-SCFAs-brain axis associated with the antidepressant activity of berberine in CUMS rats. J Affect Disord 2023, 325, 141-150. [CrossRef]
- Lu, Y.; Yu, X.; Wang, Z.; Kong, L.; Jiang, Z.; Shang, R.; Zhong, X.; Lv, S.; Zhang, G.; Gao, H.; et al. Microbiota-gut-brain axis: Natural antidepressants molecular mechanism. Phytomedicine 2024, 134, 156012. [CrossRef]
- Qian, X.H.; Xie, R.Y.; Liu, X.L.; Chen, S.D.; Tang, H.D. Mechanisms of Short-Chain Fatty Acids Derived from Gut Microbiota in Alzheimer's Disease. Aging Dis 2022, 13, 1252-1266. [CrossRef]
- Cheng, J.; Hu, H.; Ju, Y.; Liu, J.; Wang, M.; Liu, B.; Zhang, Y. Gut microbiota-derived short-chain fatty acids and depression: deep insight into biological mechanisms and potential applications. Gen Psychiatr 2024, 37, e101374. [CrossRef]
- Zhao, Z.X.; Fu, J.; Ma, S.R.; Peng, R.; Yu, J.B.; Cong, L.; Pan, L.B.; Zhang, Z.G.; Tian, H.; Che, C.T.; et al. Gut-brain axis metabolic pathway regulates antidepressant efficacy of albiflorin. Theranostics 2018, 8, 5945-5959. [CrossRef]
- Wu, D.; Chen, Q.; Chen, X.; Han, F.; Chen, Z.; Wang, Y. The blood-brain barrier: structure, regulation, and drug delivery. Signal Transduct Target Ther 2023, 8, 217. [CrossRef]
- Qin, Z.; Shi, D.D.; Li, W.; Cheng, D.; Zhang, Y.D.; Zhang, S.; Tsoi, B.; Zhao, J.; Wang, Z.; Zhang, Z.J. Berberine ameliorates depression-like behaviors in mice via inhibiting NLRP3 inflammasome-mediated neuroinflammation and preventing neuroplasticity disruption. J Neuroinflammation 2023, 20, 54. [CrossRef]
- Tang, C.F.; Wang, C.Y.; Wang, J.H.; Wang, Q.N.; Li, S.J.; Wang, H.O.; Zhou, F.; Li, J.M. Short-Chain Fatty Acids Ameliorate Depressive-like Behaviors of High Fructose-Fed Mice by Rescuing Hippocampal Neurogenesis Decline and Blood-Brain Barrier Damage. Nutrients 2022, 14. [CrossRef]
- Dudek, K.A.; Dion-Albert, L.; Lebel, M.; LeClair, K.; Labrecque, S.; Tuck, E.; Ferrer Perez, C.; Golden, S.A.; Tamminga, C.; Turecki, G.; et al. Molecular adaptations of the blood-brain barrier promote stress resilience vs. depression. Proc Natl Acad Sci U S A 2020, 117, 3326-3336. [CrossRef]
- Knox, E.G.; Aburto, M.R.; Clarke, G.; Cryan, J.F.; O'Driscoll, C.M. The blood-brain barrier in aging and neurodegeneration. Mol Psychiatry 2022, 27, 2659-2673. [CrossRef]
- Grabska-Kobyłecka, I.; Szpakowski, P.; Król, A.; Książek-Winiarek, D.; Kobyłecki, A.; Głąbiński, A.; Nowak, D. Polyphenols and Their Impact on the Prevention of Neurodegenerative Diseases and Development. Nutrients 2023, 15. [CrossRef]
- Jaeger, B.N.; Parylak, S.L.; Gage, F.H. Mechanisms of dietary flavonoid action in neuronal function and neuroinflammation. Mol Aspects Med 2018, 61, 50-62. [CrossRef]
- Calderaro, A.; Patanè, G.T.; Tellone, E.; Barreca, D.; Ficarra, S.; Misiti, F.; Laganà, G. The Neuroprotective Potentiality of Flavonoids on Alzheimer's Disease. Int J Mol Sci 2022, 23. [CrossRef]
- Pontifex, M.G.; Malik, M.; Connell, E.; Müller, M.; Vauzour, D. Citrus Polyphenols in Brain Health and Disease: Current Perspectives. Front Neurosci 2021, 15, 640648. [CrossRef]
- Martínez-Coria, H.; Arrieta-Cruz, I.; Gutiérrez-Juárez, R.; López-Valdés, H.E. Anti-Inflammatory Effects of Flavonoids in Common Neurological Disorders Associated with Aging. Int J Mol Sci 2023, 24. [CrossRef]
- Bakoyiannis, I.; Daskalopoulou, A.; Pergialiotis, V.; Perrea, D. Phytochemicals and cognitive health: Are flavonoids doing the trick? Biomed Pharmacother 2019, 109, 1488-1497. [CrossRef]
- Zagoskina, N.V.; Zubova, M.Y.; Nechaeva, T.L.; Kazantseva, V.V.; Goncharuk, E.A.; Katanskaya, V.M.; Baranova, E.N.; Aksenova, M.A. Polyphenols in Plants: Structure, Biosynthesis, Abiotic Stress Regulation, and Practical Applications (Review). Int J Mol Sci 2023, 24. [CrossRef]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des Devel Ther 2016, 10, 23-42. [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24. [CrossRef]
- Carey, C.C.; Lucey, A.; Doyle, L. Flavonoid Containing Polyphenol Consumption and Recovery from Exercise-Induced Muscle Damage: A Systematic Review and Meta-Analysis. Sports Med 2021, 51, 1293-1316. [CrossRef]
- Shi, J.; Du, P.; Xie, Q.; Wang, N.; Li, H.; Smith, E.E.; Li, C.; Liu, F.; Huo, G.; Li, B. Protective effects of tryptophan-catabolizing Lactobacillus plantarum KLDS 1.0386 against dextran sodium sulfate-induced colitis in mice. Food Funct 2020, 11, 10736-10747. [CrossRef]
- Friedman, M. Analysis, Nutrition, and Health Benefits of Tryptophan. Int J Tryptophan Res 2018, 11, 1178646918802282. [CrossRef]
- Ferrer, L.; Elsaraf, M.; Mindt, M.; Wendisch, V.F. l-Serine Biosensor-Controlled Fermentative Production of l-Tryptophan Derivatives by Corynebacterium glutamicum. Biology (Basel) 2022, 11. [CrossRef]
- Guo, Y.W.; Liu, X.J.; Yuan, J.; Li, H.J.; Mahmud, T.; Hong, M.J.; Yu, J.C.; Lan, W.J. l-Tryptophan Induces a Marine-Derived Fusarium sp. to Produce Indole Alkaloids with Activity against the Zika Virus. J Nat Prod 2020, 83, 3372-3380. [CrossRef]
- Henarejos-Escudero, P.; Méndez-García, F.F.; Hernández-García, S.; Martínez-Rodríguez, P.; Gandía-Herrero, F. Design, Synthesis and Gene Modulation Insights into Pigments Derived from Tryptophan-Betaxanthin, Which Act against Tumor Development in Caenorhabditis elegans. Int J Mol Sci 2023, 25. [CrossRef]
- Shaw, K.; Turner, J.; Del Mar, C. Tryptophan and 5-hydroxytryptophan for depression. Cochrane Database Syst Rev 2002, Cd003198. [CrossRef]
- Herrington, R.N.; Bruce, A.; Johnstone, E.C. Comparative trial of L-tryptophan and E.C.T. in severe depressive illness. Lancet 1974, 2, 731-734. [CrossRef]
- Kikuchi, A.M.; Tanabe, A.; Iwahori, Y. A systematic review of the effect of L-tryptophan supplementation on mood and emotional functioning. J Diet Suppl 2021, 18, 316-333. [CrossRef]
- Tian, P.; Chen, Y.; Zhu, H.; Wang, L.; Qian, X.; Zou, R.; Zhao, J.; Zhang, H.; Qian, L.; Wang, Q.; et al. Bifidobacterium breve CCFM1025 attenuates major depression disorder via regulating gut microbiome and tryptophan metabolism: A randomized clinical trial. Brain Behav Immun 2022, 100, 233-241. [CrossRef]
- Liang, X.; Su, T.; Wu, P.; Dai, Y.; Chen, Y.; Wang, Q.; Cao, C.; Chen, F.; Wang, Q.; Wang, S. Identification of paeoniflorin from Paeonia lactiflora pall. As an inhibitor of tryptophan 2,3-dioxygenase and assessment of its pharmacological effects on depressive mice. J Ethnopharmacol 2023, 317, 116714. [CrossRef]
- Kałużna-Czaplińska, J.; Gątarek, P.; Chirumbolo, S.; Chartrand, M.S.; Bjørklund, G. How important is tryptophan in human health? Crit Rev Food Sci Nutr 2019, 59, 72-88. [CrossRef]
- Sutanto, C.N.; Loh, W.W.; Toh, D.W.K.; Lee, D.P.S.; Kim, J.E. Association Between Dietary Protein Intake and Sleep Quality in Middle-Aged and Older Adults in Singapore. Front Nutr 2022, 9, 832341. [CrossRef]
- Sutanto, C.N.; Loh, W.W.; Kim, J.E. The impact of tryptophan supplementation on sleep quality: a systematic review, meta-analysis, and meta-regression. Nutr Rev 2022, 80, 306-316. [CrossRef]
- Mohajeri, M.H.; Wittwer, J.; Vargas, K.; Hogan, E.; Holmes, A.; Rogers, P.J.; Goralczyk, R.; Gibson, E.L. Chronic treatment with a tryptophan-rich protein hydrolysate improves emotional processing, mental energy levels and reaction time in middle-aged women. Br J Nutr 2015, 113, 350-365. [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc Natl Acad Sci U S A 2020, 117, 19376-19387. [CrossRef]
- Liu, W.; Mi, S.; Ruan, Z.; Li, J.; Shu, X.; Yao, K.; Jiang, M.; Deng, Z. Dietary Tryptophan Enhanced the Expression of Tight Junction Protein ZO-1 in Intestine. J Food Sci 2017, 82, 562-567. [CrossRef]
- Liang, H.; Dai, Z.; Kou, J.; Sun, K.; Chen, J.; Yang, Y.; Wu, G.; Wu, Z. Dietary l-Tryptophan Supplementation Enhances the Intestinal Mucosal Barrier Function in Weaned Piglets: Implication of Tryptophan-Metabolizing Microbiota. Int J Mol Sci 2018, 20. [CrossRef]
- Taleb, S. Tryptophan Dietary Impacts Gut Barrier and Metabolic Diseases. Front Immunol 2019, 10, 2113. [CrossRef]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front Cell Infect Microbiol 2018, 8, 13. [CrossRef]
- Dai, H.; Yang, H.; Wang, R.; Wang, X.; Zhang, X. Modulating Gut Microbiota with Dietary Components: A Novel Strategy for Cancer-Depression Comorbidity Management. Nutrients 2025, 17. [CrossRef]
- Trzeciak, P.; Herbet, M. Role of the Intestinal Microbiome, Intestinal Barrier and Psychobiotics in Depression. Nutrients 2021, 13. [CrossRef]
- Nunes, Y.C.; Mendes, N.M.; Pereira de Lima, E.; Chehadi, A.C.; Lamas, C.B.; Haber, J.F.S.; Dos Santos Bueno, M.; Araújo, A.C.; Catharin, V.C.S.; Detregiachi, C.R.P.; et al. Curcumin: A Golden Approach to Healthy Aging: A Systematic Review of the Evidence. Nutrients 2024, 16. [CrossRef]
- Nabavi, S.M.; Daglia, M.; Braidy, N.; Nabavi, S.F. Natural products, micronutrients, and nutraceuticals for the treatment of depression: A short review. Nutr Neurosci 2017, 20, 180-194. [CrossRef]
- Fernstrom, J.D. Effects and side effects associated with the non-nutritional use of tryptophan by humans. J Nutr 2012, 142, 2236s-2244s. [CrossRef]
- Hiratsuka, C.; Fukuwatari, T.; Sano, M.; Saito, K.; Sasaki, S.; Shibata, K. Supplementing healthy women with up to 5.0 g/d of L-tryptophan has no adverse effects. J Nutr 2013, 143, 859-866. [CrossRef]
- Zhu, X.; Tao, Q.; Sun-Waterhouse, D.; Li, W.; Liu, S.; Cui, C. γ-[Glu]n-Trp ameliorates anxiety/depression-like behaviors and its anti-inflammatory effect in an animal model of anxiety/depression. Food Funct 2019, 10, 5544-5554. [CrossRef]
- Ko, S.M.; Park, J.E.; Kim, S.H.; Park, M.K.; Seol, J.K.; Heo, I.K.; Shin, Y.U.; Kim, S.Y.; Kim, Y.H.; Son, W.C. Safety evaluation of dried L-tryptophan fermentation product in Sprague-Dawley rats. Toxicol Rep 2022, 9, 1099-1113. [CrossRef]
- Medsger, T.A., Jr. Tryptophan-induced eosinophilia-myalgia syndrome. N Engl J Med 1990, 322, 926-928. [CrossRef]
- Ko, S.M.; Park, J.E.; Heo, I.K.; Shin, Y.U.; Kim, Y.H.; Son, W.C. Safety concerns regarding impurities in L-Tryptophan associated with eosinophilia myalgia syndrome. Food Chem Toxicol 2023, 179, 113946. [CrossRef]
- Mayeno, A.N.; Gleich, G.J. Eosinophilia-myalgia syndrome and tryptophan production: a cautionary tale. Trends Biotechnol 1994, 12, 346-352. [CrossRef]
- Hill, R.H., Jr.; Caudill, S.P.; Philen, R.M.; Bailey, S.L.; Flanders, W.D.; Driskell, W.J.; Kamb, M.L.; Needham, L.L.; Sampson, E.J. Contaminants in L-tryptophan associated with eosinophilia myalgia syndrome. Arch Environ Contam Toxicol 1993, 25, 134-142. [CrossRef]
- Eidson, M.; Philen, R.M.; Sewell, C.M.; Voorhees, R.; Kilbourne, E.M. L-tryptophan and eosinophilia-myalgia syndrome in New Mexico. Lancet 1990, 335, 645-648. [CrossRef]
- Hertzman, P.A.; Blevins, W.L.; Mayer, J.; Greenfield, B.; Ting, M.; Gleich, G.J. Association of the eosinophilia-myalgia syndrome with the ingestion of tryptophan. N Engl J Med 1990, 322, 869-873. [CrossRef]
- Das, Y.T.; Bagchi, M.; Bagchi, D.; Preuss, H.G. Safety of 5-hydroxy-L-tryptophan. Toxicol Lett 2004, 150, 111-122. [CrossRef]
- Martin, R.W.; Duffy, J.; Engel, A.G.; Lie, J.T.; Bowles, C.A.; Moyer, T.P.; Gleich, G.J. The clinical spectrum of the eosinophilia-myalgia syndrome associated with L-tryptophan ingestion. Clinical features in 20 patients and aspects of pathophysiology. Ann Intern Med 1990, 113, 124-134. [CrossRef]
- Verity, M.A.; Bulpitt, K.J.; Paulus, H.E. Neuromuscular manifestations of L-tryptophan-associated eosinophilia-myalgia syndrome: a histomorphologic analysis of 14 patients. Hum Pathol 1991, 22, 3-11. [CrossRef]
- Munir, S.; Shahid, A.; Aslam, B.; Ashfaq, U.A.; Akash, M.S.H.; Ali, M.A.; Almatroudi, A.; Allemailem, K.S.; Rajoka, M.S.R.; Khurshid, M. The Therapeutic Prospects of Naturally Occurring and Synthetic Indole Alkaloids for Depression and Anxiety Disorders. Evid Based Complement Alternat Med 2020, 2020, 8836983. [CrossRef]
- Kanova, M.; Kohout, P. Serotonin-Its Synthesis and Roles in the Healthy and the Critically Ill. Int J Mol Sci 2021, 22. [CrossRef]
- Guzel, T.; Mirowska-Guzel, D. The Role of Serotonin Neurotransmission in Gastrointestinal Tract and Pharmacotherapy. Molecules 2022, 27. [CrossRef]
- Mokhtari, T. Targeting autophagy and neuroinflammation pathways with plant-derived natural compounds as potential antidepressant agents. Phytother Res 2022, 36, 3470-3489. [CrossRef]
- Jamu, I.M.; Okamoto, H. Recent advances in understanding adverse effects associated with drugs targeting the serotonin receptor, 5-HT GPCR. Front Glob Womens Health 2022, 3, 1012463. [CrossRef]
- Gibson, E.L. Tryptophan supplementation and serotonin function: genetic variations in behavioural effects. Proc Nutr Soc 2018, 77, 174-188. [CrossRef]
- Averina, O.V.; Poluektova, E.U.; Zorkina, Y.A.; Kovtun, A.S.; Danilenko, V.N. Human Gut Microbiota for Diagnosis and Treatment of Depression. Int J Mol Sci 2024, 25. [CrossRef]
- Lin, P.; Li, D.; Shi, Y.; Li, Q.; Guo, X.; Dong, K.; Chen, Q.; Lou, X.; Li, Z.; Li, P.; et al. Dysbiosis of the Gut Microbiota and Kynurenine (Kyn) Pathway Activity as Potential Biomarkers in Patients with Major Depressive Disorder. Nutrients 2023, 15. [CrossRef]
- Xie, Y.; Zhu, H.; Yuan, Y.; Guan, X.; Xie, Q.; Dong, Z. Baseline gut microbiota profiles affect treatment response in patients with depression. Front Microbiol 2024, 15, 1429116. [CrossRef]
- Liu, L.; Wang, H.; Zhang, H.; Chen, X.; Zhang, Y.; Wu, J.; Zhao, L.; Wang, D.; Pu, J.; Ji, P.; et al. Toward a Deeper Understanding of Gut Microbiome in Depression: The Promise of Clinical Applicability. Adv Sci (Weinh) 2022, 9, e2203707. [CrossRef]
- Tian, P.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium with the role of 5-hydroxytryptophan synthesis regulation alleviates the symptom of depression and related microbiota dysbiosis. J Nutr Biochem 2019, 66, 43-51. [CrossRef]
- Wang, M.; Song, Z.; Lai, S.; Tang, F.; Dou, L.; Yang, F. Depression-associated gut microbes, metabolites and clinical trials. Front Microbiol 2024, 15, 1292004. [CrossRef]
- Clerici, L.; Bottari, D.; Bottari, B. Gut Microbiome, Diet and Depression: Literature Review of Microbiological, Nutritional and Neuroscientific Aspects. Curr Nutr Rep 2025, 14, 30. [CrossRef]
- Jiang, Y.; Qu, Y.; Shi, L.; Ou, M.; Du, Z.; Zhou, Z.; Zhou, H.; Zhu, H. The role of gut microbiota and metabolomic pathways in modulating the efficacy of SSRIs for major depressive disorder. Transl Psychiatry 2024, 14, 493. [CrossRef]
- Pawlowski, T.; Malyszczak, K.; Pawlak, D.; Inglot, M.; Zalewska, M.; Grzywacz, A.; Radkowski, M.; Laskus, T.; Janocha-Litwin, J.; Frydecka, D. HTR1A, TPH2, and 5-HTTLPR Polymorphisms and Their Impact on the Severity of Depressive Symptoms and on the Concentration of Tryptophan Catabolites during Hepatitis C Treatment with Pegylated Interferon-α2a and Oral Ribavirin (PEG-IFN-α2a/RBV). Cells 2023, 12. [CrossRef]
- Duan, K.M.; Wang, S.Y.; Yin, J.Y.; Li, X.; Ma, J.H.; Huang, Z.D.; Zhou, Y.Y.; Yu, H.Y.; Yang, M.; Zhou, H.H.; et al. The IDO genetic polymorphisms and postpartum depressive symptoms: an association study in Chinese parturients who underwent cesarean section. Arch Womens Ment Health 2019, 22, 339-348. [CrossRef]
- Fasching, P.A.; Faschingbauer, F.; Goecke, T.W.; Engel, A.; Häberle, L.; Seifert, A.; Voigt, F.; Amann, M.; Rebhan, D.; Burger, P.; et al. Genetic variants in the tryptophan hydroxylase 2 gene (TPH2) and depression during and after pregnancy. J Psychiatr Res 2012, 46, 1109-1117. [CrossRef]
- MUSHTAQ, R. TRYPTOPHAN HYDROXYLASE GENE POLYMORPHISM AND 5HT TRANSPORTER GENE PROMOTER REGION POLYMORPHISM IN ANXIETY AND DEPRESSIVE DISORDER-COMPREHENSIVE LITERATURE REVIEW. INTERNATIONAL JOURNAL OF RESEARCH IN MEDICAL SCIENCES 2024, 12, 3957-3961.
- Ottenhof, K.W.; Sild, M.; Lévesque, M.L.; Ruhé, H.G.; Booij, L. TPH2 polymorphisms across the spectrum of psychiatric morbidity: A systematic review and meta-analysis. Neurosci Biobehav Rev 2018, 92, 29-42. [CrossRef]
- Tanaka, M.; Battaglia, S.; Giménez-Llort, L.; Chen, C.; Hepsomali, P.; Avenanti, A.; Vécsei, L. Innovation at the Intersection: Emerging Translational Research in Neurology and Psychiatry. Cells 2024, 13. [CrossRef]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat Rev Drug Discov 2019, 18, 379-401. [CrossRef]
- Chen, X.; Xu, D.; Yu, J.; Song, X.J.; Li, X.; Cui, Y.L. Tryptophan Metabolism Disorder-Triggered Diseases, Mechanisms, and Therapeutic Strategies: A Scientometric Review. Nutrients 2024, 16. [CrossRef]
- Siddiqui, A.J.; Adnan, M.; Saxena, J.; Alam, M.J.; Abdelgadir, A.; Badraoui, R.; Singh, R. Therapeutic Potential of Plant- and Marine-Derived Bioactive Compounds in Prostate Cancer: Mechanistic Insights and Translational Applications. Pharmaceuticals (Basel) 2025, 18. [CrossRef]
- Modoux, M.; Rolhion, N.; Mani, S.; Sokol, H. Tryptophan Metabolism as a Pharmacological Target. Trends Pharmacol Sci 2021, 42, 60-73. [CrossRef]
- Sadee, W.; Wang, D.; Hartmann, K.; Toland, A.E. Pharmacogenomics: Driving Personalized Medicine. Pharmacol Rev 2023, 75, 789-814. [CrossRef]
- Heinken, A.; Basile, A.; Hertel, J.; Thinnes, C.; Thiele, I. Genome-Scale Metabolic Modeling of the Human Microbiome in the Era of Personalized Medicine. Annu Rev Microbiol 2021, 75, 199-222. [CrossRef]
- Nowicka, A.; Tomczak, H.; Szałek, E.; Karbownik, A.; Gil, L. Microbial Crosstalk with Therapy: Pharmacomicrobiomics in AML-One Step Closer to Personalized Medicine. Biomedicines 2025, 13. [CrossRef]
- Li, D.; Yu, S.; Long, Y.; Shi, A.; Deng, J.; Ma, Y.; Wen, J.; Li, X.; Liu, S.; Zhang, Y.; et al. Tryptophan metabolism: Mechanism-oriented therapy for neurological and psychiatric disorders. Front Immunol 2022, 13, 985378. [CrossRef]
- Ge, P.Y.; Qu, S.Y.; Ni, S.J.; Yao, Z.Y.; Qi, Y.Y.; Zhao, X.; Guo, R.; Yang, N.Y.; Zhang, Q.C.; Zhu, H.X. Berberine ameliorates depression-like behavior in CUMS mice by activating TPH1 and inhibiting IDO1-associated with tryptophan metabolism. Phytother Res 2023, 37, 342-357. [CrossRef]
- Hu, M.; Shi, X.; Song, P.X. Collaborative inference for treatment effect with distributed data-sharing management in multicenter studies. Stat Med 2024, 43, 2263-2279. [CrossRef]
- Adhikari, B.M.; Jahanshad, N.; Shukla, D.; Turner, J.; Grotegerd, D.; Dannlowski, U.; Kugel, H.; Engelen, J.; Dietsche, B.; Krug, A.; et al. A resting state fMRI analysis pipeline for pooling inference across diverse cohorts: an ENIGMA rs-fMRI protocol. Brain Imaging Behav 2019, 13, 1453-1467. [CrossRef]
- Llovera, G.; Langhauser, F.; Isla Cainzos, S.; Hoppen, M.; Abberger, H.; Mohamud Yusuf, A.; Mencl, S.; Heindl, S.; Ricci, A.; Haupeltshofer, S.; et al. Stroke of Consistency: Streamlining Multicenter Protocols for Enhanced Reproducibility of Infarct Volumes in Preclinical Stroke Research. Stroke 2024, 55, 2522-2527. [CrossRef]
- Sheller, M.J.; Edwards, B.; Reina, G.A.; Martin, J.; Pati, S.; Kotrotsou, A.; Milchenko, M.; Xu, W.; Marcus, D.; Colen, R.R.; et al. Federated learning in medicine: facilitating multi-institutional collaborations without sharing patient data. Sci Rep 2020, 10, 12598. [CrossRef]
- Zhang, X.; Wang, N.; Zhang, L.; Cheng, C.W.; Liu, Z.; Liang, F.; Xiong, W.; Deng, J.; Shi, D.; Cui, W.; et al. Reporting assessment of multicenter clinical trial protocols: A cross-sectional study. J Evid Based Med 2023, 16, 16-18. [CrossRef]
- Chen, N.; Lee, J.J. Bayesian cluster hierarchical model for subgroup borrowing in the design and analysis of basket trials with binary endpoints. Stat Methods Med Res 2020, 29, 2717-2732. [CrossRef]
- Zhang, J.; Lin, R.; Chen, X.; Yan, F. Adaptive Bayesian information borrowing methods for finding and optimizing subgroup-specific doses. Clin Trials 2024, 21, 308-321. [CrossRef]
- Wang, G.; Levis, A.; Steingrimsson, J.; Dahabreh, I. Efficient estimation of subgroup treatment effects using multi-source data. arXiv preprint arXiv:2402.02684 2024.
- Tanaka, M. Parkinson's Disease: Bridging Gaps, Building Biomarkers, and Reimagining Clinical Translation. Cells 2025, 14. [CrossRef]
- Tanaka, M. From Serendipity to Precision: Integrating AI, Multi-Omics, and Human-Specific Models for Personalized Neuropsychiatric Care. Biomedicines 2025, 13. [CrossRef]
- Tanaka, M. Beyond the boundaries: Transitioning from categorical to dimensional paradigms in mental health diagnostics. Adv Clin Exp Med 2024, 33, 1295-1301. [CrossRef]
- Pagotto, G.L.O.; Santos, L.; Osman, N.; Lamas, C.B.; Laurindo, L.F.; Pomini, K.T.; Guissoni, L.M.; Lima, E.P.; Goulart, R.A.; Catharin, V.; et al. Ginkgo biloba: A Leaf of Hope in the Fight against Alzheimer's Dementia: Clinical Trial Systematic Review. Antioxidants (Basel) 2024, 13. [CrossRef]
- Tanaka, M.; Vécsei, L. A Decade of Dedication: Pioneering Perspectives on Neurological Diseases and Mental Illnesses. Biomedicines 2024, 12. [CrossRef]
- Chahla, J.; Cinque, M.E.; Piuzzi, N.S.; Mannava, S.; Geeslin, A.G.; Murray, I.R.; Dornan, G.J.; Muschler, G.F.; LaPrade, R.F. A Call for Standardization in Platelet-Rich Plasma Preparation Protocols and Composition Reporting: A Systematic Review of the Clinical Orthopaedic Literature. J Bone Joint Surg Am 2017, 99, 1769-1779. [CrossRef]
- Nueraihemaiti, N.; Dilimulati, D.; Baishan, A.; Hailati, S.; Maihemuti, N.; Aikebaier, A.; Paerhati, Y.; Zhou, W. Advances in Plant-Derived Extracellular Vesicle Extraction Methods and Pharmacological Effects. Biology (Basel) 2025, 14. [CrossRef]
- Tanaka, M. Special Issue "Translating Molecular Psychiatry: From Biomarkers to Personalized Therapies". Int J Mol Sci 2025, 26. [CrossRef]
- Lezoul, N.E.H.; Belkadi, M.; Habibi, F.; Guillén, F. Extraction Processes with Several Solvents on Total Bioactive Compounds in Different Organs of Three Medicinal Plants. Molecules 2020, 25. [CrossRef]
- Mew, E.J.; Monsour, A.; Saeed, L.; Santos, L.; Patel, S.; Courtney, D.B.; Watson, P.N.; Szatmari, P.; Offringa, M.; Monga, S.; et al. Systematic scoping review identifies heterogeneity in outcomes measured in adolescent depression clinical trials. J Clin Epidemiol 2020, 126, 71-79. [CrossRef]
- Centanni, M.; Friberg, L.E. Model-Based Biomarker Selection for Dose Individualization of Tyrosine-Kinase Inhibitors. Front Pharmacol 2020, 11, 316. [CrossRef]
- Yap, C.; Solovyeva, O.; de Bono, J.; Rekowski, J.; Patel, D.; Jaki, T.; Mander, A.; Evans, T.R.J.; Peck, R.; Hayward, K.S.; et al. Enhancing reporting quality and impact of early phase dose-finding clinical trials: CONSORT Dose-finding Extension (CONSORT-DEFINE) guidance. Bmj 2023, 383, e076387. [CrossRef]
- Carini, C.; Seyhan, A.A. Tribulations and future opportunities for artificial intelligence in precision medicine. J Transl Med 2024, 22, 411. [CrossRef]
- Chen, X.; He, R.; Chen, X.; Jiang, L.; Wang, F. Optimizing dose-schedule regimens with bayesian adaptive designs: opportunities and challenges. Front Pharmacol 2023, 14, 1261312. [CrossRef]
- Cottrell, T.R.; Lotze, M.T.; Ali, A.; Bifulco, C.B.; Capitini, C.M.; Chow, L.Q.M.; Cillo, A.R.; Collyar, D.; Cope, L.; Deutsch, J.S.; et al. Society for Immunotherapy of Cancer (SITC) consensus statement on essential biomarkers for immunotherapy clinical protocols. J Immunother Cancer 2025, 13. [CrossRef]
- Ciobanu-Caraus, O.; Aicher, A.; Kernbach, J.M.; Regli, L.; Serra, C.; Staartjes, V.E. A critical moment in machine learning in medicine: on reproducible and interpretable learning. Acta Neurochir (Wien) 2024, 166, 14. [CrossRef]
- Kim, J.W.; Choi, E.C.; Lee, K.J. Standardizing the approach to clinical-based human microbiome research: from clinical information collection to microbiome profiling and human resource utilization. Osong Public Health Res Perspect 2025, 16, 300-307. [CrossRef]
- Sharifi, S.; Mahmoud, N.N.; Voke, E.; Landry, M.P.; Mahmoudi, M. Importance of Standardizing Analytical Characterization Methodology for Improved Reliability of the Nanomedicine Literature. Nanomicro Lett 2022, 14, 172. [CrossRef]
- Arroyo-Araujo, M.; Voelkl, B.; Laloux, C.; Novak, J.; Koopmans, B.; Waldron, A.M.; Seiffert, I.; Stirling, H.; Aulehner, K.; Janhunen, S.K.; et al. Systematic assessment of the replicability and generalizability of preclinical findings: Impact of protocol harmonization across laboratory sites. PLoS Biol 2022, 20, e3001886. [CrossRef]
- Lukić, I.; Ivković, S.; Mitić, M.; Adžić, M. Tryptophan metabolites in depression: Modulation by gut microbiota. Front Behav Neurosci 2022, 16, 987697. [CrossRef]
- Hoepner, C.T.; McIntyre, R.S.; Papakostas, G.I. Impact of Supplementation and Nutritional Interventions on Pathogenic Processes of Mood Disorders: A Review of the Evidence. Nutrients 2021, 13. [CrossRef]
- Thomson, J.; Rankin, H.; Ashcroft, G.W.; Yates, C.M.; McQueen, J.K.; Cummings, S.W. The treatment of depression in general practice: a comparison of L-tryptophan, amitriptyline, and a combination of L-tryptophan and amitriptyline with placebo. Psychol Med 1982, 12, 741-751. [CrossRef]
- Kazemi, A.; Noorbala, A.A.; Azam, K.; Eskandari, M.H.; Djafarian, K. Effect of probiotic and prebiotic vs. placebo on psychological outcomes in patients with major depressive disorder: A randomized clinical trial. Clin Nutr 2019, 38, 522-528. [CrossRef]
- Jia, L.; Xiao, L.; Fu, Y.; Shao, Z.; Jing, Z.; Yuan, J.; Xie, Y.; Guo, J.; Wang, Y.; Geng, W. Neuroprotective effects of probiotics on anxiety- and depression-like disorders in stressed mice by modulating tryptophan metabolism and the gut microbiota. Food Funct 2024, 15, 2895-2905. [CrossRef]
- Donoso, F.; Cryan, J.F.; Olavarría-Ramírez, L.; Nolan, Y.M.; Clarke, G. Inflammation, Lifestyle Factors, and the Microbiome-Gut-Brain Axis: Relevance to Depression and Antidepressant Action. Clin Pharmacol Ther 2023, 113, 246-259. [CrossRef]
- Nikdasti, A.; Khodadadi, E.S.; Ferdosi, F.; Dadgostar, E.; Yahyazadeh, S.; Heidari, P.; Ehtiati, S.; Vakili, O.; Khatami, S.H. Nutritional Strategies in Major Depression Disorder: From Ketogenic Diet to Modulation of the Microbiota-Gut-Brain Axis. Mol Neurobiol 2025, 62, 2973-2994. [CrossRef]
- Merino Del Portillo, M.; Clemente-Suárez, V.J.; Ruisoto, P.; Jimenez, M.; Ramos-Campo, D.J.; Beltran-Velasco, A.I.; Martínez-Guardado, I.; Rubio-Zarapuz, A.; Navarro-Jiménez, E.; Tornero-Aguilera, J.F. Nutritional Modulation of the Gut-Brain Axis: A Comprehensive Review of Dietary Interventions in Depression and Anxiety Management. Metabolites 2024, 14. [CrossRef]
- Parletta, N.; Zarnowiecki, D.; Cho, J.; Wilson, A.; Bogomolova, S.; Villani, A.; Itsiopoulos, C.; Niyonsenga, T.; Blunden, S.; Meyer, B.; et al. A Mediterranean-style dietary intervention supplemented with fish oil improves diet quality and mental health in people with depression: A randomized controlled trial (HELFIMED). Nutr Neurosci 2019, 22, 474-487. [CrossRef]
- Opie, R.S.; O'Neil, A.; Jacka, F.N.; Pizzinga, J.; Itsiopoulos, C. A modified Mediterranean dietary intervention for adults with major depression: Dietary protocol and feasibility data from the SMILES trial. Nutr Neurosci 2018, 21, 487-501. [CrossRef]
- Firth, J.; Marx, W.; Dash, S.; Carney, R.; Teasdale, S.B.; Solmi, M.; Stubbs, B.; Schuch, F.B.; Carvalho, A.F.; Jacka, F.; et al. The Effects of Dietary Improvement on Symptoms of Depression and Anxiety: A Meta-Analysis of Randomized Controlled Trials. Psychosom Med 2019, 81, 265-280. [CrossRef]
- Xu, Y.; Zeng, L.; Zou, K.; Shan, S.; Wang, X.; Xiong, J.; Zhao, L.; Zhang, L.; Cheng, G. Role of dietary factors in the prevention and treatment for depression: an umbrella review of meta-analyses of prospective studies. Transl Psychiatry 2021, 11, 478. [CrossRef]
- Schefft, C.; Kilarski, L.L.; Bschor, T.; Köhler, S. Efficacy of adding nutritional supplements in unipolar depression: A systematic review and meta-analysis. Eur Neuropsychopharmacol 2017, 27, 1090-1109. [CrossRef]
- Thurfah, J.N.; Christine; Bagaskhara, P.P.; Alfian, S.D.; Puspitasari, I.M. Dietary Supplementations and Depression. J Multidiscip Healthc 2022, 15, 1121-1141. [CrossRef]
- Sarris, J.; Murphy, J.; Mischoulon, D.; Papakostas, G.I.; Fava, M.; Berk, M.; Ng, C.H. Adjunctive Nutraceuticals for Depression: A Systematic Review and Meta-Analyses. Am J Psychiatry 2016, 173, 575-587. [CrossRef]
- Mötteli, S.; Vetter, S.; Colla, M.; Hotzy, F. Are probiotics effective in reducing the metabolic side effects of psychiatric medication? A scoping review of evidence from clinical studies. Transl Psychiatry 2024, 14, 26. [CrossRef]
- Jacka, F.N.; O'Neil, A.; Opie, R.; Itsiopoulos, C.; Cotton, S.; Mohebbi, M.; Castle, D.; Dash, S.; Mihalopoulos, C.; Chatterton, M.L.; et al. A randomised controlled trial of dietary improvement for adults with major depression (the 'SMILES' trial). BMC Med 2017, 15, 23. [CrossRef]
- Swainson, J.; Reeson, M.; Malik, U.; Stefanuk, I.; Cummins, M.; Sivapalan, S. Diet and depression: A systematic review of whole dietary interventions as treatment in patients with depression. J Affect Disord 2023, 327, 270-278. [CrossRef]
- Ljungberg, T.; Bondza, E.; Lethin, C. Evidence of the Importance of Dietary Habits Regarding Depressive Symptoms and Depression. Int J Environ Res Public Health 2020, 17. [CrossRef]
- Marx, W.; Lane, M.; Hockey, M.; Aslam, H.; Berk, M.; Walder, K.; Borsini, A.; Firth, J.; Pariante, C.M.; Berding, K.; et al. Diet and depression: exploring the biological mechanisms of action. Mol Psychiatry 2021, 26, 134-150. [CrossRef]
- Marx, W.; Moseley, G.; Berk, M.; Jacka, F. Nutritional psychiatry: the present state of the evidence. Proc Nutr Soc 2017, 76, 427-436. [CrossRef]
- Levitan, R.D.; Shen, J.H.; Jindal, R.; Driver, H.S.; Kennedy, S.H.; Shapiro, C.M. Preliminary randomized double-blind placebo-controlled trial of tryptophan combined with fluoxetine to treat major depressive disorder: antidepressant and hypnotic effects. J Psychiatry Neurosci 2000, 25, 337-346.
- Frost, R.; Zamri, A.; Mathew, S.; Salame, A.; Bhanu, C.; Bhamra, S.K.; Bazo-Alvarez, J.C.; Heinrich, M.; Walters, K. Understanding the research landscape of over-the-counter herbal products, dietary supplements, and medications evaluated for depressive symptoms in adults: a scoping review. Front Pharmacol 2025, 16, 1609605. [CrossRef]
- Simon, M.S.; Weidinger, E.; Burger, B.; Kisla, Y.; Niedeggen, J.; Thaler, P.; Zaudig, M.; Voderholzer, U.; Schwarz, M.; Müller, N. Tryptophan metabolites predict response after cognitive behavioral therapy for depression: A single-arm trial. Journal of Affective Disorders Reports 2023, 11, 100464.
- Kuwano, N.; Kato, T.A.; Setoyama, D.; Sato-Kasai, M.; Shimokawa, N.; Hayakawa, K.; Ohgidani, M.; Sagata, N.; Kubo, H.; Kishimoto, J.; et al. Tryptophan-kynurenine and lipid related metabolites as blood biomarkers for first-episode drug-naïve patients with major depressive disorder: An exploratory pilot case-control study. J Affect Disord 2018, 231, 74-82. [CrossRef]
- Sha, Q.; Madaj, Z.; Keaton, S.; Escobar Galvis, M.L.; Smart, L.; Krzyzanowski, S.; Fazleabas, A.T.; Leach, R.; Postolache, T.T.; Achtyes, E.D.; et al. Cytokines and tryptophan metabolites can predict depressive symptoms in pregnancy. Transl Psychiatry 2022, 12, 35. [CrossRef]
- von Glischinski, M.; von Brachel, R.; Thiele, C.; Hirschfeld, G. Not sad enough for a depression trial? A systematic review of depression measures and cut points in clinical trial registrations. J Affect Disord 2021, 292, 36-44. [CrossRef]
- Dawson, G.R.; Post, A.; Smart, T.S.; Browning, M.; Harmer, C.J. Accuracy in recognising happy facial expressions is associated with antidepressant response to a NOP receptor antagonist but not placebo treatment. J Psychopharmacol 2021, 35, 1473-1478. [CrossRef]
- Hershenberg, R.; McDonald, W.M.; Crowell, A.; Riva-Posse, P.; Craighead, W.E.; Mayberg, H.S.; Dunlop, B.W. Concordance between clinician-rated and patient reported outcome measures of depressive symptoms in treatment resistant depression. J Affect Disord 2020, 266, 22-29. [CrossRef]
- Bain, E.E.; Shafner, L.; Walling, D.P.; Othman, A.A.; Chuang-Stein, C.; Hinkle, J.; Hanina, A. Use of a Novel Artificial Intelligence Platform on Mobile Devices to Assess Dosing Compliance in a Phase 2 Clinical Trial in Subjects With Schizophrenia. JMIR Mhealth Uhealth 2017, 5, e18. [CrossRef]
- Steinkamp, J.M.; Goldblatt, N.; Borodovsky, J.T.; LaVertu, A.; Kronish, I.M.; Marsch, L.A.; Schuman-Olivier, Z. Technological Interventions for Medication Adherence in Adult Mental Health and Substance Use Disorders: A Systematic Review. JMIR Ment Health 2019, 6, e12493. [CrossRef]
- Straczkiewicz, M.; Wisniewski, H.; Carlson, K.W.; Heidary, Z.; Knights, J.; Keshavan, M.; Onnela, J.P.; Torous, J. Combining digital pill and smartphone data to quantify medication adherence in an observational psychiatric pilot study. Psychiatry Res 2022, 315, 114707. [CrossRef]
- Rutherford, B.R.; Sneed, J.R.; Roose, S.P. Does study design influence outcome?. The effects of placebo control and treatment duration in antidepressant trials. Psychother Psychosom 2009, 78, 172-181. [CrossRef]
- Henssler, J.; Kurschus, M.; Franklin, J.; Bschor, T.; Baethge, C. Long-Term Acute-Phase Treatment With Antidepressants, 8 Weeks and Beyond: A Systematic Review and Meta-Analysis of Randomized, Placebo-Controlled Trials. J Clin Psychiatry 2018, 79. [CrossRef]
- Owoyemi, E.P.; Agwaza, K.P.; Doris, O.K.; Egwuatu, D.C.; Ogunye, R.O.; Anene, P.C.; Azubuike, E.O.; Ojobor, J.F.C.; Odanibeh, D.; Asenuga, O.O. Nutritional Genomics: The Use of Personalized Diets for Managing Depression. Journal of Complementary and Alternative Medical Research 2024, 25, 23-37.
- Singar, S.; Nagpal, R.; Arjmandi, B.H.; Akhavan, N.S. Personalized Nutrition: Tailoring Dietary Recommendations through Genetic Insights. Nutrients 2024, 16. [CrossRef]
- Julian, C.; Shen, N.; Molusky, M.; Hu, L.; Gopu, V.; Gorakshakar, A.; Patridge, E.; Antoine, G.; Connell, J.; Keiser, H. The effectiveness of precision supplements on depression symptoms in a US population. medRxiv 2023, 2023.2004. 2018.23288750.
- Zelada, M.I.; Garrido, V.; Liberona, A.; Jones, N.; Zúñiga, K.; Silva, H.; Nieto, R.R. Brain-Derived Neurotrophic Factor (BDNF) as a Predictor of Treatment Response in Major Depressive Disorder (MDD): A Systematic Review. Int J Mol Sci 2023, 24. [CrossRef]
- Suktas, A.; Ekalaksananan, T.; Aromseree, S.; Bumrungthai, S.; Songserm, N.; Pientong, C. Genetic polymorphism involved in major depressive disorder: a systemic review and meta-analysis. BMC Psychiatry 2024, 24, 716. [CrossRef]
- Ugartemendia, L.; Bravo, R.; Reuter, M.; Castaño, M.Y.; Plieger, T.; Zamoscik, V.; Kirsch, P.; Rodríguez, A.B. SLC6A4 polymorphisms modulate the efficacy of a tryptophan-enriched diet on age-related depression and social cognition. Clin Nutr 2021, 40, 1487-1494. [CrossRef]
- van Dalfsen, J.H.; Markus, C.R. The serotonin transporter gene-linked polymorphic region (5-HTTLPR) and the sleep-promoting effects of tryptophan: A randomized placebo-controlled crossover study. J Psychopharmacol 2019, 33, 948-954. [CrossRef]
- Nedic Erjavec, G.; Sagud, M.; Nikolac Perkovic, M.; Svob Strac, D.; Konjevod, M.; Tudor, L.; Uzun, S.; Pivac, N. Depression: Biological markers and treatment. Prog Neuropsychopharmacol Biol Psychiatry 2021, 105, 110139. [CrossRef]
- Milic, J.; Jovic, S.; Sapic, R. Advancing Depression Management Through Biomarker Discovery with a Focus on Genetic and Epigenetic Aspects: A Comprehensive Study on Neurobiological, Neuroendocrine, Metabolic, and Inflammatory Pathways. Genes (Basel) 2025, 16. [CrossRef]
- Beer, C.; Rae, F.; Semmler, A.; Voisey, J. Biomarkers in the Diagnosis and Prediction of Medication Response in Depression and the Role of Nutraceuticals. Int J Mol Sci 2024, 25. [CrossRef]
- Magzal, F.; Turroni, S.; Fabbrini, M.; Barone, M.; Vitman Schorr, A.; Ofran, A.; Tamir, S. A personalized diet intervention improves depression symptoms and changes microbiota and metabolite profiles among community-dwelling older adults. Front Nutr 2023, 10, 1234549. [CrossRef]
- Chen, L.; Liu, B.; Ren, L.; Du, H.; Fei, C.; Qian, C.; Li, B.; Zhang, R.; Liu, H.; Li, Z.; et al. High-fiber diet ameliorates gut microbiota, serum metabolism and emotional mood in type 2 diabetes patients. Front Cell Infect Microbiol 2023, 13, 1069954. [CrossRef]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut Microbiota-Derived Tryptophan Metabolites Modulate Inflammatory Response in Hepatocytes and Macrophages. Cell Rep 2018, 23, 1099-1111. [CrossRef]
- Mugo Moses, H. The Impact of Personalized Dietary Interventions on Glycemic Control and Quality of Life in Middle-Aged Adults with Type 2 Diabetes and Co-occurring Depression.
- Radford-Smith, D.E.; Anthony, D.C. Prebiotic and Probiotic Modulation of the Microbiota-Gut-Brain Axis in Depression. Nutrients 2023, 15. [CrossRef]
- Gao, K.; Mu, C.L.; Farzi, A.; Zhu, W.Y. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv Nutr 2020, 11, 709-723. [CrossRef]
- Asad, A.; Kirk, M.; Zhu, S.; Dong, X.; Gao, M. Effects of Prebiotics and Probiotics on Symptoms of Depression and Anxiety in Clinically Diagnosed Samples: Systematic Review and Meta-analysis of Randomized Controlled Trials. Nutr Rev 2025, 83, e1504-e1520. [CrossRef]
- Liu, R.T.; Walsh, R.F.L.; Sheehan, A.E. Prebiotics and probiotics for depression and anxiety: A systematic review and meta-analysis of controlled clinical trials. Neurosci Biobehav Rev 2019, 102, 13-23. [CrossRef]
- Bankah, A.Z.; Tagoe, T.A.; Darko, E.; Agoha, R.; Ametefe, E.N.; Kukuia, K.K.E.; Adjei, S. Combined Administration of Lactobacillus or Bifidobacterium Offers Enhanced Antidepressant and Anxiolytic Activity in a Dose Dependent Manner. Brain Behav 2025, 15, e70564. [CrossRef]
- Dandekar, M.P.; Palepu, M.S.K.; Satti, S.; Jaiswal, Y.; Singh, A.A.; Dash, S.P.; Gajula, S.N.R.; Sonti, R. Multi-strain Probiotic Formulation Reverses Maternal Separation and Chronic Unpredictable Mild Stress-Generated Anxiety- and Depression-like Phenotypes by Modulating Gut Microbiome-Brain Activity in Rats. ACS Chem Neurosci 2022, 13, 1948-1965. [CrossRef]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology 2017, 112, 399-412. [CrossRef]
- Fond, G.B.; Lagier, J.C.; Honore, S.; Lancon, C.; Korchia, T.; Sunhary De Verville, P.L.; Llorca, P.M.; Auquier, P.; Guedj, E.; Boyer, L. Microbiota-Orientated Treatments for Major Depression and Schizophrenia. Nutrients 2020, 12. [CrossRef]
- Chang, M.; Chang, K.T.; Chang, F. Just a gut feeling: Faecal microbiota transplant for treatment of depression - A mini-review. J Psychopharmacol 2024, 38, 353-361. [CrossRef]
- Hu, B.; Das, P.; Lv, X.; Shi, M.; Aa, J.; Wang, K.; Duan, L.; Gilbert, J.A.; Nie, Y.; Wu, X.L. Effects of 'Healthy' Fecal Microbiota Transplantation against the Deterioration of Depression in Fawn-Hooded Rats. mSystems 2022, 7, e0021822. [CrossRef]
- Rao, J.; Qiao, Y.; Xie, R.; Lin, L.; Jiang, J.; Wang, C.; Li, G. Fecal microbiota transplantation ameliorates stress-induced depression-like behaviors associated with the inhibition of glial and NLRP3 inflammasome in rat brain. Journal of Psychiatric Research 2021, 137, 147-157.
- Rao, J.; Xie, R.; Lin, L.; Jiang, J.; Du, L.; Zeng, X.; Li, G.; Wang, C.; Qiao, Y. Fecal microbiota transplantation ameliorates gut microbiota imbalance and intestinal barrier damage in rats with stress-induced depressive-like behavior. Eur J Neurosci 2021, 53, 3598-3611. [CrossRef]
- Doll, J.P.K.; Vázquez-Castellanos, J.F.; Schaub, A.C.; Schweinfurth, N.; Kettelhack, C.; Schneider, E.; Yamanbaeva, G.; Mählmann, L.; Brand, S.; Beglinger, C.; et al. Fecal Microbiota Transplantation (FMT) as an Adjunctive Therapy for Depression-Case Report. Front Psychiatry 2022, 13, 815422. [CrossRef]
- Chinna Meyyappan, A.; Forth, E.; Wallace, C.J.K.; Milev, R. Effect of fecal microbiota transplant on symptoms of psychiatric disorders: a systematic review. BMC Psychiatry 2020, 20, 299. [CrossRef]
- Cai, T.; Shi, X.; Yuan, L.Z.; Tang, D.; Wang, F. Fecal microbiota transplantation in an elderly patient with mental depression. Int Psychogeriatr 2019, 31, 1525-1526. [CrossRef]
Figure 1.
Integrated overview of major depressive disorder burden and plant-derived modulation of tryptophan pathways. This schematic summarizes the clinical burden of major depressive disorder (MDD) and highlights limitations of current selective serotonin reuptake inhibitor (SSRI) therapies, including delayed onset, sexual dysfunction, weight gain, hyponatremia, emotional blunting, and treatment resistance. Tryptophan metabolism is illustrated as a fork between the serotonin (5-HT) pathway (promoting mood regulation and emotional balance) and the kynurenine pathway (associated with inflammation, neurotoxicity, and melancholic features). Inflammatory cytokines such as interleukin (IL-6) and interferon (IFN)-γ shift tryptophan away from 5-HT production by activating Indoleamine 2,3-dioxygenase (IDO), contributing to reduced 5-HT, anhedonia, low mood, and suicidality. Mechanistic points where plant-derived compounds may intervene are shown on the right, including enhancement of Tryptophan hydroxylase (TPH) activity and 5-HT synthesis, inhibition of IDO1 activation to reduce neurotoxic kynurenine metabolites, and modulation of the gut microbiota to increase beneficial indoles and reduce inflammation. Collectively, these actions increase bioavailable tryptophan and 5-HTP, offering potential complementary strategies for improving depressive symptoms. Created using Mind the Graph (https://mindthegraph.com/, last accessed on November 2025). Abbreviations: Trp, Tryptophan; 5-HTP, 5-Hydroxytryptophan; KYN, Kynurenine; QA, Quinolinic acid.
Figure 1.
Integrated overview of major depressive disorder burden and plant-derived modulation of tryptophan pathways. This schematic summarizes the clinical burden of major depressive disorder (MDD) and highlights limitations of current selective serotonin reuptake inhibitor (SSRI) therapies, including delayed onset, sexual dysfunction, weight gain, hyponatremia, emotional blunting, and treatment resistance. Tryptophan metabolism is illustrated as a fork between the serotonin (5-HT) pathway (promoting mood regulation and emotional balance) and the kynurenine pathway (associated with inflammation, neurotoxicity, and melancholic features). Inflammatory cytokines such as interleukin (IL-6) and interferon (IFN)-γ shift tryptophan away from 5-HT production by activating Indoleamine 2,3-dioxygenase (IDO), contributing to reduced 5-HT, anhedonia, low mood, and suicidality. Mechanistic points where plant-derived compounds may intervene are shown on the right, including enhancement of Tryptophan hydroxylase (TPH) activity and 5-HT synthesis, inhibition of IDO1 activation to reduce neurotoxic kynurenine metabolites, and modulation of the gut microbiota to increase beneficial indoles and reduce inflammation. Collectively, these actions increase bioavailable tryptophan and 5-HTP, offering potential complementary strategies for improving depressive symptoms. Created using Mind the Graph (https://mindthegraph.com/, last accessed on November 2025). Abbreviations: Trp, Tryptophan; 5-HTP, 5-Hydroxytryptophan; KYN, Kynurenine; QA, Quinolinic acid.
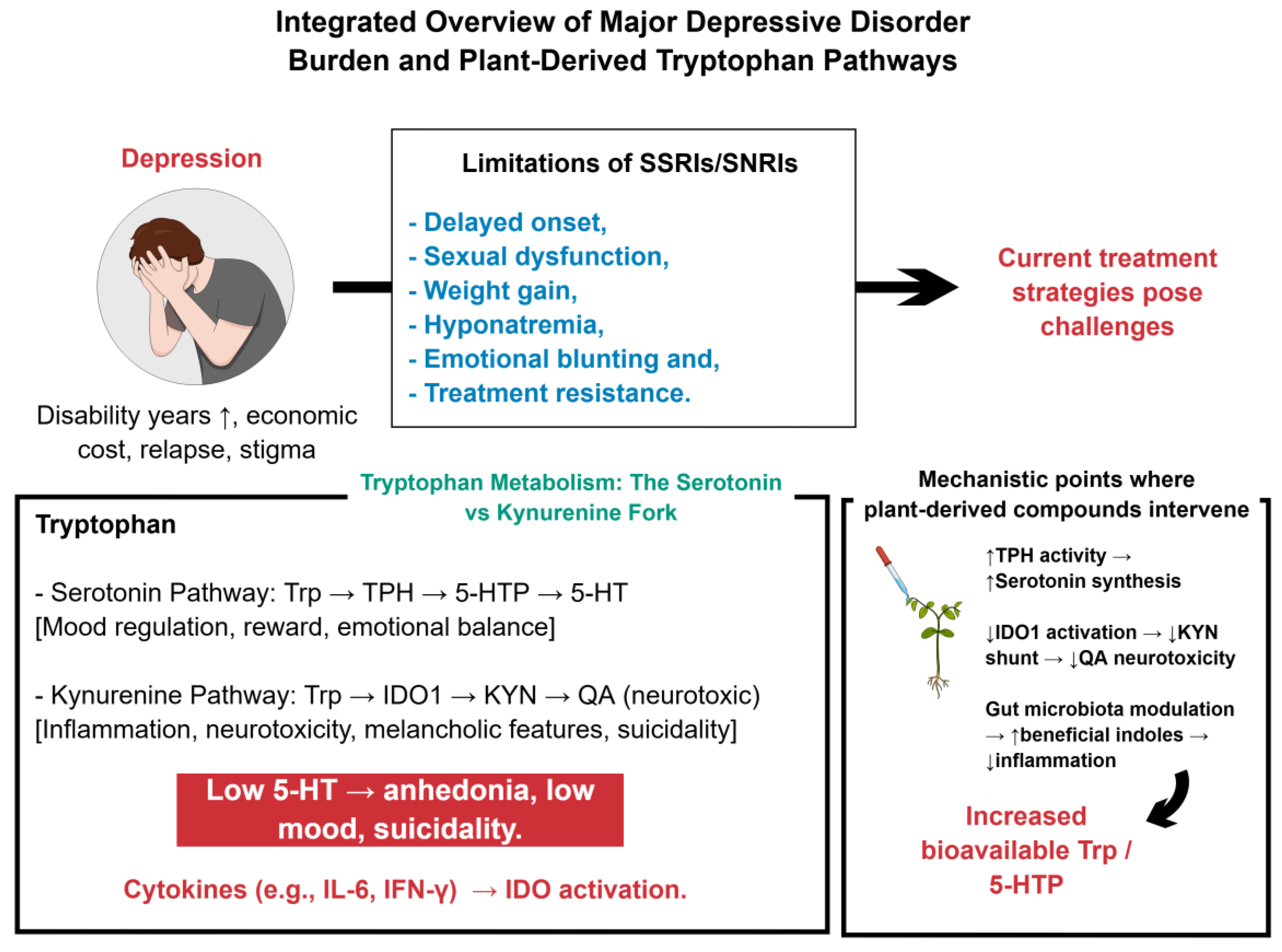
Figure 2.
Tryptophan (Trp) metabolism through the kynurenine (KYN) pathway: balancing neuroprotection and neurotoxicity in depression. This figure illustrates how immune-mediated activation of the KYN pathway disrupts Trp metabolism, linking inflammation with depression-related neuropathology. The stress hormone cortisol stimulates tryptophan 2,3-dioxygenase (TDO). Pro-inflammatory cytokines [interferon (IFN)-alpha (α), IFN-beta (ẞ), IFN-gamma (γ), tumor necrosis factor (TNF)-α] and lipopolysaccharide (LPS) induce indoleamine 2,3-dioxygenases (IDOs), diverting Trp away from serotonin (5-HT) synthesis toward KYN production. In contrast, anti-inflammatory cytokines [interleukin (IL)-4, IL-10] and superoxide dismutase (SOD) counteract this process. Oxygen molecule (O2), IFN-ẞ, IFN-γ, TNF-α stimulate and IL-4, IL-10, SOD suppress kynurenine 3-monooxydase (KMO). Furthermore, IDOs, TDO, and KMO activities are influenced by positive and negative feedback loops. Peripheral KYN crosses the blood-brain barrier, where it undergoes two competing fates: (i) conversion into kynurenic acid (KYNA), a putative neuroprotective N-Methyl-D-Aspartate (NMDA) receptor antagonist with putative antioxidant properties, or (ii) metabolism into 3-hydroxykynurenine (3-HK) and quinolinic acid (QA), which promotes reactive oxygen species (ROS), NMDA receptor overstimulation, mitochondrial dysfunction, and neuronal apoptosis, ultimately leading to neurodegeneration. The dashed inhibitory arrow indicates that enhanced KYN metabolism reduces 5-HT and melatonin synthesis, impairing neurotransmission and neuroprotection. The balance between KYNA and QA determines whether the KYN pathway promotes resilience or vulnerability to depression.
Figure 2.
Tryptophan (Trp) metabolism through the kynurenine (KYN) pathway: balancing neuroprotection and neurotoxicity in depression. This figure illustrates how immune-mediated activation of the KYN pathway disrupts Trp metabolism, linking inflammation with depression-related neuropathology. The stress hormone cortisol stimulates tryptophan 2,3-dioxygenase (TDO). Pro-inflammatory cytokines [interferon (IFN)-alpha (α), IFN-beta (ẞ), IFN-gamma (γ), tumor necrosis factor (TNF)-α] and lipopolysaccharide (LPS) induce indoleamine 2,3-dioxygenases (IDOs), diverting Trp away from serotonin (5-HT) synthesis toward KYN production. In contrast, anti-inflammatory cytokines [interleukin (IL)-4, IL-10] and superoxide dismutase (SOD) counteract this process. Oxygen molecule (O2), IFN-ẞ, IFN-γ, TNF-α stimulate and IL-4, IL-10, SOD suppress kynurenine 3-monooxydase (KMO). Furthermore, IDOs, TDO, and KMO activities are influenced by positive and negative feedback loops. Peripheral KYN crosses the blood-brain barrier, where it undergoes two competing fates: (i) conversion into kynurenic acid (KYNA), a putative neuroprotective N-Methyl-D-Aspartate (NMDA) receptor antagonist with putative antioxidant properties, or (ii) metabolism into 3-hydroxykynurenine (3-HK) and quinolinic acid (QA), which promotes reactive oxygen species (ROS), NMDA receptor overstimulation, mitochondrial dysfunction, and neuronal apoptosis, ultimately leading to neurodegeneration. The dashed inhibitory arrow indicates that enhanced KYN metabolism reduces 5-HT and melatonin synthesis, impairing neurotransmission and neuroprotection. The balance between KYNA and QA determines whether the KYN pathway promotes resilience or vulnerability to depression.
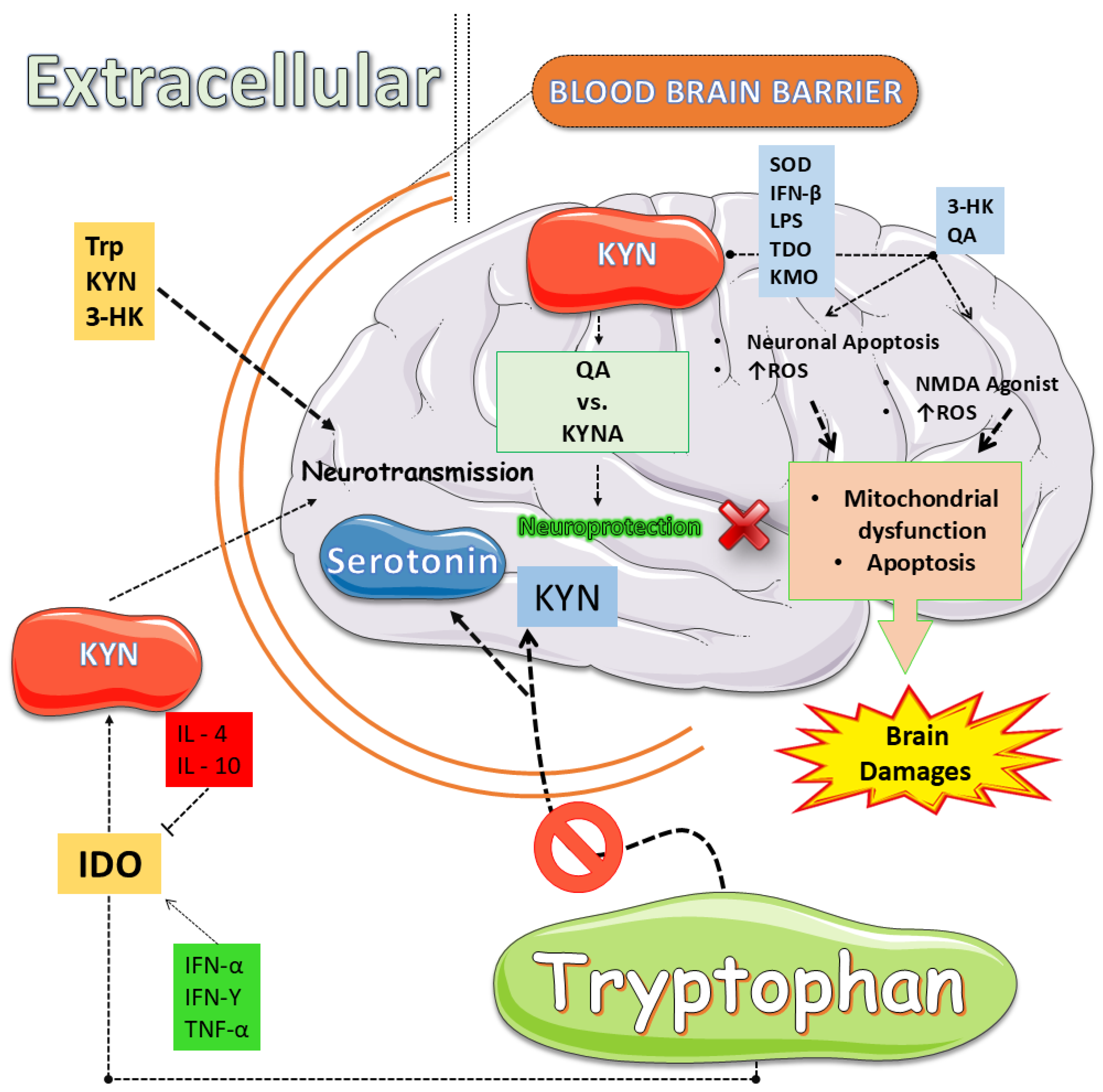
Figure 3.
Light– suprachiasmatic nucleus (SCN)–pineal pathway regulating melatonin biosynthesis. Light information from the retina reaches the suprachiasmatic nucleus (SCN), the master circadian pacemaker, via the retinohypothalamic tract. The SCN projects to the paraventricular nucleus and modulates sympathetic innervation of the pineal gland. In darkness, SCN activity stimulates pineal metabolism of Trp through a sequential enzymatic cascade: tryptophan hydroxylase (TPH) converts Trp into 5-hydroxytryptophan (5-HTP), aromatic L-amino acid decarboxylase (AADC) generates serotonin (5-HT), serotonin N-acetyltransferase (SNAT) produces N-acetylserotonin, and acetylserotonin O-methyltransferase (ASMT) catalyzes the final step to melatonin. Light exposure suppresses this pathway, reducing melatonin output. Melatonin acts as a key circadian regulator, influencing sleep, neuroendocrine rhythms, and mood regulation.
Figure 3.
Light– suprachiasmatic nucleus (SCN)–pineal pathway regulating melatonin biosynthesis. Light information from the retina reaches the suprachiasmatic nucleus (SCN), the master circadian pacemaker, via the retinohypothalamic tract. The SCN projects to the paraventricular nucleus and modulates sympathetic innervation of the pineal gland. In darkness, SCN activity stimulates pineal metabolism of Trp through a sequential enzymatic cascade: tryptophan hydroxylase (TPH) converts Trp into 5-hydroxytryptophan (5-HTP), aromatic L-amino acid decarboxylase (AADC) generates serotonin (5-HT), serotonin N-acetyltransferase (SNAT) produces N-acetylserotonin, and acetylserotonin O-methyltransferase (ASMT) catalyzes the final step to melatonin. Light exposure suppresses this pathway, reducing melatonin output. Melatonin acts as a key circadian regulator, influencing sleep, neuroendocrine rhythms, and mood regulation.
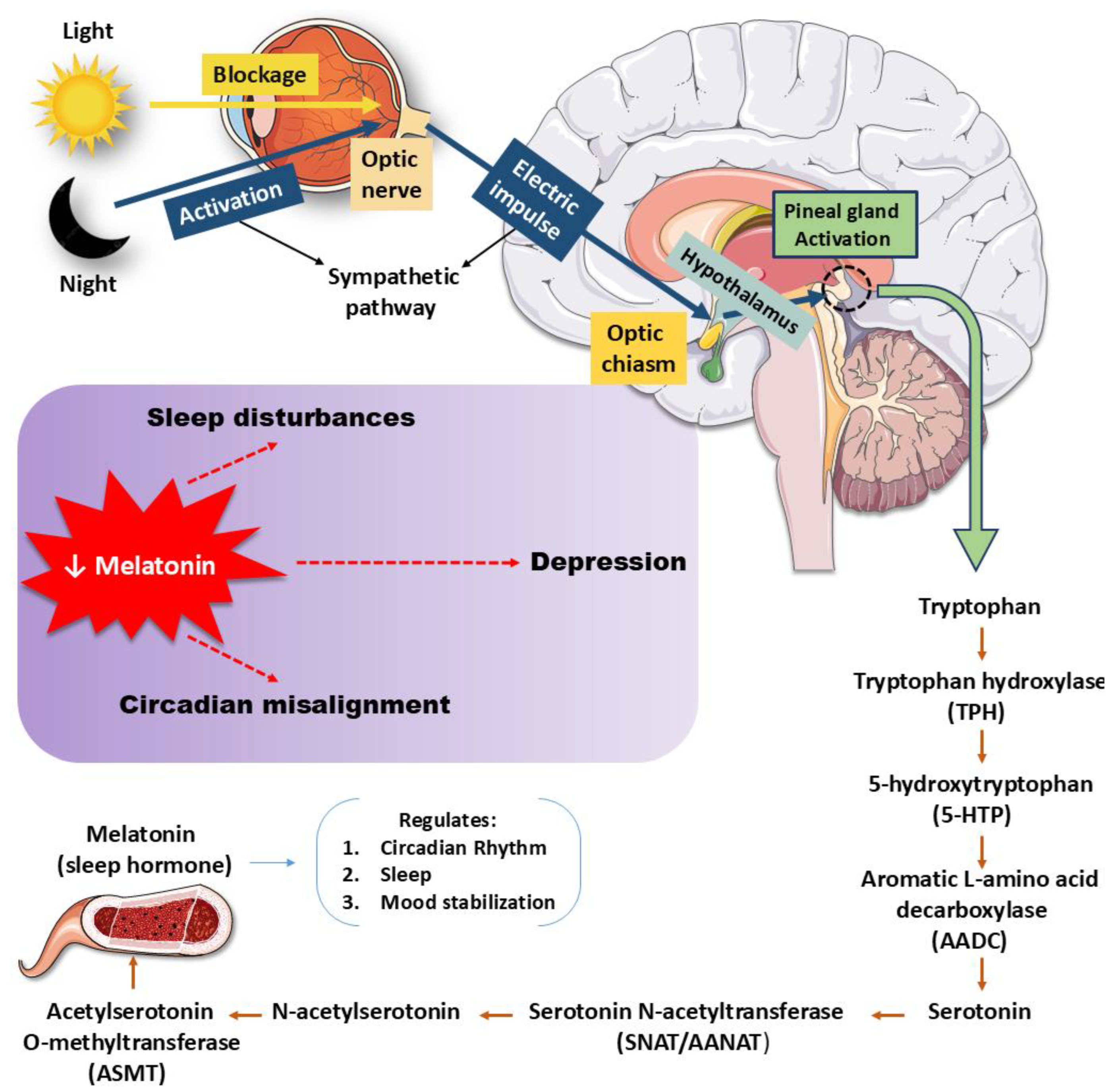
Figure 4.
Dietary tryptophan (Trp) metabolism by gut microbiota and its systemic effects. Exogenous dietary Trp, derived from protein-rich foods, is metabolized by intestinal microbiota into indole derivatives, including indole-3-aldehyde (IAld), indole-3-acetic acid (IAA), indole-3-propionic acid (IPA), and indoleacrylic acid. These metabolites enter the systemic circulation and exert pleiotropic effects on host physiology. IAld activates the aryl hydrocarbon receptor (AhR), modulating mucosal immunity, while IPA provides neuroprotection through antioxidant activity. IAA regulates intestinal motility and epithelial function, whereas indoleacrylic acid strengthens gut barrier integrity. Collectively, these microbial metabolites influence brain activity (neurotransmission, neuroinflammation, mood), bowel function (motility, barrier homeostasis), and the immune system (immune tolerance, inflammation). This highlights the central role of gut microbiota–Trp metabolism in the gut–brain–immune axis.
Figure 4.
Dietary tryptophan (Trp) metabolism by gut microbiota and its systemic effects. Exogenous dietary Trp, derived from protein-rich foods, is metabolized by intestinal microbiota into indole derivatives, including indole-3-aldehyde (IAld), indole-3-acetic acid (IAA), indole-3-propionic acid (IPA), and indoleacrylic acid. These metabolites enter the systemic circulation and exert pleiotropic effects on host physiology. IAld activates the aryl hydrocarbon receptor (AhR), modulating mucosal immunity, while IPA provides neuroprotection through antioxidant activity. IAA regulates intestinal motility and epithelial function, whereas indoleacrylic acid strengthens gut barrier integrity. Collectively, these microbial metabolites influence brain activity (neurotransmission, neuroinflammation, mood), bowel function (motility, barrier homeostasis), and the immune system (immune tolerance, inflammation). This highlights the central role of gut microbiota–Trp metabolism in the gut–brain–immune axis.
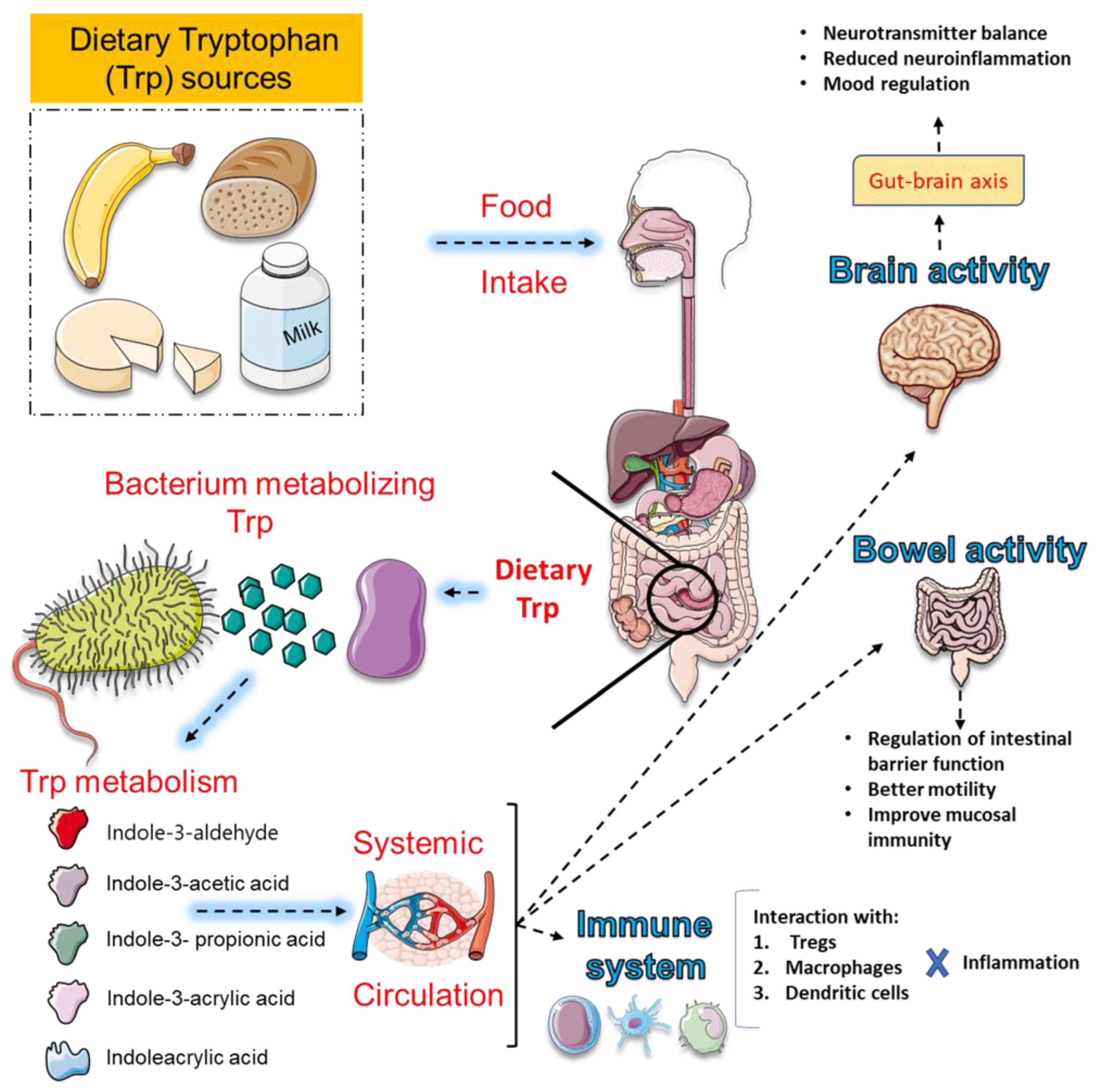
Figure 5.
Tryptophan (Trp) –indole metabolism and its pathological consequences. Gut microbiota such as Escherichia coli and Bacteroides spp. metabolize dietary Trp via tryptophanase into indole. Host and environmental factors (genetics, diet, stress, aging, physical activity) modulate this pathway. Dysregulated Trp metabolism reduces serotonin (5-HT) availability, impairing enteric nervous system signaling, metabolism, glucose absorption, and intestinal motility, contributing to chronic gastrointestinal disorders such as ulcerative colitis. Indole derivatives, particularly indoxyl sulfate (IS), accumulate when excretion is impaired, leading to systemic inflammation, oxidative stress, and cellular damage in the colon and liver. Overexpression of indoleamine 2,3-dioxygenase 1 (IDO1) and tryptophan 2,3-dioxygenase 2 (TDO2) further links altered Trp metabolism to carcinogenic processes in the brain, breast, and lungs. This highlights the central role of Trp–indole metabolism in the gut–brain–immune axis, cancer progression, and systemic disease. ENS, Enteric Nervous System.
Figure 5.
Tryptophan (Trp) –indole metabolism and its pathological consequences. Gut microbiota such as Escherichia coli and Bacteroides spp. metabolize dietary Trp via tryptophanase into indole. Host and environmental factors (genetics, diet, stress, aging, physical activity) modulate this pathway. Dysregulated Trp metabolism reduces serotonin (5-HT) availability, impairing enteric nervous system signaling, metabolism, glucose absorption, and intestinal motility, contributing to chronic gastrointestinal disorders such as ulcerative colitis. Indole derivatives, particularly indoxyl sulfate (IS), accumulate when excretion is impaired, leading to systemic inflammation, oxidative stress, and cellular damage in the colon and liver. Overexpression of indoleamine 2,3-dioxygenase 1 (IDO1) and tryptophan 2,3-dioxygenase 2 (TDO2) further links altered Trp metabolism to carcinogenic processes in the brain, breast, and lungs. This highlights the central role of Trp–indole metabolism in the gut–brain–immune axis, cancer progression, and systemic disease. ENS, Enteric Nervous System.
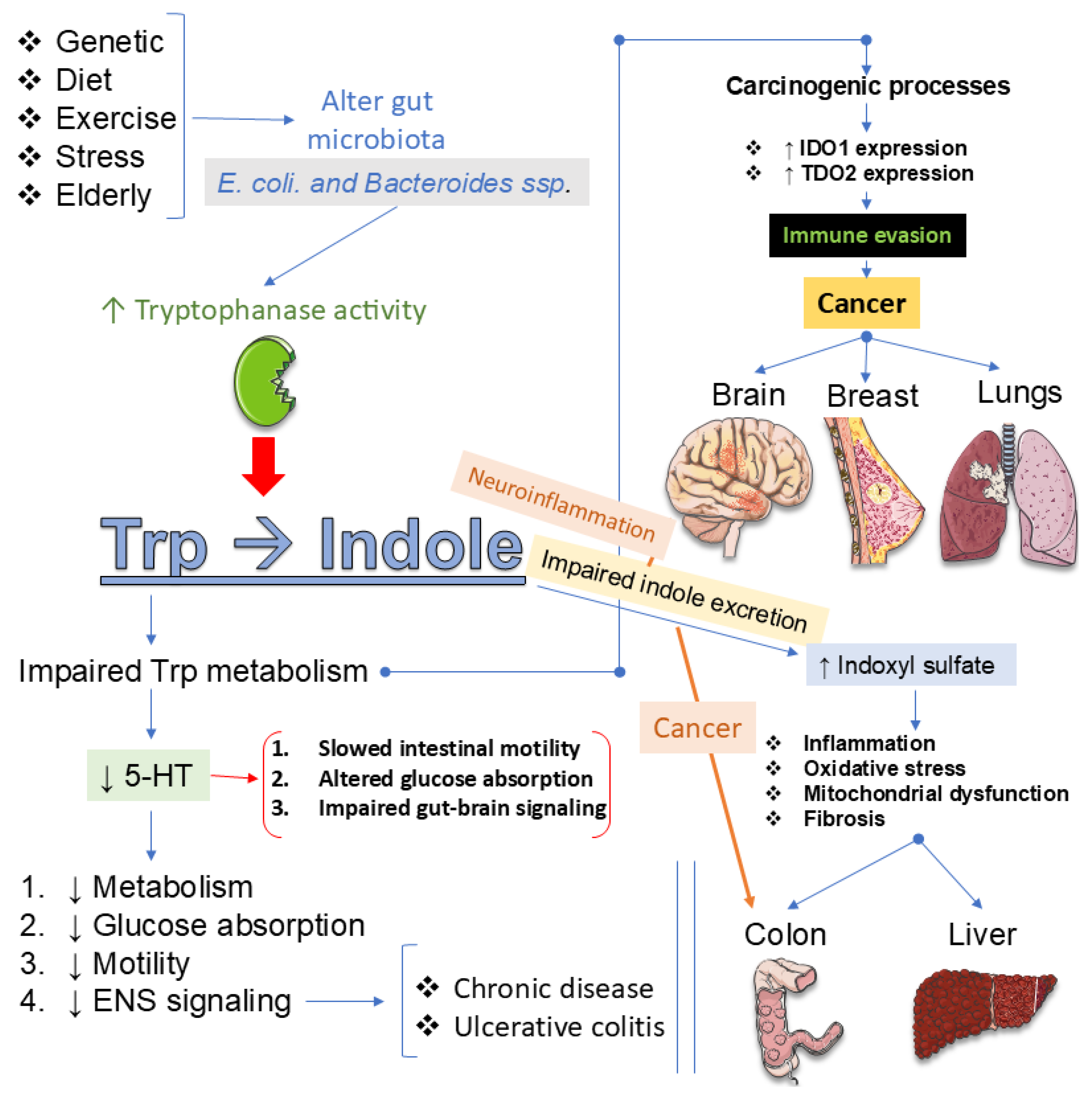
Figure 6.
Dual pathways of tryptophan (Trp) metabolism and their neuropsychiatric implications. Trp crosses the blood-brain barrier (BBB) via LAT1 transporters and follows two competing metabolic routes. One route converts Trp to serotonin (5-HT) through tryptophan hydroxylase (TPH), which subsequently gives rise to melatonin, regulating circadian rhythms and mood. Impaired 5-HT synthesis contributes to depression and sleep disturbances. Alternatively, Trp is metabolized through the kynurenine (KYN) pathway, primarily via indoleamine 2,3-dioxygenase (IDO). This generates metabolites with opposing actions: quinolinic acid (QA), a neurotoxic N-Methyl-D-Aspartate (NMDA) receptor agonist that promotes excitotoxicity, oxidative stress, and cognitive dysfunction, and kynurenic acid (KYNA), a neuroprotective NMDA receptor antagonist that counterbalances QA’s toxicity. Dysregulation of the Trp–5-HT–KYN axis disrupts neurotransmission, neuroprotection, and circadian control, ultimately contributing to depression, memory problems, and neurodegeneration.
Figure 6.
Dual pathways of tryptophan (Trp) metabolism and their neuropsychiatric implications. Trp crosses the blood-brain barrier (BBB) via LAT1 transporters and follows two competing metabolic routes. One route converts Trp to serotonin (5-HT) through tryptophan hydroxylase (TPH), which subsequently gives rise to melatonin, regulating circadian rhythms and mood. Impaired 5-HT synthesis contributes to depression and sleep disturbances. Alternatively, Trp is metabolized through the kynurenine (KYN) pathway, primarily via indoleamine 2,3-dioxygenase (IDO). This generates metabolites with opposing actions: quinolinic acid (QA), a neurotoxic N-Methyl-D-Aspartate (NMDA) receptor agonist that promotes excitotoxicity, oxidative stress, and cognitive dysfunction, and kynurenic acid (KYNA), a neuroprotective NMDA receptor antagonist that counterbalances QA’s toxicity. Dysregulation of the Trp–5-HT–KYN axis disrupts neurotransmission, neuroprotection, and circadian control, ultimately contributing to depression, memory problems, and neurodegeneration.
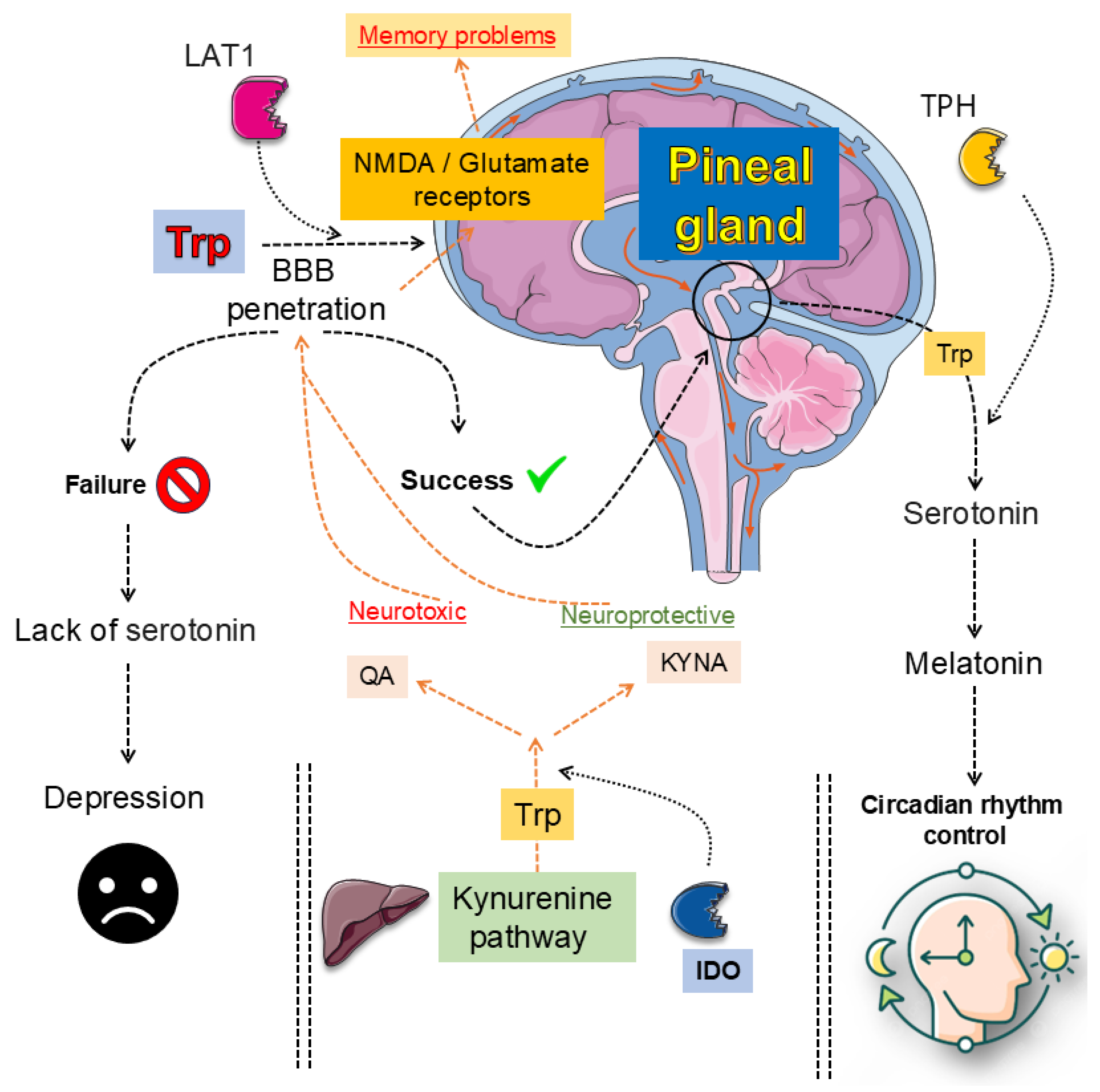
Figure 7.
Mechanistic actions of phytochemicals in depression. Phytochemicals, including 5-hydroxytryptophan (5-HTP), isoflavones, berberine, and polyphenols, exert central effects through modulation of the gut microbiome (via tryptophan (Trp) metabolism), regulation of immune signaling (attenuation of inflammation), and stimulation of neurogenesis (enhancement of plasticity and mood stabilization), collectively contributing to antidepressant outcomes.
Figure 7.
Mechanistic actions of phytochemicals in depression. Phytochemicals, including 5-hydroxytryptophan (5-HTP), isoflavones, berberine, and polyphenols, exert central effects through modulation of the gut microbiome (via tryptophan (Trp) metabolism), regulation of immune signaling (attenuation of inflammation), and stimulation of neurogenesis (enhancement of plasticity and mood stabilization), collectively contributing to antidepressant outcomes.
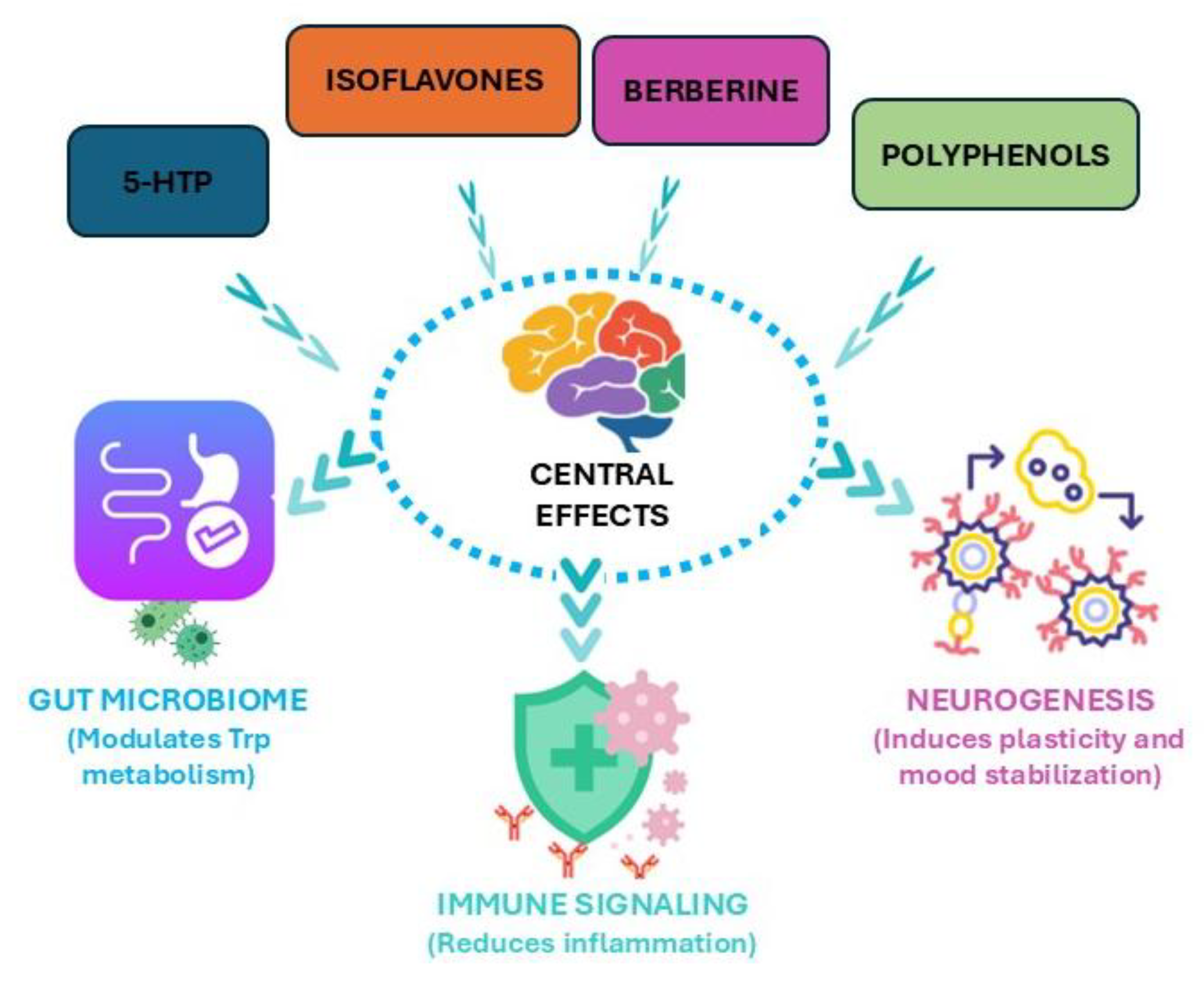

Table 1.
Summary of studies investigating dietary and non-dietary interventions in depression and related disorders and conditions. The table outlines study design, patient populations, interventions, primary outcomes, and key observations.
Table 1.
Summary of studies investigating dietary and non-dietary interventions in depression and related disorders and conditions. The table outlines study design, patient populations, interventions, primary outcomes, and key observations.
| Recovered/Remitted Depression | ||||
|---|---|---|---|---|
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| Crossover, DB, 23 recovered depressed vs. 20 healthy controls (NL) | α-lactalbumin (Trp-rich) vs. casein | Improved memory; impaired simple motor performance | No effect on mood | [224] |
| RCT, DB, crossover, 16 recovered-depressed subjects vs. 17 healthy subjects (UK and NZ) | ATD vs. placebo | No mood change overall | ATD decreased MMSE parameters in the recovered-depressed group | [225] |
| Patients with Depressive Symptoms/Depression | ||||
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| RCT, crossover, 30 subjects with MDD (MY) | Talbinah (Trp-rich barley) vs. control | ↓ depression and stress (DASS, POMS) | Multidomain improvement | [226] |
| RCT, DB, 60 subjects with MDD (IN) | 5-HTP vs. fluoxetine (8 wks) | Both ↓ HDRS (by wk 2) | Comparable efficacy | [227] |
| RCT, DB, 25 patients with Parkinson’s disease and depressive/apathy symptoms (IT) | 5-HTP (50 mg) vs. placebo (crossover: 4 wks + 4-wk washout + 4 wks) | ↓ depression (BDI-II, HDRS) | 5-HTP improved depressive symptoms but not apathy in Parkinson’s disease | [228] |
| High-Risk/Vulnerable Groups | ||||
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| Placebo-controlled, DB, 29 high stress-vulnerable vs. 29 low stress-invulnerable adults (NL) | α-lactalbumin (Trp-rich) vs. casein | Trp improved mood (POMS) and ↑Trp/LNAA ratio in the stress-vulnerable group, ↓ in cortisol in both groups | Antidepressant-like effect under stress | [229] |
| RCT, DB, 20 high vs. 18 low cognitive reactivity subjects (NL) | Trp-rich hydrolysate vs. casein | Trp improved positive mood, and ↓ cortisol after stress | Both groups benefited physiologically | [230] |
| Placebo-controlled, crossover, DB, 20 high-risk (family history) vs. 19 low-risk (CA) | ATD vs. control | High-risk: lowering mood (POMS) | ATD amplified vulnerability markers | [231] |
| Bulimia | ||||
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| RCT, DB, 26 unmedicated women with bulimia vs. 13 women with bulimia treated with SSRIs vs. 25 healthy control women (CA) | ATD vs. control (balanced amino acids) | ATD ↓ mood in all; ↑ urge to binge eat in bulimia + SSRIs | Serotonin depletion linked to bulimic relapse, especially with SSRI use | [232] |
| Healthy Volunteers | ||||
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| RCT, DB, 38 healthy adults (UK) | Trp (1g, 3x/day, 14d) vs. placebo | Trp increased recognition of happy faces, reduced recognition of disgusted expressions (females) | Positive emotional bias in the processing of emotional material with stronger effects in women | [233] |
| RCT, DB, 20 healthy adults (CA, NL, and FR) | Trp (3g/day, 3wks) vs. placebo (crossover with a 2-wk washout period) | ↑ plasma/urine kynurenine & indoles; no effect on mood and no GI symptoms | Trp triggered immunomodulatory metabolic pathways | [234] |
| RCT, DB, 47 healthy adults (NL) | Trp (2.8g/day, 6d) vs. placebo | ↑ rejection of unfair offers | Prolonged Trp may affect social decision-making | [235] |
| RCT, DB, 18 healthy adults (NL) | Trp-rich hydrolyzed protein vs. α-lactalbumin (Trp-rich) vs. casein vs. pure Trp vs. Trp-containing synthetic peptide | Hydrolyzed Trp and pure Trp improved mood (acute); hydrolyzed Trp caused an increase in plasma Trp/LNAA faster and greater than α-lactalbumin | Hydrolyzed Trp is more efficient in increasing brain Trp and 5-HT function compared with α-lactalbumin and pure Trp | [236] |
| Elderly | ||||
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| Comparative study, SB, 35 elderly subjects (ES) | Trp-enriched cereals vs. standard cereals vs. usual diet | ↑ sleep quality, ↓ anxiety & depression | Improved age alterations on the sleep/wake cycle | [237] |
| RCT, DB, 25 elderly individuals with mild cognitive impairment (IT) | DHA-phospholipids + melatonin + Trp vs. placebo, 12 weeks | ↑ cognitive function, ↑ nutrition, ↑ olfactory sensitivity | MMSE improved, positive trend for verbal fluency | [238] |
| RCT, DB, 14 elderly subjects (NL) | Trp + nicotinic acid + nicotinamide vs. control, 32 days | No effect on mitochondrial or muscle function | ↑ NAD+ metabolism, no functional benefit | [239] |
| Open-label clinical trial, 40 elderly individuals with depression/sleep disorders (PL) | Trp-rich diet (25 mg/kg/d), 12 weeks | ↓ depression and insomnia scores, improved Trp metabolism | ISI and HDRS scores halved post-intervention | [240] |
| Fibromyalgia | ||||
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| RCT, 22 women with fibromyalgia (ES) | Mediterranean diet enriched with high-dose Trp (60 mg) and Mg (60 mg) vs. standard Mediterranean diet | ↓ anxiety, ↓ mood disturbance, ↓ eating disorder symptoms, ↓ body image dissatisfaction; no improvement in sleep quality | Significant improvements in psychological variables | [217] |
| Large-Scale Observational | ||||
| Study Design and Population | Intervention/Control | Main Outcomes | Key Notes | Ref. |
| Clinical trial, 29,687 adults (US) (NHANES 2001–2012) | Mean Trp intake: 826 mg/d (observational, no intervention) | Higher Trp intake associated with ↓ depression severity (PHQ-9), ↑ sleep duration | No adverse effects on liver, kidney, or metabolism | [241] |
5-HTP, 5-Hydroxytryptophan; ↑, Increase; ↓, Decrease; ATD, Acute Tryptophan Depletion; BDI-II, Beck Depression Inventory-II; CA, Canada; DASS, Depression Anxiety Stress Scales; DB, Double-Blind; DHA, Docosahexaenoic Acid; ES, Spain; FR, France; GI, Gastrointestinal; HDRS, Hamilton Depression Rating Scale; IN, India; ISI, Insomnia Severity Index; IT, Italy; LNAA, Large Neutral Amino Acids; Mg, Magnesium; MDD, Major Depressive Disorder; MMSE, Mini-Mental State Examination; MY, Malaysia; NL, Netherlands; NZ, New Zealand; PHQ, Patient Health Questionnaire-9; PL, Poland; POMS, Profile of Mood States; RCT, Randomized Controlled Trial; SB, Simple-Blind; SSRIs, Selective Serotonin Reuptake Inhibitors; Trp, Tryptophan; UK, United Kingdom; US, United States.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



