Submitted:
01 September 2025
Posted:
02 September 2025
You are already at the latest version
Abstract
The human bladder hosts a resident, low-biomass microbial community (urobiota) that has only become subject of intense investigation in the last 15 years . The advantages that the urobiota may confer to the bladder, in contrast to the microbiota of other mucosal sites, remain to be elucidated. Alterations of the urobiota have been associated with various pathological urogenital conditions, including urinary tract infections (UTIs) and recurrent UTIs. A potential link between bladder cancer (BC), the 9th human cancer for incidence rate worldwide, and a dysbiotic urobiota is still unclear and represents an emerging field of study. The main risk factors for BC include smoking, exposure to occupational carcinogens, and schistosomiasis. In this review we focus on recent studies that not only analyzed the urobiome of BC patients using urine specimens to identify biomarkers and microbial signatures of the disease, but also to monitor therapeutic responses to therapies. We also discuss novel techniques of culturing, such as culturomics, animal models of BC, and 3D organotypic models. Furthermore, we review studies on the gut-bladder axis which, though still limited, already suggest that diet and gut-derived bacterial metabolites can influence BC progression and individual responses to therapy.
Keywords:
bladder microbiota
; bladder cancer
; gut-bladder axis
; microbial metabolites
1. Introduction
In the last decade, several studies have demonstrated the presence of a resident, specific, low-biomass, urinary tract microbiota (urobiota), thus disproving the long-lasting belief that the urinary tract above the urethra is sterile and bacteria are found only in the presence of an infection [1,2,3]. [4]The progress in our understanding of the role and composition of the urobiota is due to advancements in cultivation techniques and molecular techniques that have allowed the detection of microbial species in the urine of healthy subjects [5,6,7,8]. The urobiota has been shown to play an important role in protecting against infections and maintaining homeostasis, i.e. it supports the urinary tract epithelium, assists the immune system, and influences neurotransmission [8,9,10]. Together with the host environment, the urobiota constitutes a micro-ecological niche, the disruption of which may contribute to disease, though our understanding remains limited [9].
As for the origin of the urobiota, still many questions are open. It is suggested that a stable microbial community in the urinary tract forms within the first few years of life [11,12,13] and indeed microbial species have been detected in the urinary tract of neonates [14]. The maternal microbiome is believed to influence early microbial colonization [15] and the vaginal and gastrointestinal tracts are considered likely sources of urinary microbiota due to their anatomical proximity [16,17]. Hormonal changes during puberty may further shape the microbiota, though data on how the urobiome develops during childhood is still scarce [13,18].
As in other human body sites, bacteria benefit from the host environment, which provides nutrients, a stable temperature and a limited fluctuations in pH, which is typically mildly acidic (6.0). Yet, the full scope of the advantages that the urobiota offers to the bladder compared to that of the microbiota in other mucosal sites is still much behind and in recent years the investigations on this topic have increased [19]. Because it is well known that microbes play a key role in immune system development post-birth, similarly the urobiota may help prime the immune system and specialized immune cells in the urinary tract [9,20].
Emerging research connects microbial imbalances (dysbiosis) to a range of health issues. Spinal cord injury was associated with microbiota changes in the urinary tract of patients with/without decubitus ulcers [21], while changes in urobiota were reported in patients with lower urinary tract symptoms [22], recurrent urinary tract infections (rUTIs) [23], painful bladder syndromes, and even bladder cancer [24,25].
1.1. Effect of Gender, Age and Menopausal Changes on the Urobiome
The impact of gender and age on the urobiome (i.e. the genetic make-up of the urobiota) has been recently reviewed by others [3,19,20]. In general, in the bladder of healthy women, species belonging to the Lactobacillaceae family (i.e., Lactobacillus, Staphylococcus and Gardnerella), are by far the most abundant and are likely responsible for inhibiting uropathogens growth in the urinary tract by different mechanisms. These include the production of lactic acid which also provides a low pH environment, the impediment of the attachment of the uropathogens to the mucosal surface, the inhibition of biofilm formation, including through anti-biofilm activity, and the production of hydrogen peroxide. The bladder microbiota in men is less studied, while distal urethral, first-catch urine and urethral swabs indicate that in male the microbial composition is very similar in these specimens, regardless of collection method. The male microbiota includes Corynebacterium, Streptococcus and Lactobacillus, though the latter genus is less abundant than in women.
The alteration in the microbiome can be regarded as a secondary effect linked to the aging process. As the body undergoes various changes with age, these alterations can influence the environmental conditions of the microbiota, consequently affecting the composition and diversity of commensal microorganisms. However, in women, aging is accompanied by hormonal changes, making menopause a significant factor contributing to microbiome changes. Hormonal fluctuations in menopause can lead to environmental shifts in the vaginal microbiome, which in turn affect the urobiome. As a consequence, the urine of premenopausal women contains an abundant number of Lactobacillus, but its prevalence decreases in postmenopausal women. Conversely, abundant Mobiluncus species can be seen in postmenopausal women's urine and this shift is linked with an increased chances of UTIs and the development of rUTIs [26,27]. The decreased abundance of Lactobacillus can be regarded as the primary cause of the above infections because of the lack of their protective effects listed above.
1.2. Bladder Cancer and Influence of Microbial Factors
In the report of the International Agency for Research on Cancer of the WHO amongst the most frequently diagnosed cancers globally, bladder cancer (BC) ranks 9th for incidence rate and 13th for mortality rate. As of 2022, an estimated 614,298 new cases were reported around the world, showing a 7.1% rise compared to 2020. Moreover, recent data show that approximately 1,950,315 people of all genders are living with a BC diagnosis within the last five years, which places BC in the 7th position for prevalence. BC incidence rates are highest in Southern Europe, with countries like Greece having the highest incidence rate among men worldwide, along with Spain, Italy, Belgium, and the Netherlands in Western Europe (https://gco.iarc.who.int/today/en/dataviz/pie?mode=cancer&types=0&sexes=1&populations=900;). Indeed, when considering gender-specific data, 471,293 new BC cases in men are reported, which accounts for an age-standardized incidence rate of 9.3% of all new cancer recorded among males around the world. This puts the BC on 6th place of most diagnosed cancer in men. BC risk increases with the increase in age, and its incidence in men is almost four times higher than in women [28]. It is important to note that, approximately 75% of newly diagnosed BC patients are aged 65+ and approx. 45% are aged 75+, which includes BC as a type of cancer primarily affecting the elderly [29].
As per the guidelines of the U.S. National Comprehensive Cancer Network, BC is categorized into non-muscle-invasive bladder cancer (NMIBC) and muscle-invasive bladder cancer (MIBC) [30]. NMIBC is confined to the urothelium or lamina propria, corresponding to stages 0 and 1, while MIBC involves invasion into the muscle or deeper layers, encompassing stages 2 to 4 [31]. NMIBC shows a good prognosis, although its recurrence rate is high. In contrast to NMIBC, MIBC has a higher risk of metastasis, and its progression is linked to poor prognosis [32]. Epidemiological studies show that although men generally have a higher incidence of BC, women often present with more aggressive and advanced forms of the disease and face worse outcomes [33,34,35].
The leading risk factor for BC is smoking, mainly due to the accumulation of carcinogenic substances like aromatic amines and polycyclic aromatic hydrocarbons in the urine [36]. Such substances are present in tobacco smoke and may cause DNA damage to urothelial epithelial cells and ultimately lead to cancer development [37,38]. Other important risk factors for BC include heavy alcohol consumption and exposure to occupational carcinogens, such as benzidine, 4-chloro-o-toluidine, 4-aminobiphenyl, and 2-naphthylamine, particularly in workers from the rubber and dye industries [39]. Schistosomiasis is also a notable risk factor for BC [40].
It is estimated that globally around 20% of cancers are influenced by microbial factors and that they have a close link with the immune system and a role in tumor cell proliferation as well as in cancer metabolism [41,42,43]. Dysbiosis may decrease the presence of beneficial species involved in proper immune system functioning and in maintaining epithelial cell balance, while increasing harmful species. This imbalance can cause carcinogenesis by affecting mucosal barrier functions and even bacterial translocation to tumor sites. Although in BC the role of the microbiome is an emerging field, experimental evidence is increasing that the microbiome could be a crucial factor in susceptibility to BC [44,45]. However, analysing these findings is challenging due to the numerous factors that can directly cause change in the composition of the urinary microbiota [46]. Some inconsistent findings have been reported, related to the urobiota in BC patients. Some research shows an increased number in bacterial diversity and a differential presence of certain bacterial genera in BC patients compared to non-cancer controls [25]. Chipollini et al. [47] observed a reduction in microbial community in the BC patient’s urine and found specific taxa that were more abundant in BC patients compared to those in general clinic populations such as Faecalibacterium and Bacteroides. On the other hand, Bi et al. [48] found greater alpha diversity of the urinary microbiota in BC patients and observed more abundance of Actinomyces europaeus in these individuals. Furthermore, Bučević Popović et al. [49] did not find major differences in urobiome diversity between 12 cancer patients and 11 healthy controls but did identify difference in the abundance of specific operational taxonomic units (OTUs) between the two groups. Within BC tissues an elevated level of phylum Proteobacteria has been seen after multiple studies. Therefore, elevated levels of Proteobacteria could serve as potential markers for dysbiosis in BC diagnosis [50]. On the other hand, genera such as Ruminococcus, which are decreased in BC patients are known for their anti-inflammatory properties and their role in maintaining mucosal homeostasis [51]. A decrease in these health-promoting bacteria may create a more favorable environment for harmful bacteria, triggering inflammation and oxidative stress. The interaction between the tumor and the urobiome could involve the breakdown of normal urothelium, ultimately facilitating the attachment and proliferation of specific microbial taxa [51]. Table 1 provides a summary of studies analyzing the urobiome in BC patients vs controls. For the sake of consistency only studies where urine samples were collected and studied are reported.
In analogy to other type of cancers, the human urobiota can likely influence tumorigenesis or facilitate development of BC through different mechanisms, such as (i) direct DNA damage caused by bacterial toxins (named “genotoxins”), (ii) metabolites from microbiota that may function as potential carcinogens, (iii) bacterial-induced inflammation or biofilm formation that cause inflammation and leads to cancer promotion, and (iv) cellular microenvironment modulation by the microbiome [62,63,64]. For example, colibactin, produced by B2 Escherichia coli strains, is a toxin that causes breakage of double-stranded DNA [65]. According to Bersanelli et al. [63] the resident urinary microbiota possibly plays a concealed role in BC development. Notably, certain bacterial genera such as Acinetobacter, Stenotrophomonas, Staphylococcus, and Propionibacterium are more prevalent in BC patients as compared to healthy controls. In addition, an increased abundance of Sphingomonas spp., Acinetobacter spp., and Staphylococcus spp. can be seen in BC patients [66]. Interestingly a recent study shows that the metabolite indole-3-acetic acid, produced by the beneficial gut bacterium Parabacteroides distasonis, significantly decreased in BC patients, exerts tumor-suppressive effects by a receptor mechanism that eventually inhibits BC progression and metastasis [67].
As for inflammation, a study found increased urinary microbiome diversity and inflammatory cytokines in BC patients as well as elevated fatty acids and acylcarnitines, the latter decreasing after tumor removal [68]. Combined biomarkers from microbiome (Actinomycetaceae), metabolome (arachidonic acid), and cytokines (IL-6) showed high diagnostic accuracy and promise as a noninvasive diagnostic tool.
Pathogenic urobiota can attach to the mucosal surfaces of the bladder, leading to either continuous translocation or transient invasions of bladder tissue [69]. Many bacteria produce proteases that may act both extracellularly and intracellularly and disrupt the normal process of renewal of extracellular matrix (ECM) in the bladder. This disruption can create a modified ECM environment that can promote cancer development [69,70]. For example, the exoenzyme alkaline protease from Pseudomonas aeruginosa can degrade ECM components and impair lymphocyte proliferation by inactivating interferon gamma (IFN-ɣ) [71].
The urobiome not only may play a role in the incidence, progression, and recurrence of BC but also in treatment responses. Recent research has highlighted the role of lactic acid producing bacteria in BC involves regulating T and NK cell activities, indicating their potential relevance to immune responses [72]. For the cure of high-risk NMIBC, Bacillus Calmette-Guérin (BCG) intravesical immunotherapy is a standard treatment for this purpose [73]. However, it is not completely understood what the actual mechanism is by which microbiome of urinary tract can influence the response to BCG therapy, but it is said that it is due to the involvement of urobiome’s influence on the mucosal defensin levels. Three types of defensins, which are antimicrobial peptides, are present: human beta defensin 1 (HBD1), which is constitutively produced, and human beta defensins 2 (HBD2) and 3 (HBD3), which are induced human beta defensin. HBD1 is known to protect against BC development, while the urobiome may influence the response to BCG therapy by affecting HBD2 and HBD3 levels [61].
Contributing factors to bladder cancer
2. Culturing and Molecular Techniques to Study the Urobiota
To examine the connections between bacteria in the bladder and host health, it is important to precisely identify bacteria rapidly and at a large scale [74]. Currently, no “gold standard” has been established for the collection, preservation, and storage of urine samples for microbiome research. [75,76]. Various urine collection methods have been used in urobiome research [20] and include first-catch midstream, catheterization, and suprapubic aspiration. The latter would be more accurate to assess the microbiome composition of urine, but is invasive and not ethical in control groups. Studies suggest that there are not substantial differences in microbiome composition between catheter-based urine sample collection and suprapubic aspiration, which makes transurethral catheter-based collection more widely employed in urinary microbiome research [77,78]. Preservation and storage are also of key importance. Typically refrigeration (+4°C) is fine in the first hours after collection, but for storage longer than 24 hours freezing at -80°C is of key importance for ensuring the integrity of the microbiome [75,79]. Regardless of the collection method, Lactobacillus is commonly found in women’s urine, while men’s results vary more [80].
2.1. Culture-Dependent: Standard Urine Culture and Expanded Quantitative Urine Culture
Typically, voided urine is used in standard urine culture (SUC) technique as a diagnostic method to confirm rUTIs [81]. This aerobic protocol was mainly designed for culturing Escherichia coli to analyse the risk of pyelonephritis during pregnancy [82] and is particularly effective in culturing Enterobacteriaceae at >105 CFU/mL. As described also elsewhere [20,83], SUC is performed by inoculating 1 μL of urine onto MacConkey agar plates, then placed the plates for 24 hours at 35°C under aerobic conditions. However, a likely reason why bacteria were not detected in SUC may be assigned either to the low-biomass of the urobiota, i.e. below the culture threshold of 103 CFU/mL, or to the need of bacteria for different culturing conditions, such as incubation in an increased CO2 environment, anaerobic conditions, or extended incubation time [1]. The traditional urine culture procedures were therefore modified by 1) increasing the urine volume for plating, 2) extending the incubation time, and 3) varying the incubation atmospheric conditions. This method is called Expanded Quantitative Urine Culture (EQUC) and it is developed to detect the uropathogens that exist in low abundance, and which were unculturable previously [1,84]. For urine culture in anaerobic condition, 0.1 mL of urine can be used for inoculation onto CDC anaerobe 5% sheep blood agar (ABAP) plates, then placed under anaerobic conditions for 48 hours at 35°C [1,2,85]. Urine can also be inoculated into pre-reduced broth or Brucella blood agar plates with 5% sheep blood and vitamin K1/hemin supplementation. The plates can then be incubated in an anaerobic chamber with 85% N, 10% CO2 and 5% H2 at 37°C [86].
When bacterial counts are below 10 CFU/mL, 1.0 mL of urine can be placed in thioglycolate medium tubes and then incubated aerobically at 35°C for 5 days [2]. If bacterial growth becomes visible in the thioglycolate medium, the contents can be mixed, and a few drops can be plated on BAP and CDC Anaerobe 5% sheep blood agars for isolation. These plates can be incubated at 35°C for 48 hours under both aerobic and anaerobic conditions.
2.2. Culturomics
Culturomics is a culturing approach that employs various culture conditions, matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS), and 16S rRNA sequencing to identify bacterial species. This high-throughput culture technique initially aimed to create multiple culture environments to support the growth of fastidious bacteria, especially from the human gut. This goal was achieved by enhancing culture media with additives like blood and rumen fluid in blood culture bottles, which facilitated the growth of minority bacterial populations [87]. More recently, culturomics has integrated high-throughput sequencing and culture-dependent methods incorporating microfluidics to isolate and identify new bacterial species. This advancement led to the discovery of a new genus within the Ruminococcaceae family [88]. However, culturomics has notable limitations. It requires significant labor and cannot process as many samples as metagenomics. Despite advancements, culturomics cannot identify ‘not yet culturable’ microorganisms and does not directly yield information on gene expression or bacterial function. Functional insights require genome sequencing of newly isolated species to assess their genetic potential [89].
Culturomics has been recently applied to the study of the urobiota [90]. The extended culturomics protocol included inoculation of 0.1 mL of urine onto a large plate surface of supplemented BAP and chromogenic agar plates, which then incubation under aerobic and microaerophilic conditions at 37°C for 48 hours. BAPs can also be incubated under anaerobic conditions at 37°C for 48 h. Morphologically distinct and representative colonies, after replating, can be identified by using MALDI-TOF MS. Once (and if) the genera are identified by MALDI-TOF, they can be stored as a stock by scraping each colony from the surface of the plate and suspending into 1 mL of liquid media, with the specific media chosen based on the genus identified by MALDI-TOF. After the incubation, 1 mL of liquid culture and 1 mL 50% (v/v) glycerol are mixed to make the glycerol stocks, which can then be frozen at -80°C for later use [91].
2.3. Culture-Independent: Amplicon Sequencing and Metagenomics
Urine culture remains a routine method for detecting pathogenic microbes; however, it has limitations due to low sensitivity, making it challenging to identify all microbial species through conventional techniques [92]. This drawback is particularly notable for certain microorganisms, especially anaerobes, which may require specific nutrients or supplements and significant time to grow and thus cannot always be cultured effectively, as mentioned in 2.2. The advancements in Next-Generation Sequencing (NGS) have revolutionized molecular diagnostics, allowing researchers to quickly access extensive and diverse genetic data such as Amplicon Sequencing and Metagenome Shotgun Sequencing [20], though we should always keep in mind that sequencing does not distinguish live from dead bacteria [77].
Amplicon Sequencing, a PCR-based technique, focuses on marker genes, such as the 16S rRNA subunit, with nine hypervariable regions (V1-V9). These regions facilitate the measurement of evolutionary distances among bacterial species and provide conserved inter-regional sequences essential for primer design. This technology not only identifies bacterial species but also assesses their diversity and quantifies interrelationships within a single sample. This intra-sample diversity is referred to as alpha diversity, in contrast to beta diversity, which represents differences across multiple samples [93]. Although urinary microbiome research has advanced, 16S rRNA amplicon sequencing often limits resolution to the family or genus level, making species-level identification more challenging, especially in complex polymicrobial samples, and it cannot assess functional genes within the microbial community [94]. Furthermore, viruses, bacteriophages and fungi cannot be identified.
To analyze all the genetic material inside a (urine) sample, metagenomics, a culture-independent, high-throughput sequencing technique is used. Metagenomics enables the characterization of complete genomes within the genetic pool, achieving higher taxonomic and functional resolution. Unlike other approaches that rely on specific genetic markers, metagenomics offers an unbiased, in-depth examination of the microbiome [9]. Metagenomics can be helpful in community profiling and can improve understanding of antibiotic resistance or virulence genes associated with uropathogens [95,96,97] and provides detailed community composition analysis. Many novel uropathogens have been identified by metagenomics such as Ureaplasma, Alloscardovia, and Actinotignum, which are typically present in low abundance within the urinary tract [98]. Although sequencing the entire DNA content has clear advantages, it generally requires high sequencing depth for effective de novo assembly, especially in complex microbial communities. NGS offers significant benefits, providing a broad overview of the urinary microbiome and eliminating the need for time and labor-intensive culturing and species isolation steps. This capability has facilitated dynamic, large scale, and comprehensive analyses, allowing for the detection of microbes that are challenging to culture [1].
2.4. Animal Models and 3D Organotypic In Vitro Model
For investigating interactions between hosts and microbiota in vivo, animal models have been used as an experimental tool. However, some challenges have restricted the use of animal models such as high costs, limited reliability, and ethical concerns. Furthermore, animal models cannot fully replicate the intricate host-microbe interactions as in the human body due to variations in factors such as genetics, anatomy, diet, physiology, and life cycles of model animals and humans [99]. Despite these limitations, in the case of BC animal models N-butyl-N-(4-hydroxy butyl) nitrosamine (BBN)-induced have been established [100]. More recently, a novel mouse model of upper urinary tract urothelial carcinoma (UTUC) was obtained by treating multiple mice strains, of different sexes, with BBN: the non-engineered UTUC mouse model (only female BALB/c mice) was demonstrated to reflect human UTUC in many molecular aspects [101]. Notably, the low relative abundance of P. distasonis in the gut was reverted with a dietary intervention, which consisted mainly in removing alanine: the suppression of the TNF-related inflammatory gene expression in the upper urinary tract was observed even in the presence of BBN. The relative abundance of P. distasonis and its link, when reduced, to urothelial carcinoma likely links to the decreased levels of its metabolite indole-3-acetic acid [67], as described in section 1.2
As for in vitro studies, these often depend on primary cells (two-dimensional or 2D cell culture) and monolayer cultures of cell lines to investigate tissue function and disease pathogenesis, including infections, cancer and metastasis. However, these 2D and monolayer cultures have limitations in capturing the intricate influence of the stromal environment, which is crucial for the development of diseases. Three-dimensional (3D) organotypic models offer a promising approach to gain a deeper understanding of molecular disease development, moving closer to in vivo conditions [102,103]. An array of 3D organotypic models has been established, including to study the interaction with bacteria and microbiome [104,105]. Also in the urological field of cancer, organoid technology is witnessing significant advancements [106] and 3D bio-printed in vitro models of bladder and urethra have been developed [107,108]. However, literature is still scarce in this area of research.
3. Gut-Bladder Axis in Bladder Cancer
The interplay between the gut and the bladder microbiota (the so-called “gut-bladder axis”) is still an emerging field of research, including the study of the implication of gut microbiota (GM) and dysbiosis as etiological factor in BC. As mentioned in section 1.2, BC is a cancer that more commonly affects a population aged 65+, that, because of aging, is more at risk of dysbiosis in the gut and in the bladder.
A two-sample Mendelian randomization (MR) analysis, predicted that the genus Bifidobacterium, the phylum and class Actinobacteria and the Ruminococcus torques group in GM were associated with an increased risk of BC, while Allisonella was associated with a decreased risk [109]. Another two-sample Mendelian randomization study showed a significant causal relationship between Bilophila and BC, while Oscillibacter, Ruminococcaceae NK4A214 group and Enterobacteriales were protective against BC [110]. Whether the observed associations can be assigned to H2S production by Bilophila (a known genotoxic compound) and SCFAs production by the protective species still remain to be demonstrated. A Mendelian randomization study of GM showed that the family Pasteurellaceae, Eubacterium coprostanoligenes group and the order Pasteurellales increase significantly the risk of BC. Conversely, the genus Escherichia Shigella was associated with a decreased risk [111].
In one of the first studies where the urinary microbiota and GM of patients with BC were compared with a healthy control group, class Alphaproteobacteria, order Rhodospirillales, order Flavobacteriales, and family Flavobacteriaceae were specific only in female with BC [61]. A difference in the abundance of phylum Desulfobacterota across tumor grade was also observed, i.e. most abundant in Grade G1 and least in G2. However the study had limitations, i.e. small groups with diverse history (sociodemographic, BCG therapy, smoking), single-centered study, collection of urine by midstream[61] .
A study involving n. 142 NIMBC patients and n. 48 controls demonstrated that the GM in BC patients had a higher prevalence of Prevotella and Porphyromonas but a reduced abundance of Faecalibacterium compared with controls [112]. The alpha diversity indices in the BC group increased, while no statistically significant differences in abundances of the above bacteria were observed between genders. In the same study the differences in composition of the GM between patients who did not respond to the neoadjuvant chemotherapy (n. 57) vs controls showed higher levels of Bacteroides and Pseudomonas compared to the controls, while Lachnospirenaceae (producers of SCFAs) were more abundant in responders [112]. A more direct association between a specific bacterium P. distasonis and BC was provided in a study where the GM of n. 50 BC patients when compared to that of n. 22 matched controls: the genus Parabacteroides was more abundant in the controls [113]. Notably, in the same study it was shown that the combination of P. distasonis and anti-PD-1 monoclonal antibody significantly inhibited tumor growth and reduced its weight in bladder tumor-bearing mice, compared to treatment with anti-PD-1 antibody alone. Immunohistochemistry and RNA-sequencing showed that P. distasonis promoted anti-tumor immune responses by potentiating the efficacy of anti-PD-1 therapy through immune activation and modulation of tumor-related signaling pathways [113].
While the objective of this section was mostly on potential microbial players of the GM on the development of BC, the role of inflammation should also be taken into account. This topic is investigated [68] and has been recently reviewed [9,114].
Figure 1 provides an graphical overview of the factors contributing to BC, dealt with in this and previous sections, and in the next coming section.
4. Microbial-Derived Metabolites and Bladder Health
Our understanding of the gut-bladder axis with respect to the effect of microbial metabolites is still limited, i.e. the connection between GM, gut and/or bladder metabolites of microbial origin, and BC is still embryonic. This is confirmed by recent reviews dealing with the impact of dietary metabolites, such as SCFAs (important in regulating the function of the epithelial as well as mucosal barrier and systemic immunity), hardly mentioning the bladder in their analysis [115,116,117]. Given that a good level of understanding is available on SCFAs and this topic has been covered by the above studies, herein we focus on other metabolites that may potentially affect bladder health especially in BC. However, it is important here to mention that low levels of butyrate have been detected in the BC patients [118]. Butyrate helps in the proliferation of intestinal epithelial cells and helps in maintaining the mucosal integrity: its reduced levels compromise the epithelial homeostasis, ultimately diminishing its inhibitory effects on BC progression. The inhibitory effect is mediated by the host G-protein coupled receptors. Furthermore, butyrate can enter cells via host plasma membrane transporters and inhibits histone deacetylases inside the host cells, which leads to histone hyperacetylation and changes in gene expression, eventually leading to lowering the tumor cell proliferation and increased apoptosis [117]. It is well known that, SCFAs are by-products of fermentation, mostly produced in the colon, by GM members that degrade and retrieve energy from undigested dietary fibers [115]. Diet can therefore provide a strategic approach to prevent or treat BC patients. This is the hypothesis that guided the work by Then et al. [119] who investigated the ability of high-fiber diets to sensitize BC to irradiation. They used different dietary regimens at the same time in which mouse bladder tumor cells were inoculated subcutaneously in allograft immunoproficient mouse model; these dietary regimens included either normal chow or 0.2% cellulose, or psyllium, or psyllium + resistant starch (RS, known to contain fibers that produce butyrate), or psyllium + inulin (a fiber readily fermentable and known as radiosensitiser). Psyllum is a dietary fiber used to reduce the side effects of radiotherapy [120]. Psyllium + RS as well as psyllium + inulin significantly delayed tumor growth compared control group (i.e., 0.2% cellulose). Furthermore, when bladder tumors were irradiated psyllium + RS significantly radiosensitised the tumors compared to 0.2% cellulose and psyllium alone. The study also showed that the GM modified by the dietary fiber regimen might be required to activate immune responses at systemic level. Overall, the study confirms that the use of dietary fibers by increasing SCFAs affects the outcome of the bladder tumors. This is an effect of the shaping of the GM in favor of an increase of the relative abundance of the Lachnospiraceae family, and of Bacteroides genus abundance [119].
Amongst the microbial-derived metabolites, we should distinguish those that originate from the GM and then reach the bladder from those that are produced inside the bladder. Herein, we review some very recent reports on both aspects.
Amongst metabolites of GM origin, the metabolite 3-IAA, mentioned in sections 1.2. and 2.4 above, has been recently analyzed in great detail by Li et al. [67]. In addition to demonstrate that P. distasonis increased abundance in GM is associated with an improved prognosis in BC patients, the authors provided evidence that specifically the metabolite 3-IAA derived from P. distasonis has a tumor suppressive effect and inhibited BC cells migration in a dose-dependent manner. Given that 3-IAA is a metabolite originating from the amino acid tryptophan, the derivatives of which are known to bind the AhR receptor, activated in tumor-associated macrophages, the study demonstrated first of all that 3-IAA binds the AhR and, secondly, that 3-IAA exerts its tumor-suppressive effect by activating AhR (as demonstrated by knocking down the AhR gene). The intracellular effect consists in a reduced expression of fatty acid synthase and stearoyl-CoA desaturase. The former enzyme in particular is highly expressed in BC patients, who show a worse overall survival [67]. As a consequence of fatty acid synthase decreased expression, the incorporation of polyunsaturated fatty acids is membrane phospholipids increases and this is ultimately causing an increase of cellular sensitivity to ferroptosis.
On the other hand, some metabolites originate from the metabolism of molecules that are recognized as environmental and occupational carcinogens and represent major risk factors in BC (see section 1.2). Amongst these are N-butyl-N- (4-hydroxybutyl)-nitrosamine (BBN), a nitrosamine compound, as well as other organic contaminants related to it, that have been described to induce BC [121,122]. Using a mouse model, Roje et al. [122] provide evidence that GM may enhance the development of BC in BBN-exposed mice and that a decreased bacterial load has a positive effect, i.e. the reduced metabolism by the GM of BBN into BCPN, its oxidation product, leads to decrease in concentration of BCPN in the urine, from which it is eliminated, thereby decreasing the urothelial exposure to this carcinogen by altering it pharmacokinetics [122]. The identification of specific genera in both mouse and in human GM suggested that interpersonal differences could play a role in the individual predisposition to tumor development.
Xenobiotics transformation can be carried out not only by the GM, but also by the microbiota in other body sites, including by bladder. This is extremely important when we consider studies that have demonstrated that, out of 212 pharmaceuticals only one third is excreted through the feces, with the majority through urine and in some cases, i.e. paracetamol, acetylsalicylic acid, gabapentin (an antiepileptic drug) excretion is almost entirely through the urine [123]. Furthermore, xenobiotics are exposed in the gut to the GM, which is much more dense than the urobiota and also the exposure to xenobiotics in the gut is much longer (55 hours) than in the bladder (4 hours). Also, oxygen levels are different in the two niches. Using a genome mining approach, Marti et al. [124] analyzed the distribution of enzyme classes in the urobiota, in particular from genomes of bacteria isolated using EQUC from women samples [83]. The analysis indicated that the distribution of enzyme classes is discontinuous even within species in the same genus and that urinary Gordonia and Bacilli have a broad biotransformation potential. Amongst the EC classes, the oxidoreductases (in particular, NADP-dependent oxidoreductases/dihydropyrimidine dehydrogenases EC 1.3), transferases (EC 2) and amidases, amongst the hydrolases (EC 3), were the most represented in the urinary bacterial genomes.
5. Conclusions and Future Directions
The study of the urobiota and the urobiome in the urine of BC patients is a field of merging interest, especially in the last 15 years since the long lasting belief about urine sterility above the urethra has been demonstrated to be untrue. We focused on studies, summarized in Table 1, that analyzed the urobiome using urine specimens from BC group vs control (i.e. healthy subjects) in the last 10 years. While Actinobacteria Bacteroidetes, Firmicutes and Proteobacteria were found as the most abundant phyla in both groups, and in most of the studies, no clear signature of BC or healthy state has been demonstrated to date. This is because the studies were limited in size of recruited patients/controls, urine collection mode, gender, stage of BC, and lifestyle. A good knowledge of the microbial metabolism and enzymatic activities that take place in the bladder are extremely relevant also in terms of protection from cancer.
Notably, two recent studies [67,113] demonstrate the therapeutic potential of P. distasonis, a producer in the gut of 3-IAA, a metabolite of the amino acid tryptophan, that has a tumor suppressive effect and inhibited BC cells migration in a dose-dependent manner. Furthermore, when P. distasonis is delivered in combination with alpha-PD-1 mAb, it is effective in improving the effect of anti-PD-1 immunotherapy, likely by activating immune and antitumor related pathways. The two findings might be linked.
The observed decrease in beneficial microbial metabolites, particularly butyrate, in BC patients underlines a potentially important role of SCFAs in bladder mucosal immunity, epithelial integrity, and tumor suppression. Exploring microbiota-based SCFA restoration therapies may open unique pathways for BC prevention as well as for noninvasive therapy. The study by Then et al. [119] supports the approach of high-fiber diets in radiotherapy. In general, diet-based interventions and probiotic supplementation, is a field of great promise.
The study of relation between the bladder microbiome and the gut microbiome is still preliminary and requires comprehensive effort to uncover the full impact of these hidden and overlooked microbial communities. As for the case of P. distasonis, research in this field continues to evolve, it will open doors to potential therapeutic interventions, diagnostic biomarkers, and a deeper understanding of BC treatment and recurrence.
Author Contributions
The authors contributed equally to the conception of the review manuscript; UAB wrote the first draft of the manuscript; DDB revised the manuscript. Both authors agreed on the content of the final version of the manuscript.
Funding
This work was partially supported by grants from Sapienza University of Rome, MA120172B88445A7 and RD12318AAABE3A08.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 2D | Two-dimensional |
| 3D | Three-dimensional |
| ABAP | Anaerobe 5% sheep blood agar |
| BAP | Sheep blood agar plate |
| BC | Bladder Cancer |
| BCG | Bacillus Calmette-Guérin |
| ECM | Extracellular matrix |
| EQUC | Expanded Quantitative Urine Culture |
| GM | Gut Microbiota |
| MALDI-TOF MS | Matrix Assisted Laser Desorption Ionization-Time of Flight Mass |
| MIBC | Muscle-Invasive Bladder Cancer |
| NGS | Next-Generation Sequencing |
| NMIBC | Non-Muscle-Invasive Bladder Cancer |
| rUTIs | Recurrent urinary tract infections |
| SCFAs | Short-chain fatty acids |
| SUC | Standard urine culture |
| UTIs | Urinary tract infections |
| UTUC | urinary tract urothelial carcinoma |
References
- Hilt, E.E.; McKinley, K.; Pearce, M.M.; Rosenfeld, A.B.; Zilliox, M.J.; Mueller, E.R.; Brubaker, L.; Gai, X.; Wolfe, A.J.; Schreckenberger, P.C. Urine is not sterile: use of enhanced urine culture techniques to detect resident bacterial flora in the adult female bladder. Journal of clinical microbiology 2014, 52, 871-876. [CrossRef]
- Pearce, M.M.; Hilt, E.E.; Rosenfeld, A.B.; Zilliox, M.J.; Thomas-White, K.; Fok, C.; Kliethermes, S.; Schreckenberger, P.C.; Brubaker, L.; Gai, X.; et al. The female urinary microbiome: a comparison of women with and without urgency urinary incontinence. mBio 2014, 5, e01283-01214. [CrossRef]
- Roth, R.S.; Liden, M.; Huttner, A. The urobiome in men and women: a clinical review. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases 2023, 29, 1242-1248. [CrossRef]
- Aad, G.; Abbott, B.; Abbott, D.C.; Abed Abud, A.; Abeling, K.; Abhayasinghe, D.K.; Abidi, S.H.; AbouZeid, O.S.; Abraham, N.L.; Abramowicz, H.; et al. Measurement of the Lund Jet Plane Using Charged Particles in 13 TeV Proton-Proton Collisions with the ATLAS Detector. Physical review letters 2020, 124, 222002. [CrossRef]
- Lewis, D.A.; Brown, R.; Williams, J.; White, P.; Jacobson, S.K.; Marchesi, J.R.; Drake, M.J. The human urinary microbiome; bacterial DNA in voided urine of asymptomatic adults. Frontiers in cellular and infection microbiology 2013, 3, 41. [CrossRef]
- Brubaker, L.; Wolfe, A.J. The new world of the urinary microbiota in women. American journal of obstetrics and gynecology 2015, 213, 644-649. [CrossRef]
- Siddiqui, H.; Nederbragt, A.J.; Lagesen, K.; Jeansson, S.L.; Jakobsen, K.S. Assessing diversity of the female urine microbiota by high throughput sequencing of 16S rDNA amplicons. BMC microbiology 2011, 11, 244. [CrossRef]
- Ackerman, A.L.; Chai, T.C. The Bladder is Not Sterile: an Update on the Urinary Microbiome. Current bladder dysfunction reports 2019, 14, 331-341. [CrossRef]
- Jones-Freeman, B.; Chonwerawong, M.; Marcelino, V.R.; Deshpande, A.V.; Forster, S.C.; Starkey, M.R. The microbiome and host mucosal interactions in urinary tract diseases. Mucosal immunology 2021, 14, 779-792. [CrossRef]
- Zhang, J.; Lei, Y.; Du, H.; Li, Z.; Wang, X.; Yang, D.; Gao, F.; Li, J. Exploring urinary microbiome: insights into neurogenic bladder and improving management of urinary tract infections. Frontiers in cellular and infection microbiology 2025, 15, 1512891. [CrossRef]
- Storm, D.W.; Copp, H.L.; Halverson, T.M.; Du, J.; Juhr, D.; Wolfe, A.J. A Child's urine is not sterile: A pilot study evaluating the Pediatric Urinary Microbiome. Journal of pediatric urology 2022, 18, 383-392. [CrossRef]
- Hadjifrangiskou, M.; Reasoner, S.; Flores, V.; Van Horn, G.; Morales, G.; Peard, L.; Abelson, B.; Manuel, C.; Lee, J.; Baker, B.; et al. Defining the Infant Male Urobiome and Moving Towards Mechanisms in Urobiome Research. Research square 2023. [CrossRef]
- Kelly, M.S.; Dahl, E.M.; Jeries, L.M.; Sysoeva, T.A.; Karstens, L. Characterization of pediatric urinary microbiome at species-level resolution indicates variation due to sex, age, and urologic history. Journal of pediatric urology 2024, 20, 884-893. [CrossRef]
- Kinneman, L.; Zhu, W.; Wong, W.S.W.; Clemency, N.; Provenzano, M.; Vilboux, T.; Jane't, K.; Seo-Mayer, P.; Levorson, R.; Kou, M.; et al. Assessment of the Urinary Microbiome in Children Younger Than 48 Months. The Pediatric infectious disease journal 2020, 39, 565-570. [CrossRef]
- Robertson, R.C.; Manges, A.R.; Finlay, B.B.; Prendergast, A.J. The Human Microbiome and Child Growth - First 1000 Days and Beyond. Trends in microbiology 2019, 27, 131-147. [CrossRef]
- Komesu, Y.M.; Dinwiddie, D.L.; Richter, H.E.; Lukacz, E.S.; Sung, V.W.; Siddiqui, N.Y.; Zyczynski, H.M.; Ridgeway, B.; Rogers, R.G.; Arya, L.A.; et al. Defining the relationship between vaginal and urinary microbiomes. American journal of obstetrics and gynecology 2020, 222, 154 e151-154 e110. [CrossRef]
- Dubourg, G.; Morand, A.; Mekhalif, F.; Godefroy, R.; Corthier, A.; Yacouba, A.; Diakite, A.; Cornu, F.; Cresci, M.; Brahimi, S.; et al. Deciphering the Urinary Microbiota Repertoire by Culturomics Reveals Mostly Anaerobic Bacteria From the Gut. Frontiers in microbiology 2020, 11, 513305. [CrossRef]
- Jeries, L.M.; Sysoeva, T.A.; Karstens, L.; Kelly, M.S. Synthesis of current pediatric urinary microbiome research. Frontiers in pediatrics 2024, 12, 1396408. [CrossRef]
- Colella, M.; Topi, S.; Palmirotta, R.; D'Agostino, D.; Charitos, I.A.; Lovero, R.; Santacroce, L. An Overview of the Microbiota of the Human Urinary Tract in Health and Disease: Current Issues and Perspectives. Life 2023, 13. [CrossRef]
- Perez-Carrasco, V.; Soriano-Lerma, A.; Soriano, M.; Gutierrez-Fernandez, J.; Garcia-Salcedo, J.A. Urinary Microbiome: Yin and Yang of the Urinary Tract. Frontiers in cellular and infection microbiology 2021, 11, 617002. [CrossRef]
- Morsli, M.; Salipante, F.; Gelis, A.; Magnan, C.; Guigon, G.; Lavigne, J.P.; Sotto, A.; Dunyach-Remy, C. Evolution of the urinary microbiota in spinal cord injury patients with decubitus ulcer: A snapshot study. International wound journal 2024, 21, e14626. [CrossRef]
- Bajic, P.; Van Kuiken, M.E.; Burge, B.K.; Kirshenbaum, E.J.; Joyce, C.J.; Wolfe, A.J.; Branch, J.D.; Bresler, L.; Farooq, A.V. Male Bladder Microbiome Relates to Lower Urinary Tract Symptoms. European urology focus 2020, 6, 376-382. [CrossRef]
- Burnett, L.A.; Hochstedler, B.R.; Weldon, K.; Wolfe, A.J.; Brubaker, L. Recurrent urinary tract infection: Association of clinical profiles with urobiome composition in women. Neurourology and urodynamics 2021, 40, 1479-1489. [CrossRef]
- Stone, L. Urine microbiota differ in bladder cancer. Nature reviews. Urology 2023, 20, 7. [CrossRef]
- Wu, P.; Zhang, G.; Zhao, J.; Chen, J.; Chen, Y.; Huang, W.; Zhong, J.; Zeng, J. Profiling the Urinary Microbiota in Male Patients With Bladder Cancer in China. Frontiers in cellular and infection microbiology 2018, 8, 167. [CrossRef]
- Curtiss, N.; Balachandran, A.; Krska, L.; Peppiatt-Wildman, C.; Wildman, S.; Duckett, J. Age, menopausal status and the bladder microbiome. European journal of obstetrics, gynecology, and reproductive biology 2018, 228, 126-129. [CrossRef]
- Stapleton, A.E. Urine Culture in Uncomplicated UTI: Interpretation and Significance. Current infectious disease reports 2016, 18, 15. [CrossRef]
- Saginala, K.; Barsouk, A.; Aluru, J.S.; Rawla, P.; Padala, S.A.; Barsouk, A. Epidemiology of Bladder Cancer. Medical sciences 2020, 8. [CrossRef]
- Martin, A.; Woolbright, B.L.; Umar, S.; Ingersoll, M.A.; Taylor, J.A., 3rd. Bladder cancer, inflammageing and microbiomes. Nature reviews. Urology 2022, 19, 495-509. [CrossRef]
- Chang, S.S.; Boorjian, S.A.; Chou, R.; Clark, P.E.; Daneshmand, S.; Konety, B.R.; Pruthi, R.; Quale, D.Z.; Ritch, C.R.; Seigne, J.D.; et al. Diagnosis and Treatment of Non-Muscle Invasive Bladder Cancer: AUA/SUO Guideline. The Journal of urology 2016, 196, 1021-1029. [CrossRef]
- Lenis, A.T.; Lec, P.M.; Chamie, K.; Mshs, M.D. Bladder Cancer: A Review. Jama 2020, 324, 1980-1991. [CrossRef]
- Grabe-Heyne, K.; Henne, C.; Mariappan, P.; Geiges, G.; Pohlmann, J.; Pollock, R.F. Intermediate and high-risk non-muscle-invasive bladder cancer: an overview of epidemiology, burden, and unmet needs. Frontiers in oncology 2023, 13, 1170124. [CrossRef]
- Fernandez-Gomez, J.; Solsona, E.; Unda, M.; Martinez-Pineiro, L.; Gonzalez, M.; Hernandez, R.; Madero, R.; Ojea, A.; Pertusa, C.; Rodriguez-Molina, J.; et al. Prognostic factors in patients with non-muscle-invasive bladder cancer treated with bacillus Calmette-Guerin: multivariate analysis of data from four randomized CUETO trials. European urology 2008, 53, 992-1001. [CrossRef]
- Ingersoll, M.A.; Li, X.; Inman, B.A.; Greiner, J.W.; Black, P.C.; Adam, R.M. Immunology, Immunotherapy, and Translating Basic Science into the Clinic for Bladder Cancer. Bladder cancer 2018, 4, 429-440. [CrossRef]
- Koti, M.; Ingersoll, M.A.; Gupta, S.; Lam, C.M.; Li, X.; Kamat, A.M.; Black, P.C.; Siemens, D.R. Sex Differences in Bladder Cancer Immunobiology and Outcomes: A Collaborative Review with Implications for Treatment. European urology oncology 2020, 3, 622-630. [CrossRef]
- Burger, M.; Catto, J.W.; Dalbagni, G.; Grossman, H.B.; Herr, H.; Karakiewicz, P.; Kassouf, W.; Kiemeney, L.A.; La Vecchia, C.; Shariat, S.; et al. Epidemiology and risk factors of urothelial bladder cancer. European urology 2013, 63, 234-241. [CrossRef]
- Babjuk, M. Re: Oncological Benefit of Re-resection for T1 Bladder Cancer: A Comparative Effectiveness Study. European urology 2023, 83, 297. [CrossRef]
- Lammers, R.J.; Witjes, W.P.; Hendricksen, K.; Caris, C.T.; Janzing-Pastors, M.H.; Witjes, J.A. Smoking status is a risk factor for recurrence after transurethral resection of non-muscle-invasive bladder cancer. European urology 2011, 60, 713-720. [CrossRef]
- Kiriluk, K.J.; Prasad, S.M.; Patel, A.R.; Steinberg, G.D.; Smith, N.D. Bladder cancer risk from occupational and environmental exposures. Urologic oncology 2012, 30, 199-211. [CrossRef]
- Mostafa, M.H.; Sheweita, S.A.; O'Connor, P.J. Relationship between schistosomiasis and bladder cancer. Clinical microbiology reviews 1999, 12, 97-111. [CrossRef]
- Cao, Y.; Xia, H.; Tan, X.; Shi, C.; Ma, Y.; Meng, D.; Zhou, M.; Lv, Z.; Wang, S.; Jin, Y. Intratumoural microbiota: a new frontier in cancer development and therapy. Signal transduction and targeted therapy 2024, 9, 15. [CrossRef]
- Vogtmann, E.; Goedert, J.J. Epidemiologic studies of the human microbiome and cancer. British journal of cancer 2016, 114, 237-242. [CrossRef]
- Rajagopala, S.V.; Vashee, S.; Oldfield, L.M.; Suzuki, Y.; Venter, J.C.; Telenti, A.; Nelson, K.E. The Human Microbiome and Cancer. Cancer prevention research 2017, 10, 226-234. [CrossRef]
- Aragon-Ching, J.B.; Werntz, R.P.; Zietman, A.L.; Steinberg, G.D. Multidisciplinary Management of Muscle-Invasive Bladder Cancer: Current Challenges and Future Directions. American Society of Clinical Oncology educational book. American Society of Clinical Oncology. Annual Meeting 2018, 38, 307-318. [CrossRef]
- Zeng, J.; Zhang, G.; Chen, C.; Li, K.; Wen, Y.; Zhao, J.; Wu, P. Alterations in Urobiome in Patients With Bladder Cancer and Implications for Clinical Outcome: A Single-Institution Study. Frontiers in cellular and infection microbiology 2020, 10, 555508. [CrossRef]
- Yacouba, A.; Tidjani Alou, M.; Lagier, J.C.; Dubourg, G.; Raoult, D. Urinary microbiota and bladder cancer: A systematic review and a focus on uropathogens. Seminars in cancer biology 2022, 86, 875-884. [CrossRef]
- Chipollini, J.; Wright, J.R.; Nwanosike, H.; Kepler, C.Y.; Batai, K.; Lee, B.R.; Spiess, P.E.; Stewart, D.B.; Lamendella, R. Characterization of urinary microbiome in patients with bladder cancer: Results from a single-institution, feasibility study. Urologic oncology 2020, 38, 615-621. [CrossRef]
- Bi, H.; Tian, Y.; Song, C.; Li, J.; Liu, T.; Chen, Z.; Chen, C.; Huang, Y.; Zhang, Y. Urinary microbiota - a potential biomarker and therapeutic target for bladder cancer. Journal of medical microbiology 2019, 68, 1471-1478. [CrossRef]
- Bucevic Popovic, V.; Situm, M.; Chow, C.T.; Chan, L.S.; Roje, B.; Terzic, J. The urinary microbiome associated with bladder cancer. Scientific reports 2018, 8, 12157. [CrossRef]
- Friedrich, V.; Choi, H.W. The Urinary Microbiome: Role in Bladder Cancer and Treatment. Diagnostics 2022, 12. [CrossRef]
- Oresta, B.; Braga, D.; Lazzeri, M.; Frego, N.; Saita, A.; Faccani, C.; Fasulo, V.; Colombo, P.; Guazzoni, G.; Hurle, R.; et al. The Microbiome of Catheter Collected Urine in Males with Bladder Cancer According to Disease Stage. The Journal of urology 2021, 205, 86-93. [CrossRef]
- Xu, W.; Yang, L.; Lee, P.; Huang, W.C.; Nossa, C.; Ma, Y.; Deng, F.M.; Zhou, M.; Melamed, J.; Pei, Z. Mini-review: perspective of the microbiome in the pathogenesis of urothelial carcinoma. American journal of clinical and experimental urology 2014, 2, 57-61.
- Mai, G.; Chen, L.; Li, R.; Liu, Q.; Zhang, H.; Ma, Y. Common Core Bacterial Biomarkers of Bladder Cancer Based on Multiple Datasets. BioMed research international 2019, 2019, 4824909. [CrossRef]
- Moynihan, M.; Sullivan, T.; Provenzano, K.; Rieger-Christ, K. Urinary Microbiome Evaluation in Patients Presenting with Hematuria with a Focus on Exposure to Tobacco Smoke. Research and reports in urology 2019, 11, 359-367. [CrossRef]
- Mansour, B.; Monyok, A.; Makra, N.; Gajdacs, M.; Vadnay, I.; Ligeti, B.; Juhasz, J.; Szabo, D.; Ostorhazi, E. Bladder cancer-related microbiota: examining differences in urine and tissue samples. Scientific reports 2020, 10, 11042. [CrossRef]
- Pederzoli, F.; Ferrarese, R.; Amato, V.; Locatelli, I.; Alchera, E.; Luciano, R.; Nebuloni, M.; Briganti, A.; Gallina, A.; Colombo, R.; et al. Sex-specific Alterations in the Urinary and Tissue Microbiome in Therapy-naive Urothelial Bladder Cancer Patients. European urology oncology 2020, 3, 784-788. [CrossRef]
- Hourigan, S.K.; Zhu, W.; W, S.W.W.; Clemency, N.C.; Provenzano, M.; Vilboux, T.; Niederhuber, J.E.; Deeken, J.; Chung, S.; McDaniel-Wiley, K.; et al. Studying the urine microbiome in superficial bladder cancer: samples obtained by midstream voiding versus cystoscopy. BMC urology 2020, 20, 5. [CrossRef]
- Hussein, A.A.; Elsayed, A.S.; Durrani, M.; Jing, Z.; Iqbal, U.; Gomez, E.C.; Singh, P.K.; Liu, S.; Smith, G.; Tang, L.; et al. Investigating the association between the urinary microbiome and bladder cancer: An exploratory study. Urologic oncology 2021, 39, 370 e379-370 e319. [CrossRef]
- Ma, W.; Zhang, W.; Shen, L.; Liu, J.; Yang, F.; Maskey, N.; Wang, H.; Zhang, J.; Yan, Y.; Yao, X. Can Smoking Cause Differences in Urine Microbiome in Male Patients With Bladder Cancer? A Retrospective Study. Frontiers in oncology 2021, 11, 677605. [CrossRef]
- Qiu, J.; Liu, J.; Zhong, Y.; Liu, W.; Zhou, Z.; Li, Y.; Li, S. Analysis of Urinary Flora Characteristics in Urinary Tumor Based on 16S rRNA Sequence. BioMed research international 2022, 2022, 9368687. [CrossRef]
- Chorbinska, J.; Krajewski, W.; Nowak, L.; Bardowska, K.; Zebrowska-Rozanska, P.; Laczmanski, L.; Pacyga-Prus, K.; Gorska, S.; Malkiewicz, B.; Szydelko, T. Is the Urinary and Gut Microbiome Associated With Bladder Cancer? Clinical Medicine Insights. Oncology 2023, 17, 11795549231206796. [CrossRef]
- Poore, G.D.; Kopylova, E.; Zhu, Q.; Carpenter, C.; Fraraccio, S.; Wandro, S.; Kosciolek, T.; Janssen, S.; Metcalf, J.; Song, S.J.; et al. Microbiome analyses of blood and tissues suggest cancer diagnostic approach. Nature 2020, 579, 567-574. [CrossRef]
- Bersanelli, M.; Santoni, M.; Ticinesi, A.; Buti, S. The Urinary Microbiome and Anticancer Immunotherapy: The Potentially Hidden Role of Unculturable Microbes. Targeted oncology 2019, 14, 247-252. [CrossRef]
- Sepich-Poore, G.D.; Carter, H.; Knight, R. Intratumoral bacteria generate a new class of therapeutically relevant tumor antigens in melanoma. Cancer cell 2021, 39, 601-603. [CrossRef]
- Grasso, F.; Frisan, T. Bacterial Genotoxins: Merging the DNA Damage Response into Infection Biology. Biomolecules 2015, 5, 1762-1782. [CrossRef]
- Li, N.; Wang, L.; Yang, Q.; Li, F.; Shi, Z.; Feng, X.; Zhang, L.; Li, X.; Jin, X.; Zhu, S.; et al. Identification and Evaluation of the Urinary Microbiota Associated With Bladder Cancer. Cancer innovation 2025, 4, e70012. [CrossRef]
- Li, W.; Shangguan, W.; Huang, W.; Zhao, J.; Zhu, Y.; Xie, M.; Yu, Y.; Yang, Q.; Zheng, J.; Yang, L.; et al. Gut Parabacteroides distasonis-derived Indole-3-Acetic Acid Promotes Phospholipid Remodeling and Enhances Ferroptosis Sensitivity via the AhR-FASN Axis in Bladder Cancer. Advanced science 2025, e04688. [CrossRef]
- Wu, X.; Zou, W.; Liu, Z. Causal relationships between gut microbiota and urothelial carcinoma mediated by inflammatory cytokines and blood cell traits identified through Mendelian randomization analysis. Discover oncology 2025, 16, 1440. [CrossRef]
- Alfano, M.; Canducci, F.; Nebuloni, M.; Clementi, M.; Montorsi, F.; Salonia, A. The interplay of extracellular matrix and microbiome in urothelial bladder cancer. Nature reviews. Urology 2016, 13, 77-90. [CrossRef]
- Vollmer, P.; Walev, I.; Rose-John, S.; Bhakdi, S. Novel pathogenic mechanism of microbial metalloproteinases: liberation of membrane-anchored molecules in biologically active form exemplified by studies with the human interleukin-6 receptor. Infection and immunity 1996, 64, 3646-3651. [CrossRef]
- Horvat, R.T.; Parmely, M.J. Pseudomonas aeruginosa alkaline protease degrades human gamma interferon and inhibits its bioactivity. Infection and immunity 1988, 56, 2925-2932. [CrossRef]
- Min, K.; Kim, H.T.; Lee, E.H.; Park, H.; Ha, Y.S. Bacteria for Treatment: Microbiome in Bladder Cancer. Biomedicines 2022, 10. [CrossRef]
- Jiang, S.; Redelman-Sidi, G. BCG in Bladder Cancer Immunotherapy. Cancers 2022, 14. [CrossRef]
- Hoffman, C.; Siddiqui, N.Y.; Fields, I.; Gregory, W.T.; Simon, H.M.; Mooney, M.A.; Wolfe, A.J.; Karstens, L. Species-Level Resolution of Female Bladder Microbiota from 16S rRNA Amplicon Sequencing. mSystems 2021, 6, e0051821. [CrossRef]
- Jung, C.E.; Chopyk, J.; Shin, J.H.; Lukacz, E.S.; Brubaker, L.; Schwanemann, L.K.; Knight, R.; Wolfe, A.J.; Pride, D.T. Benchmarking urine storage and collection conditions for evaluating the female urinary microbiome. Scientific reports 2019, 9, 13409. [CrossRef]
- Bundgaard-Nielsen, C.; Ammitzboll, N.; Isse, Y.A.; Muqtar, A.; Jensen, A.M.; Leutscher, P.D.C.; Arenholt, L.T.S.; Hagstrom, S.; Sorensen, S. Voided Urinary Microbiota Is Stable Over Time but Impacted by Post Void Storage. Frontiers in cellular and infection microbiology 2020, 10, 435. [CrossRef]
- Aragon, I.M.; Herrera-Imbroda, B.; Queipo-Ortuno, M.I.; Castillo, E.; Del Moral, J.S.; Gomez-Millan, J.; Yucel, G.; Lara, M.F. The Urinary Tract Microbiome in Health and Disease. European urology focus 2018, 4, 128-138. [CrossRef]
- Wolfe, A.J.; Toh, E.; Shibata, N.; Rong, R.; Kenton, K.; Fitzgerald, M.; Mueller, E.R.; Schreckenberger, P.; Dong, Q.; Nelson, D.E.; et al. Evidence of uncultivated bacteria in the adult female bladder. Journal of clinical microbiology 2012, 50, 1376-1383. [CrossRef]
- Ng, H.H.; Ang, H.C.; Hoe, S.Y.; Lim, M.L.; Tai, H.E.; Soh, R.C.H.; Syn, C.K. Simple DNA extraction of urine samples: Effects of storage temperature and storage time. Forensic science international 2018, 287, 36-39. [CrossRef]
- Bae, S.; Chung, H. The Urobiome and Its Role in Overactive Bladder. International neurourology journal 2022, 26, 190-200. [CrossRef]
- Gaitonde, S.; Malik, R.D.; Zimmern, P.E. Financial Burden of Recurrent Urinary Tract Infections in Women: A Time-driven Activity-based Cost Analysis. Urology 2019, 128, 47-54. [CrossRef]
- Kass, E.H. Bacteriuria and the diagnosis of infections of the urinary tract; with observations on the use of methionine as a urinary antiseptic. A.M.A. archives of internal medicine 1957, 100, 709-714. [CrossRef]
- Thomas-White, K.; Forster, S.C.; Kumar, N.; Van Kuiken, M.; Putonti, C.; Stares, M.D.; Hilt, E.E.; Price, T.K.; Wolfe, A.J.; Lawley, T.D. Culturing of female bladder bacteria reveals an interconnected urogenital microbiota. Nature communications 2018, 9, 1557. [CrossRef]
- Price, T.K.; Dune, T.; Hilt, E.E.; Thomas-White, K.J.; Kliethermes, S.; Brincat, C.; Brubaker, L.; Wolfe, A.J.; Mueller, E.R.; Schreckenberger, P.C. The Clinical Urine Culture: Enhanced Techniques Improve Detection of Clinically Relevant Microorganisms. Journal of clinical microbiology 2016, 54, 1216-1222. [CrossRef]
- Price, T.K.; Wolff, B.; Halverson, T.; Limeira, R.; Brubaker, L.; Dong, Q.; Mueller, E.R.; Wolfe, A.J. Temporal Dynamics of the Adult Female Lower Urinary Tract Microbiota. mBio 2020, 11. [CrossRef]
- Hurst, R.; Meader, E.; Gihawi, A.; Rallapalli, G.; Clark, J.; Kay, G.L.; Webb, M.; Manley, K.; Curley, H.; Walker, H.; et al. Microbiomes of Urine and the Prostate Are Linked to Human Prostate Cancer Risk Groups. European urology oncology 2022, 5, 412-419. [CrossRef]
- Lagier, J.C.; Hugon, P.; Khelaifia, S.; Fournier, P.E.; La Scola, B.; Raoult, D. The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota. Clinical microbiology reviews 2015, 28, 237-264. [CrossRef]
- Ma, L.; Kim, J.; Hatzenpichler, R.; Karymov, M.A.; Hubert, N.; Hanan, I.M.; Chang, E.B.; Ismagilov, R.F. Gene-targeted microfluidic cultivation validated by isolation of a gut bacterium listed in Human Microbiome Project's Most Wanted taxa. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, 9768-9773. [CrossRef]
- Lagier, J.C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Levasseur, A.; Rolain, J.M.; Fournier, P.E.; Raoult, D. Culturing the human microbiota and culturomics. Nature reviews. Microbiology 2018, 16, 540-550. [CrossRef]
- Ugarcina Perovic, S.; Ksiezarek, M.; Rocha, J.; Cappelli, E.A.; Sousa, M.; Ribeiro, T.G.; Grosso, F.; Peixe, L. Urinary Microbiome of Reproductive-Age Asymptomatic European Women. Microbiology spectrum 2022, 10, e0130822. [CrossRef]
- Baddoo, G.; Ene, A.; Merchant, Z.; Banerjee, S.; Wolfe, A.J.; Putonti, C. Cataloging variation in 16S rRNA gene sequences of female urobiome bacteria. Frontiers in urology 2023, 3, 1270509. [CrossRef]
- Heytens, S.; De Sutter, A.; Coorevits, L.; Cools, P.; Boelens, J.; Van Simaey, L.; Christiaens, T.; Vaneechoutte, M.; Claeys, G. Women with symptoms of a urinary tract infection but a negative urine culture: PCR-based quantification of Escherichia coli suggests infection in most cases. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases 2017, 23, 647-652. [CrossRef]
- Goodrich, J.K.; Di Rienzi, S.C.; Poole, A.C.; Koren, O.; Walters, W.A.; Caporaso, J.G.; Knight, R.; Ley, R.E. Conducting a microbiome study. Cell 2014, 158, 250-262. [CrossRef]
- Deurenberg, R.H.; Bathoorn, E.; Chlebowicz, M.A.; Couto, N.; Ferdous, M.; Garcia-Cobos, S.; Kooistra-Smid, A.M.; Raangs, E.C.; Rosema, S.; Veloo, A.C.; et al. Application of next generation sequencing in clinical microbiology and infection prevention. Journal of biotechnology 2017, 243, 16-24. [CrossRef]
- Barraud, O.; Ravry, C.; Francois, B.; Daix, T.; Ploy, M.C.; Vignon, P. Shotgun metagenomics for microbiome and resistome detection in septic patients with urinary tract infection. International journal of antimicrobial agents 2019, 54, 803-808. [CrossRef]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Moller, N.; Aarestrup, F.M. Rapid whole-genome sequencing for detection and characterization of microorganisms directly from clinical samples. Journal of clinical microbiology 2014, 52, 139-146. [CrossRef]
- Mulder, H.A.; Lee, S.H.; Clark, S.; Hayes, B.J.; van der Werf, J.H.J. The Impact of Genomic and Traditional Selection on the Contribution of Mutational Variance to Long-Term Selection Response and Genetic Variance. Genetics 2019, 213, 361-378. [CrossRef]
- Moustafa, A.; Li, W.; Singh, H.; Moncera, K.J.; Torralba, M.G.; Yu, Y.; Manuel, O.; Biggs, W.; Venter, J.C.; Nelson, K.E.; et al. Microbial metagenome of urinary tract infection. Scientific reports 2018, 8, 4333. [CrossRef]
- Nguyen, T.L.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Disease models & mechanisms 2015, 8, 1-16. [CrossRef]
- Fantini, D.; Glaser, A.P.; Rimar, K.J.; Wang, Y.; Schipma, M.; Varghese, N.; Rademaker, A.; Behdad, A.; Yellapa, A.; Yu, Y.; et al. A Carcinogen-induced mouse model recapitulates the molecular alterations of human muscle invasive bladder cancer. Oncogene 2018, 37, 1911-1925. [CrossRef]
- Yamamoto, A.; Kawashima, A.; Uemura, T.; Nakano, K.; Matsushita, M.; Ishizuya, Y.; Jingushi, K.; Hase, H.; Katayama, K.; Yamaguchi, R.; et al. A novel mouse model of upper tract urothelial carcinoma highlights the impact of dietary intervention on gut microbiota and carcinogenesis prevention despite carcinogen exposure. International journal of cancer 2025, 156, 1439-1456. [CrossRef]
- Kim, R. Advanced Organotypic In Vitro Model Systems for Host-Microbial Coculture. Biochip journal 2023, 1-27. [CrossRef]
- Shin, K. Stem cells, organoids and their applications for human diseases: Special issue of BMB Reports in 2023. BMB reports 2023, 56, 1. [CrossRef]
- Charles, C.A.; Ricotti, C.A.; Davis, S.C.; Mertz, P.M.; Kirsner, R.S. Use of tissue-engineered skin to study in vitro biofilm development. Dermatologic surgery : official publication for American Society for Dermatologic Surgery [et al.] 2009, 35, 1334-1341. [CrossRef]
- Taebnia, N.; Romling, U.; Lauschke, V.M. In vitro and ex vivo modeling of enteric bacterial infections. Gut microbes 2023, 15, 2158034. [CrossRef]
- Wang, X.; Lin, D.; Feng, N. Harnessing organoid technology in urological cancer: advances and applications in urinary system tumors. World journal of surgical oncology 2025, 23, 295. [CrossRef]
- Chae, S.; Kim, J.; Yi, H.G.; Cho, D.W. 3D Bioprinting of an In Vitro Model of a Biomimetic Urinary Bladder with a Contract-Release System. Micromachines 2022, 13. [CrossRef]
- Zhang, K.; Fu, Q.; Yoo, J.; Chen, X.; Chandra, P.; Mo, X.; Song, L.; Atala, A.; Zhao, W. 3D bioprinting of urethra with PCL/PLCL blend and dual autologous cells in fibrin hydrogel: An in vitro evaluation of biomimetic mechanical property and cell growth environment. Acta biomaterialia 2017, 50, 154-164. [CrossRef]
- Mingdong, W.; Xiang, G.; Yongjun, Q.; Mingshuai, W.; Hao, P. Causal associations between gut microbiota and urological tumors: a two-sample mendelian randomization study. BMC cancer 2023, 23, 854. [CrossRef]
- Yang, H.; Jin, C.; Li, J.; Zhang, Z.; Zhao, K.; Yin, X.; Wang, Z.; Zhu, G.; Yan, X.; Jiang, Z.; et al. Causal relationship between bladder cancer and gut microbiota contributes to the gut-bladder axis: A two-sample Mendelian randomization study. Urologic oncology 2025, 43, 267 e269-267 e218. [CrossRef]
- Wang, Y.; Liu, W.; Liu, L.; He, Y.; Luo, H.; Fang, C. Causal effect of gut microbiota on the risk of cancer and potential mediation by inflammatory proteins. World journal of surgical oncology 2025, 23, 163. [CrossRef]
- Bukavina, L.; Ginwala, R.; Eltoukhi, M.; Sindhani, M.; Prunty, M.; Geynisman, D.M.; Ghatalia, P.; Valentine, H.; Calaway, A.; Correa, A.F.; et al. Role of Gut Microbiome in Neoadjuvant Chemotherapy Response in Urothelial Carcinoma: A Multi-institutional Prospective Cohort Evaluation. Cancer research communications 2024, 4, 1505-1516. [CrossRef]
- Wang, B.; Qiu, Y.; Xie, M.; Huang, P.; Yu, Y.; Sun, Q.; Shangguan, W.; Li, W.; Zhu, Z.; Xue, J.; et al. Gut microbiota Parabacteroides distasonis enchances the efficacy of immunotherapy for bladder cancer by activating anti-tumor immune responses. BMC microbiology 2024, 24, 237. [CrossRef]
- Zou, J.; Xu, B.; Luo, P.; Chen, T.; Duan, H. Non-coding RNAs in bladder cancer, a bridge between gut microbiota and host? Frontiers in immunology 2024, 15, 1482765. [CrossRef]
- Mann, E.R.; Lam, Y.K.; Uhlig, H.H. Short-chain fatty acids: linking diet, the microbiome and immunity. Nature reviews. Immunology 2024, 24, 577-595. [CrossRef]
- Thomas, R.M. Microbial molecules, metabolites, and malignancy. Neoplasia 2025, 60, 101128. [CrossRef]
- Tsvetikova, S.A.; Koshel, E.I. Microbiota and cancer: host cellular mechanisms activated by gut microbial metabolites. International journal of medical microbiology : IJMM 2020, 310, 151425. [CrossRef]
- He, C.; Li, B.; Huang, L.; Teng, C.; Bao, Y.; Ren, M.; Shan, Y. Gut microbial composition changes in bladder cancer patients: A case-control study in Harbin, China. Asia Pacific journal of clinical nutrition 2020, 29, 395-403. [CrossRef]
- Then, C.K.; Paillas, S.; Moomin, A.; Misheva, M.D.; Moir, R.A.; Hay, S.M.; Bremner, D.; Roberts Nee Nellany, K.S.; Smith, E.E.; Heidari, Z.; et al. Dietary fibre supplementation enhances radiotherapy tumour control and alleviates intestinal radiation toxicity. Microbiome 2024, 12, 89. [CrossRef]
- Then, C.K.; Paillas, S.; Wang, X.; Hampson, A.; Kiltie, A.E. Association of Bacteroides acidifaciens relative abundance with high-fibre diet-associated radiosensitisation. BMC biology 2020, 18, 102. [CrossRef]
- He, Z.; Kosinska, W.; Zhao, Z.L.; Wu, X.R.; Guttenplan, J.B. Tissue-specific mutagenesis by N-butyl-N-(4-hydroxybutyl)nitrosamine as the basis for urothelial carcinogenesis. Mutation research 2012, 742, 92-95. [CrossRef]
- Roje, B.; Zhang, B.; Mastrorilli, E.; Kovacic, A.; Susak, L.; Ljubenkov, I.; Cosic, E.; Vilovic, K.; Mestrovic, A.; Vukovac, E.L.; et al. Gut microbiota carcinogen metabolism causes distal tissue tumours. Nature 2024, 632, 1137-1144. [CrossRef]
- Lienert, J.; Burki, T.; Escher, B.I. Reducing micropollutants with source control: substance flow analysis of 212 pharmaceuticals in faeces and urine. Water science and technology : a journal of the International Association on Water Pollution Research 2007, 56, 87-96. [CrossRef]
- Marti, T.D.; Scharer, M.R.; Robinson, S.L. Microbial Biocatalysis within Us: The Underexplored Xenobiotic Biotransformation Potential of the Urinary Tract Microbiota. Chimia 2023, 77, 424-431. [CrossRef]
Figure 1.
Contributing factors to bladder cancer. Created in BioRender. De Biase, D. (2025) https://BioRender.com/17jtfaq.
Figure 1.
Contributing factors to bladder cancer. Created in BioRender. De Biase, D. (2025) https://BioRender.com/17jtfaq.
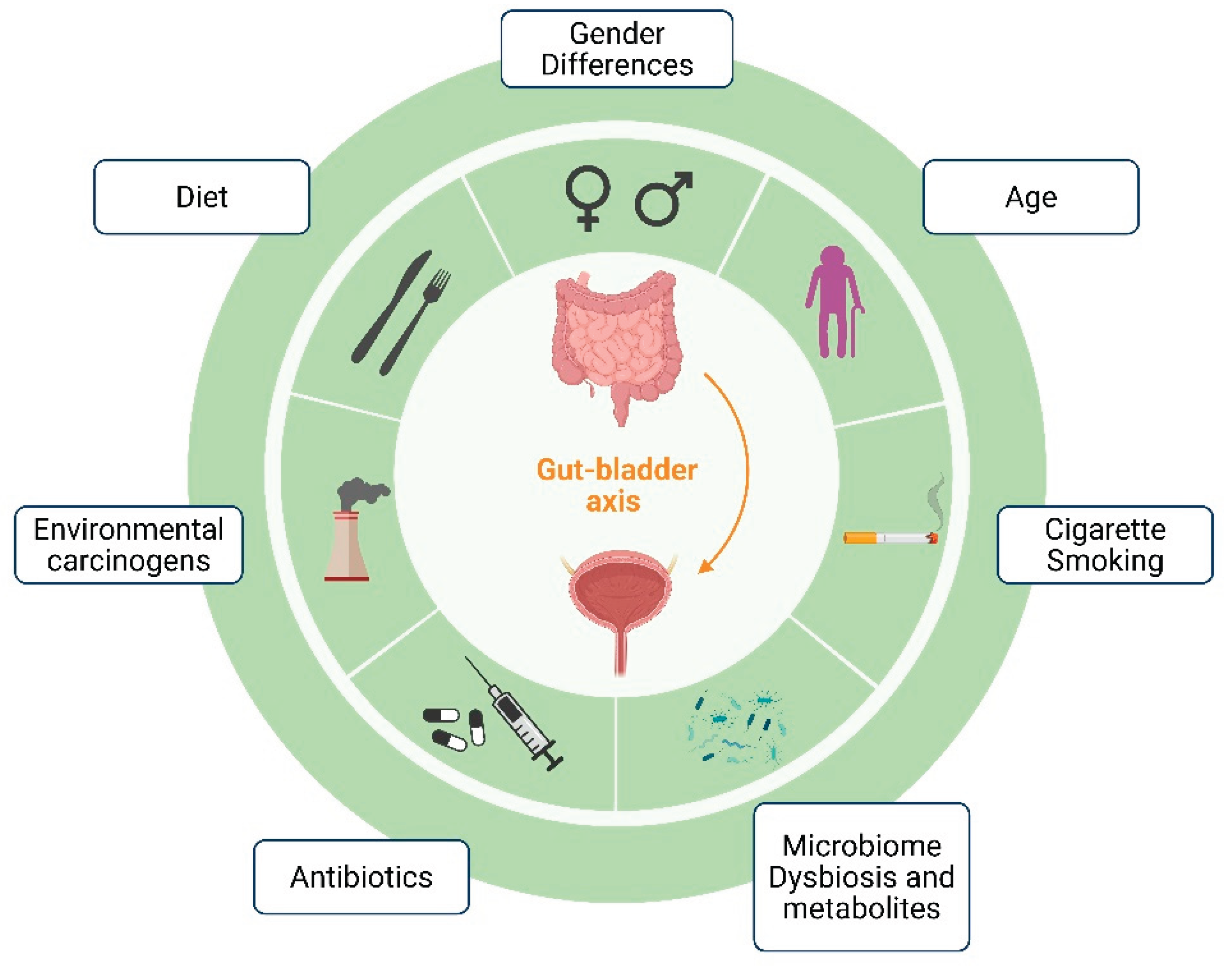

Table 1.
Summary of urobiome studies using urine specimens in BC patients.
| Study | Sample size (Cancer/Healthy) |
Cohorts and Diversity | Abundance and key findings |
|---|---|---|---|
| Xu et al. [52] | 8/6 |
|
|
| Bucevic Popovic et al. [49] | 12/11 |
|
|
| Wu et al. [25] | 31/18 |
|
|
| Bi et al. [48] | 29/26 |
|
|
| Mai et al. [53] | 24/0 |
|
|
| Moynihan et al. [54] | 35/8 |
|
|
| Mansour et al. [55] | 10/0 |
|
|
| Pederzoli et al. [56] | 49/59 |
|
|
| Zeng et al. [45] | 62/19 |
|
|
| Chipollini et al. [47] | 38/10 |
|
|
| Hourigan et al. [57] | 22/0 |
|
|
| Hussein et al. [58] | 43/10 |
|
|
| Ma et al. [59] | 15/11 |
|
|
| Oresta et al. [51] | 51/10 |
|
|
| Qiu et al. [60] | 6/4 |
|
|
| Chorbinska et al. [61] | 18/7 |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



