Submitted:
12 August 2025
Posted:
13 August 2025
You are already at the latest version
Abstract
Background: As edible insects gain importance as sustainable protein sources, their integration into the food system requires that they meet the same safety standards as conventional animal products. This includes systematic testing for residues of pharmacologically active substances, such as antibiotics. To enable such monitoring, validated analytical methods for insect matrices are essential—but currently lacking. This study evaluates whether LC-MS/MS methods already validated for conventional animal products are suitable for detecting antibiotics in edible insects. Methods: A feeding trial was conducted with Tenebrio molitor larvae using three antibiotics—tiamulin, chloramphenicol, and erythromycin—selected based on prior trace detections, known associations with insect use, or the presence of related resistance genes in insect-based products. The antibiotics were incorporated into feed, and their homogeneity and stability were confirmed. After seven days of feeding and a 24-hour fasting period, larvae were analyzed. Results: All three antibiotics were detected in the insects, although residue levels significantly declined after fasting. Conclusion: These results demonstrate the applicability of existing methods to insect matrices and highlight the need for further validation. The study provides a reproducible model for further feeding trials and discusses potential drivers of antibiotic misuse in insect farming as a foundation for developing and expanding testing methods to ensure the food safety of edible insects.
Keywords:
edible insects
; Tenebrio molitor larvae
; antibiotic residues
; LC-MS/MS analysis
; feeding trial
; food safety monitoring
1. Introduction
With the globally increasing demand for sustainable protein sources, edible insects are gaining growing attention from the scientific community, industry, and policymakers [1,2,3]. Insects for food and feed production offer numerous ecological advantages, such as high feed conversion efficiency, low greenhouse gas emissions, and the ability to be reared on organic waste streams [4,5,6]. As a potential alternative to conventional meat products, they are considered a promising solution for easing pressure on the environment and agricultural systems in the context of climate change and a growing global population [7,8,9].
At the same time, edible insects—as a novel food —face particular challenges regarding food safety and consumer acceptance [10]. Both aspects are essential and mutually reinforcing. Concerns about the uncontrolled use of substances such as antibiotics have already been raised in public discourse [11].
Indeed, the use of antibiotics such as chloramphenicol has been documented in the context of insect rearing for scientific purposes—for instance, in studies on pest insect control or in the rearing of Bombyx mori for silk production [12,13]. The use of silkworm pupae as a by-product of silk production has already been proposed as a potential additional edible insect [14]. Chloramphenicol is banned for the use in food-producing animals under Regulation (EU) No 37/2010 Annex Table 2. Accordingly, reliable detection methods are essential to monitor its potential use in the rearing of edible insects.
Moreover, the planned use of substrates such as manure entails the risk that residues from conventional livestock farming—including antibiotics—could enter insect production systems and ultimately contaminate edible insect products or insect-based feed [15,16]. Conversely, the use of insect frass as fertilizer could pose a risk for the development of antibiotic resistance in soil microbiomes if antibiotics were to be used in insect farming without proper regulation [17,18,19]. Improper feeding practices, such as the use of by-products from the meat industry, could likewise lead to contamination with antibiotics or other pharmacologically active substances [20].
These uncertainties underscore the necessity of systematically monitoring insect-based food products for residues of pharmacologically active substances, as is already standard practice for other animal-derived foods. The restriction of antibiotic use in conventional livestock production is primarily based on concerns over the emergence and spread of antibiotic resistance [21]. Initial studies have already demonstrated that edible insects can serve as carriers of antibiotic-resistant bacteria [22,23,24,25], indicating that the use of antibiotics in insect farming could similarly contribute to the selection and proliferation of resistant strains [26].
As early as 2015, the European Food Safety Authority (EFSA) recommended monitoring edible insects for residues of pharmacologically active substances [15]. Since 2022 by EU law insects have to be included in official control plans. Testing for the pharmaceutical substances listed in the plan is therefore mandatory. Due to the currently low production volumes in Germany, the number of required samples remains limited: only two samples had to be analyzed in total across 2023 and 2024. However, this number is expected to increase in the coming years. By that time at the latest, validated and appropriate analytical methods for the reliable testing of insects must be available [27].
Many questions in this context remain unresolved. Are pharmacologically active substances used at all in insect farming? If so, which active compounds are relevant for commonly farmed edible insect species? Can existing analytical methods be applied, or is there a need to develop new approaches?
As a first attempt to address these questions, our institute screened 14 samples from online retailers in 2020 for various antibiotics using detection methods already validated for animal-based matrices. In total, 75 individual compounds were analyzed, including various tetracyclines, pleuromutilins, and amphenicols. Notably, tiamulin was detected in one of the samples [28].
With the present study, we approached the open questions by initially designing a feeding experiment to gain an idea of whether antibiotics administered to insects can be detected using the analytical methods already established in our laboratories. In this context, we administered the three antibiotics tiamulin, chloramphenicol, and erythromycin to mealworms (Tenebrio molitor larvae) and analyzed them using our existing methods. Finally, we discussed various motivations for the use of pharmacologically active substances in edible insect farming, aiming to identify potential entry points for future research.
2. Results
2.1. Impact of Antibiotics on Mealworm Weight
Tiamulin administration resulted in significantly lower weight in mealworms compared to the control group (p = 0.023, Mann-Whitney U test). In contrast, erythromycin and chloramphenicol also resulted in lower weights compared to the control group, but these differences were not statistically significant (p = 0.219 and p = 0.579, respectively, Mann-Whitney U test). Moreover, while all other groups exhibited an increase in weight over the seven-day period, the group administered tiamulin experienced an average loss of 0.25 g.
Using GC-MS, we detected no residual methanol in the flour of the control groups. There was also no significant weight difference between mealworms in the methanol-treated and untreated control groups (p = 0.544, Mann-Whitney U test). The raw data can be found in the supplementary material S2.
2.2. Evaluation of Homogeneity and Stability of Antibiotic-Containing Feed Samples Using LC-MS/MS Analysis
The concentrations of all three antibiotics used exceeded the upper calibration range of the analytical method. Accordingly, the sample weight of the feed samples was adjusted, and the samples were diluted as follows: For the flour samples spiked with tiamulin, 0.5 g instead of 5 g was weighed per sample. Before solid phase extraction (SPE), a 1:20 dilution was performed. For the flour samples spiked with chloramphenicol, 0.1 g instead of 10 g was weighed per sample. We collected the residue using 2 ml instead of 0.5 ml (water/acetonitrile, 80/20, v/v), and diluted the extract 1:10 prior to the SPE measurement. For the flour samples spiked with erythromycin, 0.5 g instead of 5 g was weighed, and the extract was diluted 1:50 before SPE (0.2 ml extract + 9.8 ml McIlvaine buffer with Na-EDTA (pH 4)).
Due to these high concentrations and to establish an initial estimate of the detection limit, we reduced the amount of erythromycin in the feed by a factor of 1000 in a second feeding trial. Thus, the concentration in this case was 31.3 µg erythromycin per 100 g of flour. In this case, 1 g of the erythromycin-flour mixture was weighed for LC-MS/MS analysis.
Following Herrman et al., we assessed the homogeneity of the antibiotics in the flour by calculating the coefficient of variation (CV) of the samples (Table 1) [29].
The highest CV, and thus the lowest homogeneity, was observed in the flour spiked with chloramphenicol, with a coefficient of variation of 43.7%. The measured concentration of chloramphenicol in the flour was 119 ± 52 mg/kg. The coefficient of variation for the flour spiked with erythromycin was 19.5%, with a measured concentration of 185.8 ± 36.2 mg/kg. In the case of tiamulin and the 1:1000 diluted erythromycin, we achieved a coefficient of variation below 15%. For tiamulin, the CV was 13.3%, with a measured concentration of 325.9 ± 43.4 mg/kg in the flour. The coefficient of variation for the 1:1000 diluted erythromycin was 11.5%, with a measured concentration of 0.3 ± 0.03 mg/kg.
Furthermore, the antibiotics remained stable throughout the experiment under the given experimental conditions (Figure 1). In the control feed group, none of the administered antibiotics were detected.
2.3. Detection and Quantification of Antibiotics in Mealworms Using LC-MS/MS Analysis
We were able to detect and quantify all three antibiotics in the mealworms after 7 days of feeding and an additional 24-hour feed withdrawal period, except for the 1:1000 diluted erythromycin. In this group only traces of erythromycin below the limit of quantification were detected immediately after the 7-day feeding period. After 24 hours of feed withdrawal, it was no longer detectable. All control groups remained blank.
In the case of chloramphenicol and undiluted erythromycin, the measured concentrations exceeded the calibration curve. Therefore, a 1:100 dilution of the final extracts (10 µl extract + 990 µl water/acetonitrile) was performed for quantification. Since this dilution was still insufficient for erythromycin quantification, the already 1:100 diluted extract was further diluted 1:5.
Figure 1 presents the targeted and actual measured concentration of tiamulin, chloramphenicol, and erythromycin (undiluted) in the feed, as well as the final concentration detected in the mealworms. In the case of tiamulin the concentration detected in mealworms was 6.8 ± 0.3 mg/kg immediately after the 7-day feeding period and decreased to 0.6 ± 0.03 mg/kg after 24 hours of feed withdrawal. This corresponds to a reduction of approximately 90% after 24 hours. As previously described, chloramphenicol exhibited the highest inhomogeneity in the feed, which is evident in Figure 1 from the error indicators at Day 0. In mealworms, we detected 1.4 ± 0.2 mg/kg of chloramphenicol after the 7-day feeding period. After 24 hours of feed withdrawal, the concentration decreased to 0.2 ± 0.002 mg/kg. This means that approximately 85% of the initially detected chloramphenicol was no longer quantifiable after the fasting period. The measured concentration of erythromycin in mealworms was 224.5 ± 111 mg/kg after 7 days of feeding and decreased to 130.5 ± 0.7 mg/kg after 24 hours of feed withdrawal (Figure 1). Due to an outlier in the measurement at Day 7, the results are less clear for erythromycin. The reduction after 24 hours of fasting ranged between 10% and 57%. However, overall, the detectable amount of erythromycin in mealworms also decreased.
3. Discussion
The present study represents an initial attempt to investigate whether the analytical methods currently established in our laboratories for other matrices are suitable for detecting antibiotics in edible insects.
To generate samples that realistically reflect potential market scenarios, we designed a feeding trial using antibiotics for which (i) we have previously detected trace amounts in insects [28], (ii) documented feeding practices involving other insect species exist, such as Bombyx mori [13], or (iii) antibiotic resistance genes have been detected in commercially available edible insect products, suggesting a possible prior exposure [30]. Selection was further guided by the availability of validated detection methods in our laboratory.
The results of our trial demonstrate that the applied methods are capable of detecting tiamulin, chloramphenicol, and erythromycin in Tenebrio molitor larvae. Based on these findings, the next step will be to validate the employed methods for the insect matrix and subsequently integrate them into routine surveillance procedures.
Subsequently, we, as a state analytical institute, can expand our methodological catalogue to include additional insect-specific substances. The experimental design presented here may serve as a template for the generation of test samples. It would be advisable to include different insect species as well as various processing stages. The homogeneity of the administered antibiotics could be improved by mixing the meal in a laboratory mill after the drying phase rather than grinding it manually with a mortar.
To determine which specific substances we need to anticipate in regulatory monitoring, it is essential to examine which pharmacologically active compounds might be used in insect farming. Insight can be gained by exploring the underlying reasons and motivations for the application of such substances in insects.
Analogous to their use in pig production, these compounds could potentially be employed as growth promoters [31]. While no increase in body weight was observed, Hirose et al. reported positive effects of streptomycin on the development and lifespan of Nezara viridula [32]. All three antibiotics examined in our study tended to result in a decrease in body weight. Negative effects from the prophylactic administration of pharmacologically active compounds were also observed by Dickel et al. in Parasemia plantaginis larvae treated with fumagillin, including reduced oviposition and suppressed immune competence [33]. Webster et al. similarly reported a reduction in brood rearing in Apis mellifera following fumagillin treatment [34]. Cappellozza et al. found that the administration of chloramphenicol did not prevent the proliferation of Enterococcus mundtii and may even have promoted it by eliminating competing microorganisms [35]. Furthermore, it is possible that pharmacologically active substances reduce feed palatability, thereby indirectly counteracting weight gain. This could be investigated in analogy to Webster’s experiment, in which no effect of fumagillin on syrup intake in honeybees was observed [34]. In folivorous insects such as Locusta migratoria, the application of the leaf disc test—used by Holopainen et al. to evaluate the palatability of herbicide-treated maize for Carausius morosus—could be considered [36,37]. To assess whether and which substances might be misused as growth promoters in insect farming, these compounds would need to be examined with regard to the same mechanisms known to enhance growth in pigs and poultry. These include, for example, effects on the gut microbiota [38] or non-antibiotic, anti-inflammatory actions [39].
Another motivation for the use of pharmacologically active substances in insect farming may be their prophylactic application to prevent recurrent infections. Relevant insights into which infections are of particular concern in insect rearing are provided by studies such as those by Eilenberg et al., and Gałęcki et al. In 2015, Eilenberg et al. provided an initial overview of diseases in insect farming based on a literature review and a small-scale survey of insect producers [40]. This overview was updated in 2021 by Maciel-Vergara et al., who complemented it with an analysis of potential triggers associated with the identified diseases [41]. In addition, Gałęcki and Sokół discuss parasitic organisms relevant to insect farming, such as Nosema spp. and gregarines [42].
Beyond prophylaxis against insect diseases during rearing, reducing the microbial contamination of the final product could also be a driving factor for the use of antibiotics.
An initial risk assessment identifying human-pathogenic microorganisms potentially relevant in edible insect products was conducted by Klunder et al. They examined mealworms and crickets directly from rearing facilities and identified spore-forming bacteria as potential hazards [43]. A more comprehensive risk assessment was published by the EFSA in 2015 [15]. Additionally, Garofalo et al. provided an extensive literature review on the microbiota of insects covering the period from 2000 to 2019 [44]. As part of the present study, the generated samples were also subjected to microbiological analysis; however, no significant reduction in pathogenic microorganisms was observed in the antibiotic-treated groups compared to the control group (data not published). For this purpose, alternative decontamination methods—such as UV irradiation—may be more effective. The most recent approval of a mealworm product already includes this treatment, even though it is not directly used for microbial reduction in this case (Commission implementing regulation (EU) 2025/89).
The marked decrease in the concentrations of all three administered antibiotics in Tenebrio molitor following the 24-hour feed withdrawal period warrants further attention. This feed withdrawal has been mandated in all implementing regulations to date for the various insect products already approved under the Regulation (EU) 2015/2283 (e.g., for Tenebrio molitor larvae: Regulation (EU) 2022/169). It is essential that we are able to detect antibiotic residues even after this 24-hour withdrawal period. Since measurements of the feed over the course of the trial showed that the antibiotics remained chemically stable, the observed reduction in the insects must result from insect-internal processes. One possible explanation is that the mealworms excrete large amounts of the antibiotics unchanged. In this context, an analysis of mealworm feces would be of particular interest. Another possibility is that the antibiotics are metabolized within the insect. It is therefore necessary to determine which metabolites are formed. While the pharmacokinetics of the antibiotics used in this study have already been investigated in other livestock species [45,46,47,48,49], no corresponding studies are currently available for edible insects.
Findings from such research could serve as a scientific basis for policymakers in defining maximum residue limits (MRLs) in insects. As of now, there is a complete lack of specific legal regulations concerning the use of antibiotics in insect farming. As outlined in the introduction, no veterinary drugs are currently approved for use in the production of edible insects, and, accordingly, no MRLs have been established for insects (cf. Table 1 of Regulation (EU) No 37/2010). The use of antibiotics in edible insects is currently only described under the cascade principle, as outlined in Article 113 of Regulation (EU) 2019/6. The permissible MRLs in this context can be derived according to Implementing Regulation (EU) 2018/470, which results in applying the lowest specified MRLs to insects. For tiamulin, the MRLs are set at 100 µg/kg (target tissue: muscle in pigs, chickens, and turkeys), and for erythromycin, at 40 µg/kg (target tissue: milk). The use of chloramphenicol is entirely prohibited, as outlined in Table 2 of Regulation (EU) No 37/2010.
Conventional meat consists primarily of muscle, while insect products include the whole animal and are tested accordingly. Therefore, it is imperative to determine whether the MRLs could potentially be aligned with liver values observed in other species. Notably, in pigs, rabbits, and poultry, the highest concentrations of tiamulin are found in the liver [50]. Similarly, it can be hypothesized that the highest antibiotic residues might accumulate in the fat bodies of insects [51,52].
The prohibition of antibiotics as growth promoters in livestock farming was primarily implemented to counteract the emergence and spread of antibiotic-resistant pathogens [21]. Numerous publications have already identified resistance genes in insects and insect-derived products, suggesting that the administration of antibiotics in insects, as observed in conventional livestock, could similarly promote the selection and proliferation of antibiotic-resistant pathogens [30,53,54,55,56].
4. Materials and Methods
4.1. Antibiotics
We aimed to select antibiotics with documented associations to insects in the literature or based on our findings in routine testing, and for which a detection method exists in our laboratory. At the same time, we sought to include diverse antibiotics from different active substance groups with various spectra of activity. For our project, we opted to use pure antibiotic standards rather than complete pharmaceutical formulations. This choice was made because the standards are more readily available and free from unpredictable additives.
The antibiotics used were: Tiamulin (Sigma-Aldrich, USA), chloramphenicol (Sigma-Aldrich, USA), and erythromycin (Sigma-Aldrich, USA).
Tiamulin was previously detected in an insect product in a study conducted at our institute [28]. It is a semi-synthetic derivative of the naturally occurring antibiotic pleuromutilin and is effective against the following pathogens: Brachyspira hyodysenteriae, mycoplasma, streptococci, staphylococci, pasteurella, treponema, leptospira, and arcanobacteria [57]. Chloramphenicol has been used in silkworm breeding for some time [13,35]. Its use in edible insect production could therefore be conceivable. Chloramphenicol belongs to the amphenicol group and is effective against a broad spectrum of bacterial infections but is now used only as a reserve antibiotic due to severe side effects. Erythromycin was chosen due to its relevance in antibiotic resistance. Osimani et al. detected erythromycin resistance genes in ready-to-eat edible insects [55]. This could suggest prior exposure to the antibiotic or the widespread presence of resistance genes [58]. Both scenarios highlight the relevance of erythromycin as a target to our analysis, as its presence could further promote the proliferation of resistant bacteria within edible insects. Erythromycin is part of the macrolide antibiotic group and is used, among other applications, to treat various respiratory infections [59].
Edible insects are not explicitly mentioned in Table 1 of Regulation (EU) No 37/2010. Therefore, no veterinary medicines are approved for edible insects. However, some pharmacologically active substances are approved for all food-producing animal species (e.g., erythromycin). Prescribing veterinary drugs for insects is therefore only permissible under the cascade described in Article 113 of Regulation (EU) 2019/6 to prevent undue suffering. For antibiotics in particular, Article 107 of this regulation applies. The use of antibiotics solely for prophylaxis or growth promotion is not permitted. Commission implementing regulation (EU) 2022/1255 does not list any of the antibiotics used. Accordingly, Article 113 of Regulation 2019/6 may be applied. The antibiotics erythromycin and tiamulin used in our experiments may, therefore, be administered to insects in emergencies. Chloramphenicol, however, is prohibited under Table 2 of Regulation (EU) No 37/2010 and may not be used.
4.2. Supplementing Feed with Antibiotics
The tiamulin dose was 10 mg per 100 g of whole wheat flour (dmBio, Karlsruhe, Germany). In the absence of indications from the literature, the dose recommendation of the European Commission from Annex I of Article 35 of Directive 2001/82/EC (EMEA/V/A/137) was used as a guide. Based on Cappellozza et al., 10 mg chloramphenicol per 100 g flour was used [13]. For erythromycin, a concentration of 31.3 mg erythromycin per 100 g flour was employed. This was derived from the publication by Alippi et al. and adjusted to the weight of the mealworms determined in preliminary tests [60].
All antibiotics were initially dissolved in methanol (Merck KGaA, Germany) at a concentration of 1 mg/ml. The dissolved antibiotics were then combined with whole wheat flour in a 2:1 ratio as follows: For tiamulin and chloramphenicol we added 20 ml of the 1 mg/ml chloramphenicol resp. tiamulin solution to 10 g flour. Following the evaporation of the methanol, the flour mixtures were homogenised with a mortar and mixed with 190 g flour, which adds up to 200 g spiked flour each. For erythromycin we added 62.5 ml erythromycin solution (1 mg/ml) to 31.3 g flour. After evaporation, we added 168.7 g flour, which adds up to 200 g flour.
To ensure that the solvent methanol had no effect on the mealworms, we analyzed the control group flour (with and without methanol treatment) using headspace GC-MS (Thermo Trace 1310 GC, ThermoFisher MA, USA, detection limit: 50 mg/kg).
4.3. Feeding Experiment
Mealworms (larvae of the mealworm beetle Tenebrio molitor) were obtained from diemehlwurmfarm (Gronau, Germany) and were reared at the State Institute for Chemical Analysis and Veterinary Diagnostics Freiburg. A total of eight boxes, each measuring 15 x 11.5 x 7 cm, were used. Each box was filled with 10 g of mealworm and 10 g of whole wheat flour. Two boxes were filled with flour mixed with tiamulin, chloramphenicol, or erythromycin, respectively. Four boxes served as control groups. In two of them, the mealworms were fed only with flour, while in the other two, the flour was supplemented with methanol, which was then allowed to evaporate, mirroring the antibiotic treatment conditions. To assess the potential impact of antibiotics on the animal’s weight, we weighed the mealworms before and after seven days of antibiotic treatment. To obtain a representative average of weight development, we randomly selected and weighed 10 batches of 10 mealworms per group. All larvae were incubated at 27°C and 60–70% humidity in an incubator (Binder, Germany) for seven days. After this period, one box of each substrate type was frozen at -20°C. The remaining boxes were sieved to extract the larvae from the substrate, and the larvae were fasted in the empty boxes for another 24 h, and then frozen at -20°C. All larvae were kept at -20°C for at least 24 hours before processing to ensure all larvae were killed. The four boxes frozen after seven days were sieved to separate larvae from the substrate, as with all other boxes. The larvae were rinsed under running tap water, dried on paper towels, and homogenized in an IKA Tube Mill 100 control (IKA, Germany). Homogenates prepared from the tiamulin, chloramphenicol, erythromycin, and control groups after 0 and 24 h fasting were stored at -20 °C.
4.4. Evaluation of Homogeneity and Stability of Antibiotic-Containing Feed Samples Using LC-MS/MS Analysis
To assess the homogeneity of the antibiotic-feed mixtures, we analyzed five samples of each mixture immediately after preparation. To evaluate the stability of the antibiotics contained in the feed, we analyzed residual feed samples at the end of the feeding trial. Specifically, we examined the sieved feed residues from each experimental group, including tiamulin, chloramphenicol, and erythromycin, collected from both the 0-hour fasting group and the 24-hour fasting group. The feed analysis was conducted following the method 14.1.5 of the Association of German Agricultural Analytic and Research Institutes (Verband deutscher landwirtschaftlicher Untersuchungs- und Forschungsanstalten e.V. (VDLUFA e. V.)) [61]. The analyses were performed at the department of pharmaceutical residues of the State Institute for Chemical Analysis and Veterinary Diagnostics Karlsruhe, Germany. The only deviation from the standardized method was the use of a drying temperature of 40°C during sample concentration.
Samples were extracted using McIlvaine buffer with Na-EDTA (pH 4) and purified via SPE using Oasis HLB 6cc 200 mg cartridges (Waters, USA). The analysis was conducted using an Agilent 6460 Triple Quadrupole LC/MS system (Agilent, USA). Data evaluation was performed using the Agilent MassHunter 10 software (Agilent, USA).
4.5. Detection and Quantification of Antibiotics in Mealworms Using LC-MS/MS Analysis
The analyses were performed at the department of pharmaceutical residues at the State Institute for Chemical Analysis and Veterinary Diagnostics Karlsruhe, Germany. The method used is validated according to the Commission Implementing Regulation (EU) 2021/808 for various foodstuffs of animal origin and is used in routine testing for antibiotic residues. The adaption of this method for insects is not part of the present study. Therefore, only a brief summary is provided here (for detailed information see supplementary material S1). Homogenized and pre-weighed samples were thawed. After adding 10 mL McIlvaine buffer with Na-EDTA (pH 4), the samples were vortexed for approximately 1 min, shaken for 10 min, sonicated for 5 min, and centrifuged at 3148 g for 10 min. The supernatant was filtered through a folded filter and collected in a 50 mL centrifuge tube. The extraction was repeated with 5 mL McIlvaine buffer with Na-EDTA (pH 4), and the filter was rinsed with an additional 2 mL McIlvaine buffer with Na-EDTA (pH 4). The entire extract was then subjected to SPE. An OASIS HLB SPE cartridge (6 mL, 200 mg) was conditioned with 6 mL methanol and 6 mL deionized water. The extraction solution was loaded onto the cartridge, washed with 6 mL ultrapure water/methanol (95/5, v/v), and dried under vacuum for 10 min. Elution was performed with 6 mL methanol into a centrifuge tube. The eluate was evaporated to dryness under a nitrogen stream at 40°C. The dry residue was reconstituted in 1 mL ultrapure water/acetonitrile (90/10, v/v) using vortex mixing and ultrasonication. After centrifugation at approximately 15000 rpm, the solution was transferred to an HPLC vial for LC-MS analysis or stored frozen until measurement. The identification of target analytes in the samples was based on mass spectrometric data, specifically the detection of at least two MS2 fragment ions, and retention time comparison with corresponding standards. Each sample was initially processed and analyzed once. If a target analyte was detected, its concentration was determined using a matrix calibration curve established with control samples. In cases of critical initial findings, multiple additional sample preparations were performed and analyzed. To generate the matrix calibration curve, blank samples (e.g., five replicates) were fortified with different concentrations of the analyte, processed, and analyzed. The final result was obtained by averaging the individual measurements.
5. Conclusions
In summary, the findings of this study highlight the feasibility of detecting antibiotics in edible insects using established analytical methods. The experimental design provides a valuable foundation for future feeding trials, which will be necessary to expand the methodological repertoire to include insect-relevant substances, as well as to analyze the pharmacokinetics and pharmacodynamics in insects.
Such efforts will contribute to a robust scientific basis for developing maximum residue limits, establishing appropriate detection methods, and safeguarding public health in the context of edible insect production.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementing material S1: Test procedure for the determination of antibiotics in animal-origin foods (excluding honey) using LC-MS/MS; Supplementing material S2: Weight development of Tenebrio molitor larvae after antibiotic administration.
Author Contributions
Conceptualization, T.A. and C.W.; methodology, T.A.; Formal analysis, T.A.; Investigation, T.A.; writing—original draft preparation, T.A..; writing—review and editing, C.W. and A.V.; supervision, C.W. and A.V.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank Gudrun Kopf, Franziska Scharmann and Uta Kraus from the State Institute for Chemical Analysis and Veterinary Diagnostics Karlsruhe Weißenburger Str. 3, 76187 Karlsruhe, Germany for analyzing the samples, providing valuable advice throughout the study and for reviewing this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| EFSA | European Food Safety Authority |
| SPE | Solid Phase Extraction |
| CV | Coefficient of Variation |
| MRL | Maximum Residue Limit |
| VDLUFA | Association of German Agricultural Analytic and Research Institutes (Verband deutscher landwirtschaftlicher Untersuchungs- und Forschungsanstalten e.V.) |
References
- Angerer, M.; Dubuni, S.; Ellrott, T.; Gebhard, M.; Gebhardt, B.; Hacker, A.; Hirschfelder, G.; Gabriele, K.; Krüger, J.; Kussmann, M.; et al. Wie is(s)t Deutschland 2030?, Frankfurt am Main, 2015.
- Schiel, L.; Wind, C.; Braun, P.G.; Koethe, M. Legal framework for the marketing of food insects in the European Union. Ernahrungs Umschau 2020, 67, 76–85. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Peni, D. Growth Potential of Yellow Mealworm Reared on Industrial Residues. Agriculture 2020, 10, 599. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Harahap, I.A.; Osei-Owusu, J.; Saikia, T.; Wu, Y.S.; Fernando, I.; Perestrelo, R.; Câmara, J.S. Bioconversion of organic waste by insects – A comprehensive review. Process Safety and Environmental Protection 2024, 187, 1–25. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Itterbeeck, J.; Heetkamp, M.J.W.; van den Brand, H.; van Loon, J.J.A.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS One 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Correction: Feed Conversion, Survival and Development, and Composition of Four Insect Species on Diets Composed of Food By-Products. PLoS One 2019, 14, e0222043. [Google Scholar] [CrossRef]
- Caparros Megido, R.; Francis, F.; Haubruge, E.; Le Gall, P.; Tomberlin, J.K.; Miranda, C.D.; Jordan, H.R.; Picard, C.J.; Pino, M.; Ramos-Elordy, J.; et al. A worldwide overview of the status and prospects of edible insect production. entomologia 2024, 44, 3–27. [Google Scholar] [CrossRef]
- van Huis, A. Edible insects: Challenges and prospects. Entomological Research 2022, 52, 161–177. [Google Scholar] [CrossRef]
- van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Tettey, E.; Yunusa, B.M.; Ngah, N.; Debrah, S.K.; Yang, X.; Fernando, I.; Povetkin, S.N.; Shah, M.A. Legal situation and consumer acceptance of insects being eaten as human food in different nations across the world-A comprehensive review. Comprehensive Reviews in Food Science and Food Safety 2023, 22, 4786–4830. [Google Scholar] [CrossRef]
- Dippel, F.-W. Insekten: Das Fleisch der Zukunft? Available online: https://www.doccheck.com/de/detail/articles/42719-insekten-das-fleisch-der-zukunft (accessed on 19 May 2025).
- Favaro, R.; Lupi, D.; Jucker, C.; Cappellozza, S.; Faccoli, M. An artificial diet for rearing three exotic longhorn beetles invasive to Europe. Bulletin of Insectology 2017, 70, 91–99. [Google Scholar]
- Cappellozza, L.; Cappellozza, S.; Saviane, A.; Sbrenna, G. Artificial diet rearing system for the silkworm Bombyx mori (Lepidoptera: Bombycidae): Effect of vitamin C deprivation on larval growth and cocoon production. Appl. Entomol. Zool. 2005, 40, 405–412. [Google Scholar] [CrossRef]
- Marzoli, F.; Tata, A.; Massaro, A.; Bragolusi, M.; Passabì, E.; Cappellozza, S.; Saviane, A.; Tassoni, L.; Piro, R.; Belluco, S. Microbiological and chemical safety of Bombyx mori farmed in north-eastern Italy as a novel food source. JIFF 2023, 9, 1047–1062. [Google Scholar] [CrossRef]
- EFSA Scientific Committee. Risk profile related to production and consumption of insects as food and feed. EFSA Journal 2015, 13, 4257. [Google Scholar] [CrossRef]
- Hoek-van den Hil, E.F.; van de Schans, M.; Bor, G.; van der Fels-Klerx, H.J. Effects of veterinary drugs on rearing and safety of black soldier fly (Hermetia illucens) larvae. Journal of Insects as Food and Feed 2022, 1–10. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Li, Z.; Guo, S.; Li, K.; Xu, P.; Ok, Y.S.; Jones, D.L.; Zou, J. Antibiotics and antibiotic resistance genes in agricultural soils: A systematic analysis. Critical Reviews in Environmental Science and Technology 2023, 53, 847–864. [Google Scholar] [CrossRef]
- Marti, R.; Scott, A.; Tien, Y.-C.; Murray, R.; Sabourin, L.; Zhang, Y.; Topp, E. Impact of manure fertilization on the abundance of antibiotic-resistant bacteria and frequency of detection of antibiotic resistance genes in soil and on vegetables at harvest. Appl. Environ. Microbiol. 2013, 79, 5701–5709. [Google Scholar] [CrossRef]
- Amorim, H.C.S.; Ashworth, A.J.; Arsi, K.; Rojas, M.G.; Morales-Ramos, J.A.; Donoghue, A.; Robinson, K. Insect frass composition and potential use as an organic fertilizer in circular economies. Journal of Economic Entomology 2024, 117, 1261–1268. [Google Scholar] [CrossRef]
- Asendorf, T.; Wind, C.; Rullmann, A.; Vilcinskas, A. Comparison of DNA-based methods for the detection of meat feeding in Alphitobius diaperinus larvae. JIFF 2025, 1–12. [Google Scholar] [CrossRef]
- Tang, K.L.; Caffrey, N.P.; Nóbrega, D.B.; Cork, S.C.; Ronksley, P.E.; Barkema, H.W.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.D.; et al. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: a systematic review and meta-analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef]
- Grispoldi, L.; Karama, M.; El-Ashram, S.; Saraiva, C.M.; García-Díez, J.; Chalias, A.; Barbera, S.; Cenci-Goga, B.T. Hygienic Characteristics and Detection of Antibiotic Resistance Genes in Crickets (Acheta domesticus) Breed for Flour Production. Microbiology Research 2021, 12, 503–512. [Google Scholar] [CrossRef]
- Milanović, V.; Osimani, A.; Pasquini, M.; Aquilanti, L.; Garofalo, C.; Taccari, M.; Cardinali, F.; Riolo, P.; Clementi, F. Getting insight into the prevalence of antibiotic resistance genes in specimens of marketed edible insects. Int. J. Food Microbiol. 2016, 227, 22–28. [Google Scholar] [CrossRef]
- Osimani, A.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Ruschioni, S.; Riolo, P.; Isidoro, N.; Loreto, N.; Galarini, R.; et al. Distribution of Transferable Antibiotic Resistance Genes in Laboratory-Reared Edible Mealworms (Tenebrio molitor L.). Front. Microbiol. 2018, 9, 2702. [Google Scholar] [CrossRef]
- Osimani, A.; Garofalo, C.; Milanović, V.; Taccari, M.; Cardinali, F.; Aquilanti, L.; Pasquini, M.; Mozzon, M.; Raffaelli, N.; Ruschioni, S.; et al. Insight into the proximate composition and microbial diversity of edible insects marketed in the European Union. Eur Food Res Technol 2017, 243, 1157–1171. [Google Scholar] [CrossRef]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilbäck, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [PubMed]
- Schemmel, D.; Rukavina, M. BVL-Report 19.4: Berichte zur Lebensmittelsicherheit 2023. 2025. Available online: www.bvl.bund.de (accessed on 19 May 2025).
- Hegmanns, M.; Kopf, G.; Schiel, L.; Wind, C.; Krüger, S.; Pietsch, K.; Müller, M.; Schill, S. Ergebnisse der Untersuchungen aus dem Insekten Internetprojekt, Freiburg, 2021. Available online: https://www.ua-bw.de/pub/beitrag.asp?subid=3&Thema_ID=2&ID=3406&Pdf=No&lang=DE (accessed on 19 March 2025).
- Herrman, T.; Behnke, K.C.; Loughin, T. Mixing and clean-out properties of sulfamethazine and carbadox in swine feed (1995). Kansas Agricultural Experiment Station Research Reports 1995, 10, 120–124. [Google Scholar] [CrossRef]
- Osimani, A.; Cardinali, F.; Aquilanti, L.; Garofalo, C.; Roncolini, A.; Milanović, V.; Pasquini, M.; Tavoletti, S.; Clementi, F. Occurrence of transferable antibiotic resistances in commercialized ready-to-eat mealworms (Tenebrio molitor L.). Int. J. Food Microbiol. 2017, 263, 38–46. [Google Scholar] [CrossRef]
- Hassan, Y.I.; Lahaye, L.; Gong, M.M.; Peng, J.; Gong, J.; Liu, S.; Gay, C.G.; Yang, C. Innovative drugs, chemicals, and enzymes within the animal production chain. Vet. Res. 2018, 49, 71. [Google Scholar] [CrossRef]
- Hirose, E.; Panizzi, A.R.; Cattelan, A.J. Potential Use of Antibiotic to Improve Performance of Laboratory-Reared Nezara viridula (L.) (Heteroptera: Pentatomidae). Neotropical Entomology 2006, 35, 279–281. [Google Scholar] [CrossRef]
- Dickel, F.; Freitak, D.; Mappes, J. Long-Term Prophylactic Antibiotic Treatment: Effects on Survival, Immunocompetence and Reproduction Success of Parasemia plantaginis (Lepidoptera: Erebidae). J. Insect Sci. 2016, 16. [Google Scholar] [CrossRef]
- Webster, T. Fumagillin Affects Nosema apis and Honey Bees (Hymonopterai Apidae). Journal of Economic Entomology 1994, 87, 601–604. [Google Scholar] [CrossRef]
- Cappellozza, S.; Saviane, A.; Tettamanti, G.; Squadrin, M.; Vendramin, E.; Paolucci, P.; Franzetti, E.; Squartini, A. Identification of Enterococcus mundtii as a pathogenic agent involved in the “flacherie” disease in Bombyx mori L. larvae reared on artificial diet. J. Invertebr. Pathol. 2011, 106, 386–393. [Google Scholar] [CrossRef]
- Phillips, W.M. Modification of feeding ‘preference’ in the flea-beetle, Halitica lythri (Coleoptera, Chrysomelidae). Entomologia Exp Applicata 1977, 21, 71–80. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Kainulainen, E.; Hänninen, O. Palatability of herbicide-treated maize to the Indian stick insect (Carausius morosus). Agriculture, Ecosystems & Environment 1991, 36, 191–197. [Google Scholar] [CrossRef]
- Gaskins, H.R.; Collier, C.T.; Anderson, D.B. Antibiotics as growth promotants: mode of action. Anim. Biotechnol. 2002, 13, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Niewold, T.A. The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poult. Sci. 2007, 86, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Eilenberg, J.; Vlak, J.M.; Nielsen-LeRoux, C.; Cappellozza, S.; Jensen, A.B. Diseases in insects produced for food and feed. Journal of Insects as Food and Feed 2015, 1, 87–102. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Jensen, A.B.; Lecocq, A.; Eilenberg, J. Diseases in edible insect rearing systems. Journal of Insects as Food and Feed 2021, 7, 621–638. [Google Scholar] [CrossRef]
- Gałęcki, R.; Sokół, R. A parasitological evaluation of edible insects and their role in the transmission of parasitic diseases to humans and animals. PLoS One 2019, 14, e0219303. [Google Scholar] [CrossRef]
- Klunder, H.C.; Wolkers-Rooijackers, J.; Korpela, J.M.; Nout, M. Microbiological aspects of processing and storage of edible insects. Food Control 2012, 26, 628–631. [Google Scholar] [CrossRef]
- Garofalo, C.; Milanović, V.; Cardinali, F.; Aquilanti, L.; Clementi, F.; Osimani, A. Current knowledge on the microbiota of edible insects intended for human consumption: A state-of-the-art review. Food Res. Int. 2019, 125, 108527. [Google Scholar] [CrossRef]
- Ambros, L.; Montoya, L.; Kreil, V.; Waxman, S.; Albarellos, G.; Rebuelto, M.; Hallu, R.; San Andres, M.I. Pharmacokinetics of erythromycin in nonlactating and lactating goats after intravenous and intramuscular administration. J. Vet. Pharmacol. Ther. 2007, 30, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Switała, M.; Hrynyk, R.; Smutkiewicz, A.; Jaworski, K.; Pawlowski, P.; Okoniewski, P.; Grabowski, T.; Debowy, J. Pharmacokinetics of florfenicol, thiamphenicol, and chloramphenicol in turkeys. J. Vet. Pharmacol. Ther. 2007, 30, 145–150. [Google Scholar] [CrossRef]
- Elazab, S.T.; Elshater, N.S.; Hashem, Y.H.; Park, S.-C.; Hsu, W.H. Tissue Residues and Pharmacokinetic/Pharmacodynamic Modeling of Tiamulin Against Mycoplasma anatis in Ducks. Front. Vet. Sci. 2020, 7, 603950. [Google Scholar] [CrossRef] [PubMed]
- Laber, G.; Schütze, E. Blood level studies in chickens, turkey poults and swine with tiamulin, a new antibiotic. J. Antibiot. (Tokyo) 1977, 30, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Sartini, I.; Vercelli, C.; Lebkowska-Wieruszewska, B.; Lisowski, A.; Fadel, C.; Poapolathep, A.; Dessì, F.; Giorgi, M. Pharmacokinetics and antibacterial activity of tiamulin after single and multiple oral administrations in geese. Vet. Anim. Sci. 2023, 22, 100317. [Google Scholar] [CrossRef]
- Ciucă, V.C.; Rusănescu, C.O.; Safta, V.V. Analysis of Transfer of Tiamulin to Animal Tissue after Oral Administration: An Important Factor for Ensuring Food Safety and Environmental Protection. Pharmaceuticals (Basel) 2023, 16. [Google Scholar] [CrossRef]
- Gutierrez, E.; Wiggins, D.; Fielding, B.; Gould, A.P. Specialized hepatocyte-like cells regulate Drosophila lipid metabolism. Nature 2007, 445, 275–280. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Larson, Z.; Subramanyam, B.; Zurek, L.; Herrman, T. Diversity and antibiotic resistance of enterococci associated with stored-product insects collected from feed mills. Journal of Stored Products Research 2008, 44, 198–203. [Google Scholar] [CrossRef]
- Rawat, N.; Anjali, *!!! REPLACE !!!*; Shreyata, *!!! REPLACE !!!*; Sabu, B.; Jamwal, R.; Devi, P.P.; Yadav, K.; Raina, H.S.; Rajagopal, R. Understanding the role of insects in the acquisition and transmission of antibiotic resistance. Sci. Total Environ. 2023, 858, 159805. [Google Scholar] [CrossRef]
- Milanović, V.; Osimani, A.; Roncolini, A.; Garofalo, C.; Aquilanti, L.; Pasquini, M.; Tavoletti, S.; Vignaroli, C.; Canonico, L.; Ciani, M.; et al. Investigation of the Dominant Microbiota in Ready-to-Eat Grasshoppers and Mealworms and Quantification of Carbapenem Resistance Genes by qPCR. Front. Microbiol. 2018, 9, 3036. [Google Scholar] [CrossRef]
- Cifuentes, Y.; Glaeser, S.P.; Mvie, J.; Bartz, J.-O.; Müller, A.; Gutzeit, H.O.; Vilcinskas, A.; Kämpfer, P. The gut and feed residue microbiota changing during the rearing of Hermetia illucens larvae. Antonie Van Leeuwenhoek 2020, 113, 1323–1344. [Google Scholar] [CrossRef] [PubMed]
- Löscher, W.; Richter, A.; Potschka, H. Pharmakotherapie bei Haus- und Nutztieren; Georg Thieme Verlag: Stuttgart, 2014; ISBN 9783830412502. [Google Scholar]
- Zalewska, M.; Błażejewska, A.; Czapko, A.; Popowska, M. Antibiotics and Antibiotic Resistance Genes in Animal Manure - Consequences of Its Application in Agriculture. Front. Microbiol. 2021, 12, 610656. [Google Scholar] [CrossRef]
- Farzam, K.; Nessel, T.; Quick, J. Erythromycin. Available online: https://www.ncbi.nlm.nih.gov/books/NBK532249/ (accessed on 4 November 2024).
- Alippi, A.M.; Albo, G.N.; Leniz, D.; Rivera, I.; Zanelli, M.L.; Roca, A.E. Comparative study of tylosin, erythromycin and oxytetracycline to control American foulbrood of honey bees. Journal of Apicultural Research 1999, 38, 149–158. [Google Scholar] [CrossRef]
- Methode 14.1.5, Bestimmung ausgewählter Antibiotika in Futtermitteln mittels LC-MSMS.; VERBAND DEUTSCHER LANDWIRTSCHAFTLICHER UNTERSUCHUNGS- UND FORSCHUNGSANSTALTEN, Ed.; VDLUFA-Verlag: Darmstadt, 2018. Darmstadt.
Figure 1.
Concentration of tiamulin, chloramphenicol, and erythromycin (undiluted) measured in feed (left y-axis, blue) and in mealworms (right y-axis, red). Error bars indicate standard deviation (SD). The blue, dashed horizontal line represent the target feed concentration (left y-axis).
Figure 1.
Concentration of tiamulin, chloramphenicol, and erythromycin (undiluted) measured in feed (left y-axis, blue) and in mealworms (right y-axis, red). Error bars indicate standard deviation (SD). The blue, dashed horizontal line represent the target feed concentration (left y-axis).
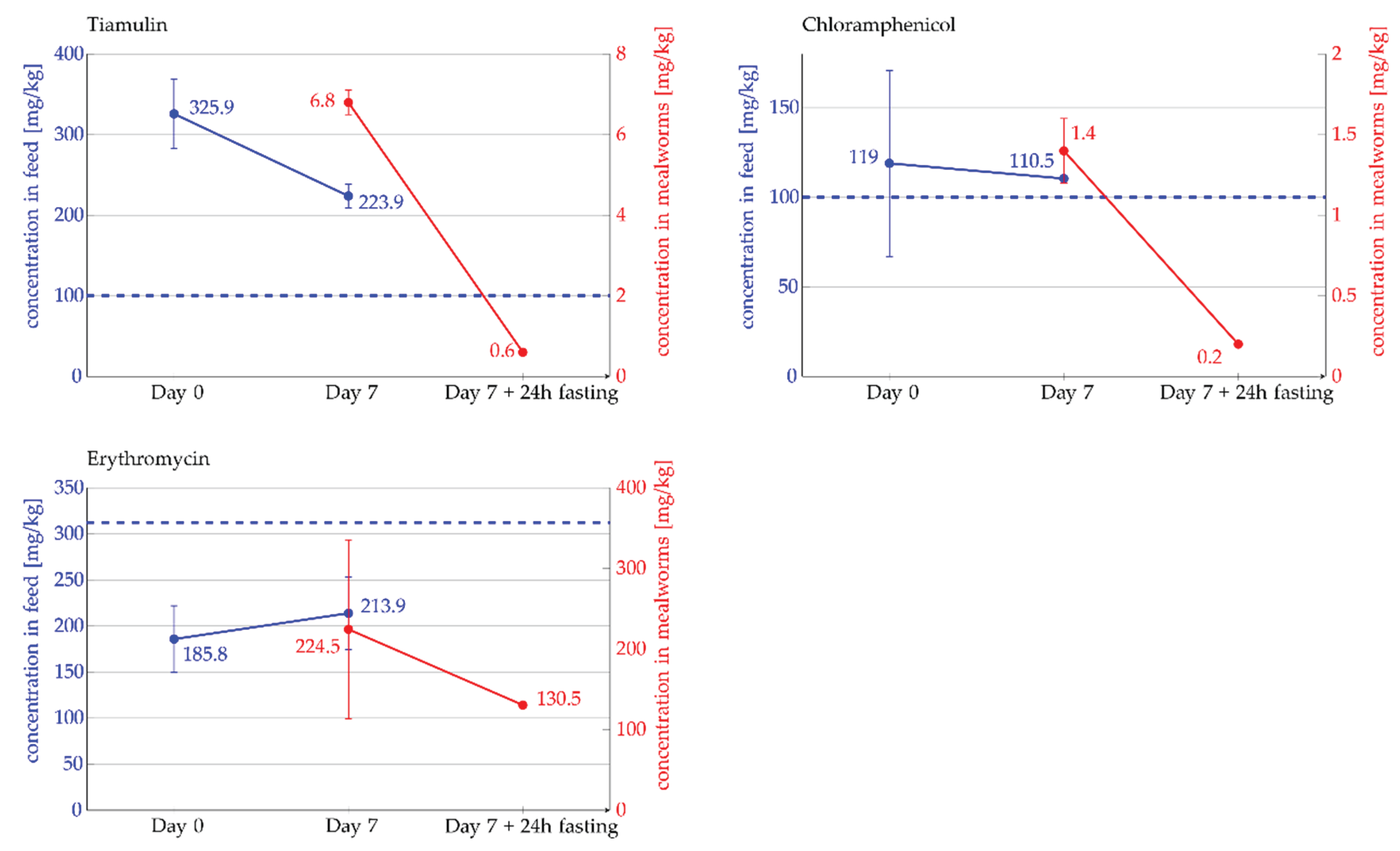

Table 1.
Homogenity of antibiotics in feed.
| Tiamulin | Chloramphenicol | Erythromycin | Erythromycin/1000 | |
|---|---|---|---|---|
| average X̅ [mg/kg] | 325.9 | 119 | 185.8 | 0.3 |
| SD [mg/kg] | 43.4 | 52 | 36.2 | 0.03 |
| coefficient of variation (CV) [%] | 13.3 | 43.7 | 19.5 | 11.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



