Submitted:
10 September 2025
Posted:
12 September 2025
You are already at the latest version
Abstract
Nitrogen is an essential element required for bacterial homeostsis. It serves as a building block for the biosynthesis of macromolecules and provides precursors for secondary metabolites. Actinomycetes have developed the ability to use various nitrogen sources and possess two central enzyme systems for nitrogen assimilation involving glutamine synthetase / glutamate synthase and glutamate dehydrogenase. Microorganisms living in habitats with changeable availability of nutrients have to cope with limited nitrogen availability in their ecological niches and developed strategies to survive under nitrogen limitation. A complex nitrogen metabolism of Actinobacteria allows the utilization of various compounds as N-sources. One such adaptation is the ability to acquire nitrogen from alternative sources like monoamines or polyamines putrescine, cadaverine, spermidine and spermine, ensuring both nutrients availability (C- and N-source) and resistance against high polyamine concentrations. Bacterial polyamine catabolism is not only important under low nitrogen availability, but it is also required to survive under high concentrations of these compounds. Such conditions can occur in diverse habitats like soil, plant tissues and human cells. Strategies of pathogenic and non-pathogenic Actinobacteria to survive in the presence of mono- and polyamines offer the possibility to combat pathogens by using their capability to metabolize polyamines as an antibiotic drug target. This work aims to summarize the knowledge on nitrogen utilization and, more specifically, catabolism of amines in actinobacterial survival and its role in nitrogen metabolism.
Keywords:
nitrogen metabolism
; polyamine utilization
; polyamine metabolism
; bacterial survival
; bacterial physiology
; Actinobacteria
1. Nitrogen Metabolism in Prokaryotes
In addition to carbon, oxygen, and hydrogen, the main component of all living cells is nitrogen (N). It makes up about 14% of the cell's dry weight (Fuchs, 2022). It is an essential component for bacterial metabolism needed for the synthesis of purines and pyrimidines that are the basic building blocks of DNA and RNA, of amino acids that are required for protein synthesis, and of amino sugars that are components of cell walls. The basic building blocks often serve as precursor molecules for the formation of secondary metabolites (Aharonowitz, 1980; Reitzer, 2001; (Ninfa et al., 2001), which may contain atoms of nitrogen. For example, Streptomyces produced molecules of the antibiotic undecylprodigiosin (Red) contain three nitrogen atoms, while the calcium-dependent antibiotic contains 14 N atoms (Hojati et al. 2002; Hopwood, 1988). The availability of nitrogen plays a crucial role in primary metabolism and in secondary metabolite formation (Krysenko & Wohlleben, 2025).
Bacteria from the phylum Actinobacteria can obtain the nitrogen they need for cellular metabolism from various sources. While soil bacteria can utilize both animal and plant remain, most enterobacteria depend on metabolites of animal metabolism. These include organic substances such as amino acids, creatinine, urea, or simple inorganic substances such as ammonium or nitrate (Neidhardt and Reitzer, 1996). The preferred nitrogen source for optimal growth in most bacteria is ammonium. For the uptake and utilization of nitrogen, bacteria have developed specific transporters along with mechanisms of assimilation and regulation. This allows them to respond quickly to changes in the environment and rapidly optimize their metabolism (Jacoby et al., 2020).
1.1. Nitrogen Uptake and Nitrogen Assimilation in Actinobacteria
Actinomycetes can utilize a wide range of substances as nitrogen sources. These include inorganic compounds such as ammonium, nitrate, and nitrite, as well as organic compounds, such as amino acids (e.g., histidine and arginine) and amino sugars (Magasanik, 1982; Hopwood, 1999). Nitrogen uptake in Actinobacteria has been extensively investigated in the model bacterium Streptomyces coelicolor. S. coelicolor lives in the soil under variable N- and C-conditions and therefore possesses the ability to metabolize a variety of different C- and N-sources, including amino sugars, amino acids, amines (mono- and polyamines), peptides, urea, NO3-, NH4. Ammonium is the preferred nitrogen source and leads to higher bacterial growth rates overall than other nitrogen sources (Merrick & Edwards, 1995). This is because ammonium can be directly absorbed into the cellular cycle.
All other inorganic compounds coming into the cell must first be reduced to ammonium at the expense of energy. Complex organic substances must be broken down by intracellular and/or extracellular enzymes. Various amino acids can be made available by deamination, which produces free ammonium, or transamination, i.e., the transfer of the amino group to, for example, 2-oxoglutarate (Voelker & Altaba, 2001). Depending on the available nutrient, bacteria can specifically synthesize or activate the necessary proteins for transport and degradation of the substances (Merrick & Edwards, 1995). Glutamine synthetases (GSs) play a key role in cellular nitrogen metabolism. Due to their high substrate specificity, these enzymes are capable of assimilating ammonium at concentrations below 0.1 mM (Magasanik, 1982).
Glutamine is formed from ammonium and glutamate using ATP. Glutamate synthase (GOGAT) then catalyzes the NADPH-dependent formation of glutamate from glutamine and 2-oxoglutarate. The GS/GOGAT pathway is ubiquitous in bacteria and is the only pathway for ammonium utilization in many organisms. Glutamine serves as both an amino acid and a nitrogen donor in the synthesis of approximately 25% of all nitrogen-containing cellular components (Reitzer & Schneider, 2001). Some bacteria have an alternative route for ammonium assimilation: the direct formation of glutamate through reductive amination of 2-oxoglutarate by glutamate dehydrogenase (GDH). However, due to its low substrate specificity, this enzyme only works effectively under high ammonium concentrations (Figure 1) (Merrick & Edwards, 1995).
1.1.2. Nitrogen Assimilation in Actinomycetes: ammonium catabolism as central catabolic route
The regulation of N assimilation in Actinomycetales differs from that in Enterobacteriaceae, but also has numerous similarities, particularly in the key enzymes. It can occur at the transcriptional and post-translational levels. Proteins such as GlnR, GlnRII, AmtR, NnaR, and Crp play an important role in transcriptional regulation. Post-translational regulation occurs through the signaling protein PII and by modulating the activity of important enzymes (GDH, GS) through adenylation/uridylation. GSI and GOGAT in Actinomycetales fulfill the same function in ammonium assimilation as GS and GOGAT in E. coli: In their overall reaction, ammonium and α-ketoglutarate are converted to glutamate with the consumption of ATP and NADPH (Figure 2) (Wray and Fisher, 1988, 1991, 1993; Fisher, 1989).
GSI is regulated by GlnE via adenylation and deadenylation (Streicher and Tyler, 1981; Fink et al., 1999; Nolden et al., 2001; Williams et al., 2013). Interestingly, there is evidence of possible transcriptional regulation by GlnE, which would make GlnE a so-called "moonlighting" protein (Rehm et al., 2010). Another form of glutamine synthetase, GSII (encoded by glnII), was first discovered in Rhizobium (Darrow and Knotts, 1977). GSII is a heat-labile octamer and shows sequence similarity to a eukaryotic GS. According to recent studies, GSII was introduced into the photosynthetic eukaryote Chloroplastida by horizontal gene transfer from gamma-proteobacteria at an early stage of plant evolution (Ghoshroy et al., 2010). GSII has been found in many members of the Actinomycetales (Edmands et al., 1987; Behrmann et al., 1990; Kumada et al., 1990), but it is absent in Amycolatopsis, Mycobacterium, and Corynebacterium (Reuther and Wohlleben, 2007). The difference from GSI is the lack of posttranslational regulation by adenylation (Hillemann et al., 1993). Instead, the GSII protein can degrade rapidly after N shock, suggesting control by proteolysis.
In addition to glutamine synthetases, bacteria possess GS-like proteins. GlnA2, GlnA3, and GlnA4 have been found in some Actinomycetes, e.g., in S. coelicolor (Bentley et al., 2002) and in M. tuberculosis (Harth et al., 2005). However, these proteins lack the amino acid residues conserved in GS, which form an adenylation motif. Therefore, a deficiency in the posttranslational regulation of GlnA2, GlnA3, and GlnA4 by adenylation was suspected. The transcription of glnA2, glnA3, and glnA4 is generally not regulated by GlnR or GlnRII in S. coelicolor, as is the case with glnA and glnII (Fink et al., 2002; Rexer et al., 2006). These proteins also lack the amino acid residues involved in the catalytic reaction of GS. Accordingly, GlnA2, GlnA3, and GlnA4 did not exhibit GS activity (Krysenko et al., 2017). The function of the GS-like protein AtdA1 in Acinetobacter sp. strain YAA lies in the ATP-dependent reaction of glutamate and aniline to ɤ-glutamylanilide, which serves for aniline degradation (Takeo et al., 2013).
On the other hand, transcription of glnA2 is upregulated in the glnR mutant of M. smegmatis (Jenkins et al., 2013; Jessberger et al., 2013). Not all bacteria function in parallel with GSI. For example, in B. subtilis, only the GS pathway is used (Deshpande et al., 1981). In contrast, in S. hygroscopicus 155, another enzyme, alanine dehydrogenase (ADH), is used instead of GDH for ammonium assimilation (Chipeva et al., 1991). ADH catalyzes the amination of pyruvate, consuming NAD+ and leading to alanine formation. Because ADH has a high Km value, it is particularly beneficial to bacteria at high N concentrations, such as 20–100 mM ammonium (Chipeva et al., 1991). ADH activity has also been detected in S. clavuligerus (Aharonowitz, 1980; Bascaran et al., 1986) and in R. leguminosarum (Allaway et al., 2000). In some Streptomyces, the activity of both enzymes, ADH and GDH, has been demonstrated (Shapiro and Vining, 1983).
In contrast to E. coli, which possesses only the NADPH-dependent GDH (Tyler, 1978), some Actinomycetes contain two GDHs. Glutamate dehydrogenase catalyzes the conversion of ammonium and α-ketoglutarate to glutamate (reductive amination) as well as the reverse reaction. In prokaryotes, ammonium assimilation is usually catalyzed by an anabolic, NADPH-dependent GDH enzyme (Hudson and Daniel, 1993). Glutamate cleavage (oxidative deamination) during ammonium dissimiliation is carried out by a catabolic, NADH-dependent glutamate dehydrogenase. The specificity of the enzymes toward their coenzymes NADP+ or NAD+ has been linked to conserved amino acid residues: an acidic amino acid at position 7 (P7) acts as an indicator of NADP+ specificity (Engel, 2014). The anabolic GDH is a homohexamer with approximately 50 kDa subunits. The catabolic prokaryotic GDH2 was discovered later than the anabolic GDH. A characteristic feature of GDH2 is its large subunit of 180 kDa (Kawakami et al., 2007).
1.1.2.1. Nitrogen Assimilation in Actinomycetales: catabolism of poor nitrogen sources
The assimilation of poor nitrogen sources requires the use of specialized enzymes involved in sophisticated systems for nitrate assimilation and respiration, for the assimilation of urea, amino acids, and other nitrogen-containing substances (Figure 3) (Hopwood et al., 1995, 1999). Nitrate can be either assimilated (extracted nitrogen is incorporated into cellular components in anabolic reactions) or dissimilated (in so-called nitrate respiration, nitrate is used as an electron acceptor instead of oxygen). Some Actinobacteria can operate both nitrate reduction pathways (Fischer et al., 2010, 2013). Nitrate uptake and utilization require special enzymes. First, nitrate molecules are transported into the cell by nitrite/nitrate transporters (NarK). There are two types of NarK transporters, NarK1 and NarK2. These are encountered as transport proteins in dissimilatory and assimilatory processes. The first is a nitrate/proton symporter, the second a nitrate/nitrite antiporter. Many proteobacteria use a mixed transporter in which NarK1 and NarK2 are fused together (Goddard et al., 2008, 2017).
After transporting into the cell, nitrate is reduced to nitrite. This is performed by a nitrate reductase. There are two classes of prokaryotic nitrate reductases: the Nar and Nas/Nap clades (Stolz and Basu, 2002). Cytosolic Nas reductase is involved in the assimilation of nitrate. In contrast, membrane-bound Nar and Nas reductases are involved in dissimilatory processes. The reducing equivalents for Nas can be derived from NAD(P)H, ferredoxin, or flavodoxin.
Transcription is induced by ammonium deficiency and nitrate presence (Stewart, 1994). Nas has the least conserved amino acid sequence among nitrate reductases, which is why it is considered a rapidly evolving protein (Stolz and Basu, 2002). In cyanobacteria, NarB functions as a cytoplasmic, assimilatory nitrate reductase. NarB, which belongs to the class of Nas reductases, has also been found in Actinomycetes. For example, NasA and NarB are present in some Streptomyces (Hsiao and Kirby, 2008; Fischer et al., 2014).
Nas, a related protein, is more flexible in its function. In most cases, it catalyzes the first step in the reduction of nitrate to ammonium. Furthermore, it can also play a role in denitrification reactions, the maintenance of redox balance, and nitrate sensing and uptake under severe N deficiency (Richardson et al., 2001). Nar is involved in the establishment of the proton gradient during nitrate respiration (Zumft, 1997).
Nitrite reductases convert nitrite produced in the reaction catalyzed by nitrate reductases into ammonium (assimilatory pathway) or into NO (dissimilatory pathway). Four types of nitrite reductases have been identified: Cu-containing NirK, found exclusively in denitrifying bacteria (Zumft, 1997; Higgins et al., 2016) and three other reductases with different heme groups (cytochrome cd1, siroheme, or multiheme) (Richter et al., 2002; Wang et al., 2000). NirS with the heme group cytochrome cd1 has been found only in denitrifying bacteria, while two nitrite reductases, NirB with siroheme and NrfA with multiheme, are present in E. coli (Wang et al., 2000; Besson et al., 2022)
In S. coelicolor, NnaR together with the global regulator GlnR, regulates the gene expression of narK, nasA, and nirB (Tiffert et al., 2011; Amin et al., 2012). The high sequence homology of NnaR in different Actinomycetales and the genetic location of nnaR, which was always found near narK, nasA, and nirB, suggest that NnaR plays a role in nitrate assimilation in all Actinomycetales.
In Actinomycetales, the genes for the ammonium transporter (encoded by amtB), the signaling protein PII (unlike in E. coli, PII is encoded by glnK instead of glnB), and the uridylyltransferase (encoded by glnD) are combined in an amtB operon (Wang et al., 2000; Jakoby et al., 2000; Fink et al., 2002). While the operon is inactive in E. coli under N excess (van Heeswijk et al., 1996), weakened expression of the amtB operon has been reported in Streptomyces under these conditions (Figure 3) (Tiffert et al., 2008).
In Streptomyces and Mycobacteria, GlnK is adenylated by GlnD under N deficiency and deadenylated again under N excess (Hesketh et al., 2002; Williams et al., 2013; Ensinck et al., 2024). In E. coli, GlnK is uridinylated and thus transmits the cellular N concentration to the ATase that regulates GSI activity (Atkinson and Ninfa, 1993, 1994, 1998, 2002; Jiang et al., 1998; Atkinson and Ninfa, 1999; Fink et al., 1999; Jiang and Ninfa, 2009; Radchenko et al., 2010, 2013). In Actinomycetales, GlnK is modified by the glutamine synthetase adenylyltransferase GlnE (ATase equivalent in Actinomycetales) (Fink et al., 1999; Forchhammer, 2007). However, signaling from GlnK to GlnE is absent in Actinomycetales (Williams et al., 2013), and the significance of GlnK adenylation is currently unknown. In addition, GlnK is Streptomyces by cleavage of the first three amino acids at the N-terminus after ammonium shock (Hesketh et al., 2002).
The functional versatility of GlnK in Actinomycetes is also evident in N metabolism systems such as the AmtR regulon of Corynebacterium (Jakoby et al., 2000) or the TnrA regulon of Bacillus (Fisher and Sonenshein, 1991). The AmtR repressor in Corynebacteria is not regulated by small effector molecules, as is common in the TetR family (Ramos et al., 2005). Instead, the repressor dissociates from the DNA after forming a complex with GlnK adenylated at Tyr51 (Jakoby et al., 2000; Nolden et al., 2001; Strösser et al., 2004; Beckers et al., 2005). Interestingly, AmtB plays a role in this process (Strösser et al., 2004). TnrA, the transcriptional regulator of N metabolism in Bacillus, is bound to a membrane-bound GlnK-AmtB complex in the absence of ATP (Heinrich et al., 2006).
1.1.2.2. Assimilation of Amines in Actinomycetes
Amines are organic compounds that contain carbon-nitrogen bonds, whereas one or more hydrogen atoms in ammonia are replaced by alkyl or aryl groups. The functional group −NH2 present in primary amines is called the amino group, which is present in biologically occurring amines, such as monoamines (e.g., ethanolamine) and polyamines (e.g., putrescine). Polyamines are cationic charged molecules with a hydrocarbon chain and multiple amino groups (Michael, 2018). They fulfill a lot of physiological functions, e.g., cell growth, maturation and proliferation, cell signaling, gene expression and others (Cohen, 1998; Igarashi & Kashiwagi, 2000; Kusano & Suzuki, 2015; Miller-Fleming et al., 2005). The monoamine ethanolamine is a short molecule which is both a primary amine and a primary alcohol and is a nitrogenous base in phospholipids and a building block of biomembranes (e.g., phosphoethanolamine) (Knaak et al., 1997). Excess of polyamines or ethanolamine is very toxic for bacterial cells and can lead to cell death (Miller-Fleming et al., 2015). Hence, polyamine and ethanolamine utilization represent bacterial survival strategies that allow survival of S. coelicolor in soil environment. Modified (e.g., glutamylated or acetylated) mono-/polyamines can be used in metabolism as sources of carbon and nitrogen.
In S. coelicolor putrescine, spermidine and diaminopropan biosynthesis have been described in the late-stationary phase in NMMP-medium, but cadaverine synthesis occurred only under iron limitation (Burell et al., 2012). Proteins with high similarity to the polyamine binding lipoprotein (PotD) and/or putrescine-binding periplasmic protein (PotF), to the amino acids/polyamine permease (PuuP) and/or putrescine importer (PlaP) were found in many phyla including Actinobacteria. SCO5667 is a predicted homolog of the putrescine-binding periplasmic protein (PotF) from E. coli with the sequence identity/similarity of 29/47%. However, transcriptional analysis of expression patterns of these genes revealed only weak expression of sco5667 and sco5671 in presence of spermidine (Krysenko et al., 2017, 2021).
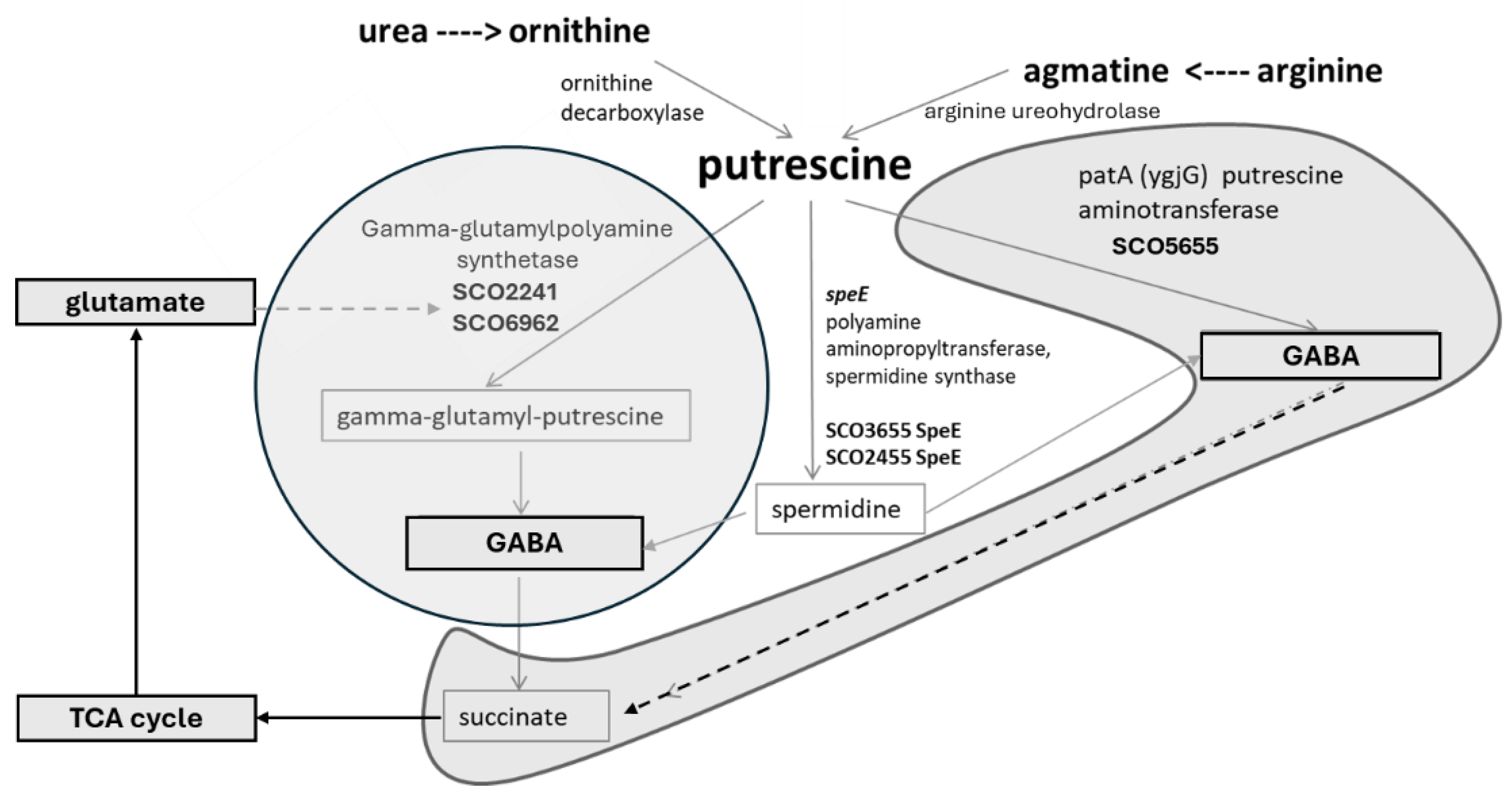
The most likely steps of the polyamine utilization pathway in S. coelicolor were reported to be similar to that known in E. coli (Kurihara et al., 2005, 2008, 2013) and P. aeruginosa (Kwon et al., 2006). The initial step of the polyamine utilization is catalyzed by gamma-glutamylpolyamine synthetases GlnA2 and/or GlnA3 resulting in glutamylated polyamines (Krysenko et al., 2017, 2022). These glutamylated products can likely be further reduced by the predicted gamma-glutamylpolyamine oxidoreductase (SCO5671). The reaction results in the production of the gamma-glutamyl-gamma-aminopentanal or gamma-glutamyl-gamma-aminobutyraldehyde. The SCO5671 enzyme is a close ortholog of the gamma-glutamylpolyamine oxidoreductases PuuB from E. coli and PauB1-B4 from P. aeruginosa. The gene sco5671 showed expression in presence of spermidine, but no expression in the presence of polyamines putrescine, cadaverine. In the next step of the utilization pathway, the predicted dehydrogenases (SCO5666 and SCO5657) might be involved. These proteins are predicted homologs of PuuC and PatD from E. coli, which are (gamma-glutamyl-) gamma-aminobutyraldehyde dehydrogenases. This step of the pathway may result in the production of the gamma-glutamyl-aminovalerate or gamma-glutamyl-GABA. The next step of polyamine utilization might require a predicted hydrolase (SCO6961) and result in the production of aminovalerate or GABA. Enhanced expression of sco5666, sco5657 and sco6961 in presence of polyamines was observed (Krysenko et al., 2017, 2021). These results suggest the role of SCO5666, SCO5657 and SCO6961 in polyamine utilization. Afterwards, SCO5676, which is a predicted homolog of the GABA aminotransferase GabT from E. coli, may be involved and catalyze the production of glutarate semialdehyde or succinate semialdehyde (Krysenko et al., 2021). It has been shown that the expression of the sco5676 gene is strong in presence of arginine (Perez-Redondo et al., 2012). Arginine is a precursor of putrescine in S. coelicolor M145. This finding as well as enhanced expression of sco5676 in presence of polyamines observed suggest the possible role of SCO5676 in the polyamine utilization. In the last step of the polyamine utilization pathway, SCO5679, which is a predicted homolog of the succinic semialdehyde dehydrogenase GabD from E. coli, may be involved. The polyamine utilization pathway ends with the succinate or glutarate that feeds the tricarboxylic acid (TCA) cycle (Figure 2 and Figure 4) (Krysenko et al., 2021).
A predicted amidotransferase (SCO5655) was identified, which is a homolog of the putrescine amidotransferase (PatA) from E. coli. In transcriptional analysis, the expression of sco5655, sco6960, sco6961 was enhanced in presence of polyamines. Moreover, sco5655 was reported to be induced by a diamide (Kallifidas et al., 2010) and not by arginine (Perez-Redondo et al., 2012). No orthologs of SCO6961 and SCO6960 were found in E. coli or P. aeruginosa. These findings suggest the possibility of an alternative polyamine utilization pathway in S. coelicolor (Figure 4) (Krysenko et al., 2017).
Metabolism of monoamine ethanolamine has been investigated in such Actinobacteria as Streptomyces and Mycobacterium (Krysenko & Wohlleben, 2022). In S. coelicolor, predicted ethanolamine permeases (SCO6014 and SCO5977) possess likely other functions, because the expression of the genes sco6014 and sco5977 was not induced by ethanolamine (Krysenko et al., 2019). Likely, an ethanolamine permease encoding gene was not required and lost in S. coelicolor during the evolution process and enough ethanolamine may enter the cells very probably through diffusion. It is also possible that S. coelicolor uses ethanolamine utilization pathway to control intracellular ethanolamine level during decomposition of membranes and to recycle the N-source.
In order to utilize ethanolamine as an N- and C-source and balance the intracellular ethanolamine pool, bacteria had to develop metabolic pathways for ethanolamine utilization. The canonical ethanolamine utilization pathway was studied in S. typhimurium and E. coli. However, alternative ethanolamine utilization pathways were reported in M. tuberculosis and C. salexigens. These pathways do not require a metabolosome and do not result in the production of toxic intermediates, such as acetaldehyde (Krysenko & Wohlleben, 2022).
It was shown in a recent study that diverse organisms ranging from the Actinobacteria to the Proteobacteria possess the capability for ethanolamine metabolism, which does not require eut genes (Brian et al., 2015; Gerlt et al., 2016). The most likely steps of the ethanolamine utilization pathway in S. coelicolor may be similar to the pathway described in studies of Brian et al., 2015 and Gerlt et al., 2016 as well as in detail in C. salexigens by Gerlt et al., 2016. After the glutamylation of ethanolamine by GlnA4 (SCO1613), a predicted gammaglutamylethanolamine dehydrogenase (SCO1611) may be required, producing gamma-glutamylacetaldehyde. In the next step of the pathway, a predicted gammaglutamylaldehyde dehydrogenase (SCO1612) may be involved, producing gammaglutamylglycine. The last step of the pathway may require a predicted gammaglutamylglycine amidohydrolase (SCO1615). The pathways may end in the production of glycine and glutamate. Further studies of SCO1611, SCO1612 and SCO1615 are required to determine their functionality in the ammaglutamylation pathway of ethanolamine in S. coelicolor (Krysenko et al., 2019, 2021).
1.1.3. Nitrogen Assimilation in Actinomycetales – Transcriptional Regulation
1.1.3.1. Regulation of Central Pathways
Nitrogen assimilation in Enterobacteriaceae is regulated by the Ntr system, however, most Actinomycetales possess the global transcriptional regulator GlnR for the same purpose (Amon et al., 2008; Tiffert et al., 2008). Homodimerization of this regulator is necessary for DNA binding and thus for regulation. In the Ntr system, phosphorylation of the conserved Asp residue by NtrB ensures NtrC dimerization. Since a GlnR with a conserved Asp residue in the N-terminal receiver domain was found in Actinomycetales, but no kinase phosphorylating this Asp residue, GlnR was long considered an "orphan" regulator. Then it was discovered that the conserved Asp residue is not phosphorylated and that the activating GlnR dimerization in A. mediterranei is caused by the ionic interaction of unphosphorylated Asp50 with the conserved Arg52 and Thr9 (Lin et al., 2014). Due to the lack of phosphorylation of Asp, GlnR is considered an atypical OmpR-like regulator. The receiver domain of one GlnR molecule forms an α4-β5-α5 surface, through which interaction with the receiver domain of the second GlnR molecule takes place (Lin et al., 2014). More than 15 genes are under the control of GlnR, most of which regulate nitrogen metabolism (Tiffert et al., 2008). GlnR exerts its activating effect on gene transcription under nitrogen deficiency by binding to the promoter regions of the regulated genes, thereby altering transcription (Tiffert et al., 2011). Thus, the amtB operon in S. coelicolor is controlled by GlnR activation (Fink et al., 2002), with a correlation between transcription and the amount of nitrogen found (Nolden et al., 2001; Fink et al., 2002). This again differs from the regulation in E. coli, where glnD expression has been described as constitutive (van Heeswijk et al., 1996). In addition to GlnR, Actinomycetes also contain GlnRII, which is less common in bacteria and has not been studied as extensively as GlnR. Like GlnR, GlnRII plays a role a regulatory role in the genes of N metabolism. However, so far, GlnRII has only been shown to affect the genes glnA, amtB, and glnII (Fink et al., 2002).
Another transcriptional regulator of N metabolism that is widespread among Actinomycetes is AmtR. In Corynebacteria, AmtR, a regulator of the TetR family that acts as a repressor of gene expression (Jakoby et al., 2000), replaces the GlnR regulator. In N excess, AmtR is bound to DNA, thus preventing the expression of key genes involved in N metabolism, such as glnA, gltB, and the amtB operon (Jakoby et al., 2000; Nolden et al., 2001; Beckers et al., 2005). In N deficiency, an adenylated GlnK gene product binds to AmtR and leads to the dissociation of the repressor from the DNA, allowing the released genes to be transcribed (Beckers et al., 2005). It was recently discovered that some members of the Corynebacterineae and Streptomyces, which possess a GlnR regulator, also possess an AmtR protein that shows low homology to the AmtR in Corynebacteria (Amon et al., 2008). Even though TetR regulators possess a highly conserved DNA-binding domain, AmtR regulators exhibit individual recognition features in the amino acid sequence (Muhl et al., 2009). Subsequent investigation of these AmtR regulators revealed that GlnR plays the main role in the regulation of nitrogen assimilation, while AmtR regulates only a subset of genes and its regulation is GlnR-dependent (Jessberger et al., 2013). This AmtR regulon includes genes for amidase, urea carboxylase, and amino acid permease (Jessberger et al., 2013).
1.1.3.2. Regulation of Catabolism of Amines in Actinomycetes
In S. coelicolor, the global regulator of the nitrogen metabolism GlnR can undergo post-translational modifications that affect its binding affinity to DNA (Amin et al., 2016). It was shown that the phosphorylation of Ser/Thr occurs under N-excess conditions. In agreement with these results, lack of phosphorylation on the Asp50 and lack of any Ser/Thr phosphorylation in GlnR isolated from S. coelicolor grown under N-limiting conditions was also demonstrated (Lin et al., 2014). Phopshorylation and acetylation were shown to influence the DNA-binding affinity of GlnR (Amin et al., 2016). GlnR regulates the glutamine synthetase encoding genes glnA and glnII at transcriptional level in dependence of N-conditions. However, no binding GlnR in promoter regions of glnA2, glnA3 and glnA4 were reported (Fink et al., 2002; Rexer et al., 2006). However, the acetylated version of GlnR binds better to the glnA2 promoter region (Krysenko et al., 2022). Interestingly, studies in the actynomycete Saccharopolyspora erythraea revealed that GlnR activates the expression of glnA3 (SACE_3095) (Yao et al., 2014).
In the genome of S. coelicolor, the sco5656 (epuRII) gene is localized close to the genes that encode predicted enzymes of polyamine utilization pathway (Krysenko et al., 2022) and is annotated as putative regulator (Bentley et al., 2002). The expression of the epuRII gene in presence of putrescine, cadaverine and spermidine was enhanced (Krysenko et al., 2017). EMSA analysis of regulatory targets of EpuRII revealed hints towards a complex regulation of several polyamine associated genes. These include glnA3, as well as sco5676 encoding a putative homolog of the 4-amino-butyrate aminotransferase GabT of E. coli K12 and sco5977 encoding a putative polyamine antiporter. The tests with EpuRII resulted in eight positive hits for EpuRII-interacting promoter sequences. The following genes seem to be regulated by EpuRII: glnA3, sco5676, coding for a putative homologue of the 4-amino-butyrate aminotransferase GabT, sco5977 encoding a putative polyamine antiporter, and sco6960 with unknown function (Krysenko et al., 2022).
There are two known regulatory mechanisms of ethanolamine utilization genes: the EutR system and the EutVW system. Regulation of ethanolamine utilization genes was well studied in S. typhimurium, E. coli and E. faecalis. In S. coelicolor, the sco1614 (epuRI) gene was annotated as putative regulator in the genome of S. coelicolor that is localized close to the gene glnA4 (sco1613) (Bentley et al., 2002). EMSA analysis revealed glnA4 gene as a potential target of EpuRI. Other genes located downstream of glnA4, namely sco1612, sco1611 and sco1610, encode predicted enzymes of the ethanolamine utilization pathway and are organized in one putative operon together with glnA4. Thus, these genes may be transcribed together with glnA4. The role of EpuRI as a negative transcriptional regulator of the ethanolamine utilization associated genes was shown in a transcriptional analysis (Krysenko et al., 2019). Interestingly, in preliminary EMSA analysis also interactions of EpuRI with promoter sequences of sco5652 and with the adjacent operon including sco5654 were observed. The sco5652 gene encodes a protein of unknown function, sco5654 encodes a putative ABC transporter. EpuRI demonstrated interactions with the promoter region of sco5657 (encoding a putative aldehyde dehydrogenase), sco1616 (a putative regulator) and the promoter sequence of epuRI itself. Regulatory network that involves EpuRI seems to be complex (Krysenko et al., 2019, 2022).
3. Conclusions and Future Perspectives
Nitrogen metabolism is essential for bacterial survival under nutrient limitation conditions in competitive ecological niches. Bacteria have developed complex metabolic networks and regulatory machinery to control intracellular pools of nitrogen. Nitrogen metabolism comprising transport, biosynthesis, utilization and regulation is a central part of bacterial primary metabolism, ensuring supply of building blocks for biomolecules and biomass generation. Catabolism of nitrogen-containing compounds, like ammonium, nitrate, amino acids, amino sugars, urea and amines ensures bacterial survival and pathogenicity. In this regard, metabolism of amines has been investigated in several human pathogenic bacteria in connection to infection process. Studies in pathogenic bacterial species including Salmonella typhimurium, Brucella abortus, Mycobacterium tuberculosis, Chlamydia pneumoniae, Legionella pneumophila, Listeria monocytogenes, and some others prove the crucial role of polyamines for their proliferation. The interconnection between the biosynthesis, uptake, and assimilation of amines remains crucial to find new therapeutic drug targets (Krysenko & Makhoba, 2025). Most human pathogens rely not only on polyamine biosynthesis, but also on polyamine detoxification to proliferate and maintain infection, targeting metabolism of amines can extend the options for combating bacterial infections (Krysenko et al., 2023, 2025). There is an urgent need to find new anti-bacterial drugs with novel modes of action that would be efficient on bacterial infections, especially in the light of emergence of resistances. Investigation of the pathways in nitrogen metabolism required for bacterial growth, survival, and pathogenicity provide new drug target candidates for development of more effective agents.
Author Contributions
Conceptualization, writing—original draft preparation, visualization, S.K.
Funding
This research received no external funding.
Acknowledgement
I thank Mariko Matsuura for proof-reading of the final version of the manuscript.
Conflicts of Interest
Sergii Krysenko is employed by the Valent BioSciences. The author declares no conflict of interest, the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Aharonowitz, Y. Nitrogen metabolite regulation of antibiotic biosynthesis. Annu Rev Microbiol 1980, 34, 209–233. [Google Scholar] [CrossRef] [PubMed]
- Allaway, D.; Lodwig, E.; Crompton, L.A.; Wood, M.; Parsons, R.; Wheeler, T.R.; Poole, P.S. Identification of alaninedehydrogenase and its role in mixed secretion of ammonium andalanine by pea bacteroids. Mol Microbiol 2000, 36, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Amin, R.; Reuther, J.; Bera, A.; Wohlleben, W.; Mast, Y. A novel GlnR target gene, nnaR, is involved in nitrate/nitrite assimilation in Streptomyces coelicolor. Microbiology 2012, 158, 1172–1182. [Google Scholar] [CrossRef]
- Amin, R.; Franz-Wachtel, M.; Tiffert, Y.; Heberer, M.; Meky, M.; Ahmed, Y.; Matthews, A.; Krysenko, S.; Jakobi, M.; Hinder, M.; Moore, J.; Okoniewski, N.; Maček, B.; Wohlleben, W.; Bera, A. Post-translational Serine/Threonine Phosphorylation and Lysine Acetylation: A Novel Regulatory Aspect of the Global Nitrogen Response Regulator GlnR in S. coelicolor M145. Front. Mol. Biosci. 2016, 3, 38. [Google Scholar] [CrossRef]
- Amon, J.; Bräu, T.; Grimrath, A.; Hänßler, E.; Hasselt, K.; Höller, M.; Jeßberger, N.; Ott, L.; Szököl, J.; Titgemeyer, F.; Burkovski, A. Nitrogen control in Mycobacterium smegmatis: Nitrogen-dependent expression of ammonium transport and assimilation proteins depends on OmpR-type regulator GlnR. J Bacteriol 2008, 190, 7108–7116. [Google Scholar] [CrossRef]
- Atkinson, M.R.; Ninfa, A.J. Mutational analysis of the bacterial signal-transducing protein kinase/phosphatase nitrogen regulator II (NRII or NtrB). J Bacteriol 1993, 175, 7016–7023. [Google Scholar] [CrossRef]
- Atkinson, M.R.; Ninfa, A.J. Role of the GlnK signal transduction protein in the regulation of nitrogen assimilation in Escherichia coli. Mol Microbiol 1998, 29, 431–447. [Google Scholar] [CrossRef]
- Atkinson, M.R.; Ninfa, A.J. Characterization of the GlnK protein of Escherichia coli. Mol Microbiol 1999, 32, 301–313. [Google Scholar] [CrossRef]
- Atkinson, M.R.; Blauwkamp, T.A.; Bondarenko, V.; Studitsky, V.; Ninfa, A.J. Activation of the glnA, glnK, and nac promoters as Escherichia coli undergoes the transition from nitrogen excess growth to nitrogen starvation. J Bacteriol 2002, 184, 5358–5363. [Google Scholar] [CrossRef]
- Atkinson, M.R.; Kamberov, E.S.; Weiss, R.L.; Ninfa, A.J. Reversible uridylylation of the Escherichia coli PII signal transduction protein regulates its ability to stimulate the dephosphorylation of the transcription factor nitrogen regulator I (NRI or NtrC). J Biol Chem 1994, 269, 28288–28293. [Google Scholar] [CrossRef]
- Bascaran, V.; Hardisson, C.; Brana, A.F. Regulation of nitrogen catabolic enzymes in Streptomyces clavuligerus. J Gen Microbiol 1989, 135, 2465–2474. [Google Scholar]
- Beckers, G.; Strösser, J.; Hildebrandt, U.; Kalinowski, J.; Farwick, M.; Krämer, R.; Burkovski, A. Regulation of AmtR-controlled gene expression in Corynebacterium glutamicum: Mechanism and characterization of the AmtR regulon. Mol. Microbiol. 2005, 58, 580–595. [Google Scholar] [CrossRef]
- Behrmann, I.; Hillemann, D.; Pühler, A.; Strauch, E.; Wohlleben, W. Overexpression of a Streptomyces viridochromogenes gene (glnII) encoding a glutamine synthetase similar to those of eucaryotes confers resistance against the antibiotic phosphinothricyl-alanyl-alanine. J. Bacteriol. 1990, 172, 5326–5334. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; A., *!!! REPLACE !!!*; et al. Complete Genome Sequence of the Model Actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Besson, S.; Almeida, M.G.; Silveira, C.M. Nitrite reduction in bacteria: A comprehensive view of nitrite reductases. Coordination Chemistry Reviews 2022, 464, 214560. [Google Scholar] [CrossRef]
- Burrell, M.; Hanfrey, C.C.; Kinch, L.N.; Elliot, K.A.; Michael, J.A. Evolution of a novel lysine decarboxylase in siderophore biosynthesis. Mol. Microbiol. 2012, 86, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Chipeva, V.; Dumanova, E.; Todorov, T.; et al. Impact of nitrogen assimilation on regulation of antibiotic production in Streptomyces hygroscopicus 155. Antibiot Khimioter 1991, 36, 5–8. [Google Scholar]
- Cohen, S.S. (1998) A Guide to Polyamines, Oxford University Press, New York.
- Darrow, R.A.; Knotts, R.R. Two forms of glutamine synthetase in free-living root-nodule bacteria. Biochem Biophys Res Commun 1977, 78, 554–559. [Google Scholar] [CrossRef]
- Deshpande, K.L.; Katze, J.R.; Kane, J.F. Effect of glutamine on enzymes of nitrogen metabolism in Bacillus subtilis. Journal of bacteriology 1981, 145, 768–774. [Google Scholar] [CrossRef]
- Edmands, J.; Noridge, N.A.; Benson, D.R. The actinorhizal root-nodule symbiont Frankia sp. strain CpI1 has two glutamine synthetases. Proc Natl Acad Sci USA 1987, 84, 6126–6130. [Google Scholar] [CrossRef]
- Engel, P.C. Glutamate dehydrogenases: The why and how of coenzyme specificity. Neurochemical research 2014, 39, 426–432. [Google Scholar] [CrossRef]
- Ensinck, D.; Gerhardt, E.C.M.; Rollan, L.; Huergo, L.F.; Gramajo, H.; Diacovich, L. The PII protein interacts with the Amt ammonium transport and modulates nitrate/nitrite assimilation in mycobacteria. Frontiers in microbiology 2024, 15, 1366111. [Google Scholar] [CrossRef]
- Fink, D.; Falke, D.; Wohlleben, W.; Engels, A. Nitrogen metabolism in Streptomyces coelicolor A3(2): Modification of glutamine synthetase I by an adenylyltransferase. Microbiology 1999, 145 Pt 9, 2313–2322. [Google Scholar] [CrossRef]
- Fink, D.; Weißschuh, N.; Reuther, J.; Wohlleben, W.; Engels, A. Two transcriptional regulators GlnR and GlnRII are involved in regulation of nitrogen metabolism in Streptomyces coelicolor A3(2). Mol Microbiol 2002, 46, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.H. Glutamate synthesis in Streptomyces coelicolor. J Bacteriol 1989, 171, 2372–2377. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.H.; Sonenshein, A.L. Control of carbon and nitrogen metabolism in Bacillus subtilis. Annu Rev Microbiol 1991, 45, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Alderson, J.; van Keulen, G.; White, J.; Sawers, R.G. The obligate aerobe Streptomyces coelicolor A3(2) synthesizes three active respiratory nitrate reductases. Microbiology (Reading, England) 2010, 156 Pt 10, 3166–3179. [Google Scholar] [CrossRef]
- Fischer, M.; Falke, D.; Sawers, R.G. A respiratory nitrate reductase active exclusively in resting spores of the obligate aerobe Streptomyces coelicolor A3(2). Molecular microbiology 2013, 89, 1259–1273. [Google Scholar] [CrossRef]
- Fischer, M.; Falke, D.; Pawlik, T.; Sawers, R.G. Oxygen-dependent control of respiratory nitrate reduction in mycelium of Streptomyces coelicolor A3(2). Journal of bacteriology 2014, 196, 4152–4162. [Google Scholar] [CrossRef]
- Forchhammer, K. Glutamine signalling in bacteria. Front Biosci 2007, 12, 358–370. [Google Scholar] [CrossRef]
- Fuchs, G. (Hrsg.), (2022) Allgemeine Mikrobiologie, 880 S, 750 Abb., Thieme, 11. vollst. überarb. Aufl., HC, ISBN: 9783132434776.
- Gerlt, J.A. Tools and strategies for discovering novel enzymes and metabolic pathways. Perspect. Sci. 2016, 9, 24–32. [Google Scholar] [CrossRef]
- Ghoshroy, S.; Binder, M.; Tartar, A.; Robertson, D.L. Molecular evolution of glutamine synthetase II: Phylogenetic evidence of a non-endosymbiotic gene transfer event early in plant evolution. BMC Evol Biol 2010, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Goddard, A.D.; Moir, J.W.; Richardson, D.J.; Ferguson, S.J. Interdependence of two NarK domains in a fused nitrate/nitrite transporter. Molecular microbiology 2008, 70, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Goddard, A.D.; Bali, S.; Mavridou, D.A.; Luque-Almagro, V.M.; Gates, A.J.; Dolores Roldán, M.; Newstead, S.; Richardson, D.J.; Ferguson, S.J. The Paracoccus denitrificans NarK-like nitrate and nitrite transporters-probing nitrate uptake and nitrate/nitrite exchange mechanisms. Molecular microbiology 2017, 103, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Harth, G.; Maslesa-Galic, S.; Tullius, M.V.; Horwitz, M.A. All four Mycobacterium tuberculosis glnA genes encode glutamine synthetase activities but only GlnA1 is abundantly expressed and essential for bacterial homeostasis. Mol Microbiol 2005, 58, 1157–1172. [Google Scholar] [CrossRef]
- Heinrich, A.; Woyda, K.; Brauburger, K.; Meiss, G.; Detsch, C.; Stülke, J.; Forchhammer, K. Interaction of the membrane-bound GlnK-AmtB complex with the master regulator of nitrogen metabolism TnrA in Bacillus subtilis. The Journal of biological chemistry 2006, 281, 34909–34917. [Google Scholar] [CrossRef]
- Hesketh, A.; Fink, D.; Gust, B.; Rexer, H.U.; Scheel, B.; Chater, K.; et al. The GlnD and GlnK homologues of Streptomyces coelicolor A3(2) are functionally dissimilar to their nitrogen regulatory system counterparts from enteric bacteria. Mol. Microbiol. 2002, 46, 319–330. [Google Scholar] [CrossRef]
- Higgins, S.A.; Welsh, A.; Orellana, L.H.; Konstantinidis, K.T.; Chee-Sanford, J.C.; Sanford, R.A.; Schadt, C.W.; Löffler, F.E. Detection and Diversity of Fungal Nitric Oxide Reductase Genes (p450nor) in Agricultural Soils. Applied and environmental microbiology 2016, 82, 2919–2928. [Google Scholar] [CrossRef]
- Hillemann, D.; Dammann, T.; Hillemann, A.; Wohlleben, W. Genetic and biochemical characterization of the two glutamine synthetases GSI and GSII of the phosphinothricyl-alanyl-alanine producer, Streptomyces viridochromogenes Tü494. Journal of general microbiology 1993, 139, 1773–1783. [Google Scholar] [CrossRef]
- Hojati, Z.; Milne, C.; Harvey, B.; Gordon, L.; Borg, M.; et al. Structure, biosynthetic origin, and engineered biosynthesis of calcium-dependent antibiotics from Streptomyces coelicolor. Chem Biol 2002, 9, 1175–1187. [Google Scholar] [CrossRef]
- Hopwood, D.A. The Leeuwenhoek Lecture, 1987: Towards an understanding of gene switching in Streptomyces, the basis of sporulation and antibiotic production. Proc R Soc Lond B Biol Sci 1988, 235, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A.; Chater, K.F.; Bibb, M.J. Genetics of antibiotic production in Streptomyces coelicolor A3(2), a model streptomycete. Biotechnology 1995, 28, 65–102. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A. Forty years of genetics with Streptomyces: From in vivo through in vitro to in silico. Microbiology 1999, 145, 2183–2202. [Google Scholar] [CrossRef]
- Hsiao, N.H.; Kirby, R. Comparative genomics of Streptomyces avermitilis, Streptomyces cattleya, Streptomyces maritimus and Kitasatospora aureofaciens using a Streptomyces coelicolor microarray system. Antonie van Leeuwenhoek 2008, 93, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.C.; Daniel, R.M. L-glutamate dehydrogenases: Distribution, properties and mechanism. Comparative biochemistry and physiology. B, Comparative biochemistry 1993, 106, 767–792. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. Polyamines: Mysterious modulators of cellular functions. Biochem. Biophys. Res. Commun. 2000, 271, 559–564. [Google Scholar] [CrossRef]
- Jakoby, M.; Nolden, L.; Meier-Wagner, J.; Krämer, R.; Burkovski, A. AmtR, a global repressor in the nitrogen regulation system of Corynebacterium glutamicum. Molecular microbiology 2000, 37, 964–977. [Google Scholar] [CrossRef]
- Jacoby, R.P.; Succurro, A.; Kopriva, S. Nitrogen Substrate Utilization in Three Rhizosphere Bacterial Strains Investigated Using Proteomics. Front. Microbiol. 2020, 11, 784. [Google Scholar] [CrossRef]
- Jenkins, V.A.; Barton, G.R.; Robertson, B.D.; Williams, K.J. Genome wide analysis of the complete GlnR nitrogen-response regulon in Mycobacterium smegmatis. BMC Genomics 2013, 14, 301. [Google Scholar] [CrossRef]
- Jeßberger, N.; Lu, Y.; Amon, J.; Titgemeyer, F.; Sonnewald, S.; Reid, S.; Burkovski, A. Nitrogen starvation-induced transcriptome alterations and influence of transcription regulator mutants in Mycobacterium smegmatis. BMC research notes 2013, 6, 482. [Google Scholar] [CrossRef]
- Knaak, J.B.; Leung, H.-W.; Stott, W.T.; Busch, J.; Bilsky, J. Toxicology of mono-, di-, and triethanolamine. Rev. Environ. Contam. Toxicol. 1997, 149, 1–86. [Google Scholar] [CrossRef] [PubMed]
- Krysenko, S.; Okoniewski, N.; Kulik, A.; Matthews, A.; Grimpo, J.; Wohlleben, W.; Bera, A. Gamma-Glutamylpolyamine Synthetase GlnA3 is involved in the first step of polyamine degradation pathway in Streptomyces coelicolor M145. Front. Microbiol. 2017, 8, 726. [Google Scholar] [CrossRef] [PubMed]
- Krysenko, S.; Matthews, A.; Busche, T.; Bera, A.; Wohlleben, W. Poly- and Monoamine Metabolism in Streptomyces coelicolor: The New Role of Glutamine Synthetase-Like Enzymes in the Survival under Environmental Stress. Microb. Physiol. 2021, 31, 233–247. [Google Scholar] [CrossRef]
- Krysenko, S.; Matthews, A.; Okoniewski, N.; Kulik, A.; Girbas, M.G.; Tsypik, O.; Meyners, C.S.; Hausch, F.; Wohlleben, W.; Bera, A. Initial metabolic step of a novel ethanolamine utilization pathway and its regulation in Streptomyces coelicolor M145. mBio. 2019, 10, e00326–19. [Google Scholar] [CrossRef]
- Krysenko, S.; Okoniewski, N.; Nentwich, M.; Matthews, A.; Bäuerle, M.; Zinser, A.; Busche, T.; Kulik, A.; Gursch, S.; Kemeny, A.; et al. A Second Gamma-Glutamylpolyamine Synthetase, GlnA2, Is Involved in Polyamine Catabolism in Streptomyces coelicolor. Int. J. Mol. Sci. 2022, 23, 3752. [Google Scholar] [CrossRef]
- Krysenko, S.; Lopez, M.; Meyners, C.; Purder, P.L.; Zinser, A.; Hausch, F.; Wohlleben, W. A novel synthetic inhibitor of polyamine utilization in Streptomyces coelicolor. FEMS microbiology letters 2023, 370, fnad096. [Google Scholar] [CrossRef]
- Krysenko, S.; Emani, C.S.; Bäuerle, M.; Oswald, M.; Kulik, A.; Meyners, C.; Hillemann, D.; Merker, M.; Prosser, G.; Wohlers, I.; Hausch, F.; Brötz-Oesterhelt, H.; Mitulski, A.; Reiling, N.; Wohlleben, W. GlnA3Mt is able to glutamylate spermine but it is not essential for the detoxification of spermine in Mycobacterium tuberculosis. Journal of bacteriology 2025, 207, e0043924. [Google Scholar] [CrossRef]
- Krysenko, S.; Wohlleben, W. Role of Carbon, Nitrogen, Phosphate and Sulfur Metabolism in Secondary Metabolism Precursor Supply in Streptomyces spp. Microorganisms 2024, 12, 1571. Microorganisms 2024, 12, 1571. [Google Scholar] [CrossRef]
- Kumada, Y.; Benson, D.R.; Hillemann, D.; Hosted, T.J.; Rochefort, D.A.; Thompson, C.J.; Wohlleben, W.; Tateno, Y. Evolution of the glutamine synthetase gene, one of the oldest existing and functioning genes. Proc. Natl. Acad. Sci. USA 1993, 90, 3009–3013. [Google Scholar] [CrossRef]
- Kurihara, S.; Oda, S.; Tsuboi, Y.; Kim, H.G.; Oshida, M.; Kumagai, H.; Suzuki, H. γ-Glutamylputrescine synthetase in the putrescine utilization pathway of Escherichia coli K-12. J Biol Chem 2008, 283, 19981–19990. [Google Scholar] [CrossRef]
- Kurihara, S.; Sakai, Y.; Suzuki, H.; Muth, A.; Ot, P.; Rather, P.N. Putrescine importer PlaP contributes to swarming motility and urothelial cell invasion in Proteus mirabilis. J Biol Chem 2013, 288, 15668–15676. [Google Scholar] [CrossRef]
- Kurihara, S.; Oda, S.; Kato, K.; Kim, H.G.; Koyanagi, T.; Kumagai, H.; Suzuki, H. A novel putrescine utilization pathway involves γ-glutamylated intermediates of Escherichia coli K-12. J Biol Chem 2005, 280, 4602–4608. [Google Scholar] [CrossRef] [PubMed]
- Kusano, T.; Suzuki, H. (eds). (2015) Polyamines: A Universal Molecular Nexus for Growth, Survival, and Specialized Metabolism. Tokyo: Springer.
- Kawakami, R.; Sakuraba, H.; Ohshima, T. Gene cloning and characterization of the very large NAD-dependent l-glutamate dehydrogenase from the psychrophile Janthinobacterium lividum, isolated from cold soil. Journal of bacteriology 2007, 189, 5626–5633. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Lu, C.D. Polyamines induce resistance to cationic peptide, aminoglycoside, and quinolone antibiotics in Pseudomonas aeruginosa PAO1. Antimicrob. Agents Chemother 2006, 50, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Wang, Y.; Han, X.; Zhang, Z.; Wang, C.; Wang, J.; et al. Atypical OmpR/PhoB subfamily response regulator GlnR of actinomycetes functions as a homodimer, stabilized by the unphosphorylated conserved Asp-focused charge interactions. J. Biol. Chem. 2014, 289, 15413–15425. [Google Scholar] [CrossRef]
- Magasanik, B. Genetic control of nitrogen assimilation in bacteria. Annual review of genetics 1982, 16, 135–168. [Google Scholar] [CrossRef]
- Makhoba, X.H.; Krysenko, S. Drug Target Validation in Polyamine Metabolism and Drug Discovery Advancements to Combat Tuberculosis. Future Pharmacology 2025, 5, 32. [Google Scholar] [CrossRef]
- Merrick, M.J.; Edwards, R.A. Nitrogen control in bacteria. Microbiological reviews 1995, 59, 604–622. [Google Scholar] [CrossRef]
- Michael, A.J. Polyamine function in archaea and bacteria. J. Biol. Chem. 2018, 293, 18693–18701. [Google Scholar] [CrossRef]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef]
- Muhl, D.; Jessberger, N.; Hasselt, K.; Jardin, C.; Sticht, H.; Burkovski, A. DNA binding by Corynebacterium glutamicum TetR-type transcription regulator AmtR. BMC molecular biology 2009, 10, 73. [Google Scholar] [CrossRef]
- Neidharth, F.C.; Reitzer, L.J. (1996). Regulation of nitrogen utilization Escherichia coli and Salmonella: Cellular and Molecular Biology (pp. 1344–1356). Washington, DC: ASM Press.
- Ninfa, A.J.; Jiang, P.; Atkinson, M.R. Integration of antagonistic signals in the regulation of nitrogen assimilation in Escherichia coli. Curr Top Cell Regul 2001, 36, 31–75. [Google Scholar] [CrossRef]
- Nolden, L.; Farwick, M.; Kramer, R.; Burkovski, A. Glutamine synthetases of Corynebacterium glutamicum: Transcriptional control and regulation of activity. FEMS Microbiol Lett 2001, 201, 91–98. [Google Scholar] [CrossRef]
- Perez-Redondo, R.; Rodriguez-Garcia, A.; Botas, A.; Santamarta, I.; Martin, J.F.; Liras, P. ArgR of Streptomyces coelicolor is a versatile regulator. PLoS ONE. 2012, 7, e32697. [Google Scholar] [CrossRef]
- Radchenko, M.V.; Thornton, J.; Merrick, M. Control of AmtB-GlnK complex formation by intracellular levels of ATP, ADP, and 2-oxoglutarate. J Biol Chem 2010, 285, 31037–31045. [Google Scholar] [CrossRef] [PubMed]
- Radchenko, M.V.; Thornton, J.; Merrick, M. P(II) signal transduction proteins are ATPases whose activity is regulated by 2-oxoglutarate. Proc Natl Acad Sci USA 2013, 110, 12948–12953. [Google Scholar] [CrossRef] [PubMed]
- Rehm, N.; Buchinger, S.; Strosser, J.; Dotzauer, A.; Walter, B.; Hans, S.; Bathe, B.; Schomburg, D.; Kramer, R.; Burkovski, A. Impact of adenylyltransferase GlnE on nitrogen starvation response in Corynebacterium glutamicum. J Biotechnol 2010, 145, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Reitzer, L. Nitrogen assimilation and global regulation in Escherichia coli. Annu Rev Microbiol 2003, 57, 155–176. [Google Scholar] [CrossRef]
- Reitzer, L.; Schneider, B.L. Metabolic context and possible physiological themes of σ54-dependent genes in Escherichia coli. Microbiol Mol Biol Rev 2001, 65, 422–444. [Google Scholar] [CrossRef]
- Reuther, J.; Wohlleben, W. Nitrogen metabolism in Streptomyces coelicolor: Transcriptional and post-translational regulation. J Mol Microbiol Biotechnol 2007, 12, 139–146. [Google Scholar] [CrossRef]
- Rexer, H.U.; Schäberle, T.; Wohlleben, W.; Engels, A. ; Investigation of the functional properties and regulation of three glutamine synthetase-like genes in Streptomyces coelicolor A3(2). Arch Microbiol 2006, 186, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.J.; Berks, B.C.; Russell, D.A.; Spiro, S.; Taylor, C.J. Functional, biochemical and genetic diversity of prokaryotic nitrate reductases. Cellular and molecular life sciences: CMLS 2001, 58, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.D.; Allen, J.W.; Higham, C.W.; Koppenhofer, A.; Zajicek, R.S.; Watmough, N.J.; Ferguson, S.J. Cytochrome cd1, reductive activation and kinetic analysis of a multifunctional respiratory enzyme. The Journal of biological chemistry 2002, 277, 3093–3100. [Google Scholar] [CrossRef] [PubMed]
- San Francisco, B.; Zhang, X.; Whalen, K.; Gerlt, K. A novel pathway for bacterial ethanolamine metabolism. FASEB J. 2015, 29, 573.45. [Google Scholar] [CrossRef]
- Shapiro, S.; Vining, L.C. Nitrogen metabolism and chloramphenicol production in Streptomyces venezuelae. Canadian journal of microbiology 1983, 29, 1706–1714. [Google Scholar] [CrossRef]
- Stewart, V. Dual interacting two-component regulatory systems mediate nitrate- and nitrite-regulated gene expression in Escherichia coli. Res Microbiol 1994, 145, 450–454. [Google Scholar] [CrossRef]
- Stolz, J.F.; Basu, P. Evolution of nitrate reductase: Molecular and structural variations on a common function. Chembiochem: A European journal of chemical biology 2002, 3, 198–206. [Google Scholar] [CrossRef]
- Streicher, S.L.; Tyler, B. Regulation of glutamine synthetase activity by adenylylation in the Gram-positive bacterium Streptomyces cattleya. Proc Natl Acad Sci USA 1981, 78, 229–233. [Google Scholar] [CrossRef]
- Strösser, J.; Ludke, A.; Schaffer, S.; Kramer, R.; Burkovski, A. Regulation of GlnK activity: Modification, membrane sequestration and proteolysis as regulatory principles in the network of nitrogen control in Corynebacterium glutamicum. Mol Microbiol 2004, 54, 132–147. [Google Scholar] [CrossRef]
- Takeo, M.; Ohara, A.; Sakae, S.; Okamoto, Y.; Kitamura, C.; Kato, D.; Negoro, S. Function of a glutamine synthetase-like protein in bacterial aniline oxidation via γ-glutamylanilide. Journal of bacteriology 2013, 195, 4406–4414. [Google Scholar] [CrossRef]
- Tiffert, Y.; Franz-Wachtel, M.; Fladerer, C.; Nordheim, A.; Reuther, J.; Wohlleben, W.; Mast, Y. Proteomic analysis of the GlnR-mediated response to nitrogen limitation in Streptomyces coelicolor M145. Appl Microbiol Biotechnol 2011, 89, 1149–1159. [Google Scholar] [CrossRef]
- Tiffert, Y.; Supra, P.; Wurm, R.; Wohlleben, W.; Wagner, R.; Reuther, J. The Streptomyces coelicolor GlnR regulon: Identification of new GlnR targets and evidence for a central role of GlnR in nitrogen metabolism in actinomycetes. Mol Microbiol 2008, 67, 861–880. [Google Scholar] [CrossRef]
- Tyler, B. Regulation of the assimilation of nitrogen compounds. Annu Rev Biochem 1978, 47, 1127–1162. [Google Scholar] [CrossRef] [PubMed]
- van Heeswijk, W.C.; Hoving, S.; Molenaar, D.; Stegeman, B.; Kahn, D.; Westerhoff, H.V. An alternative PII protein in the regulation of glutamine synthetase in Escherichia coli. Mol Microbiol 1996, 21, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Voelker, F.; Altaba, S. Nitrogen source governs the patterns of growth and pristinamycin production in 'Streptomyces pristinaespiralis'. Microbiology (Reading, England) 2001, 147 Pt 9, 2447–2459. [Google Scholar] [CrossRef]
- Wang, H.; Gunsalus, R.P. The nrfA and nirB nitrite reductase operons in Escherichia coli are expressed differently in response to nitrate than to nitrite. Journal of bacteriology 2000, 182, 5813–5822. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.J.; Bennett, M.H.; Barton, G.R.; Jenkins, V.A.; Robertson, B.D. Adenylylation of mycobacterial Glnk (PII) protein is induced by nitrogen limitation. Tuberculosis (Edinb) 2013, 93, 198–206. [Google Scholar] [CrossRef]
- Wray, L.V., Jr.; Fisher, S.H. Cloning and nucleotide sequence of the Streptomyces coelicolor gene encoding glutamine synthetase. Gene 1988, 71, 247–256. [Google Scholar] [CrossRef]
- Wray, L.V., Jr.; Fisher, S.H. The Streptomyces coelicolor glnR gene encodes a protein similar to other bacterial response regulators. Gene 1993, 130, 145–150. [Google Scholar] [CrossRef]
- Wray, L.V., Jr.; Atkinson, M.R.; Fisher, S.H. Identification and cloning of the glnR locus, which is required for transcription of the glnA gene in Streptomyces coelicolor A3(2). J Bacteriol 1991, 173, 7351–7360. [Google Scholar] [CrossRef]
- Yao, L.L.; Liao, C.H.; Huang, G.; Zhou, Y.; Rigali, S.; Zhang, B.; Ye, B.C. GlnR-mediated regulation of nitrogen metabolism in the actinomycete Saccharopolyspora erythraea. Applied microbiology and biotechnology 2014, 98, 7935–7948. [Google Scholar] [CrossRef]
- Zalkin, H.; Smith, J.L. Enzymes utilizing glutamine as an amide donor. Adv Enzymol Relat Areas Mol Biol 1998, 72, 87–144. [Google Scholar] [CrossRef]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiology and molecular biology reviews: MMBR 1997, 61, 533–616. [Google Scholar] [CrossRef]
Figure 1.
Central reactions of ammonium assimilation. At low concentrations, ammonium can be introduced into metabolism via the GS/GOGAT pathway. In the net reaction, glutamate is formed from 2-oxoglutarate and ammonium, consuming ATP and NADPH. At high concentrations, GDH catalyzes ammonium assimilation. All nitrogen-containing cellular components and metabolites, such as purines, pyrimidines, amino sugars, amino acids, and proteins, are produced from glutamine and glutamate (Zalkin and Smith, 1998). GS: glutamine synthetase, GOGAT: glutamate synthase, GDH: glutamate dehydrogenase.
Figure 1.
Central reactions of ammonium assimilation. At low concentrations, ammonium can be introduced into metabolism via the GS/GOGAT pathway. In the net reaction, glutamate is formed from 2-oxoglutarate and ammonium, consuming ATP and NADPH. At high concentrations, GDH catalyzes ammonium assimilation. All nitrogen-containing cellular components and metabolites, such as purines, pyrimidines, amino sugars, amino acids, and proteins, are produced from glutamine and glutamate (Zalkin and Smith, 1998). GS: glutamine synthetase, GOGAT: glutamate synthase, GDH: glutamate dehydrogenase.
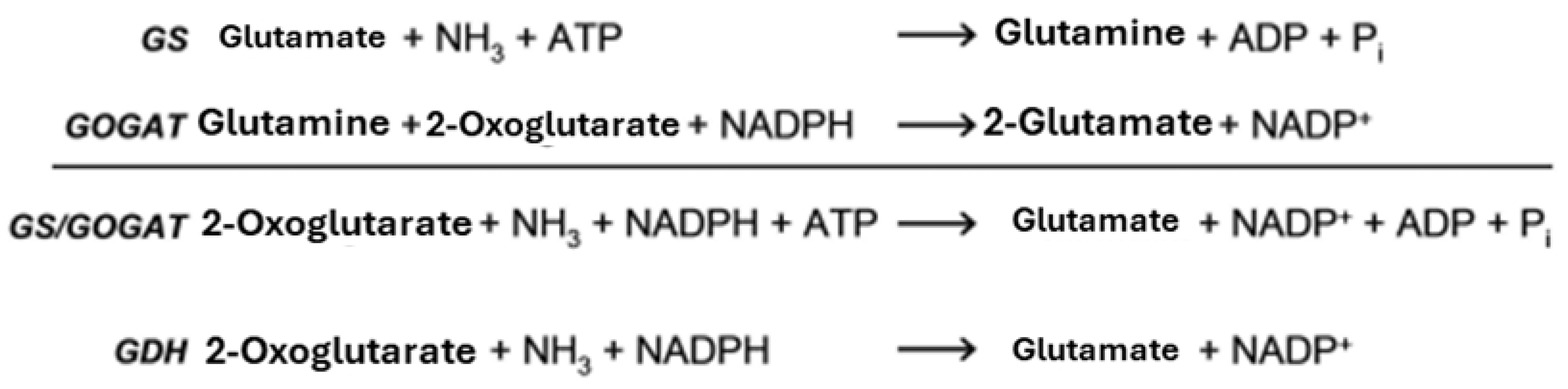
Figure 2.
Krebs cycle or tricarboxylic acid (TCA) cycle with reactions of the anabolic and catabolic glutamate dehydrogenases (modified after Voelker & Altaba, 2001; Merrick & Edwards, 1995).
Figure 2.
Krebs cycle or tricarboxylic acid (TCA) cycle with reactions of the anabolic and catabolic glutamate dehydrogenases (modified after Voelker & Altaba, 2001; Merrick & Edwards, 1995).
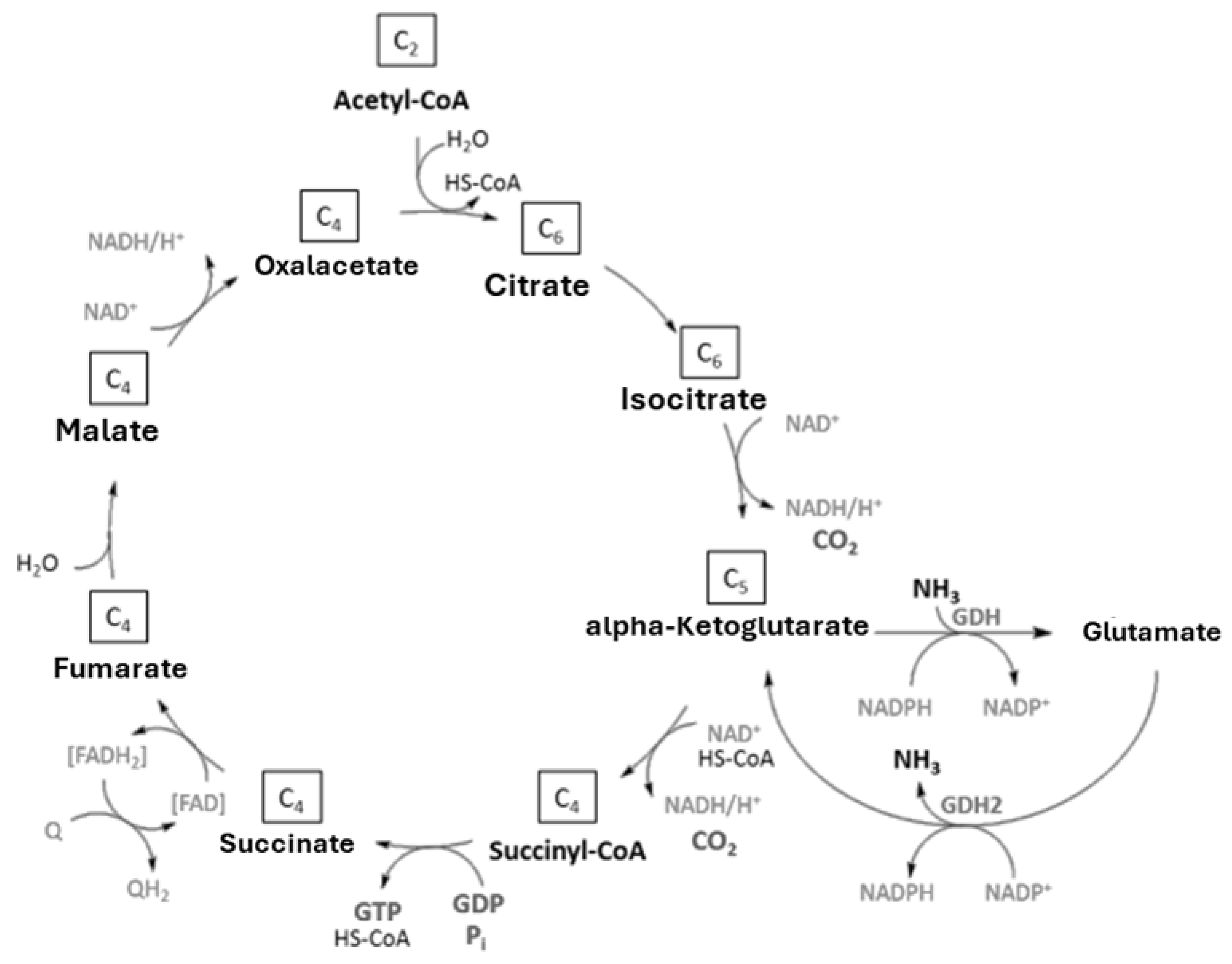
Figure 3.
Pathways of ammonium, nitrate, urea and amino acids assimilation in Streptomyces. The uptake of various nitrogen sources into the metabolic cycle occurs via conversion to ammonium and its assimilation via the central GS/GOGAT or GDH pathways. GS: Glutamine synthetase, GOGAT: Glutamate synthase, GDH: Glutamate dehydrogenase (modified after Krysenko & Wohlleben, 2022).
Figure 3.
Pathways of ammonium, nitrate, urea and amino acids assimilation in Streptomyces. The uptake of various nitrogen sources into the metabolic cycle occurs via conversion to ammonium and its assimilation via the central GS/GOGAT or GDH pathways. GS: Glutamine synthetase, GOGAT: Glutamate synthase, GDH: Glutamate dehydrogenase (modified after Krysenko & Wohlleben, 2022).
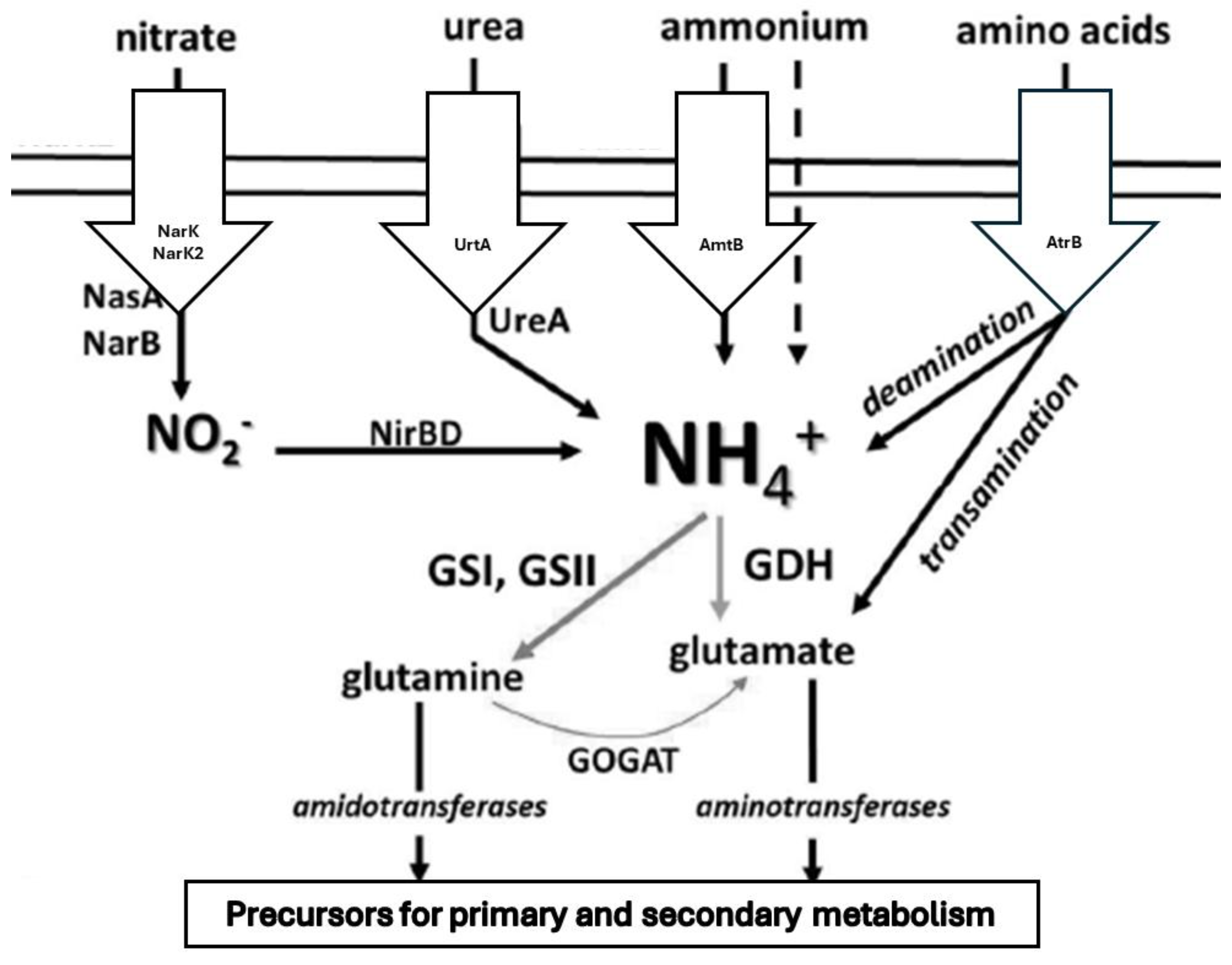
Figure 4.
Model of the polyamine putrescine utilization in Actinomycetes with links to polyamine biosynthesis and TCA cycle - the case study model system S. coelicolor (modified after Krysenko & Wohlleben, 2022).
Figure 4.
Model of the polyamine putrescine utilization in Actinomycetes with links to polyamine biosynthesis and TCA cycle - the case study model system S. coelicolor (modified after Krysenko & Wohlleben, 2022).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



