
Submitted:
25 August 2025
Posted:
26 August 2025
You are already at the latest version
Abstract
In this work, the activity of extracts from S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 was evaluated against in vitro and in silico models related to leukemia. The cytotoxicity of extracts was analyzed by the crystal violet assay, whereas their effect on apoptosis was evaluated through flow cytometry approaches. The effect of treatment on the DNA fragmentation of representative leukemia cell lines was determined by electrophoresis. In silico modeling consisted of assessing the binding affinities between major components from the obtained extracts against CCR2. Results revealed that extracts from Sechium spp. can decrease the viability of J774 and P388 cell lines upon exposure to IC50 0.93-1.38 mg/mL. It was noted that extract from S. edule var. nigrum spinosum exhibited the highest capacity to induce the late apoptosis of J774 cells, whereas treatment with extract from S. chinantlense exerted the highest capability to cause early apoptosis of P388 cells. It was also determined that extracts caused the DNA fragmentation of the cell lines tested in this study after 68 h exposure to treatment. In silico evaluation evidenced that metabolites from extracts poses high affinity for chemokine-type receptors involved in the initiation and progression of leukemia exhibiting binding energies ranging from -7.89 to -9.49 kcal/mol. The retrieved evidence demonstrated the therapeutic capacity of Sechium spp. against leukemia considering in vitro and in silico mechanisms of action.
Keywords:
traditional medicine
; cancer research
; Sechium spp.
; leukemia
; anticancer mechanisms
1. Introduction
Leukemia is a malignant hematological disorder characterized by the abnormal proliferation of immature white blood cells within the bone marrow and peripheral blood. It is broadly categorized into four primary types: acute lymphoblastic leukemia (ALL), acute myeloid leukemia (AML), chronic lymphocytic leukemia (CLL), and chronic myeloid leukemia (CML) [1]. Risk factors for leukemia include genetic predisposition, exposure to ionizing radiation, certain chemical agents such as benzene, and prior chemotherapy treatment. Although leukemia can affect individuals across all age groups, specific types are more prevalent in certain age demographics [2]; for example, ALL is more common in children [3], whereas CLL predominantly affects older adults [4]. Treatment regimens typically involve a combination of chemotherapy, targeted therapy, immunotherapy [5], and stem cell transplantation, tailored to the specific leukemia type, disease stage, and patient characteristics [6].
Molecularly, there are various receptors involved in the initiation and progression of leukemia and its subtypes. For instance, the C-C chemokine receptor type 2 (CCR2) is a G protein-coupled receptor (GPCR) with a crucial role in the immune system by mediating the migration and activation of monocytes and other immune cells [7]. Even though CCR2 is primarily expressed on various immune cell types (e.g., monocytes and T dendritic cells) to orchestrate inflammatory phenomena, recent scientific evidence has suggested that, when it binds to ligands (e.g., CCL2, CCL7, or CCL13), it can initiate intracellular signaling cascades that influence cell behavior, including migration, proliferation, and cytokine production [8]. In leukemia, the role of CCR2 is multifaceted since it has been associated with progression and metastasis events, together with facilitation of tumor growth[9], immune evasion, and modulation of signaling pathways such as the NF-κB/MAPK pathway [10].
Despite notable advancements in the treatment of leukemia, current methodologies encounter several limitations that impede their efficacy and patient outcomes. Chemotherapy, although effective in inducing remission, frequently results in severe side effects and long-term complications due to its non-specific nature, impacting both malignant and healthy cells [11]. The development of drug resistance remains a significant challenge, particularly in relapsed or refractory cases, necessitating the exploration of alternative therapeutic strategies [12]. Targeted therapies, while more specific, are constrained by the heterogeneity of leukemic cells and the rapid emergence of resistance mechanisms [13]. Stem cell transplantation, although potentially curative, is associated with considerable risks, including graft-versus-host disease and opportunistic infections, and is not suitable for all patients due to age restrictions or the lack of suitable donors [14]. Furthermore, the high cost and limited accessibility of novel therapies, such as CAR T-cell therapy, present substantial barriers to their widespread implementation [15]. These limitations underscore the urgent need for more effective, less toxic, and more accessible treatment approaches in the management of leukemia.
Traditional medicine has increasingly attracted attention in cancer treatment research, offering potential complementary approaches to conventional therapies [16]. Various herbal remedies and natural compounds have demonstrated promising anti-cancer properties in preclinical studies, targeting multiple cellular pathways involved in cancer progression [16]. For instance, certain plant-derived compounds have shown the ability to induce apoptosis in cancer cells, inhibit angiogenesis, and modulate the immune response, together with signaling pathways involved in cancer progression [17]. Evaluating these mechanisms, as well as potential material genetic damage, and modulation of signaling pathways, is crucial when evaluating plant extracts to provide critical insights into their potential efficacy and capacity to participate in drug design and development [18].
Sechium spp., belonging to the Cucurbitaceae family, encompasses economically and culturally significant species. The genus Sechium is native to Mesoamerica and has been cultivated for centuries, playing a crucial role in traditional cuisines and medicinal practices [19]. In traditional medicine, species from the genus Sechium are valued for their edible fruits, leaves, and roots, contributing to food security and agricultural diversity in many regions [20]. Recent research has highlighted the potential of Sechium spp. in cancer treatment due to their phytochemical content. The phytochemical content of Sechium spp. is based, predominantly, on flavonoids, phenolic acids, and cucurbitacins. Against in vitro and in vivo cancer-based models, the presence of such compounds has been reported in extracts from S. chinantlense, which has been reported to exhibit antiproliferative and pro-apoptotic effects on cell lines representative of cervical cancer [21]. Similar results have been associated with extracts from S. edule var. Madre Negra, which has been reported to decrease the viability of breast cancer cells [22]. The application of extracts from Sechium spp. has been reported in only two studies, which evaluated the activity of the methanol extract of Sechium edule var. nigrum spinosum against P388 and mononuclear bone marrow cells [23]. Another study considered a similar approach, but evaluated a Sechium spp. hybrid which was designated as H-837-07-GISeM® [24].
Given the importance of extracts from Sechium spp. in preventing, managing, and treating various types of cancer, this study aimed to investigate the effect of polar extracts from other varieties or hybrids, such as S. compositum, S. chinantlense, and the hybrid H-387-07, against representative cancer cell lines of leukemia. The activity of an extract from S. edule var. nigrum spinosum was also included to compare the retrieved results. Cytotoxicity assays were performed by the crystal violet assay, which was also utilized to determine the half-maximal inhibitory concentration (IC50). Flow cytometry was implemented to unveil the capacity of extracts of leading leukemia cell lines to death by apoptosis. Electrophoresis was also considered to determine the capability of extracts to cause DNA fragmentation among the cultured cell lines. In silico modeling was utilized to identify the affinity of major constituents of extracts towards CCR2.
2. Materials and Methods
2.1. Extract Obtention and Phytochemical Composition Analyses
The extracts from S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 were obtained, processed, and phytochemically characterized as published [23,25,26]. Briefly, fruits were cut and dried in a RIOSSA E-33 convection oven (Modena, Italy) until a constant weight was achieved. After this, they were powdered, macerated with methanol for 48 h, and filtered with cellulose filter paper. The solvent was removed utilizing a Buchi rotavapor R-114 (Flawil, Switzerland) at 50 °C. The phytochemical content of extracts S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 was determined by high-performance liquid chromatography (HPLC) and assessed in previous reports by Grupo Interdisciplinario de Sechium edule en Mexico (GISeM).
2.2. Experimental Animals
CD-1 mice (male and female), aged 10 to 12 weeks, were kept in the animal facility at Facultad de Estudios Superiores (FES) Zaragoza from Universidad Nacional de México. Male and female mice received a sterile standard powdered rodent diet available ad libitum. All experimental procedures were approved by the Ethics Committee of FES-Zaragoza and conducted following the "Guide for the Care and Use of Laboratory Animals, Eighth Edition" from the National Institutes of Health, which is considered for research methodologies and evaluation protocols involving traditional medicine. Additionally, experimental procedures were executed considering the national regulations for the care and use of experimental animals (NOM-062-ZOO-1999).
2.3. Cell Lines Culture
The J774 and P388 cell lines were cultured with Iscove's Modified Dulbeco's Medium (IMDM) (Gibco BRL, NY, USA) supplemented with 10% deactivated fetal bovine serum (FBS) (Gibco BRL, NY, USA) at 37°C in a humidified atmosphere supplemented with 5% CO2. The cell density was maintained at 1x105 cells/mL for P388 and 5x104 cells/mL for J774. The same protocol was implemented for culturing the cells extracted from the bone marrow of CD-1 mice. For obtaining the bone marrow mononuclear cells from CD-1 mice, the femurs were harvested, and total bone marrow cells were collected by flowing IMDM (Gibco BRL, USA) supplemented with 10% FBS (Gibco BRL, USA). After this, cells were then separated using a density gradient (ficol density = 1.077 g/mL) (Sigma, St. Louis, USA). The cells were cultured in IMDM supplemented with 15% FBS (Gibco BRL, USA), 5% equine serum, and recombinant mouse interleukin-3 at a density of 100,000 cells/mL in 5-mL plates (Corning, NY, USA).
2.4. Cytotoxicity Assay
The effect of extracts from S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 on the viability of the J774 and P388 cell lines was evaluated using the crystal violet assay. Briefly, 1x105 P388 cells/mL and 5x104 J774 cells/mL were seeded onto 96-well plates (Corning Costar, St. Louis, MO, USA). After this, cells were fixed with glutaraldehyde (1.1%) and stained with crystal violet solution (0.1%). The excess of the latter was removed by washing the cells with distilled water, and it was solubilized with an acetic acid solution (10%). The absorbance of wells was recorded utilizing a Tecan Spectra microplate reader (Grödig, Austria) at 570 nm. The IC50 of each extract was assessed according to the percentage of dead cells and the tested concentrations.
2.5. Early Apoptosis Analysis
The P388 and J774 cell lines were cultured with IMDM (Gibco BRL, NY, USA) supplemented with 10% deactivated FBS (Gibco BRL, NY, USA) at 37°C in a humidified atmosphere supplemented with 5% CO2. The cell density was maintained at 1x105 cells/mL for P388 and 5x104 cells/mL for J774. The effect of treatment with extracts from species of the genus Sechium was evaluated when P388 and J774 reached an average of 70% saturation. The tested extract concentrations consisted of their half-maximal inhibitory values (IC50). The tested concentrations of S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 against the J774 cell line were 0.93, 1.96, and 0.26 μg/mL, respectively. The evaluated concentrations of extracts against the P388 cell line were 1.18, 0.98, 0.98, and 1.38 μg/mL, respectively. The effect of treatment on the early apoptosis of J774 and P388 cell lines was evaluated using the Annexin V Kit (BD pharmagen™ PE Annexin V Apoptosis Detection Kit). Briefly, cells were washed with phosphate buffer solution, and cell counting was performed. After this, Annexin V buffer was then added. These cells were removed and placed in a 5 mL Falcon tube. Fluorescein isothiocyanate (FITC) was then added, which is positive for cells undergoing early apoptosis and cells undergoing late apoptosis, or cells that were dead, either due to apoptosis or necrosis. Then, they were incubated for 15 minutes at room temperature in the dark. Annexin V buffer was then added, and the cells were placed in the dark and on ice. Finally, they were analyzed by flow cytometry on a cytometer (BD FACSAria II) within one hour.
2.6. Oligonucleosomal DNA Fragmentation Analysis
The effect of extracts from S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 in the DNA fragmentation of P388 and J774 cell lines, and bone marrow mononuclear cells from CD-1 mice was evaluated, considering the mentioned IC50 values in section 2.3 at 48, 60, and 68 h. Briefly, Cells were collected by conventional centrifugation at 1,500 rpm. Then, 600 µL of lysis buffer and 20 µL of the stock solution (4 mg/mL) of proteinase K (Promega, Madison, WI, USA) were added, and the cells were incubated at 37°C for 4 h. Next, 7.5 µL of RNAse (Sigma-Aldrich, USA) was added, and the cells were incubated for one hour at 37°C. After this, 0.9 mL of 3M sodium acetate (pH 5.2) was added, followed by 0.3 mL of chloroform-phenol-isoamyl alcohol, and the pellet was centrifuged at 18,000 rpm at 4°C. The upper (aqueous) phase was then transferred to a new Eppendorf tube containing 600 µL of isopropanol (Sigma-Aldrich, USA) at 4°C. The tube was centrifuged, and the supernatant was decanted. 30 µL of TE buffer was added to hydrate the pellet. The DNA obtained was analyzed on a 1.5% UltraPure Agarose gel (Invitrogen) and stained with 40 µL of the stock (0.5 µg/mL) of ethidium bromide (Sigma-Aldrich) in an electrophoresis chamber (Gibco BRL, Horizon 58) and analyzed using a transilluminator (Foto/Eclipse, FotoDyne).
2.7. Antagonistic Effect Analysis
The possible antagonistic activity of gallic acid, quercetin, and cucurbitacin B on the receptor CCR2 was investigated to determine their influence on modulating inflammatory phenomena, pro-inflammatory cytokine release, apoptosis regulation, and cell proliferation. Briefly, the chemical structures of gallic acid (CID: 4678042), cucurbitacin B (CID: 5281316), and quercetin (CID: 5280343) were obtained from Pubchem (https://pubchem.ncbi.nlm.nih.gov) in 2D SDF format. The crystal structure of CCR2 was obtained from the RCSB protein database (www.rcsb.org) corresponding to the PDB ID: 6GPS, originally used in the research to modulate the response. Subsequently, the ligand charges were minimized using the MMFF94 algorithm in Avogadro at a pH of 7.3. Then, the Autodock 4.2 program was used to prepare the protein by eliminating waters, assigning polar hydrogens, obtaining native ligand coordinates, and adding Kollman charges. The spatial coordinates of the native ligand MK-0812 (F7N) were 8.27 (center_x), -10.42 (center_y), -12.43 (center_z), 56 (size_x), 54 (size_y), and 50 (size_z). The ligands of interest were oriented to the same orthosteric site, performing 10 iterations for each ligand to calculate binding energies, average standard deviation, normality tests, homoscedasticity, group comparison, and post hoc tests.
2.8. Statistical Analysis
The data retrieved from in vitro and in silico experiments were analyzed utilizing a one-way analysis of variance (ANOVA) followed by Tukey’s mean separation test.
3. Result
3.1. Cytotoxicity Assay
The results obtained from the application of the extracts showed a reduction in cell density in the mouse myeloid leukemic lines P388 and J774 (observed under a 10x light microscope) after 68 and 60 hours, respectively, compared to the positive control and the vehicle, in the presence of the IC50, recording a reduction in cell density close to that induced by the commercial antineoplastic agent Ara-C (Cytarabine®) (see Figure 1 and Figure 2). The results obtained after using the highest concentrations of the plant extracts on normal mouse bone marrow cells showed that the proliferation of normal mouse bone marrow mononuclear cells was reduced, especially in Ara-c® and S. edule var. nigrum spinosum (Figure 3A,B). In the specific cases of H-387-07, S. compositum, and S. chinantlense (see Figure 3D,F,G), cell density was observed to be very similar to that of the controls (Figure 3A,B), which could indicate that these extracts do not significantly affect the proliferation of normal cells. Regarding S. edule var. nigrum spinosum (see Figure 3E), although it was observed to reduce cell density in normal cells, it did not reach the levels of reduction observed in the leukemic lines observed in Figure 1E and Figure 2 E.
3.2. Flow Cytometry Assay
It was observed that control and vehicle cells clustered in quadrant Q3, indicating the absence of PS translocation, which is interpreted as viable cells (untreated). With the addition of the experimental IC50 of each plant extract, the population migrated to quadrant Q4 and Q2, representing PS translocation, indicative of early or late apoptosis, respectively (see Figure 3 and Figure 4D,E,F,G). Regarding the J774 and P388 cell lines under S. edule var. nigrum spinosum conditions, it was observed that the PS-positive cell population migrated to the Q2 region, indicating late apoptosis. One possible explanation for this is the reduced cell density, since the extract from this genotype had the highest concentration. Therefore, the damage caused to the cells was much more severe. Another point worth highlighting is that the data obtained by flow cytometry indicated that at least 50% of the total population in both cell lines treated with the extracts was undergoing either early (Q4) or late (Q2) apoptosis (see Table 2 and Table 3).
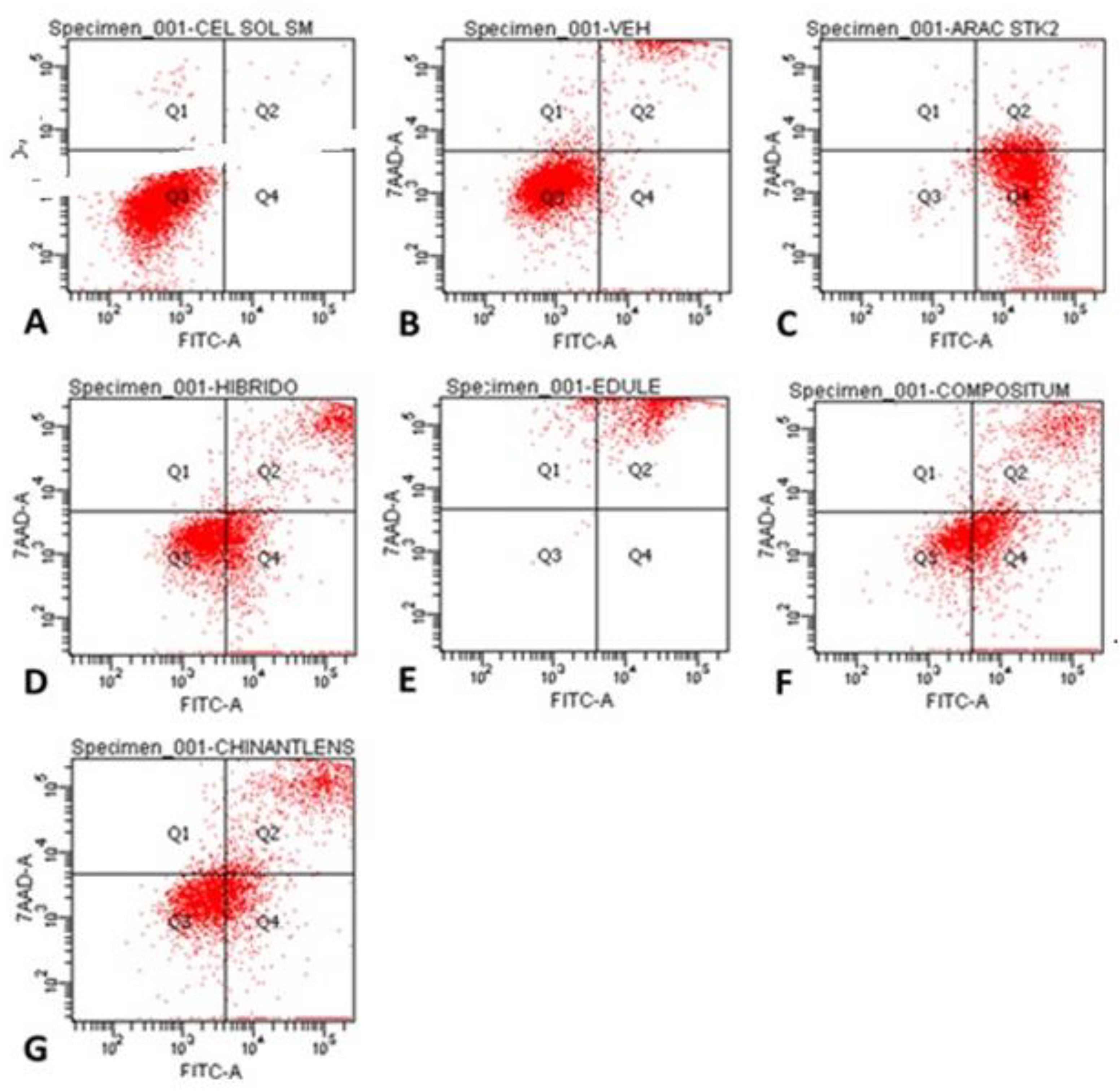
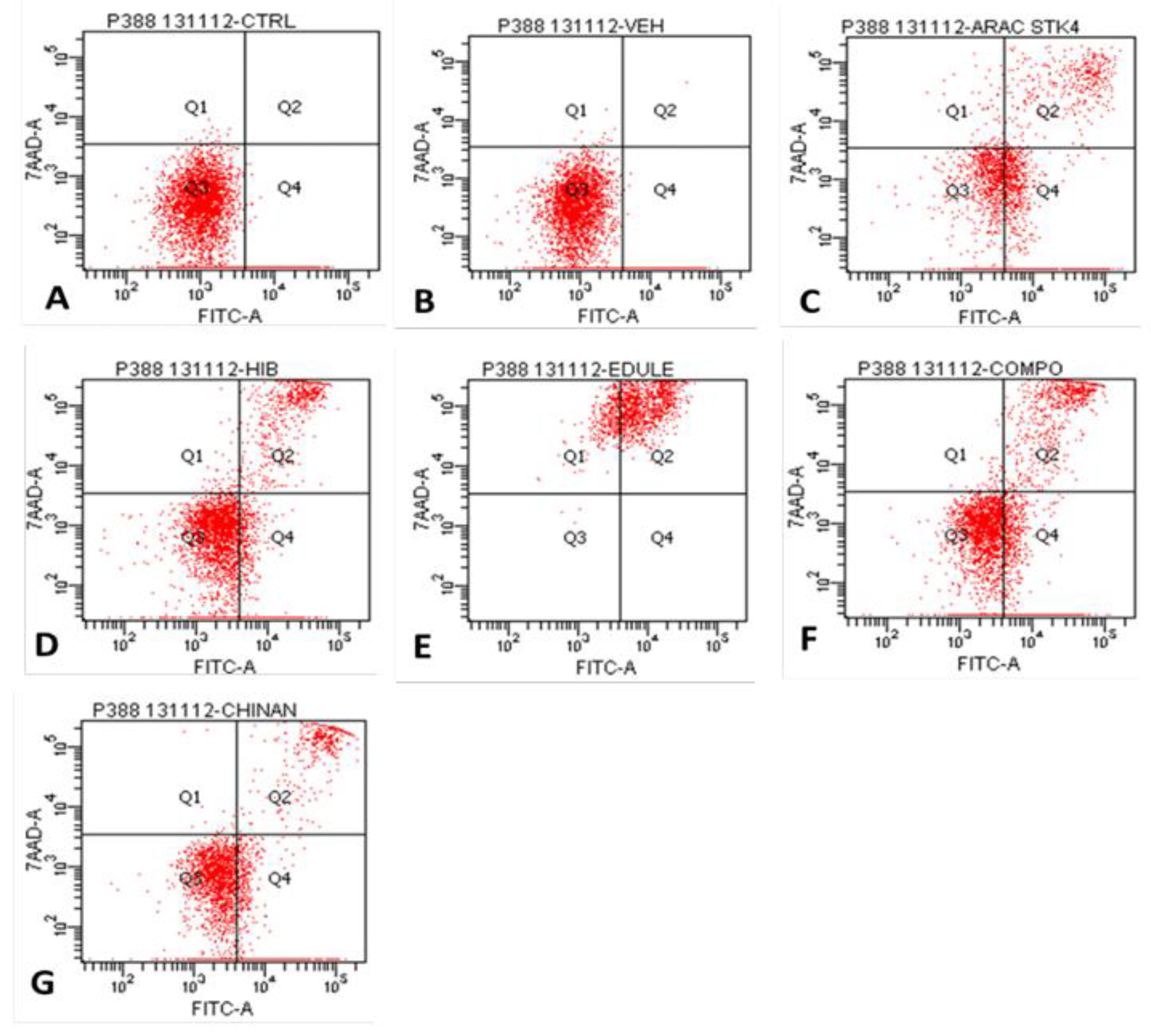
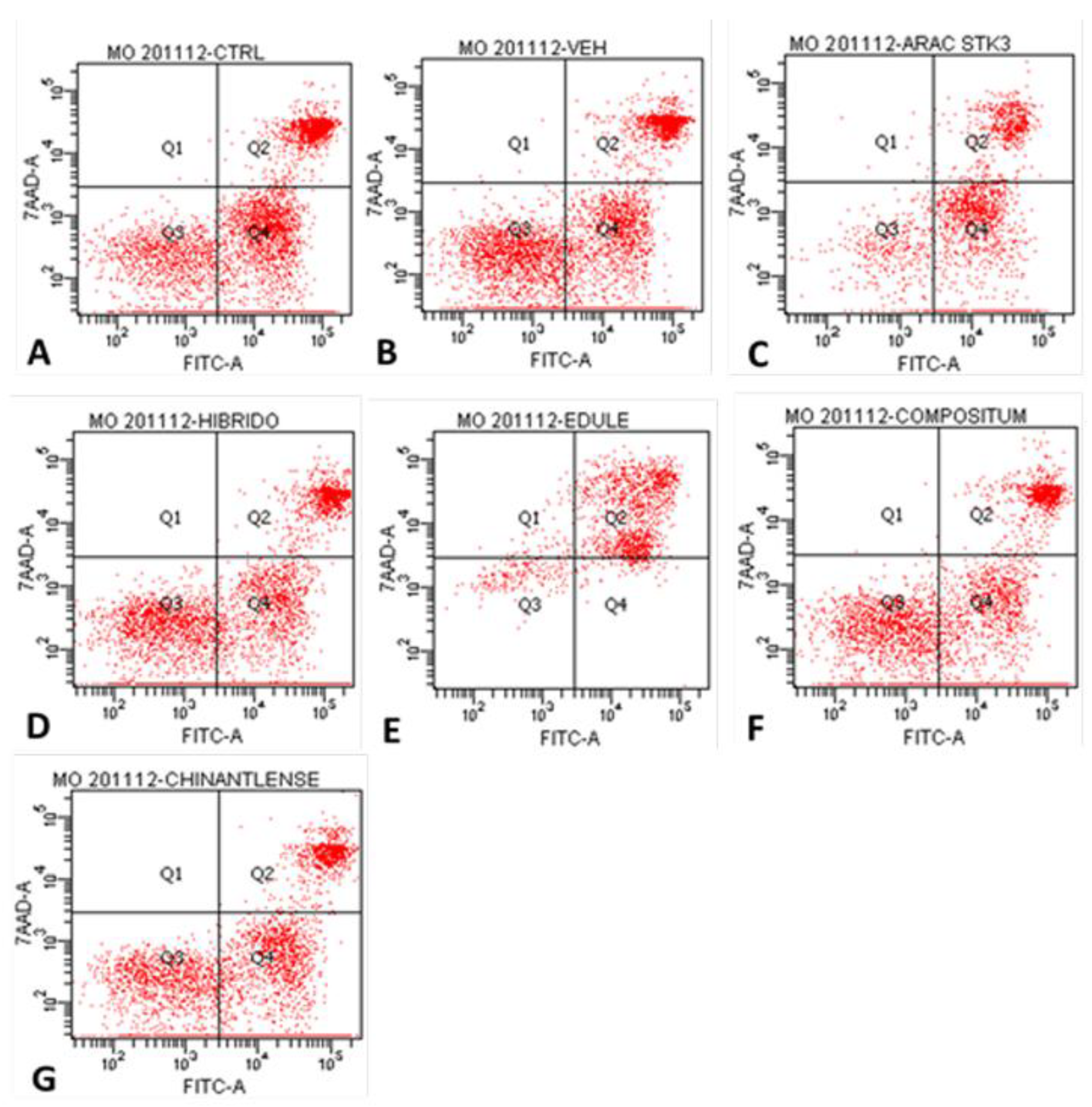
Figure 3.
Detection of phosphatidylserine expression in J774 cells at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 3.
Detection of phosphatidylserine expression in J774 cells at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.

Table 1.
Percentage of cells positive for Annexin V-conjugated FITC labeling bound to PS per quadrant in J774 cells for each treatment. Q1 represents necrosis; Q2 cells are in late apoptosis; Q3 represent normal cells; and Q4 represent cells in early apoptosis.
Table 1.
Percentage of cells positive for Annexin V-conjugated FITC labeling bound to PS per quadrant in J774 cells for each treatment. Q1 represents necrosis; Q2 cells are in late apoptosis; Q3 represent normal cells; and Q4 represent cells in early apoptosis.
| Cells per quadrant (%) | ||||
| Treatment | Q1 | Q2 | Q3 | Q4 |
| Control | 0.5 | 0.2 | 99.2 | 0.1 |
| Vehicle (PBS) | 1.7 | 9.9 | 86.1 | 2.3 |
| Ara-C (Cytarabine®) | 0.1 | 6.5 | 0.8 | 92.6 |
| H-387-07 | 0.6 | 17.5 | 53.2 | 28.7 |
| S. edule var. nigrum spinosum | 4.4 | 95.5 | 0.1 | 0.0 |
| S. compositum | 0.6 | 21.0 | 33.2 | 45.1 |
| S. chinantlense | 2.1 | 23.9 | 48.9 | 25.0 |
Figure 4.
Detection of phosphatidylserine expression in P388 cells at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 4.
Detection of phosphatidylserine expression in P388 cells at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Table 2.
Percentage of cells positive for Annexin V-conjugated FITC labeling bound to PS per quadrant in P388 cells for each treatment. Q1 represents necrosis; Q2 cells are in late apoptosis; Q3 represent normal cells; and Q4 represent cells in early apoptosis.
Table 2.
Percentage of cells positive for Annexin V-conjugated FITC labeling bound to PS per quadrant in P388 cells for each treatment. Q1 represents necrosis; Q2 cells are in late apoptosis; Q3 represent normal cells; and Q4 represent cells in early apoptosis.
| Cells per quadrant (%) | ||||
| Treatment | Q1 | Q2 | Q3 | Q4 |
| Control | 0.1 | 0.0 | 82.0 | 17.9 |
| Vehicle (PBS) | 0.1 | 0.1 | 84.1 | 15.7 |
| Ara-C (Cytarabine®) | 10.0 | 18.2 | 34.1 | 37.7 |
| H-387-07 | 0.8 | 12.8 | 63.3 | 23.0 |
| S. edule var. nigrum spinosum | 17.1 | 82.8 | 0.1 | 0.0 |
| S. compositum | 0.7 | 16.2 | 50.6 | 32.6 |
| S. chinantlense | 0.2 | 9.0 | 51.6 | 39.3 |
Figure 5.
Detection of phosphatidylserine expression in mouse bone marrow mononuclear cells CD-1, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 5.
Detection of phosphatidylserine expression in mouse bone marrow mononuclear cells CD-1, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
3.2. Oligonucleosomal DNA Fragmentation Analysis
The induction of DNA fragmentation is considered the technique par excellence for detecting apoptosis [27], given that the apoptotic process is reversible as long as it does not reach the point of DNA fragmentation, which is a point of no return for the cell and therefore definitive evidence of apoptotic death [28]. The application of the plant extracts to J774 and P388 cells induced DNA fragmentation, due to the expression of a "ladder" pattern [29]. This indicates that, under the conditions of the present study, the extracts lead the cells to a point of irreversible damage (see Figure 6 and Figure 7), and consequently to a complete apoptosis process. The "ladder" pattern was formed by oligonucleosomal fragments of multiples between 180 and 200 base pairs (bp), which is a prominent biochemical marker of apoptotic cells [30] (see Figure 6 and Figure 7).
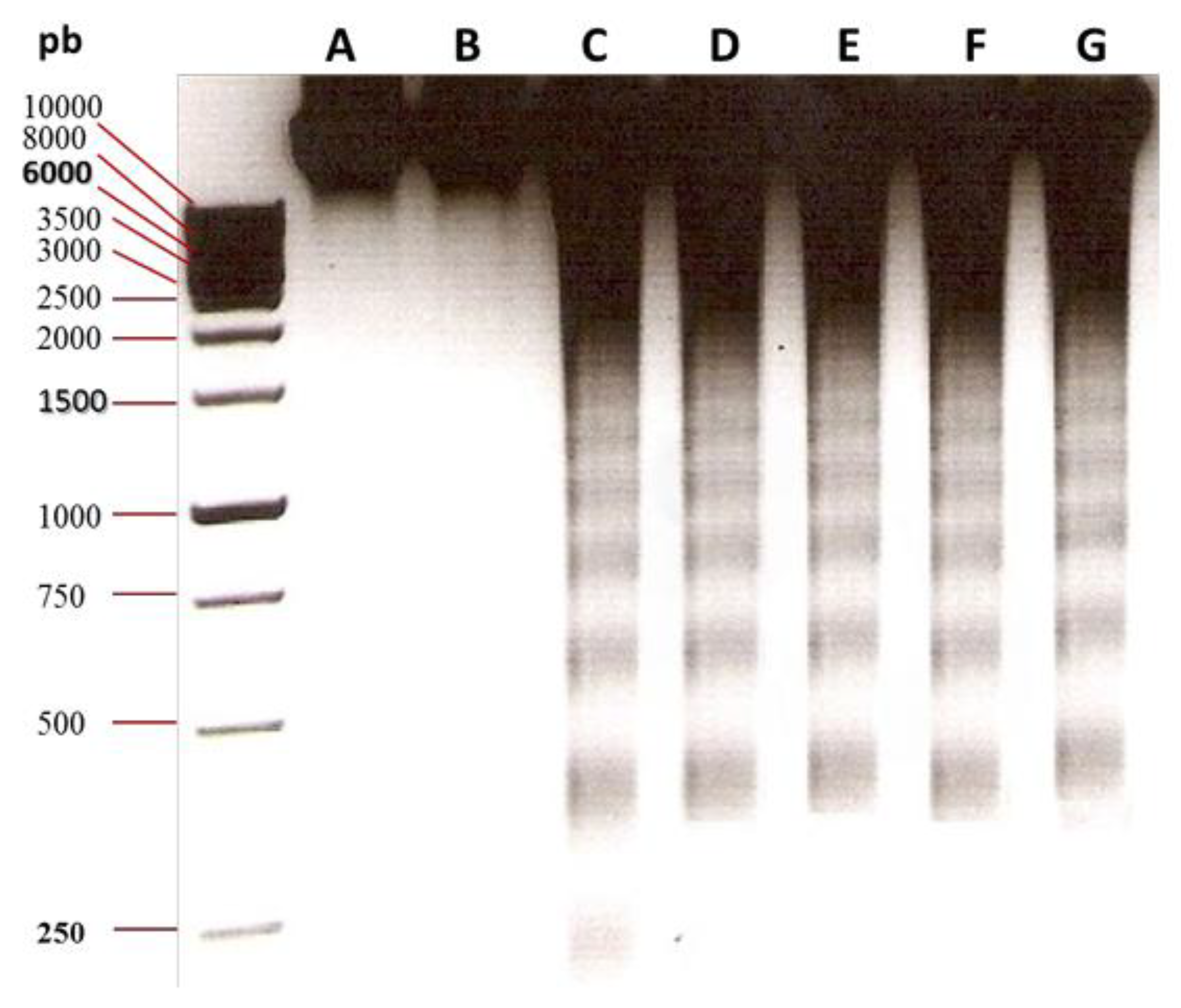
Figure 5.
Detection of DNA fragmentation in J774 cells, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 5.
Detection of DNA fragmentation in J774 cells, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
After observing that the extracts induced PS translocation in normal cells, it was essential to evaluate whether the extracts led to DNA fragmentation in normal bone marrow cells. What was observed was that the plant extracts did not affect the genetic material of normal mouse bone marrow cells, as none of the concentrations of the plant extracts used induced DNA fragmentation, except for the commercial anticancer agent Ara-c® (see Figure 8).
3.3. Antagonistic Effect Analysis
As shown in Table 4, binding energies showed significant differences between groups (Kruskal-Wallis, H = 23.47, p < 0.001; ε2 = 0.795), demonstrating a robust effect depending on the ligand and affinity. Post hoc tests (Mann-Whitney with Bonferroni correction) showed that the ligands gallic acid and cucurbitacin B presented significantly higher affinity than quercetin (p < 0.001), but not between them, which means a better binding profile against the orthosteric site of CCR2.
Comparison of contact residues revealed key differences between the binding pattern of the native ligand (F7N) and the selected ligands. The native ligand interacts with a broad set of amino acids, which according to includes amino acids directly involved in the physiological binding site [31], such as Glu291, His121, Tyr120, Trp98, and Cys190 (see Figure 8A). In comparison, gallic acid showed a higher number of matches with the control (see Figure 8B), including Glu291, Trp98, Gln288, Ser101, Ala102, Tyr120, His121, Cys190, Gly191, Pro192, and Met295, suggesting a possible competition with the physiological binding site of CCL2.
Quercetin was second with 13 shared residues: Trp98, Cys190, Glu291, Tyr120, Thr117, Ser101, Ala102, and Pro192 (see Figure 9C), showing partial binding to the site of interest within the receptor, less stable according to the affinity data in Table 4. Similarly, cucurbitacin B exhibited the lowest number of interactions compared to the control, with 7 matches. Notably, there was no binding of interest with Glu291 or His121, indicating a distance from the site, which may suggest a role as an allosteric modulator (see Figure D). Finally, a comparison of structural similarities between the natural and native ligands was performed using a hierarchical cluster analysis using the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) [32], for which a presence (1) and absence (0) matrix of amino acids was constructed, grouping the ligands according to their coincidence with the control, to which the value of 1 was assigned to all residues. In this way, the degree of overlap with the physiological binding site of the CCR2 receptor could be determined. The dendrogram showed a wide separation between quercetin and the compounds gallic acid and cucurbitacin B, which showed greater closeness to the control, thus demonstrating the natural closeness of gallic acid with the binding mode of the native ligand, followed by cucurbitacin B and finally quercetin (see Figure 9E).
4. Discussion
Leukemia remains a significant public health concern, accounting for approximately 3.5% of all new cancer cases and 4% of cancer-related deaths globally. Traditional medicine has historically posed a pivotal role in the treatment and prevention of various diseases, including leukemia. The advantages of traditional medicine in leukemia treatment include its potential to induce apoptosis in cancer cells, inhibit angiogenesis, modulate immune responses, and target multiple cellular pathways involved in leukemia progression. Sechium spp. is a genus of flowering plants belonging to the Cucurbitaceae family. Among species from the genus Sechium, S. edule var. nigrum spinosum, S. compositum, and S. chinantlense are known for their potential therapeutic properties, and climbing vines, lobed leaves, and green- or dark-colored spiny fruits. Plant hybrids are the result of crossbreeding between two different plant varieties or species, and it is a convenient approach for obtaining species with suitable yield, disease resistance, environmental adaptability, and biological performance. Here, the hybrid H-387-07 was evaluated for its activity towards leukemia and obtained from the hybridization between S. edule var. amarus sylvestris and S. edule var. virens levis by the GISeM.
Cytotoxicity alludes to the capability of organic and inorganic substances to induce cell damage or death. Extracts from natural sources can exert cytotoxicity among cancer cell lines due to their capacity to interfere with the generation of reactive oxygen species (ROS), modulate cell cycle progression, promote the disruption of cellular membranes, interfere with metabolic pathways, and upregulate apoptotic signaling cascades [33]. Here, it was noted that treatment with extracts from S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 reduced the viability of the J774 and P388 cell lines after 68 h of exposure to treatment. Phytochemically, previous reports have demonstrated that the biological activities of extracts from S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 are associated with their flavonoid, phenolic acid, and cucurbitacin content.
In cancer treatment, it has been noted that phenolic acids such as gallic acid can lead cancer cells to death (e.g., HeLa) by modulating the levels of P53/Bax in combination with chemotherapy agents such as doxorubicin [34]. In the same regard, it has been documented that phenolic acids such as caffeic acid can sensitize mutated breast cancer cells to radiation while exerting mitochondrial membrane depolarization (∆Ψm) capacity [35]. Against leukemia-based models, treatment with caffeic acid has been also determined to enhance the activity of imatinib, a targeted therapy agent indicated for CML, in K562 cells while exerting cytotoxicity through the upregulation of the expression of cell cycle repressor genes (CDKN1A and CHES1), and the induction of apoptosis [36]. The anti-leukemia capability of cucurbitacins has been also unveiled over the last years. For instance, recent studies have reported that cucurbitacin D can hamper cell cycle progression, decrease the expression of oncogenes such as ZNF217, and contribute to the apoptosis of chronic B-cell lymphocytic leukemia (MEC1, PGA1), monocytic lymphoma (U937), acute T-cell lymphoblastic leukemia (Jurkat), and AML (OCI-AML3) cell lines [37].
Apoptosis is a form of programmed cell death that is required in maintaining cellular homeostasis, development, and tissue remodeling [38]. Molecularly, apoptosis is characterized by specific morphological and biochemical changes, including cell shrinkage, membrane blebbing, chromatin condensation, and DNA fragmentation [39]. The factors that regulate apoptosis include intrinsic and extrinsic pathways together with cellular stress, growth factors and cytokines, and genetic factors [40]. In leukemia, the regulation of apoptosis exhibits a series of aberrations regarding the upregulation of anti-apoptotic proteins, downregulation of pro-apoptotic proteins, and altered signaling pathways (e.g., PI3K/Akt, NF-κB, and MAPK) involved in membrane permeabilization, apoptosome formation, and activation of downstream effects such as caspases. Here, it was noted that treatment with S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 occurred in the late and early apoptosis of J774 cells being the extract from S. edule var. nigrum spinosum the treatment with the highest effect in late apoptosis (95.5%), followed by S. chinantlense (23.9%), S. compositum (21.0%), and the hybrid H-387-07 (17.5%), respectively.
Similar results were obtained towards the P388 cell line, where it was observed that the extract from S. edule var. nigrum spinosum exerted the highest effect in late apoptosis (82.8%), but followed by S. compositum (16.2%), S. chinantlense (9.0%), and the hybrid H-387-07 (12.8%), respectively. Still, it was also recorded that treatment with extracts exhibited higher capacity to induce early apoptosis in P388 cells. This was the case of treatment with S. compositum (32.6%), S. chinantlense (39.3%), and the hybrid H-387-07 (23.0%). Early apoptosis is the initial reversible phase of programmed cell death, characterized by eliciting intrinsic and extrinsic pathways to lead cells to death and the translocation of PS. Contrary, late apoptosis is the advanced stage of the apoptotic process where irreversible cellular changes such as DNA fragmentation leads to cell death. According to the data enlisted in Table 2 and 3, extracts from Sechium spp. can induce the late or early apoptosis of J774 and P388 cells, which can be related to their phytochemical content [41]. For example, flavonoids such as kaempferol have been documented to induce the early apoptosis of K562 by promoting the translocation of PS upon exposure to 100 μM. In another study, it has been revealed that flavonoids such as quercetin in combination with targeted therapy agents such as venetoclax, can also cause late apoptosis of the KG-1 cell line by inducing poly (ADP-ribose) polymerase cleavage, and cell cycle arrest, together with Bax and Bcl-2 expression [42].
DNA fragmentation refers to the process by which DNA molecules are broken into smaller pieces. This can occur because of normal cellular processes such as apoptosis, as well as from external factors like radiation or chemical exposure [43]. Among cell lines derived from leukemia models, it has been noted that natural products can induce DNA fragmentation by increasing the generation of ROS, which can cause base modifications and cross-linking with DNA strands [44]. In the same context, it has been mentioned that extracts can inhibit the activity of topoisomerases or DNA polymerases, prevent proper repair, replication, or transcription, and lead to the accumulation of DNA breaks and fragmentation [45]. Here, it was observed that exposure to treatment after 68 h occurred in the DNA fragmentation of J744 and P388 cells, as well as mouse bone marrow mononuclear cells. Despite the molecular and cellular variabilities between the cultured cell lines, it was observed in the agarose gels a laddering pattern, which is characteristic of apoptotic events, where activation of caspases leads to the cleavage of nuclear proteins and DNA fragmentation. As mentioned in previous paragraphs, the capacity of extracts from S. edule var. nigrum spinosum, S. compositum, S. chinantlense, and the hybrid H-387-07 to cause DNA fragmentation can be associated with their phytochemical content. For instance, it has been noted that cucurbitacin B can decrease the viability of K562 cells by inducing DNA expression, an effect associated with the increased expression of nuclear H2AX foci, increased intracellular calcium ion concentration, ∆Ψm, and G2/M phase arrest [46]. Similar effects have been reviewed for flavonoids such as quercetin which can induce DNA fragmentation of leukemia cell lines by increasing the ROS levels, while reducing glutathione levels and ∆Ψm [47].
In silico methods involve computational-based approaches to simulate biological processes and predict the interactions between molecules and receptors involved in pathophysiological phenomena. Representative techniques for evaluating this encompass molecular docking, virtual screening, quantitative structure-activity relationship (QSAR) modeling, and molecular dynamics simulations [48]. When evaluating extracts from natural sources, the importance of in silico analysis relies on its capacity to rapidly screen large libraries of compounds for bioactivity against specific cancer targets, and to determine the possibility of utilizing them in future studies involving other in vitro, in vivo, and ex vivo models by initially predicting their binding affinities to target proteins [49]. Here, the interaction between three main constituents of extracts from Sechium spp. were analyzed for their affinity to CCR2, a chemokine type receptor involved in chronic inflammation, tumorigenesis, and poor prognosis among leukemia patients. It was observed that gallic acid (-9.49 kcal/mol), cucurbitacin B (-9.53 kcal/mol), and quercetin (-7.89 kcal/mol) exerted affinity towards the orthosteric site of CCR2. Contrary to cucurbitacin B, and quercetin, it was found that gallic acid exerted the highest number of interactions, especially at Glu291, Trp98, Gln288, Ser101, Ala102, Tyr120, His121, Cys190, Gly191, Pro192, and Met295 residues. Still, cucurbitacin B also exerted higher affinity than quercetin, but lower shared interactions (7 matches). Considering their natural closeness with the binding mode of the native ligand F7N, a UPGMA analysis was performed to categorize the ligands by means of their coincidence with F7N. Thereby, it was recorded that gallic acid exhibited higher closeness with the binding mode of the native ligand, followed by cucurbitacin B, and quercetin.
5. Conclusions
This study reported the capacity of extracts from Sechium spp. decrease the viability of leukemia cell lines by inducing apoptosis and DNA fragmentation. It was noted that the identification of Sechium spp. extracts induce DNA fragmentation in leukemic cells but not in normal cells, indicating that they exhibit selectivity toward tumor cells, unlike the clinically used anticancer drug Ara-C. This is relevant because drugs used to combat cancer specifically target cellular damage, which is not selective for tumor cells. This affects all cells in the body, resulting in adverse toxic effects for the patient. Hence, the importance of having agents that induce apoptosis requires very small doses and exhibit selectivity, as is the case with the Sechium spp. extracts used in the present study. Regarding the evaluation performed in silico, it can be concluded that interrupting the binding of the native CCL2 ligand to CCR2 through the interaction of natural compounds could favor the modulation of the chronic inflammatory response by inhibiting the NF-κB and JAK/STAT pathways, which are associated with the expression of proinflammatory and antiapoptotic cytokines and cell proliferation, especially in tumor contexts. Thus, negative modulation by the selected compounds could also improve apoptotic processes. Further studies are required to validate in vitro and in vivo the capability of extracts from Sechium spp. to modulate cell signaling pathways involved in leukemia initiation and progression.
Supplementary Materials
None.
Author Contributions
Conceptualization, J.D.C.-Z., J.C.-I., and J.L.M.-M.; validation, J.C.-I., E.S.-O., I.A.-S., M.I.I.-L., D.A.C.-Z., I.V., E.S.-A., J.M.S.-J., and J.L.M.-M.; investigation, J.D.C.-Z., J.C.-I., E.S.-O., J.M.S.-J., and J.L.M.-M.; writing—original draft preparation, J.D.C.-Z., J.C.-I., I.V., E.S.-A., J.M.S.-J., and J.L.M.-M; writing—review and editing, E.S.-O., I.A.-S., M.I.I.-L., D.A.C.-Z., I.V., E.S.-A., J.M.S.-J., and J.L.M.-M.; visualization, J.D.C.-Z., J.C.-I., E.S.-O., I.V., and J.M.S.-J.; supervision, J.C.-I.; funding acquisition; J.C.-I., E.S.-A., J.M.S.-J., and J.L.M.-M; project administration, J.C.-I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data generated from this study can be consulted with the authors for correspondence upon reasonable request.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gower, M.; Li, X.; Aguilar-Navarro, A.G.; Lin, B.; Fernandez, M.; Edun, G.; Nader, M.; Rondeau, V.; Arruda, A.; Tierens, A.; et al. An Inflammatory State Defines a High-Risk T-Lineage Acute Lymphoblastic Leukemia Subgroup. Science Translational Medicine 2025, 17, eadr2012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Guo, C.; Liang, Y.; Qing, S.; Liang, Z.; Wang, Y.; Zhang, C. The Burden Dynamics of Leukemia in China from 1990 to 2021: An Epidemiological Analysis of Trends, Risk Factors, and Projections to 2036. Preventive Medicine Reports 2025, 57, 103191. [Google Scholar] [CrossRef] [PubMed]
- Ghouraba, R.F.; EL-Desouky, S.S.; El-Shanshory, M.R.; Kabbash, I.A.; Metwally, N.M. Early Diagnosis of Acute Lymphoblastic Leukemia Utilizing Clinical, Radiographic, and Dental Age Indicators. Sci Rep 2025, 15, 12376. [Google Scholar] [CrossRef]
- Duchesneau, E.D.; McNeill, A.M.; Schary, W.; Pate, V.; Lund, J.L. Prognosis of Older Adults with Chronic Lymphocytic Leukemia: A Surveillance, Epidemiology, and End Results-Medicare Cohort Study. Journal of Geriatric Oncology 2023, 14, 101602. [Google Scholar] [CrossRef]
- Zehtabcheh, S.; Soleimani Samarkhazan, H.; Asadi, M.; Zabihi, M.; Parkhideh, S.; Mohammadi, M.H. Insights into KMT2A Rearrangements in Acute Myeloid Leukemia: From Molecular Characteristics to Targeted Therapies. Biomark Res 2025, 13, 73. [Google Scholar] [CrossRef]
- Shahzad, M.; Iqbal, Q.; Amin, M.K.; Irfan, S.; Warraich, S.Z.; Anwar, I.; Dave, P.; Basharat, A.; Hebishy, A.; Faisal, M.S.; et al. Outcomes of Hematopoietic Stem Cell Transplantation in Primary Plasma Cell Leukemia: A Systematic Review and Meta-Analysis. Leukemia Research 2025, 148, 107640. [Google Scholar] [CrossRef]
- Grzegorzewska, W.; Zarobkiewicz, M.; Jastrzębska-Pawłowska, K.; Lehman, N.; Tomczak, W.; Mizerska-Kowalska, M.; Bojarska-Junak, A.; Roliński, J. MLR Corresponds to the Functional Status of Monocytes in Chronic Lymphocytic Leukemia. International Journal of Inflammation 2025, 2025, 4443773. [Google Scholar] [CrossRef]
- Avila-Rodríguez, D.; Ibarra-Sánchez, A.; Sosa-Garrocho, M.; Vázquez-Victorio, G.; Caligaris, C.; Anaya-Rubio, I.; Segura-Villalobos, D.; Blank, U.; González-Espinosa, C.; Macias-Silva, M. An Autocrine Regulator Loop Involving Tumor Necrosis Factor and Chemokine (C-C Motif) Ligand-2 Is Activated by Transforming Growth Factor-β in Rat Basophilic Leukemia-2H3 Mast Cells. International Journal of Molecular Sciences 2025, 26, 4263. [Google Scholar] [CrossRef]
- Cardona, C.I.; Rodriguez, A.; Torres, V.C.; Sanchez, A.; Torres, A.; Vazquez, A.E.; Wagler, A.E.; Brissette, M.A.; Bill, C.A.; Vines, C.M. C-C Chemokine Receptor 7 Promotes T-Cell Acute Lymphoblastic Leukemia Invasion of the Central Nervous System via Β2-Integrins. International Journal of Molecular Sciences 2024, 25, 9649. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Ma, L.; Huang, T.; Wang, T.; Zhou, F.; Liu, X. Azelaic Acid Attenuates CCL2/CCR2 Axis-Mediated Skin Trafficking of Acute Myeloid Leukemia Cells through NF-κB/MAPK Signaling Modulation in Keratinocytes. BMC Cancer 2025, 25, 1250. [Google Scholar] [CrossRef]
- Zhang, L.; Lei, L.; Zhao, Z.; Yang, G.; Wang, K.; Wang, L.; Zhang, N.; Ai, Y.; Ma, X.; Liu, G.; et al. Enhanced Venetoclax Delivery Using L-Phenylalanine Nanocarriers in Acute Myeloid Leukemia Treatment. Chinese Chemical Letters 2025, 36, 110316. [Google Scholar] [CrossRef]
- Korbecki, J.; Bosiacki, M.; Stasiak, P.; Snarski, E.; Brodowska, A.; Chlubek, D.; Baranowska-Bosiacka, I. Clinical Aspects and Significance of β-Chemokines, γ-Chemokines, and δ-Chemokines in Molecular Cancer Processes in Acute Myeloid Leukemia (AML) and Myelodysplastic Neoplasms (MDS). Cancers 2024, 16, 3246. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Long, Z.; Lei, M.; Ding, R.; Chen, M. Integrated Genomics Reveal Potential Resistance Mechanisms of PANoptosis-Associated Genes in Acute Myeloid Leukemia. Molecular Carcinogenesis 2025, 64, 801–815. [Google Scholar] [CrossRef]
- Gras, E.; Azoyan, L.; Monzó-Gallo, P.; Garcia-Vidal, C.; Lanternier, F.; Brissot, E.; Guitard, J.; Lacombe, K.; Dechartres, A.; Surgers, L. Risk Factors for Invasive Mould Infections in Adult Patients with Hematological Malignancies and/or Stem Cell Transplant: A Systematic Literature Review and Meta-Analysis. Journal of Infection 2025, 106574. [Google Scholar] [CrossRef] [PubMed]
- Oluwole, O.O.; Ray, M.D.; Ma, H.; Sharma, R.; Patel, A.R.; Smith, N. Health and Economic Impact of Vein-to-Vein Time in CAR T-Cell Therapy in the Second-Line Treatment of Relapsed/Refractory Large B-Cell Lymphoma: A US Cost-Effectiveness Analysis. Transplantation and Cellular Therapy 2025. [Google Scholar] [CrossRef] [PubMed]
- Gyi, K.K.; Anuchapreeda, S.; Intasai, N.; Tungjai, M.; Okonogi, S.; Iwasaki, A.; Usuki, T.; Tima, S. Anti-Leukemia Activity of the Ethyl Acetate Extract from Gynostemma Pentaphyllum (Thunb.) Leaf against FLT3-Overexpressing AML Cells and Its Phytochemical Characterization. BMC Complement Med Ther 2025, 25, 172. [Google Scholar] [CrossRef]
- Cavalloro, V.; Malacrida, A.; Miloso, M.; Ronchi, D.; Porta, A.; Fossati, A.; Gheza, G.; De Siervi, S.; Mantovani, S.; Oliviero, B.; et al. From Lichen to Organoids: Usnic Acid Enantiomers Show Promise against Cholangiocarcinoma via MNK2 Targeting and MAPK Pathway Modulation. Biomedicine & Pharmacotherapy 2025, 188, 118208. [Google Scholar] [CrossRef]
- Javrushyan, H.; Ginovyan, M.; Harutyunyan, T.; Gevorgyan, S.; Karabekian, Z.; Maloyan, A.; Avtandilyan, N. Elucidating the Impact of Hypericum Alpestre Extract and L-NAME on the PI3K/Akt Signaling Pathway in A549 Lung Adenocarcinoma and MDA-MB-231 Triple-Negative Breast Cancer Cells. PLoS ONE 2025, 20, e0303736. [Google Scholar] [CrossRef]
- Martínez-García, M.; Pascual-Pineda, L.A.; Rascón-Díaz, M.P.; Quintanilla-Carvajal, M.X.; Jiménez-Fernández, M. Physicochemical, Technological, and Structural Properties and Sensory Quality of Bread Prepared with Wheat Flour and Pumpkin (Cucurbita Argyrosperma), Chayotextle (Sechium Edule Root) and Jinicuil (Inga Paterno Seeds) Flour. ACS Food Sci. Technol. 2025, 5, 512–524. [Google Scholar] [CrossRef]
- Osorio, E.-S.; María-Guadalupe, R.-P.; Víctor-Manuel, M.-N.; Jorge, C.-Í.; Marcos, S.-H.; Juana, R.-P.; Ernesto, R.-L.; Benny, W.-S.; Graciela, G.-G.; Taide-Laurita, A.-U.; et al. Hepatoprotective Effect of the Sechium HD-Victor Hybrid Extract in a Model of Liver Damage Induced by Carbon Tetrachloride in Mice. Biomedicine & Pharmacotherapy 2025, 183, 117831. [Google Scholar] [CrossRef]
- Salazar-Aguilar, S.; Ruiz-Posadas, L.D.M.; Cadena-Iñiguez, J.; Soto-Hernández, M.; Santiago-Osorio, E.; Aguiñiga-Sánchez, I.; Rivera-Martínez, A.R.; Aguirre-Medina, J.F. Sechium Edule (Jacq.) Swartz, a New Cultivar with Antiproliferative Potential in a Human Cervical Cancer HeLa Cell Line. Nutrients 2017, 9, 798. [Google Scholar] [CrossRef]
- Cadena-Iñiguez, J.; Aguiñiga-Sánchez, I.; Uriostegui-Arias, M.T.; Santiago-Osorio, E.; Ruiz-Posadas, L.d.M.; Soto-Hernández, M. Antiproliferative Effect of Sechium Edule (Jacq.) Sw., Cv. Madre Negra Extracts on Breast Cancer In Vitro. Separations 2022, 9, 230. [Google Scholar] [CrossRef]
- Aguiñiga-Sánchez, I.; Cadena-Íñiguez ,Jorge; Santiago-Osorio ,Edelmiro; Gómez-García ,Guadalupe; Mendoza-Núñez ,Víctor Manuel; Rosado-Pérez ,Juana; Ruíz-Ramos ,Mirna; Cisneros-Solano ,Víctor Manuel; Ledesma-Martínez ,Edgar; Delgado-Bordonave ,Angel de Jesus; et al. Chemical Analyses and in Vitro and in Vivo Toxicity of Fruit Methanol Extract of Sechium Edule Var. Nigrum Spinosum. Pharmaceutical Biology 2017, 55, 1638–1645. [CrossRef]
- Aguiñiga-Sánchez, I.; Soto-Hernández, M.; Cadena-Iñiguez, J.; Ruíz-Posadas, L.d.M.; Cadena-Zamudio, J.D.; González-Ugarte, A.K.; Weiss Steider, B.; Santiago-Osorio, E. Fruit Extract from A Sechium Edule Hybrid Induce Apoptosis in Leukaemic Cell Lines but Not in Normal Cells. Nutrition and Cancer 2015, 67, 250–257. [Google Scholar] [CrossRef]
- Cadena-Iñiguez, J.; Arévalo-Galarza, M.d.L.; Santiago-Osorio, E.; Aguiñiga-Sánchez, I.; Cadena-Zamudio, J.D.; Soto-Hernández, M.; Ramírez-Rodas, Y.C.; Ruiz-Posadas, L.d.M.; Salazar-Aguilar, S.; Cisneros-Solano, V.M. Genotypes of Sechium Spp. as a Source of Natural Products with Biological Activity. Life 2025, 15, 15. [Google Scholar] [CrossRef]
- Rivera-Martínez, A.R.; Aguiñiga-Sánchez, I.; Cadena-Iñiguez, J.; Soto-Cruz, I.; Monroy-García, A.; Gómez-García, G.; Ledesma-Martínez, E.; Weiss-Steider, B.; Santiago-Osorio, E. Fruit Extract of Sechium Chinantlense (Lira & F. Chiang) Induces Apoptosis in the Human Cervical Cancer HeLa Cell Line. Nutrients 2023, 15, 667. [Google Scholar] [CrossRef]
- Mustafa, N.N.; El-Desouky, M.A.; Shawush, N.A.; Hanna, D.H. Apoptosis Induction in Ascorbic Acid Treated Human Colorectal Cancer Cell Lines (Caco-2). Journal of Biologically Active Products from Nature 2025, 15, 56–71. [Google Scholar] [CrossRef]
- Ismail, D.F.; El-Keey, M.M.; Elgendy, S.M.; Hessien, M. Impregnation of Mesenchymal Stem Cell Conditioned Media with Wortmannin Enhanced Its Antiproliferative Effect in Breast Cancer Cells via PI3K/Akt/mTOR Pathway. BMC Res Notes 2025, 18, 93. [Google Scholar] [CrossRef]
- Hussain, S.; Ashafaq, M.; Alshahrani, S.; Qadri, M.; Khardali, A.; Mawkili, W.; Hassan, D.A.; Alam, M.I.; Almoshari, Y.; Elhassan Taha, M.M.; et al. Synergistic Effect of Piperine on Curcumin in Cisplatin-Induced Nephrotoxicity through DNA Fragmentation and Cytokines Gene Expressions. Drug and Chemical Toxicology 0, 1–10. [CrossRef]
- Abd El-Raheem, H.; Alawam, A.S.; Rudayni, H.A.; Allam, A.A.; Helim, R.; Fafa, S.; Yahia, S.; Mahmoud, R.; Alahmad, W. Emerging Electrochemical Approaches for the Early Detection of Programmed Cell Death. ACS Omega 2025, 10, 34106–34122. [Google Scholar] [CrossRef]
- Apel, A.-K.; Cheng, R.K.Y.; Tautermann, C.S.; Brauchle, M.; Huang, C.-Y.; Pautsch, A.; Hennig, M.; Nar, H.; Schnapp, G. Crystal Structure of CC Chemokine Receptor 2A in Complex with an Orthosteric Antagonist Provides Insights for the Design of Selective Antagonists. Structure 2019, 27, 427–438.e5. [Google Scholar] [CrossRef]
- Loewenstein, Y.; Portugaly, E.; Fromer, M.; Linial, M. Efficient Algorithms for Accurate Hierarchical Clustering of Huge Datasets: Tackling the Entire Protein Space. Bioinformatics 2008, 24, i41–i49. [Google Scholar] [CrossRef]
- Doan, C.C.; Le, T.L.; Ho, N.Q.C.; Nguyen, T.T.; Hoang, N.Q.H.; Le, P.C.; Le, N.T.L.; Tran, T.L.G.; Nguyen, T.P.T.; Hoang, N.S. Cytotoxic Effects of the Standardized Extract from Curcuma Aromatica Salisb. Rhizomes via Induction of Mitochondria-Mediated Caspase-Dependent Apoptotic Pathway and P21-Mediated G0/G1 Cell Cycle Arrest on Human Gastric Cancer AGS Cells. Journal of Toxicology and Environmental Health, Part A 2025, 88, 227–249. [Google Scholar] [CrossRef] [PubMed]
- Sarı, U.; Zaman, F.; Özdemir, İ.; Öztürk, Ş.; Tuncer, M.C. Gallic Acid Induces HeLa Cell Lines Apoptosis via the P53/Bax Signaling Pathway. Biomedicines 2024, 12, 2632. [Google Scholar] [CrossRef]
- Prades-Sagarra, È.; Geurts, F.A.P.; Biemans, R.; Lieuwes, N.G.; Yaromina, A.; Dubois, L.J. The Radiosensitizing Effect of Caffeic Acid Phenethyl Ester in Breast Cancer Is Dependent on P53 Status. Radiotherapy and Oncology 2025, 209, 110945. [Google Scholar] [CrossRef] [PubMed]
- Feriotto, G.; Tagliati, F.; Giriolo, R.; Casciano, F.; Tabolacci, C.; Beninati, S.; Khan, M.T.H.; Mischiati, C. Caffeic Acid Enhances the Anti-Leukemic Effect of Imatinib on Chronic Myeloid Leukemia Cells and Triggers Apoptosis in Cells Sensitive and Resistant to Imatinib. International Journal of Molecular Sciences 2021, 22, 1644. [Google Scholar] [CrossRef]
- Adorisio, S.; Fierabracci, A.; Cham, B.T.; Hoang, V.D.; Thuy Linh, N.T.; Nhung, L.T.H.; Martelli, M.P.; Ayroldi, E.; Ronchetti, S.; Rosati, L.; et al. Modulatory Effect of Cucurbitacin D from Elaeocarpus Hainanensis on ZNF217 Oncogene Expression in NPM-Mutated Acute Myeloid Leukemia. Pharmaceuticals 2024, 17, 1561. [Google Scholar] [CrossRef]
- Liu, C.; Jin, X.; An, Y.; Li, W. A Predictive Model Based on Program Cell Death Genes for Prognosis and Therapeutic Response in Early Stage Hepatocellular Carcinoma. Sci Rep 2025, 15, 13937. [Google Scholar] [CrossRef]
- Agrawal, M.; Saxena, A.K.; Agrawal, S.K. Essential Oil from Ocimum Carnosum Induces ROS Mediated Mitochondrial Dysfunction and Intrinsic Apoptosis in HL-60 Cells. Toxicology in Vitro 2025, 104, 105988. [Google Scholar] [CrossRef]
- Wu, X.; Gu, R.; Tang, M.; Mu, X.; He, W.; Nie, X. Elucidating the Dual Roles of Apoptosis and Necroptosis in Diabetic Wound Healing: Implications for Therapeutic Intervention. BURNS TRAUMA 2025, 13, tkae061. [Google Scholar] [CrossRef]
- Saraei, R.; Rahman, H.S.; Soleimani, M.; Asghari-Jafarabadi, M.; Naimi, A.; Hassanzadeh, A.; Solali, S. Kaempferol Sensitizes Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand-Resistance Chronic Myelogenous Leukemia Cells to Apoptosis. Mol Biol Rep 2022, 49, 19–29. [Google Scholar] [CrossRef]
- Kawakatsu, R.; Tadagaki, K.; Yamasaki, K.; Kuwahara, Y.; Nakada, S.; Yoshida, T. The Combination of Venetoclax and Quercetin Exerts a Cytotoxic Effect on Acute Myeloid Leukemia. Sci Rep 2024, 14, 26418. [Google Scholar] [CrossRef]
- Kagiya, G.; Ogawa, R.; Matsumoto, T.; Hyodo, F.; Abe, N.; Yuzawa, A.; Takeuchi, H.; Aoyagi, M.; Sato, A.; Yamashita, K.; et al. Real-Time Imaging Reveals Radiation-Induced Intratumor Apoptosis via Nutrient and Oxygen Deprivation Following Vascular Damage. Molecular Therapy Oncology 2025, 33. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lan, L. Transcription-Coupled DNA Repair Protects Genome Stability upon Oxidative Stress-Derived DNA Strand Breaks. FEBS Letters 2025, 599, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhang, X.; Zeng, F. Biological Functions and Health Benefits of Flavonoids in Fruits and Vegetables: A Contemporary Review. Foods 2025, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhao, W.; Hao, W.; Ren, G.; Lu, J.; Chen, X. Cucurbitacin B Induces DNA Damage, G2/M Phase Arrest, and Apoptosis Mediated by Reactive Oxygen Species (ROS) in Leukemia K562 Cells. Anti-Cancer Agents in Medicinal Chemistry 2014, 14, 1146–1153. [Google Scholar] [CrossRef]
- Moriello, C.; De Rosa, C.; D’Angelo, S.; Pasquale, P. Polyphenols and Chronic Myeloid Leukemia: Emerging Therapeutic Opportunities. Hemato 2025, 6, 28. [Google Scholar] [CrossRef]
- S. Lakshmi, B.; K. Bidarur, J.; G. Anilkumar, H.; S. Ravindranath, B. Prioritization of Phytochemical Isolation and Characterization against HER2 as a Breast Cancer Target Based on Chromatographic Methods, DFT Studies, 3D-QSAR Analysis, and Molecular Docking Simulations. RSC Advances 2025, 15, 25103–25114. [CrossRef]
- Millan-Casarrubias, E.J.; García-Tejeda, Y.V.; González-De la Rosa, C.H.; Ruiz-Mazón, L.; Hernández-Rodríguez, Y.M.; Cigarroa-Mayorga, O.E. Molecular Docking and Pharmacological In Silico Evaluation of Camptothecin and Related Ligands as Promising HER2-Targeted Therapies for Breast Cancer. Current Issues in Molecular Biology 2025, 47, 193. [Google Scholar] [CrossRef]
Figure 1.
Reduction in cell density in the J774 cell line was observed under a microscope (10X) after 68 h of stimulation with the IC50 of plant extracts of Sechium spp. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 1.
Reduction in cell density in the J774 cell line was observed under a microscope (10X) after 68 h of stimulation with the IC50 of plant extracts of Sechium spp. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
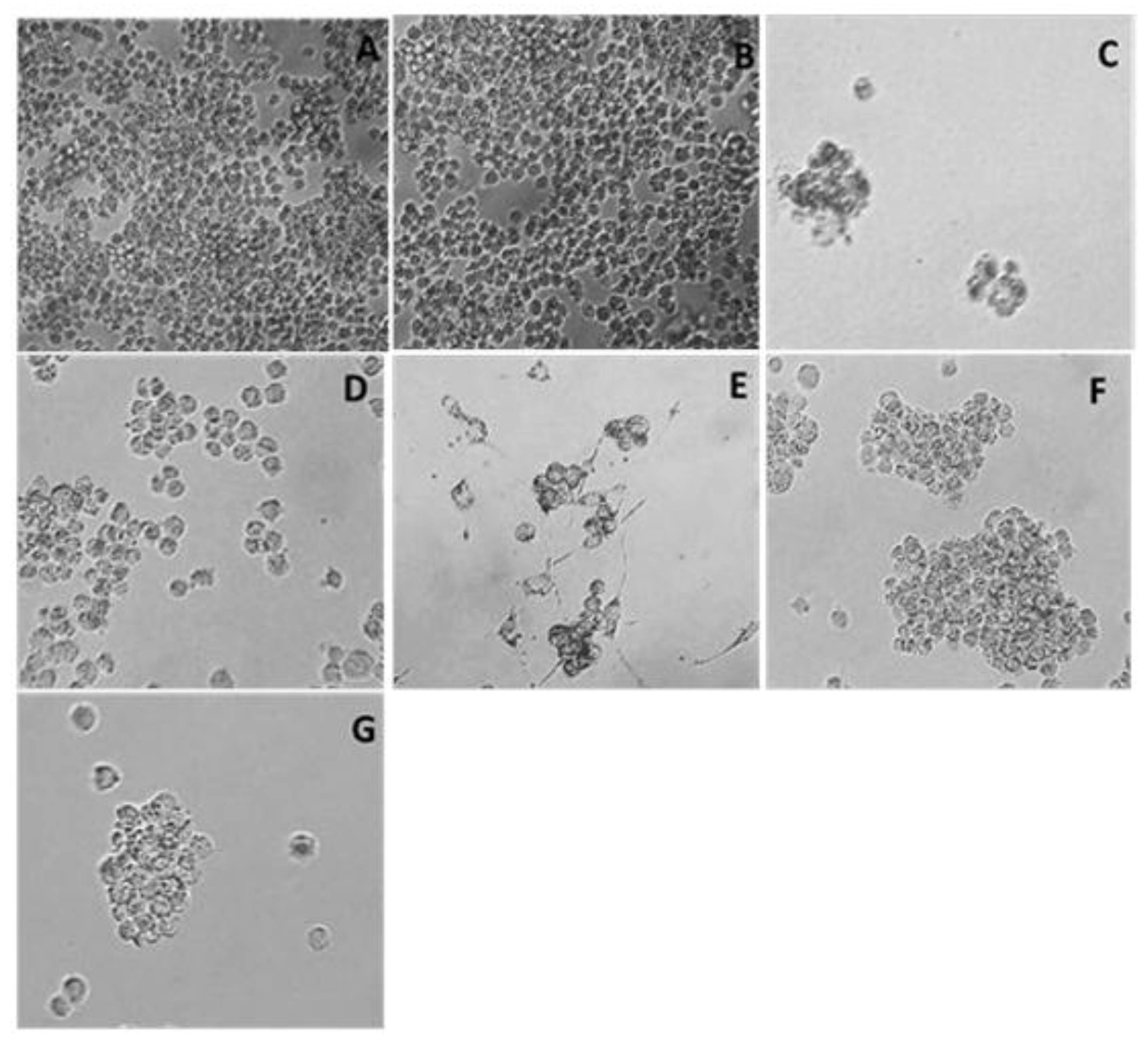
Figure 2.
Reduction in cell density in the P388 cell line observed under a microscope (10X) after 60 h of stimulation with the IC50 of plant extracts of Sechium spp. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 2.
Reduction in cell density in the P388 cell line observed under a microscope (10X) after 60 h of stimulation with the IC50 of plant extracts of Sechium spp. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
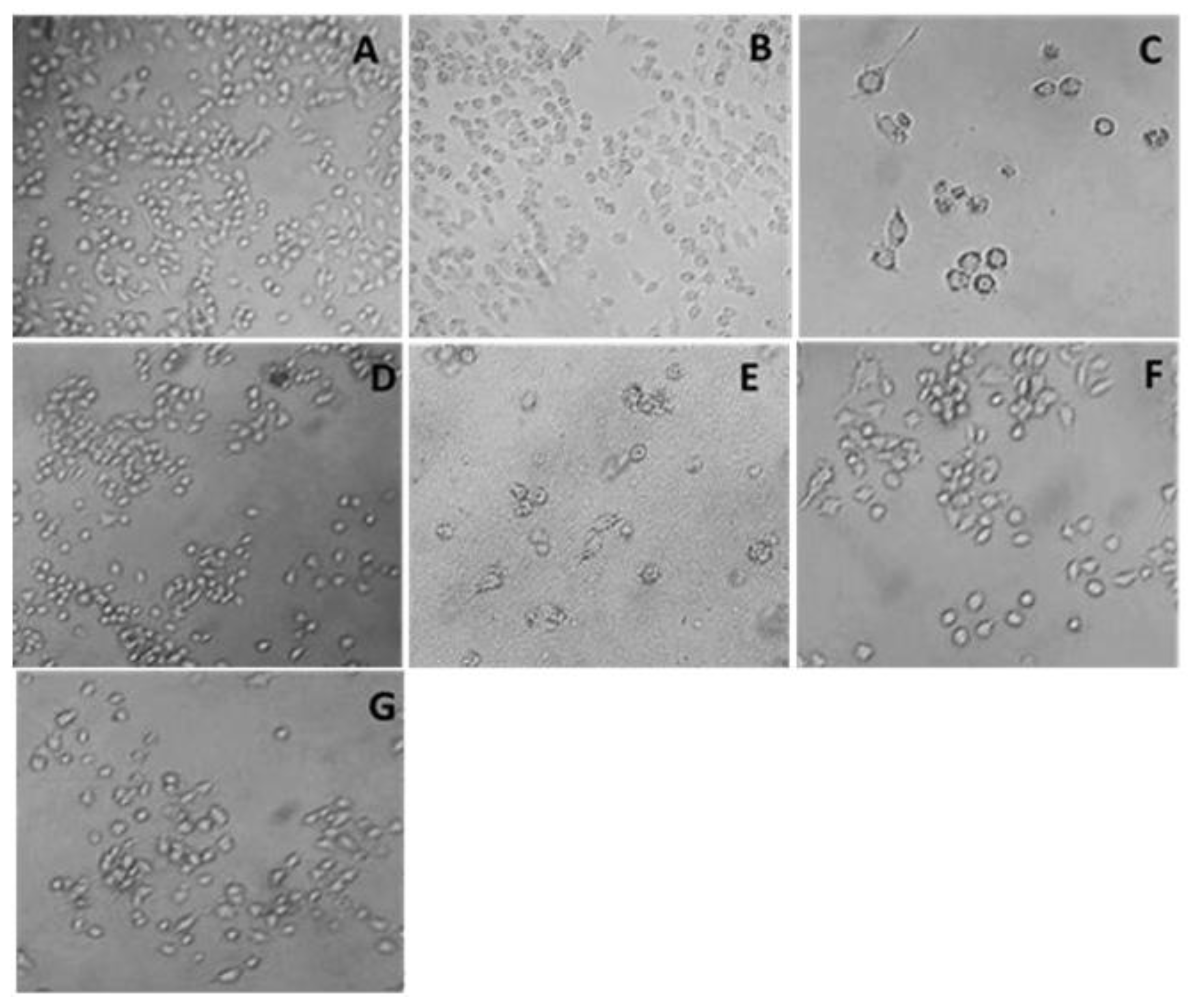
Figure 6.
Detection of DNA fragmentation in P388 cells, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 6.
Detection of DNA fragmentation in P388 cells, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
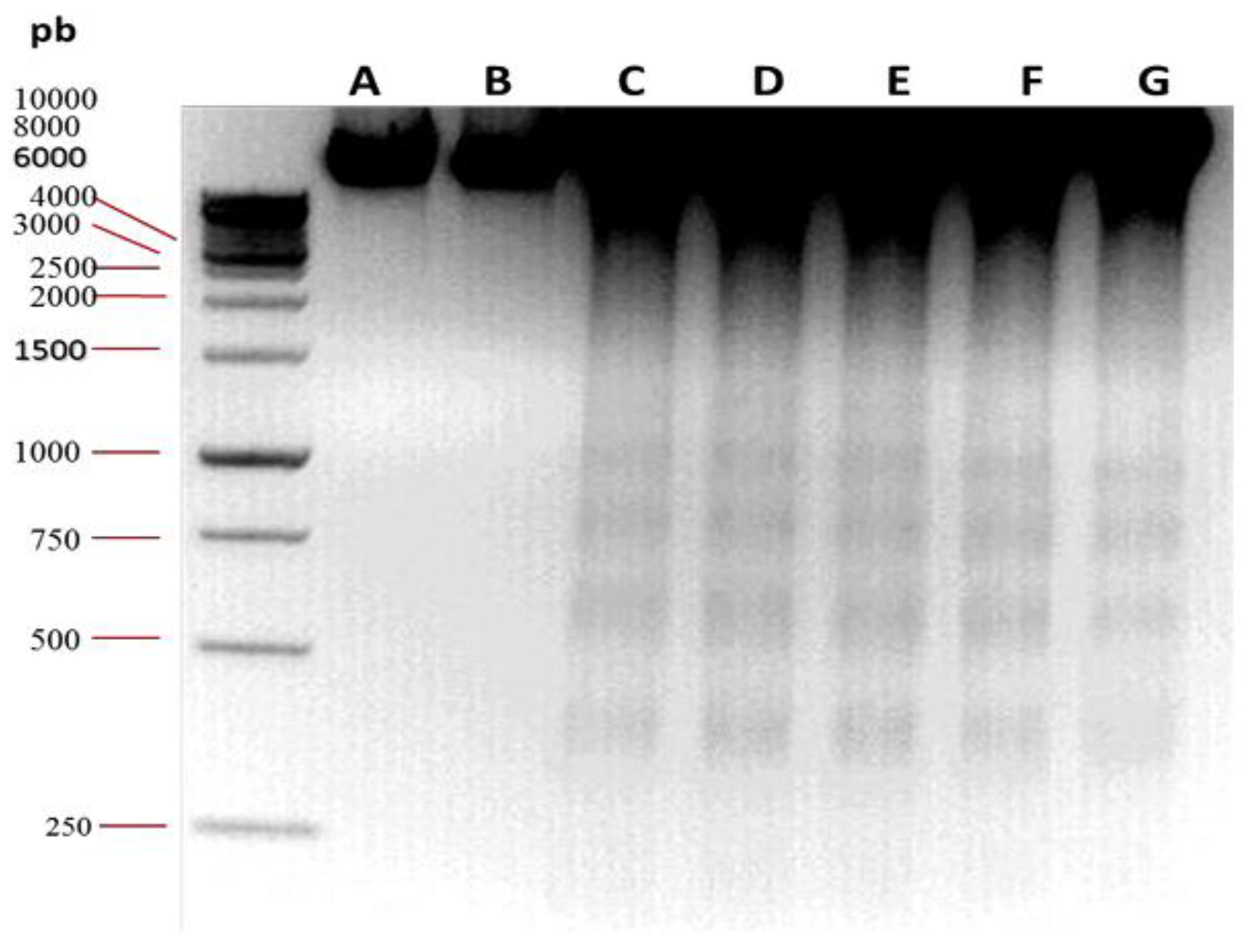
Figure 7.
Detection of DNA fragmentation in mouse bone marrow mononuclear cells CD-1 cells, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
Figure 7.
Detection of DNA fragmentation in mouse bone marrow mononuclear cells CD-1 cells, at 68 h of stimulation with the IC50 of Sechium spp. plant extracts. (A) Control, (B) Vehicle (PBS), (C) Ara-C (Cytarabine®) (5 μM), (D) H-387-07, (E) S. edule var. nigrum spinosum, (F) S. compositum, and (G) S. chinantlense.
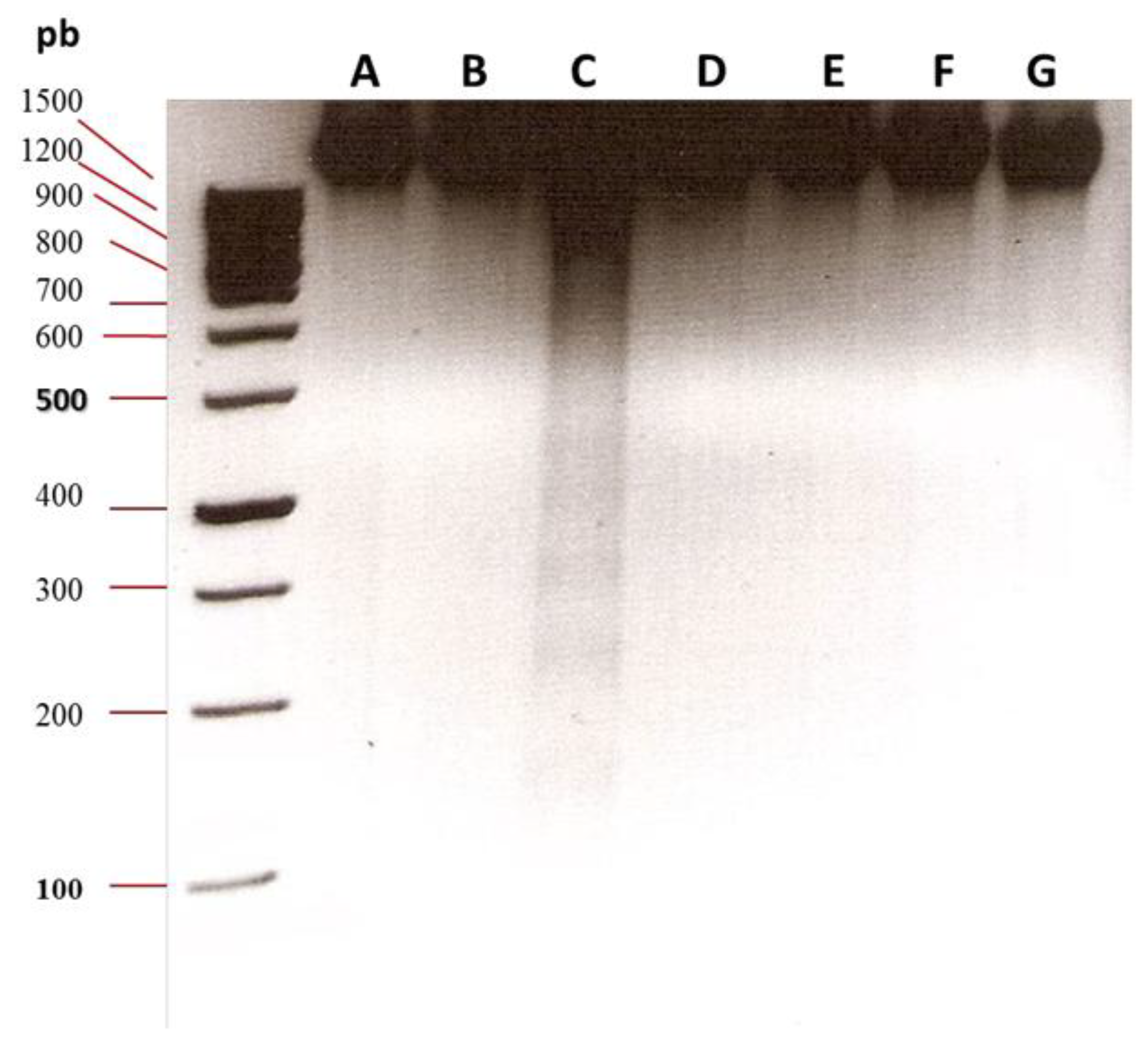
Figure 8.
In silico analysis of (A) F7N, (B) gallic acid, (C) quercetin, and (D) cucurbitacin B towards the CCR2 receptor.
Figure 8.
In silico analysis of (A) F7N, (B) gallic acid, (C) quercetin, and (D) cucurbitacin B towards the CCR2 receptor.
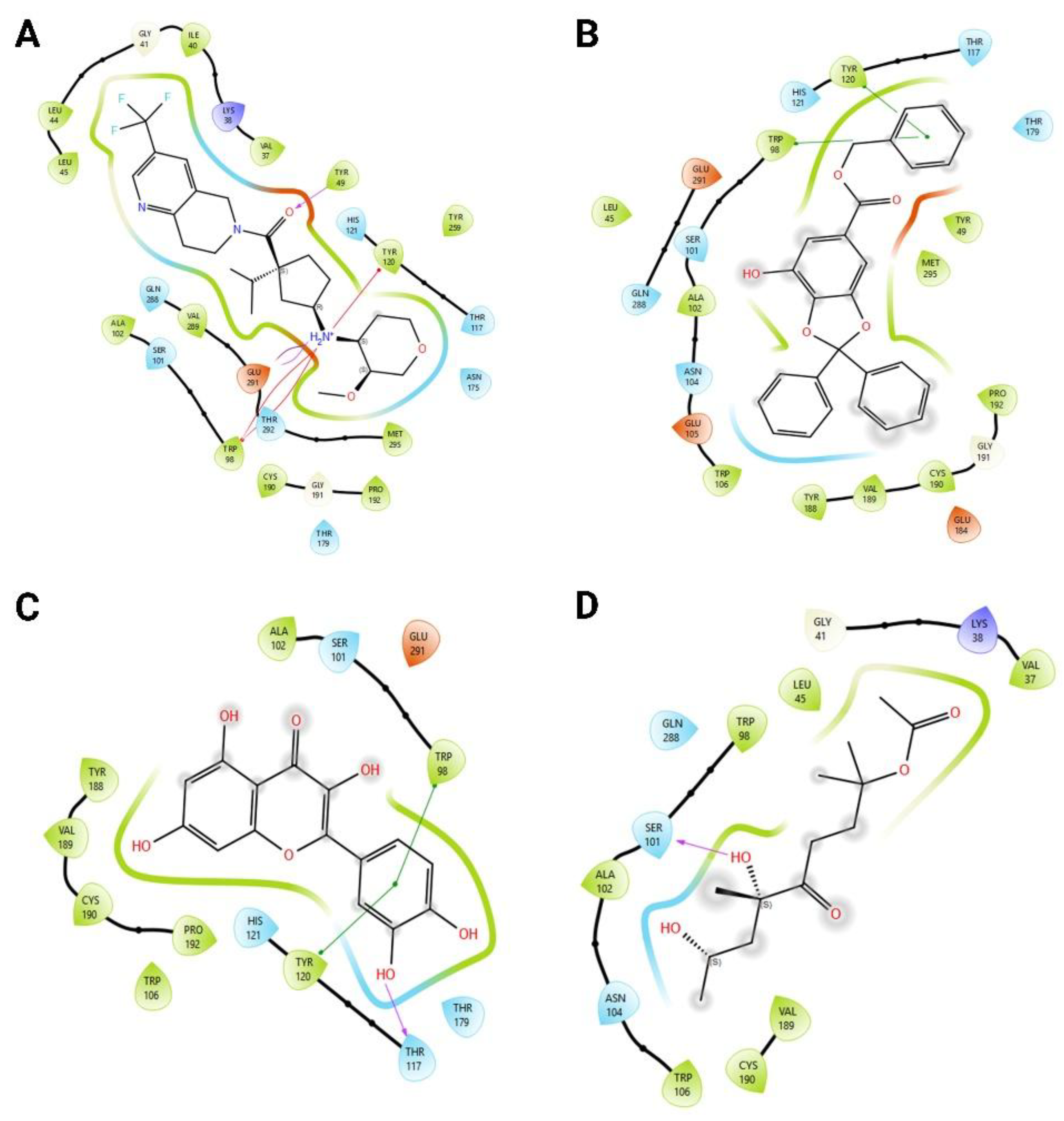
Figure 9.
Shared amino acid cluster analysis between F7N, gallic acid, quercetin, and cucurbitacin B.
Figure 9.
Shared amino acid cluster analysis between F7N, gallic acid, quercetin, and cucurbitacin B.
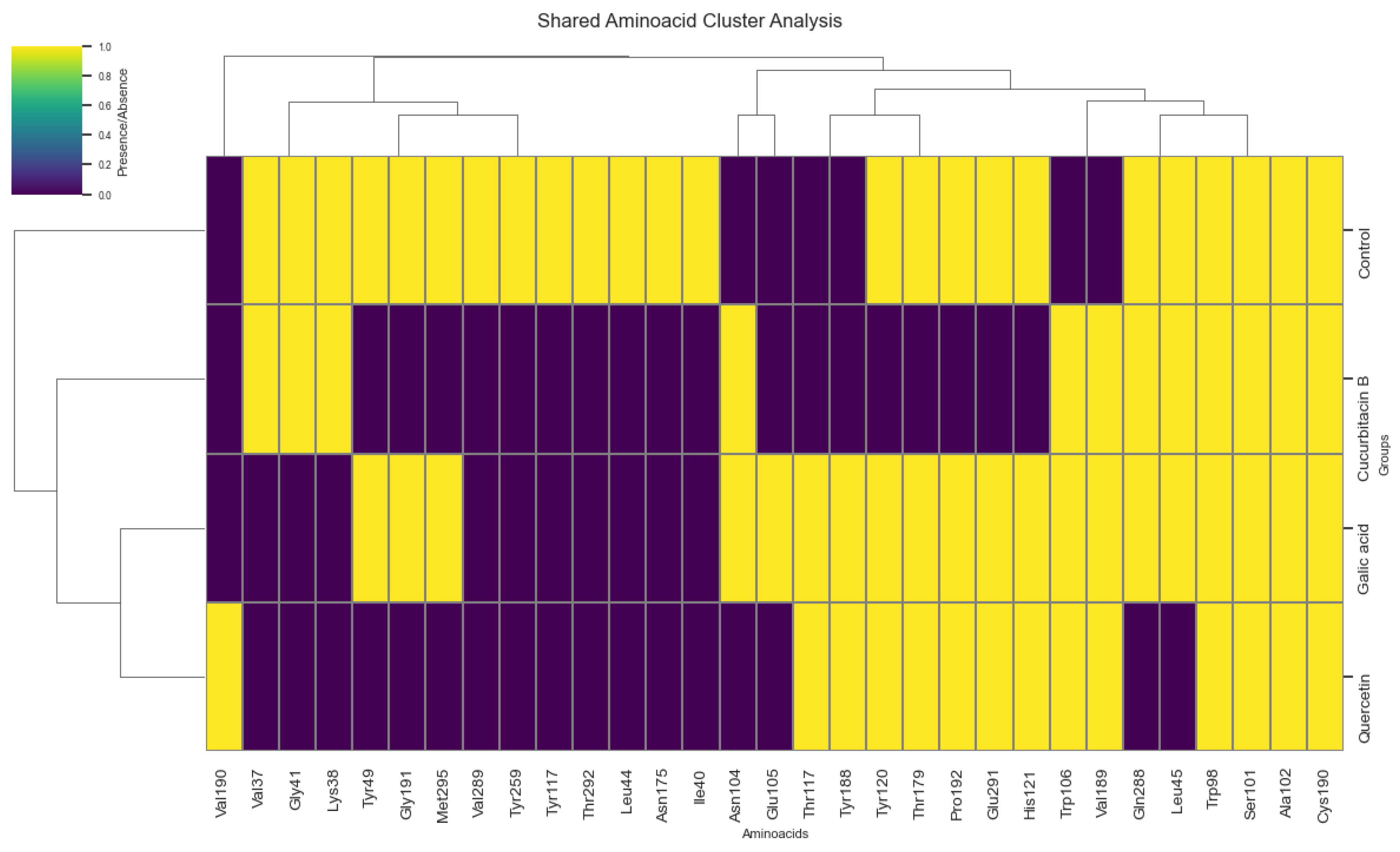
Table 3.
Percentage of cells positive for Annexin V-conjugated FITC labeling bound to PS per quadrant in mouse bone marrow mononuclear cells CD-1 for each treatment. Q1 represents necrosis; Q2 cells are in late apoptosis; Q3 represent normal cells; and Q4 represent cells in early apoptosis.
Table 3.
Percentage of cells positive for Annexin V-conjugated FITC labeling bound to PS per quadrant in mouse bone marrow mononuclear cells CD-1 for each treatment. Q1 represents necrosis; Q2 cells are in late apoptosis; Q3 represent normal cells; and Q4 represent cells in early apoptosis.
| Cells per quadrant (%) | ||||
| Treatment | Q1 | Q2 | Q3 | Q4 |
| Control | 0.1 | 25.1 | 24.2 | 50.7 |
| Vehicle (PBS) | 0.1 | 21.9 | 37.0 | 41.1 |
| Ara-C (Cytarabine®) | 0.4 | 24.7 | 15.1 | 59.8 |
| H-387-07 | 0.0 | 20.8 | 35.2 | 43.9 |
| S. edule var. nigrum spinosum | 3.2 | 77.7 | 11.4 | 8.3 |
| S. compositum | 0.1 | 19.2 | 35.7 | 45.0 |
| S. chinantlense | 0.0 | 17.6 | 30.2 | 52.2 |
Table 4.
Descriptive analysis of ligand binding energies.
| Ligand | Average (kcal/mol) | Standard deviation | n |
| Gallic acid | -9.49 | 0.001 | 10 |
| Cucurbitacin B | -9.53 | 0.016 | 10 |
| Quercetin | -7.89 | 0.001 | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



