Submitted:
01 September 2025
Posted:
02 September 2025
You are already at the latest version
Abstract
Background: Pediatric thrombosis (PT) represents a rare condition that can manifest from neonatal life to adolescence, encompassing life-threatening complications. Its pathogenesis is attributed to immature hemostasis in conjunction with environmental and genetic factors, predominantly including those resulting in increased levels of plasminogen activator inhibitor 1 (PAI-1), the principal inhibitor of fibrinolysis, subject to upstream regulation by ACE (angiotensin-converting enzyme). Although the implication of microRNAs (miRNAs), epigenetic modulators of gene-expression, is demonstrated in adult-thrombosis, evidence is lacking in the pediatric setting. Here, we investigated the involvement of two miRNA-regulators of PAI-1 in PT, in relation to clinical and genetic parameters that induce PAI-1 fluctuations. Methods: Following bioinformatic target-prediction, miRNA expression was assessed by quantitative real-time PCR, in serum-samples of 19 pediatric patients with thrombosis (1-18 months post-incident), and 19 controls. Patients were genotyped for the PAI-1-4G/5G and ACE-I/D polymorphisms by PCR-based assays. Genotypic and thrombosis-related clinical data were analyzed in relation to miRNA-expression. Results: Two miRNAs (miR-145-5p, miR-34a-5p) were identified to target PAI-1 mRNA, with miR-34a additionally targeting ACE mRNA. The expression of miR-34a was significantly decreased in patients compared to controls (p=0.029), while no difference was observed for miR-145. Within patients, miR-34a demonstrated a peak 1-3 months post-thrombosis and was diminished upon treatment-completion (p=0.031), followed by a slight long-term increase. MiR-34a expression differed significantly by thrombosis site (p=0.019), while a significant synergistic interaction between site and onset-type (provoked/unprovoked) was detected (p=0.016). Finally, an epistatic modification was observed in cerebral cases, since double homozygosity (4G/4G+D/D) led to miR-34a diminishing, with DD carriership reversing the 4G4G-induced upregulation of miR-34a (p=0.006). Conclusions: Our findings suggest that in pediatric thrombosis, downregulation of miR-34a is indicative of impaired fibrinolytic capacity, attributed to deficient regulation of the inhibitory ACE/PAI-1 axis.
Keywords:
thrombosis in childhood
; adolescent thrombosis
; fibrinolysis
; coagulation
; thrombophilia
; stroke
; microRNA
; miR-34a-5p
; miR-145-5p
; epigenetics
; SERPINE1 4G/5G polymorphism
; ACE I/D polymorphism
1. Introduction
Thrombosis is characterized by abnormal clot formation within a blood vessel, obstructing blood flow and resulting from substantial imbalance between coagulation and fibrinolysis. The developed thrombotic complexes mostly comprise of red blood cells, fibrin, as well as platelets and leukocytes [1,2]. Although conditions such as an ischemic stroke (IS) or pulmonary embolism (PE) caused by thrombus development, are typically seen in adults, thrombosis is also a serious health complication of pediatric populations and can be classified as venous or arterial, depending on the affected vessels, with venous thromboembolism (VTE) scoring the highest rates [3,4,5].
While pediatric thrombosis is considered rare, corresponding to up to 14/10,000 annual admissions, its incidence rates have demonstrated a significant increase by 70%, with a 7-fold increase only corresponding to strokes affecting newborns and children, with newborns significantly higher thrombotic rates, due to unstable hemostasis [1,4,5,6,7]. Thrombotic incidents in pediatric patients most frequently present as cerebral thrombosis, followed by limb incidents and entail life-threatening complications, such as pulmonary embolism and post-thrombotic syndrome [4,8,9], while early detection and thorough anticoagulant/thrombolytic treatment and subsequent chemoprophylaxis can be determinant to patients’ survival, complication occurrence and overall therapeutic outcomes, while thorough and multifactorial evaluation of a child’s risk is crucial for their clinical management as to prevention of secondary thrombosis, post-thrombotic syndrome, and overall recurrence [4,6,8,10].
Although pediatric thrombosis is multifactorial and its manifestation is affected by possible risk factors such as hypoxia, inflammation-involving conditions, malignancy and injury, it demonstrates a strong and well-established genetic background that increases a child’s vulnerability to thrombi formation [9,11,12,13]. Inherited thrombophilia is considered a solid risk factor in pediatric thrombosis and includes the presence of genetic variants causing functional or quantitative imbalances in factors involved in hemostasis and fibrinolysis, such as factor V (FV) Leiden, prothrombin (PTH), methylenetetrahydrofolate reductase (MTHFR), plasminogen activator inhibitor-1 (PAI-1) and angiotensin-converting enzyme (ACE) [11,14]. Amongst the above, PAI-1 has recently been reported as the predominant inherited genetic risk-factor for pediatric thrombosis [9].
PAI-1 (or SERPINE1) is the most robust inhibitor of fibrinolysis, a process that acts at the final hemostatic stage by dissolving developed thrombi, through the enzymatic activity of fibrin, thus restoring normal blood circulation [13,15,16,17]. PAI-1 is predominantly secreted by endothelial cells and negatively regulates of the fibrinolytic system by covalent binding to tissue plasminogen activator (tPA), obstructing the conversion of plasminogen to plasmin, which ultimately catalyzes the degradation of fibrin, the main stabilizer of blood clots [2,13,18,19]. The ratio of tPA/PAI-1 is unfavorable for effective fibrinolysis during childhood, since tPA levels are significantly reduced at birth and until late adolescence, compared to the increased levels of its primary inhibitor PAI-1 [19].
It has been demonstrated that increased PAI-1 levels ultimately lead to diminished fibrinolytic activity and a hypercoagulable state, therefore skyrocketing the risk of both venous and arterial thrombosis, while the generation of thrombin itself acts as an inducer of additional PAI-1 secretion [13,15,16,17,20]. It is experimentally supported that upregulated PAI-1 levels not only promote thrombotic development, but may also contribute to resistance against thrombolytics [19,21]. SERPINE1 expression has been demonstrated to increase prior to thrombus development, while maximum PAI-1 levels are reached during the acute phase of thrombosis and finally decrease post treatment, which includes tPA in life-threatening incidents [5,15,22,23]. However, following a thrombotic incident PAI-1 has been shown to remain increased for prolonged periods of time, with a minimum of three months, contributing to proportional PAI-1/t-PA abnormalities and therefore decreased fibrinolytic capacity in up to 54% patients [24].
PAI-1 levels are highly influenced by the 4G/5G (rs1799768) 1bp insertion/deletion functional polymorphism, in the promoter region of the SERPINE1 coding gene [15,25,26]. The presence of the 4G polymorphic allele in either homozygous or heterozygous state is strongly associated with higher levels of circulating PAI-1, resulting in hypofibrinolysis and coagulation promotion, while the normal 5G normal allele has been shown to exert a protective effect against thrombotic recurrence [25,26]. The PAI-1 4G/5G polymorphism is reported to be the most frequently detected genetic variant within children that have undergone thrombosis, compared to other predisposing factors [9]. Moreover, PAI-1 levels are also regulated by the renin-angiotensin system (RAS), which indirectly suppresses fibrinolysis via angiotensin II (angII) that triggers a significant PAI-1 increase [27,28,29]. The angiotensin-converting enzyme (ACE), primarily responsible for direct angII formation, serves as the bridge between the two systems, thus acting as an indirect regulator of PAI-1. The insertion/deletion (I/D) functional polymorphism (rs1799752) of the ACE gene is widely known for increasing circulating ACE levels in presence of the D (deletion) allele (II<ID<DD), with the DD genotype associated with maximum concentrations and enzymatic activity for angII formation [30,31]. The presence of the D allele and primarily DD homozygosity has also been demonstrated to indirectly increase PAI-1 levels, resulting in fibrinolytic impairment [32]
In addition to key proteinic regulators of thrombosis, such as PAI-1, evidence has been mounting around the implication of microRNAs (miRNAs) in the delicate regulatory interplays of adult thrombosis[33]. MiRNAs are a class of single-stranded, small (18-25nt), non-coding RNA molecules that act as profound negative regulators of gene expression at a post-transcriptional level, through the translational inhibition of mRNA targets. Each miRNA retains the capacity to regulate the expression of multiple gene-targets at once, through the binding of its seed sequence to recognized complimentary binding sites within the 3′ untranslated region (3′-UTR) of their mRNAs [33,34,35] . Even after their extracellular release, miRNAs remain remarkably stable, due to their binding with Argonaute proteins and/or their encapsulation in extracellular vesicles (EVs), which provide protection against RNase-mediated degradation [36]. Hence, and given that their expressional fluctuations (up- or down-regulation) can be highly reflective of a pathology, they hold great promise both as biomarkers, but also as crucial elements of disease pathogenicity by the abnormal regulation of the implicated genes [34,37].
In the present study, we have examined the expression levels of two miRNAs regulators of PAI-1 (miR-145-5p and miR-34a-5p) in pediatric patients with thrombosis, 1-18 months post-incident, compared to age- and gender-matched healthy controls. Expressional patterns of the studied miRNAs were furtherly evaluated within distinct subpopulations of the patient cohort, in relation to factors such as post-thrombotic time, PAI-1 4G/5G genotypes and treatment status, which have been demonstrated to induce anticipated changes in PAI-1 levels.
This study in pediatric patients was designed under the guiding hypothesis of a possible regulatory interplay between miR-145 and miR-34a expression and PAI-1 fluctuations, which are documented in the literature for adults [5,15,22,23], across different stages of post-thrombotic course, and treatment, as well as different genotypes of the 4G/5G functional polymorphism. The present research is, to our knowledge, the first globally to investigate miRNA expression profiles in the context of pediatric thrombosis.
2. Materials and Methods
2.1. Sample Collection
This study was bioethically approved by the appropriate Committee of the “Aghia Sophia” Children’s University Hospital (protocol 21330/2020). Whole blood samples of gender and age matched patients and controls were collected at the Haemophilia Center for Children and Adolescents/Haemostasis and Thrombosis Unit of “Aghia Sophia” Children’s University Hospital. Patients were recruited during post-thrombosis routine follow-up visits. After thorough informative discussion, the parents of the enrolled participants provided written consent for their offspring’s blood samples to be collected and processed for subsequent molecular analyses, as well as for selected clinical data to be co-evaluated. In addition, a thorough four-generation family history of the patients was obtained. Blood samples that were intended for serum separation were drawn directly into collection tubes containing clot-activator, while samples intended for genomic analysis were collected in EDTA-containing tubes.
2.2. Serum Separation and RNA Extraction
Whole blood-samples with clot-activator remained in room temperature for 15-30 minutes, to allow clotting to take place. The samples were followingly centrifuged at 3,000 rpm, in 4°C for 10 minutes, using a swing-bucket rotor centrifuge. Supernatant serum phase was carefully collected and was centrifuged again for additional sample-purity.
RNA extraction was performed using the “NucleoSpin miRNA Plasma, Mini kit for circulating miRNA” (MACHEREY-NAGEL, Germany), according to the manufacturer’s protocol for serum-samples and RNA elution in the minimum recommended volume, in order to obtain maximum miRNA concentrations. The concentration (ng/μL) and purity of the eluted RNA were assessed using a BioSpec-nano Spectrophotometer for Life Science (Shimadzu Corporation, Japan). RNA was reverse-transcribed into cDNA directly after extraction, quantificaction and normalization across sample-concentrations to ensure template comparability.
2.3. Bioinformatic Target-Prediction Analyses
Target prediction and retrieval of experimentally-validated miRNA/mRNA interactions were combinatorially performed via the utilization of TargetScanHuman, miRNet 2.0, as well as TarBase.v.9 (incorporating DIANA-microT-CDS) computational algorithms.
2.4. Reverse Transcription and Quantitative Real-Time PCR
Reverse Transcription (RT) of miRNAs was performed using the miRCURY LNA RT Kit (Qiagen, Germany) for miRNA polyadenylation and first-strand cDNA synthesis in a single-tube reaction of 10μL (2μL 5xRT SYBR-Green Reaction Buffer, 2μL RNA template, 1μL 10xRT Enzyme-Mix and 5μL RNase-free water). The RT reaction was performed using a Gradient Thermal Cycler (Takara Bio, Japan) and involved incubation at 40°C for 60 minutes, an inactivation-step at 95°C for 5 minutes, followed by subsequent cooling of the reaction at 4°C. The cDNA-samples were stored at -20°C overnight and processed the following day.
Due to the fact that the studied cohorts included children from an age-range of 0.6-16 years (Table 1), which corresponds to various develpmental stages, the use of a conventional reference-gene for relative quantification was deemed ureliable due to possible lack of stability. Therefore, absolute quantification of miRNA expression (copies/μL) was performed using standard curves, produced by 2-fold serial dilutions of reverse-transcribed synthetic oligonucleotides (Qiagen, Germany).
The quantification of miRNA levels was conducted by SYBR® Green-based, real-time quantitative PCR (qPCR), performed on a LightCycler 480ii real-time PCR instrument (Roche, Switzerland). Each reaction of 10μL contained 5μL of 2XmiRCURY SYBR-Green Master Mix, 4μL cDNA and 1μL miRCURY LNA miRNA PCR Assay (Qiagen, Germany) for the respective miRNA. The qPCR protocol involved heat activation at 95°C for 10 minutes, followed by 45 cycles of 95°C for 15s and 60°C for 1 minute and finally a Melting-Curve analysis cycle ranging from 60°C to 95°C, with a ramp rate of 0.03°C/s and continuous fluorescence acquisition.
Threshold cycles (Ct values) were obtained through the instrument’s software by employing the “Absolute Quantification/2nd Derivative Max” analysis. Although the experimental protocol incorporated 45 cycles of amplification, a threshold of 40 cycles was set for reliable detection.
2.5. PAI-1 Genotyping for the PAI-1 (SERPINE1) 4G/5G Polymorphism
Genomic DNA was extracted from white blood cells of the patients’ EDTA-containing whole blood samples, using the NucleoSpin Blood kit (MACHEREY-NAGEL, Germany). Genotyping of the PAI-1 4G/5G polymorphism was conducted by PCR amplification, performed on a Gradient Thermal Cycler (Takara Bio, Japan), using the following primers: 5′- CACAGAGAGAGTCTGGCCACGT-3′ and R: 5′-CCAACAGAGGACTCTTGGTCT-3′. Thermocycling included initial denaturation at 94°C for 4 min, followed by 35 cycles consisting of a denaturation step at 94°C for 0.5 minutes, subsequent annealing at 57°C for 1 minute and an elongation step at 72°C for 1 minute. The final 5-minute elongation step was set at 72°C, ultimately resulting in an amplified fragment of 99 bp [38].
The obtained PCR products underwent overnight incubation with the BslI restriction enzyme at 37°C, which resulted in a 77bp and a 22bp fragment in presence of the 5G allele. Genotyping of the studied samples was carried out through the visualization of the DNA fragments under UV-light, following agarose gel electrophoresis and nucleic-acid staining with GelRed fluorescent stain (Biotium, USA).
2.6. ACE Genotyping for the ACE I/D Polymorphism
Genotyping of the ACE I/D polymorphism was conducted by PCR amplification, performed on a Gradient Thermal Cycler (Takara Bio, Japan), using the following primers for I or D allele amplification: F: 5′-CTGGAGACCACTCCCATCCTTTCT-3′ and R: 5′-GATGTGGCCATCACATTCGTCAGAT-3′.
Th thermocycling protocol included initial denaturation at 94°C for 4 min, followed by 35 cycles consisting of a denaturation step at 94°C for 0.5 minutes, subsequent annealing at 60°C for 1 minute and a 1-minute elongation step at 72°C. The final 5-minute elongation step was set at 72°C. The PCR amplification resulted in two DNA fragments of 490 and 190 bp, respectively corresponding to the I and D alleles [38]. Genotyping was performed under UV-light, following agarose gel electrophoresis and nucleic-acid staining.
2.7. Statistical Analysis
The total of statistical analyses was performed using IBM SPSS Statistics, Version 29.0.2.0. Normality was assessed using the Shapiro-Wilk test for continuous values, according to sample size. Two-sample comparisons were conducted by employing Independent-Samples T-test or Mann-Whitney U test, in respect to the distribution of values. Likewise, comparisons between more than two groups were conducted using “one-way ANOVA” or the Kruskal-Wallis test, as appropriate. Additionally, Jonckheere-Terpstra test was applied for ordered group comparisons and subsequent pairwise analyses. Finally, to examine the effect of the interaction between time and treatment on miRNA expression, General Linear Models were employed. The level of statistical significance was set at p<0.05 (two-tailed).
3. Results
3.1. Group Characteristics
A total of 38 participants were included in the present study, including 19 pediatric patients having undergone thrombosis 1-18 months ago, and 19 healthy controls. Summarized in Table 1 are the characteristics of both studied groups (age and gender), as well as the relevant clinical data of the studied patients, including type (AT or DVT) and classification of thrombotic incident (provoked/unprovoked), site of thrombosis, time intervals between thrombosis and sample collection, treatment status and finally genotypic data as to the 4G/5G polymorphism of the PAI-1 (SERPINE1) gene, as well as to the ACE I/D polymorphism. Man-Whitney U test was applied for the comparison of the quantitative variable of age, yielding no statistically-significant difference between the two groups (p=0.124). Gender-wise, a Chi-Squared Test was performed and also did not reveal a statistically significant difference (p=0.163). As to the type of presented thrombosis, VT represented the vast majority of cases, while only 5% corresponded to AT, consistent to epidemiological data for pediatric thrombosis in the general population, as stated in the literature [3,4,5]. Nearly 50% of the enrolled cases had developed cerebral thrombosis, followed by thrombosis of the lower limbs in 30% of patients and pulmonary thromboembolism (PE) in around 20%. It was estimated that 68% of the studied thrombotic incidents were unprovoked, while a 32% was injury-induced (provoked).
A positive family history for hereditary thrombosis was reported for 63% of patients, which was defined by the occurrence of at least one affected individual per generation (across four generations) through a single parental line in vertical inheritance, or at least two affected individuals when both the paternal and maternal lineages are involved. As to PAI-1 genotyping for the 4G/5G promoter functional polymorphism, 84% of patients were carriers of at least one 4G allele, which is associated with higher SERPINE1 levels and risk of thrombosis [8,14,24,25], while only around 16% carried the normal 5G allele in a homozygous state. In relation to the ACE-I/D functional polymorphism, around 95% of patients carried at least one D allele, associated with elevated ACE levels, while homozygotes for the normal I allele only accounted for 5% of the patient population. (Table 1.).
3.2. Bioinformatic Target-Prediction Analyses
Target prediction analyses suggest that both miR-145-5p and miR-34a-5p hold conserved binding sites on positions 32-38 and 959-966 of the 3′-UTR of SERPINE1 mRNA, respectively. Both score perfect complementarity of the miRNA seed region, while in the case of miR-34a, the mRNA binding site retains an adenine (A), opposite of miRNA position 1 of the seed sequence, thus additionally enhancing the stability of the predicted miRNA/mRNA complex (Figure 1). Both miRNAs demonstrated high microT-scores (>0.7), supporting the validity of predictions. Experimentally-validated negative regulation of SERPINE1 by both miR-145 and miR-34a has been demonstrated through 5 and 21 experiments respectively according to DIANA-microT-CDS analysis, including both direct and indirect validation in various cell types, including endothelial cells in the case of miR-34a.
In addition, target prediction was performed to investigate the possible complementarity and regulatory relationship between the aforementioned miRNAs and the mRNA of the ACE gene, which acts as the indirect controller of PAI-1 levels. The analysis yielded positive results only for miR-34a, which holds a binding site at position 772-779 of ACE 3′ UTR, demonstrating a perfect 7/7 complimentarity and an additional A opposite to position 1 U of the seed sequence, which furtherly stabilizes the miRNA/mRNA complex. The miR-34a/ACE regulatory relationship has been validated in 8 direct experiments and it presents with a microT score of 0.78.
3.3. MiRNA Expression Profiling Between Patients and Controls
The results of the expressional quantification (copies/μL) of the studied miRNAs between patients and controls revealed that the levels of miR-34a-5p were significantly decreased by around 40% in the group of patients, compared to controls (FC=0.61; p=0.029). As to the expression of miR-145-5p, a similar trend towards higher levels in the control population was observed, but did not reach statistical significance (p=0.068) (Table 2, Figure 2).
3.4. MiRNA Expression Profiling Within Patients in Relation to Different Factors
3.4.1. Time Post-Thrombosis
Within the group of patients, in order to assess whether the levels of the studied miRNAs fluctuate over time following thrombosis, the omnibus tests of one-way ANOVA and Kruskal-Wallis were performed, in respect to the distribution of variables. Additionally, to evaluate monotonic trends across the time-groups of patients and subsequent exploratory pairwise comparisons, the Jonckheere-Terpstra (JT) test for ordered variables was applied. The expression levels of miR-145 and miR-34a did not exhibit significant overall significance across time post-thrombosis (p>0.05). However, the 1-3 and 6-12-month time groups differed significantly as to the expression of miR-34a, with the latter exhibiting approximately 45% lower expression compared to the 1-3 group (FC=0.55; p=0.029) (Figure 3), which was additionally confirmed by the conduction of a t-test for independent samples (p=0.031). Pairwise comparisons, by the employment of independent-samples t-test, were also performed for all the other time-groups, which furtherly confirmed the lack of significance as to the expression of both miR-145 and miR-34a (p>0.05). Although not statistically significant, miR-34a remains around 39% reduced during the final 12-18 post-thrombotic interval of the patients, compared to its mean expression in the control group (FC=0.61; p=0.348) (Table 3, Figure 3).
3.4.2. PAI-1 Genotype
Given the fact that both miR-145 and miR-34a are regulators of PAI-1, their expression was evaluated across the different genotypic groups, among the patients in regards to the gene’s 4G/5G (rs1799768) promoter polymorphism. All performed tests (One-way ANOVA, Kruskal-Wallis and Jonckheere-Terpstra) indicated no statistical significance in the expression of the two miRNAs among the genotypic groups (5G5G, 4G5G and 4G4G) (p>0.05), while additional analysis under a dominant model (carriers versus non-carriers of the 4G allele), also yielded insignificant results (Mann-Whitney-U/Independent-Samples t-test, p>0.05). Similarly, pairwise comparisons did not identify any significant differences in miRNA expression among genotypes. Despite the absence of statistical significance, box-plots for miR-34a indicate a mild increase in light of increasing 4G-allele count (Figure 4), while miR-145 did not exhibit any consistent visual trend. This pattern in miR-34a, although highly consistent with biological plausibility and above findings, did not reach statistical significance, probably due to the markedly imbalanced genotypic distributions, since the vast majority of the patients (84%) were carriers of the pathological 4G allele, not allowing for even comparisons across groups.
3.4.3. Treatment Status
Patients were collected in variable timeframes following their thrombotic incidents. Therefore, almost half of the participants (47.4%) within the patient group were under active treatment (including chemoprophylaxis) at the time of sample collection, while the other half (52.6%) were not, either due to recent termination (1-3 months) or long-term discontinuation. Hence, in order to assess whether the treatment affects the expression of the studied miRNAs, the two groups were compared as to miRNA levels factored by treatment status (on/off), as shown in Figure 5. The expression of miR-145-5p did not differ significantly according to therapy status (Mann-Whitney U test, p=0.902), while the respective result for miR-34a was also not significant but exhibited a slightly decreasing trend within the actively treated patients (Independent-Samples t-test, p=0.06). Finally, General Linear Models were used to evaluate the possible effect of the interaction between time post-thrombosis and treatment-status on miRNA expression, which also yielded non-significant results (p>0.05).
3.4.4. Recent Treatment Completion
As yielded from previous analyses, the lowest miR-34a expression was observed in the 6-12 months group, which aligns with recent therapy completion (1-3 months) for the respective patients, a time-point during which the levels of its target (PAI-1) are diminished, according to literature [5,15,22,23]. Hence, recent treatment completion was also examined as a binary variable in the overall addressing of therapy status for both miRNAs within the patient-group (Figure 5). The expression of miR-145 did not exhibit any statistically significant difference between the examined groups, according to the Mann-Whitney U test (p=0.375). On the other hand, miR-34a exhibited a significant 38% downregulation in the group that had recently completed treatment compared to the total of those actively receiving treatment, as well as those who had discontinued treatment for more extended periods and were collected during long-term follow-ups (FC=0.62; p=0.048) (Figure 1, Figure 5).
3.5. MiR-34a Expression Profiling Within Patients in Relation to Different Factors
As per its yielded significant patterns in the above analyses and overall significance between patients and controls, coupled with supporting bioinformatic evidence indicating strong regulatory relationships with PAI-1 and its positive regulator, ACE, we opted to analyze the expression of miR-34a across additional parameters, in pursuit of a clearer elucidation of its role in pediatric thrombosis.
3.5.1. Family History
Within the patient group, miR-34a levels were assessed in relation to the presence of a thrombosis-positive family history, in accordance with the respective set criteria. In both cases no statistical significance was yielded by the performed Mann-Whitney U test and T-test independent-samples, respectively (p>0.05), thus indicating no effect on miRNA expression.
3.5.2. Anatomical Site and Thrombosis Onset Type
To address potential site-specific expressional patterns of miR-34a, patients were stratified according to the anatomical site of the developed thrombi in three groups: cerebral, pulmonary and lower-limb thrombosis. An omnibus one-way ANOVA, comparing miR-34a expression across all anatomically-defined groups, revealed an overall significant difference (p=0.019), hence underlining a strong correlation between thrombus site and miRNA expression (Figure 6). The most significant disparity was noted between the subgroups of cerebral and pulmonary thrombosis, with cerebral cases demonstrating a 76% increase in miR-34a expression, compared to those of pulmonary thromboembolism (FC=1.76; p=0.007).
The cerebral subgroup incorporated both unprovoked and injury-following thrombotic incidents, while the majority (75%) of PE cases were unprovoked. Hence a Mann-Whitney U test was conducted for the evaluation of miR-34a expression in relation to thrombosis onset type (provoked/unprovoked), which alone did not reach statistical significance (p=0.357). However, when a generalized linear model was employed to assess the possible synergistic interaction between anatomical site and thrombosis onset type, a significant effect was detected, with injury-induced cerebral cases demonstrating a 37% increase in miR-34a levels, compared to respective cerebral unprovoked subgroup (p=0.016; FC=1.37).
3.5.3. ACE Genotype
Following the genotyping of the patients’ samples for the functional I/D polymorphism of the ACE gene, an upstream positive regulator of PAI-1 levels, we opted to evaluate miR-34a overall expression across the three genotypes (II, ID and DD), which correspond to ascending levels of ACE [28]. The performed one-way ANOVA and JT tests indicated no significant difference across genotypes (p>0.05); however, a clear visual trend for increasing miR-34a expression across rising count of the ACE-elevating D allele was observed (Figure 7). Since homozygosity for the normal I allele corresponded to only 5% of the studied patients, not adequately allowing for proper omnibus and post-hoc comparisons, the comparison between the I/D and D/D was separately evaluated by an independent-samples T-test. Indeed, patients carrying the DD genotype presented with 22% increased miR-34a expression, although not reaching statistical significance despite the observed trend (FC=1.22; p=0.355).
3.5.4. Combined Effects of Onset Type, ACE Genotype and Thrombosis Site
Within the studied patients, a general linear model analysis revealed a significant three-way interaction between thrombosis anatomical site, onset type and the genotypic presence of at least one D allele of the ACE I/D polymorphism (p=0.035), thus underpinning that miR-34a levels were highly influenced by the combined effect of these anatomical, environmental and genetic variables, rather than a direct environmental/genetic interaction between onset type and D carriage. In fact, cerebral provoked cases, carrying at least on D allele demonstrated a 46% increase in miR-34a expression compared to cerebral unprovoked cases carrying the II genotype (FC=1.46), and a 5.2-fold higher expression (421% increase; FC=5.22), compared to unprovoked incidents in any other anatomical site (lungs and lower limbs) and no presence of the D allele. The GLM model explained one third of the observed variance in miR-34a expression, thus indicating a robust, multifactorial effect on miRNA expression (R2=0.516; adj. R2=0.330).
3.5.5. Combined Effects of 4G4G (PAI-1) and DD (ACE) Genotypes
A general linear model was also applied to assess the possible combinatorial effect of the 4G4G and DD polymorphic genotypes of the PAI-1 and ACE genes, which are associated to the maximum levels of the respective encoded factors. Neither 4G or D homozygosity per se significantly influenced miR-34a expression (p=0.247). However, a parallel univariate GLM analysis including the anatomical site of thrombus development demonstrated a highly significant three-way interaction between PAI-1 4G homozygosity (4G4G), ACE D homozygosity (DD) and thrombosis site (cerebral vs other), that notably affects miRNA expression (p=0.006). Within the patient group, nearly 50% of the expressional heterogeneity of miR-34a was captured by this model (R2=0.663; adj. R2=0.494).
Interaction plots indicated that among patients who had undergone cerebral thrombosis, the double homozygotes (4G4G + DD) exhibited the lowest levels of miR-34a expression. That effect was reversed in 4G4G carriers under the absence of DD homozygosity (II and ID genotypes), demonstrating that DD homozygosity of the ACE gene is a critical determinant of whether the 4G allele of PAI-1 positively or negatively affects miR-34a expression (Figure 8).
4. Discussion
Childhood thrombosis, including the development of venous and arterial thrombi that obstruct circulation, is a rare condition with its etiological origins lying in the disproportionate imbalance between coagulation and fibrinolytic mechanisms [1,2]. Thrombotic incidents can occur at any stage during a child’s development, from the neonatal period to late adolescence, due to the interplay of environmental risk factors, including hypoxia, inflammation and injury, alongside a robust genetically-predesposing background for thrombophilia [9,11,12,13]. The above risk factors contribute to thrombosis development, in a child’s normally immature hemostatic system with a markedly low fibrinolytic capacity, compared to that of adults [1,4,5,6]. Pediatric patients that develop thrombosis come across acute life-threating or long-term complications, secondary incidents, as well as future thrombotic recurrences, typically managed through anticoagulant/fibrinolytic treatment, chemoprophylaxis, but also by a comprehensive assesment of the children’s overall risk that includes their genetic makeup [4,6,8,10].
Among the recognized genetic risk factors associated with thrombophilia, PAI-1 (SERPINE1) elevated gene expression has been identified as the most robust in pediatric populations affected by thrombosis [9]. PAI-1, mainly released by endothelial cells is the predominant inhibitor of fibrinolysis, since its binding to its antagonist, tPA, thus blocking the conversion of plasminogen to active plasmin, which in turn normally degrades fibrin, a proteinic polymer that stabilizes the thrombus [2,13,18,19]. Higher levels of PAI-1 lead to hypercoagulability by diminishing fibrinolysis, thus increasing the risk for thrombus-development, while thrombin itself stimulates additional PAI-1 release in a coagulation-enhancing feedback loop [13,15,16,17,20]. PAI-1 levels demonstrate a progressive rise a priori to the thrombotic incident, peak during the acute phase and thereafter decline after the completion of administrated treatment regimen that corresponds to successful thrombus-resolution and additional prophylaxis [5,15,22,23]. Fibrinolytic capacity of more than 50% of patients remains long-term impaired, for at least 3 months after the thrombotic event, due to PAI-1/t-PA disproportion, which can be attributable to prolonged PAI-1 upregulation [24].
The functional 4G/5G promoter polymorphism of the PAI-1 (SERPINE1) gene is strongly associated with increased circulating PAI-1 levels, leading to subsequent hypofibrinolysis and hypercoagulation, in light of the 4G allele, and has been recently demonstrated as the most common predisposing variant in childhood thrombosis, which can additionally excarbarate already impaired childhood fibrinolysis [9,25,26]. Further to the functional genetic variation of PAI-1 itself, its levels are positively mediated by the ACE, through its active proteolytic product angII, that elevates PAI-1 expression, therefore resulting in hypercoagulability [27,28,29]. The functional I/D polymorphism of the ACE gene that elevates its expression (peaking at D homozygosity) and activity, has been strongly correlated to significant PAI increase, through upstream positive regulation [30,31,32].
In addition to PAI-1, the levels of which are highly indicative of thrombotic development and progression, the implication of miRNAs in adult thrombosis is being actively explored, with highly promising results [33]. However, their role in pediatric thrombosis remains completely uninvestigated, with no relevant studies published to far, possibly due to the rarity of the condition, which restricts sample collection and subsequent research.
In the present study we evaluated serum expression levels of miR-145-5p and miR-34a-5p miRNAs in pediatric patients aged between 0.6 and 16 years, who had undergone thrombosis during the past 1-18 months (Table 1, Figure 1). Within the patient cohort, we assessed the expressional profiles across distinct post-thrombotic time groups and in relation to treatment status, including recent completion of treatment or administered chemoprophylaxis. Potential associations between miRNA expression and the patients’ genotypes for the PAI-1 4G/5G, functional polymorphism were explored. The expressional patterns of miR-34a were furtherly evaluated in relation to the family history of thrombosis, anatomical site of thrombus-development, type of onset (provoked/unprovoked), as well as in regards to ACE I/D genotypes of the patients.
Both studied miRNAs were found to hold conserved binding sites in the 3′-UTR of the PAI-1 gene with perfect complimentarity and have been experimentally shown to negatively regulate its expression through direct experiments. Between these two PAI-1 regulators, miR-34a was bioinformatically shown to form an even more stable complex with PAI-1 mRNA, and also appeared to target the mRNA of the ACE gene as well (Figure 1).
Following RNA extraction and reverse-transcription, the levels of miR-145 and miR-34a were quantified by real-time qPCR. Considering the characteristics of the enrolled study groups, which exhibit age heterogeneity spanning from neonatal life to adolescence and therefore encompasses different develeopmental stages, miRNA expression was evaluated through absolute quantification (copies/μL), in order to alleviate bias induced from possibly unstable reference-genes, the stability of which is required for reliable qPCR Relative-Quantification under the comparative ΔΔCt method.
The guiding hypothesis for the present research was based on an anticipated regulatory interplay between the levels of miR-145 and miR-34a expression and PAI-1 fluctuations, as documented in the literature, across different stages of thrombotic course, clinical parameters, but also different genotypes of the 4G/5G functional polymorphism of the PAI-1, which affect its levels.
Our findings suggest that the expression of serum miR-34a was significantly reduced by approximately 40% in the patient group, compared to the studied healthy children (FC=0.61; p=0.029). On the other hand, the expression of miR-145 did not achieve statistical significance between the two groups, although exhibiting an analogous trend of downregulation (p=0.068).
In regards to different post-thrombotic time groups (Table 1), the 1-3 and 6-12-month groups noted a significant difference, with the latter exhibiting 45%-reduced expression as to miR-34a (p=0.031). Statistical significance was not met in comparisons of the rest of post-thrombotic time intervals; nevertheless, the visual inspection of the overall miR-34a expression patterns suggested a characteristic trajectory. The highest levels within patients were observed during the 1–3-month period, which as the shortest after the thrombotic incidence and PAI-1 expression is expected around its peak. Followingly, a gradual decline of miR-34a was observed, which ultimately reached its lowest point at 6-12 months. This expressional nadir coincided with the recent completion of treatment or chemoprophylaxis (1-3 months) within this specific patient subpopulation, therefore aligning with the anticipated diminishing in PAI-1 levels. Thereafter, reaching the 12-18-month group, the expression of miR-34a demonstrated a subtle increase, which is 39% lower than its respective expression in the control population (Figure 1).
Neither ongoing treatment per se, nor the interaction of treatment and post-thrombotic time seemed to pose an influence on miRNA levels. However, the recent completion of treatment or chemoprophylaxis induced a significant 38% decrease of miR-34a levels (p=0.048), thus suggesting a temporarily dependent effect, possibly reflecting an absence of necessity for PAI-1 regulation, owing to its typically diminished state following a successful treatment course [5,15,22,23].
Taken together, the above observations are highly consistent with a regulatory role, whereby miR-34a acts in a compensational manner, in order to reactively counterbalance the quantitative rise of its target. Its significant downregulation in the patient population compared to controls, which persists even 12-18 months post-incident and prolonged treatment discontinuation, thus suggesting a partial recovery that reflects the long-term impairment of its regulatory capacity. Further support to this conclusion comes from miRNA studies in adult thrombosis, placing miR-34a among the reactively upregulated molecules during the acute phase of thrombotic events, such as an ischemic stroke [39,40], where PAI-1 is known to steeply rise [5,15,22,23].
As to the possible influence of PAI-1 genetic variability on miRNA levels, which was assessed under both general and dominant models, neither miR-145 nor miR-34a, displayed notable differences amongst the PAI-1 genotypes for the 4G/5G polymorphism, with miR-34a however noting a mild expressional increase with rising 4G count and therefore genetically predetermined higher PAI-1 levels [25,26]. Although this increasing trend of miR-34a is biologically plausible, it is possible that statistical significance might have been influenced by imbalanced genotypic distributions, as 84% of patients carried at least one 4G allele, thereby not allowing for balanced group comparisons and posing an interpretational limitation.
Another limitation of the present study is the relatively small sample size, largely attributable to the rarity of cases, which may adversely affect statistical power, thereby partly elucidating the lack of significance in certain overall comparisons, despite consistent patterns and significant pairwise differences (p<0.05). Moreover, the wide dispersion in the expression values, commonly reported in miRNA studies, furtherly contributes to this matter, thus possibly reducing the power of omnibus tests (one-way ANOVA, Kruskal Wallis and Jonckheere Terpstra). Therefore, biological plausibility should be cautiously co-evaluated in light of results demonstrating contradiction between statistical significance and clearly observed trends, before definite conclusions are drawn.
Given that miR-34a repeatedly exhibited significant and biologically plausible findings in our study, we opted to furtherly explore its observed expression within the patient population, across additional clinical and thrombosis-related genomic parameters. Its expression demonstrated a significant association with the anatomical site of the developed thrombi (p=0.019), with a peak at cerebral thrombosis, which was 76% higher compared to the expressional nadir of pulmonary embolism (p=0.007). The synergistic interaction of cerebral involvement and disease-onset type (provoked/unprovoked thrombosis) was also shown to significantly influence miR-34a levels, with post-injury cerebral strokes demonstrating a notable increase (37%) when compared to the respective cerebral unprovoked incidents, thus indicating that the onset exerts differential effects, depending on the thrombosis-affected organ.
The robust effect of injury-induced cerebral involvement is completely in line with both the established upregulation of miR-34a during the acute phase of adult ischemic-stroke, but also with its functionally-demonstrated role to epigenetically counterbalance injury-induced inflammation, thus acting as a protective mechanism [41,42]. In fact, not only it is strongly involved in the acute response to mechanical injury, consistent with the provoked pediatric-stroke cases we analyzed, but also its higher serum overtime-levels have been reportedly associated to successful remission in cases with spinal cord injury. The overall serum levels of injured patients were significantly downregulated compared to non-injured individuals, in accordance with our thrombosis-related overtime-findings, while their assessment has been suggested as a monitoring approach for spinal injury status [43]. These observations agree with the expressional serum-level trajectory of miR-34a that was detected in our study, with its highest levels observed in the time period closest to the thrombotic event, reflecting a compensatory response, followed by its sharp, significant downregulation with treatment-completion, in the setting of pharmacologically induced remission, where its target pathways are externally suppressed. Finally, its tendency to increase in light of long-term absence of treatment confirms its normally protective pro-fibrinolytic role. However, the expressional decrease remains both in the initial reactive response but also long-term after thrombosis, when compared to healthy children with expectedly normal miR-34a function. The latter strongly points out the substantial inadequacy of the miRNA to normally function as an anti-thrombotic protective mechanism, through the effective regulation of the key pediatric thrombosis factor PAI-1 and its upstream indirect activator, ACE. This conclusion gains further strength as the overwhelming majority of studied patients (84%) carried at least one PAI-1 4G allele, which directly increases the mainly implicated PAI-1 gene at all times, while nearly all patients (95%) harbored at least one ACE D allele that has been renown for significantly increasing both levels and activity of its positive regulator and results in even greater levels of pro-thrombotic PAI-1. Therefore, this cannot be regarded as a solely epigenetic observation, rather than an indication of a strong genetic-epigenetic interaction, which may also reflect insight on disease-pathogenesis itself, in complete accordance with the multifactorial nature of thrombosis.
This tentative conclusion is ultimately sealed by the two final genetically-driven combinatorial findings that emerged from our data analyses. It was revealed that the manner in which miR-34a levels behaved within the patient sample not solely depended on either D-allele presence or the onset type across anatomically different thrombotic incidents, but was attributed to their three-way-interaction (p=0.035), with cerebral provoked cases that carried at least on D allele, demonstrating an almost 50% increase in miR-34a expression compared to cerebral unprovoked cases carrying the normal II ACE-lowering genotype, while a tremendous 5.2-fold expressional increase in miR-34a levels was observed when compared to unprovoked non-cerebral incidents (PE or lower-limb thrombosis).
The final and highly significant combinatorial finding involves the interaction of genetic epistasis with an environmental driver that contributes to an apparently already-impaired epigenetic dysregulation and accounts for nearly 50% of miR-34a expressional heterogeneity in the studied patient-sample. The abundance of miR-34a is highly dependent on the three-way interaction between PAI-1 4G homozygosity, ACE D homozygosity and cerebral thrombotic involvement (p=0.006). It was shown that double homozygotes (4G4G + DD) exhibited the lowest levels of miR-34a expression, which additionally reflects the deficiency of miR-34a to adequately increase in light of two genetically predisposed, upregulated targets, with a thrombosis-promoting synergistic effect. The epistatic interaction of the two genes on miR-34a expression was clearly elucidated in light of a complete reverse in miRNA expression in 4G4G carriers lacking the DD homozygosity, whose miR-34a levels were the maximum observed. Hence, the ability of the 4G allele of the PAI-1 gene to exert an anticipated substantial increase on miR-34a levels is not autonomous but emerges only in absence of ACE DD genotype, which acts as the genetic switch. This leads to the conclusion that probably, only in light of double 4G4G + DD homozygosity, where the maximum possible resultant levels of PAI-1 are secreted, the already impaired miR-34a, which is significantly lower to post-thrombotic patients than controls, although not quantitatively enough, is forced to regulate both increased targets that synergistically lead to PAI-1 overflooding. This inadequate response allows for ultimate fibrinolytic dysregulation in children with genetically-determined thrombophilia, which when combined with their immature hemostatic system can pave the way for thrombosis development.
This possibly defective regulation could also provide a mechanistic hint that miR-34a might normally function as a regulatory “emergency brake” in thrombosis driven by PAI-1-induced hypofibrinolysis. By failing to adequately control the expression of PAI-1 due to its insufficient expression, it allows for its target’s unchecked elevation, which is already renown to drive thrombosis development.
Moreover, our findings suggest that the genetic epistasis between ACE and PAI-1 (4G4G + DD) as the main determinant for lack of the normally anticipated anti-thrombotic response of miR-34a, might provide a combinatorial genetic/epigenetic predisposing profile, which is highly possible to account for thrombosis development, by acting as a multivariate, molecular switch in light of an environmental trigger, such as injury. This conclusion is furtherly supported by the fact that PAI-1 4G/5G polymorphism has been recognized as the predominant risk factor for childhood thrombophilia among many established risk factors that are considered much more important in adult thrombosis [9]. Since to our knowledge the present exploratory piece of research is the first so far conducted in the context of pediatric thrombosis and miRNA profiling, we anticipate that it will lay the groundwork for the future broadening of predisposition from a solely genetic term to a genetic/epigenetic dynamic concept.
Author Contributions
IG conceived and designed the particular study, performed the experiments and bioinformatic analyses, authored the original manuscript and co-revised the final draft, performed statistical analysis and drew the figures. KB, AD, EM, and EP actively participated in clinical evaluation and sample collection of patients and controls. DV, AT, IP assisted in study supervision. GPC participated in study design and performed critical text reviewing. CY is the approved protocol scientific coordinator, supervised and designed the overall study, coordinated and performed sample collection, secured the study’s conduction under bioethical standards, collected extended family history and clinical data, standardized the PAI-1 and ACE genotyping protocols, and revised the final draft of the manuscript.
Funding
The present research was funded by the University Research Institute of Maternal and Child Health and Precision Medicine, as to the consumables used to perform sample collection and molecular analyses.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and was bioethically approved by the respective Committee of the “Aghia Sophia” Children’s University Hospital (protocol 21330/2020).
Informed Consent Statement
Informed written consent was obtained from the parents of all minors involved in the study.
Data Availability Statement
Original data presented in this research shall be available upon request to the corresponding author. Certain data may be subjected to limited availability in accordance to confidentiality and privacy protecting protocols or temporary ongoing study conduction.
Acknowledgments
The authors would like to thank all participants and their parents for contributing to this research.
Conflicts of Interest
There are no conflicts of interest to declare.
References
- Celkan, T.; Dikme, G. Thrombosis in Children: Which Test to Whom, When and How Much Necessary? Turk Pediatri Ars 2018, 53, 1–9. [Google Scholar] [CrossRef]
- Alkarithi, G.; Duval, C.; Shi, Y.; Macrae, F.L.; Ariëns, R.A.S. Thrombus Structural Composition in Cardiovascular Disease. Arterioscler Thromb Vasc Biol 2021, 41, 2370–2383. [Google Scholar] [CrossRef]
- Abdelghani, E.; Agarwal, S.; Stanek, J.; Sankar, A.; Kerlin, B.A.; Rodriguez, V. Pediatric Arterial Thrombosis: A Single-Institution Cohort Study of Patient Characteristics and Thrombosis Outcomes. Pediatr Blood Cancer 2024, 71, e30756. [Google Scholar] [CrossRef]
- Audu, C.O.; Wakefield, T.W.; Coleman, D.M. Pediatric Deep Venous Thrombosis. J Vasc Surg Venous Lymphat Disord 2019, 7, 452–462. [Google Scholar] [CrossRef]
- Bacciedoni, V.; Attie, M.; Donato, H.; Comité Nacional de Hematología, O.y.M.T. . Thrombosis in Newborn Infants. Arch Argent Pediatr 2016, 114, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Branchford, B.R.; Betensky, M.; Goldenberg, N.A. Pediatric Issues in Thrombosis and Hemostasis: The How and Why of Venous Thromboembolism Risk Stratification in Hospitalized Children. Thromb Res 2018, 172, 190–193. [Google Scholar] [CrossRef]
- Papadogeorgou, P.; Valsami, S.; Boutsikou, M.; Pergantou, E.; Mantzou, A.; Papassotiriou, I.; Iliodromiti, Z.; Sokou, R.; Bouza, E.; Politou, M.; et al. Coagulation Profile in Neonates with Congenital Heart Disease: A Pilot Study. Medicina (Kaunas) 2024, 60. [Google Scholar] [CrossRef] [PubMed]
- Engel, E.R.; Nguyen, A.T.H.; Amankwah, E.K.; Albisetti, M.; Brandão, L.R.; Goldenberg, N.A.; Betensky, M. Predictors of Postthrombotic Syndrome in Pediatric Thrombosis: A Systematic Review and Meta-Analysis of the Literature. J Thromb Haemost 2020, 18, 2601–2612. [Google Scholar] [CrossRef]
- Sava, C.; Iuhas, A.; Balmoș, A.; Niulaș, L.; Marinău, C.; Futaki, Z.; Bei, D.; Kozma, K.; Ritli, L.; Szilagyi, A. Thrombosis in Children: A Retrospective Study from a Single-Center Database. In Vivo 2025, 39, 473–481. [Google Scholar] [CrossRef]
- Gerstl, L.; Bonfert, M. V; Heinen, F.; Olivieri, M.; Schroeder, A.S.; Tacke, M.; Kurnik, K.; Bidlingmaier, C.; Reiter, K.; Borggraefe, I. Childhood Arterial Ischaemic Stroke: Clinical Presentation, Risk Factors and Management. Hamostaseologie 2020, 40, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Guzoglu, N.; Albayrak, M.; Aliefendioglu, D. Evaluation of Patients with Neonatal Thrombosis. Indian J Pediatr 2023, 90, 615–617. [Google Scholar] [CrossRef]
- Krleza, J.L.; Coen Herak, D.; Đakovic, I.; Vulin, K.; Roic, G.; Tripalo Batoš, A.; Čeri, A.; Zadro, R.; Đuranovic, V. Inherited Thrombophilia Associated With Ischemic Pediatric Stroke in Parent-Child Pairs. Pediatr Neurol 2023, 146, 119–128. [Google Scholar] [CrossRef]
- Huebner, B.R.; Moore, E.E.; Moore, H.B.; Gonzalez, E.; Kelher, M.R.; Sauaia, A.; Banerjee, A.; Silliman, C.C. Thrombin Stimulates Increased Plasminogen Activator Inhibitor-1 Release from Liver Compared to Lung Endothelium. J Surg Res 2018, 225, 1–5. [Google Scholar] [CrossRef]
- Saedi, S.; Yazzaf, R.; Parsaee, M.; Maleki, M. Inherited Thrombophilia in Mechanical Valve Malfunction. Ann Cardiol Angeiol (Paris) 2020, 69, 213–215. [Google Scholar] [CrossRef]
- AL, M.-V. Role of Polymorphisms in Factor V (FV Leiden), Prothrombin, Plasminogen Activator Inhibitor Type-1 (PAI-1), Methylenetetrahydrofolate Reductase (MTHFR) and Cystathionine β-Synthase (CBS) Genes as Risk Factors for Thrombophilias. Mini Rev Med Chem 2012, 12, 997–1006. [Google Scholar] [CrossRef]
- Seheult, J.N.; Chibisov, I. A Case of Unexplained Cerebral Sinus Thrombosis in a 22-Year-Old Obese Caucasian Woman. Lab Med 2016, 47, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Zhang, G.; Yan, D.; Yang, H.; Ma, T.; Ye, Z. Modulation of Plasminogen Activator Inhibitor-1 (PAI-1) by the Naphthoquinone Shikonin. Fitoterapia 2016, 113, 117–122. [Google Scholar] [CrossRef]
- Weisel, J.W. Fibrinogen and Fibrin. Adv Protein Chem 2005, 70, 247–299. [Google Scholar] [CrossRef] [PubMed]
- Tarango, C.; Manco-Johnson, M.J. Pediatric Thrombolysis: A Practical Approach. Front Pediatr 2017, 5, 260. [Google Scholar] [CrossRef] [PubMed]
- Basurto, L.; Manuel-Apolinar, L.; Robledo, A.; O’Leary, S.; Martínez-Murillo, C.; Medina-Ortíz, L.I.; Montes Osorio, M.G.; Zarazua, J.; Balcázar-Hernández, L.; Anda-Garay, J.C. Thrombotic Risk Assessed by PAI-1 in Patients with COVID-19: The Influence of Hyperglycemia and Diabetes Mellitus. Clin Investig Arterioscler 2024, 36, 201–209. [Google Scholar] [CrossRef]
- Brown, N.J. Therapeutic Potential of Plasminogen Activator Inhibitor-1 Inhibitors. Ther Adv Cardiovasc Dis 2010, 4, 315–324. [Google Scholar] [CrossRef]
- Wang, N.; Gao, Y.; Wang, Y.; Dai, Y.; Tang, Y.; Huang, J.; Sun, L.; Qian, G.; Ma, J.; Li, X.; et al. Plasma Proteomic Profiling Reveals That SERPINE1 Is a Potential Biomarker Associated with Coronary Artery Lesions in Kawasaki Disease. Int Immunopharmacol 2024, 139. [Google Scholar] [CrossRef]
- Mo, J.W.; Zhang, D.F.; Ji, G.L.; Liu, X.Z.; Fan, B. TGF-Β1 and Serpine 1 Expression Changes in Traumatic Deep Vein Thrombosis. Genet Mol Res 2015, 14, 13835–13842. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.; Horellou, M.H.; Kruithof, E.K.; Conard, J.; Samama, M.M. Residual Plasminogen Activator Inhibitor Activity after Venous Stasis as a Criterion for Hypofibrinolysis: A Study in 83 Patients with Confirmed Deep Vein Thrombosis. Blood 1988, 72, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Glueck, C.J.; Bell, H.; Vadlamani, L.; Gupta, A.; Fontaine, R.N.; Wang, P.; Stroop, D.; Gruppo, R. Heritable Thrombophilia and Hypofibrinolysis. Possible Causes of Retinal Vein Occlusion. Arch Ophthalmol 1999, 117, 43–49. [Google Scholar] [CrossRef]
- Sundquist, K.; Wang, X.; Svensson, P.J.; Sundquist, J.; Hedelius, A.; Lönn, S.L.; Zöller, B.; Memon, A.A. Plasminogen Activator Inhibitor-1 4G/5G Polymorphism, Factor V Leiden, Prothrombin Mutations and the Risk of VTE Recurrence. Thromb Haemost 2015, 114, 1156–1164. [Google Scholar] [CrossRef]
- Moore, J.H.; Smolkin, M.E.; Lamb, J.M.; Brown, N.J.; Vaughan, D.E. The Relationship between Plasma T-PA and PAI-1 Levels Is Dependent on Epistatic Effects of the ACE I/D and PAI-1 4G/5G Polymorphisms. Clin Genet 2002, 62, 53–59. [Google Scholar] [CrossRef]
- Gintoni, I.; Adamopoulou, M.; Yapijakis, C. The Angiotensin-Converting Enzyme Insertion/Deletion Polymorphism as a Common Risk Factor for Major Pregnancy Complications. In Vivo 2021, 35, 95–103. [Google Scholar] [CrossRef]
- Felmeden, D.C.; Lip, G.Y. The Renin-Angiotensin-Aldosterone System and Fibrinolysis. J Renin Angiotensin Aldosterone Syst 2000, 1, 240–244. [Google Scholar] [CrossRef]
- Vairaktaris, E.; Yapijakis, C.; Serefoglou, Z.; Vylliotis, A.; Ries, J.; Nkenke, E.; Wiltfang, J.; Derka, S.; Vassiliou, S.; Springer, I.; et al. Plasminogen Activator Inhibitor-1 Polymorphism Is Associated with Increased Risk for Oral Cancer. Oral Oncol 2006, 42, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Gintoni, I.; Adamopoulou, M.; Yapijakis, C. The Impact of ACE and ACE2 Gene Polymorphisms in Pulmonary Diseases Including COVID-19. In Vivo 2022, 36, 13–29. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, S.; Lei, J.; Zhang, J. [Association of Angiotensin I-Converting Enzyme Gene Polymorphism with ACE and PAI-1 Levels in Guangdong Chinese Han Patients with Essential Hypertension]. Nan Fang Yi Ke Da Xue Xue Bao 2007, 27, 1681–1684. [Google Scholar]
- Yapijakis, C. The Role of MicroRNAs in Thrombosis. Adv Exp Med Biol 2021, 1339, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Gintoni, I.; Vassiliou, S.; Chrousos, G.P.; Yapijakis, C. Identification of Stage-Specific MicroRNAs That Govern the Early Stages of Sequential Oral Oncogenesis by Strategically Bridging Human Genetics with Epigenetics and Utilizing an Animal Model. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Gintoni, I.; Vassiliou, S.; Chrousos, G.P.; Yapijakis, C. Review of Disease-Specific MicroRNAs by Strategically Bridging Genetics and Epigenetics in Oral Squamous Cell Carcinoma. Genes (Basel) 2023, 14, 1578. [Google Scholar] [CrossRef]
- Muth, D.C.; Powell, B.H.; Zhao, Z.; Witwer, K.W. MiRNAs in Platelet-Poor Blood Plasma and Purified RNA Are Highly Stable: A Confirmatory Study. BMC Res Notes 2018, 11, 273. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv Exp Med Biol 2020, 1253, 3–55. [Google Scholar] [CrossRef] [PubMed]
- Yapijakis, C.; Serefoglou, Z.; Voumvourakis, C. Common Gene Polymorphisms Associated with Thrombophilia. In Thrombosis, Atherosclerosis and Atherothrombosis—New Insights and Experimental Protocols; InTech, 2015. [Google Scholar]
- Wei, G.-J.; Yuan, M.-Q.; Jiang, L.-H.; Lu, Y.-L.; Liu, C.-H.; Luo, H.-C.; Huang, H.-T.; Qi, Z.-Q.; Wei, Y.-S. A Genetic Variant of MiR-34a Contributes to Susceptibility of Ischemic Stroke Among Chinese Population. Front Physiol 2019, 10, 432. [Google Scholar] [CrossRef]
- Ansari Dezfouli, M.; Esmati, M.; Rashidi, S.K.; Rafie, S.; Mahmoudian-Sani, M.-R.; Behzad, E. SIRT1 and MiR-34a as Potential Plasma Biomarkers in the Acute Phase of Ischemic Stroke. Int J Mol Cell Med 2025, 14, 714–725. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Zhang, Y.; Chen, W. [MiR-34a Alleviates Podocyte Injury in Mice with Diabetic Nephropathy by Targeted Downregulation of Notch Signaling Pathway]. Nan Fang Yi Ke Da Xue Xue Bao 2022, 42, 1839–1845. [Google Scholar] [CrossRef]
- Zhou, J.; Shuang, O.; Li, J.; Cai, Z.; Wu, C.; Wang, W. MiR-34a Alleviates Spinal Cord Injury via TLR4 Signaling by Inhibiting HMGB-1. Exp Ther Med 2019, 17, 1912–1918. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Gu, H.; Yang, H. MiR-34a May Function as Potential Biomarker for Remission after Traumatic Spinal Cord Injury. Clin Lab 2020, 66. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Graphical abstract of study design and key findings upon miR-34a expression in pediatric post-thrombotic course, as well as visualized outputs from target-prediction in-silico analyses (Targetscan, miRNet) presented alongside experimental results. Bioinformatic analyses revealed. Target-prediction analyses yielded that miR-145-5p and miR-34a-5p hold conserved binding sites on the 3′-UTR of SERPINE1 (PAI-1) mRNA, both scoring perfect complementarity of the miRNA seed region, while in the case of miR-34a, mRNA binding exhibits possibly increased stability due to the presence of an adenine (A), opposite of miRNA position 1 of the seed sequence. MiR-34a additionally holds a 7/7 binding site on the 3′ UTR of the mRNA of the ACE gene, which codes for the upstream positive regulator of PAI-1. opposite to seed-sequence position 1. The expression of miR-34a was significantly decreased in patients, compared to controls, while no difference was observed for the levels of miR-145, between the two studied cohorts. The lower panel presents a combinatory graph that illustrates the expressional trajectory of miR-34a across the post-thrombotic course with a peak at 1-3 months post-thrombosis, a time when PAI-1 is anticipated to be significantly increased. Its expression follows a gradual decline and finally a nadir, shortly after treatment-completion, when PAI-1 has been demonstrated to diminish. At the long-term post-thrombotic stage of 12-18 months, where PAI expression is expected recover, miR-34a strikes a slight, non-significant increase reaching lower levels than those of the studied controls. These findings are indicative of a dynamic, yet impaired regulatory relationship between miR-34a and PAI-1, as well as its upstream controller ACE. Hence, the downregulation of miR-34a in pediatric thrombosis might possibly lead to deficient fibrinolytic capacity through the inadequate suppression of the antifibrinolytic ACE/PAI-1 axis, the upregulation of which is critical for thrombosis development.
Figure 1.
Graphical abstract of study design and key findings upon miR-34a expression in pediatric post-thrombotic course, as well as visualized outputs from target-prediction in-silico analyses (Targetscan, miRNet) presented alongside experimental results. Bioinformatic analyses revealed. Target-prediction analyses yielded that miR-145-5p and miR-34a-5p hold conserved binding sites on the 3′-UTR of SERPINE1 (PAI-1) mRNA, both scoring perfect complementarity of the miRNA seed region, while in the case of miR-34a, mRNA binding exhibits possibly increased stability due to the presence of an adenine (A), opposite of miRNA position 1 of the seed sequence. MiR-34a additionally holds a 7/7 binding site on the 3′ UTR of the mRNA of the ACE gene, which codes for the upstream positive regulator of PAI-1. opposite to seed-sequence position 1. The expression of miR-34a was significantly decreased in patients, compared to controls, while no difference was observed for the levels of miR-145, between the two studied cohorts. The lower panel presents a combinatory graph that illustrates the expressional trajectory of miR-34a across the post-thrombotic course with a peak at 1-3 months post-thrombosis, a time when PAI-1 is anticipated to be significantly increased. Its expression follows a gradual decline and finally a nadir, shortly after treatment-completion, when PAI-1 has been demonstrated to diminish. At the long-term post-thrombotic stage of 12-18 months, where PAI expression is expected recover, miR-34a strikes a slight, non-significant increase reaching lower levels than those of the studied controls. These findings are indicative of a dynamic, yet impaired regulatory relationship between miR-34a and PAI-1, as well as its upstream controller ACE. Hence, the downregulation of miR-34a in pediatric thrombosis might possibly lead to deficient fibrinolytic capacity through the inadequate suppression of the antifibrinolytic ACE/PAI-1 axis, the upregulation of which is critical for thrombosis development.
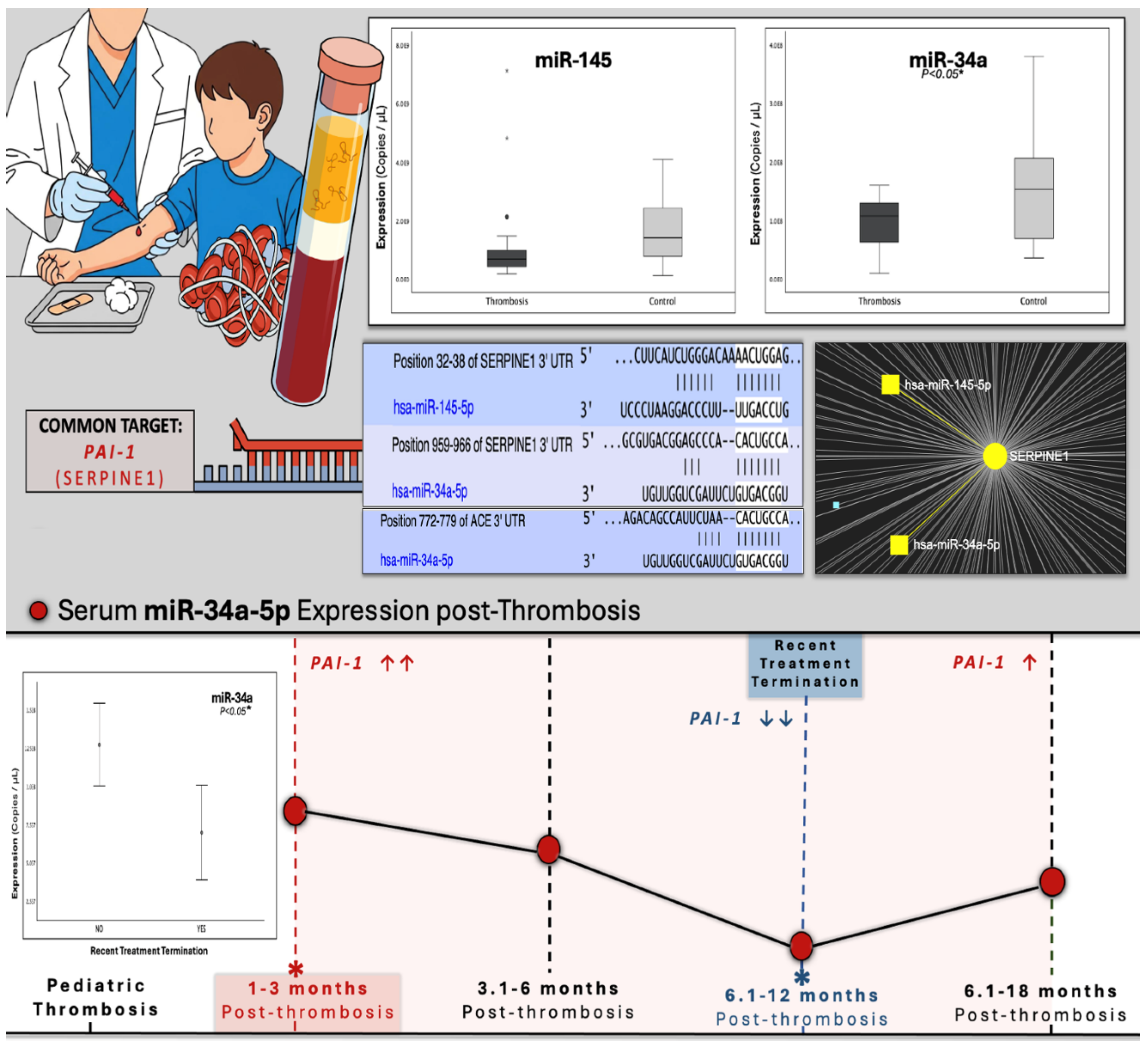
Figure 2.
The graphs depict the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p that was studied in serum samples of pediatric thrombosis patients (1-18 months post-incident) and healthy controls. It was shown that miR-34a was significantly downregulated (p<0.05) in the patient population compared to controls, while the expression of miR-145-5p exhibited no statistical difference between the two groups.
Figure 2.
The graphs depict the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p that was studied in serum samples of pediatric thrombosis patients (1-18 months post-incident) and healthy controls. It was shown that miR-34a was significantly downregulated (p<0.05) in the patient population compared to controls, while the expression of miR-145-5p exhibited no statistical difference between the two groups.

Figure 3.
Graphs depicting the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p that was studied across four time-groups following thrombosis occurrence. (c), Error bars of miR-34a expression across all time-groups, highlighting its trajectory through the post-thrombotic course and (d), the comparison between the 1-3 and 6-12 time-groups. Statistical significance was set at p<0.05.
Figure 3.
Graphs depicting the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p that was studied across four time-groups following thrombosis occurrence. (c), Error bars of miR-34a expression across all time-groups, highlighting its trajectory through the post-thrombotic course and (d), the comparison between the 1-3 and 6-12 time-groups. Statistical significance was set at p<0.05.
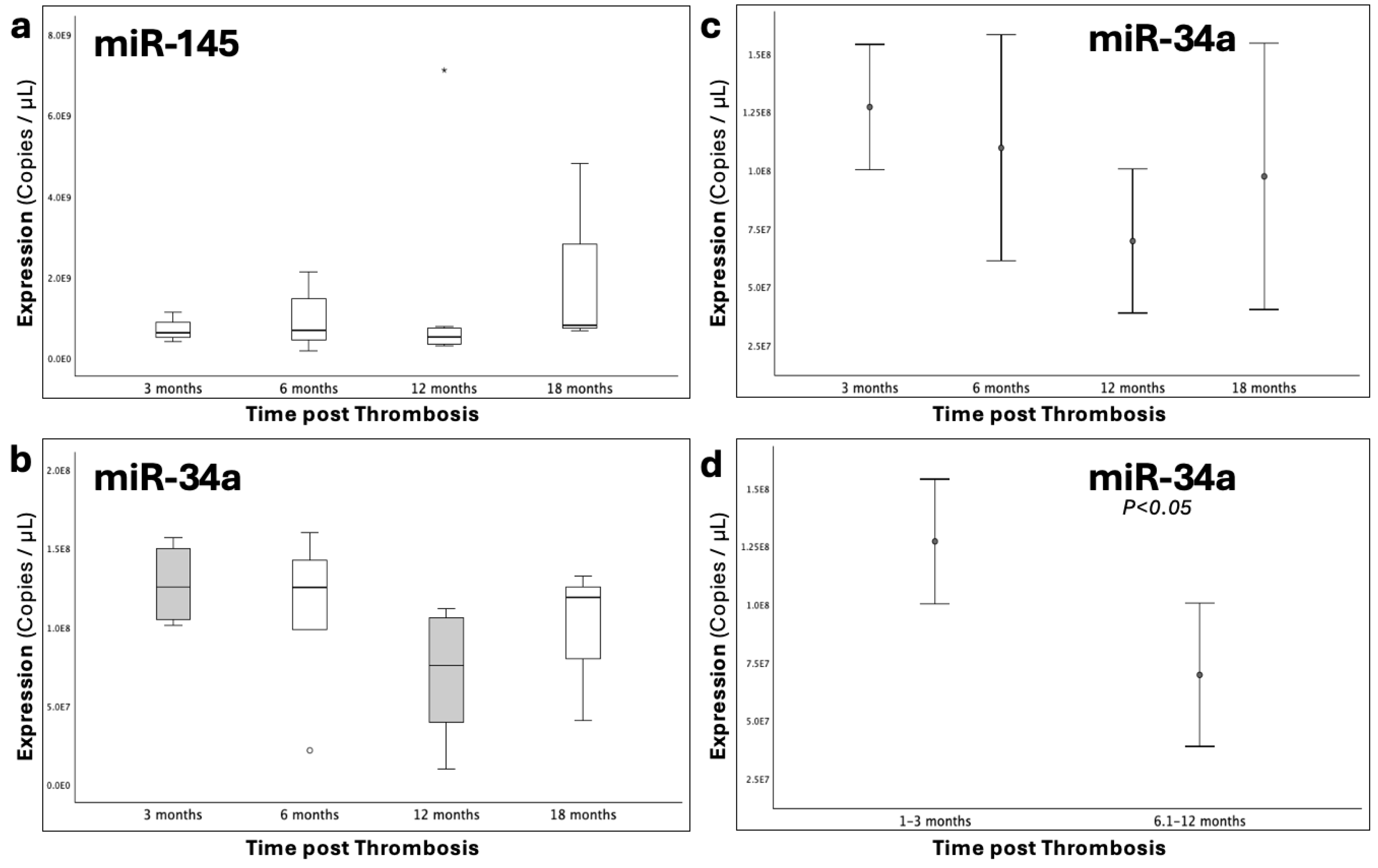
Figure 4.
Boxplots depicting the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p that was studied across genotypic groups for the 4G/5G functional polymorphism of the PAI-1 gene, which increases the expression of PAI-1 in light of the 4G allele.
Figure 4.
Boxplots depicting the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p that was studied across genotypic groups for the 4G/5G functional polymorphism of the PAI-1 gene, which increases the expression of PAI-1 in light of the 4G allele.

Figure 5.
The graphs depict the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p across patient subgroups receiving and not actively receiving treatment following thrombosis. c), Error bars of miR-34a expression between patients that had recently completed treatment regimen (1-3 months), and patients either actively receiving or having long-term terminated treatment course. Statistical significance was set at p<0.05.
Figure 5.
The graphs depict the expression (copies/μL) of (a), miR-145-5p and (b), miR-34a-5p across patient subgroups receiving and not actively receiving treatment following thrombosis. c), Error bars of miR-34a expression between patients that had recently completed treatment regimen (1-3 months), and patients either actively receiving or having long-term terminated treatment course. Statistical significance was set at p<0.05.
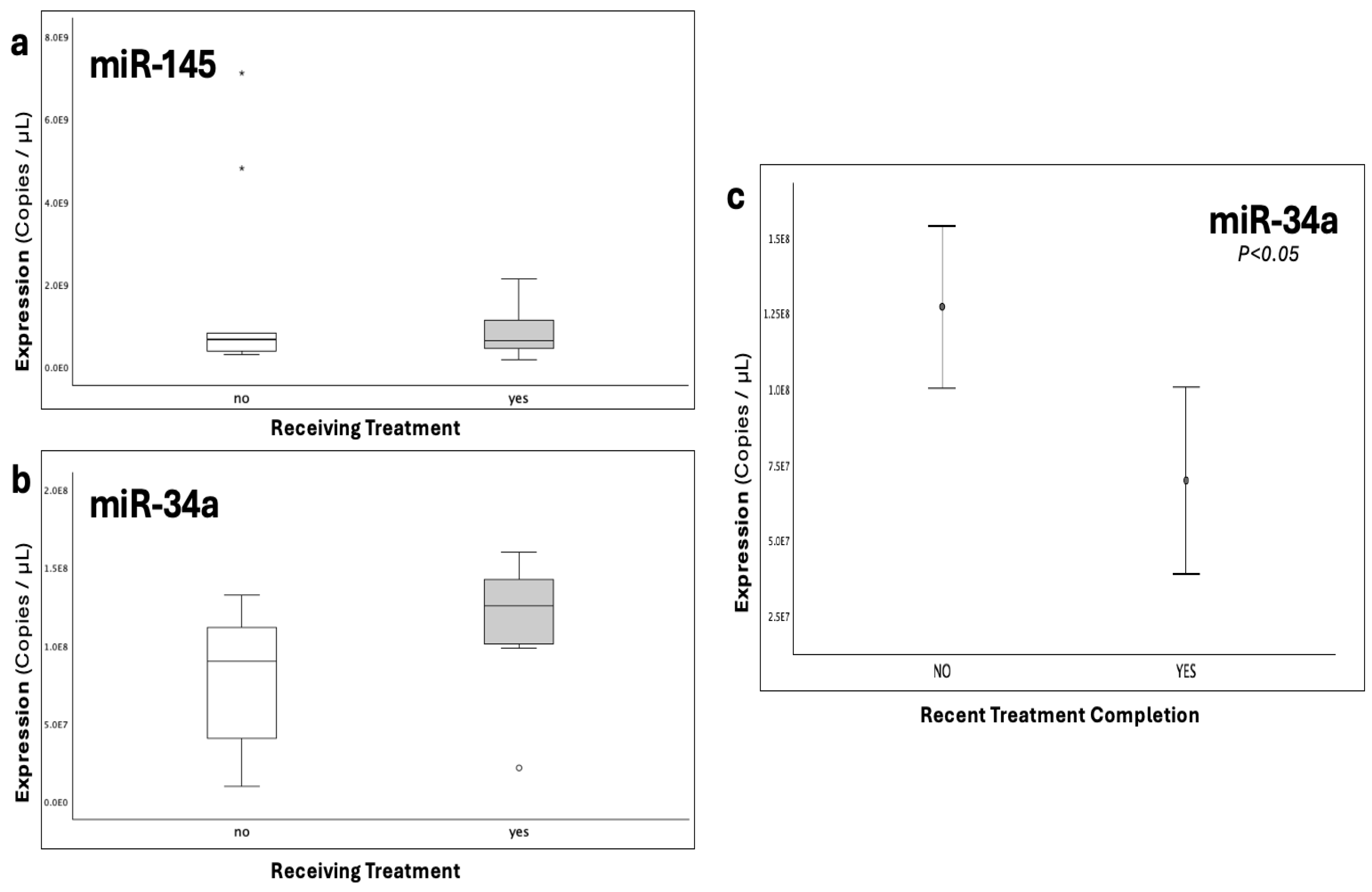
Figure 6.
The graphs present the expression (copies/μL) of (a), miR-34a-5p across different anatomical sites, where the thrombi were developed, including cerebral thrombosis (brain), pulmonary embolism (lung) and lower-limb thrombosis; and (b), miR-34a-5p under the combined influence of cerebral involvement and onset-type (provoked/unprovoked).
Figure 6.
The graphs present the expression (copies/μL) of (a), miR-34a-5p across different anatomical sites, where the thrombi were developed, including cerebral thrombosis (brain), pulmonary embolism (lung) and lower-limb thrombosis; and (b), miR-34a-5p under the combined influence of cerebral involvement and onset-type (provoked/unprovoked).
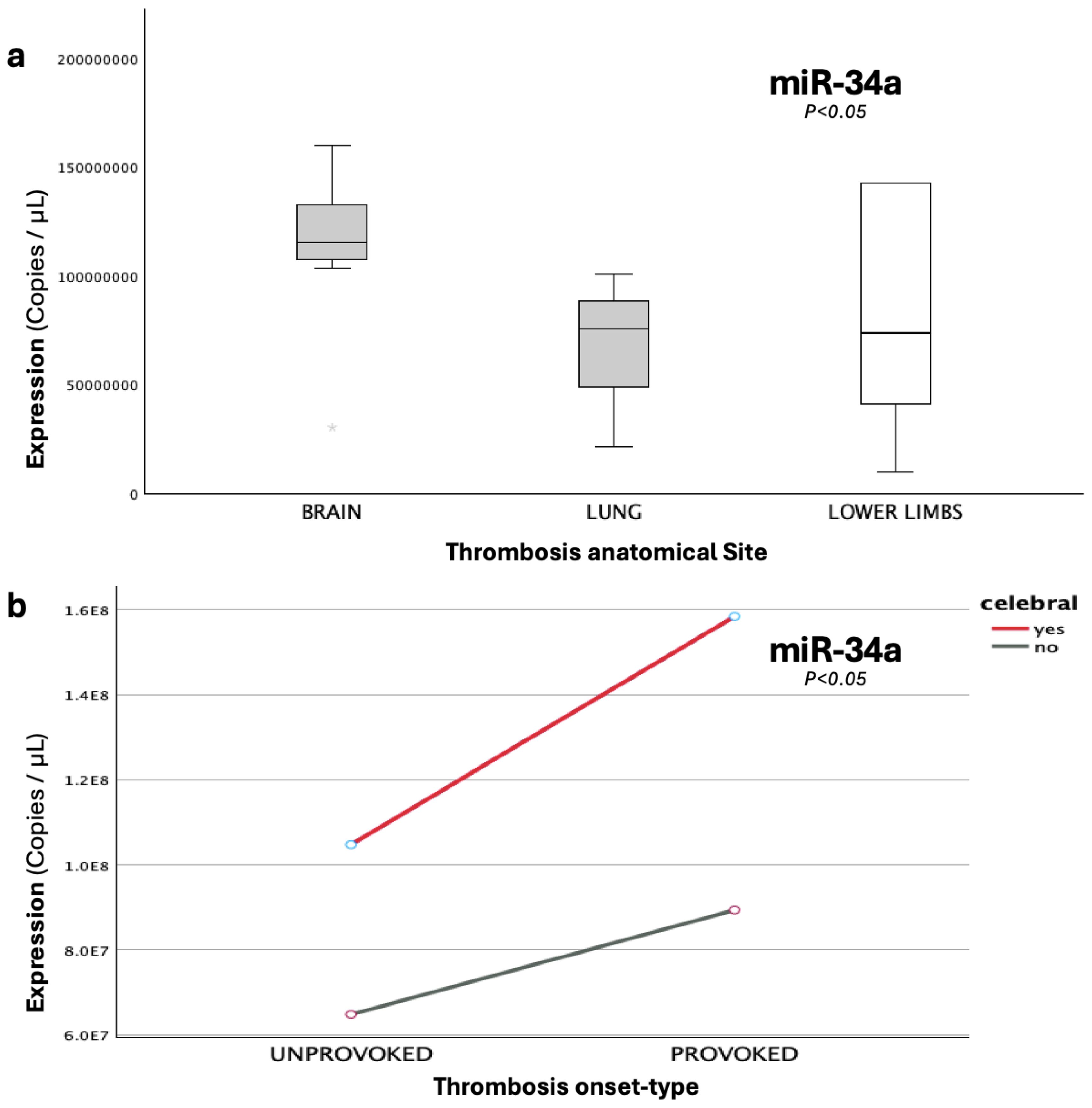
Figure 7.
Boxplots illustrating the expression (copies/μL) of miR-34a-5p across genotypic groups for the I/D functional polymorphism of the ACE gene, which increases the expression of ACE in light of the D allele, thus indirectly upstream increasing PAI-1 levels.
Figure 7.
Boxplots illustrating the expression (copies/μL) of miR-34a-5p across genotypic groups for the I/D functional polymorphism of the ACE gene, which increases the expression of ACE in light of the D allele, thus indirectly upstream increasing PAI-1 levels.
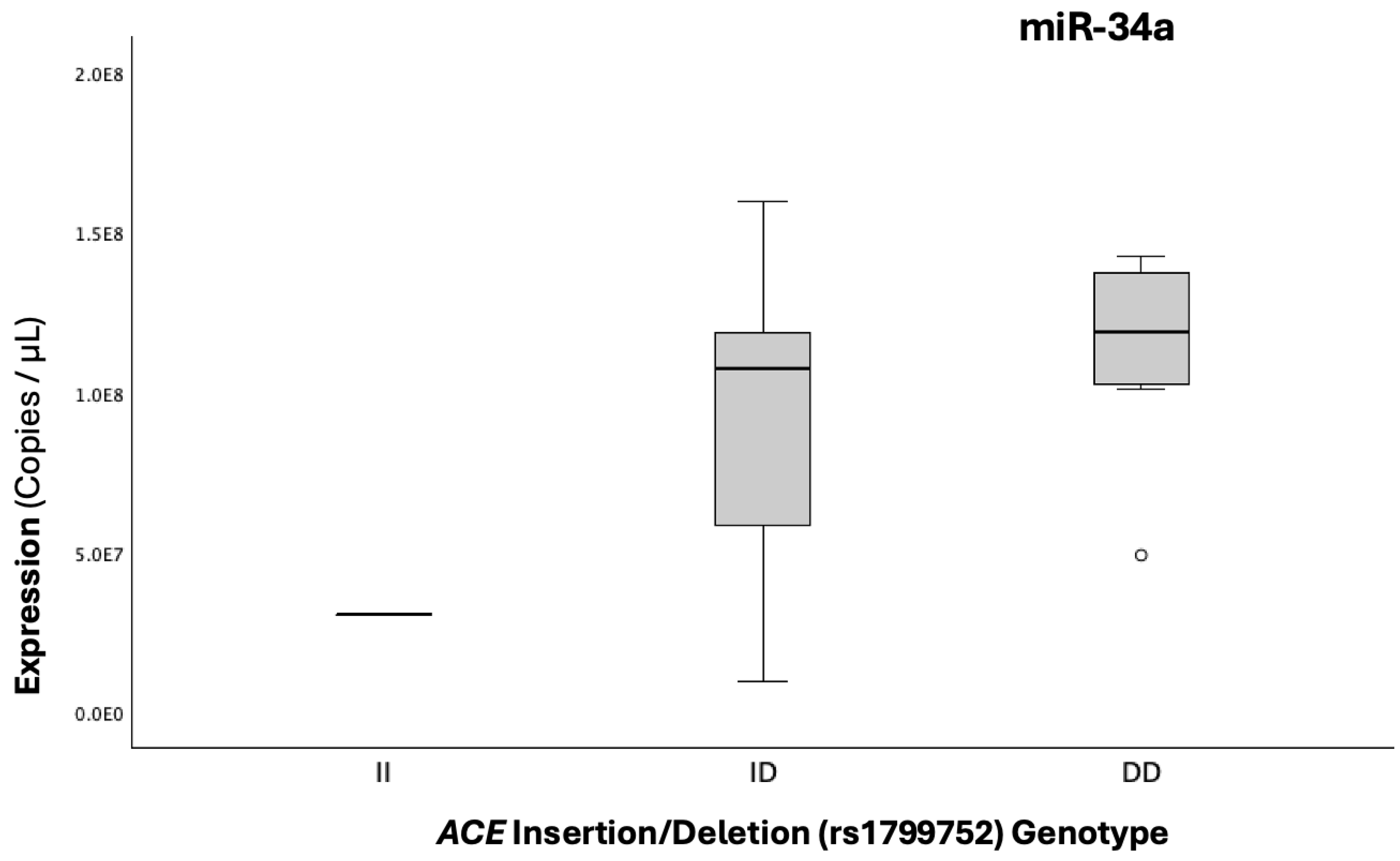
Figure 8.
The graph illustrates the epistatic modification, which was observed in cerebral cases, with double homozygosity (4G/4G+D/D), for the PAI-1-4G/5G and ACE-I/D gene functional polymorphisms, leading to miR-34a diminishing, while solely the 4G4G (in absence of DD carriership) exerts the reverse effect of significant miR-34a upregulation (p=0.006).
Figure 8.
The graph illustrates the epistatic modification, which was observed in cerebral cases, with double homozygosity (4G/4G+D/D), for the PAI-1-4G/5G and ACE-I/D gene functional polymorphisms, leading to miR-34a diminishing, while solely the 4G4G (in absence of DD carriership) exerts the reverse effect of significant miR-34a upregulation (p=0.006).
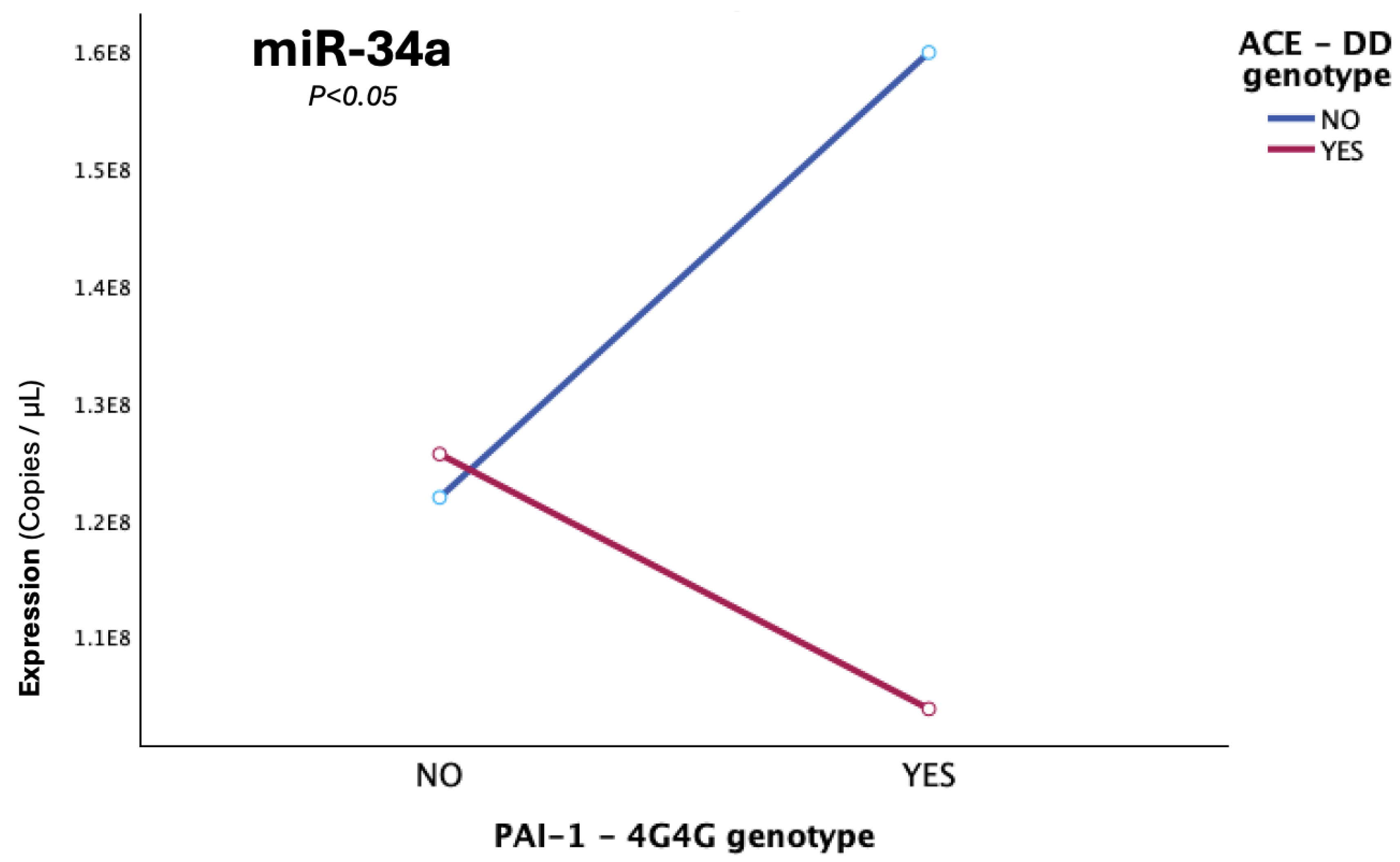

Table 1.
Characteristics and clinical data of the studied patient and control groups. Variables such as age and gender were statistically compared between the two groups by utilization of the appropriate statistical tests. The level of statistical significance was set at p<0.05. The studied groups did not differ significantly as to the variables of age and gender. 1 Mean (Range); a Mann-Whitney U test (2-sided), b Pearson Chi-Square (2-sided), c Independent Samples T-test.
Table 1.
Characteristics and clinical data of the studied patient and control groups. Variables such as age and gender were statistically compared between the two groups by utilization of the appropriate statistical tests. The level of statistical significance was set at p<0.05. The studied groups did not differ significantly as to the variables of age and gender. 1 Mean (Range); a Mann-Whitney U test (2-sided), b Pearson Chi-Square (2-sided), c Independent Samples T-test.
| Variable | Patients (n=19) | Controls (n=19) | P-value | |||||
| Age (years) 1 | 9.51 (0.6-16) | 7.03 (0.6-15) | 0.124a | |||||
|
Gender n (%) |
Male | Female | Male | Female | 0.163b | |||
| 15 (78.9%) | 4 (21.1%) |
11 (57.9%) | 8 (42.1%) |
|||||
|
Thrombosis Patients (n=19) | ||||||||
| Type of Thrombosis (n,%) | ||||||||
| Arterial (AT) | Venous (VTE) | |||||||
| 1 (5.3%) | 18 (94.7%) | |||||||
| Anatomical Site of Thrombosis (n,%) | ||||||||
| Cerebral | Lower Limbs | Pulmonary | ||||||
| 9 (47.4%) | 6 (31.6%) | 4 (21.0%) | ||||||
| Thrombotic Incident Classification (n,%) | ||||||||
| Provoked | Unprovoked | |||||||
| 6 (31.6%) | 13 (68.4%) | |||||||
| Months post-Thrombosis 1 | ||||||||
| 7.63 (1-18) | ||||||||
|
Time Groups (months; n,%) |
1-3 | 3.1-6 | 6.1-12 | 12.1-18 | ||||
| 4 (21.1%) | 5 (26.32%) | 7 (36.8%) | 3 (15.79%) | |||||
| Treatment | Yes (47.37%) | No (52.63%) | ||||||
| Family History of Thrombosis (n,%) | P-value | |||||||
| Positive | Negative | |||||||
| 12 (63.2%) | 7 (36.8%) | 0.710c | ||||||
| PAI-1(rs1799889) Genotype (n,%) | ||||||||
| 5G5G | 4G5G | 4G4G | ||||||
| 3 (15.8%) | 12 (63.2%) | 4 (21.1%) | ||||||
| ACE I/D (rs1799752) Genotype (n,%) | ||||||||
| II | ID | DD | ||||||
| 1 (5.3%) | 11 (57.9%) | 7 (36.8%) | ||||||
Table 2.
Absolute expression of miR-145-5p and miR-34a-5p in the group of patients that underwent thrombotic incidents and in the group of healthy controls. The expression of the under-study miRNAs is presented in copies per mL of qRT-PCR reaction. Statistical significance was set at p<0.05*. The expression of miR-34a-5p exhibited statistically significant downregulation in the group of patients compared to controls (p=0.029, < 0.05), while the two groups did not significantly differ as to the expression of miR-145-5p (p=0.068, >0.05). 1 Mean (Range); a Mann-Whitney U test; b Independent-Samples T test.
Table 2.
Absolute expression of miR-145-5p and miR-34a-5p in the group of patients that underwent thrombotic incidents and in the group of healthy controls. The expression of the under-study miRNAs is presented in copies per mL of qRT-PCR reaction. Statistical significance was set at p<0.05*. The expression of miR-34a-5p exhibited statistically significant downregulation in the group of patients compared to controls (p=0.029, < 0.05), while the two groups did not significantly differ as to the expression of miR-145-5p (p=0.068, >0.05). 1 Mean (Range); a Mann-Whitney U test; b Independent-Samples T test.
| Expression (copies/μL) 1 |
Patients (n=19) | Controls (n=19) | P-value |
|---|---|---|---|
| miR-145-5p | 1.26 x 109 (1.72 x 108 − 7.12 x 109) |
1.63 x 109 (9.90 x 107 − 4.09 x 109) |
0.068 a |
| miR-34a-5p | 9.66 x 107 (9.63 x 106 − 1.6 x 108) |
1.59 x 108 (3.59 x 107 − 3.80 x 108) |
0.029*b |
Table 3.
Comparative overview of pairwise and omnibus comparisons of miRNA expression for single variables across distinctly defined groups, showing 1 mean expression (copies/μL), a fold changes (FC, log2FC), b expressional pattern (up- or down-regulation of miRNA in the first listed group), alongside p-values for significant differences. Statistical significance was set at p<0.05*.
Table 3.
Comparative overview of pairwise and omnibus comparisons of miRNA expression for single variables across distinctly defined groups, showing 1 mean expression (copies/μL), a fold changes (FC, log2FC), b expressional pattern (up- or down-regulation of miRNA in the first listed group), alongside p-values for significant differences. Statistical significance was set at p<0.05*.
| miRNA | Expression1 (copies/μL) | FC a | Log2FC | Pattern b | P-value | ||||
| Groups | Patients | Controls | |||||||
| miR-145-5p | 1.26 x 109 | 1.63 x 109 | 0.77 | -0.38 | Down ↓ | ||||
| miR-34a-5p | 9.66 x 107 | 1.59 x 108 | 0.61 | -0.71 | Down ↓ | 0.029* | |||
| Within Thrombosis Patients | |||||||||
|
Time-Groups (post-Thrombosis) |
1-3 months |
3-6 months |
6-12 months |
12-18 months |
|||||
| miR-34a-5p | 1.27 x 108 | 1.1 x 108 | 7.0 x 107 | 9.7 x 107 | |||||
| miRNA | Expression 1 (copies/μL) | FC | Log2FC | Pattern | P-value | ||||
|
Recent Treatment Completion |
yes | no | |||||||
| miR-34a-5p | 7.0 x 107 | 1.2 x 108 | 0.62 | -0.69 | Down ↓ | 0.048* | |||
| PAI-1 4G carrier status | 4G/5G; 4G/4G | 5G/5G | |||||||
| miR-34a-5p | 1.0 x 108 | 7.8 x 107 | 1.29 | 0.37 | Down ↓ | ||||
| ACE D carrier status | D/D | I/D | |||||||
| miR-34a-5p | 1.1 x 108 | 9.2 x 107 | 1.22 | 0.29 | Up ↑ | ||||
| Type of incident | Provoked | Unprovoked | |||||||
| miR-34a-5p | 1.2 x 108 | 8.9 x 107 | 1.22 | 0.29 | Up ↑ | ||||
| Thrombosis site | cerebral | pulmonary | Lower-limbs | P-value | |||||
| miR-34a-5p | 1.3 x 108 | 5.7 x 107 | 8.1 x 107 | 0.019* | |||||
| cerebral | non-cerebral | ||||||||
| miR-34a-5p | 1.3 x 108 | 7.1 x 107 | 1.76 | 0.82 | Up ↑ | 0.007* | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



