Submitted:
02 September 2025
Posted:
04 September 2025
You are already at the latest version
Abstract
MicroRNAs (miRNAs) are small non-coding RNAs that play pivotal roles in post-transcriptional gene regulation, influencing development, differentiation, and disease pathogenesis. Since their discovery in 1993, miRNAs have been recognized for their evolutionary conservation and pleiotropic effects, with recent Nobel Prize awards underscoring their significance in RNA interference (RNAi). This review synthesizes the complete life cycle of miRNAs —from transcription and processing to function and de-cay— emphasizing the regulatory mechanisms that govern these processes and their implications in human diseases, particularly cancer. We discuss how epitranscriptomic modifications influence miRNA biogenesis and activity, explore their nuclear and mi-tochondrial functions, and address emerging challenges in miRNA-based therapeutics, including the expanding small RNA landscape, such as tRNA-derived small RNAs (tsRNAs), and Argonaute (AGO)-independent activities. Despite hurdles such as modest multi-target effects and unvalidated specificity, miRNAs remain promising as both biomarkers and therapeutic agents, underscoring the need for sustained research to bridge preclinical insights with clinical applications.
Keywords:
miRNA biogenesis
; Argonaute
; TDMD
; miRNA modification
; tsRNA
1. Introduction
MicroRNAs (miRNAs) were first identified in 1993 by Victor Ambros and Gary Ruvkun in Caenorhabditis elegans [1]. Initially dismissed as anomalies, their importance became evident with the discovery of homologous sequences across species, revealing their evolutionary conservation and essential function in post-transcriptional gene regulation [2,3]. These ~22-nucleotide (nt) non-coding RNAs are now established as master regulators of gene expression, modulating processes such as development, cellular differentiation, and homeostasis [3].
The profound biological and clinical impact of miRNAs was highlighted by the 2024 Nobel Prize in Physiology or Medicine [4], building on the 2006 award for small interfering RNA (siRNA) discoveries. However, miRNA biology remains complex, characterized by pleiotropic effects, context-dependent functions, and unresolved therapeutic challenges. Despite their short length, miRNAs exhibit distinct functional regions: they typically begin with a 5'-phosphorylated uridine, followed by a 2–7 nt "seed sequence" that is complementary to target RNAs [5]. MiRNAs sharing the same seed belong to families, often regulating overlapping targets. This is succeeded by a central bulge and a 3'-supplementary sequence that partially pairs with targets, influencing regulatory efficacy [5]. Variations in base pairing yield diverse outcomes, positioning each miRNA as a three-dimensional entity whose interactions with proteins and RNAs dictate specific structures and effects.
Extensive studies —from genetic screens to animal models [6,7]— have implicated miRNAs in human diseases, including cancer [8], neurodegeneration [9], and autoimmune disorders [10]. Dysregulation at any biogenesis stage —transcription, processing, export, loading, or decay —can drive pathogenesis. This review integrates recent advances across the miRNA life cycle, focusing on novel regulatory mechanisms and disease implications, especially in oncology. We highlight structure-specific determinants beyond sequence alone, explore epitranscriptomic layers, and address translational opportunities and barriers in miRNA therapeutics.
2. Transcription of MiRNA
Approximately half of all currently identified miRNAs are intragenic —located within introns (and occasionally exons) of protein-coding genes— and are typically co-transcribed with their host genes [11,12]. The highly enriched human miRNA hairpins reside in intronic regions, suggesting that introns serve as evolutionary hotspots for miRNA emergence [11].
The remaining miRNAs are intergenic and transcribed independently under the control of their own promoters. Some miRNAs are organized into genomic clusters and are transcribed as a single long primary transcript (pri-miRNA), often sharing seed sequences and forming functional families [13], such as miR-17-92 cluster [14], miR-106a-363 cluster. MiRNAs from same family can also distribute to different gene loci, such as let-7 family, whose members located in seven different chromosomes [15].
Most miRNAs are transcribed by RNA polymerase II (Pol II) and, like other Pol II transcripts, carry a 5′ cap and a 3′ poly(A) tail [16,17]. However, a subset —including miR-515-1, miR-517a, and miR-519a-1 within the chromosome 19 miRNA cluster— is transcribed by RNA polymerase III [18]. Regardless of polymerase origin, all pri-miRNAs undergo multistep processing to become mature, functional miRNAs [16].
Many studies report that the overall level miRNA expression is downregulated in tumor tissues compared to normal counterparts [8,19,20,21], suggesting that disruptions in miRNA biogenesis —including transcriptional defects— may contribute to oncogenesis. While miRNAs can act as oncogenes or tumor suppressors depending on their cellular context and target genes, their expression is generally regulated at the transcriptional level, where they are initially generated.
2.1. Transcriptional Regulation
The transcription of miRNA genes is tightly controlled by transcription factors (TFs) and epigenetic modifications. Over 2,600 human miRNAs have been cataloged (e.g., in miRbase until 2025) and their expression is highly tissue- and cell-type-specific [22,23]. Although around 10% miRNA can be found in all tissues, no miRNA is uniformly expressed across all cell types and regarded as housekeeping [23]. About 10% miRNAs exhibit tissue-restricted expression which means they only express in specific cell types [23]. And in certain tissues, specific miRNAs dominate the miRNA pool [24] —for example, miR-290–295 in embryonic stem cells (ESCs) [25,26], miR-122 in the liver [24,27], and miR-1 in muscle [24,28].
The tissue specific is firstly taken account for their transcription by tissue specific TFs [29,30], such as MYOD, REST and OCT4. These TFs are critical for development and cell differentiation processes through regulating miRNAs. For example, MYOD induces muscle cell differentiation by transcriptionally upregulating the expression of miR-1, miR-133, and miR-206, which in turn target downstream genes such as PAX3 and UTRN, inhibitors of muscle differentiation [31,32]. While REST, a repressor of neuronal genes, can inhibit the expression of brain-specific miRNAs like miR-124a, miR-9, and miR-132 in non-neuronal cells, influencing neuronal identity [33,34]. In pluripotent stem cells, OCT4, SOX2, and KLF4 suppress differentiation-promoting miRNAs such as miR-145 to maintain self-renewal [35,36].
The transcription of miRNAs is under the control of epigenetic regulation. Both DNA methylation and histone modifications modulate miRNA promoter activity [37]. Super-enhancers (SEs) are large clusters of enhancers that play a crucial role in gene expression by recruiting high levels of transcription factors and co-activators like the Mediator complex [38]. They are also characterized by specific epigenetic modifications, including histone modifications like H3K27ac and H3K4me1 and are central to the maintenance of identity of cancer cells [39,40]. SE-miRNAs and typical-enhancer-associated miRNAs dominated about 60% and 20%, respectively, of the total miRNAs pool in mouse ES cells [38]. Change of epigenetic modification especially in SE region could significantly switch miRNA profile and drive the cellular identification switch. (Figure 1A)
2.2. Mutations of Transcriptional Regulators
Genetic mutations affecting miRNA loci or their transcriptional regulators can profoundly disrupt miRNA expression and contribute to disease, particularly in cancer. Although most current studies on tumor mutations have focused on protein-coding genes, substantial evidence indicates that mutations in miRNA genes and in genes regulating miRNAs also play important roles in tumor initiation and progression [41].
Deletions or point mutations in miRNA loci directly impair miRNA expression. For instance, miR-15a and miR-16-1, which are located at 13q14, are deleted in approximately 70% of chronic lymphocytic leukemia cases, leading to dysregulation of pro-survival genes [42].
More commonly, mutations or dysregulation in transcription factors and epigenetic regulators have broader effects on the miRNA landscape. Mutations in transcription factors like MYC are commonly associated with altered miRNA expression in various tumors, it activates the transcription of the miR-17-92 cluster to maintain survival, autonomous proliferation, and self-renewal [43]. At same time, MYC represses tumor-suppressive let-7 family members by binding upstream of the let-7a-1/let-7f-1/let-7d cluster [44]. The tumor suppressor p53 upregulates the transcription of the miR-34 family. MiR-34s act as tumor suppressors by targeting oncogenes, affecting cell proliferation, apoptosis, and metastasis [45,46]. In addition, ZEB1 and ZEB2, which are known as key activators to promote the epithelial-mesenchymal transition (EMT), repress transcription of the miR-200 family. Mir-200 is a tumor-suppressors that targets a series of EMT related genes including ZEB1/2, SNAIL, VIM and its expression is highly related to cancer progression in various cancer [47]. (Figure 1A)
Epigenetic dysregulation alone is able to induce cancer in the absence of driver mutations [48] and also able to induce cancer-related miRNAs expression. Several studies have shown that tumor suppressor miRNAs, such as miR-124a in acute lymphoblastic leukemia, can be silenced by aberrant methylation of CpG islands adjacent to their promoters [49]. And the frequency of human miRNA gene methylation is nearly one order of magnitude higher than that of the protein-encoding genes [50]. Epigenetic modifiers such as HDAC and TET enzymes are often mutated or mis-regulated in cancer, making them attractive therapeutic targets [51]. HDACs are overexpressed in Chronic lymphocytic leukemia and mediate the epigenetic silencing of miR-15a, miR-16, and miR-29b. HDAC inhibition have been found trigger the accumulation of the transcriptionally activating chromatin modification H3K4me2 and restored the expression of miR-15a, miR-16, and miR-29b in approximately 35% of patient samples [52].
Environmental chemicals can induce epigenetic modifications and change miRNA profiles. Exposure to heavy metals like cadmium and arsenic can alter DNA methylation patterns and impact miRNA expression, potentially contributing to cancer development. For example, increased expression of miR-146a was linked to cadmium exposure in steel workers [53]. The reversible nature of epigenetic alterations and the widespread impact of miRNA dysregulation make them attractive therapeutic targets [51].
In summary, miRNA transcription is controlled by a complex interplay of transcription factors and epigenetic modifications. Mutations or dysregulation at any of these levels can alter miRNA expression, contributing to oncogenesis. The high tissue specificity of miRNA transcription also makes them valuable biomarkers and potential therapeutic targets in cancer.
3. Processing in the Nucleus
Following transcription, primary miRNAs (pri-miRNAs) undergo essential maturation steps within the nucleus. These pri-miRNAs are typically long transcripts containing one or more stem-loop structures, which are recognized and cleaved by the Microprocessor complex —a minimal unit composed of the RNase III enzyme DROSHA and the double-stranded RNA-binding protein DGCR8 [16]. (Figure 1B)
MiRNA processing by the Microprocessor complex is referred to as the canonical pathway, which is responsible for producing the majority of miRNAs. In addition to this dominant route, several non-canonical pathways also contribute to miRNA biogenesis. These include mirtrons, which account for approximately 15% of human miRNAs and originate from spliced introns, bypassing DROSHA processing [54,55], and simtrons, which depend on DROSHA but are independent of splicing [56]. Despite the existence of alternative pathways, the canonical route remains predominant as knockout of DGCR8 results in more than a 95% global reduction in mature miRNAs in the skin cells, highlighting the essential role of the Microprocessor [57].
3.1. The Microprocessor Complex
The Microprocessor complex initiates pri-miRNA maturation in the nucleus. DGCR8 first binds to the stem-loop region of the pri-miRNA and recruits DROSHA, which cleaves the RNA to produce a ~70 nt precursor miRNA (pre-miRNA) with a characteristic 2-nt 3′ overhang. This cleavage event represents a key commitment step in miRNA biogenesis [16].
Structurally, the Microprocessor is a ~364 kDa heterotrimer composed of one DROSHA and two DGCR8 subunits. DROSHA (~160 kDa) serves as the catalytic core and contains multiple functional domains: an N-terminal proline- and RS-rich region for nuclear localization, a central conserved domain including a platform and PAZ-like domain, a connector region, two RNase III domains (RIIIDa and RIIIDb), and a C-terminal double strand RNA-binding domain (dsRBD). These elements allow DROSHA to precisely recognize the basal junction of the pri-miRNA and cleave ~11 basepair (bp) from the base and ~22 bp from the apical loop [16,58,59].
DGCR8 (~86 kDa per monomer) stabilizes and activates DROSHA. It contains a heme-binding domain (RHED), which is essential for dimerization and activity, as well as two dsRBDs that recognize the conserved UGU motif in the apical loop. DGCR8 also directly interacts with DROSHA’s C-terminal region to ensure precise positioning of the cleavage site [59].
Beyond its core components, some proteins are considered as auxiliary components of Microprocessor complex. For instance, hnRNP A1 facilitates the processing of miR-18a from the polycistronic miR-17–92 cluster through binds to the terminal loop of pri-miR-18a and induces structural changes that make the cleavage site more accessible to DROSHA [60]. The efficiency of processing is also influenced by specific RNA sequence motifs (e.g., UG, CNNC, UGUG) and secondary structure, though most human pri-miRNAs lack all optimal features [16,61]. These limitations are often compensated by regulatory RNA-binding proteins (RBPs), such as QKI5 and NONO, which modulate the processing of certain miRNAs depending on their binding preferences [62].
3.2. Regulation of Nuclear Processing
The activity of the Microprocessor complex is finely regulated at multiple levels, including transcription, complex assembly, autoregulatory loops, and post-translational modifications (PTMs). A feedback mechanism exists between DROSHA and DGCR8: DROSHA cleaves a hairpin structure in the 5′ UTR of DGCR8 mRNA to reduce its expression, while DGCR8 stabilizes DROSHA protein levels. This reciprocal regulation ensures homeostatic control of Microprocessor complex activity [63].
RBPs can both promote and inhibit miRNA processing in the nuclear [62]. NF90 can inhibit the processing of certain miRNA precursors through compete pri-miRNA with Microprocessor [64]. And in some cancer, dysregulation of certain RBPs can alter the processing efficiency of miRNAs. LIN28 is overexpressed in various cancers, promoting cell growth and proliferation by blocking the processing of let-7 precursors into mature, functional miRNAs [65].
PTMs play a crucial role in tuning the activity and stability of Microprocessor components. For DROSHA, phosphorylation at Ser300/Ser302 by GSK3β and PLK1 enhances nuclear localization, promotes DGCR8 binding, and increases the catalytic activity of miRNAs such as miR-1248, miR-1306-5p, miR-2277-5p [66,67]. Acetylation protects DROSHA from ubiquitin-mediated degradation and is enhanced by HDAC inhibitors, such as Trichostatin A and nicotinamide, which also increase the expression of miRNA-143 [68]. Conversely, ubiquitination by the E3 ligase MDM2, downstream of mTORC1 signaling, targets DROSHA for degradation, leading to a global reduction in miRNA levels [69]. (Figure 1B)
DGCR8 is likewise subject to multilayered regulation. Phosphorylation at Tyr267 by ABL kinase enhances its processing activity of miR-34c in response to DNA damage [70]. SUMOylation at Lys707 stabilizes DGCR8 and improves its pri-miRNA binding capacity, promoting cancer cell migration and transformation [71]. Moreover, DGCR8 can bind heme via its RHED domain. Heme binding is essential for DGCR8’s proper structural conformation and productive interaction with DROSHA [72,73]. (Figure 1B)
Mutations in DROSHA or DGCR8 impair Microprocessor complex function, leading to a global reduction in miRNA levels in various cancers [74]. While some mutations can also alter their processing preference, leading to increased expression of oncogenic miRNAs [75]. DROSHA and DGCR8 can be either upregulated or downregulated in cancers, often depending on the cancer type, which suggests the tissue- and cell-specificity of miRNAs and the diverse roles they play in cancer development [8].
In summary, the Microprocessor complex functions as a central regulatory hub in miRNA biogenesis. Its activity is modulated by intrinsic structural domains, PTMs, binding partners, and homeostatic feedback. Disruption of any of these mechanisms can profoundly reshape the miRNA landscape, often promoting cancer progression.
3.3. Nuclear Export of Pre-miRNA
Following nuclear processing, pre-miRNAs with a 2-nt 3′ overhang and 5′ monophosphate are exported to the cytoplasm by Exportin-5 (XPO5) both through Ran-GTP–dependent and independent mechanisms [76]. XPO5 recognizes stem-loop structures longer than 16 bps. While variations at the 3′ end are tolerated, the presence of a 5′ overhang significantly impairs binding and export, underscoring the importance of precise Microprocessor cleavage [77,78].
Although XPO5 is not strictly essential for all pre-miRNA export, its knockout leads to a notable, albeit partial, reduction in mature miRNA levels [79]. This suggests some degree of functional redundancy, potentially involving other RNA-binding proteins or alternative exportins.
XPO5 activity is modulated by upstream signaling pathways. The ATM/AKT pathway enhances pre-miRNA export by promoting phosphorylation of NUP153, a nuclear pore protein that interacts with XPO5 [80]. In contrast, ERK-mediated phosphorylation of XPO5 inhibits its function, resulting in widespread suppression of miRNA maturation [81]. Importantly, XPO5 expression is frequently dysregulated in cancer and may function either as an oncogene or tumor suppressor, depending on the cellular context and cancer type [82]. (Figure 2C)
In summary, nuclear processing and export of miRNAs are complex and tightly regulated events. The Microprocessor complex, accessory RNA-binding proteins, and nuclear export machinery together ensure efficient and accurate miRNA maturation. Disruptions at any stage of this process can lead to aberrant miRNA profiles and contribute to pathogenesis, particularly in cancer.
4. Processing in the Cytoplasm
After export from the nucleus, pre-miRNAs enter the cytoplasm where they undergo final processing steps essential for producing mature, functional miRNAs. This stage is mainly orchestrated by the RNase III enzyme DICER and its cofactors, which generate a ~22 nt miRNA duplex that is subsequently loaded into AGO proteins to form the RNA-induced silencing complex (RISC). The precise processing of pre-miRNAs at this step critically determines the length, sequence, and strand selection of mature miRNAs, influencing their stability and gene regulatory function.
4.1. DICER-Mediated Cleavage
DICER is a large, multi-domain endoribonuclease responsible for converting pre-miRNAs into miRNA duplexes. It recognizes the characteristic hairpin structure of pre-miRNAs, which typically contain a 2-nt 3′ overhang and a ~70 nt stem-loop [16]. (Figure 2A)
The enzyme’s structure enables highly specific substrate recognition and precise cleavage. The N-terminal DExD/H-box helicase domain engages the terminal loop of the pre-miRNA, clamping and positioning the RNA for accurate processing. The adjacent DUF283 domain, structurally resembling dsRBD, may assist in binding single-stranded RNA regions and contribute to substrate recognition. The PAZ domain binds the 3′ overhang of the pre-miRNA, anchoring it firmly and functioning as a molecular ruler that measures a fixed distance (~22 nt) from the end to the cleavage site. Two RNase III domains (IIIa and IIIb) dimerize to catalyze staggered cuts on each RNA strand, generating the characteristic 2-nt 3′ overhang on the miRNA duplex. Finally, a C-terminal dsRBD further stabilizes the interaction with dsRNA substrates, enhancing cleavage fidelity. The spatial arrangement of these domains ensures DICER cleaves pre-miRNAs at consistent positions, producing duplexes of defined length crucial for downstream strand selection and function [83,84].
Structural variations within pre-miRNAs, such as differences in terminal loop size, 3′ overhang length, and internal bulges or mismatches, influence DICER’s binding and cleavage precision [83,85]. These features can create subtle shifts in cleavage sites, producing isomiRs—miRNA variants with sequence or length differences—which expand the regulatory repertoire of miRNAs.
The biological importance of DICER is supported by strong genetic evidence. Dicer1 knockout mice exhibit embryonic lethality, while conditional knockouts display various developmental defects [86,87,88,89]. Somatic mutations in the Dicer1 gene, especially in the RNase IIIb domain, have been identified in several human cancers including Wilms tumor, pleuropulmonary blastoma, and others [8,90]. These mutations disrupt processing of miRNAs, skewing miRNA populations and contributing to oncogenesis. Beyond cancer, altered Dicer expression or mutations are implicated in neurodegenerative diseases, metabolic disorders, and developmental defects, emphasizing the enzyme’s broad physiological importance [91].
Moreover, DICER’s activity and stability are dynamically regulated by PTMs. Phosphorylation via signaling pathways like ERK and mTORC2 can modulate DICER’s localization and catalytic efficiency, linking miRNA processing to extracellular cues and cellular states [92,93]. Ubiquitination, SUMOylation and glycosylation also regulate DICER turnover, allowing cells to finely tune miRNA production in response to stress, differentiation, or disease [94,95]. (FFigure 2A)
4.2. Regulation of Cytoplasmic Processing by TRBP and PACT
DICER does not act alone. It forms complexes with cofactors such as TRBP and PACT, which bind primarily to the helicase and DUF283 domains. These cofactors stabilize the DICER-pre-miRNA complex, enhance cleavage efficiency, and can influence cleavage site selection, thereby affecting miRNA isoform diversity [96,97,98].
TRBP contains three dsRNA-binding domains, although only the first two are primarily responsible for RNA binding. It enhances DICER’s catalytic efficiency by stabilizing the interaction between DICER and its pre-miRNA substrates [96]. It also serves as a molecular bridge facilitating the transfer of the miRNA duplex from DICER to AGO proteins, promoting efficient RISC assembly [99]. Beyond catalysis, TRBP modulates substrate selectivity, fine-tuning miRNA expression profiles [100,101]. TRBP’s role in miRNA processing also links it to antiviral defense mechanisms, as it interacts with the kinase PKR—a key sensor of dsRNA—thereby inhibiting PKR activation and preventing excessive cellular stress responses [102].
PACT, also a dsRBP with three dsRNA-binding domains, forms a functional complex with DICER and supports its catalytic activity. [97]. Structural studies show that PACT binds to DICER in a manner similar to TRBP, but through mutual exclusion, leading each to selectively promote miRNA processing [96]. At the same time, PACT is a known activator of PKR, promoting antiviral and stress-response signaling pathways. Within the cell, TRBP and PACT compete for PKR and DICER binding, creating a finely balanced system that coordinates stress and immune signaling with miRNA processing [103].
Interestingly, the DICER–TRBP and DICER–PACT complexes have distinct effects on miRNA biogenesis. For some pre-miRNA substrates, such as pre-miR-200a and pre-miR-34c, these complexes produce different isomiR patterns by varying cleavage sites. Experiments swapping their RNA-binding domains confirmed that these functional differences largely depend on their N-terminal dsRNA-binding domains. Moreover, TRBP generally promotes DICER stability and miRNA dicing, while PACT may inhibit processing under certain conditions, illustrating a nuanced regulatory interplay [98].
In addition to TRBP and PACT, many other RBPs have been reported to regulate DICER-mediated processing. For example, LIN28 bind to the terminal loop of pre-let-7, inhibiting DICER cleavage and promoting its degradation [104,105]. YB-1 , whose is often overexpressed in various cancers, binds the terminal loop of pre-miR-29b-2 and suppress the tumor-suppressing effects of miR-29b [106].
Thus, the DICER cofactors TRBP and PACT serve as critical regulators of cytoplasmic miRNA maturation, linking RNA processing to cellular stress responses and expanding the functional diversity of the miRNA pool. In addition, many cytoplasmic RBPs can regulate the maturation of specific miRNAs and contribute to cancer development.
4.3. MiRNA Loading and Strand Selection
Once DICER generates the miRNA duplex, the next step is to load one strand —the guide strand— into an AGO protein to form RISC, while the other, known as the passenger strand (miRNA*), is typically degraded. (Figure 2B)
Humans express four AGO proteins (AGO1–4), which share approximately 80% amino acid identity and have largely overlapping miRNA binding profiles [107,108]. These proteins have conserved domains (N-terminal, PAZ, MID, PIWI), forming a bilobed structure that interacts with miRNAs. AGO2 is unique in its catalytic “slicer” activity and critical role during early development [109], making it the most studied.
MiRNA loading is not a random process and strand selection depends on two main principles: AGO preferences and the thermodynamic asymmetry of the duplex ends [16,108,110]. AGO proteins preferentially bind miRNA strands whose 5′ end is less stably paired [111]. The MID domain specifically anchors the 5′ phosphate of the guide strand and has a preference for uridine at this position [112], which further biases strand selection.
Thermodynamic asymmetry arises because the two ends of the miRNA duplex differ in base-pairing stability; the strand with the weaker 5′ end is more readily loaded into AGO. This asymmetric loading ensures that the correct strand guides target recognition [110].
However, strand selection is more complex than these rules alone. In different tissues or under various physiological or pathological states, arm switching can occur, where the dominant strand loaded into AGO changes [113]. This phenomenon is influenced by RBPs, post-transcriptional miRNA modifications[114], and cellular chaperones such as Hsc70 and Hsp90, which facilitate the ATP-dependent loading of miRNA duplexes into AGO [115,116].
Notably, cancer-associated miRNA mutations or altered expression of miRNA regulators frequently disrupt strand selection and promote arm switching, contributing to oncogenic deregulation [110,117]. For example, miR-574-5p and miR-574-3p are inversely expressed and play opposite roles in gastric cancer progression, with a high 5p/low 3p expression pattern significantly associated with advanced TNM stages and poor patient prognosis [118]. This dynamic regulation of miRNA strand loading adds another layer of control to miRNA-mediated gene silencing and highlights potential diagnostic and therapeutic targets.
The cytoplasmic processing of miRNAs involves tightly regulated steps from DICER-mediated cleavage to RISC assembly. The multi-domain enzyme DICER works with cofactors TRBP and PACT to generate precise miRNA duplexes. Subsequent strand selection and loading into AGO proteins involve complex recognition of structural features and cellular context, ensuring functional miRNAs guide gene silencing. Disruption of any of these steps leads to widespread dysregulation implicated in cancer, neurodegeneration, and other diseases. Understanding these processes offers insights into miRNA biology and potential avenues for therapeutic intervention.
5. Function of MiRNA in RISC
Once mature miRNAs are loaded into AGOs to form RISC —a multiprotein complex centered on AGOs and guide RNAs— they become potent regulators of gene expression. About 60% of all the whole protein-coding gene population have at least one miRNA target site [119]. This percentage increases to over 80% among cancer-related genes [120]. The canonical role of RISC is to repress target mRNAs post-transcriptionally, primarily in the cytoplasm. However, recent advances have revealed diverse noncanonical functions extending beyond traditional mRNA silencing, including nuclear activities and epigenetic regulation. Moreover, mutations and PTMs of AGO proteins critically influence miRNA function and are increasingly linked to human diseases.
5.1. Canonical Function: Translational Inhibition and mRNA Destabilization
Although in many organisms the cleavage activity of AGOs function in immunity or regulation of endogenous gene expression, these functions seem to be largely absent in mammals [121]. Even though human AGO2 and AGO3 retain their catalytic activity in miRNA processing, it rarely cleaves target RNAs in vivo. Studies from other organisms and siRNA research have shown that target cleavage occurs only when the guide RNA and target exhibit near-perfect base pairing [122,123], even though recent high-throughput analyses suggest that the cleavage mechanism of human AGO2 is more complex than previously thought [124,125] and may require further investigation from a structural perspective of the complex, rather than viewing it solely as a simple linear base-pairing process. Notably, since miRNAs primarily regulate their targets through the seed sequence, their function is not heavily dependent on the cleavage activity of AGO.
In mammalian cells, miRNAs primarily function to repress gene expression by guiding RISC to partially complementary sequences typically located in the 3′ untranslated region (UTR) of target mRNAs [8,108]. This targeting leads to translational inhibition and mRNA destabilization, two intertwined mechanisms that together reduce protein output effectively. (Figure 3A)
Translational inhibition can be achieved through several mechanisms [126], most notably by blocking the initiation phase of protein synthesis. RISC interferes with the assembly of the eukaryotic initiation factor 4F (eIF4F) complex and prevents ribosome recruitment, thus reducing the rate at which target mRNAs are translated into protein [127]. This mode of repression can be rapid and reversible, allowing cells to fine-tune protein production in response to stimuli.
Alongside translational repression, miRNA-loaded RISC recruits GW182 proteins, which serve as scaffolds to recruit deadenylase complexes such as CCR4-NOT. The deadenylation of the mRNA poly(A) tail triggers decapping and exonucleolytic degradation, thereby decreasing mRNA stability and reducing transcript abundance [126,128]. This dual action ensures both immediate and sustained downregulation of target gene expression.
Dysregulation of these canonical pathways is a hallmark of many cancers. Tumor-suppressive miRNAs like the let-7 family repress oncogenes such as RAS and MYC, and their loss contributes to unchecked cell proliferation [129,130]. Conversely, oncomiRs such as miR-21 target tumor suppressor genes including PTEN and TPM1, promoting tumor cell survival and growth. The balance of miRNA-mediated gene repression is thus critical for maintaining cellular homeostasis and preventing malignancy [131,132].
5.2. Functions in the Nucleus, Mitochondria and Condensate
Beyond their well-known cytoplasmic functions, miRNAs and AGO proteins also operate within the nucleus, exerting regulatory effects on transcription, chromatin modification, and RNA processing.
Nuclear AGOs, directed by miRNAs, can localize to gene promoters and enhancers to influence chromatin accessibility and transcriptional activity. Histone methyltransferase euchromatic histone lysine methyltransferase 2 (EHMT2), which suppresses the expression of fumarate hydratase in nasopharyngeal carcinoma is recruited along with EZH2 by miR-584-3p in an AGO2-dependent manner to reduce matrix metalloproteinase 14 (MMP-14) expression in gastric cancer [133]. Endogenous nuclear miR-589 activates expression of COX-2 through sense promoter RNA-mediated histone modification by WDR5 [134].
MiRNAs also serve as enhancer trigger in the nucleus. MiR-24-1 can recruit transcriptional activators —RNA polymerase II, p300/CBP, and enhancer RNAs— to its neighboring genes, FBP1 and FANCI, through AGO2-dependent localization [135]. Many miRNAs have been found to influence the expression of their neighboring genes, highlighting their role in the cell-specific regulation of 3D chromatin organization [136].
MiRNAs also modulate alternative splicing by interacting with intronic sequences or splicing factors, thereby altering splice site selection and expanding proteomic diversity. AGOs have been detected binding intronic sequences, suggesting that nuclear RISC can associate with pre-mRNAs and regulate their splicing [137,138,139]. AGO3 can form a complex with SF3B3 to regulate pre-mRNA splicing, thereby restraining type 2 immunity [140]. In addition, miR-10b promotes glioma progression, in part, by binding U6 snRNA, a core component of the spliceosome, thereby affecting the splicing of genes such as CDC42 [141].
Besides nuclear RNAi, several miRNAs have also been identified in mitochondria, termed mitochondrial microRNAs (mitomiRs) [142]. MitomiRs such as miR-696, miR-532, miR-690, and miR-345-3p may play important roles in the early stages of heart failure. [143]. MitomiRs such as miR-92a-2-5p and let-7b-5p contribute to impaired mitochondrial gene expression and increased ROS production. [144]. Since cardiomyocytes are rich in mitochondria and identifying the mitomiRs will aid in early and accurate diagnosis, which could result in the initiation of an effective therapy and a reduced mortality rate.
RISCs often assemble into cytoplasmic non-membranous bodies, such as P-bodies and stress granules [145], which are driven by liquid–liquid phase separation with the assistance of other proteins [126]. In P-bodies, GW182-AGOs interactions promote phase separation and contribute to mRNA decay [146]. Under stress conditions (e.g., heat shock, nutrient deprivation, or viral infection), stress granules help cells survive adverse environments by temporarily storing some components and providing a site for RNA repair [147]. Due to the highly dynamic nature of biomolecular condensates [148], miRNAs can modulate mRNA structure, thereby facilitating the translation of specific transcripts. (Figure 3B)
Advanced RNA detection technologies have enabled the precise identification of miRNA subcellular localization, expanding our understanding of RNAi. RNAi can occur in both the nucleus and cytoplasm, where distinct molecular environments may confer different functional mechanisms. Even within the cytoplasm, the discovery of phase separation has revealed that miRNAs can exert diverse effects on gene regulation in response to a variety of environmental stimuli.
5.3. Post-Translational Regulation of RISC in Diseases
The function of RISC is tightly controlled by genetic and post-translational mechanisms, and their disruption contributes to various diseases, including cancer, neurological disorders. Beyond AGOs, several other RISC components —such as GEMIN3/4 and GW182— have also been found to undergo mutations or dysregulation in various diseases [74].
Mutations in AGOs have been identified in some diseases especially neurodevelopmental disorders [149,150,151] and their expression with associated with various cancer [152,153]. While AGO expression can be either upregulated or downregulated, consistent with other miRNA processing factors, this suggests that miRNA function is highly tissue- and stage-specific.
PTMs, particularly phosphorylation and ubiquitination, dynamically regulate AGO2 function under both physiological and pathological conditions. AGO2 undergoes a phosphorylation cycle on residues Ser824–834 regulated by CK1α and reversed by the Protein Phosphatase 6 complex (PP6). CK1α mediated phosphorylation alters AGO2’s interaction with target mRNAs, reducing its ability to recruit the silencing machinery. This phosphorylation promotes target mRNA release and remodeling of RISC activity. Conversely, PP6 dephosphorylates AGO2, restoring target binding and silencing capacity. Together, the CK1α–PP6 cycle acts a reversible switch that fine-tunes miRNA-mediated gene silencing in response to cellular signals. [154]. In additional, phosphorylation of neuronal AGO2 at Ser387 via AKT3 downstream of NMDA receptor activation enhances miRNA-mediated translational repression critical for synaptic plasticity and memory [155]. Hypoxia-induced ubiquitination of AGO2 is mediated by LUBAC, which catalyzes M1-linked ubiquitination that can be removed by OTULIN. This ubiquitination stabilizes mRNAs by interfering with miRNA-mediated silencing, thereby promoting tumor progression under low-oxygen conditions. Furthermore, AGO ubiquitination by ZSWIM8 can be triggered by the binding of miRNAs to specific target RNAs, a mechanism that will be discussed later. (Figure 2B)
Collectively, genetic and post-translational regulatory mechanisms act as molecular switches that controlling AGO2’s interaction, activity, and localization, with significant implications for development, cancer, and neuronal function.
In summary, miRNAs exert their gene regulatory functions predominantly through the RISC complex by mediating translational inhibition and mRNA destabilization, which constitute their canonical roles essential for maintaining cellular homeostasis. Beyond these classical functions, miRNAs and AGO proteins participate in diverse noncanonical activities within the nucleus and mitochondria, including transcriptional and epigenetic regulation as well as RNA processing, expanding their influence in gene expression control and cancer biology. Moreover, mutations and dynamic PTMs of AGO proteins critically shape miRNA functionality, with profound impacts on disease processes such as tumor progression, neurological disorders. Understanding these multilayered mechanisms provides valuable insights into miRNA biology and offers promising avenues for therapeutic intervention.
6. MiRNA Decay
MicroRNAs undergo a complex lifecycle, starting from biogenesis to eventual degradation. While various biogenesis byproducts are generally rapidly degraded, mature miRNAs are relatively stable. Their longevity primarily results from tight association with AGO proteins, which shield them from exonucleolytic attack and maintain their function within RISC. Nonetheless, miRNAs are not permanent cellular components; their turnover is controlled by regulated decay pathways in both the cytoplasm and nucleus, allowing dynamic modulation of gene expression.
6.1. Degradation of MiRNA Intermediates and Mature MiRNAs
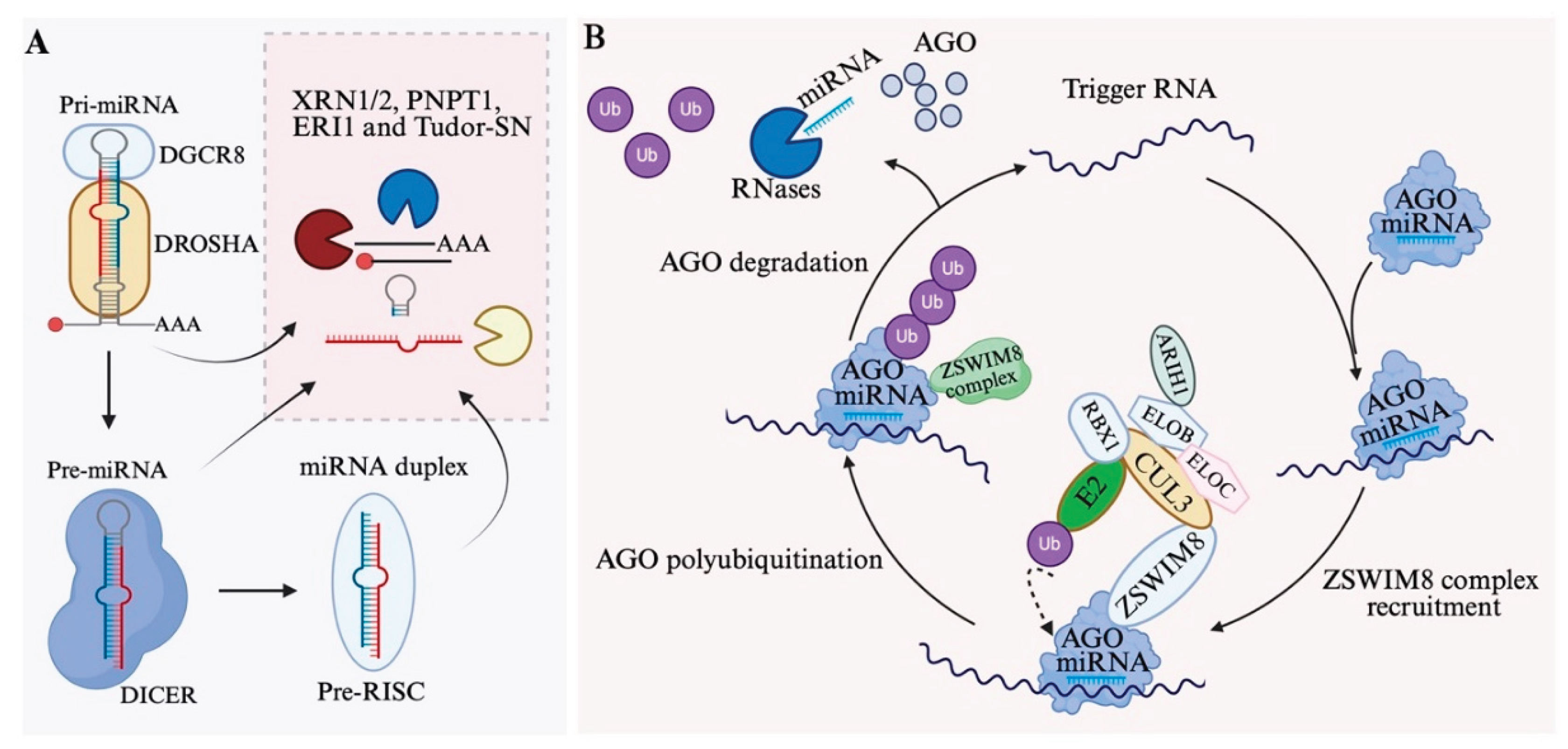
During miRNA maturation, several non-functional intermediates are produced, including the cleaved 5′ or 3′ fragments of pri-miRNAs, the terminal loop of pre-miRNAs, and the passenger strand from the miRNA duplex. These RNA fragments are rapidly eliminated by cellular RNA degradation machinery [79,156], such as 5′-to-3′ exoribonuclease 1/2 (XRN1/2) [157,158], 3′-to-5′ exoribonucleases PNPT1 [159] and ERI1 [160], and the endoribonuclease Tudor-SN [161]. Prompt clearance prevents unwanted interactions with RBPs and maintains cellular RNA homeostasis. (Figure 4A)
In contrast, mature miRNAs generally exhibit much longer half-lives, ranging from hours to days, owing to their stable incorporation into AGO proteins [156,162]. MiRNA half-lives are tightly regulated and display strong tissue specificity. MiRNAs are protected by AGO proteins, with their 5′ ends anchored in the MID domain, their internal regions shielded by surrounding AGOs to limit RNase accessibility, and their 3′ ends generally secured within the PAZ domain.
However, interactions with target RNAs or chemical modifications at the 3′ end can trigger conformational changes of RISCs, leading to the release of the miRNA 3′ end [125,163,164] and initiating its degradation through multiple mechanisms. The released 3′ end can undergo trimming by exonucleases or tailing by terminal nucleotidyl transferases. Terminal nucleotidyl transferases (TENTs) can modify the 3′ end of miRNAs, such as 3′ uridylation by TUT4/7, which promotes degradation by the U-specific exonuclease DIS3L2 [165].
6.2. Target-Directed miRNA Degradation
TDMD occurs when target RNAs have extensive complementarity to the miRNA's 3' supplementary sequence in addition to the seed region. This extended pairing extracts the 3' end of the miRNA from the PAZ domain and stabilizes a conformation of RISC that is distinct from that which occurs when the AGO–miRNA complex binds with less complementarity to a target [167]. ZSWIM8, a substrate receptor of a Cullin-RING E3 ubiquitin ligase (CRL), is able to recognize the RISC in TDMD and assembling with elongins B and C (ELOB and ELOC), cullin 3 (CUL3), ARIH1, and RBX1 to catalyze the transfer of an activated ubiquitin from an E2 ubiquitin-conjugating enzyme to AGO. Polyubiquitinated AGO is then degraded by the proteasome, releasing both the miRNA and its trigger RNA [168,169,170]. The released miRNAs are generally highly sensitive to RNases and been degraded soon. (Figure 4B)
TDMD is a conserved mechanism of miRNA decay from C.elegans to human [170]. A wide range of transcripts can serve as TDMD triggers and induce certain phenotypes through inhibiting miRNAs. The long noncoding RNA Cyrano directs efficient degradation of miR-7, thereby regulating neural activity [171]. Certain mRNAs, such as Serpine1, have been found to trigger the degradation of miR-30b/c [172], while BCL2L11 promotes miR-222 degradation, enhancing BIM-induced apoptosis in cancer cells [173]. Virus-derived snRNA-like RNA HSUR1 reduces T cell activation by triggering the degradation of miR-27a [166].
The importance of TDMD is underscored by genetic studies. Knockout models of Zswim8 exhibit severe developmental defects and perinatal lethality in mice, accompanied by aberrant accumulation of more than 50 miRNAs across 12 tissues [174,175]. Notably, deletion of two miRNAs, miR-322 and miR-503, can rescue growth of Zswim8-null embryos [175,176], illustrating the physiological relevance of tightly controlled miRNA degradation.
Although ZSWIM8 has been identified as a key factor in recognizing RISC during TDMD, the molecular features that target AGO for degradation remain to be elucidated, including how RNA–RNA pairing induces structural changes in AGO. Furthermore, additional studies are needed to catalog ZSWIM8 loss-of-function phenotypes and to determine the extent to which loss of TDMD contributes to these phenotypes.
7. RNA Modifications Add a New Regulatory Layer to MiRNA Biogenesis and Function
Recent research has uncovered a growing repertoire of chemical RNA modifications —collectively referred to as the epitranscriptome— that significantly influence miRNA biogenesis, stability, and function. Beyond well-characterized 3′ uridylation and adenylation, an expanding set of modifications —including adenosine-to-inosine (A-to-I) editing [177], N6-methyladenosine (m⁶A) [178], 5-methylcytosine (m5C) [179], 7-methylguanosine (m7G) [180], 2'-O-methylation [181], N4-acetylcytidine (ac4C) [182], 8-oxoguanine (8-oxoG) [183], 5′-O-methylation [184]—have been identified. As detection technologies continue to evolve [185], it is likely that many additional miRNA modifications remain to be uncovered.
MiRNA modifications function as regulatory switches superimposed on the miRNA pathway, dynamically modulating gene silencing in response to developmental signals, cellular stress, and disease states [16,186]. As many as 16% of human pri-miRNAs are subject to A-to-I editing, which can have a significant impact on miRNA processing [177]. And at least 6% of miRNAs show A-to-I editing that could change miRNA binding specificity [187]. These modifications exhibit strong tissue specificity and play critical roles in development and tissue differentiation. Their dysregulation has been observed in various diseases, including cancer, where they contribute to disease onset and progression.
7.1. RNA Modifications During MiRNA Biogenesis
RNA modifications, both internal and terminal, regulate the efficiency and accuracy of miRNA maturation from primary transcripts to functional strands loaded into AGO proteins through fine-tune their interaction with miRNA biogenesis regulators.
Adenosine deaminases acting on RNA (ADARs) catalyze A-to-I editing within double-stranded regions of pri- or pre-miRNAs. For example, editing of pri-miR-142 by ADAR1/2 inhibits DROSHA cleavage [188], while editing of pre-miR-151 prevents DICER processing [189]. Intriguingly, ADAR knockout selectively increases the levels of certain miRNAs, such as miR-142, because ADAR1 not only edits RNA but also interacts with DICER to promote miRNA processing through an editing-independent mechanism [190].
METTL3 enhances pri-miRNA processing by installing m⁶A marks on pri-miRNAs [178]. Mechanistically, distinct m⁶A reader proteins recognize these modified pri-miRNAs and recruit the Microprocessor complex to promote their cleavage —for example, hnRNPC facilitates the processing of pri-miR-21 [191], while hnRNPA2B1 promotes pri-miR-106b [192]. This modification not only boosts processing efficiency but may also facilitate nuclear export. NAT10-mediated ac⁴C on pri-miRNAs improves DGCR8 binding and promotes efficient processing. Elevated ac⁴C levels are found in some cancers, where this modification enhances miRNA maturation and contributes to altered gene expression [182].
Beyond internal modifications, both the 3′ and 5′ termini of miRNAs can undergo chemical alterations. At the 3′ end, tailing by TENTs introduces non-templated nucleotides —most commonly uridines or adenosines— onto pre- or mature miRNAs, which can either inhibit or promote their processing. For example, in pre-let-7 and miR-105 with a 1-nt 3′ overhang, mono-uridylation restores the canonical 2-nt overhang, thereby facilitating efficient DICER processing and miRNA biogenesis [193]. At the 5′ end, the monophosphate group is critical for AGO loading. Enzymes such as CLP1 restore 5′ phosphorylation of miR-34 in response to DNA damage, enabling its rapid activation [194]. Conversely, 5′-O-methylation by BCDIN3D neutralizes this charge and impairs DICER processing, acting as a gatekeeper in miRNA maturation [184].
7.2. Modifications Impacting miRNA function and stability
Beyond processing, RNA modifications can also directly influence miRNA function by altering seed sequences or modifying terminal ends. Most A-to-I editing sites fall within the seed sequences [195] and weaken complementarity with original targets, often resulting in reduced repression. At the same time, they can redirect the miRNA to novel targets, generating distinct regulatory outputs that can profoundly influence disease progression [196]. Reduced A-to-I editing of miR-376a in high-grade gliomas leads to an accumulation of the unedited form (miR-376a*A). This unedited form promotes glioma cell migration and invasion, whereas the edited form (miR-376a*G) suppresses these features. Specifically, unedited miR-376a*A targets RAP2A, promoting glioma cell invasion, while the edited miR-376a*G targets AMFR, suppressing invasion [197].
2′-O-methylation and m⁶A can enhance miRNA association with AGO or improve inhibition efficacy through enhance RISC stability. For instance, 2′-O-methylated miR-21-5p by HENMT1 exhibits greater AGO2 affinity and resistance to exonucleases, promoting sustained silencing [181]. MiRNA stability is also strongly influenced by 3′ terminal modifications. 2′-O-methylation generally protects miRNAs from exonucleolytic degradation, whereas oligo-uridylation typically serves as a degradation signal. For instance, LIN28 binds the terminal loop of pre-let-7 and recruits TUT4/7 to induce oligo-uridylation (rather than mono-uridylation), leading to pre-let-7 degradation by the 3′–5′ exonuclease DIS3L2 [198,199]. LIN28-mediated inhibition of let-7 plays a crucial role in regulating developmental timing, pluripotency, and glucose metabolism. Notably, Lin28 is frequently overexpressed in various cancers, resulting in elevated expression of oncogenes such as RAS, MYC, and HMGA2, which are normally targeted by let-7 [65].
Reactive oxygen species (ROS)-induced 8-oxoG formation in miRNAs can disrupt target pairing and alter their functional roles. For example, oxidized miR-184 misrecognizes BCL-xL and BCL-w, leading to their downregulation and influencing apoptosis [183]. Specifically, 8-oxoG can be predominantly introduced at position 7 of miR-1, and this single modification is sufficient to induce cardiac hypertrophy in mice. Notably, targeted inhibition of 8-oxoG–modified miR-1 in mouse cardiomyocytes attenuates hypertrophy [200]. In gliomas and hepatocellular carcinoma, oxidative modifications of miRNAs such as let-7 or miR-122 can shift their activity from tumor-suppressive to oncogenic —or vice versa— depending on the extent of oxidation [201]. ROS, generated in response to extracellular or intracellular stress, can contribute to disease—including cancer and neurodegenerative disorders—through miRNA modifications.
RNA modifications introduce a dynamic and adaptable layer of control over miRNA biogenesis and function. These modifications not only diversify the regulatory potential of miRNAs but also link environmental cues and cellular states to gene silencing programs. As our understanding of the epitranscriptome deepens, targeting these chemical switches holds promise for diagnostics and RNA-based therapeutics, particularly in cancer, neurodegeneration, and immune disorders.
8. Advances in MiRNA Research and Challenges in MiRNA Therapeutics
Many miRNAs have been identified in humans and other species, classical and noncanonical biogenesis pathways have been elucidated, various mechanisms and targets have been discovered, and their diverse roles in development and disease continue to be explored. These achievements contributed to the awarding of the Nobel Prize for miRNA research. However, compared with siRNAs —which were discovered earlier, received the Nobel Prize sooner, and have shown remarkable success in disease therapy— no miRNA-based therapy has yet advanced to phase III clinical trials or received FDA approval. This reflects both the complexity of miRNAs and our incomplete understanding of them. Recent discoveries have expanded our knowledge of miRNAs and other small RNAs, providing new insights for their potential clinical applications, including diagnostics and therapeutics.
8.1. Expansion of the Small RNA Landscape
For years, miRNAs were considered the predominant class of small regulatory RNAs. Although other types of small RNAs, such as tRNA-derived small RNAs (tsRNAs) [202] and rRNA-derived small RNAs (rsRNAs) [203], were occasionally detected in the early small RNA sequencing datasets, they were often dismissed as sequencing noise. However, biochemical analyses (e.g., PAGE gels) consistently reveal small RNA bands outside the canonical 20–24 nt miRNA range, particularly around 30–35 nt, where tsRNAs are enriched [204].
Traditional small RNA sequencing methods, which are biased RNAs with a 5′ phosphate and 3′ hydroxyl group, greatly favored miRNA detection. Emerging sequencing technologies, such as PANDORA-seq [205], overcome biases introduced by terminal and internal RNA modifications and have revealed a more complex small RNA landscape [204]. These studies demonstrate that housekeeping derived RNAs-including tsRNA and rsRNA-could dominate the small RNA pools across most tissues and cell types [206]. These RNAs are also present in AGO immunoprecipitants [207], suggesting that they may function through RISC or participate in the regulation of RNAi.
This broader small RNA spectrum raises critical questions about the role of miRNAs. Given that the copy number of AGO proteins in mammalian cells is approximately 1.5 × 10⁵ molecules per cell [208], miRNA abundance typically ranges from a few thousand to tens of thousands per cell [209], depending on the cell type. In contrast, tsRNAs and rsRNAs can be present at levels ten times higher than miRNAs, suggesting that they may occupy a substantial proportion of AGO proteins. In many miRNA studies, synthetic miRNAs are often overexpressed in cells, which can exaggerate their function and even cause off-target effects, rendering the results potentially questionable.
The function of small RNAs requires comprehensive investigation, as exemplified by reproductive studies highlighting the role of miRNAs in embryonic development. Intriguingly, tsRNAs is extremely enriched in mature mouse sperm [210]. Embryonic injection of small RNAs from mature sperm can mediate transgenerational phenotypes, with different classes of small RNAs exerting distinct biological effects: tsRNAs induce metabolic disorders in offspring, miRNAs can trigger embryonic lethality, whereas injection of total small RNAs allows embryos to survive, producing offspring with phenotypes resembling those induced by tsRNAs alone [211]. Although the precise mechanisms remain to be elucidated [212], tsRNAs may act as nutrient sensors [213,214] and regulate miRNA activity by competing for AGO loading, thereby modulating miRNA-mediated target repression, whose dysregulation can lead to embryonic lethality. Additionally, tsRNAs are generally heavily modified, a feature required for transgenerational inheritance [215], which may also differentially impact RISC function. These findings suggest that housekeeping-derived small RNAs provide a previously underappreciated layer of regulation, indirectly influencing miRNA function.
TsRNAs and rsRNAs are evolutionarily conserved small RNAs [216] that have been found to be dysregulated in cancer [217], neurodegeneration [218] and immune disorders [219,220]. Studying them from the perspective of miRNA regulation could provide novel insights into the miRNA field and help explain the inefficiencies observed in miRNA-based therapeutics.
8.2. AGO-Independent MiRNA Functions
MiRNA function has traditionally been understood through its association with AGO proteins, primarily mediating translational repression and mRNA degradation. However, emerging evidence indicates that miRNAs can also exert regulatory effects independent of AGO.
One example is miR-328, which functions as an RNA decoy by binding to hnRNP E2 and lifting its translational repression of CEBPA mRNA during myeloid differentiation [221]. Certain miRNAs, such as let-7, can interact with Toll-like receptors 7/8 (TLR7/8), key components of innate immunity [222]. These GU-rich miRNAs can act as ligands for TLR7/8, triggering pro-inflammatory responses [223]. Certain miRNAs can be secreted in extracellular vesicles (EVs) and modulate immune or neuronal activity in recipient cells [224]. When dysregulated, these mechanisms may contribute to autoimmune pathologies, particularly in the presence of elevated RBP-specific autoantibodies [225].
Additionally, some miRNAs appear to function as RNA aptamers, adopting specific conformations that enable them to bind directly to proteins. For example, miR-711 can bind ion channel TRPA1 extracellularly in sensory neurons and induce chronic itch [226], while miR-1 binds to cardiac potassium channel KIR2.1 in heart and modulates electrophysiological properties [227]. These findings suggest that miRNA secondary or tertiary structures, beyond their sequence, confer diverse functional capacities.
Given the structural flexibility of RNA, miRNA functionality can extend beyond classical RNAi, which relies heavily on linear base pairing. Induced modifications can stabilize their structure, enabling highly specific targeting. Exploring miRNAs as aptamers opens the door to precise modulation of receptors or ion channels, broadening their potential applications beyond gene silencing.
8.3. Clinical Outlook and Challenges
Despite the vast body of research and promising preclinical data, the clinical application of miRNA therapeutics lags behind that of siRNAs [228,229]. Several obstacles hinder miRNA drug development. The first challenge is the mild and multi-targeted activity of miRNAs. Typically, miRNAs exert modest effects on numerous targets, which complicates therapeutic design and dose optimization. Moreover, their specificity requires careful validation. Identifying direct miRNA targets remains difficult, as interactions observed in vitro often fail to replicate in vivo, and many detected changes may instead represent indirect effects [230]. Like a coin with two sides, although miRNAs can simultaneously target multiple genes, this also gives them the potential to regulate multiple pathogenic genes at once. By introducing miRNA mimics, it may be possible to simultaneously reduce the expression of several genes, thereby exerting unique effects in certain diseases or at specific stages of disease progression. Conversely, introducing miRNA inhibitors to reduce miRNA levels may restore the expression of certain genes, contributing to disease recovery. In summary, the clinical potential of miRNAs still depends on our understanding of diseases, particularly research on the specific functions of miRNAs within them.
In recent years, with the clinical application of mRNA and siRNA therapeutics, research on RNA delivery methods in vivo and on modulating their immunogenicity through chemical modifications has advanced rapidly [230,231]. These developments can also be applied to the field of miRNA-based therapeutics; however, miRNAs differ from other RNA drugs in their mechanisms of action, and it remains to be further evaluated whether such modification strategies would affect their functions [232].
Compared with the challenges faced by miRNAs in clinical therapeutics, they have shown greater potential in clinical diagnostics. MiRNAs have been found to be present in the biofluids, especially encapsulated within EVs [233]. As mentioned earlier, miRNAs display strong cell- and tissue-specific expression patterns, and under both physiological and pathological conditions, they can exist in different isoforms. These variations are often highly sensitive to disease progression, making miRNAs effective diagnostic biomarkers [233]. For example, serum-derived miR-205-5p has been identified as a promising biomarker candidate, capable of distinguishing between patients with pancreatitis and those with pancreatic cancer, with a reported accuracy of 91.5% [234]. Nevertheless, compared with circulating DNA detection, current methods for circulating RNA still require improvement in both accuracy and practicality, including sample storage [235].
In summary, miRNAs remain at the preclinical stage in both therapeutic and diagnostic applications. While their intrinsic characteristics currently limit their therapeutic use within the existing disease classification framework, their high sensitivity to disease states and the presence of distinct variants offer greater clinical potential in diagnostics.
9. Conclusions and Outlook
Over three decades since their discovery, miRNAs have evolved from obscure genetic elements to central pillars of gene regulation, with far-reaching implications for biology and medicine. This review has traversed their intricate life cycle —from transcription and multi-step biogenesis to canonical and non-canonical functions, decay pathways, and epitranscriptomic modifications— underscoring their roles in development, disease, and emerging therapeutic paradigms. Key insights include the tissue-specific transcriptional control, regulatory hubs like the Microprocessor and DICER complexes, diverse RISC-mediated silencing mechanisms extending to nuclear and mitochondrial compartments, and dynamic modifications that fine-tune miRNA activity in response to cellular cues.
Yet, challenges persist: the expanding small RNA landscape reveals miRNAs as part of a broader regulatory network, where tsRNAs and rsRNAs may modulate their potency. AGO-independent functions, such as decoy roles and aptamer-like interactions, further diversify miRNA biology, while therapeutic hurdles —like mild multi-target effects, delivery inefficiencies, and immunogenicity— have stalled clinical progress compared to siRNAs. Nonetheless, miRNAs' high sensitivity to disease states, tissue-specific expression, and isoform variations positions them as superior diagnostic biomarkers, with potential in liquid biopsies for early detection.
Future advances hinge on refined sequencing technologies, structural biology, and delivery innovations borrowed from mRNA therapeutics. By addressing these gaps, miRNA research can unlock targeted interventions for cancer, neurodegeneration, and beyond, transforming preclinical promise into clinical reality.
Figure 1.
Regulation of miRNA biogenesis in the nucleus. (A) Transcriptional regulation of miRNA genes. Transcription factors (TFs) and super-enhancers (SEs), together with epigenetic modifications, modulate the activity of RNA polymerase II/III at miRNA promoters. Representative examples include MYC induces miR-17–92 cluster and inhibits let-7 family), p53 induces miR-34), and ZEB1/2 inhibits miR-200. (B) Regulation of the Microprocessor complex. Pri-miRNAs are processed by DROSHA and its cofactor DGCR8. Representative examples of DROSHA/DGCT8 modifications: DROSHA nuclear localization is enhanced by phosphorylation at S300/S302 via GSK3β/PLK1, whereas MDM2-mediated ubiquitination promotes its degradation. DGCR8 activity is regulated by ABL-mediated phosphorylation at Y267 and heme binding, both of which promote pri-miRNA processing. (C) Nuclear export of pre-miRNAs. Exportin-5 (XPO5) transports pre-miRNAs from the nucleus to the cytoplasm. Representative examples of XPO5 regulation: XPO5 function is regulated by signaling pathways including AKT/ATM via NUP153 and ERK-mediated phosphorylation.
Figure 1.
Regulation of miRNA biogenesis in the nucleus. (A) Transcriptional regulation of miRNA genes. Transcription factors (TFs) and super-enhancers (SEs), together with epigenetic modifications, modulate the activity of RNA polymerase II/III at miRNA promoters. Representative examples include MYC induces miR-17–92 cluster and inhibits let-7 family), p53 induces miR-34), and ZEB1/2 inhibits miR-200. (B) Regulation of the Microprocessor complex. Pri-miRNAs are processed by DROSHA and its cofactor DGCR8. Representative examples of DROSHA/DGCT8 modifications: DROSHA nuclear localization is enhanced by phosphorylation at S300/S302 via GSK3β/PLK1, whereas MDM2-mediated ubiquitination promotes its degradation. DGCR8 activity is regulated by ABL-mediated phosphorylation at Y267 and heme binding, both of which promote pri-miRNA processing. (C) Nuclear export of pre-miRNAs. Exportin-5 (XPO5) transports pre-miRNAs from the nucleus to the cytoplasm. Representative examples of XPO5 regulation: XPO5 function is regulated by signaling pathways including AKT/ATM via NUP153 and ERK-mediated phosphorylation.
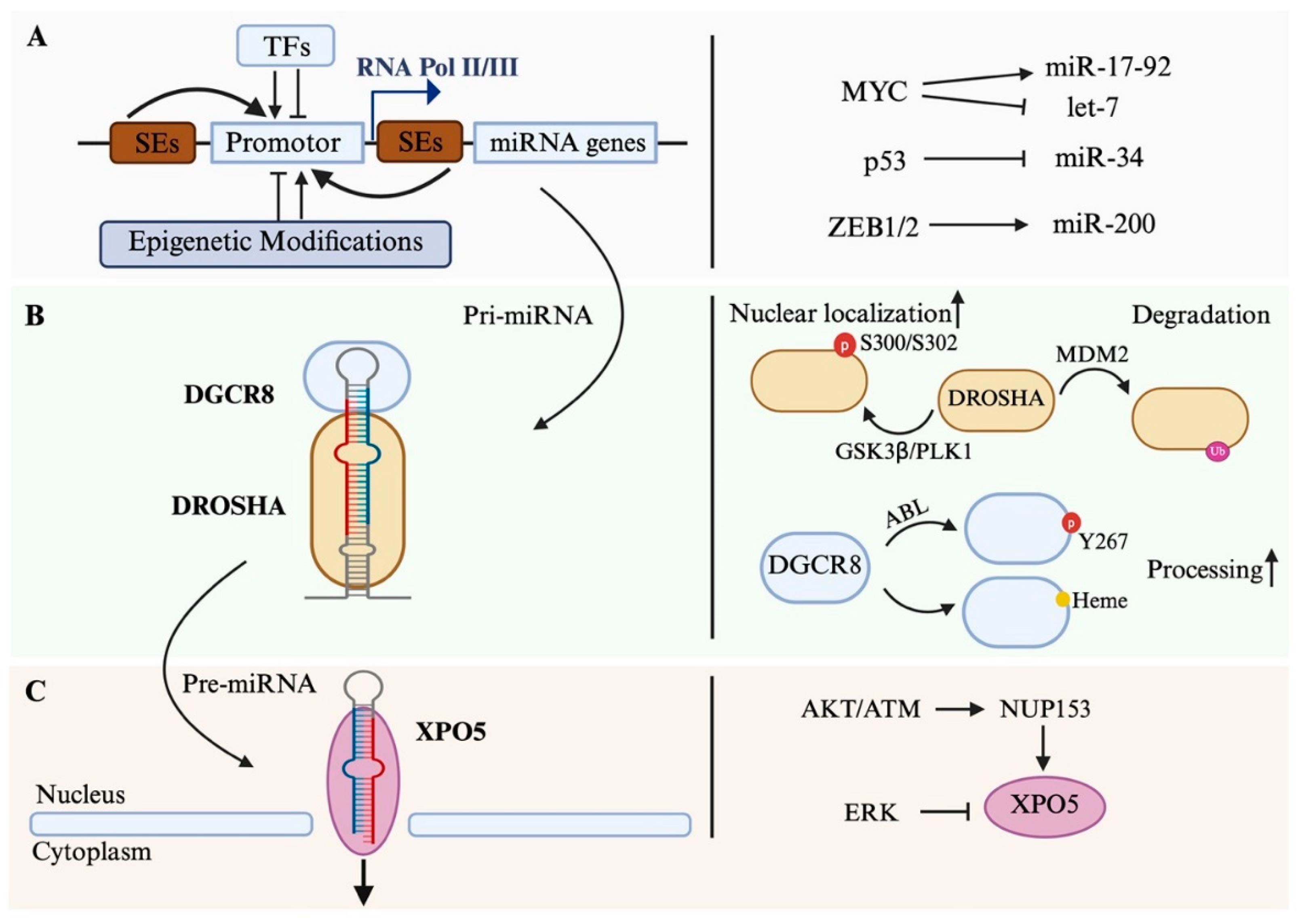
Figure 2.
Regulation of miRNA biogenesis in the cytoplasm. (A) Regulation of DICER. Pre-miRNAs are processed by DICER to generate mature miRNA duplexes. DICER catalytic efficiency is enhanced by ERK- and mTOR-mediated phosphorylation. Conversely, post-translational modifications such as SUMOylation and ubiquitination promote DICER degradation, reducing miRNA processing capacity. (B) Regulation of AGO and RISC assembly. After DICER cleavage, duplex miRNAs are loaded onto AGO to form a pre-RISC, followed by passenger strand removal to generate a mature RISC capable of target recognition. AGO function is dynamically regulated by post-translational modifications: (i) a CK1α–PP6 phosphorylation cycle at residues S824–834 controls target interaction and RISC remodeling; (ii) AKT3-mediated phosphorylation at S387 enhances miRNA-mediated translational inhibition; and (iii) ubiquitination mediated by LUBAC or ZSWIM8 promotes AGO degradation, reducing miRNA silencing activity.5. Conclusions
Figure 2.
Regulation of miRNA biogenesis in the cytoplasm. (A) Regulation of DICER. Pre-miRNAs are processed by DICER to generate mature miRNA duplexes. DICER catalytic efficiency is enhanced by ERK- and mTOR-mediated phosphorylation. Conversely, post-translational modifications such as SUMOylation and ubiquitination promote DICER degradation, reducing miRNA processing capacity. (B) Regulation of AGO and RISC assembly. After DICER cleavage, duplex miRNAs are loaded onto AGO to form a pre-RISC, followed by passenger strand removal to generate a mature RISC capable of target recognition. AGO function is dynamically regulated by post-translational modifications: (i) a CK1α–PP6 phosphorylation cycle at residues S824–834 controls target interaction and RISC remodeling; (ii) AKT3-mediated phosphorylation at S387 enhances miRNA-mediated translational inhibition; and (iii) ubiquitination mediated by LUBAC or ZSWIM8 promotes AGO degradation, reducing miRNA silencing activity.5. Conclusions
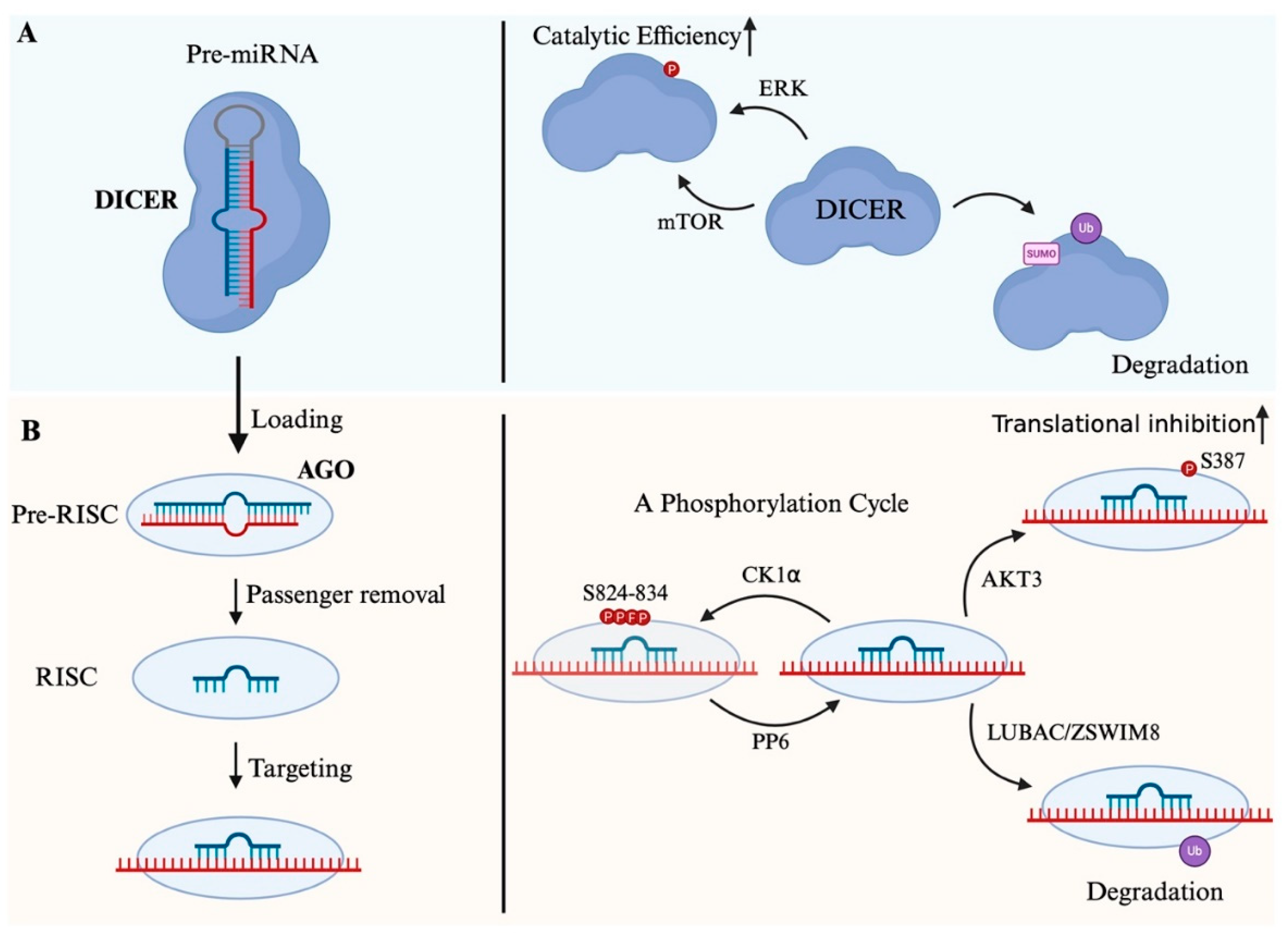
Figure 3.
Mechanisms of miRNA-mediated gene regulation. (A) Translational inhibition and mRNA destabilization: Translational inhibition occurs when miRNA-AGO complexes block ribosome initiation or post-initiation, while mRNA destabilization involves decapping (via DCP1/DCP2) and deadenylation (via CCR4-NOT, PAN2-PAN3, PABP) leading to decay. (B) Subcellular localization of RISC: RISCs can function in the nucleus and mitochondria, and can also form condensates in the cytoplasm.
Figure 3.
Mechanisms of miRNA-mediated gene regulation. (A) Translational inhibition and mRNA destabilization: Translational inhibition occurs when miRNA-AGO complexes block ribosome initiation or post-initiation, while mRNA destabilization involves decapping (via DCP1/DCP2) and deadenylation (via CCR4-NOT, PAN2-PAN3, PABP) leading to decay. (B) Subcellular localization of RISC: RISCs can function in the nucleus and mitochondria, and can also form condensates in the cytoplasm.

Figure 4.
MiRNA decay mechanisms. (A) Byproducts from miRNA processing are degraded by exonucleases (XRN1/2, PNPT1, ERI1) and endonucleases (Tudor-SN). (B) ZSWIM8 mediated Target-directed miRNA degradation (TDMD): AGO-miRNA extended pairing with trigger RNA (e.g., mRNA or lncRNA) extracts the miRNA 3' end from the AGO PAZ domain, stabilizing a distinct AGO conformation. This process, mediated by the ZSWIM8-Cullin-RING E3 ubiquitin ligase (ZSWIM8-CRL) complex (including ELOB, ELOC, CUL3, RBX1, and ARIH1), leads to AGO polyubiquitination and proteasomal degradation. The released miRNA is degraded by RNases, while the trigger RNA can bind a new AGO-miRNA complex, enabling multiple turnover-like activity and suprastoichiometric miRNA level reduction. Tailing or trimming of the miRNA 3' end may occur but is not required for TDMD.
Figure 4.
MiRNA decay mechanisms. (A) Byproducts from miRNA processing are degraded by exonucleases (XRN1/2, PNPT1, ERI1) and endonucleases (Tudor-SN). (B) ZSWIM8 mediated Target-directed miRNA degradation (TDMD): AGO-miRNA extended pairing with trigger RNA (e.g., mRNA or lncRNA) extracts the miRNA 3' end from the AGO PAZ domain, stabilizing a distinct AGO conformation. This process, mediated by the ZSWIM8-Cullin-RING E3 ubiquitin ligase (ZSWIM8-CRL) complex (including ELOB, ELOC, CUL3, RBX1, and ARIH1), leads to AGO polyubiquitination and proteasomal degradation. The released miRNA is degraded by RNases, while the trigger RNA can bind a new AGO-miRNA complex, enabling multiple turnover-like activity and suprastoichiometric miRNA level reduction. Tailing or trimming of the miRNA 3' end may occur but is not required for TDMD.
Acknowledgments
Illustrative figures are created with icon elements from Biorender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| RNAi | RNA interference |
| TF | Transcription factors |
| ESC | Embryonic stem cell |
| SE | Super enhancers |
| EMT | Epithelial-mesenchymal transition |
| dsRBD | Double strand RNA-binding domain |
| RBP | RNA-binding protein |
| PTM | Post-translational modification |
| RISC | RNA-induced silencing complex |
| UTR | Untranslated region |
| MitomiR | Mitochondrial microRNA |
| PP6 | Protein Phosphatase 6 complex |
| TENT | Terminal nucleotidyl transferase |
| TDMD | Target-Directed miRNA Degradation |
| ROS | Reactive oxygen species |
| tsRNA | tRNA-derived small RNA |
| rsRNA | rRNA-derived small RNA |
| EV | Extracellular vesicle |
References
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Almeida, M.I.; Reis, R.M.; Calin, G.A. MicroRNA history: discovery, recent applications, and next frontiers. Mutat Res 2011, 717, 1–8. [Google Scholar] [CrossRef]
- Callaway, E.; Sanderson, K. Medicine Nobel awarded for gene-regulating 'microRNAs'. Nature 2024, 634, 524–525. [Google Scholar] [CrossRef]
- Kilikevicius, A.; Meister, G.; Corey, D.R. Reexamining assumptions about miRNA-guided gene silencing. Nucleic Acids Res 2022, 50, 617–634. [Google Scholar] [CrossRef]
- Park, C.Y.; Choi, Y.S.; McManus, M.T. Analysis of microRNA knockouts in mice. Hum Mol Genet 2010, 19, R169–175. [Google Scholar] [CrossRef]
- Park, C.Y.; Jeker, L.T.; Carver-Moore, K.; Oh, A.; Liu, H.J.; Cameron, R.; Richards, H.; Li, Z.; Adler, D.; Yoshinaga, Y.; et al. A resource for the conditional ablation of microRNAs in the mouse. Cell Rep 2012, 1, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat Rev Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Eacker, S.M.; Dawson, T.M.; Dawson, V.L. Understanding microRNAs in neurodegeneration. Nat Rev Neurosci 2009, 10, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Pauley, K.M.; Cha, S.; Chan, E.K. MicroRNA in autoimmunity and autoimmune diseases. J Autoimmun 2009, 32, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Marsico, A.; Huska, M.R.; Lasserre, J.; Hu, H.; Vucicevic, D.; Musahl, A.; Orom, U.; Vingron, M. PROmiRNA: a new miRNA promoter recognition method uncovers the complex regulation of intronic miRNAs. Genome Biol 2013, 14, R84. [Google Scholar] [CrossRef] [PubMed]
- Ghorai, A.; Ghosh, U. miRNA gene counts in chromosomes vary widely in a species and biogenesis of miRNA largely depends on transcription or post-transcriptional processing of coding genes. Front Genet 2014, 5, 100. [Google Scholar] [CrossRef]
- Altuvia, Y.; Landgraf, P.; Lithwick, G.; Elefant, N.; Pfeffer, S.; Aravin, A.; Brownstein, M.J.; Tuschl, T.; Margalit, H. Clustering and conservation patterns of human microRNAs. Nucleic Acids Res 2005, 33, 2697–2706. [Google Scholar] [CrossRef] [PubMed]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: a comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ 2013, 20, 1603–1614. [Google Scholar] [CrossRef]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef]
- Kim, H.; Lee, Y.Y.; Kim, V.N. The biogenesis and regulation of animal microRNAs. Nat Rev Mol Cell Biol 2025, 26, 276–296. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA polymerase III transcribes human microRNAs. Nat Struct Mol Biol 2006, 13, 1097–1101. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Calin, G.A.; Lopez-Berestein, G.; Sood, A.K. miRNA Deregulation in Cancer Cells and the Tumor Microenvironment. Cancer Discov 2016, 6, 235–246. [Google Scholar] [CrossRef]
- Kumar, M.S.; Lu, J.; Mercer, K.L.; Golub, T.R.; Jacks, T. Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat Genet 2007, 39, 673–677. [Google Scholar] [CrossRef]
- Keller, A.; Groger, L.; Tschernig, T.; Solomon, J.; Laham, O.; Schaum, N.; Wagner, V.; Kern, F.; Schmartz, G.P.; Li, Y.; et al. miRNATissueAtlas2: an update to the human miRNA tissue atlas. Nucleic Acids Res 2022, 50, D211–D221. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stahler, C.; Meese, E.; et al. Distribution of miRNA expression across human tissues. Nucleic Acids Res 2016, 44, 3865–3877. [Google Scholar] [CrossRef] [PubMed]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr Biol 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Wang, Y.; Baskerville, S.; Shenoy, A.; Babiarz, J.E.; Baehner, L.; Blelloch, R. Embryonic stem cell-specific microRNAs regulate the G1-S transition and promote rapid proliferation. Nat Genet 2008, 40, 1478–1483. [Google Scholar] [CrossRef]
- Wu, D.R.; Gu, K.L.; Yu, J.C.; Fu, X.; Wang, X.W.; Guo, W.T.; Liao, L.Q.; Zhu, H.; Zhang, X.S.; Hui, J.; et al. Opposing roles of miR-294 and MBNL1/2 in shaping the gene regulatory network of embryonic stem cells. EMBO Rep 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Valdmanis, P.N.; Kim, H.K.; Chu, K.; Zhang, F.; Xu, J.; Munding, E.M.; Shen, J.; Kay, M.A. miR-122 removal in the liver activates imprinted microRNAs and enables more effective microRNA-mediated gene repression. Nat Commun 2018, 9, 5321. [Google Scholar] [CrossRef]
- Zhelankin, A.V.; Iulmetova, L.N.; Ahmetov, II; Generozov, E.V.; Sharova, E.I. Diversity and Differential Expression of MicroRNAs in the Human Skeletal Muscle with Distinct Fiber Type Composition. Life (Basel) 2023, 13. [CrossRef]
- Hobert, O. Common logic of transcription factor and microRNA action. Trends Biochem Sci 2004, 29, 462–468. [Google Scholar] [CrossRef]
- Guo, Z.; Maki, M.; Ding, R.; Yang, Y.; Zhang, B.; Xiong, L. Genome-wide survey of tissue-specific microRNA and transcription factor regulatory networks in 12 tissues. Sci Rep 2014, 4, 5150. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat Genet 2006, 38, 228–233. [Google Scholar] [CrossRef]
- Rosenberg, M.I.; Georges, S.A.; Asawachaicharn, A.; Analau, E.; Tapscott, S.J. MyoD inhibits Fstl1 and Utrn expression by inducing transcription of miR-206. J Cell Biol 2006, 175, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Wanet, A.; Tacheny, A.; Arnould, T.; Renard, P. miR-212/132 expression and functions: within and beyond the neuronal compartment. Nucleic Acids Res 2012, 40, 4742–4753. [Google Scholar] [CrossRef]
- Conaco, C.; Otto, S.; Han, J.J.; Mandel, G. Reciprocal actions of REST and a microRNA promote neuronal identity. Proc Natl Acad Sci U S A 2006, 103, 2422–2427. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Papagiannakopoulos, T.; Pan, G.; Thomson, J.A.; Kosik, K.S. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell 2009, 137, 647–658. [Google Scholar] [CrossRef]
- Divisato, G.; Passaro, F.; Russo, T.; Parisi, S. The Key Role of MicroRNAs in Self-Renewal and Differentiation of Embryonic Stem Cells. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. Epigenetic alteration and microRNA dysregulation in cancer. Front Genet 2013, 4, 258. [Google Scholar] [CrossRef]
- Suzuki, H.I.; Young, R.A.; Sharp, P.A. Super-Enhancer-Mediated RNA Processing Revealed by Integrative MicroRNA Network Analysis. Cell 2017, 168, 1000–1014 e1015. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Rai, S.N. Predicting the Effect of miRNA on Gene Regulation to Foster Translational Multi-Omics Research-A Review on the Role of Super-Enhancers. Noncoding RNA 2024, 10. [Google Scholar] [CrossRef]
- Tan, H.; Huang, S.; Zhang, Z.; Qian, X.; Sun, P.; Zhou, X. Pan-cancer analysis on microRNA-associated gene activation. EBioMedicine 2019, 43, 82–97. [Google Scholar] [CrossRef]
- Urbanek-Trzeciak, M.O.; Galka-Marciniak, P.; Nawrocka, P.M.; Kowal, E.; Szwec, S.; Giefing, M.; Kozlowski, P. Pan-cancer analysis of somatic mutations in miRNA genes. EBioMedicine 2020, 61, 103051. [Google Scholar] [CrossRef]
- Bonci, D.; Coppola, V.; Musumeci, M.; Addario, A.; Giuffrida, R.; Memeo, L.; D'Urso, L.; Pagliuca, A.; Biffoni, M.; Labbaye, C.; et al. The miR-15a-miR-16-1 cluster controls prostate cancer by targeting multiple oncogenic activities. Nat Med 2008, 14, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Choi, P.S.; Casey, S.C.; Dill, D.L.; Felsher, D.W. MYC through miR-17-92 suppresses specific target genes to maintain survival, autonomous proliferation, and a neoplastic state. Cancer Cell 2014, 26, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.C.; Yu, D.; Lee, Y.S.; Wentzel, E.A.; Arking, D.E.; West, K.M.; Dang, C.V.; Thomas-Tikhonenko, A.; Mendell, J.T. Widespread microRNA repression by Myc contributes to tumorigenesis. Nat Genet 2008, 40, 43–50. [Google Scholar] [CrossRef]
- Hermeking, H. p53 enters the microRNA world. Cancer Cell 2007, 12, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Chai, B.; Li, L.; Lu, Z.; Ma, Z. p53/MicroRNA-34 axis in cancer and beyond. Heliyon 2023, 9, e15155. [Google Scholar] [CrossRef]
- Jo, H.; Shim, K.; Jeoung, D. Potential of the miR-200 Family as a Target for Developing Anti-Cancer Therapeutics. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Parreno, V.; Loubiere, V.; Schuettengruber, B.; Fritsch, L.; Rawal, C.C.; Erokhin, M.; Gyorffy, B.; Normanno, D.; Di Stefano, M.; Moreaux, J.; et al. Transient loss of Polycomb components induces an epigenetic cancer fate. Nature 2024, 629, 688–696. [Google Scholar] [CrossRef]
- Agirre, X.; Vilas-Zornoza, A.; Jimenez-Velasco, A.; Martin-Subero, J.I.; Cordeu, L.; Garate, L.; San Jose-Eneriz, E.; Abizanda, G.; Rodriguez-Otero, P.; Fortes, P.; et al. Epigenetic silencing of the tumor suppressor microRNA Hsa-miR-124a regulates CDK6 expression and confers a poor prognosis in acute lymphoblastic leukemia. Cancer Res 2009, 69, 4443–4453. [Google Scholar] [CrossRef]
- Strmsek, Z.; Kunej, T. MicroRNA Silencing by DNA Methylation in Human Cancer: a Literature Analysis. Noncoding RNA 2015, 1, 44–52. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Koldobskiy, M.A.; Gondor, A. Epigenetic modulators, modifiers and mediators in cancer aetiology and progression. Nat Rev Genet 2016, 17, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Sampath, D.; Liu, C.; Vasan, K.; Sulda, M.; Puduvalli, V.K.; Wierda, W.G.; Keating, M.J. Histone deacetylases mediate the silencing of miR-15a, miR-16, and miR-29b in chronic lymphocytic leukemia. Blood 2012, 119, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Bollati, V.; Marinelli, B.; Apostoli, P.; Bonzini, M.; Nordio, F.; Hoxha, M.; Pegoraro, V.; Motta, V.; Tarantini, L.; Cantone, L.; et al. Exposure to metal-rich particulate matter modifies the expression of candidate microRNAs in peripheral blood leukocytes. Environ Health Perspect 2010, 118, 763–768. [Google Scholar] [CrossRef]
- Berezikov, E.; Chung, W.J.; Willis, J.; Cuppen, E.; Lai, E.C. Mammalian mirtron genes. Mol Cell 2007, 28, 328–336. [Google Scholar] [CrossRef]
- Salim, U.; Kumar, A.; Kulshreshtha, R.; Vivekanandan, P. Biogenesis, characterization, and functions of mirtrons. Wiley Interdiscip Rev RNA 2022, 13, e1680. [Google Scholar] [CrossRef]
- Havens, M.A.; Reich, A.A.; Duelli, D.M.; Hastings, M.L. Biogenesis of mammalian microRNAs by a non-canonical processing pathway. Nucleic Acids Res 2012, 40, 4626–4640. [Google Scholar] [CrossRef]
- Yi, R.; Pasolli, H.A.; Landthaler, M.; Hafner, M.; Ojo, T.; Sheridan, R.; Sander, C.; O'Carroll, D.; Stoffel, M.; Tuschl, T.; et al. DGCR8-dependent microRNA biogenesis is essential for skin development. Proc Natl Acad Sci U S A 2009, 106, 498–502. [Google Scholar] [CrossRef]
- Kwon, S.C.; Nguyen, T.A.; Choi, Y.G.; Jo, M.H.; Hohng, S.; Kim, V.N.; Woo, J.S. Structure of Human DROSHA. Cell 2016, 164, 81–90. [Google Scholar] [CrossRef]
- Partin, A.C.; Zhang, K.; Jeong, B.C.; Herrell, E.; Li, S.; Chiu, W.; Nam, Y. Cryo-EM Structures of Human Drosha and DGCR8 in Complex with Primary MicroRNA. Mol Cell 2020, 78, 411–422 e414. [Google Scholar] [CrossRef] [PubMed]
- Guil, S.; Caceres, J.F. The multifunctional RNA-binding protein hnRNP A1 is required for processing of miR-18a. Nat Struct Mol Biol 2007, 14, 591–596. [Google Scholar] [CrossRef]
- Garg, A.; Shang, R.; Cvetanovic, T.; Lai, E.C.; Joshua-Tor, L. The structural landscape of Microprocessor-mediated processing of pri-let-7 miRNAs. Mol Cell 2024, 84, 4175–4190 e4176. [Google Scholar] [CrossRef]
- Hynes, C.; Kakumani, P.K. Regulatory role of RNA-binding proteins in microRNA biogenesis. Front Mol Biosci 2024, 11, 1374843. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Pedersen, J.S.; Kwon, S.C.; Belair, C.D.; Kim, Y.K.; Yeom, K.H.; Yang, W.Y.; Haussler, D.; Blelloch, R.; Kim, V.N. Posttranscriptional crossregulation between Drosha and DGCR8. Cell 2009, 136, 75–84. [Google Scholar] [CrossRef]
- Grasso, G.; Higuchi, T.; Mac, V.; Barbier, J.; Helsmoortel, M.; Lorenzi, C.; Sanchez, G.; Bello, M.; Ritchie, W.; Sakamoto, S.; et al. NF90 modulates processing of a subset of human pri-miRNAs. Nucleic Acids Res 2020, 48, 6874–6888. [Google Scholar] [CrossRef]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/let-7 Pathway in Cancer. Front Genet 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Li, M.; Tucker, L.; Ramratnam, B. Glycogen synthase kinase 3 beta (GSK3beta) phosphorylates the RNAase III enzyme Drosha at S300 and S302. PLoS One 2011, 6, e20391. [Google Scholar] [CrossRef]
- Fletcher, C.E.; Taylor, M.A.; Bevan, C.L. PLK1 Regulates MicroRNA Biogenesis through Drosha Phosphorylation. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Wen, S.; Zheng, D.; Tucker, L.; Cao, L.; Pantazatos, D.; Moss, S.F.; Ramratnam, B. Acetylation of drosha on the N-terminus inhibits its degradation by ubiquitination. PLoS One 2013, 8, e72503. [Google Scholar] [CrossRef]
- Ye, P.; Liu, Y.; Chen, C.; Tang, F.; Wu, Q.; Wang, X.; Liu, C.G.; Liu, X.; Liu, R.; Liu, Y.; et al. An mTORC1-Mdm2-Drosha axis for miRNA biogenesis in response to glucose- and amino acid-deprivation. Mol Cell 2015, 57, 708–720. [Google Scholar] [CrossRef]
- Tu, C.C.; Zhong, Y.; Nguyen, L.; Tsai, A.; Sridevi, P.; Tarn, W.Y.; Wang, J.Y. The kinase ABL phosphorylates the microprocessor subunit DGCR8 to stimulate primary microRNA processing in response to DNA damage. Sci Signal 2015, 8, ra64. [Google Scholar] [CrossRef]
- Zhu, C.; Chen, C.; Huang, J.; Zhang, H.; Zhao, X.; Deng, R.; Dou, J.; Jin, H.; Chen, R.; Xu, M.; et al. SUMOylation at K707 of DGCR8 controls direct function of primary microRNA. Nucleic Acids Res 2015, 43, 7945–7960. [Google Scholar] [CrossRef]
- Quick-Cleveland, J.; Jacob, J.P.; Weitz, S.H.; Shoffner, G.; Senturia, R.; Guo, F. The DGCR8 RNA-binding heme domain recognizes primary microRNAs by clamping the hairpin. Cell Rep 2014, 7, 1994–2005. [Google Scholar] [CrossRef]
- Partin, A.C.; Ngo, T.D.; Herrell, E.; Jeong, B.C.; Hon, G.; Nam, Y. Heme enables proper positioning of Drosha and DGCR8 on primary microRNAs. Nat Commun 2017, 8, 1737. [Google Scholar] [CrossRef]
- Galka-Marciniak, P.; Urbanek-Trzeciak, M.O.; Nawrocka, P.M.; Kozlowski, P. A pan-cancer atlas of somatic mutations in miRNA biogenesis genes. Nucleic Acids Res 2021, 49, 601–620. [Google Scholar] [CrossRef] [PubMed]
- Torrezan, G.T.; Ferreira, E.N.; Nakahata, A.M.; Barros, B.D.; Castro, M.T.; Correa, B.R.; Krepischi, A.C.; Olivieri, E.H.; Cunha, I.W.; Tabori, U.; et al. Recurrent somatic mutation in DROSHA induces microRNA profile changes in Wilms tumour. Nat Commun 2014, 5, 4039. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lee, J.E.; Riemondy, K.; Yu, Y.; Marquez, S.M.; Lai, E.C.; Yi, R. XPO5 promotes primary miRNA processing independently of RanGTP. Nat Commun 2020, 11, 1845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, F.; Yang, F.; Meng, Z.; Zeng, Y. Selectivity of Exportin 5 binding to human precursor microRNAs. RNA Biol 2021, 18, 730–737. [Google Scholar] [CrossRef]
- Zeng, Y.; Cullen, B.R. Structural requirements for pre-microRNA binding and nuclear export by Exportin 5. Nucleic Acids Res 2004, 32, 4776–4785. [Google Scholar] [CrossRef]
- Kim, Y.K.; Kim, B.; Kim, V.N. Re-evaluation of the roles of DROSHA, Export in 5, and DICER in microRNA biogenesis. Proc Natl Acad Sci U S A 2016, 113, E1881–1889. [Google Scholar] [CrossRef]
- Wan, G.; Zhang, X.; Langley, R.R.; Liu, Y.; Hu, X.; Han, C.; Peng, G.; Ellis, L.M.; Jones, S.N.; Lu, X. DNA-damage-induced nuclear export of precursor microRNAs is regulated by the ATM-AKT pathway. Cell Rep 2013, 3, 2100–2112. [Google Scholar] [CrossRef]
- Sun, H.L.; Cui, R.; Zhou, J.; Teng, K.Y.; Hsiao, Y.H.; Nakanishi, K.; Fassan, M.; Luo, Z.; Shi, G.; Tili, E.; et al. ERK Activation Globally Downregulates miRNAs through Phosphorylating Exportin-5. Cancer Cell 2016, 30, 723–736. [Google Scholar] [CrossRef]
- Wu, K.; He, J.; Pu, W.; Peng, Y. The Role of Exportin-5 in MicroRNA Biogenesis and Cancer. Genomics Proteomics Bioinformatics 2018, 16, 120–126. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Lee, H.; Kim, H.; Kim, V.N.; Roh, S.H. Structure of the human DICER-pre-miRNA complex in a dicing state. Nature 2023, 615, 331–338. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, J.; Cheng, H.; Ke, X.; Sun, L.; Zhang, Q.C.; Wang, H.W. Cryo-EM Structure of Human Dicer and Its Complexes with a Pre-miRNA Substrate. Cell 2018, 173, 1191–1203 e1112. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Trinh, T.A.; Bao, S.; Nguyen, T.A. Secondary structure RNA elements control the cleavage activity of DICER. Nat Commun 2022, 13, 2138. [Google Scholar] [CrossRef]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer is essential for mouse development. Nat Genet 2003, 35, 215–217. [Google Scholar] [CrossRef]
- Chen, J.F.; Murchison, E.P.; Tang, R.; Callis, T.E.; Tatsuguchi, M.; Deng, Z.; Rojas, M.; Hammond, S.M.; Schneider, M.D.; Selzman, C.H.; et al. Targeted deletion of Dicer in the heart leads to dilated cardiomyopathy and heart failure. Proc Natl Acad Sci U S A 2008, 105, 2111–2116. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, S.; Lee, M.; Akimoto, T. Conditional Deletion of Dicer in Adult Mice Impairs Skeletal Muscle Regeneration. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- De Cauwer, A.; Loustau, T.; Erne, W.; Pichot, A.; Molitor, A.; Stemmelen, T.; Carapito, R.; Orend, G.; Bahram, S.; Georgel, P. Dicer1 deficient mice exhibit premature aging and metabolic perturbations in adipocytes. iScience 2022, 25, 105149. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.C.; Jorcyk, C.L.; Oxford, J.T. DICER1 Syndrome: DICER1 Mutations in Rare Cancers. Cancers (Basel) 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Theotoki, E.I.; Pantazopoulou, V.I.; Georgiou, S.; Kakoulidis, P.; Filippa, V.; Stravopodis, D.J.; Anastasiadou, E. Dicing the Disease with Dicer: The Implications of Dicer Ribonuclease in Human Pathologies. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Chou, P.C.; Rajput, S.; Zhao, X.; Patel, C.; Albaciete, D.; Oh, W.J.; Daguplo, H.Q.; Patel, N.; Su, B.; Werlen, G.; et al. mTORC2 Is Involved in the Induction of RSK Phosphorylation by Serum or Nutrient Starvation. Cells 2020, 9. [Google Scholar] [CrossRef]
- Reyes-Castro, R.A.; Chen, S.Y.; Seemann, J.; Kundu, S.T.; Gibbons, D.L.; Arur, S. Phosphorylated nuclear DICER1 promotes open chromatin state and lineage plasticity of AT2 tumor cells in lung adenocarcinomas. Sci Adv 2023, 9, eadf6210. [Google Scholar] [CrossRef]
- Lai, H.H.; Li, J.N.; Wang, M.Y.; Huang, H.Y.; Croce, C.M.; Sun, H.L.; Lyu, Y.J.; Kang, J.W.; Chiu, C.F.; Hung, M.C.; et al. HIF-1alpha promotes autophagic proteolysis of Dicer and enhances tumor metastasis. J Clin Invest 2018, 128, 625–643. [Google Scholar] [CrossRef]
- Kurzynska-Kokorniak, A.; Koralewska, N.; Pokornowska, M.; Urbanowicz, A.; Tworak, A.; Mickiewicz, A.; Figlerowicz, M. The many faces of Dicer: the complexity of the mechanisms regulating Dicer gene expression and enzyme activities. Nucleic Acids Res 2015, 43, 4365–4380. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.C.; Tambe, A.; Kidwell, M.A.; Noland, C.L.; Schneider, C.P.; Doudna, J.A. Dicer-TRBP complex formation ensures accurate mammalian microRNA biogenesis. Mol Cell 2015, 57, 397–407. [Google Scholar] [CrossRef]
- Lee, Y.; Hur, I.; Park, S.Y.; Kim, Y.K.; Suh, M.R.; Kim, V.N. The role of PACT in the RNA silencing pathway. EMBO J 2006, 25, 522–532. [Google Scholar] [CrossRef]
- Lee, H.Y.; Zhou, K.; Smith, A.M.; Noland, C.L.; Doudna, J.A. Differential roles of human Dicer-binding proteins TRBP and PACT in small RNA processing. Nucleic Acids Res 2013, 41, 6568–6576. [Google Scholar] [CrossRef]
- Chendrimada, T.P.; Gregory, R.I.; Kumaraswamy, E.; Norman, J.; Cooch, N.; Nishikura, K.; Shiekhattar, R. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005, 436, 740–744. [Google Scholar] [CrossRef]
- Fukunaga, R.; Han, B.W.; Hung, J.H.; Xu, J.; Weng, Z.; Zamore, P.D. Dicer partner proteins tune the length of mature miRNAs in flies and mammals. Cell 2012, 151, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Doudna, J.A. TRBP alters human precursor microRNA processing in vitro. RNA 2012, 18, 2012–2019. [Google Scholar] [CrossRef]
- Haase, A.D.; Jaskiewicz, L.; Zhang, H.; Laine, S.; Sack, R.; Gatignol, A.; Filipowicz, W. TRBP, a regulator of cellular PKR and HIV-1 virus expression, interacts with Dicer and functions in RNA silencing. EMBO Rep 2005, 6, 961–967. [Google Scholar] [CrossRef]
- Heyam, A.; Lagos, D.; Plevin, M. Dissecting the roles of TRBP and PACT in double-stranded RNA recognition and processing of noncoding RNAs. Wiley Interdiscip Rev RNA 2015, 6, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Rybak, A.; Fuchs, H.; Smirnova, L.; Brandt, C.; Pohl, E.E.; Nitsch, R.; Wulczyn, F.G. A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nat Cell Biol 2008, 10, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Heo, I.; Joo, C.; Cho, J.; Ha, M.; Han, J.; Kim, V.N. Lin28 mediates the terminal uridylation of let-7 precursor MicroRNA. Mol Cell 2008, 32, 276–284. [Google Scholar] [CrossRef]
- Wu, S.L.; Fu, X.; Huang, J.; Jia, T.T.; Zong, F.Y.; Mu, S.R.; Zhu, H.; Yan, Y.; Qiu, S.; Wu, Q.; et al. Genome-wide analysis of YB-1-RNA interactions reveals a novel role of YB-1 in miRNA processing in glioblastoma multiforme. Nucleic Acids Res 2015, 43, 8516–8528. [Google Scholar] [CrossRef]
- Dueck, A.; Ziegler, C.; Eichner, A.; Berezikov, E.; Meister, G. microRNAs associated with the different human Argonaute proteins. Nucleic Acids Res 2012, 40, 9850–9862. [Google Scholar] [CrossRef]
- Nakanishi, K. Anatomy of four human Argonaute proteins. Nucleic Acids Res 2022, 50, 6618–6638. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Carmell, M.A.; Rivas, F.V.; Marsden, C.G.; Thomson, J.M.; Song, J.J.; Hammond, S.M.; Joshua-Tor, L.; Hannon, G.J. Argonaute2 is the catalytic engine of mammalian RNAi. Science 2004, 305, 1437–1441. [Google Scholar] [CrossRef]
- Medley, J.C.; Panzade, G.; Zinovyeva, A.Y. microRNA strand selection: Unwinding the rules. Wiley Interdiscip Rev RNA 2021, 12, e1627. [Google Scholar] [CrossRef]
- Schwarz, D.S.; Hutvagner, G.; Du, T.; Xu, Z.; Aronin, N.; Zamore, P.D. Asymmetry in the assembly of the RNAi enzyme complex. Cell 2003, 115, 199–208. [Google Scholar] [CrossRef]
- Frank, F.; Sonenberg, N.; Nagar, B. Structural basis for 5'-nucleotide base-specific recognition of guide RNA by human AGO2. Nature 2010, 465, 818–822. [Google Scholar] [CrossRef]
- Pinhal, D.; Goncalves, L.B.; Campos, V.F.; Patton, J.G. Decoding microRNA arm switching: a key to evolutionary innovation and gene regulation. Cell Mol Life Sci 2025, 82, 197. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.; Yu, S.; Lee, Y.Y.; Park, J.; Choi, R.J.; Yoon, S.J.; Kang, S.G.; Kim, V.N. A Mechanism for microRNA Arm Switching Regulated by Uridylation. Mol Cell 2020, 78, 1224–1236 e1225. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K. Anatomy of RISC: how do small RNAs and chaperones activate Argonaute proteins? Wiley Interdiscip Rev RNA 2016, 7, 637–660. [Google Scholar] [CrossRef]
- Iwasaki, S.; Kobayashi, M.; Yoda, M.; Sakaguchi, Y.; Katsuma, S.; Suzuki, T.; Tomari, Y. Hsc70/Hsp90 chaperone machinery mediates ATP-dependent RISC loading of small RNA duplexes. Mol Cell 2010, 39, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Ohanian, M.; Humphreys, D.T.; Anderson, E.; Preiss, T.; Fatkin, D. A heterozygous variant in the human cardiac miR-133 gene, MIR133A2, alters miRNA duplex processing and strand abundance. BMC Genet 2013, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Pi, J.; Zou, D.; Wang, X.; Xu, J.; Yu, S.; Zhang, T.; Li, F.; Zhang, X.; Zhao, H.; et al. microRNA arm-imbalance in part from complementary targets mediated decay promotes gastric cancer progression. Nat Commun 2019, 10, 4397. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Tian, S.; Wang, J.; Zhang, F.; Wang, D. Comparative Analysis of microRNA Binding Site Distribution and microRNA-Mediated Gene Expression Repression of Oncogenes and Tumor Suppressor Genes. Genes (Basel) 2022, 13. [Google Scholar] [CrossRef]
- Vidigal, J.A. A catalytic function for mammalian Argonautes. Nat Rev Mol Cell Biol 2024, 25, 339. [Google Scholar] [CrossRef]
- Hutvagner, G.; Zamore, P.D. A microRNA in a multiple-turnover RNAi enzyme complex. Science 2002, 297, 2056–2060. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Becker, W.R.; Ober-Reynolds, B.; Jouravleva, K.; Jolly, S.M.; Zamore, P.D.; Greenleaf, W.J. High-Throughput Analysis Reveals Rules for Target RNA Binding and Cleavage by AGO2. Mol Cell 2019, 75, 741–755 e711. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Wang, P.Y.; Bartel, D.P.; Vos, S.M. The structural basis for RNA slicing by human Argonaute2. Cell Rep 2025, 44, 115166. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.O.; Tomari, Y. Life of RISC: Formation, action, and degradation of RNA-induced silencing complex. Mol Cell 2022, 82, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Fukaya, T.; Iwakawa, H.O.; Tomari, Y. MicroRNAs block assembly of eIF4F translation initiation complex in Drosophila. Mol Cell 2014, 56, 67–78. [Google Scholar] [CrossRef]
- Chen, C.Y.; Zheng, D.; Xia, Z.; Shyu, A.B. Ago-TNRC6 triggers microRNA-mediated decay by promoting two deadenylation steps. Nat Struct Mol Biol 2009, 16, 1160–1166. [Google Scholar] [CrossRef]
- Johnson, S.M.; Grosshans, H.; Shingara, J.; Byrom, M.; Jarvis, R.; Cheng, A.; Labourier, E.; Reinert, K.L.; Brown, D.; Slack, F.J. RAS is regulated by the let-7 microRNA family. Cell 2005, 120, 635–647. [Google Scholar] [CrossRef]
- Kim, H.H.; Kuwano, Y.; Srikantan, S.; Lee, E.K.; Martindale, J.L.; Gorospe, M. HuR recruits let-7/RISC to repress c-Myc expression. Genes Dev 2009, 23, 1743–1748. [Google Scholar] [CrossRef]
- Meng, F.; Henson, R.; Wehbe-Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wu, H.; Wu, F.; Nie, D.; Sheng, S.; Mo, Y.Y. MicroRNA-21 targets tumor suppressor genes in invasion and metastasis. Cell Res 2008, 18, 350–359. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, Y.; Ye, L.; Jiao, W.; Song, H.; Mei, H.; Li, D.; Yang, F.; Li, H.; Huang, K.; et al. miRNA-584-3p inhibits gastric cancer progression by repressing Yin Yang 1- facilitated MMP-14 expression. Sci Rep 2017, 7, 8967. [Google Scholar] [CrossRef]
- Matsui, M.; Chu, Y.; Zhang, H.; Gagnon, K.T.; Shaikh, S.; Kuchimanchi, S.; Manoharan, M.; Corey, D.R.; Janowski, B.A. Promoter RNA links transcriptional regulation of inflammatory pathway genes. Nucleic Acids Res 2013, 41, 10086–10109. [Google Scholar] [CrossRef]
- Xiao, M.; Li, J.; Li, W.; Wang, Y.; Wu, F.; Xi, Y.; Zhang, L.; Ding, C.; Luo, H.; Li, Y.; et al. MicroRNAs activate gene transcription epigenetically as an enhancer trigger. RNA Biol 2017, 14, 1326–1334. [Google Scholar] [CrossRef]
- Chen, D.; Fu, L.Y.; Zhang, Z.; Li, G.; Zhang, H.; Jiang, L.; Harrison, A.P.; Shanahan, H.P.; Klukas, C.; Zhang, H.Y.; et al. Dissecting the chromatin interactome of microRNA genes. Nucleic Acids Res 2014, 42, 3028–3043. [Google Scholar] [CrossRef]
- Chu, Y.; Yokota, S.; Liu, J.; Kilikevicius, A.; Johnson, K.C.; Corey, D.R. Argonaute binding within human nuclear RNA and its impact on alternative splicing. RNA 2021, 27, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Tarallo, R.; Giurato, G.; Bruno, G.; Ravo, M.; Rizzo, F.; Salvati, A.; Ricciardi, L.; Marchese, G.; Cordella, A.; Rocco, T.; et al. The nuclear receptor ERbeta engages AGO2 in regulation of gene transcription, RNA splicing and RISC loading. Genome Biol 2017, 18, 189. [Google Scholar] [CrossRef]
- Ameyar-Zazoua, M.; Rachez, C.; Souidi, M.; Robin, P.; Fritsch, L.; Young, R.; Morozova, N.; Fenouil, R.; Descostes, N.; Andrau, J.C.; et al. Argonaute proteins couple chromatin silencing to alternative splicing. Nat Struct Mol Biol 2012, 19, 998–1004. [Google Scholar] [CrossRef]
- Guidi, R.; Wedeles, C.; Xu, D.; Kolmus, K.; Headland, S.E.; Teng, G.; Guillory, J.; Zeng, Y.J.; Cheung, T.K.; Chaudhuri, S.; et al. Argonaute3-SF3B3 complex controls pre-mRNA splicing to restrain type 2 immunity. Cell Rep 2023, 42, 113515. [Google Scholar] [CrossRef] [PubMed]
- El Fatimy, R.; Zhang, Y.; Deforzh, E.; Ramadas, M.; Saravanan, H.; Wei, Z.; Rabinovsky, R.; Teplyuk, N.M.; Uhlmann, E.J.; Krichevsky, A.M. A nuclear function for an oncogenic microRNA as a modulator of snRNA and splicing. Mol Cancer 2022, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- Bandiera, S.; Ruberg, S.; Girard, M.; Cagnard, N.; Hanein, S.; Chretien, D.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. Nuclear outsourcing of RNA interference components to human mitochondria. PLoS One 2011, 6, e20746. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, C.; Zhou, X.; Han, X.; Li, J.; Wang, Z.; Shang, H.; Liu, Y.; Cao, H. Mitochondria Associated MicroRNA Expression Profiling of Heart Failure. Biomed Res Int 2017, 2017, 4042509. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dai, B.; Fan, J.; Chen, C.; Nie, X.; Yin, Z.; Zhao, Y.; Zhang, X.; Wang, D.W. The Different Roles of miRNA-92a-2-5p and let-7b-5p in Mitochondrial Translation in db/db Mice. Mol Ther Nucleic Acids 2019, 17, 424–435. [Google Scholar] [CrossRef]
- Karginov, F.V.; Hannon, G.J. Remodeling of Ago2-mRNA interactions upon cellular stress reflects miRNA complementarity and correlates with altered translation rates. Genes Dev 2013, 27, 1624–1632. [Google Scholar] [CrossRef]
- Sheu-Gruttadauria, J.; MacRae, I.J. Phase Transitions in the Assembly and Function of Human miRISC. Cell 2018, 173, 946–957 e916. [Google Scholar] [CrossRef]
- Zhou, Y.; Panhale, A.; Shvedunova, M.; Balan, M.; Gomez-Auli, A.; Holz, H.; Seyfferth, J.; Helmstadter, M.; Kayser, S.; Zhao, Y.; et al. RNA damage compartmentalization by DHX9 stress granules. Cell 2024, 187, 1701–1718 e1728. [Google Scholar] [CrossRef]
- Cai, C.; Yu, J.; Zhang, X.; Zhou, T.; Chen, Q. A model for propagation of RNA structural memory through biomolecular condensates. Nat Cell Biol 2025. [Google Scholar] [CrossRef]
- Duan, Y.; Li, L.; Panzade, G.P.; Piton, A.; Zinovyeva, A.; Ambros, V. Modeling neurodevelopmental disorder-associated human AGO1 mutations in Caenorhabditis elegans Argonaute alg-1. Proc Natl Acad Sci U S A 2024, 121, e2308255121. [Google Scholar] [CrossRef]
- Schalk, A.; Cousin, M.A.; Dsouza, N.R.; Challman, T.D.; Wain, K.E.; Powis, Z.; Minks, K.; Trimouille, A.; Lasseaux, E.; Lacombe, D.; et al. De novo coding variants in the AGO1 gene cause a neurodevelopmental disorder with intellectual disability. J Med Genet 2022, 59, 965–975. [Google Scholar] [CrossRef]
- Lessel, D.; Zeitler, D.M.; Reijnders, M.R.F.; Kazantsev, A.; Hassani Nia, F.; Bartholomaus, A.; Martens, V.; Bruckmann, A.; Graus, V.; McConkie-Rosell, A.; et al. Germline AGO2 mutations impair RNA interference and human neurological development. Nat Commun 2020, 11, 5797. [Google Scholar] [CrossRef]
- Fawzy, M.S.; Toraih, E.A.; Alelwani, W.; Kattan, S.W.; Alnajeebi, A.M.; Hassan, R. The prognostic value of microRNA-biogenesis genes Argonaute 1 and 2 variants in breast cancer patients. Am J Transl Res 2020, 12, 1994–2006. [Google Scholar]
- Nowak, I.; Sarshad, A.A. Argonaute Proteins Take Center Stage in Cancers. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Golden, R.J.; Chen, B.; Li, T.; Braun, J.; Manjunath, H.; Chen, X.; Wu, J.; Schmid, V.; Chang, T.C.; Kopp, F.; et al. An Argonaute phosphorylation cycle promotes microRNA-mediated silencing. Nature 2017, 542, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Paradis-Isler, N.; Boehm, J. NMDA receptor-dependent dephosphorylation of serine 387 in Argonaute 2 increases its degradation and affects dendritic spine density and maturation. J Biol Chem 2018, 293, 9311–9325. [Google Scholar] [CrossRef]
- Kingston, E.R.; Bartel, D.P. Global analyses of the dynamics of mammalian microRNA metabolism. Genome Res 2019, 29, 1777–1790. [Google Scholar] [CrossRef] [PubMed]
- Zangari, J.; Ilie, M.; Rouaud, F.; Signetti, L.; Ohanna, M.; Didier, R.; Romeo, B.; Goldoni, D.; Nottet, N.; Staedel, C.; et al. Rapid decay of engulfed extracellular miRNA by XRN1 exonuclease promotes transient epithelial-mesenchymal transition. Nucleic Acids Res 2017, 45, 4131–4141. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, Y.; Chen, E.; Li, X.; Lv, B.; Vikis, H.G.; Liu, P. XRN2 promotes EMT and metastasis through regulating maturation of miR-10a. Oncogene 2017, 36, 3925–3933. [Google Scholar] [CrossRef]
- Das, S.K.; Sokhi, U.K.; Bhutia, S.K.; Azab, B.; Su, Z.Z.; Sarkar, D.; Fisher, P.B. Human polynucleotide phosphorylase selectively and preferentially degrades microRNA-221 in human melanoma cells. Proc Natl Acad Sci U S A 2010, 107, 11948–11953. [Google Scholar] [CrossRef]
- Thomas, M.F.; Abdul-Wajid, S.; Panduro, M.; Babiarz, J.E.; Rajaram, M.; Woodruff, P.; Lanier, L.L.; Heissmeyer, V.; Ansel, K.M. Eri1 regulates microRNA homeostasis and mouse lymphocyte development and antiviral function. Blood 2012, 120, 130–142. [Google Scholar] [CrossRef]
- Elbarbary, R.A.; Miyoshi, K.; Myers, J.R.; Du, P.; Ashton, J.M.; Tian, B.; Maquat, L.E. Tudor-SN-mediated endonucleolytic decay of human cell microRNAs promotes G(1)/S phase transition. Science 2017, 356, 859–862. [Google Scholar] [CrossRef]
- Reichholf, B.; Herzog, V.A.; Fasching, N.; Manzenreither, R.A.; Sowemimo, I.; Ameres, S.L. Time-Resolved Small RNA Sequencing Unravels the Molecular Principles of MicroRNA Homeostasis. Mol Cell 2019, 75, 756–768 e757. [Google Scholar] [CrossRef]
- Yuan, Y.R.; Pei, Y.; Ma, J.B.; Kuryavyi, V.; Zhadina, M.; Meister, G.; Chen, H.Y.; Dauter, Z.; Tuschl, T.; Patel, D.J. Crystal structure of A. aeolicus argonaute, a site-specific DNA-guided endoribonuclease, provides insights into RISC-mediated mRNA cleavage. Mol Cell 2005, 19, 405–419. [Google Scholar] [CrossRef]
- Ameres, S.L.; Horwich, M.D.; Hung, J.H.; Xu, J.; Ghildiyal, M.; Weng, Z.; Zamore, P.D. Target RNA-directed trimming and tailing of small silencing RNAs. Science 2010, 328, 1534–1539. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Shao, T.J.; Bofill-De Ros, X.; Lian, C.; Villanueva, P.; Dai, L.; Gu, S. AGO-bound mature miRNAs are oligouridylated by TUTs and subsequently degraded by DIS3L2. Nat Commun 2020, 11, 2765. [Google Scholar] [CrossRef]
- Cazalla, D.; Yario, T.; Steitz, J.A. Down-regulation of a host microRNA by a Herpesvirus saimiri noncoding RNA. Science 2010, 328, 1563–1566. [Google Scholar] [CrossRef]
- Sheu-Gruttadauria, J.; Pawlica, P.; Klum, S.M.; Wang, S.; Yario, T.A.; Schirle Oakdale, N.T.; Steitz, J.A.; MacRae, I.J. Structural Basis for Target-Directed MicroRNA Degradation. Mol Cell 2019, 75, 1243–1255 e1247. [Google Scholar] [CrossRef] [PubMed]
- Kleaveland, B. SnapShot: Target-directed miRNA degradation. Cell 2023, 186, 5674–5674 e5671. [Google Scholar] [CrossRef]
- Shi, C.Y.; Kingston, E.R.; Kleaveland, B.; Lin, D.H.; Stubna, M.W.; Bartel, D.P. The ZSWIM8 ubiquitin ligase mediates target-directed microRNA degradation. Science 2020, 370. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; LaVigne, C.A.; Jones, B.T.; Zhang, H.; Gillett, F.; Mendell, J.T. A ubiquitin ligase mediates target-directed microRNA decay independently of tailing and trimming. Science 2020, 370. [Google Scholar] [CrossRef]
- Kleaveland, B.; Shi, C.Y.; Stefano, J.; Bartel, D.P. A Network of Noncoding Regulatory RNAs Acts in the Mammalian Brain. Cell 2018, 174, 350–362 e317. [Google Scholar] [CrossRef]
- Ghini, F.; Rubolino, C.; Climent, M.; Simeone, I.; Marzi, M.J.; Nicassio, F. Endogenous transcripts control miRNA levels and activity in mammalian cells by target-directed miRNA degradation. Nat Commun 2018, 9, 3119. [Google Scholar] [CrossRef]
- Li, L.; Sheng, P.; Li, T.; Fields, C.J.; Hiers, N.M.; Wang, Y.; Li, J.; Guardia, C.M.; Licht, J.D.; Xie, M. Widespread microRNA degradation elements in target mRNAs can assist the encoded proteins. Genes Dev 2021, 35, 1595–1609. [Google Scholar] [CrossRef]
- Shi, C.Y.; Elcavage, L.E.; Chivukula, R.R.; Stefano, J.; Kleaveland, B.; Bartel, D.P. ZSWIM8 destabilizes many murine microRNAs and is required for proper embryonic growth and development. Genome Res 2023, 33, 1482–1496. [Google Scholar] [CrossRef]
- Jones, B.T.; Han, J.; Zhang, H.; Hammer, R.E.; Evers, B.M.; Rakheja, D.; Acharya, A.; Mendell, J.T. Target-directed microRNA degradation regulates developmental microRNA expression and embryonic growth in mammals. Genes Dev 2023, 37, 661–674. [Google Scholar] [CrossRef]
- Kingston, E.R.; Blodgett, L.W.; Bartel, D.P. Endogenous transcripts direct microRNA degradation in Drosophila, and this targeted degradation is required for proper embryonic development. Mol Cell 2022, 82, 3872–3884 e3879. [Google Scholar] [CrossRef]
- Kawahara, Y.; Megraw, M.; Kreider, E.; Iizasa, H.; Valente, L.; Hatzigeorgiou, A.G.; Nishikura, K. Frequency and fate of microRNA editing in human brain. Nucleic Acids Res 2008, 36, 5270–5280. [Google Scholar] [CrossRef]
- Alarcon, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Cheray, M.; Etcheverry, A.; Jacques, C.; Pacaud, R.; Bougras-Cartron, G.; Aubry, M.; Denoual, F.; Peterlongo, P.; Nadaradjane, A.; Briand, J.; et al. Cytosine methylation of mature microRNAs inhibits their functions and is associated with poor prognosis in glioblastoma multiforme. Mol Cancer 2020, 19, 36. [Google Scholar] [CrossRef] [PubMed]
- Pandolfini, L.; Barbieri, I.; Bannister, A.J.; Hendrick, A.; Andrews, B.; Webster, N.; Murat, P.; Mach, P.; Brandi, R.; Robson, S.C.; et al. METTL1 Promotes let-7 MicroRNA Processing via m7G Methylation. Mol Cell 2019, 74, 1278–1290 e1279. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Jiao, Z.; Rong, W.; Qu, S.; Liao, Z.; Sun, X.; Wei, Y.; Zhao, Q.; Wang, J.; Liu, Y.; et al. 3'-Terminal 2'-O-methylation of lung cancer miR-21-5p enhances its stability and association with Argonaute 2. Nucleic Acids Res 2020, 48, 7027–7040. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lu, R.; Huang, J.; Li, L.; Cao, Y.; Huang, C.; Chen, R.; Wang, Y.; Huang, J.; Zhao, X.; et al. N4-acetylcytidine modifies primary microRNAs for processing in cancer cells. Cell Mol Life Sci 2024, 81, 73. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Gao, J.; Ding, S.L.; Wang, K.; Jiao, J.Q.; Wang, Y.; Sun, T.; Zhou, L.Y.; Long, B.; Zhang, X.J.; et al. Oxidative Modification of miR-184 Enables It to Target Bcl-xL and Bcl-w. Mol Cell 2015, 59, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Xhemalce, B.; Robson, S.C.; Kouzarides, T. Human RNA methyltransferase BCDIN3D regulates microRNA processing. Cell 2012, 151, 278–288. [Google Scholar] [CrossRef]
- Yuan, X.; Su, Y.; Johnson, B.; Kirchner, M.; Zhang, X.; Xu, S.; Jiang, S.; Wu, J.; Shi, S.; Russo, J.J.; et al. Mass Spectrometry-Based Direct Sequencing of tRNAs De Novo and Quantitative Mapping of Multiple RNA Modifications. J Am Chem Soc 2024, 146, 25600–25613. [Google Scholar] [CrossRef]
- Zhang, X.; Cozen, A.E.; Liu, Y.; Chen, Q.; Lowe, T.M. Small RNA Modifications: Integral to Function and Disease. Trends Mol Med 2016, 22, 1025–1034. [Google Scholar] [CrossRef]
- Blow, M.J.; Grocock, R.J.; van Dongen, S.; Enright, A.J.; Dicks, E.; Futreal, P.A.; Wooster, R.; Stratton, M.R. RNA editing of human microRNAs. Genome Biol 2006, 7, R27. [Google Scholar] [CrossRef]
- Yang, W.; Chendrimada, T.P.; Wang, Q.; Higuchi, M.; Seeburg, P.H.; Shiekhattar, R.; Nishikura, K. Modulation of microRNA processing and expression through RNA editing by ADAR deaminases. Nat Struct Mol Biol 2006, 13, 13–21. [Google Scholar] [CrossRef]
- Kawahara, Y.; Zinshteyn, B.; Chendrimada, T.P.; Shiekhattar, R.; Nishikura, K. RNA editing of the microRNA-151 precursor blocks cleavage by the Dicer-TRBP complex. EMBO Rep 2007, 8, 763–769. [Google Scholar] [CrossRef]
- Ota, H.; Sakurai, M.; Gupta, R.; Valente, L.; Wulff, B.E.; Ariyoshi, K.; Iizasa, H.; Davuluri, R.V.; Nishikura, K. ADAR1 forms a complex with Dicer to promote microRNA processing and RNA-induced gene silencing. Cell 2013, 153, 575–589. [Google Scholar] [CrossRef]
- Park, Y.M.; Hwang, S.J.; Masuda, K.; Choi, K.M.; Jeong, M.R.; Nam, D.H.; Gorospe, M.; Kim, H.H. Heterogeneous nuclear ribonucleoprotein C1/C2 controls the metastatic potential of glioblastoma by regulating PDCD4. Mol Cell Biol 2012, 32, 4237–4244. [Google Scholar] [CrossRef]
- Alarcon, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef]
- Heo, I.; Ha, M.; Lim, J.; Yoon, M.J.; Park, J.E.; Kwon, S.C.; Chang, H.; Kim, V.N. Mono-uridylation of pre-microRNA as a key step in the biogenesis of group II let-7 microRNAs. Cell 2012, 151, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Salzman, D.W.; Nakamura, K.; Nallur, S.; Dookwah, M.T.; Metheetrairut, C.; Slack, F.J.; Weidhaas, J.B. miR-34 activity is modulated through 5'-end phosphorylation in response to DNA damage. Nat Commun 2016, 7, 10954. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Yu, S.; Jeong, K.J.; Zhou, Z.; Han, L.; Tsang, Y.H.; Li, J.; Chen, H.; Mangala, L.S.; et al. Systematic characterization of A-to-I RNA editing hotspots in microRNAs across human cancers. Genome Res 2017, 27, 1112–1125. [Google Scholar] [CrossRef]
- Paul, D.; Sinha, A.N.; Ray, A.; Lal, M.; Nayak, S.; Sharma, A.; Mehani, B.; Mukherjee, D.; Laddha, S.V.; Suri, A.; et al. A-to-I editing in human miRNAs is enriched in seed sequence, influenced by sequence contexts and significantly hypoedited in glioblastoma multiforme. Sci Rep 2017, 7, 2466. [Google Scholar] [CrossRef]
- Choudhury, Y.; Tay, F.C.; Lam, D.H.; Sandanaraj, E.; Tang, C.; Ang, B.T.; Wang, S. Attenuated adenosine-to-inosine editing of microRNA-376a* promotes invasiveness of glioblastoma cells. J Clin Invest 2012, 122, 4059–4076. [Google Scholar] [CrossRef] [PubMed]
- Heo, I.; Joo, C.; Kim, Y.K.; Ha, M.; Yoon, M.J.; Cho, J.; Yeom, K.H.; Han, J.; Kim, V.N. TUT4 in concert with Lin28 suppresses microRNA biogenesis through pre-microRNA uridylation. Cell 2009, 138, 696–708. [Google Scholar] [CrossRef]
- Thornton, J.E.; Chang, H.M.; Piskounova, E.; Gregory, R.I. Lin28-mediated control of let-7 microRNA expression by alternative TUTases Zcchc11 (TUT4) and Zcchc6 (TUT7). RNA 2012, 18, 1875–1885. [Google Scholar] [CrossRef]
- Seok, H.; Lee, H.; Lee, S.; Ahn, S.H.; Lee, H.S.; Kim, G.D.; Peak, J.; Park, J.; Cho, Y.K.; Jeong, Y.; et al. Position-specific oxidation of miR-1 encodes cardiac hypertrophy. Nature 2020, 584, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.; Peak, J.; Park, J.; Ahn, S.H.; Cho, Y.K.; Jeong, Y.; Lee, H.S.; Lee, J.; Ignatova, E.; Lee, S.E.; et al. Widespread 8-oxoguanine modifications of miRNA seeds differentially regulate redox-dependent cancer development. Nat Cell Biol 2023, 25, 1369–1383. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, Y.; Zhou, T.; Chen, Q. tsRNAs: The Swiss Army Knife for Translational Regulation. Trends Biochem Sci 2019, 44, 185–189. [Google Scholar] [CrossRef]
- Shi, J.; Zhou, T.; Chen, Q. Exploring the expanding universe of small RNAs. Nat Cell Biol 2022, 24, 415–423. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, Y.; Li, Y.; Zhang, L.; Zhang, X.; Yan, M.; Chen, Q.; Zhang, Y. Optimized identification and characterization of small RNAs with PANDORA-seq. Nat Protoc 2025. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, Y.; Tan, D.; Zhang, X.; Yan, M.; Zhang, Y.; Franklin, R.; Shahbazi, M.; Mackinlay, K.; Liu, S.; et al. PANDORA-seq expands the repertoire of regulatory small RNAs by overcoming RNA modifications. Nat Cell Biol 2021, 23, 424–436. [Google Scholar] [CrossRef]
- Hernandez, R.; Shi, J.; Liu, J.; Li, X.; Wu, J.; Zhao, L.; Zhou, T.; Chen, Q.; Zhou, C. PANDORA-Seq unveils the hidden small noncoding RNA landscape in atherosclerosis of LDL receptor-deficient mice. J Lipid Res 2023, 64, 100352. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.; Feng, N.; Zhai, Q. Discovery of the major 15-30 nt mammalian small RNAs, their biogenesis and function. Nat Commun 2023, 14, 5796. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Z.; O'Loughlin, E.; Lee, T.; Houel, S.; O'Carroll, D.; Tarakhovsky, A.; Ahn, N.G.; Yi, R. Quantitative functions of Argonaute proteins in mammalian development. Genes Dev 2012, 26, 693–704. [Google Scholar] [CrossRef]
- Bissels, U.; Wild, S.; Tomiuk, S.; Holste, A.; Hafner, M.; Tuschl, T.; Bosio, A. Absolute quantification of microRNAs by using a universal reference. RNA 2009, 15, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Shi, J.; Zhang, Y.; Zhang, H.; Liao, S.; Li, W.; Lei, L.; Han, C.; Ning, L.; Cao, Y.; et al. A novel class of tRNA-derived small RNAs extremely enriched in mature mouse sperm. Cell Res 2012, 22, 1609–1612. [Google Scholar] [CrossRef]
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.H.; Peng, H.; Zhang, X.; Zhang, Y.; et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef]
- Chen, Q.; Yan, W.; Duan, E. Epigenetic inheritance of acquired traits through sperm RNAs and sperm RNA modifications. Nat Rev Genet 2016, 17, 733–743. [Google Scholar] [CrossRef]
- Tomar, A.; Gomez-Velazquez, M.; Gerlini, R.; Comas-Armangue, G.; Makharadze, L.; Kolbe, T.; Boersma, A.; Dahlhoff, M.; Burgstaller, J.P.; Lassi, M.; et al. Epigenetic inheritance of diet-induced and sperm-borne mitochondrial RNAs. Nature 2024, 630, 720–727. [Google Scholar] [CrossRef]
- Cai, C.; Chen, Q. Father's diet influences son's metabolic health through sperm RNA. Nature 2024, 630, 571–573. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Shi, J.; Tuorto, F.; Li, X.; Liu, Y.; Liebers, R.; Zhang, L.; Qu, Y.; Qian, J.; et al. Dnmt2 mediates intergenerational transmission of paternally acquired metabolic disorders through sperm small non-coding RNAs. Nat Cell Biol 2018, 20, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, X.; Shi, J.; Yan, M.; Zhou, T. Origins and evolving functionalities of tRNA-derived small RNAs. Trends Biochem Sci 2021, 46, 790–804. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Shi, J.; Liu, H.; Zhang, X.; Zhou, J.J.; Li, M.; Zhou, D.; Li, R.; Lv, J.; Wen, G.; et al. Peripheral blood non-canonical small non-coding RNAs as novel biomarkers in lung cancer. Mol Cancer 2020, 19, 159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Trebak, F.; Souza, L.A.C.; Shi, J.; Zhou, T.; Kehoe, P.G.; Chen, Q.; Feng Earley, Y. Small RNA modifications in Alzheimer's disease. Neurobiol Dis 2020, 145, 105058. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Shi, J.; Zhang, H.; Cao, Z.; Gao, X.; Ren, W.; Ning, Y.; Ning, L.; Cao, Y.; et al. Identification and characterization of an ancient class of small RNAs enriched in serum associating with active infection. J Mol Cell Biol 2014, 6, 172–174. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, J.; Chen, Q. tsRNAs: new players in mammalian retrotransposon control. Cell Res 2017, 27, 1307–1308. [Google Scholar] [CrossRef]
- Eiring, A.M.; Harb, J.G.; Neviani, P.; Garton, C.; Oaks, J.J.; Spizzo, R.; Liu, S.; Schwind, S.; Santhanam, R.; Hickey, C.J.; et al. miR-328 functions as an RNA decoy to modulate hnRNP E2 regulation of mRNA translation in leukemic blasts. Cell 2010, 140, 652–665. [Google Scholar] [CrossRef]
- Lehmann, S.M.; Kruger, C.; Park, B.; Derkow, K.; Rosenberger, K.; Baumgart, J.; Trimbuch, T.; Eom, G.; Hinz, M.; Kaul, D.; et al. An unconventional role for miRNA: let-7 activates Toll-like receptor 7 and causes neurodegeneration. Nat Neurosci 2012, 15, 827–835. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, H.; Jin, F.; Yan, X.; Xu, G.; Hu, H.; Liang, G.; Zhan, S.; Hu, X.; Zhao, Q.; et al. Injured liver-released miRNA-122 elicits acute pulmonary inflammation via activating alveolar macrophage TLR7 signaling pathway. Proc Natl Acad Sci U S A 2019, 116, 6162–6171. [Google Scholar] [CrossRef]
- Park, C.K.; Xu, Z.Z.; Berta, T.; Han, Q.; Chen, G.; Liu, X.J.; Ji, R.R. Extracellular microRNAs activate nociceptor neurons to elicit pain via TLR7 and TRPA1. Neuron 2014, 82, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, X.; Cai, C.; Zhou, T.; Chen, Q. Small RNA and Toll-like receptor interactions: origins and disease mechanisms. Trends Biochem Sci 2025, 50, 385–401. [Google Scholar] [CrossRef]
- Han, Q.; Liu, D.; Convertino, M.; Wang, Z.; Jiang, C.; Kim, Y.H.; Luo, X.; Zhang, X.; Nackley, A.; Dokholyan, N.V.; et al. miRNA-711 Binds and Activates TRPA1 Extracellularly to Evoke Acute and Chronic Pruritus. Neuron 2018, 99, 449–463 e446. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wan, X.; Dennis, A.T.; Bektik, E.; Wang, Z.; Costa, M.G.S.; Fagnen, C.; Venien-Bryan, C.; Xu, X.; Gratz, D.H.; et al. MicroRNA Biophysically Modulates Cardiac Action Potential by Direct Binding to Ion Channel. Circulation 2021, 143, 1597–1613. [Google Scholar] [CrossRef]
- Seyhan, A.A. Trials and Tribulations of MicroRNA Therapeutics. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- What will it take to get miRNA therapies to market? Nat Biotechnol 2024, 42, 1623–1624. [CrossRef]
- Segal, M.; Slack, F.J. Challenges identifying efficacious miRNA therapeutics for cancer. Expert Opin Drug Discov 2020, 15, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhu, L.; Wang, X.; Jin, H. RNA-based therapeutics: an overview and prospectus. Cell Death Dis 2022, 13, 644. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, J.; Canfield, J.; Mong, E.F.; VanWye, J.; Totary-Jain, H. Nucleotide Modification Alters MicroRNA-Dependent Silencing of MicroRNA Switches. Mol Ther Nucleic Acids 2019, 14, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Groot, M.; Lee, H. Sorting Mechanisms for MicroRNAs into Extracellular Vesicles and Their Associated Diseases. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Wartmann, T.; Accuffi, S.; Al-Madhi, S.; Perrakis, A.; Kahlert, C.; Link, A.; Venerito, M.; Keitel-Anselmino, V.; Bruns, C.; et al. Integrating a microRNA signature as a liquid biopsy-based tool for the early diagnosis and prediction of potential therapeutic targets in pancreatic cancer. Br J Cancer 2024, 130, 125–134. [Google Scholar] [CrossRef]
- Metcalf, G.A.D. MicroRNAs: circulating biomarkers for the early detection of imperceptible cancers via biosensor and machine-learning advances. Oncogene 2024, 43, 2135–2142. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



