Submitted:
18 August 2025
Posted:
18 August 2025
You are already at the latest version
Abstract
Rhodoliths are calcareous red algae considered indicators of ocean acidification and key biodiversity hotspots due to their ability to host a variety of species within their three-dimensional structures. This work aims to review the available scientific on rhodolith-forming species: reports from literature, the Symbiota digital taxonomic inven-tory, field observations, and nucleotide databases. A total of 21 species is reported, pre-dominantly from the Corallinaceae family and the Lithophylloideae subfamily. Rhodo-liths have been reported in Bocas del Toro, the Gulf of Chiriqui, Coiba National Park (PNC), the Gulf of Panama, and at Las Perlas Archipelago. This review represents the first step in raising awareness about the presence of these organisms along Panama’s coast and advocating for their inclusion in the management plans of protected areas, such as PNC, a UNESCO World Heritage Site, where rhodoliths are not yet part of the recorded algae species list or the park´s conservation targets despite its ecological relevance. Knowledge remains limited, and their conservation status is uncertain, but the increasing sampling efforts, and integration of morphological and molecular studies will open new opportunities to improve the estimation of rhodolith diversity in Panama.
Keywords:
Coiba
; coralline algae
; rhodoliths beds
; checklist
; conservation
1. Introduction
Rhodoliths are free-living calcareous red algae primarily composed of calcium carbonate. These algae are considered habitat modifiers or oceanic bioengineers as they provide a stable habitat for communities of other marine species within their three-dimensional branched and interlaced thalli [1]. As such, their ecological importance has drawn increasing attention to the need for their conservation.
Accoding to Tuya et al. 2023 [36], rhodolith beds are globally distributed, occurring from tropical to polar regions, and they cover an estimated area of 4.12 million km2 worldwide—approximately 20% larger than the estimated global area of tropical coral reefs, and between 2.5 to 30 times greater than other well-studied coastal habitats such as kelp forests, seagrass meadows, and mangroves. Despite these figures, rhodolith-bed science still lags behind other coastal ecosystems in terms of research efforts and ecological understanding
Interest in the conservation of rhodoliths in other countries has increased due to their role as indicators of ocean acidification [1,2,3,4,5,6,7]. Rhodolith beds are considered threatened and protected in coastal habitats of New Zealand, Europe, Australia, Brazil and Mexico. However, in Central America, only Costa Rica has initiated research efforts on them. Notably, studies conducted around Isla del Coco have revealed the presence of extensive rhodolith beds, as documented by Díaz-Licona [8]. These beds not only provide structural habitat and support high biodiversity but also play a crucial role in calcium carbonate production and the delivery of essential ecosystem services such as sediment generation, carbon cycling, and benthic habitat stabilization. These findings highlight the ecological importance of rhodolith beds in the Eastern Tropical Pacific and the urgent need for expanded conservation and research across Central America.
In contrast, Panama remains largely unexplored regarding rhodolith presence and diversity. This gap is particularly evident in the Coiba National Park (PNC), a UNESCO World Heritage Site and a critical protected area that harbors a significant role in the conservation of marine biodiversity in the region. Located off the southwest coast of Panama, in the Gulf of Chiriquí, the PNC harbors significant marine habitats, including coral/algal beds, yet rhodoliths remain underexplored.
This lack of recognition is further evidenced by the most recent management plan for the PNC, which does not include rhodoliths in any of its prioritized conservation categories, such as species or ecosystems [9]. Although the plan acknowledges the significant extent of coral/algal coverage in its shallow bottoms, the absence of rhodoliths as a conservation object highlights a gap in the area’s conservation efforts, despite their ecological importance.
Although the Isthmus of Panama provides suitable substrates for rhodoliths development, research and documentation on these ecosystems remain scarce. In this review, we provide a comprehensive overview of the calcareous red algal species that form rhodoliths in Panama, including georeferenced field observations, herbarium records, available DNA sequences, and other ecological data on rhodoliths worldwide. This information is essential for advancing future taxonomy, biogeography, conservation, phylogeography, genetics, and ecology of rhodoliths in the region. Our study contributes to the expanding body of literature on Central America, helping to reduce knowledge gaps and encourage further scientific attention to these overlooed habitats.
2. Materials and Methods
2.1. Literature Review
A review of the literature from 1910 to 2024 was conducted, including reports of rhodolith species for Panama, the digital taxonomic inventory Symbiota from the Smithsonian Tropical Research Institute (https://panamabiota.org/stri/projects/index.php?pid=18), the macroalgal herbarium consortium website of the U.S. National Science Foundation and several field observations. For molecular data, the NCBI website (https://www.ncbi.nlm.nih.gov/) and BOLD Systems (http://www.barcodinglife.org/) were consulted. Georeferencing maps were created using ArcGIS® software, and updates to scientific name updates were cross-referenced with the AlgaeBase database (https://www.algaebase.org).
2.2. General Settings of Rodolith Beds at Coiba National Park
A random sampling was conducted through underwater survey using standard SCUBA techniques in an area approximately 250-500 m2 at sites ranging from 8 to 17 meters in depth. To measure the percentage of coverage of the rhodolith beds, three photographs were taken per site using a Canon EOS R6 camera with an 85 mm macro lens. The images were processed using the ImageJ software, where the “particle analysis” tool was applied to quantify the coverage. The percentage of coverage was determined by comparing the area of the rhodolith beds to the total area of each image, and an average coverage was determined for each site. Additionally, observations on the associated marine fauna and flora, as well as depth were documented.
3. Results and Discussion
3.1. Historical Review
The history of rhodoliths research in Panama dates back more than a century ago (Figure 1a), when in 1910, Marshall Howe ventured into the Isthmus and first documented Corallinaceae species in the Bay of Panama. Although the known marine flora was limited at that time, Howe found rhodoliths in sites such as Isla Taboga, Urava, and Taboguilla, as well as in the Canal Zone [10]. Eight years later, in 1918, Howe returned to the region and reported four species of Lithothamiaceae in the Panama Canal, highlighting the geological significance of his findings, as well as their relevance to fossil species [11].
Interest in these algae continued to grow over the following decates. In 1929, the naturalist Lemoine, as part of a British expedition on the Saint George cruiser, documented several species of Corallinaceae on the Pacific coast of Panama, specifically in Taboga, Perlas, Coiba, and Jicaron. It was during this expedition that Lithophyllum coibense, a new species, was described [12]. Later, in 1945, Taylor conducted a detailed study on Isla Secas and other areas of the Panamanian Pacific, reporting rodoliths-forming species, such as Lithothamnion, which was found covering corals subtrates in Bahia Honda, Veraguas [13].
In the following decades, other researchers, incuding Dawson in the 1960s [14] and Sylvia Earle in 1972, futher expanded the records on calcareous algae capable of forming rhodoliths. Earle, for example, reported a total of 17 rhodolith species for the Pacific coast and six for the Caribbean coast [15]. It is important to note that all taxonomic classifications during this period were based exclusively on morphological characteristics, as molecular methods were not yet available.
Thirty-two years later, in 2004, a bathymetric survey revealed extensive rhodoliths beds in the shallow waters of Coiba National Park [16], although the specific species remained unidentified. Later, in 2008, Littler and Littler expanded the knowledge of biodiversity in the Gulf of Chiriqui, by documenting large rhodolith beds dominated by species such as Lithophyllum divaricatum and Lithothamnion indicum [17]. More recently, research has taken a more technical and integrative approach. Since 2013, anatomical and molecular studies have started to shape a new perspective on rhodoliths in Panama. Martínez [18], reported three species of Lithophyllum in the Caribbean coast, based on morphometric data, thereby expanding knowledge on their distribution. Other molecular studies, such as those by Richards and collaborators, have included samples from Panama, proving new insights in understanding genetic diversity of these calcareous marine algae [19,20,21,22].
Over the decades, research on rhodoliths in Panama has progressed gradually, with most reports resulting from expeditions and literature reviews (Figure 2a). Each new discovery has shed light on the ecological importance of these calcareous algae, which continue to attract a growing interest in marine science community. In recent years, the number of publications providing molecular information on Panama’s rhodoliths has been steadily increasing (Figure 2b), further enhancing our understanding of their biological relevance.
3.2. Checklist of Rhodoliths Species in Panama
A total of 21 rhodoliths-forming species has been reported for Panama, including 11 from the family Corallinaceae, five from Hapalidiaceae, and two from Sporolithaceae. Table 1 shows the list of rodoliths species based on published literature and student theses. The taxa are organized alphabetically by family and subfamily.
3.3. Reports to Be Confirmed
Field observation and morphological studies are considered as records requiring confirmation (Table 2), due to the limitation such as detalied taxonomic identification, the absence of molecular evidence to support the morphological observations, or the need for additional studies to verify the presence of these species in the reported locations. Additionally, some of these records are based on non-systematic observations, which hinders the ability to validate them conclusively without more comprehensive and rigorous analyses
Several studies have documented various species of calcareous algae that form rhodoliths in Panama; however, some of these species still require confirmation. For example, the genera Fosliella has been recorded as a rhodolith-forming species, yet studies from the South Pacific do not include this genus among rhodolith-associated taxa. Similarly, Phymatholiton masonianum exhibits anatomical characteristics consistent with specimens from Australia, but genetic information from the type species is crucial to validate this identification.
Other species, such as Hydrolithon breviclavium and Lithophyllum corallinae, have not been previously reported for Central America, making their presence in Panama uncertain until verified through detailed molecular and anatomical studies. Although these species are not officially registered for Panama, recent field observations suggest L. corallinae may be present along the Pacific coast [33].
3.4. Misreportings
Some reports have been considered invalid, due to incorrect distribution data. For instance, species such as Lithophyllum fetum, Lithophyllum lividum, and Lithophyllum propinquum var. cocosicum were listed for the Pacific of Panama in the review lists of Fernández García et al. [24] and Earle [15]. However, these species were initially reported from Isla del Coco, Costa Rica [12].
Similarly, Dermatolithon saxicola, also listed by Earle [15] for the Pacific of Panama, was recorded by Lemoine’s [12] at Isla del Coco, Costa Rica. Likewise, Lithothamnion indicum var. subtilis and Lithothamnion mesomorphum, mistakenly reported by Earle [15] to Panama, were first recorded from Isla Gorgona, Colombia [12], not Panama.
These misreportings highlight the need of verifying species records, especially for taxes with overlapping geographical distributions. Comprehensive molecular and anatomical studies are essential to clarify the true occurrence of these species in Panama and ensure accurate biogeographical mapping.
3.5. Localization and Diversity
Georeferencing data provided by the Symbiota digital database of the Smithsonian Tropical Research Institute, along with records from other international collections, are important to identifying the distribution and extent of rhodolith beds in Panama (Supplementary data, S1).
Based on these records, four major regions within the Isthmus of Panama have been identified as important areas for rhodolith occurrence: the Bocas del Toro Archipelago in the Caribbean, and the Gulf of Chiriqui, Coiba National Park and the Pearl Islands Archipelago in the Pacific. Additional records from Colon, Taboga, the Canal Zone and San Blas, each with at least four georeferenced point suggest that these locations warrant to further sampling efforts to enhance the current understanding of rhodolith distribution in the region (Figure 3).
The greatest diversity is found in Panama’s Pacific (Figure 3), with the Coiba National Park standing out as the area with the highest species richness (Figure 4). The vast sandy and rocky, soft-bottom substrates predominant in PNC provide ideal conditions for the establishment of rhodolith communities, forming extensive beds [9,16].
Ten genera of rhodoliths-forming species are reported in Panama. Similar to observations by Robinson et al. [34] for the Tropical Eastern Pacific, the order Corallinales dominates the assemblage, accounting for 66.7% of the species (Figure 5a), with the subfamily Lithophylloideae representing 44% (Figure 5b). Within this group, the genus Lithophyllum emerges as the most frequently recorded along Panama’s coasts.
3.6. Molecular Studies Data
Twenty-eight genetic sequences of calcareous rhodolith-forming algae species from Panama have been recorded, corresponding to the genes co1, cox2, lsu, upa, rbcl, and psba, and retrieved from the NCBI and BOLD Systems database. The psba gene has been the most used for species characterization s in the country (Appendix A1).
Recent studies have incorporated Panamanian species into their molecular analyses. Richards et al. [22] included Panama’s sequences in a phylogenetic analysis of rhodolith diversity in the northwest Gulf of Mexico. In 2021, they confirmed the presence of Harveylithon munitum in Panama and suggested a potential phylogenetic relationship with Harveylithon maris-bahiensis from Brazil [21].
In addition, Richards et al. [20], confirmed the occurrence of Sporolithon episporum in the Caribbean coast of Panama and proposed that Sporolithon samples from the Gulf of Chiriquí may correspond to either S. howei or S. pacificum.
Richards et al. [19] provided both molecularly and morphologically characterization of two Lithothamnion specimens from Panama’s Pacific, though their precise identification requires further analysis using additional molecular markers. Robinson [31] was the to report Lithophyllum okamurae in the Las Perlas Archipelago, based on combined morphological and molecular evidence.
These studies have expended the of molecular dataset available for rhodolith-forming species in Panama. Further research using other molecular markers is needed to resolve incomplete phylogenetic relationships and understand the genetic diversity of these organisms. The inclusion of bioinformatics tools will be key for advancing the interpretation of rhodolith diversity patterns in the region.
3.7. Rhodolith Beds at Coiba National Park (PNC)
According to the updated 2024 management plan for PNC, coral/algal assemblages dominate the marine benthos, covering 35.17% of the total protected area (equivalent to 2,877.84 hectares) [9]. This extensive coverage plays a critical role in the structure and function of reef ecosystems, offering essential habitats for a wide range of marine species [9,16].
Substrate distribution analyses from the PNC management plan show that these benthic habitats, found at depths of up to 10 meters around the Coiba island, are key in maintaining ecological structure of the protected marine area [9]. The benthic structure of the area reflects a combination of corals and algae assemblages that promote marine biodiversity and the ecological dynamics of the region.
Notably, the benthic structures currently referred to as “coral/algal” in the 2024 plan were previously described as “rhodolith beds” in the 2014 management plan, a classification supported by our field observations.
Our surveys confirm the presence of rhodolith beds at the NE of the PNC (Table 3, Figure 6), with coverage ranging between 46% and 69%, depending on the location. The Canales de Afuera exhibited a rhodolith coverage of 46%, while in Buffet the coverage was 49% and in Don Juan reached 65%. The highest coverage to date was found at Iglesias, with 69% of rhodoliths coverage.
The sediments associated with the beds in the study sites are mainly composed of gravelly, muddy sand and basaltic rocks. The rhodolith beds to the NE of the PNC host an associated diversity of fauna and other macroalgae (Figure 6 and Supplementary data, video, S2).
3.8. Ecological Role, Threats and Conservation
Rodoliths beds are recognized as both Ecologically or Biologically significant Marine Areas (EBSA) and Small Natural Features (SNF) due to their exceptional role in marine ecosystems. EBSAs are considered areas of significant ecological value because they contribute to biodiversity conservation and play a vital role in maintaining the health and function of marine ecosystems [37]. As SNFs, they are classified as small but ecologically crucial units that have a significant impact on their surrounding environments [38].
In addition to their role in seabed stabilization and habitat, rhodoliths provide a wide range of ecosystem services [39,40]. They act as biodiversity hotspots [1,17,31,40,41,42], support the growth and development of other species (i.e., commercial species) [31,40,41,43,45] and contribute to coastal sediment production [1,44].
Also offer valuable insights into paleoclimatic predictions [31,47], ocean acidification [2,46,48] and serve as important areas for recreation and tourism [39]. Furthermore, they help prevent and moderate disturbances [1,40], playing a critical role in maintaining ecosystem resilience.
Despite their ecological importance, rhodoliths face various threats. Anthropogenic impacts such as coastal pollution, urban development in coastal areas, and trawling fishing practices alter water quality, increase sedimentation, and damage the physical structures of rhodoliths beds [2,40,49,50,51]. Climate change, in turn, causes ocean acidification and rising sea temperatures, affecting the availability of calcium carbonate essential for their growth which can compromise their survival [2,52]. Additionally, the lack of research and monitoring in areas such as Panama hampers a full understanding of their ecology, delaying the implementation of adaptive conservation measures.
To address these threats, we propose the following conservation strategies based on Coiba National Park management plan [8] and other documents whose refers frameworks for the study and conservation of rhodoliths in other latitudes [34,39,40,53,54,55] (Figure 7):
• Integration into management plans: Incorporate rhodoliths as objectives of conservation into the management plans of marine protected areas where their presence is known, as well as other marine areas.
• Research and Monitoring: Promote research to understand the diversity of rhodoliths and to explore the ecology, distribution, and genetics of rhodoliths beds in Panama. Establish long-term monitoring programs to assess the condition of these algae and their response to threats, as well as evaluate the conservation status of their populations.
• Education: Raise awareness about the ecological importance of rhodoliths among decision-makers, local communities, and the public.
• Collaboration and funding: Encourage collaboration between researchers, conservation organizations, and government agencies to address the conservation challenges of rhodoliths beds. Seek funding to support research, monitoring, and conservation initiatives.
• Regulation and fisheries management: Implement fisheries regulations to protect rhodoliths beds from trawling in marine that are not protected. In protected areas, strengthen and enforce regulations to maintain ecosystem sustainability.
In contrast, in Panama’s Caribbean side of Panama shows a notably lower number of reported species, likely due to limited sampling efforts in the region However, favorable conditions in the region, such as depth, temperature, site accessibility and high diversity of other marine algae species [30,35,36], suggest that species richness may be underestimated. Recent studies have documented rhodolith specimens from the Bocas del Toro Archipelago [20,21,26] and areas near Colón [18], indicating that further exploration could reveal greater diversity in the Caribbean sector.
4. Conclusions
A total of 21 species of calcareous algae that form rhodoliths have been reported for Panama. Coiba National Park is the site with the highest occurrence of reports and extensive rhodoliths bed areas. Despite their importance and predominant abundance along Panama’s Pacific coast, rhodoliths beds remain an underexplored ecosystem. As such, significant research efforts are needed to better understand the diversity of these calcareous algae. The various species reports must be studied not only to clarify the correct application of scientific names but also to ensure that these names are properly applied to Panama’s material through morphological, anatomical, and molecular studies. This will provide a more definitive and reliable species list. Including Panama’s rhodolith beds in the list of threatened habitats and developing appropriate conservation strategies for these species should be the goal of future research.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Georeferencing available data of rhodoliths reports for Panama; Video S2: Rhodoliths, ocean bioengineers from Coiba.
Author Contributions
N.L., wrote the original draft, funding acquisition, carry out the research; C.F.G, I.V. and E.D.F. wrote, review and editing, B.W., provision of database, collecting and planned some collecting expeditions, C.F.G, provision of literature, supervision and participated of the collecting expedition organized by B.W., I.V. critical review and E.D.F, supervision and funding acquisition.
Funding
This study was funded by Coiba AIP Scientific Station, “Mar del Sur” thesis grant program and University of Panama grant for research VIP-01-04-07-2023-29.
Institutional Review Board Statement
Not applicable
Data Availability Statement
Supplementary data are available in http://dx.doi.org/10.13140/RG.2.2.35769.89441. DNA sequence can be accessed in GenBank under accession numbers register in appendix A.1.
Acknowledgments
We can acknowledge Eduardo Estrada and Sara Justo for the support in field trip and Rosa Leon for help with the georeferencing map.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| PNC | Coiba National Park |
| CMAR | Eastern Tropical Pacific Marine Corridor |
| EBSA | Ecologically or Biologically significant Marine Areas |
| SNF | Small Natural Features |
| NE | North East |
Appendix A

Table A1.
Localities and GenBank access numbers for available sequences of rhodolith-forming species for Panama.
Table A1.
Localities and GenBank access numbers for available sequences of rhodolith-forming species for Panama.
| # | Specie | Locality | ID | GenBank Accesion | |||||
|---|---|---|---|---|---|---|---|---|---|
| COX2 | LSU | COI | rbcL | UPA | psbA | ||||
| 1 | Harveylithon sp. | Wild Cay, BT | PHYKOS 7053 | ---- | ---- | ---- | ---- | ---- | MW452886 |
| 2 | H. munitum | Escudo de Veraguas, BT | PHYKOS_3593 | ---- | MW457636.1 | ---- | MF979962 | ---- | ---- |
| 3 | Lithophyllum sp. | Cebaco Island, VE | LAF7219 | ---- | KJ412333.1 | KJ418417.1 | ---- | ---- | KJ418411.1 |
| 4 | Lithophyllum sp. 3 | Swan Cay, BT | FBCS12912 | KJ801356.1 | ---- | ---- | ---- | ---- | ---- |
| 5 | Lithophyllum sp.3 | Sand Fly Bay, BT | FBCS12913 | KJ801357.1 | ---- | ---- | ---- | ---- | ---- |
| 6 | Lithothamnion sp. 4 | Swan Cay, BT | FBCS12920 | KJ801364.1 | ---- | ---- | ---- | ---- | ---- |
| 7 | Lithothamnion sp. D | Gulf of Chiriqui | LAF6631 | ---- | ---- | ---- | ---- | KU519740 | KU557500 |
| 8 | Lithothmanion 1 | Swan Cay, BT | FBCS12917 | KJ801365.1 | ---- | ---- | ---- | ---- | ---- |
| 9 | Lithothmnion sp. J | Tintorera Island, VE | PHYKOS7249 | ---- | KR075891.1 | KU504277 | ---- | KU504275 | KP844865 |
| 10 | L. neocongestum | Bocas del Toro | NCU 598862 | ---- | ---- | ---- | KX020485 | ---- | ---- |
| 11 | L. neocongestum | Bocas del Toro | US223011 | ---- | ---- | ---- | KX020484.1 | ---- | KX020466 |
| 12 | L. neocongestum | Bocas del Toro | US169412 | ---- | ---- | ---- | ---- | ---- | KX020486 |
| 13 | L. neocongestum | , Flat Rock Beach, BT | US170968 | ---- | ---- | ---- | ---- | ---- | KX020440 |
| 14 | L. neocongestum | Sand Fly Bay, BT | US170967 | ---- | ---- | ---- | ---- | ---- | KX020441 |
| 15 | Neogoniolithon sp | Panama | VPF00177 | ---- | ---- | KM392370.1 | ---- | ---- | ---- |
| 16 | S. episporum | Punta Toro, Colón | NY_900041 | ---- | ---- | ---- | KY994125.1 | ---- | ---- |
| 17 | S. episporum | Bocas del Toro | NCU_598843 | ---- | ---- | KY994113.1 | KY994124.1 | ---- | MF034547.1 |
| 18 | Sporolithon sp. | Mono Feliz, GC | PHYKOS_4623 | ---- | ---- | ---- | ---- | ---- | MF034548.1 |
References
- Riosmena-Rodríguez, R. Chapter 1: Natural History of Rhodolith/Maërl Beds: Their Role in Near-Shore Biodiversity and Management. In Rhodolith/maërl beds: a global perspective, 1st ed.; R. Riosmena- Rodríguez, W. Nelson, J. Aguirre, Eds.; Springer, Suiza, 2017; pp. 3–26.
- Fredericq, S.; Krayesky, S.; Sauvage, T.; Richards, J.; Kittle, R.; Arakaki, N.; Schmidt, W. The Critical Importance of Rhodoliths in the Life Cycle Completion of Both Macro- and Microalgae, and as Holobionts for the Establishment and Maintenance of Marine Biodiversity. Front. Mar. Sci 2018, 5, 505. [Google Scholar] [CrossRef]
- Kamenos, N.; Burdett, H.; Darrenougue, N. Chapter 2: Coralline Algae as Recorders of Past Climatic and Environmental Conditions. In Rhodolith/maërl beds: a global perspective, 1st ed.; R. Riosmena- Rodríguez, W. Nelson, J. Aguirre, Eds.; Springer, Suiza, 2017; pp.27-53.
- Martin, S.; Hall-Spencer, J. Chapter 3: Effects of Ocean Warming and Acidification on Rhodolith/Maërl Beds. In Rhodolith/maërl beds: a global perspective, 1st ed.; R. Riosmena- Rodríguez, W. Nelson, J. Aguirre, Eds.; Springer, Suiza, 2017; pp. 55–85.
- Ragazzola, F.; Foster, L. C.; Form, A.; Anderson, P. S.; Hansteen, T. H.; Fietzke, J. Ocean acidification weakens the structural integrity of coralline algae. Glob. Change Biol. 2012, 18, 2804–2812. [Google Scholar] [CrossRef]
- Andersson, A. J.; Bates, N. R. .; Mackenzie, F. T. Dissolution of carbonate sediments under rising pCO2 and ocean acidification: observations from Devil’s Hola, Bermuda. Aquat. Geochem. [CrossRef]
- Morse, J. .; Andersson, A.; Mackenzie, F. Initial responses of carbonate-rich shelf sediments to rising atmospheric pCO2 and “ocean acidification”: Role of high Mg-calcites. Geochim. Cosmochim. Acta 2006, 70, 5814–3580. [Google Scholar] [CrossRef]
- Diaz-Licona, C.; Schubert, N.; González-Gamboa, V.; Tuya, F.; Azofeifa-Solano, J.; Fernández-García, C. Rhodolith beds in the Eastern Tropical Pacific: Habitat structure and associated biodiversity. Aquat. Bot. 2025, 2025 201, 103914. [Google Scholar] [CrossRef]
- Ministerio de Ambiente. Plan de Manejo del Parque Nacional Coiba 2024. Panamá. https://miambiente.gob.pa/download/plan-de-manejo-del-parque-nacional-coiba-borrador-final-may24/.
- Howe, M. Report on a botanical visit to the Isthmus of Panama. J. N Y Bot. Gard. 1910, 11, 30–44. [Google Scholar]
- Howe, M. On some fossil and recent Lithothamnieae of the Panama Canal Zone. Bull. U.S. Nat. Mus. 1918, 103, 1–13. [Google Scholar]
- Lemoine, M. P. Les Corallinacees de l’ Archipel des Galapagos et du Golfe de Panama. Arch. Mus. Hist. Nat. 1929, 6, 47–88. [Google Scholar]
- Taylor, W. R. Pacific marine algae of the Allan Hancock expeditions to the Galapagos Islands. Allan Hancock Pac. Exp. 1945, 12, 1–528. [Google Scholar]
- Dawson, E. New records of marine algae from Pacific Mexico and Central America. Pac. Nat. 1960, 1, 31–52. [Google Scholar]
- Earle, S. A review of the marine plants of Panama. Bull. Biol. Soc. Wash. 1972, 2, 69–87. [Google Scholar]
- Guzmán, H.; Guevara, C.; Breddy, O. Distribution, diversity, and conservation of coral reefs and coral communities in the largest marine protected area of Pacific Panama (Coiba Island). Environ. Conserv. 2004, 31, 111–121. [Google Scholar] [CrossRef]
- Littler, M.; Litller, D. Coralline algal rhodoliths form extensive benthic communities in the Gulf of Chiriqui, Pacific Panama. Coral Reefs 2008, 27(3), 553. [Google Scholar] [CrossRef]
- Matrínez, J. Extensiones de rango y nuevas especies de algas coralinas (Corallinales, Rodophyta) formadoras de mantos de rodolitos en el Caribe. Bachelor Thesis, Universidad Autónoma de Baja California Sur, La Paz, Mexico, 2013.
- Richards, J.; Vieira-Pinto, T.; Schmidt, W.; Sauvage, T.; Gabrielson, P.; Oliveira, M.; Fredericq, S. Molecular and Morphological Diversity of Lithothamnion spp. (Hapalidiales, Rhodophyta) from Deepwater Rhodolith Beds in the Northwestern Gulf of Mexico. Phytotaxa 2016, 278, 81–114. [Google Scholar] [CrossRef]
- Richards, J.; Sauvage, T.; Schmidt, W.; Fredericq, S.; Hughey, J.; Gabrielson, P. The coralline genera Sporolithon and Heydrichia (Sporolithales, Rhodophyta) clarified by sequencing type material of their generitypes and other species. J. Phycol 2017, 53, 1044–1059. [Google Scholar] [CrossRef]
- Richards, J.; Schmidt, W.; Fredericq, S.; Sauvage, T.; Peña, V.; Gall, L. ; Mateo-Cid, L;, Mendoza-González, A.; Hughey, J.; Gabrielson, P. DNA sequencing of type material and newly collected specimens reveals two heterotypic synonyms for Harveylithon munitum (Metagoniolithoideae, Corallinales, Rhodophyta) and three new species. J. Phycol. 2021, 57. [Google Scholar] [CrossRef]
- Richards, J.; Kittle, R.; Schmidt, W.; Sauvage, T.; Gurgel, C.; Gabriel, D.; Fredericq, S. Assessment of Rhodolith Diversity in the Northwestern Gulf of Mexico Including the Description of Sporolithon gracile sp. nov. (Sporolithales, Rhodophyta), and Three New Species of Roseolithon (Hapalidiales, Rhodophyta). Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Dawson, E. Y. A guide to the literature and distributions of Pacific benthic algae from Alaska to the Galapagos Islands. Pac. Sci. 1961, 15, 370–461. [Google Scholar]
- Fernández-García, C.; Riosmena-Rodríguez, R.; Wysor, B.; Tejeda O., L.; Cortés, J. Checklist of the Pacific marine macroalgae of Central America. Bot. Mar. 2011, 2011. 54, 53–57. [Google Scholar] [CrossRef]
- Taylor, W. Caribbean Marine Algae of the Allan Hancock Expedition, Rep. Allan Hancock Atlant. Exped. 1939, 2, 193. [Google Scholar]
- Hernandez-Kantun, J.; Gabrielson, P.; Hughey, J.; Pezzolesi, L.; Rindi, F.; Robinson, N.; Peña, V.; Riosmena-Rodriguez, R.; Gall, L.; Adey, W. Reassessment of branched Lithophyllum spp. (Corallinales, Rhodophyta) in the Caribbean Sea with global implications. Phycolog. 2016, 55, 619–639. [Google Scholar] [CrossRef]
- Taylor, W. Marine Algae of the Eastern Tropical and Subtropical Coasts of the Americas. The University of Michigan Press, Ann Arbor, 1960, pp. 870.
- Taylor, W. R. Notas sobre las algas del Océano Atlántico tropical. Revista americana de botánica 1929, 621-630.
- Gabrielson. P.; Hughey. R.; Diaz-Pulido, G. Genomics reveals abundant speciation in the coral reef building alga Porolithon onkodes (Corallinales, Rhodophyta) J. Phycol. 2018, 54, 429–434. [CrossRef]
- Gómez, D.; Pérez, H. Estimación de la riqueza y abundancia de macroalgas en los arrecifes de Punta Galeta, 15 años después de un derrame de petróleo. Bachelor Thesis, Escuela de Biología, Universidad de Panamá, 2004. [Google Scholar]
- Robinson, N. Sistemática de las especies de algas coralinas (Corallinophycidae, Rodophyta) formadoras de mantos de rodolitos en el Pacífio Tropical Oriental. Master Thesis, Universidad Autónoma de Baja California Sur, México, 2013. [Google Scholar]
- Dawson, E. Marine algae from the 1958 Cruise of the Stella Polaris in the Gulf of California. Los Angeles County Mus.Contr. Sci. 1959, 27, 1–39. [Google Scholar] [CrossRef]
- Caracterización morfoestructural y molecular de rodolitos (Corallinales ) del Parque Nacional Coiba. XII Congreso de Ficología de Latinoamérica y el Caribe, Costa Rica, November 18-22, 2024.
- Robinson, N.; Fernández-García, C.; Riosmena-Rodríguez, R.; Rosas-Alquicira, E.; Konar, B.; Chenelot, H.; Jewett, S.; Melzer, R.; Meyer, R.; Försterra, G.; Häussermann, V.; Macaya, E. Chapter 13: Eastern Pacific. In Rhodolith/maërl beds: a global perspective, 1st ed.; R. Riosmena- Rodríguez, W. Nelson, J. Aguirre, Eds.; Springer, Suiza, 2017; pp. 319–333.
- Wysor, B.; Kooistra, W. An annotated list of marine Chlorophyta from the Caribbean coast of the Republic of Panama. Nova Hedwig. Beih 2003, 77, 487–523. [Google Scholar] [CrossRef]
- Wysor, B. An annotated list of marine Chlorophyta from the Pacific Coast of the Republic of Panama with a comparison to Caribbean Panama species. Nova Hedwig. Beih 2004, 78, 209–241. [Google Scholar] [CrossRef]
- Tuya, F.; Schubert, N.; Aguirre, J.; Basso, D.; Bastos, E.; Berchez, F.; Bernardino, A.; Bosch, N.; Burdett, H. ; Espino,F.; Fernández-Garcia, C.; Francini-Filho, R.; Gagnon, P.; Hall-Spencer, J.; Haroun, R.; Hofmann, L.; Horta, P.; Kamenos, N.; Le Gall, L.; Magris, R.; Martin, S.; Nelson, W.; Neves, P.; Olivé, I.; Otero-Ferrer, F.; Peña, V.; Pereira-Filho, G.; Ragazzola, F.; Rebelo, A.; Ribeiro, C.; Rinde, E.; Schoenrock, K.; Silva, J.; Sissini, M.; Tâmega, F. Levelling-up rhodolith-bed science to address global-scale conservation challenges. Sci. Total Environ, 1648. [Google Scholar] [CrossRef]
- Hunter, M.; Acuña, V.; Bauer, D.; Bell, K.; Calhoun, A.; Felipe-Lucia, M.; Fitzsimons, J.; González, E.; Kinnison, M.; Lindenmayer, D.; Lundquist, C.; Medellin, R.; Nelson, E.; Poschlod, P. Conserving small natural features with large ecological roles: A synthetic overview. Biol. Conserv. 2017, 211(B), 88–95. [Google Scholar] [CrossRef]
- Amado-Filho, G.; Bahia, R; Guilherme, H.; Pereira-Filho, G.; Longo, L. Chapter 12: South Atlantic Rhodolith Beds: Latitudinal Distribution, Species Composition, Structure and Ecosystem Functions, Threats and Conservation Status. In Rhodolith/maërl beds: a global perspective, 1st ed.; R. Riosmena- Rodríguez, W. Nelson, J. Aguirre, Eds.; Springer, Suiza, 2017; pp. 299–314.
- Díaz Licona, C. Mantos de rodolitos (rhodophyta) del pacífico costarricense: caracterización e identificación de posibles servicios ecosistémicos para generar recomendaciones de manejo. Master Thesis, Universidad de Costa Rica, 2024.
- Amado-Filho, G. M.; Maneveldt, G. W.; Pereira-Filho, G. H.; Manso, R. C.; Bahia, R. G.; Barros-Barreto, M. B.; Guimarães, S. M. Seaweed diversity associated with a Brazilian tropical rhodolith bed. Cien. Mar. 2010, 36, 371–391. [Google Scholar] [CrossRef]
- Solano-Barquero, A; Sibaja-Cordero, J; Cortés, J. Macrofauna associated with a rhodolith bed at an oceanic island in the Eastern Tropical Pacific (Isla del Coco National Park, Costa Rica). Front. Mar. Sci. 2022, 9. [CrossRef]
- Documentando a los rodolitos:bioingenieros del Parque Nacional Coiba. XIX Congreso Nacional de Ciencia y Tecnología, Panama, September 27, 2023.
- Bassi, D.; Simone, L.; Nebelsick, J. Chapter 6: Re-sedimented Rhodoliths in Channelized Depositional Systems. In Rhodolith/maërl beds: a global perspective, 1st ed.; R. Riosmena- Rodríguez, W. Nelson, J. Aguirre, Eds.; Springer, Suiza, 2017; pp. 139–168.
- Acuña, F. .; Cortés, J.; Garese, A.; González-Muñoz, R. The sea anemone Exaiptasia diaphana (Actiniaria: Aiptasiidae) associated to rhodoliths at Isla del Coco National Park, Costa Rica. Rev. Biol. Trop 2020, 68, S283–S288. [Google Scholar] [CrossRef]
- Simon-Nutbrown, C.; Hollingsworth, P.; Fernandes, T.; Kamphausen, L.; Baxter, J.; Burdett, H. Species Distribution Modeling Predicts Significant Declines in Coralline Algae Populations Under Projected Climate Change with implications for conservation Policy. Front. Mar. Sci. 2020, 7, 575825. [Google Scholar] [CrossRef]
- Kamenos, N.; Burdett, H.; Darrenougue, N. Chapter 2: Coralline Algae as Recorders of Past Climatic and Environmental Conditions. In Rhodolith/maërl beds: a global perspective, 1st ed.; R. Riosmena- Rodríguez, W. Nelson, J. Aguirre, Eds.; Springer, Suiza, 2017; pp. 27–53.
- Ragazzola, F.; Foster, L.; Form, A.; Anderson, P.; Hansteen, T.; Fietzke, J. Ocean acidification weakens the structural integrity of coralline algae. Glob. Change Biol. 2012, 18, 2804–2812. [Google Scholar] [CrossRef]
- Birkett, D.; Maggs, C.; Dring, M. An overview of assessing and sensitivity characteristics for conservation management of marine SACs1998, Scottish Association for Marine Science (UK Marine SACs Project).
- Hall-Spencer, J. Conservation issues relating to maerl beds as habitats for molluscs. J. Conchol. 1998, 2, 271–286. [Google Scholar]
- Hall-Spencer, J.; Grall, J.; Moore, P.; Atkinson, R. Bivalve fishing and maerl-bed conservation in France and the UK-retrospect and prospect. Aquat. Conserv. 2003, 13, S33–S41. [Google Scholar] [CrossRef]
- Adey, W.; Halfar, J.; Williams, B. The coralline genus Clathromorphum Foslie emend. Adey; biological, physiological and ecological factors controlling carbonate production in an Arctic/Subarctic climate archive. Smithsonian Contr. Mar. Sc. 2013. 40, 1–83. [CrossRef]
- Rebelo, A.; Teichert, S.; Bracchi, V.; Rasser, M.; Basso, D. Crustose coralline red algae frameworks and rhodoliths: Past and present. Front. Earth Sci. 2022, 10, 1090091. [Google Scholar] [CrossRef]
- Antunez Horta, P.; Riul, P.; Amado Filho, G.; Gurgel, C.; Berchez, F.; de Castro, J.; Figueiredo, M. Rhodoliths in Brazil: Current knowledge and potential impacts of climate change. Braz. J. Oceanogr. 2016, 64, 117–136. [Google Scholar] [CrossRef]
- Alvarado, J. J.; Herrera, B.; Corrales, L.; Asch, J.; Paaby, P. Identificación de las prioridades de conservación de la biodiversidad marina y costera en Costa Rica, Rev. Biol. Trop 2011, 59, 829–842. [Google Scholar] [CrossRef]
Figure 1.
Timeline about rodoliths research in Panama.
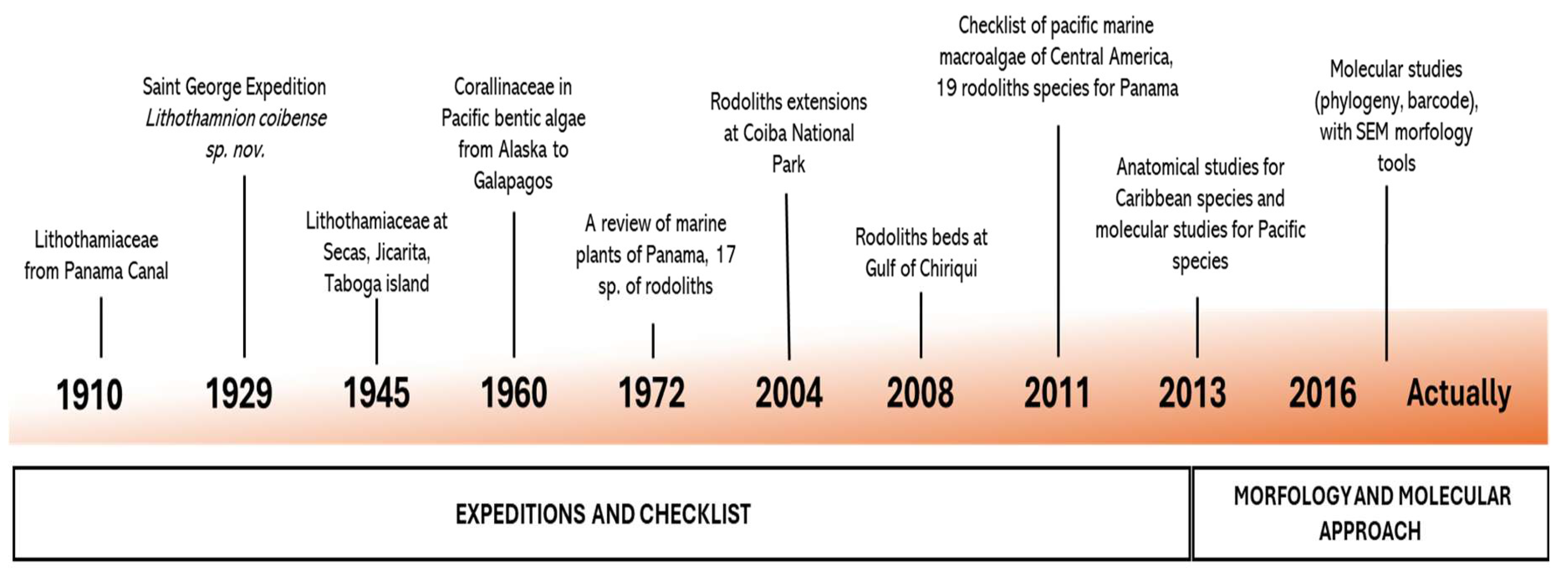
Figure 2.
(a) number of reported species in both stages of panamenian rhodolith research; and (b) number of different publised literature with panamenian rhodoliths reports.
Figure 2.
(a) number of reported species in both stages of panamenian rhodolith research; and (b) number of different publised literature with panamenian rhodoliths reports.
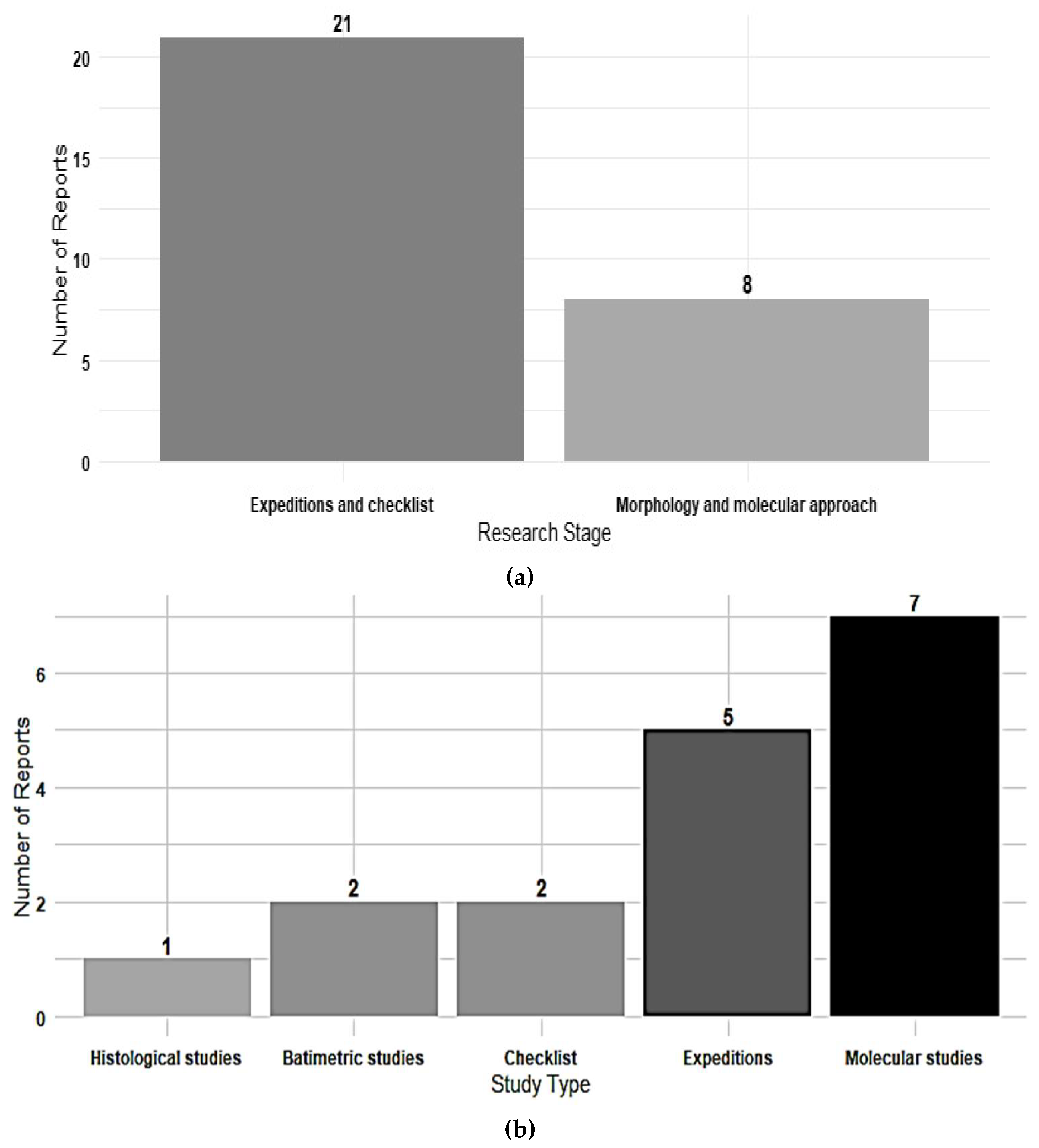
Figure 3.
Key areas with important extensions of rhodoliths in Panama; a) Coiba, b) Bocas del Toro, c) Gulf of Chiriqui and d) Las Perlas Archipelago. Squares with dotted lines in white represent sites with one or two reports.
Figure 3.
Key areas with important extensions of rhodoliths in Panama; a) Coiba, b) Bocas del Toro, c) Gulf of Chiriqui and d) Las Perlas Archipelago. Squares with dotted lines in white represent sites with one or two reports.
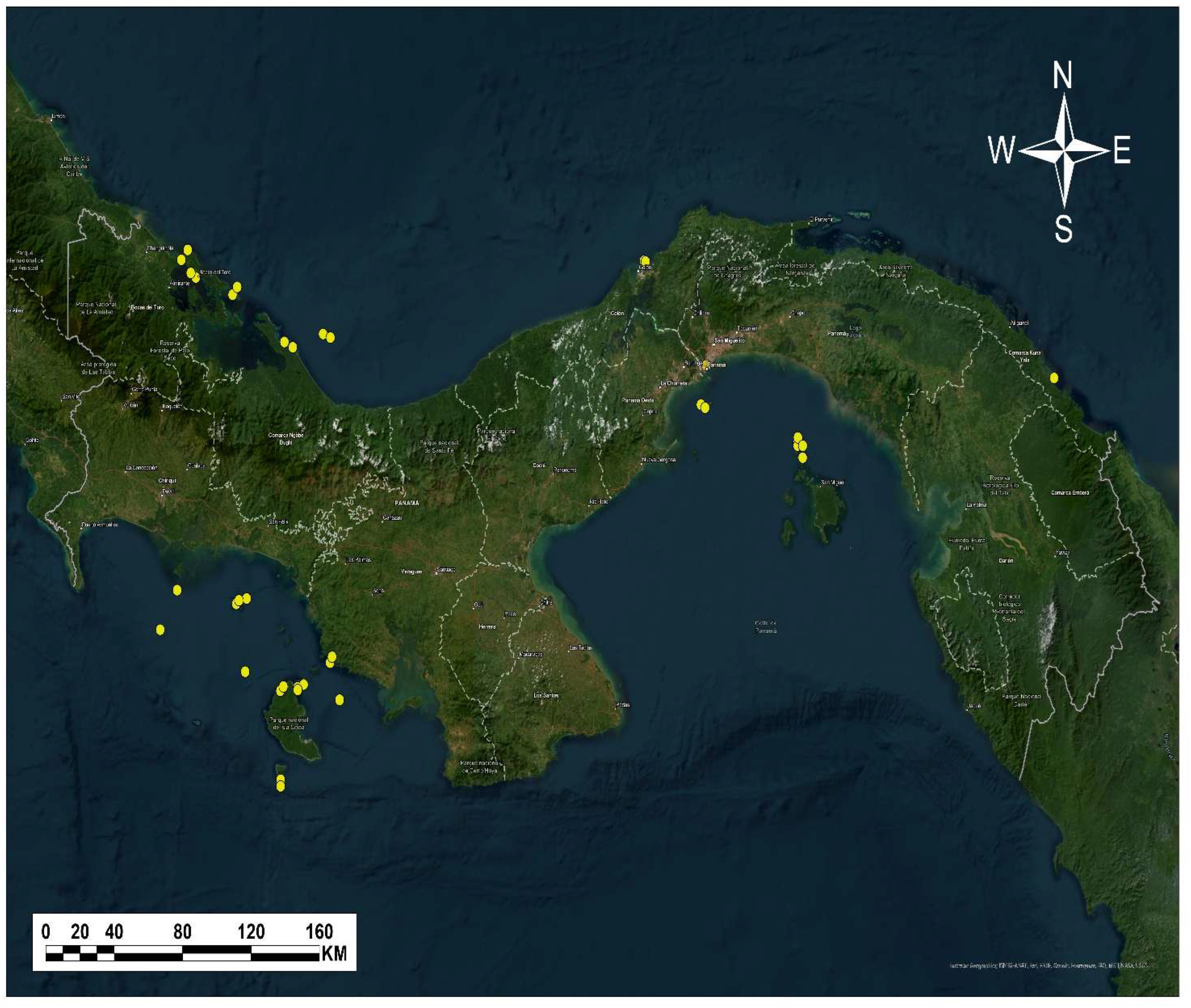
Figure 4.
Porcentage of rhodoliths reports by sites.
Figure 5.
Diversity of rhodoliths reported for Panama (a) by order and (b) by sub-family. LI= Lithophylloideae, ME= Melobesioideae, CH= Chamberlainoideae, SPO= Sporolithaceae, MA= Mastophoroideae, MET= Metagoniolithoideae, NE= Neogoniolithoideae.
Figure 5.
Diversity of rhodoliths reported for Panama (a) by order and (b) by sub-family. LI= Lithophylloideae, ME= Melobesioideae, CH= Chamberlainoideae, SPO= Sporolithaceae, MA= Mastophoroideae, MET= Metagoniolithoideae, NE= Neogoniolithoideae.
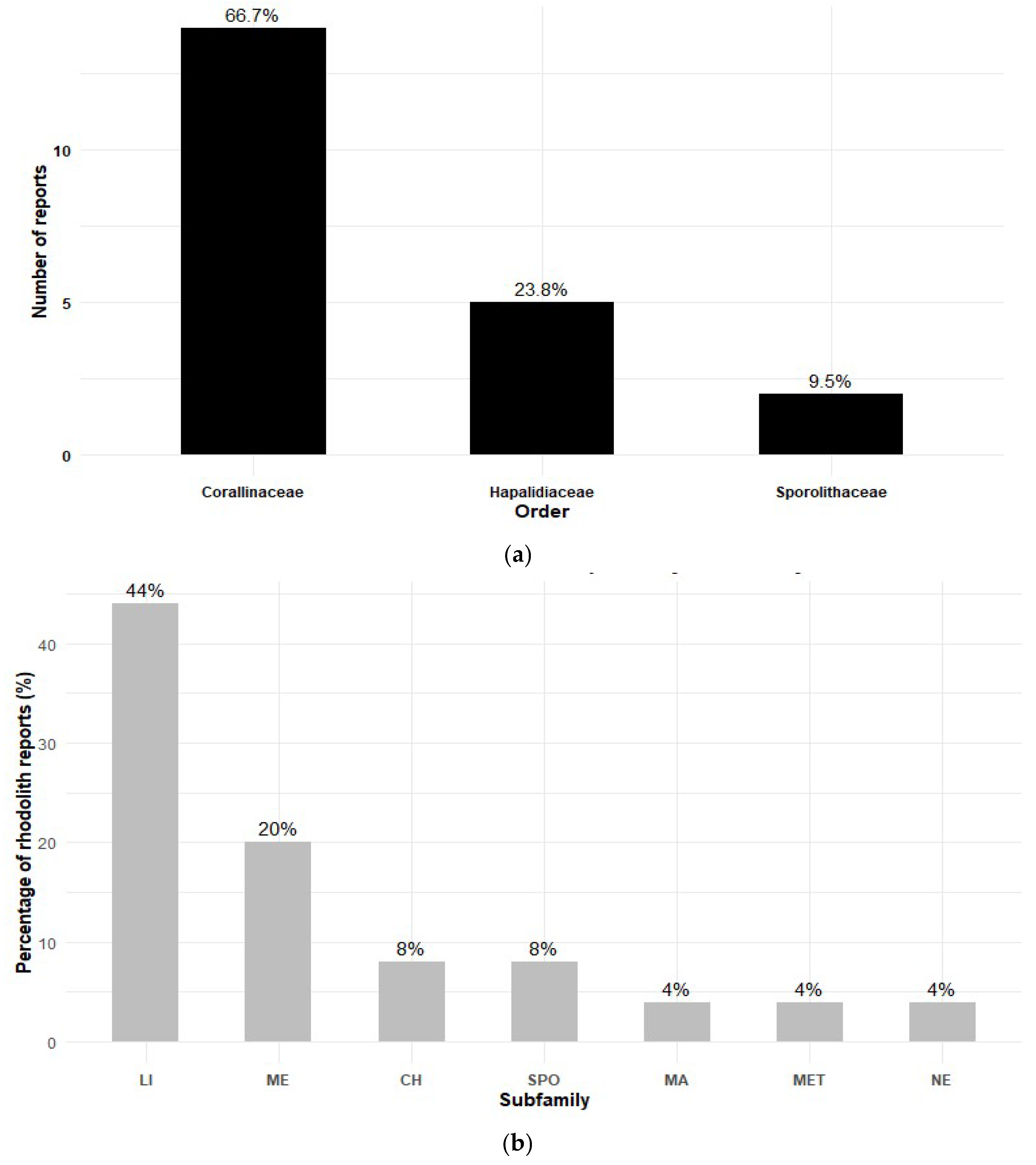
Figure 6.
Rhodolith bed at four sites of northeast of Coiba National Park.

Figure 7.
Conservation strategies for rhodoliths in Panama. Created with MindMeister web tool.
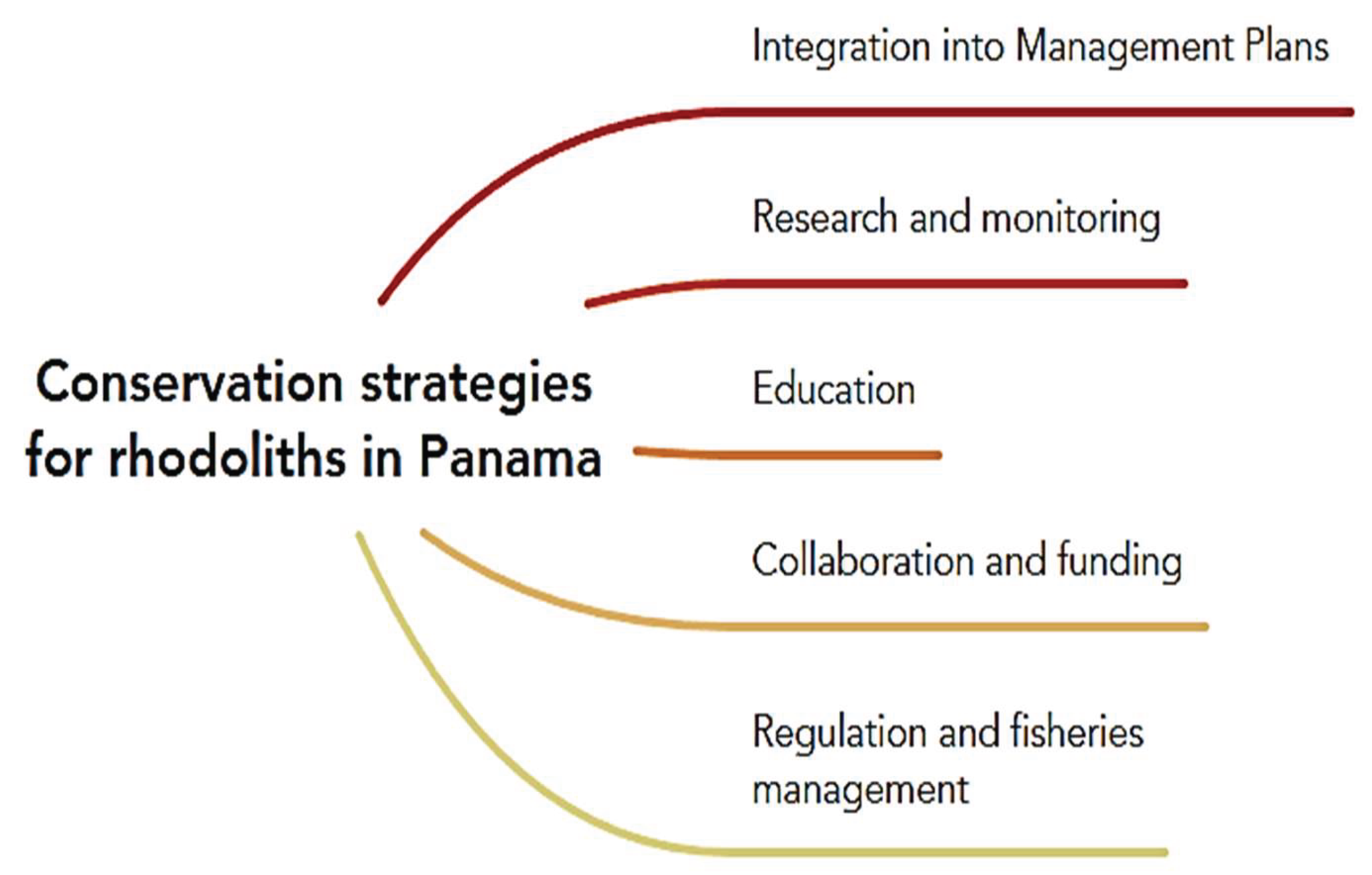
Table 1.
List of rhodolith-forming species reported for Panama.
| Family/Sub-family/Species | Locality |
|---|---|
|
Corallinaceae Sub-family Chamberlainoideae |
|
| Chamberlainium decipiens (Foslie) Caragnano, Foetisch, Maneveldt & Payri (as Spongites decipiens) [23,24] | Pacific |
| Pneophyllum confervicola (Kützing) YMChamberlain: (as Heteroderma minutulum) [23,24] | Pacific |
|
Sub-family Lithophylloideae |
|
| Lithophyllum coibense Me. Lemoine [12,24] | Pacific |
| Lithophyllum brachiatum (Heydrich) Me.Lemoine [12,24,25] | Pacific |
| Lithophyllum alternans Me.Lemoine [17,24] | Pacific |
| Lithophyllum okamurae Foslie [20] | Pacific |
| Lithophyllum prototypum (Foslie) Foslie (as Goniolithon tessellatum) [15,23,24] | Pacific |
| Lithophyllum pallescens (Foslie) Foslie [23,24] | Pacific |
| Lithophyllum divaricatum M. Lemoine [13,24] | Pacific |
| Lithophyllum neocongestum JJHernandez-Kantun, WHAdey & PWGabrielson [26] | Caribbean |
| Titanoderma pustulatum (JVLamouroux) Nägeli [15,27,28] | Caribbean |
| Sub-family Mastophoroideae | |
|
Goniolithon decutescens (Heydrich) Foslie ex M.Howe [20,25] |
Caribbean |
| Sub-family Metagoniolithoideae | |
|
Harveylithon munitum (Foslie & M.Howe) A.Rösler, Perfectti, V.Peña & JCBraga [21,29] |
Caribbean |
| Sub-family Neogoniolithoideae | |
|
Neogoniolithon trichotomum (Heydrich)Setchell et L.R. Mason [23,24] |
Pacific |
| Hapalidiaceae | |
|
Sub-family Melobesioideae |
|
| Lithothamnion australe Foslie [23,24] | Pacific |
| Lithothamnion australe f.americanum Foslie [13,24] | Pacific |
| Lithothamnion crispatum Hauck (as L. indicum) [12,17,24] | Pacific |
| Lithothamnion australe f. minutulum Foslie (as Mesophyllum australe var. minutula ) [12] | Pacific |
|
Mesophyllum australe var. tualense (Foslie) Mc. Lemoine [12,24] |
Pacific |
| Sporolithaceae | |
| Sporolithon episporum (M.Howe) EYDawson [12,15,17,27] | Caribbean |
|
Sporolithon howei (Lemoine) N.Y. Yamaguishi-Tomita ex M-J. Wynne: (asArchaeolithothamnion howei) [12,14,24] |
Pacific |
Table 2.
List of rhodolith-forming species reports from Panama to be confirmed.
| Locality/Species |
|---|
|
CARIBBEAN Ϯ Clathromorphum Foslie |
| Hydrolithon farinosum (J.V.Lamouroux) Penrose & Y.M.Chamberlain (as Fosliella farinosa) [15,25,27] |
| Lithophyllum corallinae (P.Crouan y H.Crouan) Heydrich [18] |
| Ϯ Lithophyllum kaiseri (Heydrich) Heydrich |
| Lithophyllum stictaeforme (Areschoug) Hauck [19] |
| Ϯ Mesophyllum mesomorphum (Foslie) WHAdey |
| Neogoniolithon spectabile (Foslie) Setchell & LRMason [30] |
| Ϯ Neogoniolithon strictum (Foslie) Setchell y LRMason |
|
Porolithon sp. Foslie [15] |
| PACIFIC |
| Fosliella fertilis (M. Lemoine) Segonzac [17,24] |
| Fosliella minuta W.R. Taylor [13,15,24] |
| Ϯ Hydrolithon boergesenii (Foslie) Foslie |
| Ϯ Hydrolithon breviclavium (Foslie) Foslie |
| ϮHydrolithon boergesenii (Foslie) Foslie |
| Ϯ Lithophyllum imitans Foslie |
| Ϯ Lithophyllum kotschyanum Foslie |
| Ϯ Phymatolithon lenormandii (Areschoug) WHAdey |
| Ϯ Mesophyllum engelhartii (Foslie) WHAdey |
| Phymatolithon masonianum Wilks & Woelkerling [31] |
| Ϯ Porolithon onkodes (Heydrich) Foslie [24,32] |
| Ϯ Porolithon sonorense EY Dawson |
| ϮSpongites fruticulosus Kützing [33] |
Ϯ Field observations or herbarium specimens reviewed in the Symbiota database, without related publications.
Table 3.
Sites performed at NE of Coiba National Park with the indication of the coordinates, coverage of rhodoliths and depth.
Table 3.
Sites performed at NE of Coiba National Park with the indication of the coordinates, coverage of rhodoliths and depth.
| SITE | Latitude °N | Longitude°W | Coverage (%) | Depth (ft) |
|---|---|---|---|---|
| Canales de Afuera | 7.68888 | -81.63419 | 46 | 45 |
| Buffet | 7.68537 | -81.61061 | 49 | 55 |
| Don Juan | 7.39809 | -81.63869 | 65 | 42 |
| Iglesias | 7.64542 | -81.69166 | 69 | 32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



