Submitted:
12 August 2025
Posted:
19 August 2025
You are already at the latest version
Abstract
Emerging evidence highlights the transformative potential of transcranial focused ultrasound in enhancing brain function and repair. Low-intensity focused ultrasound (LIFUS) enables precise, non-invasive modulation of neuronal activity, promoting cellular repair and brain plasticity. By targeting specific brain regions, LIFUS facilitates the release of neurotrophic factors, strengthens synaptic connectivity, and modulates molecular and cellular pathways essential for neural recovery. These mechanisms collectively support neuronal proliferation, differentiation, and functional integration, leading to cognitive improvements and neuroprotection without causing thermal damage. This review provides a comprehensive overview of LIFUS's effects on brain function, emphasizing its role in neuromodulation, cellular adaptation, and long-term viability for treating brain injuries and neurodegenerative diseases. The discussion covers optimized ultrasound parameters, efficacy in cellular and behavioral models, and its therapeutic potential for brain repair and functional recovery. By extending its scope beyond isolated mechanisms, LIFUS emerges as a versatile neuromodulation tool with significant clinical promise for enhancing brain resilience and cognitive outcomes.
Keywords:
ultrasound modulation
; neurogenesis
; cognitive repair
; ultrasound parameters
; neurotrophic factors
Introduction
Neurodegenerative diseases are a leading cause of disability and death worldwide, driven largely by population growth and aging [1]. Neurodegeneration is marked by the progressive loss of nerve cells from specific brain regions, through either chronic degeneration or acute injury [2]. Aging nervous systems are less capable of neuronal regeneration and axonal repair due to inhibitory microenvironments and various pathological mechanisms, contributing to the onset and progression of neurodegenerative diseases [2,3]. For instance, neuroinflammation and mitochondrial dysfunction disrupt the survival and proliferation of neural stem cells (NSCs), impeding neurogenesis [3,4,5,6]. In addition, oxidative damage from reactive oxygen species and inadequate antioxidant protection impairs the proliferation and differentiation of NSCs [7,8,9]. Neurotoxicity also prevents repair by causing harmful protein aggregates, including mutant huntingtin in Huntington's disease [10], alpha-synuclein in Parkinson's disease [11], and amyloid-beta (Aβ) in Alzheimer's disease [12], as well as other proteins in conditions like traumatic brain injury (TBI) [13]. These diseases and aging further contribute to reducing the phagocytic capacity of microglia, trigger astrogliosis, and alter the microvasculature of the dentate gyrus (DG). Consequently, neuronal plasticity is affected by neurodegeneration, disrupting hippocampal function while leading to psychiatric comorbidities and cognitive impairments [14]. These physiological abnormalities impede phenotypic changes, including altered eating and metabolism, autonomic nervous system dysfunction, sleep disturbances, cognition decline, and movement impairment, collectively leading to life-threatening complications [15].
Traditional therapy predominantly relies on pharmacological agents that lack long-term efficacy and safety, due to aging-related comorbidities, drug resistance, and tolerability [16,17]. Conversely, surgical interventions for neurodegenerative diseases pose significant risks, particularly in the elderly, due to their invasiveness [18,19]. While these traditional modalities may relieve symptoms or slow disease progression, they eventually become ineffective due to the continuous loss of neurons. Thus, the restoration of impaired repair or possible neurogenesis could be a promising therapeutic strategy for brain diseases characterized by neuronal cell loss [20]. Recent advancement in therapeutic modalities interfacing with the central and peripheral nervous systems aim to be less invasive with minimal adverse side effects. Ultrasound technology, in particular, has shown efficiency in creating precise lesions, eliminating protein aggregates, improving drug absorption, and controlling neuronal activity [21]. Ultrasound has been used in various medical conditions for the treatment of tremors, especially in the fields of neurology and psychiatry [22,23]. Current neuromodulation techniques often rely on electrical stimulation to treat chronic pain through implantable electrodes targeting deep brain structures, offering lasting effects despite their invasive nature [24,25]. Several alternative modalities address the constraints of electrical stimulation, such as transcranial direct current stimulation and repetitive transcranial magnetic stimulation; however, they are limited in terms of cortical surface area and focus [26]. Recently, a growing number of registered human trials have explored focused ultrasound for treating Alzheimer's and Parkinson's diseases demonstrating minimal side effects [27].
In particular, transcranial low-intensity focused ultrasound (LIFUS) has emerged as a promising treatment option, offering high spatial resolution while minimizing invasiveness and financial burdens on patients [26,28,29]. Notably, transcranial focused ultrasound (tFUS) can remotely stimulate intact brain circuits without requiring exogenous factors or surgery. By enhancing sharp-wave ripple oscillations, LIFUS facilitates endogenous brain plasticity and hippocampal-dependent memory consolidation [30]. Research has also explored combining focused ultrasound with intravenously injected contrast agents (ICAs) to deliver drugs to specific brain regions in humans and various animal species, thereby opening the blood-brain barrier (BBB) [31,32]. For instance, scanning ultrasound combined with microbubbles (MB) successfully opened the BBB and fully restored long-term potentiation (LTP) induction in the DG of the hippocampus while the same treatment without MB proved even more effective, not only restoring LTP but also ameliorating the spatial learning deficits of the aged mice [33]. In turn, LIFUS directly modulates neuronal activity by targeting brain regions at any depth without the need for pharmacotherapeutic drugs [34,35]. Importantly, LIFUS promotes neuronal repair and functional recovery by modulating the extracellular matrix, enhancing neuronal growth, and strengthening neuroprotection, ultimately leading to long-term cognitive improvements [31,36,37]. These attributes have highlighted LIFUS for its neuromodulatory effects, given its safety and potential benefits for learning, memory, and overall brain function [38,39]. While further research is needed to fully elucidate its underlying cellular mechanisms, studies suggest LIFUS may aid in treating psychiatric disorders and promoting neuronal recovery after stroke [40,41,42].
In this review, we explore the role of LIFUS beyond its neuromodulatory effects, specifically its link to promoting plasticity, neuronal repair, and functional recovery in the adult brain. We assess the evidence supporting this emerging approach across various settings, spanning both in-vitro and in-vivo applications. Our investigation identifies specific LIFUS parameters that modulate neural activity and contribute to brain plasticity, with a focus on the cellular mechanisms underlying structural and functional adaptations. Furthermore, we aim to assess the effectiveness of these state-of-the-art approaches by exploring LIFUS-induced changes in neural circuitry and cellular dynamics. We also discuss the challenges that impede their clinical translation for treating brain injuries and neurological disorders. The review is organized as follows: First, we describe adult neurogenesis, the process through which new neurons are generated and differentiated in the brain, as well as associated key brain regions and biomarkers. Second, we provide an in-depth overview of the physical properties of ultrasound technology and the molecular mechanisms involved in modulating neural function through ultrasound sonication. Following that, we highlight state-of-the-art applications of LIFUS, particularly its role in enhancing cognitive function and neural network remodeling in-vivo. Additionally, comprehensive research on the ultrasound parameters achieving significant results will be reported. Lastly, we address the current limitations of existing approaches and propose future research directions to advance therapeutic applications.
Adult Neurogenesis: Neurogenic Niches and Biomarkers
Many research studies have highlighted the brain's remarkable capacity for plasticity and self-repair following injury or disease [43]. These processes involve various mechanisms, including the integration of newly generated cells into existing neural circuits, which contribute to cognitive and affective functions [42,44]. Historically, Ramon y Cajal suggested that neuronal regeneration was restricted to embryonic development, fostering the belief that the adult brain lacked the ability to produce new neurons [45]. However, recent findings demonstrate that specialized regions of the brain, known as neurogenic niches, support cellular regeneration throughout life. These include the subgranular zone (SGZ) of the hippocampal DG and the subventricular zone (SVZ), which contribute to neural circuit remodeling and adaptive responses to environmental changes [46,47,48,49]. Figure 1A highlights these key regions in the adult rat brain, while Figure 2B outlines the progression of NSCs toward functional integration, along with the biomarkers and signaling pathways involved in this process.
Hippocampal and SVZ Brain Plasticity
A In the hippocampus, the SGZ of the DG exhibits a remarkable capacity for cellular plasticity and repair, as demonstrated in several rodent studies including our group [50,51,52,53,54]. This region is highly interconnected through rich synaptic networks and is influenced by neural circuit activity regulated by various neurotransmitters [55,56]. Cellular regeneration in this area supports the remodeling and adaptation of existing hippocampal networks, leading to changes in their functionality and contribution to cognitive processes [45]. Neural precursor cells in the SGZ progress through distinct stages of differentiation, expanding precursor populations, and undergoing maturation. Immature cells play roles in establishing functional connectivity within hippocampal circuits, with a subset achieving full integration into these networks [57]. These immature neurons play a role in neuronal maturation and functional connectivity within the hippocampus. Eventually, granule cells achieve complete integration into the hippocampal circuitry, contributing to overall brain function [58,59]. Similarly, the SVZ is surrounded by existing neurons and physically separated from the olfactory bulb, where newly generated neurons integrate into circuits responsible for processing olfactory information [55,56]. NSCs in the SVZ give rise to neurons through defined stages: beginning as type-B1 cells, resembling astrocytes, they differentiate into neurons via asymmetric division, then transition into type-A neuroblasts and migrate towards the olfactory bulb along the rostral migratory stream [60]. Once in the olfactory bulb, they integrate into the granule cell layer, contributing to plasticity and learning processes. It is notable that only a small portion of these neuroblasts survives to become mature granule cells, establishing connections crucial for brain function [61,62]. Recent discoveries, such as Alvarez-Buylla’s identification of a pathway within the infant human brain where immature cells migrate to the cerebral cortex and differentiate into functional neurons, further highlight the dynamic nature of cellular plasticity and regeneration in the brain [63]. While the hippocampal SGZ and SVZ share certain characteristics, such as their role in supporting brain adaptability, they also exhibit distinct mechanisms and functions. Both regions rely on diverse cellular and molecular processes to support brain function and plasticity [55,56,64].
Molecular Biomarkers
Currently, a diverse array of molecular biomarkers (Table 1), indicative of various stages in neurogenesis and neuronal repair, have been discovered and developed. Beginning with 5-bromo-2’-deoxyuridine (BrdU) and proliferating cell nuclear antigen (PCNA), which are essential in identifying actively dividing cells during DNA replication. Importantly, BrdU crosses the BBB and can be delivered by intracerebroventricular (i.c.v.), intraperitoneal (i.p.), or intravenous (i.v.) injection, or orally for studying adult neurogenesis [65]. These features render BrdU invaluable for detecting cell proliferation. However, research has also shown that BrdU can be integrated into DNA through various mechanisms, including cell differentiation [66,67,68]. To accurately characterize cell populations, researchers often combine BrdU labeling with immunohistochemistry or other molecular techniques, targeting specific markers for different cell types or developmental stages [65]. In addition, PCNA is expressed throughout all phases of the cell cycle, including repair processes [69]. Furthermore, doublecortin (DCX) is a protein crucial for microtubule polymerization, primarily expressed in migrating neuroblasts and immature neurons, making it an essential factor of neuronal development. Mutations in DCX can disrupt neuronal migration, contributing to various pathological conditions [70]. While DCX expression is associated with specific regions such as the hippocampus and olfactory system, it is notably absent in newly generated neurons in the neocortex, suggesting the involvement of alternative mechanisms in governing neuronal migration and maturation in this brain region [71]. On the other hand, neurons become more susceptible to degeneration and are less capable of repairing damage or forming new connections without adequate neurotrophic support [72,73,74]. For instance, BDNF or brain-derived neurotrophic factor, along with its high-affinity receptor TrkB, regulates various stages of neurogenesis from the proliferation of neural progenitor cells to their differentiation into mature neurons, indicating the importance of BDNF-TrkB signaling in maintaining neuronal populations [67]. Additionally, BDNF maintains an enriched environment conducive to neurogenesis, particularly in the granule layer of the DG and other hippocampal regions [75]. In turn, glial cell line-derived neurotrophic factor (GDNF) indirectly facilitates proliferation and differentiation of certain types of neurons, particularly dopamine-producing neurons in the brain [76,77]. GDNF has garnered significant attention for its potential therapeutic applications, particularly in neurodegenerative diseases like Parkinson's disease [78]. It is primarily produced by glial cells, and functions by binding to specific receptors on target neurons. This activates signaling pathways that promote their survival during the critical period following their birth [78,79]. On the other hand, vascular endothelial growth factor (VEGF) is a signaling protein, also known as a cytokine, playing a crucial role in angiogenesis, the formation of new blood vessels from existing ones [80]. Studies have suggested that VEGF can promote the proliferation and differentiation of NSCs in the hippocampus, contributing to the cognitive function of the brain [81]. Likewise, nerve growth factor (NGF) activates signaling pathways necessary for cell proliferation, differentiation, and survival by phosphorylating extracellular signal-regulated kinases 1 and 2 (ERK1/2) [82,83]. Additionally, NGF orchestrates the differentiation of NSCs into mature neural phenotypes, specifically inducing the development of neurons and astrocytes, while not promoting the differentiation of NSCs into oligodendrocytes [84,85,86].
Recent findings have opened new avenues, highlighting the potential of focused ultrasound to influence both neuronal activity and neural repair in the adult brain [87]. Ultrasound activates IL-10/IL-10R signaling pathways, contributing to neural repair and improved neurobehavioral outcomes after ischemic stroke [88]. Consequently, LIFUS has been shown to enhance spatial memory functions in animal models compared to the non-sonicated animals [33]. Moreover, LIFUS has been found to stimulate astrocytes and upregulate their production of BDNF [40]. Both neural stimulation and increased levels of BNDF are recognized key contributors to neuronal growth, synaptic plasticity, and functional recovery [89,90]. Thus, the process of adult neurogenesis can be significantly enhanced through LIFUS neuromodulation via influencing cell behaviors such as proliferation, differentiation, and modulation of synapse functionality and neuronal networks [91].
Ultrasound Effect on Neurons: Then and Now
The initial reports of the bioelectric impact of ultrasound on neural tissue traces back to 1929, when Harvey recorded muscular twitching in a frog's gastrocnemius muscle following ultrasound sonication of the sciatic nerve [64,92,93]. Subsequently, during the 1950s, the Fry brothers conducted ultrasound sonication on brain areas in cats and observed reversible modulation of visually evoked potentials [94]. Ever since, studies have investigated the impact of focused ultrasound on neural excitability, using low-intensity pulses targeted at different areas of the brain, specifically cortical and thalamic regions, in rats, rabbits, and cats [95,96]. Indeed, studies showed a change in the electrical activity of the targeted brain regions with a consistent movement of electrical charges towards a more negative state, enhancing a widespread suppression of neuronal activity and conductivity. In the early 2000s, a series of pioneering studies revealed that low-intensity ultrasound could directly trigger potential action firing in cultured hippocampal neurons and isolated mouse brains [97,98]. Recent studies have confirmed ultrasound's capacity to stimulate brain circuits, notably those associated with movement. This was demonstrated by eliciting motor responses in rodents and rabbits following ultrasound application to somatomotor cortical regions. These investigations extensively explored various stimulation parameters, aiming to enhance our understanding of ultrasound's interaction with brain tissue [30,99,100,101,102]. More recently, applying low-intensity high-frequency ultrasound, to the developing pyramidal neurons of mouse fetuses significantly increased dendritic branching, suggesting ultrasound as a promising modality in regenerative medicine for individuals with neurodevelopmental disorders characterized by a reduction in the dendrite count [103]. Translationally, larger models including non-human primates (NHPs) and humans were considered for investigating the neuromodulatory effects of low-intensity ultrasound in the brain, with precise targeting facilitated by multiple imaging technologies such as computed tomography (CT) and magnetic resonance imaging (MRI). The firing rate of individual neurons significantly changed when applying LIFUS to the frontal eye field in NHPs, consequently influencing visual processing and decision-making behavior [104,105]. When targeted at the somatosensory cortex, LIFUS could modulate sensory networks, as detected by functional MRI [106]. Moreover, in human studies, LIFUS directed at the somatosensory cortex could modify electroencephalogram (EEG) dynamics [107,108], as well as induce tactile sensations in the limbs [109]. Additionally, directing LIFUS towards the frontal-temporal cortex, with a high dependence on the precise targeting of the ultrasound waves, could produce non-specific mood effects, underscoring a localized impact on neural structures [110]. Concurrently, longer sonication protocols were investigated to study the long-lasting effects of ultrasound on the brain. Prolonged application of LIFUS to the pre-frontal cortex in humans has been associated with changes in functional connectivity [111]. In NHPs, LIFUS targeted at the amygdala and supplementary motor area resulted in significant alterations in the connectivity patterns of the sonicated regions [112,113].
Nowadays, clinical neuromodulation techniques primarily rely on electrical stimulation. While these approaches have advanced significantly, they often involve surgical procedures and the implantation of specialized devices. In contrast, delivering ultrasound pulses to targeted brain regions presents a promising, less invasive alternative with potential benefits for cognitive function and brain repair after injury or disease [21]. Ultrasound sonication through the skull, especially at low-intensity, can impact biological processes in living tissue through thermal and mechanical mechanisms [91]. Subsequently, LIFUS stimulation-based approaches have been shown to induce adult neurogenesis and cognition by modulating various molecular substrates [114]. Published work indicates that LIFUS induces proliferation and secretion of nutritional factors in NSCs without triggering apoptosis [115]. LIFUS has been found to effectively enhance the production of neurotrophic factors [38], namely BDNF [30,38,39,41,90,116,117,118,119,120,121], GDNF [38,116,122], and VEGF [38,40,118,120,123], all of which contribute to neuronal survival and functional recovery. LIFUS activates integrin receptor signaling pathways that regulate the production of neurotrophic factors, supporting key stages of neuronal development, including growth, differentiation, and migration [40,116]. Research has reported that LIFUS activates the ERK-CREB pathway, leading to increased CREB phosphorylation and subsequent neuronal adaptation [87,124]. Additionally, transcranial LIFUS enhances the activation of the TrkB/Akt-CREB pathway, which is essential for neuroprotection and synaptic plasticity [118].
Ongoing research aims to elucidate how LIFUS enhances neuronal electrical activity through the modulation of sodium (Na+) and calcium (Ca2+) channel gates, thereby indirectly supporting neurogenesis [38,39]. Moreover, LIFUS treatment has demonstrated neuroprotective potential and may be effective in preventing recurrent strokes [121]. The modulation of the TLR4/NF-κB signaling pathway further suggests the role of LIFUS in reducing inflammation and promoting neural repair [90]. Thus, LIFUS has the potential to drive structural and functional recovery in brain injury models. As a result, it is considered an optimal neuromodulation technology capable of achieving highly adequate spatial and temporal neuromodulation while minimizing invasiveness and circumventing the need for any genetic manipulation [30].
Physical Properties
Ultrasound comprises acoustic waves oscillating at frequencies higher than the human hearing range (20 Hz – 20 kHz). These sound waves cause mechanical disruption to the media particles, which deviate from their equilibrium position due to the pressure field created. Ultrasonic waves propagate through a medium while generating sequential compressions and rarefactions, which dynamically alter the local pressure at a specific frequency denoted as the carrier frequency (fc). The lower the acoustic frequency, the broader the focal region of the acoustic waves at the targeted area, registering deeper penetration as the waves experience less scattering and absorption along their propagation. In addition, the kinetic energy carried out by a propagating acoustic wave is typically quantified through intensity I (in W/m2). It represents the energy transmitted across a unit area per unit time, taking into consideration the peak pressure P amplitude (in Pa) and the impedance of the medium characterized by its density ρ (in Kg/m3) and speed of sound c (in m/s). For a propagating sinusoidal acoustic wave, the intensity is described by equation (1) [125,126,127].
Generally, soft tissues closely resemble water in terms of density and compressibility, leading to comparable propagation speeds of c = 1500 m/s. This indicates that the velocity of acoustic waves decreases as materials become denser and more compressible. The ultrasonic intensity is calculated as the spatial-peak pulse-average intensity (ISPPA) computed at the spatial peak across the tone burst duration (TBD) or as the spatial-peak temporal-average intensity (ISPTA) computed at the spatial peak across the pulse repetition period (PRP) when ultrasound is applied in the pulsed mode; LIFUS operates at low ISPPA in the 1 – 102 W/cm2 range. Moreover, ultrasound waves (Figure 2A) are applied for a sonication duration (SD) ranging from a few milliseconds (ms) to several minutes, with an interstimulus interval (ISI) between each SD. Ultrasonic pulses are delivered for a TBD, repeated at a pulse repetition frequency (PRF), stated in equation (2), for a given duty cycle (DC). The DC defines the percentage of SD during which ultrasound pulses are being delivered. Consequently, the efficacy and neuromodulatory effects of LIFUS on biological tissues are highly dependent on the acoustic parameters shown in Figure 2A.
During tFUS, the ultrasonic waveform gets attenuated as it passes through the skull, with the degree of attenuation depending on the bone density [125,128]. For safe human tFUS, the fc generally used lies in the range of 200 – 650 kHz. Most ultrasound stimulations are conducted at acoustic intensities and pressures well below the FDA-approved acoustic outputs typically employed for transcranial Doppler imaging. Mainly, ISPTA ≤ 720 mW/cm2 and either a mechanical index (MI) ≤ 1.9 or ISPPA ≤ 190 W/cm [129]. The latest guidelines for low-intensity tFUS advise using short acoustic pulses in the 0.2 – 100 ms range at a moderate to high PRF ranging from 10 Hz to 2 kHz, with a low DC maintained at less than 50%, along with an ISPPA below 30 mW/cm2 [125].
Mechanism of LIFUS Effect on Neurons
Despite an increasing body of empirical studies, questions persist regarding the specific mechanisms through which ultrasound influences neural structures including effects on excitability. This lack of precise understanding stems from the multitude of physical effects ultrasound has on biological tissue, coupled with sub-cellular structures potentially involved in converting pressure waves into electrical responses. Upon sonication, an acoustic wave interacts with the neural membrane through various mechanisms, altering the state of the membrane. Ultrasound introduces energy to the lipid membrane and initiates a localized disturbance that can transform into temporary modified states, subsequently propagating along the membrane. As structures revert to their lowest energy state, the membrane adjusts its mechanical attributes in reaction to an ultrasonic wave. Here, we propose three commonly stated mechanisms for the mechano-electrical coupling of ultrasound with the neural membrane, altering its excitability. These mechanisms, illustrated in Figure 2B, C, and D, can operate simultaneously or interchangeably with different sensitivities depending on the chosen ultrasound parameters and target region.
Intramembrane Cavitation
Upon sonication, low-intensity ultrasound could form microscopic gaseous bubbles within the intramembrane space of the lipid bilayer leaflets due to their continuous inflection and deflection (Figure 2B). Once these cavities form, they experience periodic oscillations at the frequency of the ultrasound waves [130], a process known as intramembrane cavitation. This cavitation could then lead to changes to the membrane capacitance and membrane potential by enhancing the lipid bilayer permeability due to the creation of membrane-bound physical pores that facilitate ion transport. Consequently, cavitation leads to depolarization of the membrane, thus inducing neural firing [131,132]. During the compression phase, the intramembrane bubble shrinks and compresses its gaseous core when subjected to positive acoustic pressure, while during the rarefaction phase, the low pressure facilitates the expansion of the bubble as its gaseous core decompresses. This continuous cycle, known as stable cavitation, boosts the absorption of acoustic energy locally while creating microstreaming in the surrounding fluid, leading to intense shear stresses that may harm cells severely. As a result, viscous stresses are generated in the surrounding medium, making stable cavitation particularly effective at lower ultrasonic frequencies. When exposed to higher pressure amplitudes, the generated microbubbles expand more during the rarefaction phase. However, their ability to compress is naturally limited due to their gaseous core. Accordingly, the response of the microbubble to sonication becomes highly nonlinear, causing it to expand further with each rarefaction cycle. This continual expansion eventually leads to the collapse of the bubble itself, a phenomenon known as inertial cavitation. This type of cavitation can create significant increases in acoustic pressure and temperatures locally while generating shock waves. These effects can have profound implications for the surrounding tissue.
Mechanosensitive Ion Channels
Ultrasound sonication exerts pressure on surrounding tissues and cell membranes, potentially leading to the opening of ion channels that are sensitive to mechanical changes, thus depolarizing the membrane [133] (Figure 2C). Numerous in-vitro studies have reported this effect in mechanosensitive channels at various degrees [134,135], including the two-pore-domain potassium channel family such as TREK-1 and TRAAK channels [136], as well as in Na+ and Ca2+ voltage-gated ion channels involved in generating action potentials [98,137]. Transient receptor potential (TRP) channels are activated via ultrasound-induced membrane stretching, ultimately changing the mechanism of ion transport [138]. For instance, Yoo et al. demonstrated that LIFUS targeted at cultured mouse primary cortical neurons activated specific mechanosensitive channels, leading to a Ca2+ influx, which in turn activated calcium-dependent sodium channels, triggering a train of action potentials [139]. However, the variable expression of such ion channels across neural structures [140] poses challenges in formulating a comprehensive theory explaining their interaction with ultrasound upon activation.
Membrane Conformational Changes
Ultrasound sonication causes localized changes to the mechanical properties of the neuronal plasma membrane by modifying its geometry, especially membrane thickness [141], curvature [142], and overall membrane electrical state [143] (Figure 2D). Ultrasound mechanical energy exerted on the neural membrane takes the form of an acoustic radiation force (ARF), which results from the transfer of momentum from the ultrasound wave field to the surrounding medium. ARF induces a steady localized pressure throughout the SD of ultrasound. This sustained pressure can lead to the stretching of a cell membrane to a degree that affects the ion channel reconfiguration, thus influencing the membrane's fluidity and permeability [144]. Such force is enhanced at higher acoustic frequencies and can cause irreversible damage due to the generated acoustic streaming. Another form of the electro-mechanical coupling of ultrasound with the neural membrane is direct flexoelectricity [145]. Similar to the phenomenon of piezoelectricity observed in solid crystals, the concept of mechano-electric flexoelectricity in liquid crystals refers to the generation of an electrical current resulting from strain-induced oscillations in the membrane curvature as the ultrasound waves cause a variation in the membrane’s area [146].
Moving forward, LIFUS is regarded as non-thermogenic and non-destructive, whereas its mechanical strains may result in positive biochemical events on the cellular level [91,147]. While several possible explanations exist, the mechanisms of how these events occur remain unclear. LIFUS may excite or inhibit neuronal activity by acting on mechanically sensitive ion channels, which induces neuronal morphologic changes by activating stretch-activated ion channels and Ca2+ messenger ions [88,147]. More specifically, it may regulate ion channels and change the intracellular Ca2+ level. This results in a variety of cell behavioral responses, including neuron proliferation and differentiation [91]. Such exposure consequently increases intracellular Ca2+ concentration, leading to the expression of genes that are important to dendritic development, neuronal survival, and synaptic plasticity [124]. These mechanisms also facilitate molecule passage across membranes, increase enzyme activity, and modify cell metabolism, thereby promoting cell membrane fluidity and selectivity [148]. Collectively, LIFUS-induced changes in ion channel activity and membrane properties may enhance neuronal plasticity and foster conditions conducive to neurogenesis.
Ultrasound Induced-Neurogenesis
As previously reported, the use of ultrasound at low intensity elicits diverse responses in neurons, astroglia, and the extracellular matrix, creating an environment conducive to cell proliferation and neuronal plasticity by modulating various molecular substrates [31,114]. GDNF and VEGF, along with other biomarkers, contribute to brain plasticity by supporting the growth and development of neural cells and blood vessels in the brain [118,120]. Notably, LIFUS enhances the release and synthesis of neurotrophic factors by astrocytes which play crucial roles in neuroprotection, neural regeneration, and long-term cognitive improvements [31]. These effects are initiated by targeted acoustic waves that trigger intracellular signaling events through the activation of mechanosensitive receptors, membrane cavitation and other changes. While some studies focus specifically on the effects of ultrasound on neurogenesis, others examine its broader impact on brain function and pathology, investigating factors involved in generating new neurons and their integration within the existing network. This section will discuss the responsiveness of various cell types to LIFUS and examine how different parameters of ultrasound stimulation react with targeted regions of animal brains, affecting both neuronal activity and behavior.
In-Vitro Applications
Recent findings highlight the benefits of using ultrasound stimulation in modulating cultured human NSCs (Table 2). One major application is stem cell transplantation, which holds a promising future for individuals suffering from central nervous system injuries, neurodegenerative diseases, or mental illnesses [114]. Various cell types, including NSCs, mesenchymal stem cells (MSCs), Schwann cells, and others, have contributed to advancements in stem cell transplantation [149]. While focusing on promoting proliferation and differentiation, researchers have investigated the therapeutic effects of ultrasound stimulation on NSCs in-vitro at different frequencies while maintaining low- intensities. For instance, Lv et al. employed 1 MHz low-intensity pulsed ultrasound (LIPUS) with an intensity of 500 mW/cm², revealing enhanced cell viability, proliferation, and neural differentiation, along with up-regulated gene and protein expressions of NF-M, Tuj1, S100b, and GFAP in induced pluripotent stem cells-derived neural crest stem cells (iPSCs– NCSCs) [150]. In another study, Lee et al. investigated the impact of low-intensity ultrasound exposure on neural stem/progenitor cell (NSPC) cultures by utilizing a central frequency of 1.8 MHz and intensities up to 500 mW/cm2. They demonstrated that ultrasound stimulation maintains NSPC viability and promotes their attachment, differentiation, and neurite outgrowth [91]. Additionally, dual-frequency ultrasound stimulation with frequencies at 560 kHz and 1138 kHz was found to promote NSPC differentiation more effectively than single-frequency stimulation at 1138 kHz alone, suggesting its ability to enhance Ca2+ influx and stimulate cell membrane mechanochannels [151]. On the other hand, Wu et al. explored 1 MHz LIPUS with a comparably lower intensity of 69.3 mW/cm2, inducing NSCs proliferation while increasing the expression of nutritional factors NGF, NT-3, and BDNF and modulating the Notch signaling pathway [115]. However, Schuster et al. suggested that applying continuous ultrasound ranging from 510 kHz – 3 W/cm2 to 4.36 MHz – 25 W/cm2, caused in-vitro NSCs expansion without directly affecting further differentiation into neurons or glial cells [152].
In a related context, Xie et al. investigated the proliferation of human bone marrow mesenchymal stem cells (hBM-MSC) using LIPUS with an intensity of 50 or 60 mW/cm2 along a frequency of 1.5 MHz, showing a significant effect on cell proliferation[153]. They noted that LIPUS sonication for 5 minutes enhanced hBM-MSC proliferation through the activation of the PI3K/AKt signaling pathway and increased expression of cyclin D1. However, they noticed that prolonged exposure may inhibit cell growth [154]. In another study, cell proliferation was assessed, revealing that 0.25 MHz LIPUS of 30 mW/cm2 intensity significantly promoted human amnion-derived mesenchymal stem cell (hAD-MSC) proliferation through activation of the ERK1/2 and PI3K-Akt signaling pathways, indicating their potential to differentiate into multiple types of cells [155]. Contrarily, Ning et al. investigated the application of LIPUS to enhance rat bone marrow mesenchymal stem cell (BMSC). Notably, LIPUS upregulated BDNF and NGF, increased BMSC viability, and inhibited gliosis differentiation without promoting neuronal differentiation. The optimal cell proliferation rate was achieved under 1 MHz LIPUS stimulation at an intensity of 50 mW/cm2 [149].
Lastly, low-intensity ultrasound has been used to promote the proliferation and differentiation of Schwann cells, a persistent source of neurotrophic factors, obtained from rats under varying conditions. Tsuang et al. highlighted that subjecting Schwann cells to 1 MHz ultrasound treatment at an intensity of 0.3 mW/cm2 resulted in cell viability and proliferation, particularly in serum-deprived culture [148]. A subsequent study explored LIPUS treatment at an intensity of 27.37 mW/cm2, indicating increased Schwann cell viability and proliferation, accompanied by upregulation of NGF, BDNF, GDNF, and Cyclin D1, suggesting potential for promoting neuronal repair after injury [156]. Another in-vitro study by Guo et al. revealed that 1.5 MHz LIPUS stimulation of intensity 15 mW/cm2 increased the viability of neuron-like SH-SY5Y cells and primary glial cells when delivered daily for 5 minutes over the scope of 3 days [41]. A very recent study utilized the human SH-SY5Y neuroblastoma cell line as an NSCs model to be treated with single or triple 20 minutes 1.5 MHz LIPUS exposures with intensity of 60 mW/cm2. The results show that LIPUS exposure(s) induced cell proliferation, as evidenced by an increase in the numbers of NSCs. Additionally, the study indicated a regulatory role for ERK signaling in LIPUS-induced NSC proliferation [157]. In a different context, Zhao group combined LIPUS and NGF at a concentration of 50 ng/mL on neurite outgrowth in a clonal cell line derived from rat pheochromocytoma (PC12) to investigate their effects on the employed cells. They administered LIPUS at a frequency of 1 MHz, with low intensities of 50 mW/cm2, administered every other day for 50 minutes alongside NGF treatment. The outcomes revealed that the combination of LIPUS and NGF significantly promoted neurite outgrowth, exhibiting a comparable effect to NGF treatment alone at a higher concentration (100 ng/mL) [147].
In summary, low-intensity ultrasound leads to enhanced cell viability, proliferation, and differentiation of NSCs and various other cell types in-vitro. Nevertheless, triple 1 MHz LIPUS stimulation on astrocyte function in the rat brain, totaling 15 minutes of stimulation, led to increased levels of essential neurotrophic factors such as BDNF, GDNF, VEGF, and GLUT1 in rat brain astrocytes, highlighting its therapeutic potential in supporting brain plasticity and cellular repair [116]. Similarly, rat cerebral cortex astrocytes following 1 MHz LIPUS stimulation revealed a significant increase in cell growth within 24 hours compared to control cells, indicating that LIPUS can enhance BDNF expression [158]. Moving forward, several of these investigations have been complemented by in-vivo experiments aimed at directing NSC migration to damaged areas of the brain, thereby promoting neural repair and regeneration. Specifically, Ling et al.'s study on the transplantation of ultrasound-stimulated BMSCs in rats with spinal cord injuries (SCI) highlighted improved functional recovery [149].
In-Vivo Applications
New approaches (summarized in Table 3) emerged when employing in-vivo ultrasound stimulation to study its potential applications, particularly in the enhancement of different injured animal models. This section highlights the efficacy of LIFUS in the absence of ICAs, such as MB or drugs. Blackmore et al. demonstrated that 1 MHz scanning ultrasound without MBs exhibited a significant increase in DCX levels in the DG compared to all other treatment groups while reducing perineuronal nets (PNN) and improving cognitive function in physiologically aged mice [33]. Tufail et al. investigated the impact of transcranial LIPUS with a frequency of 0.35 MHz and an applied intensity of 36.20 mW/cm2, targeting and stimulating unilateral hippocampi in healthy mice. Their findings suggested that LIPUS modulated neuronal activity in the intact mouse hippocampus, causing elevated BDNF protein expression levels and fostering endogenous brain plasticity [30]. Similarly, 1 MHz focused ultrasound with a low intensity of 5 mW/cm2 could activate neurons and promote adult neurogenesis in the DG of healthy mouse brains. This treatment led to doubled DCX levels compared to non-treated controls, highlighting its potential to enhance remodeling, repair and neuronal activity [87].
Numerous studies have demonstrated the effect of ultrasound stimulation on neural differentiation, particularly within the frequency range of 1 – 1.8 MHz and a power density of 100 – 500 mW/cm2 [124]. Researchers have employed animal models with induced injuries to investigate the therapeutic capabilities of ultrasound stimulation. Lin et al. explored the effect of LIPUS stimulation on an Alzheimer’s disease rat model. Applying 1 MHz LIPUS with an intensity of 528 mW/cm2 revealed an increase in BDNF, GDNF, and VEGF in the hippocampus and further improved cognitive dysfunction [38]. Similarly, Chen et al. demonstrated that LIPUS treatment caused increases in Aβ, amyloid precursor protein (APP), and CREB/BDNF expression, thereby attenuating memory impairment and amyloidogenesis in a mouse model simulating Alzheimer's disease [90]. By employing the same parameters for ultrasound stimulation (1 MHz, 528 mW/cm2), Sung et al. demonstrated that LIPUS stimulation on the right striatum in a rat model of Parkinson's disease induced the number of TH-positive neurons, GDNF, and BDNF expression in the rat’s SNpc [122]. Yang et al. explored the effects of LIPUS on bilateral rat hemispheres with multiple sclerosis (MS). Their application significantly enhanced protein levels of BDNF, GDNF, VEGF, and GLUT1 in rat brain astrocytes [116]. In addition, Huang et al. demonstrated that LIPUS treatment upregulated BDNF expression and improved learning and memory abilities in rats with vascular dementia (VD) [117]. Su et al. investigated the effect of LIPUS on TBI in mice, enhancing BDNF and VEGF expression while inhibiting apoptosis [118]. Moreover, Chen et al. and Wu et al. investigated, respectively, the effects of LIPUS treatment against cerebral ischemia/reperfusion injury of middle cerebral artery occlusion (MCAO). Notably, Chen et al. found that LIPUS significantly reduced neuronal cell apoptosis, downregulated apoptosis-related signaling molecules, and upregulated BDNF expression [120]. Correspondingly, Wu et al. reported neurological score and motor activity improvement, attenuation of apoptotic neuronal cells, and upregulation of Bax/Bcl-2 protein BDNF expression ratios in the brain of recurrent stroke mice [159]. In a very recent study investigating the impact of tFUS on cognitive function in Parkinson's disease, researchers assessed memory ability, revealing a significant improvement after 4 days of stimulation. The main parameters of the output ultrasound were 1 MHz and maximum sound pressure of 0.51 MPa [160].
Moving forward, many studies shifted to employing higher frequencies up to 3.3 MHz, while others employed even lower frequencies, down to 0.04 MHz [124]. As such, Guo et al. applied 1.5 MHz LIPUS to cuprizone-treated mice and investigated its effects on their whole brain. They found that LIPUS increased levels of DCX in the cortex and BDNF in the hippocampus, indicating enhanced neurogenesis [41]. Further, Mooney et al. employed 1.68 MHz tFUS at different pressure amplitudes to study the mechanisms of neurogenesis by stimulating neurons. Their method potentially increased the density of BDNF-positive puncta in the hippocampus [89]. Furthermore, Eguchi et al. explored the therapeutic effects of whole brain 1.875 MHz LIPUS therapy in both dementia and Alzheimer's disease models. Improved cognitive function and cerebral blood flow, enhanced endothelial cells, and expressions of endothelial nitric oxide synthase (eNOS) and neurotrophins were reported [123]. Lastly, Tranmontin et al. evaluated 3.3 MHz LIPUS treatment in a rat model of Alzheimer's disease, marking the highest frequency used on neural regeneration in literature. Their results showed that LIPUS improved recognition memory and increased NGF and BDNF levels in the hippocampus and cortex [39].
On the other hand, Wang et al. employed 0.8 MHz LIPUS with an average pulse intensity of 3.0 W/cm2, which promoted hippocampal neurogenesis and enhanced neural plasticity in the CA1 subregion of the hippocampus, restoring neuronal function and cognitive capabilities in TBI mice [161]. In addition, Liu et al. applied 0.5 MHz ultrasound stimulation at 30 minutes after distal MCAO in rat models, indicating strong positive expression of BDNF in cells of the LIPUS group [119]. Wang et al. explored the effect of 0.5 MHz focused ultrasound on a similar mouse model, indicating improved neurobehavioral tests, increased M2 microglia activation, and upregulated IL-10 signaling [162]. One year later, Ichuji et al. revealed that LIPUS therapy improved neurological functions and increased DCX neurons in the ischemic brain region of the same mouse model. Interestingly, they examined the influence of eNOS ablation in the MCAO model using eNOS-deficient mice to demonstrate that LIPUS therapy had no effect on the number of DCX neuroblasts nor on the expression levels of VEGF, CXCR4, and ERK [40], contradicting previous studies. Similarly, Cho et al. demonstrated that ultrasound stimulation at a very low frequency (0.5 MHz) and low intensity (50 mW/cm2) induced neural differentiation in hBM-MSCs without causing cell death or neurogenesis in a mouse stroke model [124].
In addition to varying frequencies and average intensities, researchers investigated different sonication treatments spanning from a few hours [119] to days [30,40,87,118,120,123], weeks [33,89,90,121,128], and months [39,41,116,117,122,123,124,155]. Conversely, many studies employ behavioral studies within a multi-treatment paradigm spanning several days to weeks before and after ultrasound treatment to evaluate animal cognition. These tests included the Morris water maze to assess spatial [38,90], the accelerated APA paradigm [33], the CPZ model [41], the Y maze [123,161], the wire-hanging task [30], and the novel object recognition test [123]. Additionally, motor functions were assessed using the rotarod, corner test [30,40,162], neurological severity scores [161,162], and elevated body swing test [120,121,162]. Consequently, none of these studies reported a decrease in behavior or cognition but they found improved cognitive functioning [117].

Table 2.
In-vitro applications.
| Study | Cell Culture | F MHz |
PRF Hz |
DC % |
TBD ms |
Ispta mW/cm2 |
Isppa W/cm2 |
SD min |
Outcome |
|---|---|---|---|---|---|---|---|---|---|
| [148] | Schwann cells | 1 | 20 | - | - | 0.3 | 3 | ↑IL-1, ↓IL-6, ↓TNF-α | |
| [150] | iPSCs–NCSCs | 1 | 100 | 20 | - | 500 | - | 10 | ↑NF-M, ↑S100β, ↑GFAP |
| [91] | NSPC cultures | 1.8 | 20 | - | - | 100 to 500 | - | 5 | ↑NGF, ↑MAP-2, ↑GFAP, ↑NO |
| [116] | Rat brain astrocyte cells | 1 | 10 | 50 | 50 | 528 | - | 15 | ↑BDNF, ↑GDNF, ↑VEGF ↑GLUT1 |
| [124] | hBM-MSC | 0.04 | - | - | - | 50 | - | 60 | ↑MAP2, ↑ND1, ↑NF-L,↑Tau, ↑DCX ↑NESTIN, ↑p-CREB, ↑p-ERK |
| [147] | PC12 | 1 | 100 | 20 | - | 50 | - | 10 | ↑p-ERK1/2 ↑p-Akt ↑p-CREB ↑Trx-1 |
| [155] | hAD-MSC | 0.25 | 1 | 20 | 0.2 | 30 | - | 30 | ↑ERK1/2, ↑Akt, ↑Cyclin D1, ↑Cyclin E1, ↑Cyclin A2, and ↑Cyclin B1 |
| [158] | Rat astrocyte cell line | 1 | 10 | 50 | 50 | 110 | - | 15 | ↑BDNF, ↑NF-κB, ↑ TrkB/PI3K/Akt, ↑Ca/CaMK |
| [152] | NSCs | 4.36 | - | - | - | 496 | - | 10 | no differentiation |
| [156] | Schwann cells | 1 | - | - | - | 27.37 | - | 10 | ↑FGF, ↑NGF, ↑BDNF, ↑GDNF, ↑Cyclin D1 |
| [149] | Rat BMSC | 1 | - | - | - | 50 | - | 3 | ↑BDNF, ↑NGF |
| [163] | Human umbilical vein endothelial cells | 1.5 | 1000 | - | 0.2 | 30 +- 5.0 | - | 20 | ↑YAP/TAZ |
| [154] | hBM-MSC | 1.5 | 1000 | 20 | - | 50 or 60 | - | 5 | ↑PI3K/AKt ↑Cyclin D1 |
| [115] | NSCs | 1 | - | - | - | 69.3 | - | 5 | ↑NeuN, ↓GFAP, ↑NGF, ↑NT-3, ↑BDNF |
| [41] | SH-SY5Y and primary glial cells | 1.5 | 1000 | 20 | - | 15 | - | 5 | ↑BDNF, ↑DCX |
| [157] | human SH-SY5Y neuroblastoma | 1.5 | 100 | 20 | - | 60 | - | 20 | ↑ERK1/2, ↑p-ERK |
Summary of in-vitro applications of low-intensity focused ultrasound on various cell culture lines. iPSCs–NCSCs: Pluripotent stem cells-derived neural crest stem cells; NSPC: neural stem/progenitor cell; hBM-MSC: human bone marrow mesenchymal stem cells; hAD-MSC: human amnion-derived mesenchymal stem cell; NSCs: neural stem cells; BMSC: bone marrow mesenchymal stem cell; F: frequency; PRF: pulse repetition frequency; DC: duty cycle; TBD: tone burst duration; Ispta: spatial-peak temporal-average intensity; Isppa: spatial-peak pulse-average intensity; SD: sonication duration, ↑: increase; ↓: decrease; =no changes.
Table 3.
In-vivo applications.
| Study | Animal Model | Brain Targets |
F MHz |
PRF Hz | DC % | TBD ms | Ispta mW/cm2 | Isppa W/cm2 | SD mins | Outcome |
| [30] | Healthy wild-type mice | Unilateral hippocampi | 0.35 | 1500 | - | 0.0002 | 36.2 | - | 30 | ↑BDNF+ |
| [38] | AD in SD rats | Bilateral hippocampi | 1 | 1 | 5 | 50 | 528 | - | 15 | ↑BDNF, ↑GDNF, ↑VEGF ↓AChE, ↓Aβ |
| [116] | MS in male SD rats | Bilateral hemispheres | 1 | 1 | 5 | 50 | 528 | - | 15 | ↑BDNF, and ↑GDNF, =VEGF, =GLUT1 |
| [124] | Ischemic model in C57BL/6N mice | Whole brain | 0.04 | - | - | - | 50 | - | 20 | ↑p-ERK, ↑p-CREB |
| [89] | Healthy C57BL/6 mice | Unilateral hippocampi | 1.68 | 1 | 1 | 10 | 81.12 | - | 2 | =BrdU-NeuN or DCX |
| [117] | BCCAO in rats | Bilateral hemispheres | 1 | 1 | 5 | 50 | 528 | - | 15 | ↑BDNF |
| [118] | TBI in male C57BL/6 J mice | Injured cortical areas | 1 | 1 | 5 | 50 | 528 | - | 5 | ↑BDNF, ↑VEGF, ↑TrkB/Akt-CREB, =GDNF |
| [120] | MCAO in C57BL/6 mice | Injured cortical area | 1 | 1 | 5 | 50 | 528 | - | 15 | ↑BDNF |
| [123] | VD in male C57BL/6 mice | Whole brain | 1.875 | 6000 | - | - | 90 | - | 20 | ↑t-eNOS, ↑CXCR4, ↑FGF2, ↑VEGF, ↑NGF |
| AD in 5XFAD transgenic mice | Whole brain | 1.875 | 6000 | - | - | 90 | 20 | ↑t-eNOS, ↓Aβ, ↑Hsp 90, ↑NGF, ↑pro-BDNF, =VEGF | ||
| [119] | dMCAO in adult male SD rats | Whole brain | 0.5 | 1000 | - | 0.5 | - | 2.6 | 10 | ↑ BDNF |
| [121] | MCAO in C57BL6J mice | Injured cortical area | 1 | 1 | 50 | - | 528 | - | 15 | ↑BDNF and ↑Bcl-2/ Bax |
| [90] | AD in male C57BL/6 J mice | Hippocampus and Cortex | 1 | 1 | 5 | - | 528 | - | 15 | ↑TLR4/NF-κB, ↑CREB/BDNF, ↓TNF-α, ↓IL-1β, ↓IL-6 |
| [162] | MCAO in adult male ICR mice | Ipsilateral hemisphere | 0.5 | 1000 | - | 0.5 | 120 | - | 10 | ↑M2 microglia, ↑IL-10, ↑IL-10R |
| [41] | Female C57BL/6 mice treated with demyelinating drug CPZ | Whole brain | 1.5 | 1000 | 20 | - | 25 | - | 20 | ↑DCX, ↑BDNF |
| [39] | AD in Wistar male rats | Whole brain | 3.3 | 100 | 50 | 5 | - | 0.8 | 5 | ↑BDNF, ↑NGF-β, ↑IL-10 |
| [40] | MCAO in Wild-type mice | Whole brain | 0.5 | 780 | - | 0.064 | 193 | - | 20 | ↑VEGF, ↑eNOS, ↑DCX+, ↑SDF-1α, ↑CXCR4 |
| [33] | Aged C57BL/6 wild-type mice | Whole brain | 1 | 10 | 10 | 20000 | 120 | - | 4 | ↑DCX |
| [87] | Healthy wildtype mice | Whole brain | 1 | 1000 | 1 | 5 | - | 5 | ↑DCX, ↑p-ERK | |
| [122] | PD model in Female SD rats | Right striatum | 1 | 1 | 5 | 50 | 528 | - | 5 | ↑GDNF, ↓LCN2, ↓IL-1β |
| [161] | TBI in male C67BL/6 N rat | Hippocampus | 0.8 | - | - | - | - | 3 | 10 | ↑Brdu+NeuN+, ↑Dcx |
| [160] | PD in C57BL/6 wild-type mice | CA1 area | 1 | 1 | 5 | 50 | 8.67 | - | 25 | ↑LFP |
Summary of in-vivo applications of LIFUS on animal models. AD: Alzheimer disease; MS: Multiple sclerosis; BCCAO: Bilateral common carotid artery occlusion; TBI: MCAO: Middle cerebral artery occlusion; VD: Vascular dementia; dMCAO: distal middle cerebral artery occlusion; CPZ: Caprizone; PD: Parkinson disease; F: frequency; PRF: pulse repetition frequency; DC: duty cycle; TBD: tone burst duration; Ispta: spatial-peak temporal-average intensity; Isppa: spatial-peak pulse-average intensity; SD: sonication duration; ↑: increase; ↓: decrease; =no changes.
These findings support the hypothesis that neuromodulation-based LIFUS therapy offers unique advantages in promoting neural repair, enhancing neuronal plasticity, and improving cognition, without demonstrating any adverse effects. However, researchers have extensively explored various stimulation paradigms, including parameters and experimental protocols, prioritizing their therapeutic effects after an injury or disease. Table 4 maps the frequency-intensity parameters employed by various studies reviewed in this work. Many studies have focused on a 1 MHz frequency, due to its effective penetration and interaction with brain tissue. This frequency is used in combination with varying intensities, emerging as the most employed approach. Across these intensities, the majority remained below 720 mW/cm2, aligning with FDA recommendations to minimize thermal damage risks associated with higher intensities. Alternatively, some studies have explored a 1.5 MHz frequency with intensities below 50 mW/cm2. In contrast, other studies have employed a lower frequency of 0.5 MHz but with higher intensities reaching 2600 mW/cm2, compromising penetration depth and potential thermal effects. On the other hand, LIFUS with optimal parameters may offer non-invasive cell stimulation to enhance growth factors and neuronal viability. Therefore, ultrasound parameters must be carefully selected to achieve the desired biological effects, cell survival, and proliferation [91]. Subsequently, an overexposure in terms of duration or intensity could destroy the cell membrane, reducing neuronal survival and proliferation [148]. Based on our analysis, claiming an optimal parameter set proves challenging due to diverse experimental setups and study goals. Further studies investigating acceptable frequency and intensity ranges are needed. Establishing standardized protocols for coupling these parameters would reduce variability, enhance reproducibility across studies, and advance our understanding of ultrasound interactions with the adult brain.
Ultrasound Parameters
The studies reviewed in this work utilized a range of frequencies from 0.04 MHz to 4.36 MHz. Lower frequencies (up to 0.5 MHz) allow deeper penetration and broader focal regions, while higher frequencies (3.3 and 4.36 MHz) provide more precise targeting but with limited depth penetration. This limitation restricts high-frequency ultrasound to superficial or near-surface brain regions, making it less suitable for conditions requiring deeper brain stimulation. The tradeoff makes 1 MHz suitable for targeting deeper brain structures without excessive attenuation, which is a frequency more commonly used in these studies (Table 4). The intensity of ultrasound is another crucial parameter influencing the targeted tissue’s biological effects. LIFUS, up to 100 mW/cm2, has been associated with increased cell viability and proliferation without causing significant thermal or mechanical damage. This intensity range typically avoids exceeding the thermal index (TI), thus minimizing the risk of tissue heating and related damage. Moderate-intensity ultrasound, ranging from 100 to 528 mW/cm², appears to strike an optimal balance for promoting cellular activity and repair in most studies. Higher-intensity ultrasound, above 800 mW/cm², may enhance cellular responses by increasing activity, upregulating growth factors, and promoting the expression of genes involved in cellular repair and neuroprotection. However, this increased efficacy comes with a heightened risk of thermal and mechanical damage. The latter can be measured via TI, which is a dimensionless parameter that indicates the potential for tissue heating due to ultrasound exposure. Generally, TI > 1 indicates that the ultrasound exposure could potentially raise tissue temperature by more than 1°C, which may still cause thermal damage if maintained for extended periods [163,164]. Although some reviewed studies exceeded TI > 2.5, all showed neurogenesis except the study with the highest TI [152,165]. For mechanical damage, the MI provides an estimate of the likelihood of non-thermal, mechanical effects such as cavitation that can potentially damage tissue. MI < 0.3 is considered safe with minimal risk of cavitation, commonly used in diagnostic ultrasound. For MI between 0.3 and 1, it is an intermediate range where the potential for cavitation exists, particularly in tissues with pre-existing microbubbles or gas bodies [146,166]. That being said, all studies did not exceed an MI of 0.3, and no tissue damage has been reported or observed. The balance between efficacy and safety is a recurring theme in the application of LIFUS. Figure 3 illustrates the variability of studies based on TI and Mechanical Index MI parameters. Maintaining TI and MI below recommended thresholds minimizes the risk of thermal and tissue damage during treatment. Further studies are needed to tailor frequency and intensity settings to enhance efficacy and safety, especially considering the long-term effects of repeated ultrasound exposure on brain tissue.
Future Perspectives
To date, ultrasound neuromodulation has shown promise in enhancing brain plasticity and supporting neuronal repair, particularly following disease or injury. In various animal models, including those with injuries or neurodegenerative diseases, studies have reported positive outcomes in promoting neural recovery and improving cognitive function. However, the biomechanical mechanisms and treatment efficacy of LIFUS still require thorough investigation to establish a reliable, non-invasive treatment suitable for clinical applications. Additionally, ultrasound parameters must adhere to biosafety standards to ensure safety and efficacy before clinical translation. More systematic studies are needed to elucidate underlying neural mechanisms to optimize ultrasound's effectiveness in both in-vivo and in-vitro settings at the cellular and molecular levels. The wide range of ultrasound parameters used underscores the need for a comprehensive investigation of LIFUS effects on the adult brain. While behavioral tests have demonstrated ultrasound's efficacy in enhancing cognition, its long-term behavioral impact remains poorly understood. Therefore, further research is needed to consider ultrasound parameters that maximize modulation efficacy while minimizing potential side effects and tissue damage.
Acknowledgements
The authors would like to thank the Neural Engineering and Nanobiosensors Group at the American University of Beirut for their support during the preparation of this manuscript.
Conflict of Interest
The authors declare no conflict of interest.
References
- Kirmani, B.F., L.A. Shapiro, and A.K. Shetty, Neurological and neurodegenerative disorders: novel concepts and treatment. Aging and disease, 2021. 12(4): p. 950. [CrossRef]
- Rahman, M.M., et al., Stem cell transplantation therapy and neurological disorders: current status and future perspectives. Biology, 2022. 11(1): p. 147. [CrossRef]
- Lamptey, R.N., et al., A review of the common neurodegenerative disorders: current therapeutic approaches and the potential role of nanotherapeutics. International journal of molecular sciences, 2022. 23(3): p. 1851. [CrossRef]
- Cenini, G., A. Lloret, and R. Cascella, Oxidative stress in neurodegenerative diseases: from a mitochondrial point of view. Oxidative medicine and cellular longevity, 2019. 2019. [CrossRef]
- Elfawy, H.A. and B. Das, Crosstalk between mitochondrial dysfunction, oxidative stress, and age related neurodegenerative disease: Etiologies and therapeutic strategies. Life sciences, 2019. 218: p. 165-184. [CrossRef]
- Russo, I., S. Barlati, and F. Bosetti, Effects of neuroinflammation on the regenerative capacity of brain stem cells. Journal of neurochemistry, 2011. 116(6): p. 947-956. [CrossRef]
- Pizzino, G., et al., Oxidative stress: harms and benefits for human health. Oxidative medicine and cellular longevity, 2017. 2017. [CrossRef]
- Christen, Y., Oxidative stress and Alzheimer disease. The American journal of clinical nutrition, 2000. 71(2): p. 621S-629S.
- Yuan, T.-F., et al., Oxidative stress and adult neurogenesis. Stem Cell Reviews and Reports, 2015. 11: p. 706-709. [CrossRef]
- Rubinsztein, D.C., How does the Huntington's disease mutation damage cells? Science of Aging Knowledge Environment, 2003. 2003(37): p. pe26-pe26.
- Lücking, C. and A. Brice*, Alpha-synuclein and Parkinson's disease. Cellular and Molecular Life Sciences CMLS, 2000. 57: p. 1894-1908.
- Bayés-Genis, A., A. González, and J. Lupón, ST2 in Heart Failure. 2018. p. e005582.
- Winner, B., Z. Kohl, and F.H. Gage, Neurodegenerative disease and adult neurogenesis. European Journal of Neuroscience, 2011. 33(6): p. 1139-1151. [CrossRef]
- Terreros-Roncal, J., et al., Impact of neurodegenerative diseases on human adult hippocampal neurogenesis. Science, 2021. 374(6571): p. 1106-1113. [CrossRef]
- Ahmed, R.M., et al., Physiological changes in neurodegeneration—mechanistic insights and clinical utility. Nature Reviews Neurology, 2018. 14(5): p. 259-271. [CrossRef]
- Rezaei, S., et al., Current therapies for neurological disorders and their limitations, in Phytonutrients and Neurological Disorders. 2023, Elsevier. p. 107-130.
- Vashist, A., et al., Recent advances in nanotherapeutics for neurological disorders. ACS Applied Bio Materials, 2023. 6(7): p. 2614-2621.
- Beisteiner, R., M. Hallett, and A.M. Lozano, Ultrasound neuromodulation as a new brain therapy. Advanced Science, 2023. 10(14): p. 2205634. [CrossRef]
- Ha, S., et al., Direct inductive stimulation for energy-efficient wireless neural interfaces. Conf Proc IEEE Eng Med Biol Soc, 2012. 2012: p. 883-6.
- Cheyuo, C., M. Aziz, and P. Wang, Neurogenesis in Neurodegenerative Diseases: Role of MFG-E8. Front Neurosci, 2019. 13: p. 569. [CrossRef]
- Krauss, J.K., et al., Technology of deep brain stimulation: current status and future directions. Nature Reviews Neurology, 2021. 17(2): p. 75-87. [CrossRef]
- Elias, W.J., et al., A randomized trial of focused ultrasound thalamotomy for essential tremor. New England Journal of Medicine, 2016. 375(8): p. 730-739. [CrossRef]
- Bond, A.E., et al., Safety and efficacy of focused ultrasound thalamotomy for patients with medication-refractory, tremor-dominant Parkinson disease: a randomized clinical trial. JAMA neurology, 2017. 74(12): p. 1412-1418. [CrossRef]
- Croft, P.G., The assessment of pain perception. Journal of Mental Science, 1952. 98(412): p. 427-432.
- Kubanek, J., Neuromodulation with transcranial focused ultrasound. Neurosurgical focus, 2018. 44(2): p. E14. [CrossRef]
- Zhang, T., et al., Excitatory-inhibitory modulation of transcranial focus ultrasound stimulation on human motor cortex. CNS Neuroscience & Therapeutics, 2023. 29(12): p. 3829-3841. [CrossRef]
- Meng, Y., K. Hynynen, and N. Lipsman, Applications of focused ultrasound in the brain: from thermoablation to drug delivery. Nature Reviews Neurology, 2021. 17(1): p. 7-22. [CrossRef]
- Collins, M.N. and K.A. Mesce, A review of the bioeffects of low-intensity focused ultrasound and the benefits of a cellular approach. Frontiers in physiology, 2022. 13: p. 1047324. [CrossRef]
- Badawe, H.M., P. Raad, and M.L. Khraiche, High-resolution acoustic mapping of tunable gelatin-based phantoms for ultrasound tissue characterization. Front Bioeng Biotechnol, 2024. 12: p. 1276143. [CrossRef]
- Tufail, Y., et al., Transcranial pulsed ultrasound stimulates intact brain circuits. Neuron, 2010. 66(5): p. 681-694. [CrossRef]
- Blackmore, D.G., D. Razansky, and J. Götz, Ultrasound as a versatile tool for short-and long-term improvement and monitoring of brain function. Neuron, 2023. 111(8): p. 1174-1190. [CrossRef]
- Badawe, H.M., et al., Experimental and Computational Analysis of High-Intensity Focused Ultrasound Thermal Ablation in Breast Cancer Cells: Monolayers vs. Spheroids. Cancers (Basel), 2024. 16(7). [CrossRef]
- Blackmore, D.G., et al., Low-intensity ultrasound restores long-term potentiation and memory in senescent mice through pleiotropic mechanisms including NMDAR signaling. Molecular Psychiatry, 2021. 26(11): p. 6975-6991. [CrossRef]
- Khraiche, M.L., et al. Ultrasound induced increase in excitability of single neurons. in 2008 30th Annual International Conference of the IEEE Engineering in Medicine and Biology Society. 2008. IEEE.
- Blackmore, J., et al., Ultrasound neuromodulation: a review of results, mechanisms and safety. Ultrasound in medicine & biology, 2019. 45(7): p. 1509-1536. [CrossRef]
- Badawe, H., J.P. Harouz, and M. Khraiche. Effect of Ultrasound-Induced Temperature on the Dynamics of the Hodgkin-Huxley Neuron. in 2024 46th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC). 2024. IEEE.
- Badawe, H. and M. Khraiche, Modeling of Calcium-dependent Low Intensity Low Frequency Ultrasound Modulation of a Hodgkin–Huxley Neuron. Proceedings of EMBC 2024 46th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, 2023/7.
- Lin, W.-T., et al., Protective effects of low-intensity pulsed ultrasound on aluminum-induced cerebral damage in Alzheimer's disease rat model. Scientific reports, 2015. 5(1): p. 9671. [CrossRef]
- dos Santos Tramontin, N., et al., Effects of low-intensity transcranial pulsed ultrasound treatment in a model of Alzheimer's disease. Ultrasound in Medicine & Biology, 2021. 47(9): p. 2646-2656. [CrossRef]
- Ichijo, S., et al., Low-intensity pulsed ultrasound therapy promotes recovery from stroke by enhancing angio-neurogenesis in mice in vivo. Scientific Reports, 2021. 11(1): p. 4958. [CrossRef]
- Guo, H., et al., Exploratory study on neurochemical effects of low-intensity pulsed ultrasound in brains of mice. Medical & Biological Engineering & Computing, 2021. 59: p. 1099-1110. [CrossRef]
- Habli, Z., et al., Single-cell fluid-based force spectroscopy reveals near lipid size nano-topography effects on neural cell adhesion. Lab Chip, 2024. 24(4): p. 707-718. [CrossRef]
- Leal-Galicia, P., et al., Adult neurogenesis: a story ranging from controversial new neurogenic areas and human adult neurogenesis to molecular regulation. International journal of molecular sciences, 2021. 22(21): p. 11489. [CrossRef]
- Alonso, M., A.-C. Petit, and P.-M. Lledo, The impact of adult neurogenesis on affective functions: of mice and men. Molecular Psychiatry, 2024: p. 1-16. [CrossRef]
- Zhao, J., et al., Versatile strategies for adult neurogenesis: Avenues to repair the injured brain. Neural Regeneration Research, 2024. 19(4): p. 774-780. [CrossRef]
- Urbán, N. and F. Guillemot, Neurogenesis in the embryonic and adult brain: same regulators, different roles. Frontiers in cellular neuroscience, 2014. 8: p. 396. [CrossRef]
- Lie, D.C., et al., Neurogenesis in the adult brain: new strategies for central nervous system diseases. Annu. Rev. Pharmacol. Toxicol., 2004. 44: p. 399-421. [CrossRef]
- Alvarez-Buylla, A. and D.A. Lim, For the long run: maintaining germinal niches in the adult brain. Neuron, 2004. 41(5): p. 683-686.
- Lledo, P.-M., M. Alonso, and M.S. Grubb, Adult neurogenesis and functional plasticity in neuronal circuits. Nature Reviews Neuroscience, 2006. 7(3): p. 179-193. [CrossRef]
- Sahay, A., et al., Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature, 2011. 472(7344): p. 466-470. [CrossRef]
- Li, Y.-D., et al., Hypothalamic modulation of adult hippocampal neurogenesis in mice confers activity-dependent regulation of memory and anxiety-like behavior. Nature neuroscience, 2022. 25(5): p. 630-645. [CrossRef]
- Chamaa, F., et al., Thalamic stimulation in awake rats induces neurogenesis in the hippocampal formation. Brain stimulation, 2016. 9(1): p. 101-108. [CrossRef]
- Chamaa, F., et al., Long-term stimulation of the anteromedial thalamus increases hippocampal neurogenesis and spatial reference memory in adult rats. Behavioural brain research, 2021. 402: p. 113114. [CrossRef]
- Chamaa, F., et al., Sustained activation of the anterior thalamic neurons with low doses of Kainic Acid boosts hippocampal neurogenesis. Cells, 2022. 11(21): p. 3413. [CrossRef]
- Ming, G.-l. and H. Song, Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron, 2011. 70(4): p. 687-702. [CrossRef]
- Fontán-Lozano, Á., et al., To become or not to become tumorigenic: subventricular zone versus hippocampal neural stem cells. Frontiers in Oncology, 2020. 10: p. 602217. [CrossRef]
- Kempermann, G., H. Song, and F.H. Gage, Neurogenesis in the adult hippocampus. Cold Spring Harbor perspectives in biology, 2015. 7(9): p. a018812.
- Ehninger, D. and G. Kempermann, Neurogenesis in the adult hippocampus. Cell and tissue research, 2008. 331: p. 243-250.
- Christian, K.M., H. Song, and G.-l. Ming, Functions and dysfunctions of adult hippocampal neurogenesis. Annual review of neuroscience, 2014. 37: p. 243-262. [CrossRef]
- Lim, D.A. and A. Alvarez-Buylla, The adult ventricular–subventricular zone (V-SVZ) and olfactory bulb (OB) neurogenesis. Cold Spring Harbor perspectives in biology, 2016. 8(5): p. a018820. [CrossRef]
- Nissant, A., et al., Adult neurogenesis promotes synaptic plasticity in the olfactory bulb. Nature neuroscience, 2009. 12(6): p. 728-730. [CrossRef]
- Hack, M.A., et al., Neuronal fate determinants of adult olfactory bulb neurogenesis. Nature neuroscience, 2005. 8(7): p. 865-872. [CrossRef]
- Sanai, N., et al., Corridors of migrating neurons in the human brain and their decline during infancy. Nature, 2011. 478(7369): p. 382-386. [CrossRef]
- Khraiche, M.L., et al., Sustained elevation of activity of developing neurons grown on polyimide microelectrode arrays (MEA) in response to ultrasound exposure. Microsystem Technologies, 2017. 23: p. 3671-3683. [CrossRef]
- Taupin, P., BrdU immunohistochemistry for studying adult neurogenesis: paradigms, pitfalls, limitations, and validation. Brain research reviews, 2007. 53(1): p. 198-214. [CrossRef]
- Duque, A. and R. Spector, A balanced evaluation of the evidence for adult neurogenesis in humans: implication for neuropsychiatric disorders. Brain Structure and Function, 2019. 224(7): p. 2281-2295. [CrossRef]
- Zheng, Y., et al., Increased BrdU incorporation reflecting DNA repair, neuronal de-differentiation or possible neurogenesis in the adult cochlear nucleus following bilateral cochlear lesions in the rat. Experimental brain research, 2011. 210: p. 477-487. [CrossRef]
- Duque, A. and P. Rakic, Different effects of bromodeoxyuridine and [3H] thymidine incorporation into DNA on cell proliferation, position, and fate. Journal of Neuroscience, 2011. 31(42): p. 15205-15217.
- Maga, G. and U. Hubscher, Proliferating cell nuclear antigen (PCNA): a dancer with many partners. Journal of cell science, 2003. 116(15): p. 3051-3060. [CrossRef]
- Ayanlaja, A.A., et al., Distinct features of doublecortin as a marker of neuronal migration and its implications in cancer cell mobility. Frontiers in molecular neuroscience, 2017. 10: p. 199. [CrossRef]
- Dayer, A.G., et al., New GABAergic interneurons in the adult neocortex and striatum are generated from different precursors. The Journal of cell biology, 2005. 168(3): p. 415-427. [CrossRef]
- Zuccato, C. and E. Cattaneo, Brain-derived neurotrophic factor in neurodegenerative diseases. Nature Reviews Neurology, 2009. 5(6): p. 311-322. [CrossRef]
- Dawbarn, D. and S. Allen, Neurotrophins and neurodegeneration. Neuropathology and applied neurobiology, 2003. 29(3): p. 211-230.
- Fumagalli, F., et al., Neurotrophic factors in neurodegenerative disorders: potential for therapy. CNS drugs, 2008. 22: p. 1005-1019.
- Rossi, C., et al., Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. European Journal of Neuroscience, 2006. 24(7): p. 1850-1856. [CrossRef]
- Tseng, K.-Y., et al., MANF promotes differentiation and migration of neural progenitor cells with potential neural regenerative effects in stroke. Molecular therapy, 2018. 26(1): p. 238-255. [CrossRef]
- Cortés, D., et al., The non-survival effects of glial cell line-derived neurotrophic factor on neural cells. Frontiers in Molecular Neuroscience, 2017. 10: p. 258. [CrossRef]
- d’Anglemont de Tassigny, X., A. Pascual, and J. López-Barneo, GDNF-based therapies, GDNF-producing interneurons, and trophic support of the dopaminergic nigrostriatal pathway. Implications for Parkinson’s disease. Frontiers in neuroanatomy, 2015. 9: p. 10.
- Wang, F., et al., GDNF-pretreatment enhances the survival of neural stem cells following transplantation in a rat model of Parkinson's disease. Neuroscience research, 2011. 71(1): p. 92-98. [CrossRef]
- Duffy, A.M., D.J. Bouchier-Hayes, and J.H. Harmey, Vascular endothelial growth factor (VEGF) and its role in non-endothelial cells: autocrine signalling by VEGF, in Madame Curie Bioscience Database [Internet]. 2013, Landes Bioscience. [CrossRef]
- Barth, K., et al., EGFL7 loss correlates with increased VEGF-D expression, upregulating hippocampal adult neurogenesis and improving spatial learning and memory. Cellular and Molecular Life Sciences, 2023. 80(2): p. 54.
- Zhang, L., H. Jiang, and Z. Hu, Concentration-dependent effect of nerve growth factor on cell fate determination of neural progenitors. Stem Cells and Development, 2011. 20(10): p. 1723-1731. [CrossRef]
- Wang, B., et al., Erk1/2 promotes proliferation and inhibits neuronal differentiation of neural stem cells. Neuroscience letters, 2009. 461(3): p. 252-257. [CrossRef]
- Choi, K.-C., et al., Effect of single growth factor and growth factor combinations on differentiation of neural stem cells. Journal of Korean Neurosurgical Society, 2008. 44(6): p. 375. [CrossRef]
- Nakajima, M., et al., Combinatorial protein display for the cell-based screening of biomaterials that direct neural stem cell differentiation. Biomaterials, 2007. 28(6): p. 1048-1060. [CrossRef]
- Levenberg, S., et al., Neurotrophin-induced differentiation of human embryonic stem cells on three-dimensional polymeric scaffolds. Tissue engineering, 2005. 11(3-4): p. 506-512. [CrossRef]
- Lim, J., et al., ASIC1a is required for neuronal activation via low-intensity ultrasound stimulation in mouse brain. Elife, 2021. 10: p. e61660. [CrossRef]
- Wang, Z., et al., Transcranial ultrasound stimulation directly influences the cortical excitability of the motor cortex in Parkinsonian mice. Movement Disorders, 2020. 35(4): p. 693-698. [CrossRef]
- Mooney, S.J., et al., Focused ultrasound-induced neurogenesis requires an increase in blood-brain barrier permeability. PloS one, 2016. 11(7): p. e0159892. [CrossRef]
- Chen, T.-T., T.-H. Lan, and F.-Y. Yang, Low-intensity pulsed ultrasound attenuates LPS-induced neuroinflammation and memory impairment by modulation of TLR4/NF-κB signaling and CREB/BDNF expression. Cerebral Cortex, 2019. 29(4): p. 1430-1438. [CrossRef]
- Lee, I.-C., et al., Differentiation of neural stem/progenitor cells using low-intensity ultrasound. Ultrasound in medicine & biology, 2014. 40(9): p. 2195-2206. [CrossRef]
- Harvey, E.N., The effect of high frequency sound waves on heart muscle and other irritable tissues. American Journal of Physiology-Legacy Content, 1929. 91(1): p. 284-290. [CrossRef]
- Khraiche, M.L., et al., Ultrasound induced increase in excitability of single neurons. Annu Int Conf IEEE Eng Med Biol Soc, 2008. 2008: p. 4246-9.
- Fry, F., H. Ades, and W. Fry, Production of reversible changes in the central nervous system by ultrasound. Science, 1958. 127(3289): p. 83-84. [CrossRef]
- Koroleva, V., N. Vykhodtseva, and V. Elagin, Spreading depression in the cortex and subcortical structures of the brain of the rat induced by exposure to focused ultrasound. Neirofiziologiia= Neurophysiology, 1986. 18(1): p. 55-61.
- Velling, V. and S. Shklyaruk, Modulation of the functional state of the brain with the aid of focused ultrasonic action. Neuroscience and behavioral physiology, 1988. 18: p. 369-375. [CrossRef]
- Muratore, R., et al. Bioeffective ultrasound at very low doses: Reversible manipulation of neuronal cell morphology and function in vitro. in AIP Conference Proceedings. 2009. American Institute of Physics.
- Tyler, W.J., et al., Remote excitation of neuronal circuits using low-intensity, low-frequency ultrasound. PloS one, 2008. 3(10): p. e3511. [CrossRef]
- Kim, H., et al., Noninvasive transcranial stimulation of rat abducens nerve by focused ultrasound. Ultrasound in medicine & biology, 2012. 38(9): p. 1568-1575. [CrossRef]
- Kim, H., et al., Suppression of EEG visual-evoked potentials in rats through neuromodulatory focused ultrasound. Neuroreport, 2015. 26(4): p. 211-215. [CrossRef]
- King, R.L., et al., Effective parameters for ultrasound-induced in vivo neurostimulation. Ultrasound in medicine & biology, 2013. 39(2): p. 312-331. [CrossRef]
- King, R.L., J.R. Brown, and K.B. Pauly, Localization of ultrasound-induced in vivo neurostimulation in the mouse model. Ultrasound in medicine & biology, 2014. 40(7): p. 1512-1522. [CrossRef]
- Papp, T., et al., Ultrasound Used for Diagnostic Imaging Facilitates Dendritic Branching of Developing Neurons in the Mouse Cortex. Frontiers in Neuroscience, 2022. 16: p. 803356. [CrossRef]
- Hameroff, S., et al., Transcranial ultrasound (TUS) effects on mental states: a pilot study. Brain stimulation, 2013. 6(3): p. 409-415. [CrossRef]
- Kubanek, J., et al., Remote, brain region–specific control of choice behavior with ultrasonic waves. Science advances, 2020. 6(21): p. eaaz4193.
- Yang, P.-F., et al., Neuromodulation of sensory networks in monkey brain by focused ultrasound with MRI guidance and detection. Scientific reports, 2018. 8(1): p. 7993. [CrossRef]
- Legon, W., et al., Transcranial focused ultrasound modulates the activity of primary somatosensory cortex in humans. Nature neuroscience, 2014. 17(2): p. 322-329. [CrossRef]
- Mueller, J., et al., Transcranial focused ultrasound modulates intrinsic and evoked EEG dynamics. Brain stimulation, 2014. 7(6): p. 900-908. [CrossRef]
- Lee, W., et al., Image-guided transcranial focused ultrasound stimulates human primary somatosensory cortex. Scientific reports, 2015. 5(1): p. 8743. [CrossRef]
- Deffieux, T., et al., Low-intensity focused ultrasound modulates monkey visuomotor behavior. Current Biology, 2013. 23(23): p. 2430-2433. [CrossRef]
- Sanguinetti, J.L., et al., Transcranial focused ultrasound to the right prefrontal cortex improves mood and alters functional connectivity in humans. Frontiers in human neuroscience, 2020. 14: p. 494085. [CrossRef]
- Folloni, D., et al., Manipulation of subcortical and deep cortical activity in the primate brain using transcranial focused ultrasound stimulation. Neuron, 2019. 101(6): p. 1109-1116. e5. [CrossRef]
- Verhagen, L., et al., Offline impact of transcranial focused ultrasound on cortical activation in primates. Elife, 2019. 8: p. e40541. [CrossRef]
- Yuan, T.-F., et al., Neuromodulation-based stem cell therapy in brain repair: recent advances and future perspectives. Neuroscience Bulletin, 2021. 37: p. 735-745. [CrossRef]
- Wu, Y., et al., Low-intensity pulsed ultrasound regulates proliferation and differentiation of neural stem cells through notch signaling pathway. Biochemical and biophysical research communications, 2020. 526(3): p. 793-798. [CrossRef]
- Yang, F.-Y., et al., Enhancement of neurotrophic factors in astrocyte for neuroprotective effects in brain disorders using low-intensity pulsed ultrasound stimulation. Brain stimulation, 2015. 8(3): p. 465-473. [CrossRef]
- Huang, S.-L., et al., Protective effect of low-intensity pulsed ultrasound on memory impairment and brain damage in a rat model of vascular dementia. Radiology, 2017. 282(1): p. 113-122. [CrossRef]
- Su, W.-S., et al., Transcranial ultrasound stimulation promotes brain-derived neurotrophic factor and reduces apoptosis in a mouse model of traumatic brain injury. Brain Stimulation, 2017. 10(6): p. 1032-1041. [CrossRef]
- Liu, L., et al., Readout-segmented echo-planar diffusion-weighted MR at 3.0 T for the evaluation the effect of low-intensity transcranial ultrasound on stroke in a rat model. Magnetic Resonance Imaging, 2020. 67: p. 79-84. [CrossRef]
- Chen, C.-M., et al., Preventive effect of low intensity pulsed ultrasound against experimental cerebral ischemia/reperfusion injury via apoptosis reduction and brain-derived neurotrophic factor induction. Scientific reports, 2018. 8(1): p. 5568. [CrossRef]
- Wu, C.-T., et al., Low intensity pulsed ultrasound prevents recurrent ischemic stroke in a cerebral ischemia/reperfusion injury mouse model via brain-derived neurotrophic factor induction. International Journal of Molecular Sciences, 2019. 20(20): p. 5169. [CrossRef]
- Sung, C.-Y., et al., Low-intensity pulsed ultrasound enhances neurotrophic factors and alleviates neuroinflammation in a rat model of Parkinson’s disease. Cerebral Cortex, 2022. 32(1): p. 176-185. [CrossRef]
- Eguchi, K., et al., Whole-brain low-intensity pulsed ultrasound therapy markedly improves cognitive dysfunctions in mouse models of dementia-Crucial roles of endothelial nitric oxide synthase. Brain Stimulation, 2018. 11(5): p. 959-973. [CrossRef]
- Cho, S.-E., et al., The effect of ultrasound for increasing neural differentiation in hBM-MSCs and inducing neurogenesis in ischemic stroke model. Life Sciences, 2016. 165: p. 35-42. [CrossRef]
- Tyler, W.J., S.W. Lani, and G.M. Hwang, Ultrasonic modulation of neural circuit activity. Current opinion in neurobiology, 2018. 50: p. 222-231. [CrossRef]
- Hassan, R.E., T.E. Chemaly, and M. Khraiche. Towards A Biomechanical Model for Ultrasound Effect on Neural Excitability. in 2018 IEEE International Multidisciplinary Conference on Engineering Technology (IMCET). 2018.
- Khraiche, M.L., J. Rogul, and J. Muthuswamy, Design and Development of Microscale Thickness Shear Mode (TSM) Resonators for Sensing Neuronal Adhesion. Front Neurosci, 2019. 13: p. 518. [CrossRef]
- Khraiche, M. and J. Muthuswamy, Multi-modal biochip for simultaneous, real-time measurement of adhesion and electrical activity of neurons in culture. Lab Chip, 2012. 12(16): p. 2930-41. [CrossRef]
- Legon, W., et al., A retrospective qualitative report of symptoms and safety from transcranial focused ultrasound for neuromodulation in humans. Scientific reports, 2020. 10(1): p. 5573. [CrossRef]
- Krasovitski, B., et al., Intramembrane cavitation as a unifying mechanism for ultrasound-induced bioeffects. Proceedings of the National Academy of Sciences, 2011. 108(8): p. 3258-3263. [CrossRef]
- Plaksin, M., S. Shoham, and E. Kimmel, Intramembrane cavitation as a predictive bio-piezoelectric mechanism for ultrasonic brain stimulation. Physical review X, 2014. 4(1): p. 011004. [CrossRef]
- Plaksin, M., E. Kimmel, and S. Shoham, Cell-type-selective effects of intramembrane cavitation as a unifying theoretical framework for ultrasonic neuromodulation. eneuro, 2016. 3(3). [CrossRef]
- Tyler, W.J., Noninvasive neuromodulation with ultrasound? A continuum mechanics hypothesis. The Neuroscientist, 2011. 17(1): p. 25-36. [CrossRef]
- Ibsen, S., et al., Sonogenetics is a non-invasive approach to activating neurons in Caenorhabditis elegans. Nature communications, 2015. 6(1): p. 8264. [CrossRef]
- Qiu, Z., et al., Targeted neurostimulation in mouse brains with non-invasive ultrasound. Cell reports, 2020. 32(7). [CrossRef]
- Kubanek, J., et al., Ultrasound modulates ion channel currents. Scientific reports, 2016. 6(1): p. 24170. [CrossRef]
- Morris, C.E. and P.F. Juranka, Nav channel mechanosensitivity: activation and inactivation accelerate reversibly with stretch. Biophysical journal, 2007. 93(3): p. 822-833. [CrossRef]
- Venkatachalam, K. and C. Montell, TRP channels. Annu. Rev. Biochem., 2007. 76: p. 387-417.
- Yoo, S., et al., Focused ultrasound excites cortical neurons via mechanosensitive calcium accumulation and ion channel amplification. Nature communications, 2022. 13(1): p. 493. [CrossRef]
- Sukharev, S. and D.P. Corey, Mechanosensitive channels: multiplicity of families and gating paradigms. Science's STKE, 2004. 2004(219): p. re4-re4. [CrossRef]
- Heimburg, T. and A.D. Jackson, On soliton propagation in biomembranes and nerves. Proceedings of the National Academy of Sciences, 2005. 102(28): p. 9790-9795. [CrossRef]
- Petrov, A.G., Flexoelectricity of model and living membranes. Biochimica et Biophysica Acta (BBA)-Biomembranes, 2002. 1561(1): p. 1-25.
- Prieto, M.L., et al., Dynamic response of model lipid membranes to ultrasonic radiation force. PLoS One, 2013. 8(10): p. e77115. [CrossRef]
- Taylor, G.J., et al., Capacitive detection of low-enthalpy, higher-order phase transitions in synthetic and natural composition lipid membranes. Langmuir, 2017. 33(38): p. 10016-10026. [CrossRef]
- Badawe, H.M., R.H. El Hassan, and M.L. Khraiche, Modeling ultrasound modulation of neural function in a single cell. Heliyon, 2023. 9(12). [CrossRef]
- Badawe, H.M. and M.L. Khraiche. Modeling of Calcium-dependent Low Intensity Low Frequency Ultrasound Modulation of a Hodgkin–Huxley Neuron. in 2023 45th Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC). 2023. IEEE.
- Zhao, L., et al., Low-intensity pulsed ultrasound enhances nerve growth factor-induced neurite outgrowth through mechanotransduction-mediated ERK1/2–CREB–Trx-1 signaling. Ultrasound in medicine & biology, 2016. 42(12): p. 2914-2925. [CrossRef]
- Tsuang, Y.H., et al., Effects of low intensity pulsed ultrasound on rat Schwann cells metabolism. Artificial organs, 2011. 35(4): p. 373-383. [CrossRef]
- Ning, G.Z., et al., Bone marrow mesenchymal stem cells stimulated with low-intensity pulsed ultrasound: better choice of transplantation treatment for spinal cord injury: treatment for SCI by LIPUS-BMSCs transplantation. CNS Neuroscience & Therapeutics, 2019. 25(4): p. 496-508.
- Lv, Y., et al., Effects of low-intensity pulsed ultrasound on cell viability, proliferation and neural differentiation of induced pluripotent stem cells-derived neural crest stem cells. Biotechnology letters, 2013. 35: p. 2201-2212. [CrossRef]
- Lee, I.-C., H.-J. Wu, and H.-L. Liu, Dual-frequency ultrasound induces neural stem/progenitor cell differentiation and growth factor utilization by enhancing stable cavitation. ACS Chemical Neuroscience, 2019. 10(3): p. 1452-1461. [CrossRef]
- Schuster, A., et al., Neural stem cells influenced by ultrasound: Frequency and energy density dependencies. Physics in Medicine, 2017. 4: p. 8-16. [CrossRef]
- El Hassan, R.H., et al., Frequency dependent, reversible focused ultrasound suppression of evoked potentials in the reflex arc in an anesthetized animal. J Peripher Nerv Syst, 2022. [CrossRef]
- Xie, S., et al., Low-intensity pulsed ultrasound promotes the proliferation of human bone mesenchymal stem cells by activating PI3K/AKt signaling pathways. Journal of Cellular Biochemistry, 2019. 120(9): p. 15823-15833.
- Ling, L., et al., Low-intensity pulsed ultrasound activates ERK 1/2 and PI 3K-Akt signalling pathways and promotes the proliferation of human amnion-derived mesenchymal stem cells. Cell proliferation, 2017. 50(6): p. e12383. [CrossRef]
- Ren, C., et al., Low-intensity pulsed ultrasound promotes Schwann cell viability and proliferation via the GSK-3β/β-catenin signaling pathway. International journal of biological sciences, 2018. 14(5): p. 497. [CrossRef]
- Al-Maswary, A.A., et al., Low-intensity pulsed ultrasound induces proliferation of human neural stem cells. Clinical and Translational Discovery, 2024. 4(2): p. e259. [CrossRef]
- Liu, S.-H., et al., Ultrasound enhances the expression of brain-derived neurotrophic factor in astrocyte through activation of TrkB-Akt and calcium-CaMK signaling pathways. Cerebral cortex, 2017. 27(6): p. 3152-3160. [CrossRef]
- Adelegan, O.J., et al., Fabrication of 2D Capacitive Micromachined Ultrasonic Transducer (CMUT) Arrays on Insulating Substrates With Through-Wafer Interconnects Using Sacrificial Release Process. Journal of Microelectromechanical Systems, 2020. 29(4): p. 553-561. [CrossRef]
- Zhao, Z., et al., Transcranial Ultrasound Stimulation Improves Memory Performance of Parkinsonian Mice. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 2024. 32: p. 1284-1291. [CrossRef]
- Wang, W., et al., LIPUS-induced neurogenesis: A potential therapeutic strategy for cognitive dysfunction in traumatic brain injury. Experimental Neurology, 2024. 371: p. 114588. [CrossRef]
- Wang, J., et al., Transcranial focused ultrasound stimulation improves neurorehabilitation after middle cerebral artery occlusion in mice. Aging and disease, 2021. 12(1): p. 50. [CrossRef]
- Şen, T., O. Tüfekçioğlu, and Y. Koza, Mechanical index. Anatolian journal of cardiology, 2015. 15(4): p. 334.
- Khraiche, M.L., et al., Ultra-High Photosensitivity Silicon Nanophotonics for Retinal Prosthesis: Electrical Characteristics. Conf Proc IEEE Eng Med Biol Soc, 2011.
- Tarhini, A., et al., The effect of temperature on the electrical and thermal conductivity of graphene-based polymer composite films. Journal of Applied Polymer Science, 2022. 139(14): p. 51896. [CrossRef]
- Chou, Z., et al., Bidirectional neural interface: Closed-loop feedback control for hybrid neural systems. Conf Proc IEEE Eng Med Biol Soc, 2015. 2015: p. 3949-52.
Figure 1.
Traditional neurogenic niches and schematic representation of the main stages of adult neurogenesis. (A) The regions that exhibit distinct processes in adult neurogenesis are the SVZ along the walls of the LV and the SGZ within the hippocampus of a rat. (B) The main stages of neurogenesis occur in both the SGZ and SVZ regions of the adult brain. The schematic representation illustrates the expressed markers and molecular mechanisms involved in these regions, from neural stem cell activation, proliferation, differentiation into neural progenitors, migration, and maturation into mature neurons essential for brain function. DG: Dentate Gyrus; LV: lateral ventricles; SGZ: subgranular zone; GCL: granule cell layer, ML: molecular layer, SVZ: subventricular zone, RMS: rostal migratory stream, OB: olfactory bulb.
Figure 1.
Traditional neurogenic niches and schematic representation of the main stages of adult neurogenesis. (A) The regions that exhibit distinct processes in adult neurogenesis are the SVZ along the walls of the LV and the SGZ within the hippocampus of a rat. (B) The main stages of neurogenesis occur in both the SGZ and SVZ regions of the adult brain. The schematic representation illustrates the expressed markers and molecular mechanisms involved in these regions, from neural stem cell activation, proliferation, differentiation into neural progenitors, migration, and maturation into mature neurons essential for brain function. DG: Dentate Gyrus; LV: lateral ventricles; SGZ: subgranular zone; GCL: granule cell layer, ML: molecular layer, SVZ: subventricular zone, RMS: rostal migratory stream, OB: olfactory bulb.
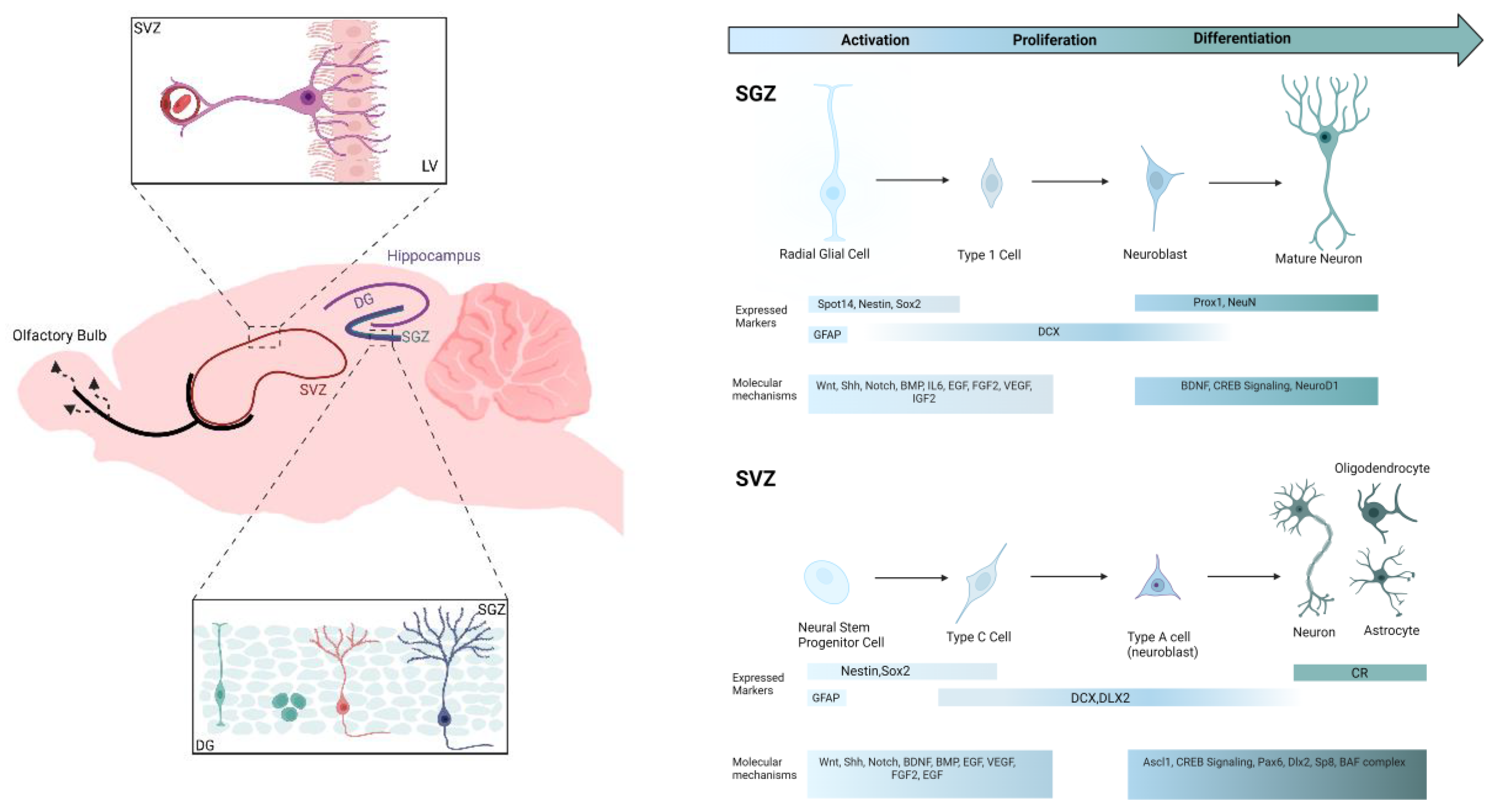
Figure 2.
Ultrasound physical properties and its neuromodulation mechanisms on the cellular level. (A) Ultrasound pulsation scheme at a specified SD depending on the DC, i.e., the ratio of TBD over the PRP with a given ISI. (B) Intramembrane cavitation characterized by the formation of bubbles in the intramembrane space between the lipid bilayer leaflets leading to a change in the membrane capacitance and consequently membrane potential difference. (C) Opening of mechanosensitive ion channels that can detect pressure changes leading to a calcium influx that triggers a train of action potentials. (D) Membrane conformational changes due to a difference in the induced pressure between the intracellular and extracellular fluids leading to a change in membrane geometry, thickness and curvature. The ultrasound-induced mechanical strain causes dipoles redistribution and surface polarization known as direct flexoelectricity. SD: Sonication duration; DC: Duty cycle; TBD: Tone burst duration; PRP: Pulse repetition period; ISI: Inter-stimulus-interval.
Figure 2.
Ultrasound physical properties and its neuromodulation mechanisms on the cellular level. (A) Ultrasound pulsation scheme at a specified SD depending on the DC, i.e., the ratio of TBD over the PRP with a given ISI. (B) Intramembrane cavitation characterized by the formation of bubbles in the intramembrane space between the lipid bilayer leaflets leading to a change in the membrane capacitance and consequently membrane potential difference. (C) Opening of mechanosensitive ion channels that can detect pressure changes leading to a calcium influx that triggers a train of action potentials. (D) Membrane conformational changes due to a difference in the induced pressure between the intracellular and extracellular fluids leading to a change in membrane geometry, thickness and curvature. The ultrasound-induced mechanical strain causes dipoles redistribution and surface polarization known as direct flexoelectricity. SD: Sonication duration; DC: Duty cycle; TBD: Tone burst duration; PRP: Pulse repetition period; ISI: Inter-stimulus-interval.
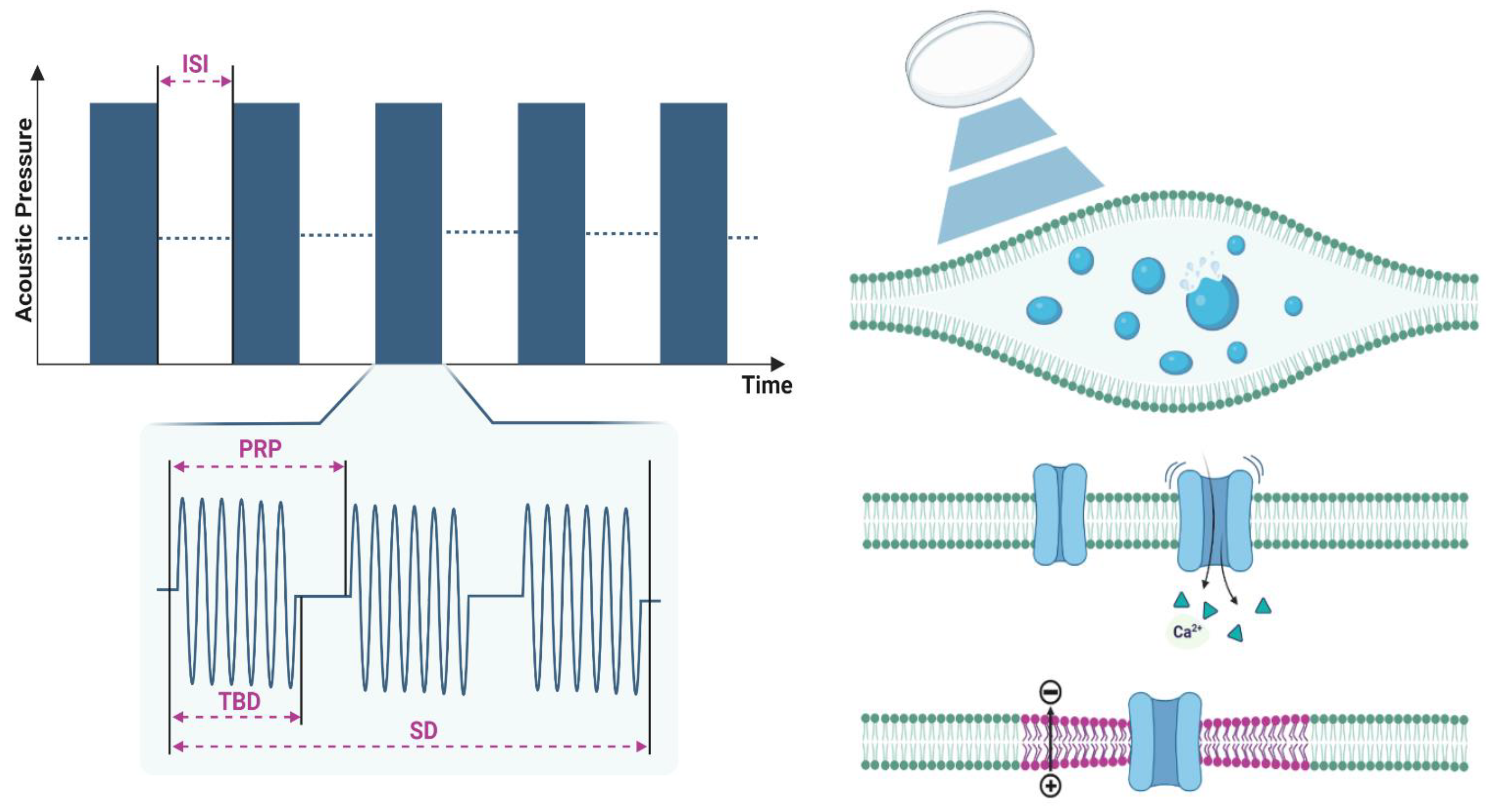
Figure 3.
Impact of frequency and intensity on thermal and mechanical indices. Controlling these indices IS crucial in ensuring the safety and effectiveness of ultrasound applications. (A) Intensity vs. frequency with TI displaying the temperature increase across studies. (B) Intensity vs. frequency with MI indication throughout studies.
Figure 3.
Impact of frequency and intensity on thermal and mechanical indices. Controlling these indices IS crucial in ensuring the safety and effectiveness of ultrasound applications. (A) Intensity vs. frequency with TI displaying the temperature increase across studies. (B) Intensity vs. frequency with MI indication throughout studies.
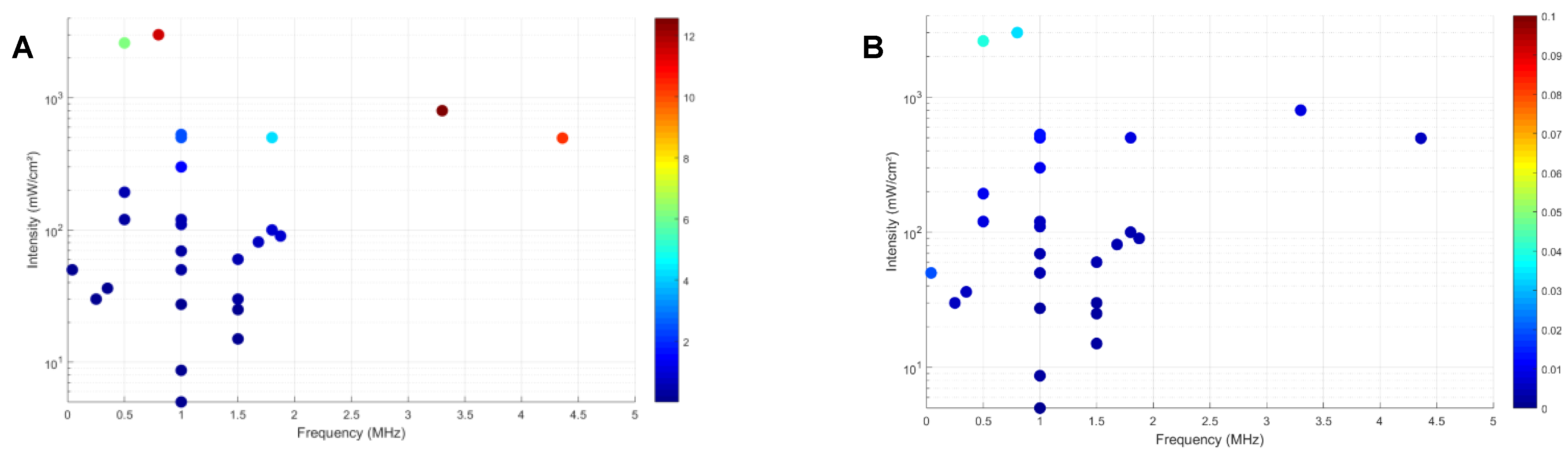
Table 1.
Key biomarkers and their relevant function in the context of neurogenesis.
| Biomarkers | Function |
|---|---|
| Nestin | Supports neural progenitor cells during early development |
| DCX | Indicates ongoing neurogenesis in immature neurons |
| NeuN | Identifies mature neurons (not all types) |
| GFAP | Supports neuron function and health via astrocytes |
| BrdU | Tracks cell proliferation |
| BDNF | Promotes neuron survival, growth, and synapse formation |
| CREB | Regulates neuronal plasticity, learning, and memory |
| PCNA | Supports DNA synthesis and repair in proliferating cells |
| Tuj1 | Identifies early post-mitotic neurons |
| VEGF | Promotes angiogenesis and neuroprotection |
| NGF | Supports neuron survival, growth, and neuroplasticity |
| GLUT1 | Facilitates glucose transport for brain energy |
NESTIN: NSC marker nestin; DCX: Doublecortin; NeuN: Neuronal nuclei; GFAP: Glial fibrillary acidic protein; BrdU: Bromodeoxyuridine; BDNF: Brain-derived neurotrophic factor; CREB: cAMP response element-binding protein; Tuj1: Neuron-specific class III beta-Tubulin; PCNA: Proliferating cell nuclear antigen; VEGF: Vascular endothelial growth factor; NGF: Nerve growth factor; GLUT1: Glucose transporter 1.
Table 4.
Map of frequency and intensity parameters across all studies. Overview of the frequencies and intensities employed, highlighting the diverse approaches used in LIFUS research.
Table 4.
Map of frequency and intensity parameters across all studies. Overview of the frequencies and intensities employed, highlighting the diverse approaches used in LIFUS research.
| Intensity (mW/cm2) | 3000 | [151] | |||||||||||
| 2600 | [114] | ||||||||||||
| 800 | [33] | ||||||||||||
| 528 | [111] | ||||||||||||
| [117] | |||||||||||||
| [86] | |||||||||||||
| [116] | |||||||||||||
| [115] | |||||||||||||
| [113] | |||||||||||||
| [112] | |||||||||||||
| [111] | |||||||||||||
| [32] | |||||||||||||
| 500 | [142] | [87] | |||||||||||
| 496 | [144] | ||||||||||||
| 300 | [140] | ||||||||||||
| 193 | [34] | ||||||||||||
| 120 | [152] | [29] | |||||||||||
| 110 | [149] | ||||||||||||
| 100 | |||||||||||||
| 90 | [118] | ||||||||||||
| 81.12 | [85] | ||||||||||||
| 69.3 | [110] | ||||||||||||
| 60 | [148] | ||||||||||||
| [145] | |||||||||||||
| 50 | [119] | [141] | |||||||||||
| [139] | |||||||||||||
| 36.2 | [27] | ||||||||||||
| 30 | [146] | [153] | |||||||||||
| 27.37 | [147] | ||||||||||||
| 25 | [35] | ||||||||||||
| 15 | [35] | ||||||||||||
| 8.67 | [150] | ||||||||||||
| 5 | [83] | ||||||||||||
| 0.04 | 0.25 | 0.35 | 0.5 | 0.8 | 1 | 1.5 | 1.68 | 1.8 | 1.875 | 3.3 | 4.36 | ||
| Frequency (MHz) | |||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



