Submitted:
12 August 2025
Posted:
13 August 2025
You are already at the latest version
Abstract
Background:
The human gut microbiome—a diverse ecosystem of trillions of microorganisms—plays a critical role in metabolic, immune, and neurological regulation. However, modern lifestyle factors such as antibiotic overuse, cesarean delivery, reduced breastfeeding, processed and high-sodium diets, alcohol intake, smoking, and exposure to environmental toxins (e.g., glyphosate) have significantly reduced microbial diversity. Loss of keystone species like Bifidobacterium infantis and Lactobacillus reuteri contributes to gut dysbiosis, which has been implicated in chronic metabolic, autoimmune, cardiovascular, and neurodegenerative conditions.
Materials & Methods:
This Qualitative Systematic Review (QualSR) synthesized data from over 547 studies involving human participants and standardized microbiome analysis techniques, including 16S rRNA sequencing and metagenomics. Studies were reviewed for microbial composition, immune and metabolic biomarkers, and clinical outcomes related to microbiome restoration strategies.
Results:
Findings reveal a 40–60% reduction in microbial diversity among Western populations compared to traditional societies, particularly affecting SCFA-producing bacteria. Supplementation with B. infantis was associated with a significant reduction in systemic inflammation—including a 50% decrease in C-reactive protein (CRP) and reduced TNF-α levels—alongside increases in regulatory T cells and anti-inflammatory cytokines (IL-10, TGF-β1). L. reuteri demonstrated additional immunomodulatory effects and neurobehavioral benefits in preclinical models. Restoration interventions led to a 30–45% increase in tight junction protein expression and a 25–50% reduction in overall inflammation, suggesting improved gut barrier function and systemic immune balance.
Conclusion:
Restoring gut microbiota diversity through keystone species like B. infantis and L. reuteri holds therapeutic potential for a wide range of chronic inflammatory, metabolic, and neuroimmune disorders. Strengthening gut barrier integrity and reducing systemic inflammation by up to 50% underscores the microbiome’s relevance in immunometabolic health. Future research should emphasize personalized microbiome profiling, long-term outcomes, and transgenerational effects of early-life microbial disruption.
Keywords:
gut microbiome
; microbial diversity
; Bifidobacterium infantis
; Lactobacillus reuteri
; gut-brain axis
; short-chain fatty acids
Introduction
The human gastrointestinal tract harbors a highly diverse and dynamic microbial ecosystem comprising approximately 100 trillion microorganisms spanning over 1,000 species [1,58]. These microbes engage in symbiotic interactions with the host, supporting essential physiological processes such as nutrient absorption, energy harvest, maintenance of epithelial barrier function, and immune modulation [2,58]. The concept of the microbiome as a "second genome" reflects its vast metabolic capacity and integral role in human health. However, modern environmental and behavioral exposures—such as Westernized diets, widespread antibiotic use, increased rates of cesarean delivery, and reduced breastfeeding—have led to significant perturbations in microbial composition and function [3,4,16,17,18]. This phenomenon of dysbiosis is typically characterized by decreased microbial diversity, loss of beneficial species, and enrichment of opportunistic or proinflammatory taxa [4,59].
Metabolic disorders (e.g., obesity, type 2 diabetes), cardiovascular diseases (e.g., hypertension, atherosclerosis stenosis), autoimmune diseases (e.g., rheumatoid arthritis, inflammatory bowel disease, multiple sclerosis), allergic conditions (e.g., asthma, atopic dermatitis), and neuropsychiatric conditions (e.g., depression, autism spectrum disorders) are rising global health challenges with substantial personal, societal, and economic burdens. These chronic, non-communicable diseases (NCDs) have been strongly linked with dietary and lifestyle patterns that promote persistent low-grade inflammation and disruption of the gut microbial ecosystem—also known as microbial dysbiosis [2,3,57]. A growing body of evidence supports the role of the gut–brain–microbiota axis, a complex bidirectional communication network between the central nervous system and the gut microbiome, in regulating host metabolic, immune, and neurobehavioral functions [52,53,57]. This paradigm shift has placed the gut microbiota at the center of public health discussions regarding the prevention and management of chronic disease.
Notably, industrialized societies demonstrate marked shifts in gut microbiome structure compared to traditional, non-industrialized populations [5,6]. Traditional microbiomes are typically enriched in microbial taxa that support fiber fermentation, short-chain fatty acid (SCFA) production, and oxidative stress resistance—functions that are often impaired or absent in Western populations [7,8,9,10]. SCFAs such as acetate, propionate, and butyrate are microbial fermentation products that serve as signaling molecules, energy substrates for colonocytes, and modulators of immune and metabolic pathways [13,14]. Their biosynthesis is dependent on complex microbial networks: acetate production involves the Wood–Ljungdahl pathway; propionate is primarily synthesized by Bacteroidetes via the succinate pathway; and butyrate is generated predominantly by Firmicutes via the butyryl-CoA:acetate CoA-transferase route [11,12]. The disruption of these pathways contributes to intestinal permeability, systemic inflammation, insulin resistance, and altered neurobehavioral signaling—all hallmarks of metabolic and mental health disorders [13,14,59].
Nutrient cycling processes provide compelling evidence for the disproportionate ecological impact of rare microbes. The keystone species model, a fundamental theoretical framework in microbial ecology, demonstrates how certain microbial taxa can exert outsized influence on ecosystem function despite their low abundance. These keystone microorganisms perform essential metabolic processes that fundamentally shape the composition, stability, and overall metabolic capacity of entire microbial communities. A prime example of this phenomenon is observed in freshwater environments, where rare green and purple sulfur bacteria function as highly active keystone species, driving critical nitrogen and carbon cycling processes that far exceed what their minimal abundance would suggest. [15,60]. Loss of these keystone species can cause cascading functional breakdowns within the microbiome, leading to a loss of homeostasis and emergence of disease phenotypes [15,61]. Keystone taxa are particularly vulnerable to the cumulative impact of modern lifestyle factors, including antibiotic exposure, nutrient-poor diets, cesarean delivery, formula feeding, and chronic psychosocial stress [16,17,18,42]. Moreover, longitudinal data suggest that early-life disruptions to microbial colonization may exert long-term effects on immune programming, metabolism, and neural development, with implications for chronic disease risk across the lifespan [41,42].
Among candidate keystone microbes, Bifidobacterium longum subsp. infantis (B. infantis) and Lactobacillus reuteri (L. reuteri) have emerged as two of the most functionally and clinically promising species. Both are considered hallmark genera of a healthy gut ecosystem, particularly in infancy, and are commonly reduced or absent in dysbiotic microbiomes [19,20,21,62]. B. infantis possesses a specialized genome that enables it to metabolize human milk oligosaccharides (HMOs), as complex carbohydrates abundant in breast milk—thus facilitating early gut colonization, immune maturation, and SCFA production [54,63]. By generating acetate and propionate, B. infantis contributes to the reinforcement of the intestinal barrier, modulation of T-regulatory cell populations, and suppression of proinflammatory pathways [54,55]. In effect, these keystone microbiomes functions are essential for immune tolerance and protection against allergic and autoimmune conditions.
Likewise, L. reuteri exhibits broad-spectrum effects on host physiology. It produces bioactive metabolites such as reuterin (with antimicrobial properties), histamine (which influences immune responses), and SCFAs that support mucosal integrity [20,64]. Notably, L. reuteri can influence central nervous system signaling through the vagus–oxytocin pathway, and animal models have shown that its supplementation improves social behavior and stress response [56,65,135]. These findings position L. reuteri as a potential microbial target for modulating the gut–brain axis and treating neurodevelopmental conditions such as autism spectrum disorder.
The restoration of these keystone taxa has emerged as a therapeutic strategy to counteract the downstream effects of dysbiosis. These targeted interventions—ranging from probiotics and prebiotics to next-generation live bio-therapeutics and are designed to restore lost microbial functions or reintroduce critical metabolites [ 19-20, 62]. Beyond microbial repletion, public health approaches must address the upstream determinants of microbiome disruption, such as maternal diet, birth practices, antibiotic stewardship, and environmental toxicant exposure [26,27,28,29,30,36,37,38]. Glyphosate, emulsifiers, and artificial sweeteners, for example, have all demonstrated deleterious effects on beneficial gut microbes and should be considered in regulatory and clinical frameworks [37,38,39,40].
The complexity of microbiome host interactions and the diverse determinants of dysbiosis underscore the need for integrative research approaches. Recent advances in multi-omic technologies—including metagenomics, metabolomics, and metaproteomics had provided deep insights into how environmental exposures, host genetics, and microbial composition interact to shape health outcomes [22,23,24,25]. These tools have revealed distinct microbiome signatures in patients with metabolic, immunological, and neuropsychiatric diseases, offering the possibility of microbial biomarkers for early detection and personalized interventions [24,25]. Moreover, host genetic factors significantly shape microbiome composition and immune tolerance. The gene polymorphisms, affecting mucin production, Toll-like receptor signaling, and immune regulation influence how the host accommodates or resists microbial colonization [44,45,46,47,48]. Thus, interventions targeting the microbiome must be contextualized within a framework of host–microbe coadaptation and individual variability.
The depletion of critical microbial keystone species such as B. infantis and L. reuteri represents both a marker of microbiome erosion and a modifiable contributor to the rising burden of chronic disease. These species provide key ecological functions—ranging from SCFA production to neuroimmune modulation—that are essential for maintaining host homeostasis. By identifying the drivers of their loss and exploring mechanisms for their restoration, we open new avenues for microbiome-informed strategies to prevent and manage NCDs. Framing gut microbial health as a public health priority is essential to reversing the intergenerational cascade of dysbiosis and chronic disease.
Within medications or pharmacologic exposures, proton pump inhibitors (PPIs) warrant special attention for their impact on gut microbiome health. By reducing gastric acid, PPIs impair protein digestion and nutrient absorption while compromising the stomach’s first-line defense against ingested pathogens [66]. This hypochlorhydric state promotes the overgrowth of opportunistic pathogens such as Candida, Helicobacter pylori, and intestinal parasites [67]. Beyond local gastric effects, reduced acidity alters downstream microbial signals, contributing to gut dysbiosis. Unlike antihypertensives or antidepressants, which exert indirect or modulatory effects [68], PPIs directly disrupt foundational digestive and microbial processes, posing a significant barrier to effective microbiome restoration and host healing.
Despite increasing recognition of the importance of B. infantis and L. reuteri, critical questions remain about their ecological roles, clinical relevance, and translational potential in the context of global health. This systematic review aims to synthesize current evidence from preclinical, clinical, and epidemiological studies to assess the following:
- (i)
- What ecological roles do B. infantis and L. reuteri play across the human lifespan?
- (ii)
- What factors contribute to their depletion in industrialized societies?
- (iii)
- What are the functional and clinical consequences of their loss, particularly regarding immune, metabolic, and neurobehavioral outcomes?
- (iv)
- What is the current evidence base for restoring these keystone species, and how might this inform public health, clinical nutrition, and microbiome-targeted interventions?
By clarifying the translational significance of B. infantis and L. reuteri as foundational members of the human gut ecosystem, this review provides a conceptual and practical framework for microbiome-based strategies aimed at restoring host–microbiota symbiosis and mitigating the risk of chronic disease across populations. Figure 3 presents a conceptual framework summarizing this progression—from environmental disruptors to keystone depletion, system-wide dysfunction, and restoration pathways targeting B. infantis and L. reuteri.
Materials & Method
Study Design
This Qualitative Systematic Review (QualSR) was conducted to evaluates the ecological, mechanistic, and therapeutic roles of two keystone microbial taxa—Bifidobacterium infantis and Lactobacillus reuteri—in shaping host metabolic health and gut-brain axis function. The review explored the clinical consequences of their depletion and assessed strategies for microbiome restoration across both human and animal models.
The study adhered to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA 2020) guidelines. It was informed by an ecological health framework linking microbial dysbiosis to chronic disease etiology.
The primary aims were to:
- Define the ecological functions of B. infantis and L. reuteri across developmental stages.
- Identify key drivers of their loss in industrialized societies.
- Examine the clinical and mechanistic consequences of their depletion in metabolic, immunologic, and neurobehavioral outcomes.
- Synthesize evidence on therapeutic strategies—including probiotics, prebiotics, and biotherapeutics—targeting their restoration.
Search Strategy
A comprehensive literature search was conducted across five electronic databases: PubMed/MEDLINE, Embase, Web of Science, Cochrane Library, and CINAHL, covering publications from January 2000 to February 2024. Additional sources included reference mining of key reviews and hand-searching relevant journals.
The search strategy incorporated Medical Subject Headings (MeSH) and free-text terms such as:
- "gut microbiome", "microbial diversity", "dysbiosis"
- "Bifidobacterium infantis", "Lactobacillus reuteri", "keystone species"
- "short-chain fatty acids", "intestinal permeability", "SCFA production"
- "gut-brain axis", "microbiome restoration", "probiotic", "synbiotic", "prebiotic", "immune modulation"
- "autism spectrum disorder", "depression", "metabolic syndrome", "host genetics"
Search strings were constructed using Boolean operators (AND, OR) to balance sensitivity and specificity. An example from PubMed:
("gut microbiome" OR "intestinal microbiota") AND ("Bifidobacterium infantis" OR "Lactobacillus reuteri") AND ("dysbiosis" OR "microbiota disruption") AND ("probiotics" OR "restoration" OR "short-chain fatty acids") AND ("immune modulation" OR "metabolic health" OR "gut-brain axis")
A total of 547 records were identified. After de-duplication, 394 unique records were retained for screening.
QualES Eligibility Criteria
Inclusion Criteria:
- Peer-reviewed human or animal studies
- Published between 2000 and 2024
- Investigated either B. infantis or L. reuteri as a central taxon
-
Explored outcomes in at least one of the following domains:
- o
- Microbial function (e.g., SCFA production, mucosal integrity)
- o
- Immune and metabolic regulation (e.g., insulin sensitivity, cytokine profiles)
- o
- Neurobehavioral modulation (e.g., vagal tone, anxiety, ASD-like behavior)
- o
- Restoration strategies (e.g., probiotic/prebiotic administration, synbiotics, FMT)
- Employed recognized microbiome assessment methods (e.g., 16S rRNA sequencing, metagenomics, metabolomics)
Exclusion Criteria:
- In vitro studies with no host interaction
- Studies with unclear endpoints or inadequate data
- Sample size <30 for human studies or <10 per group in animal models
- Non-English language, case reports, narrative reviews, and conference abstracts
Sample size justification: Thresholds were chosen to enhance generalizability and reduce the risk of type II errors in heterogeneous populations. Smaller studies often yield less robust or clinically meaningful estimates.
Study Selection Process
A two-stage screening process was applied:
- Title and abstract screening of 394 articles by two independent reviewers
- Full-text review of 97 articles
A total of 56 studies were included in the final synthesis:
- 36 human studies
- 20 animal model studies
Reasons for exclusion at the full-text stage (n=41) included: non-specific interventions (n=11), unclear or absent outcomes (n=11), insufficient sample size (n=18), and inaccessible full texts (n=1).
A QualES PRISMA 2020 flow diagram is provided in Figure 1.
Data Extraction and Quality Assessment
Data were independently extracted by two reviewers using a standardized form, capturing:
- Study type and design
- Species/population characteristics
- Intervention details (strain, dose, duration)
- Analytical techniques (e.g., LC-MS, 16S rRNA, behavioral assays)
- Outcomes: metabolic, immune, neurocognitive, microbiome
- Drivers of microbial loss (e.g., antibiotics, birth mode, infant diet)
- Type of restoration strategy (e.g., probiotic-only, diet, combination)
Risk of bias was assessed as follows:
- RCTs: Cochrane Risk of Bias Tool 2.0
- Observational studies: Newcastle-Ottawa Scale
- Animal studies: SYRCLE Risk of Bias tool
- Multi-omics and mechanistic studies: STROBE-Omics checklist
Studies were rated as low, moderate, or high quality using validated appraisal tools, and low-quality studies were excluded from the thematic synthesis to preserve interpretive reliability. Detailed quality scoring is presented in Supplementary Table S1, while the characteristics of all included studies—such as design, strain, condition, and tools used—are summarized in Supplementary Table S2. A synthesis of the major therapeutic outcomes associated with B. infantis and L. reuteri across various domains is presented in Table 3.
Data Synthesis
Due to the heterogeneity of models, interventions, and outcomes, a narrative synthesis approach was applied.
Studies were grouped into five thematic domains:
- Functional role and ecological significance of B. infantis and L. reuteri
- Drivers of microbial depletion in industrialized contexts
- Consequences of loss on host immune, metabolic, and neurobehavioral health
- Mechanisms of restoration and clinical efficacy of interventions
- Host–microbe coadaptation and microbiome-targeted personalization
Quantitative data (e.g., SCFA levels, behavior scores, inflammatory markers) were summarized descriptively where reported, but meta-analysis was not feasible due to variability in endpoints, analytical tools, and reporting standards.
Results
This Qualitative Systematic Review (QualSR) synthesized findings from 56 eligible studies (36 human, 20 animal) to evaluate the ecological roles, clinical significance, and restoration strategies for Bifidobacterium infantis (B. infantis) and Lactobacillus reuteri (L. reuteri) as keystone taxa within the gut microbiome. Studies were categorized into four core domains: (i) microbial depletion patterns, (ii) metabolic and immune impacts, (iii) neurobehavioral outcomes, and (iv) efficacy of restoration strategies.
1. Patterns of Microbial Depletion in Industrialized Populations
Cross-sectional data consistently showed a 40–60% reduction in overall gut microbial diversity among Western populations compared to non-industrialized cohorts (p < 0.001), driven by the loss of ancestral taxa (e.g., Prevotellaceae, Spirochaetaceae) critical for fiber metabolism [28,69]. This depletion disproportionately affects SCFA-producing genera, including Bifidobacterium, Lactobacillus, and Prevotella, which are diminished in industrialized gut ecosystems [70,71,72]. Specifically, B. infantis and L. reuteri—species essential for butyrate/propionate synthesis and mucosal immunity—are frequently undetectable in dysbiotic microbiomes [72,73,74]. Recent studies show that the gut microbiome and its short-chain fatty acid (SCFA) metabolites play a key role in metabolic health [75]. Bidirectional Mendelian Randomization (MR) analysis revealed that a host genetic-driven increase in gut butyrate production is causally linked to improved insulin response after an oral glucose tolerance test (P = 9.8 × 10⁻⁵) [76]. In contrast, impaired production or absorption of propionate is causally associated with a higher risk of type 2 diabetes (P = 0.004) [76]. Butyrate improves insulin response, while propionate is linked to a higher risk of type 2 diabetes [76,77]. Gut composition, shaped by diet and environment, influences SCFA output and chronic disease risk [78,79].
Drivers of Microbial Depletion in Dysbiotic Gut Microbiomes
Mechanistic analyses identified the following key factors associated with depletion of beneficial taxa (e.g., Bifidobacterium infantis, Lactobacillus reuteri) and reduced microbial richness:
-
Pharmaceutical agents: Antibiotic exposure, both acute and cumulative, significantly alters microbiome composition and reduces microbial diversity. Perinatal antibiotic exposure triggered an initial suppression of microbial phylogenetic diversity (P < 0.0001 at birth) followed by compensatory hyper-restoration, with richness recovery rates exceeding untreated controls by 12 months [42,80]. Non-steroidal anti-inflammatory drugs (NSAIDs) were associated with:
- Exacerbated mucosal injury when co-administered with PPIs. Endoscopic evaluation revealed significantly higher rates of small bowel mucosal injury in patients receiving nonselective NSAID-PPI combination therapy compared to controls (60-80% vs 16.7%, P = .04) [83]. COX-2 inhibitor/PPI coadministration demonstrated intermediate toxicity (44%) [83]. Lesion burden varied by anatomical site, with the jejunum showing particular vulnerability (P = 0.03 for injury severity) [83]. The observed dose-response relationship (P = 0.02 for erosion count gradient) supports a synergistic damaging mechanism between gastric acid suppression and NSAID-mediated mucosal injury [83].
- Perinatal factors, including cesarean delivery and reduced breastfeeding duration, were linked to early-life microbial deficits [84]. In a cohort of 102 infants, gut microbiota composition at 6 weeks was significantly associated with delivery mode (P < .001; Q < .001) and feeding method (P = .01; Q < .001) [85]. Vaginal delivery (vs cesarean) was linked to increased Bacteroides abundance (P < .001; Q = .02) [85]. Cesarean birth caused greater shifts in microbial profiles than feeding differences (P = .003) [85]. Mixed-fed infants resembled formula-fed peers (P = .002) [85].
- Dietary shifts toward low-fiber and high-processed foods correlated with decreased SCFA-producing taxa [86,87]. Analysis of 64 studies (n = 2099) showed that dietary fiber supplementation significantly increased the relative abundance of Bifidobacterium spp. (SMD = 0.64; 95% CI: 0.42–0.86; P < .00001) and Lactobacillus spp. (SMD = 0.22; 95% CI: 0.03–0.41; P = .02), alongside modest gains in fecal butyrate levels (SMD = 0.24; 95% CI: 0.00–0.47; P = .05), compared to placebo or low-fiber controls [88].
- Environmental pollutants (e.g., glyphosate, emulsifiers, artificial sweeteners) exhibited dose-dependent inhibitory effects on commensal bacteria [89]. Environmental pollutants are known to disrupt the balance of gut microbiota, leading to dysbiosis, and can consequently exert various detrimental effects on overall health [89,90]. Food-borne toxicants and additives disrupt gut microbiota function, compromising intestinal barrier integrity and promoting metabolic disease development [91]. Targeting microbe-toxicant interactions through interventions like fermentable fiber may mitigate these metabolic disruptions [91]. Research consistently demonstrates that these pollutants can specifically inhibit the beneficial functions and composition of the gut microbiota [90,91].
- Gastric acid suppression (proton pump inhibitors, PPIs) was uniquely associated with a reduction in microbial richness [92]. Among 211 PPI users, stool microbiome analysis revealed a significant reduction in Shannon diversity and alterations in approximately 20% of bacterial taxa (FDR < 0.05) [92]. PPI use was associated with increased abundance of oral-origin genera including Rothia (p = 9.8×10⁻³⁸), as well as elevated levels of Enterococcus, Streptococcus, Staphylococcus, and Escherichia coli [92]. Using one-tailed Wilcoxon rank sum tests on 1827 individuals, with a significance threshold of P<.05, findings revealed a significant reduction in gut microbiome diversity among Proton Pump Inhibitor (PPI) users compared to non-users [93]. This indicates a notable impact of PPIs on the gut microbiota [93].
- 2.
- Metabolic and Immune Outcomes After Keystone Restoration
Recent findings underscore the role of early-life gut microbiome modulation in promoting infant immune health optimization. In a randomized trial, Lactobacillus reuteri DSM 17938 supplementation in formula-fed infants delivered via cesarean section shifted their gut microbiota composition to resemble that of vaginally delivered infants [94]. L. reuteri was also shown to enhance mucosal barrier function, exert antimicrobial effects, and modulate immune activity [95]. Similarly, supplementation with Bifidobacterium infantis EVC001 in exclusively breastfed infants significantly reduced intestinal inflammation, as indicated by lower levels of fecal pro-inflammatory cytokines and calprotectin [96]. B. infantis utilizes human milk oligosaccharides (HMOs) and is associated with reduced systemic inflammation and improved immune regulation in early life [97]. Notably, its metabolite, indole-3-lactic acid, has been shown to induce galectin-1 expression in T helper cells, contributing to immune tolerance [98,99]. Together, these studies demonstrate that targeted early-life supplementation with L. reuteri DSM 17938 and B. infantis EVC001 can modulate gut microbiota composition, reduce intestinal inflammation, and enhance immunoregulatory responses in infants.
In a post hoc analysis, approximately 70% of individuals with ulcerative colitis (UC), chronic fatigue syndrome (CFS), or psoriasis who received Bifidobacterium infantis 35624 supplementation for 6–8 weeks showed marked reductions across all three measured systemic inflammatory biomarkers [100]. Specifically, plasma TNF-α levels were significantly lower in B. infantis 35624-fed subjects compared to placebo controls in both psoriasis (p = 0.04) and CFS (p = 0.02), suggesting a broad anti-inflammatory effect potentially mediated through regulatory T cell activity [100].
Ex vivo studies in healthy individuals supported these findings. Following B. infantis 35624 supplementation, secretion of IL-6 and TNF-α from lipopolysaccharide (LPS)-stimulated peripheral blood mononuclear cells (PBMCs) was significantly reduced compared to placebo-fed controls [100]. In vivo, B. infantis 35624 consistently lowered systemic pro-inflammatory biomarkers in both gastrointestinal and extraintestinal inflammatory disorders, including UC, CFS, and psoriasis [100].
Additional evidence from a clinical trial in diabetic patients showed that a multispecies probiotic formulation containing B. infantis significantly decreased serum high-sensitivity C-reactive protein (hs-CRP) levels and increased plasma glutathione, indicating both anti-inflammatory and antioxidant effects [101].
Mechanistic studies in animal models further reinforced these observations. In both wild-type and IL-10 knockout mice, B. infantis 35624 reduced interferon-γ levels in Peyer's patches, suggesting that its immunomodulatory actions may occur independently of IL-10 signaling [102]. These findings indicate that B. infantis 35624 can modulate immune responses at both systemic and mucosal levels in these experimental models. A consolidated overview of these physiological benefits is presented in Table 2, summarizing the immunologic, metabolic, neurobehavioral, and gastrointestinal outcomes linked to B. infantis and L. reuteri across various populations and models.
In a murine model of dextran sodium sulfate (DSS)-induced colitis, Bifidobacterium infantis supplementation exerted significant dose-dependent protective effects [103]. Mice receiving B. infantis showed increased body weight and significantly reduced disease activity index (DAI) and histological damage scores compared to the DSS-only group (all p<0.05) [103]. Treatment significantly elevated the expression of the regulatory T cell marker Foxp3 (p<0.05) and anti-inflammatory cytokines IL-10 and TGF-β1 in colon tissue (both p<0.05) [103]. PD-L1 expression was also significantly upregulated in the treatment groups (p<0.05), with a dose-dependent relationship observed for both Foxp3 and PD-L1 expression [103]. Correlation analysis revealed that PD-L1 levels were positively associated with Foxp3, IL-10, and TGF-β1 expression [103]. These results suggest that B. infantis mitigates intestinal inflammation and promotes immune tolerance through Treg cell activation and anti-inflammatory cytokine production.
Consistent with these findings, B. infantis has demonstrated protective effects in other murine models of inflammatory bowel disease (IBD), primarily by increasing Treg populations and suppressing pro-inflammatory Th1 and Th17 responses [104]. This strain enhances colonic expression of Foxp3, IL-10, and TGF-β1, and upregulates PD-L1, which positively correlates with anti-inflammatory mediators [103]. Beyond IBD, B. infantis has also been shown to inhibit NF-κB activation in response to Salmonella typhimurium infection and LPS exposure. This effect is Treg-mediated and associated with reduced secretion of pro-inflammatory cytokines, lower T cell proliferation, and decreased dendritic cell activation [105].
- 3.
- Neurobehavioral and Gut-Brain Axis Effects
Specific probiotics have shown potential in modulating the microbiota–gut–brain axis and altering neurobehavioral outcomes. In murine models, Lactobacillus reuteri administration significantly attenuated depressive-like behaviors by modulating gut microbiota composition and enhancing serotonin metabolism [106]. A synbiotic formulation of L. reuteri and inulin improved autism spectrum disorder (ASD)-like social impairments, supporting gut–brain axis involvement [107]. Similarly, Bifidobacterium infantis reversed behavioral deficits and normalized immune responses in a rat depression model [108].
Several Bifidobacterium and Lactobacillus strains have been shown to modulate anxiety, depression, and memory in both animals and humans through vagus nerve signaling and gut-derived neurotransmitter pathways [108,109,110,111]. These findings align with previous reports demonstrating that Lactobacillus rhamnosus and Bifidobacterium longum influence CNS biochemistry and behavior via vagal mechanisms [112,113]. In particular, B. infantis has shown consistent immunomodulatory and behavioral benefits in depressive models, indicating broader therapeutic potential.
Disruptions in gut microbiota have also been shown to significantly alter neurodevelopment, as demonstrated in germ-free, antibiotic-fed, and fecal microbiota-transplanted mouse models [114]. These findings support a broader gut–immune–brain axis, involving neuroimmune circuits such as brain-resident T cells in CNS regulation [114]. In line with these observations, L. reuteri and L. bulgaricus fermented yogurt significantly alleviated anxiety- and depression-like behaviors in mice exposed to chronic unpredictable mild stress (CUMS) [115]. These effects were associated with hippocampal neuroprotection, preserved cerebral activity, and normalization of monoamine neurotransmitters. The treatment also inhibited NF-κB activation while enhancing CREB–BDNF signaling and reducing inflammatory cytokines [115]. Changes in gut microbial composition and metabolites further supported a microbiota–gut–brain axis mechanism [115].
Complementary studies have shown that gut microbiota alterations affect mood, pain, and cognition via immune, hormonal, metabolic, and neural pathways [116,117]. Early-life gut colonization influences immune and endocrine development, potentially shaping CNS function [117]. In pediatric populations, L. reuteri has reduced crying time in infantile colic, though evidence for other neurogastroenterological disorders remains limited [118]. Microbial production of neurotransmitters and modulation of central neurochemistry further underscore the therapeutic potential of microbiota-based interventions in neuropsychiatric conditions, including anxiety, depression, and ASD [116,119].
- 4.
- Efficacy of Restoration Strategies
Lactobacillus reuteri has demonstrated therapeutic potential across metabolic, gastrointestinal, and inflammatory conditions. In type 2 diabetes patients, supplementation with L. reuteri DSM 17938 significantly improved insulin sensitivity and glucose metabolism [120]. In four randomized controlled trials (RCTs) involving 347 children, L. reuteri reduced the duration of acute diarrhea compared to placebo or no treatment (mean difference [MD] = −0.87 days; 95% CI: −1.43 to −0.31) [121]. Meta-analysis of three RCTs (n = 284) showed a reduction in hospitalization duration (MD = −0.54 days; 95% CI: −1.09 to 0.00) [121]. A separate controlled study (n = 127) further confirmed these effects: significantly more children receiving L. reuteri were diarrhea-free at 24 hours (50% vs. 5%, p < 0.001), 48 hours (69% vs. 11%, p < 0.001), and 72 hours (69% vs. 11%, p < 0.001) compared to controls. Mean hospitalization duration was shorter in the L. reuteri group (4.31 ± 1.3 days vs. 5.46 ± 1.77 days; p < 0.001), and no cases of prolonged diarrhea were observed in this group (0% vs. 17% in controls) [122]. No adverse events were reported, highlighting its favorable safety profile.
In murine models of colitis, L. reuteri I5007 reduced intestinal inflammation, modulated gut microbiota, and improved associated metabolic dysfunction [123]. Strain FYNLJ109L1 demonstrated systemic benefits, including improved glucose tolerance, better lipid profiles, and decreased pro-inflammatory cytokine levels in mice with metabolic syndrome [124]. Additional studies confirmed L. reuteri’s immunoregulatory effects and ability to restore microbial balance in colitis models [123]. Other Lactobacillus and Bifidobacterium strains also exhibit therapeutic potential in conditions such as eczema, obesity, and hypercholesterolemia, through mechanisms including enhanced epithelial barrier integrity and cholesterol-lowering effects [125]. These effects appear strain-specific, reinforcing the importance of precise probiotic selection.
Collectively, these findings support gut microbiota modulation—particularly through targeted L. reuteri strains—as a promising strategy for managing inflammatory and metabolic disorders. This aligns with broader evidence implicating gut dysbiosis in the pathophysiology of obesity and type 2 diabetes, and supports the therapeutic value of microbiome restoration [126]. In a randomized trial of 120 ulcerative colitis patients, participants received probiotics (Bifidobacterium longum, 2 × 10⁹ CFU/day), prebiotics (8 g psyllium/day), or a synbiotic combination [300]. While all groups showed improvement in Inflammatory Bowel Disease Questionnaire scores, the synbiotic group demonstrated statistically significant gains (from 168 to 176; P = 0.03) [300]. Specific functional domains improved across groups: emotional function (probiotics, P = 0.03), bowel function (prebiotics, P = 0.04), and both systemic (P = 0.008) and social function (synbiotics, P = 0.02) [127]. Notably, only the synbiotic group exhibited a significant decrease in C-reactive protein (0.59 to 0.14 mg/dL; P = 0.04), with no reported adverse events [127]. These results indicate that synbiotic therapy provides greater clinical and anti-inflammatory benefit than either component alone.
Discussion
Principal Findings and Clinical Implications
This QualSR supports evidence of gut dysbiosis in industrialized populations, including a 40–60% reduction in microbial diversity and consistent depletion of key short-chain fatty acid (SCFA)-producing taxa such as Bifidobacterium infantis and Lactobacillus reuteri. A multi-cohort study demonstrated that gut microbial diversity was highest in traditional rural communities and progressively declined among first- and second-generation U.S. immigrants [128]. This pattern reflects a broader ecological collapse of keystone taxa that regulate immune, metabolic, and neurobehavioral homeostasis via SCFA production and gut-brain axis signaling. Mendelian randomization studies support the clinical relevance of SCFAs: increased butyrate production improves insulin sensitivity, while impaired propionate metabolism raises the risk of type 2 diabetes [76,129]. This cascade is illustrated in Figure 2, depicting the pathway from dysbiosis to chronic disease and potential restoration strategies.
Figure 3.
Conceptual Framework: From Dysbiosis to Disease—The Role of Keystone Microbial Species. This framework illustrates the progression from modern environmental exposures to microbial keystone depletion, and the cascading biological and clinical effects that follow. The figure outlines: (A) lifestyle disruptors such as antibiotics, cesarean delivery, and poor diet; (B) loss of B. infantis and L. reuteri; (C) impacts on gut barrier, immune signaling, short-chain fatty acid (SCFA) production, and neurobehavioral function; and (D) potential restoration pathways leading to improved host outcomes. Notes and Abbreviations: SCFAs = short-chain fatty acids; PPIs = proton pump inhibitors; B. infantis = Bifidobacterium infantis; L. reuteri = Lactobacillus reuteri.
Figure 3.
Conceptual Framework: From Dysbiosis to Disease—The Role of Keystone Microbial Species. This framework illustrates the progression from modern environmental exposures to microbial keystone depletion, and the cascading biological and clinical effects that follow. The figure outlines: (A) lifestyle disruptors such as antibiotics, cesarean delivery, and poor diet; (B) loss of B. infantis and L. reuteri; (C) impacts on gut barrier, immune signaling, short-chain fatty acid (SCFA) production, and neurobehavioral function; and (D) potential restoration pathways leading to improved host outcomes. Notes and Abbreviations: SCFAs = short-chain fatty acids; PPIs = proton pump inhibitors; B. infantis = Bifidobacterium infantis; L. reuteri = Lactobacillus reuteri.

This QualES of 56 studies found that B. infantis and L. reuteri are frequently undetectable in Western guts, but their targeted supplementation yields consistent therapeutic benefits across diverse clinical contexts. In trials involving patients with ulcerative colitis, chronic fatigue, and psoriasis, B. infantis 35624 significantly reduced systemic inflammation by lowering C-reactive protein and pro-inflammatory cytokines (TNF-α, IL-6) by up to 50% (p < 0.001) [100]. In animal models, B. infantis promoted regulatory immunity and improved epithelial barrier integrity, with synbiotic treatment increasing ZO-1, occludin, and claudin-1 expression by 30–40% in colitis models [103,130].
The L. reuteri supplementation produced comparable benefits. In diabetic patients, strain DSM 17938 enhanced GLP-1 and GLP-2 secretion and increased insulin output by approximately 50% over four weeks [131]. Pediatric randomized controlled trials (RCTs) demonstrated a ~0.9-day reduction in diarrhea duration (MD –24.8 h; 95% CI: –38.8 to –10.8) and improved early recovery rates [132]. In mouse models, strain NK33 improved mood and reduced colitis via hippocampal BDNF upregulation and NF-κB inhibition [133]. Together, these data support the clinical utility of B. infantis and L. reuteri for immune, metabolic, and neurobehavioral dysregulation associated with modern dysbiosis.
Mechanistic Insights
Mechanistic evidence explains how these two keystone taxa exert outsized biological effects. Figure 2 illustrates the key mechanistic pathways through which B. infantis and L. reuteri modulate host physiology—ranging from SCFA biosynthesis and mucosal immune regulation to gut–brain signaling and neuroendocrine modulation. B. infantis is uniquely adapted to human milk oligosaccharides (HMOs), fermenting them into acetate and propionate—metabolites that feed colonocytes and expand Foxp3+ regulatory T cells (Tregs). In colitis models, B. infantis drove dose-dependent increases in Foxp3+ Tregs and IL-10/TGF-β1 levels and activated the PD-1/PD-L1 immune checkpoint pathway, supporting its anti-inflammatory profile [103,130]. It also improved gut barrier function by upregulating tight junction proteins such as ZO-1, occludin, and claudin-1 [130].
The L. reuteri synthesizes bioactive compounds like reuterin and histamine, which modulate immunity by suppressing NF-κB and lowering TNF-α and IL-6 while increasing IL-10 systemically and in mucosal tissues [100,103]. Meta-analyses show that L. reuteri is among the most effective probiotics at restoring tight junction integrity—particularly claudin-1 and ZO-1—in inflamed gut tissue [134]. These species also engage the gut-brain axis. L. reuteri signals via the vagus nerve to stimulate hypothalamic oxytocin neurons, restoring social behaviors in autism models, even in the absence of adaptive immunity [135]. Strain NK33 improved anxiolytic behavior through hippocampal BDNF/CREB activation and NF-κB suppression [135].
Metabolically, B. infantis and L. reuteri increase SCFA levels that act on host G-protein-coupled receptors (GPRs) and inhibit histone deacetylases (HDACs), improving glucose tolerance and lipid handling. Mendelian-randomization studies confirm a causal role: genetically higher butyrate predicts improved insulin response, whereas poor propionate utilization increases diabetes risk [133].
Clinical and Public Health Implications
These findings have immediate translational value in population-based studies. In inflammatory diseases, probiotic formulations containing B. infantis or L. reuteri may serve as adjunct therapies. For example, B. infantis combined with psyllium improved quality of life and reduced CRP in IBD patients [130]. In extraintestinal inflammation (psoriasis, CFS, UC), B. infantis 35624 supplementation for 6–8 weeks lowered systemic CRP and TNF-α [100], an effect mirrored by increased Tregs in colonic tissue [100].
For metabolic dysfunction, L. reuteri’s enhancement of incretin hormones (GLP-1/GLP-2) and insulin secretion [103] suggests potential for managing prediabetes and T2D. Although no change in insulin sensitivity was observed over 4 weeks in healthy volunteers [131], longer trials in dysmetabolic populations are justified. Co-administering L. reuteri DSM 17938 with metformin in clinical practice may be safe and potentially synergistic.
In pediatric gastroenterology, L. reuteri DSM 17938 consistently shortened diarrhea duration by ~1 day in acute gastroenteritis across multiple RCTs (n ≈ 300), with no serious adverse events [131]. This has major implications for resource-limited settings by accelerating rehydration and discharge. Preliminary data support neurodevelopmental applications. L. reuteri reversed social deficits in ASD mouse models via vagus-mediated gut–brain signaling [132]. Given the unmet need in ASD treatment, pilot trials using L. reuteri-rich foods or supplements in affected children are warranted. For mood disorders, multi-strain probiotics containing Lactobacillus and Bifidobacterium have modest anxiolytic/antidepressant effects. Strain-specific data—especially for NK33 and DSM 17938—support future trials. In infantile colic, L. reuteri reduces crying duration, indicating neuromodulatory effects [135].
Strain specificity is essential. Clinicians should use evidence-backed strains such as B. infantis 35624 or EVC001, and L. reuteri DSM 17938 or NK33. Synbiotic approaches may amplify effects: B. infantis combined with xylooligosaccharide or psyllium further increased IL-10 and tight junction expression [133]. Co-prescribing prebiotics like inulin or GOS can also enhance colonization. Personalized protocols, ideally guided by microbiome profiling, are the future; in the interim, evidence-based strain selection should guide clinical use.
Public health policy and implementation must address the erosion of these keystone species. Promoting vaginal delivery and exclusive breastfeeding helps seed B. infantis and shape early immunity [88,130]. Antibiotic stewardship—especially in pregnant women, infants, and children—can reduce collateral loss of these beneficial taxa. Regulatory bodies should consider labeling additives (emulsifiers, glyphosate, artificial sweeteners) that harm gut microbes, given their link to reduced Bifidobacterium and epithelial disruption [100,103].
Diet is a modifiable determinant of microbial composition. Fiber-rich diets containing inulin, FOS, and GOS substantially increase the abundance of Bifidobacterium and Lactobacillus [136]. National programs should promote whole plant foods and include prebiotic supplementation in at-risk groups (infants, elderly, institutionalized). Schools and worksites could provide synbiotic-rich foods like yogurt and fortified bread. Proton pump inhibitors, which reduce diversity and promote opportunistic overgrowth [88], should be prescribed with caution and time limited.
Together, clinical, dietary, obstetric, and pharmaceutical strategies can facilitate microbiome restoration and preserve keystone microbial species, potentially mitigating the rising burden of chronic diseases associated with microbiome collapse.
QualES Limitations
The QualEs remains remarkable based on the implicated findings, however there are several limitations which constrain the interpretation and clinical application of our findings. The reviewed studies varied widely in design, probiotic strain selection, dosing, delivery formats, and outcome measures, which precluded formal meta-analysis and limits the precision of effect estimates. Many of the mechanistic insights—especially those involving brain–behavior modulation—were derived from animal models. For instance, while L. reuteri reversed autism-like social deficits in rodents [135], comparable human trials are lacking.
Short-term studies dominate the literature. This raises concerns about the sustainability of therapeutic effects and the potential for adaptive host or microbial responses that diminish efficacy over time. Furthermore, many studies relied on 16S rRNA sequencing, which limits taxonomic and functional resolution. Broader use of metagenomics and metabolomics is necessary to better characterize keystone species’ functions and interactions.
Population diversity is another limitation. Most trials were conducted in Western, homogeneous cohorts, reducing generalizability to other ethnic and geographic populations. Host-specific factors—such as baseline microbiota composition, dietary habits, and genetics—likely influence probiotic efficacy, making single-trial findings difficult to extrapolate.
Publication bias is also a concern. Positive findings tend to be overrepresented, while underpowered studies with null results (e.g., L. reuteri in antibiotic-associated diarrhea) are less frequently published [132]. Additionally, the review focused on B. infantis and L. reuteri, which simplifies the complexity of microbiome restoration. While these are keystone taxa, broader community dynamics and diet–microbe interactions are likely essential for lasting clinical benefit.
Finally, clinical translation is hampered by a lack of standardization. Variation in probiotic preparation, dosing protocols, strain specificity, and delivery matrices create inconsistencies in outcomes. Long-term safety, including risks such as small intestinal bacterial overgrowth or horizontal gene transfer, remains insufficiently studied.
Conclusion and Future Directions
This systematic review highlights the therapeutic potential of restoring keystone microbial species—particularly Bifidobacterium infantis and Lactobacillus reuteri—as a viable strategy to counteract the chronic disease burden driven by modern lifestyle-induced dysbiosis. The evidence from mechanistic studies, preclinical models, and human trials supports their role in modulating host immunity, metabolism, and gut-brain signaling across various disease contexts.
For successful clinical translation, key challenges remain in optimizing microbiome-targeted interventions for human disease. These include the development of personalized microbial restoration protocols, refinement of delivery systems to enhance colonization and persistence, and integration of microbiome-targeted interventions into existing clinical and public health frameworks. Given the complex, bidirectional nature of host-microbe interactions, advancing this field will require interdisciplinary collaboration across microbiology, immunology, neuroscience, and systems biology.
It is essential that microbiome restoration must not be limited to therapeutic supplementation alone, however a comprehensive approach should also address upstream environmental determinants such as antibiotic overuse, toxicant exposure, dietary patterns, and early-life microbial disruptions. The keystone species restoration paradigm provides a scientifically grounded and translationally relevant model for reversing microbial depletion and its systemic consequences.
This review highlights how precision microbiome medicine, through epigenomic modulation, supports the development of targeted, sustainable, and personalized interventions to restore human–microbe symbiosis and improve long-term health outcomes.
Microbiomes Implication in Disease Causation, Health Outcomes and Future Directions
Future research should involve integrative, translational approach to address these gaps and advance the clinical utility of B. infantis and L. reuteri.
- Personalized Interventions: Development of diagnostic tools using microbiome sequencing, SCFA profiling, or immune biomarkers will allow precision targeting. Trials should stratify patients based on baseline microbial or immunologic markers to assess differential responses to probiotic therapy.
- Longitudinal and Intergenerational Studies: Extended follow-up is needed to determine the durability of benefits and their effects across generations. For example, supplementing pregnant women with B. infantis could be studied for its impact on neonatal microbiota and early immune development.
- Next-Generation Therapeutics: Engineered strains or microbial consortia could be designed to deliver targeted metabolites or immunomodulators. For instance, a modified B. infantis that overproduces acetate, or L. reuteri strains that optimize oxytocin signaling, could enhance therapeutic precision.
- Mechanistic Human Studies: Future trials should integrate multi-omics with host physiology data. Measuring changes in host gene expression, epigenetics, inflammatory markers, neuroimaging, and vagal tone alongside microbiome dynamics will clarify causal pathways.
- Population-Level Research: Real-world interventions—such as fiber subsidies, fermented food promotion, or early-life microbial seeding—should be evaluated for their capacity to restore keystone taxa and reduce chronic disease incidence.
- Regulatory and Safety Frameworks: As probiotics become clinical tools, robust long-term safety monitoring is essential. Registries and pharmacovigilance systems can track rare adverse events or ecological risks. Regulatory agencies should also evolve to include microbiome endpoints in risk-benefit assessments.
Pursuing these research directions requires collaborative, symbiotic, mutualistic, transdisciplinary, dynamic and translational initiatives and application of scientific data across microbiology, immunology, epigenomics, genomics, clinical medicine, and public health. This translational initiatives as basic medical sciences, clinical correlates and population-based findings is to optimize these findings in microbiomes diversity beyond correlation and causal inference, hence B. infantis and L. reuteri as translational initiatives in restoring host–microbe homeostasis as well as human health improvement and optimization across all populations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef]
- Rook, G.; Bäckhed, F.; Levin, B.R.; McFall-Ngai, M.J.; McLean, A.R. Evolution, human-microbe interactions, and life history plasticity. Lancet 2017, 390, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Hills, Ronald D., Benjamin A. Pontefract, Hillary R. Mishcon, Cody A. Black, Steven C. Sutton, and Cory R. Theberge. "Gut microbiome: profound implications for diet and disease." Nutrients 11, no. 7 (2019): 1613.
- Shelton, C.D.; Byndloss, M.X.; Richardson, A.R. Gut Epithelial Metabolism as a Key Driver of Intestinal Dysbiosis Associated with Noncommunicable Diseases. Infect. Immun. 2020, 88, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, E.D.; Sonnenburg, J.L. The ancestral and industrialized gut microbiota and implications for human health. Nat. Rev. Microbiol. 2019, 17, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Bicknell, B.; Liebert, A.; Borody, T.; Herkes, G.; McLachlan, C.; Kiat, H. Neurodegenerative and Neurodevelopmental Diseases and the Gut-Brain Axis: The Potential of Therapeutic Targeting of the Microbiome. Int. J. Mol. Sci. 2023, 24, 9577. [Google Scholar] [CrossRef]
- Tett, A.; Pasolli, E.; Masetti, G.; Ercolini, D.; Segata, N. Prevotella diversity, niches and interactions with the human host. Nat. Rev. Microbiol. 2021, 19, 585–599. [Google Scholar] [CrossRef]
- Duncan, S.H.; Richardson, A.J.; Kaul, P.; Holmes, R.P.; Allison, M.J.; Stewart, C.S. Oxalobacter formigenes and Its Potential Role in Human Health. Appl. Environ. Microbiol. 2002, 68, 3841–3847. [Google Scholar] [CrossRef]
- Camara, A.; Konate, S.; Alou, M.T.; Kodio, A.; Togo, A.H.; Cortaredona, S.; Henrissat, B.; Thera, M.A.; Doumbo, O.K.; Raoult, D.; et al. Clinical evidence of the role of Methanobrevibacter smithii in severe acute malnutrition. Sci. Rep. 2021, 11, 5426. [Google Scholar] [CrossRef]
- Belkacemi, Souad, Maryam Tidjani Alou, Matthieu Million, Anthony Levasseur, Saber Khelaifia, And Didier Raoult. "Prevalence of Treponema species in the Gut Microbiome is Linked to Bifidobacterium sp. and Bacteroides sp." (2020).
- Tomás-Pejó, E.; González-Fernández, C.; Greses, S.; Kennes, C.; Otero-Logilde, N.; Veiga, M.C.; Bolzonella, D.; Müller, B.; Passoth, V. Production of short-chain fatty acids (SCFAs) as chemicals or substrates for microbes to obtain biochemicals. Biotechnol. Biofuels Bioprod. 2023, 16, 96. [Google Scholar] [CrossRef]
- Louis, P.; Young, P.; Holtrop, G.; Flint, H.J. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environ. Microbiol. 2010, 12, 304–314. [Google Scholar] [CrossRef]
- Zhang, Dan, Yong-Ping Jian, Yu-Ning Zhang, Yao Li, Li-Ting Gu, Hui-Hui Sun, Ming-Di Liu, Hong-Lan Zhou, Yi-Shu Wang, and Zhi-Xiang Xu. "Short-chain fatty acids in diseases." Cell Communication and Signaling 21, no. 1 (2023): 212.
- Ikeda, Takako, Akari Nishida, Mayu Yamano, and Ikuo Kimura. "Short-chain fatty acid receptors and gut microbiota as therapeutic targets in metabolic, immune, and neurological diseases." Pharmacology & therapeutics 239 (2022): 108273.
- Banerjee, S.; Schlaeppi, K.; Van Der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Brushett, S.; Sinha, T.; Reijneveld, S.A.; de Kroon, M.L.A.; Zhernakova, A. The Effects of Urbanization on the Infant Gut Microbiota and Health Outcomes. Front. Pediatr. 2020, 8, 408. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cao, Q.; Cheng, Y.; Zhao, D.; Wang, Z.; Yang, H.; Wu, Q.; You, L.; Wang, Y.; Lin, Y.; et al. Chronic stress promotes colitis by disturbing the gut microbiota and triggering immune system response. Proc. Natl. Acad. Sci. 2018, 115, E2960–E2969. [Google Scholar] [CrossRef] [PubMed]
- Abjani, F.; Madhavan, P.; Chong, P.P.; Chinna, K.; Rhodes, C.A.; Lim, Y.A.L. Urbanisation and its associated factors affecting human gut microbiota: where are we heading to? Ann. Hum. Biol. 2023, 50, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Li, Mingjie, Jiuhong Ding, Catherine Stanton, R. Paul Ross, Jianxin Zhao, Bo Yang, and Wei Chen. "Bifidobacterium longum subsp. infantis FJSYZ1M3 ameliorates DSS-induced colitis by maintaining the intestinal barrier, regulating inflammatory cytokines, and modifying gut microbiota." Food & Function 14, no. 1 (2023): 354-368.
- Wang, Luanfeng, Bo Ren, Shufeng Wu, Haizhao Song, Ling Xiong, Fang Wang, and Xinchun Shen. "Current research progress, opportunities, and challenges of limosillactobacillus reuteri-based probiotic dietary strategies." Critical Reviews in Food Science and Nutrition (2024): 1-21.
- Ağagündüz, D.; Bingöl, F.G.; Çelik, E.; Cemali, Ö.; Özenir, Ç.; Özoğul, F.; Capasso, R. Recent developments in the probiotics as live biotherapeutic products (LBPs) as modulators of gut brain axis related neurological conditions. J. Transl. Med. 2022, 20, 460. [Google Scholar] [CrossRef]
- Satya, Shalni, Sakshi Sharma, Garima Choudhary, and Garima Kaushik. "Advances in Environmental Microbiology: A Multi-omic Perspective." In Microbial Omics in Environment and Health, pp. 175-204. Singapore: Springer Nature Singapore, 2024.
- Seneviratne, C.J.; Suriyanarayanan, T.; Widyarman, A.S.; Lee, L.S.; Lau, M.; Ching, J.; Delaney, C.; Ramage, G. Multi-omics tools for studying microbial biofilms: current perspectives and future directions. Crit. Rev. Microbiol. 2020, 46, 759–778. [Google Scholar] [CrossRef]
- Li, N.; Cai, Q.; Miao, Q.; Song, Z.; Fang, Y.; Hu, B. High-Throughput Metagenomics for Identification of Pathogens in the Clinical Settings. Small Methods 2020, 5, 2000792. [Google Scholar] [CrossRef]
- Xiong, W.; Abraham, P.E.; Li, Z.; Pan, C.; Hettich, R.L. Microbial metaproteomics for characterizing the range of metabolic functions and activities of human gut microbiota. Proteomics 2015, 15, 3424–3438. [Google Scholar] [CrossRef]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef]
- Martinez, K.B.; Leone, V.; Chang, E.B. Western diets, gut dysbiosis, and metabolic diseases: Are they linked? Gut Microbes 2017, 8, 130–142. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Sonnenburg, E.D. Vulnerability of the industrialized microbiota. Science 2019, 366, eaaw9255. [Google Scholar] [CrossRef]
- Lange, Kathleen, Martin Buerger, Andreas Stallmach, and Tony Bruns. "Effects of antibiotics on gut microbiota." Digestive diseases 34, no. 3 (2016): 260-268.
- Cusumano, Gaia, Giancarlo Angeles Flores, Roberto Venanzoni, and Paola Angelini. "The Impact of Antibiotic Therapy on Intestinal Microbiota: Dysbiosis, Antibiotic Resistance, and Restoration Strategies." Antibiotics 14, no. 4 (2025): 371.
- Schwartz, D.J.; Langdon, A.E.; Dantas, G. Understanding the impact of antibiotic perturbation on the human microbiome. Genome Med. 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjölund-Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-Term Antibiotic Treatment Has Differing Long-Term Impacts on the Human Throat and Gut Microbiome. PLOS ONE 2010, 5, e9836. [Google Scholar] [CrossRef] [PubMed]
- Marlicz, W.; Łoniewski, I.; Grimes, D.S.; Quigley, E.M. Nonsteroidal Anti-inflammatory Drugs, Proton Pump Inhibitors, and Gastrointestinal Injury: Contrasting Interactions in the Stomach and Small Intestine. Mayo Clin. Proc. 2014, 89, 1699–1709. [Google Scholar] [CrossRef]
- Wallace, J.L.; Syer, S.; Denou, E.; de Palma, G.; Vong, L.; McKnight, W.; Jury, J.; Bolla, M.; Bercik, P.; Collins, S.M.; et al. Proton Pump Inhibitors Exacerbate NSAID-Induced Small Intestinal Injury by Inducing Dysbiosis. Gastroenterology 2011, 141, 1314–1322.e5. [Google Scholar] [CrossRef] [PubMed]
- Le Bastard, Q.; Al-Ghalith, G.A.; Grégoire, M.; Chapelet, G.; Javaudin, F.; Dailly, E.; Batard, E.; Knights, D.; Montassier, E. Systematic review: human gut dysbiosis induced by non-antibiotic prescription medications. Aliment. Pharmacol. Ther. 2017, 47, 332–345. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.L.Y.; Petersen, C.; Turvey, S.E. Reduce, reinforce, and replenish: safeguarding the early-life microbiota to reduce intergenerational health disparities. Front. Public Heal. 2024, 12, 1455503. [Google Scholar] [CrossRef]
- Puigbò, P.; Leino, L.I.; Rainio, M.J.; Saikkonen, K.; Saloniemi, I.; Helander, M. Does Glyphosate Affect the Human Microbiota? Life 2022, 12, 707. [Google Scholar] [CrossRef]
- Li, P.; Qu, R.; Li, M.; Sheng, P.; Jin, L.; Huang, X.; Xu, Z.Z. Impacts of food additives on gut microbiota and host health. Food Res. Int. 2024, 196, 114998. [Google Scholar] [CrossRef]
- Laudisi, F.; Stolfi, C.; Monteleone, G. Impact of Food Additives on Gut Homeostasis. Nutrients 2019, 11, 2334. [Google Scholar] [CrossRef]
- Inan-Eroglu, E.; Ayaz, A. Effects of food additives on gut microbiota: friend or foe? Nutr. Food Sci. 2019, 49, 955–964. [Google Scholar] [CrossRef]
- Brockway, M. The role of antibiotic exposure and the effects of breastmilk and human milk feeding on the developing infant gut microbiome. Front. Public Heal. 2024, 12, 1408246. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.D.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef] [PubMed]
- Thompson, Amanda L. "Developmental origins of obesity: early feeding environments, infant growth, and the intestinal microbiome." American Journal of Human Biology 24, no. 3 (2012): 350-360.
- Sartor, R.B. Genetics and Environmental Interactions Shape the Intestinal Microbiome to Promote Inflammatory Bowel Disease Versus Mucosal Homeostasis. Gastroenterology 2010, 139, 1816–1819. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, K.; Witkiewicz, W. Correlations of Host Genetics and Gut Microbiome Composition. Front. Microbiol. 2016, 7, 1357. [Google Scholar] [CrossRef]
- Dharmani, P.; Srivastava, V.; Kissoon-Singh, V.; Chadee, K. Role of Intestinal Mucins in Innate Host Defense Mechanisms against Pathogens. J. Innate Immun. 2009, 1, 123–135. [Google Scholar] [CrossRef]
- Janssens, S.; Beyaert, R. Role of Toll-Like Receptors in Pathogen Recognition. Clin. Microbiol. Rev. 2003, 16, 637–646. [Google Scholar] [CrossRef]
- Bauer, Stefan, Thomas Müller, and Svetlana Hamm. "Pattern recognition by Toll-like receptors." Target Pattern Recognition in Innate Immunity (2009): 15-34.
- Bonder, M.J.; Kurilshikov, A.; Tigchelaar, E.F.; Mujagic, Z.; Imhann, F.; Vila, A.V.; Deelen, P.; Vatanen, T.; Schirmer, M.; Smeekens, S.P.; et al. The effect of host genetics on the gut microbiome. Nat. Genet. 2016, 48, 1407–1412. [Google Scholar] [CrossRef]
- Cuomo, Paola, Rosanna Capparelli, Marco Alifano, Antonio Iannelli, and Domenico Iannelli. "Gut microbiota host-gene interaction." International Journal of Molecular Sciences 23, no. 22 (2022): 13717.
- Marwaha, K.; Cain, R.; Asmis, K.; Czaplinski, K.; Holland, N.; Mayer, D.C.G.; Chacon, J. Exploring the complex relationship between psychosocial stress and the gut microbiome: implications for inflammation and immune modulation. J. Appl. Physiol. 2025, 138, 518–535. [Google Scholar] [CrossRef]
- Zhu, X.; Han, Y.; Du, J.; Liu, R.; Jin, K.; Yi, W. Microbiota-gut-brain axis and the central nervous system. Oncotarget 2017, 8, 53829–53838. [Google Scholar] [CrossRef]
- Montagnani, M.; Bottalico, L.; Potenza, M.A.; Charitos, I.A.; Topi, S.; Colella, M.; Santacroce, L. The Crosstalk between Gut Microbiota and Nervous System: A Bidirectional Interaction between Microorganisms and Metabolome. Int. J. Mol. Sci. 2023, 24, 10322. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, Daniela, Marjorie K. De la Fuente, Glauben Landskron, María Julieta González, Rodrigo Quera, Gerard Dijkstra, Hermie JM Harmsen, Klaas Nico Faber, and Marcela A. Hermoso. "Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases." Frontiers in immunology 10 (2019): 277.
- Shin, Y.; Han, S.; Kwon, J.; Ju, S.; Choi, T.G.; Kang, I.; Kim, S.S. Roles of Short-Chain Fatty Acids in Inflammatory Bowel Disease. Nutrients 2023, 15, 4466. [Google Scholar] [CrossRef]
- Sarkar, A.; Harty, S.; Johnson, K.V.; Moeller, A.H.; Carmody, R.N.; Lehto, S.M.; Erdman, S.E.; Dunbar, R.I.M.; Burnet, P.W.J. The role of the microbiome in the neurobiology of social behaviour. Biol. Rev. 2020, 95, 1131–1166. [Google Scholar] [CrossRef] [PubMed]
- Severino, A.; Tohumcu, E.; Tamai, L.; Dargenio, P.; Porcari, S.; Rondinella, D.; Venturini, I.; Maida, M.; Gasbarrini, A.; Cammarota, G.; et al. The microbiome-driven impact of western diet in the development of noncommunicable chronic disorders. Best Pr. Res. Clin. Gastroenterol. 2024, 72, 101923. [Google Scholar] [CrossRef]
- Lloyd-Price, Jason, Galeb Abu-Ali, and Curtis Huttenhower. "The healthy human microbiome." Genome medicine 8 (2016): 1-11.
- Safarchi, Azadeh, Ghanyah Al-Qadami, Cuong D. Tran, and Michael Conlon. "Understanding dysbiosis and resilience in the human gut microbiome: biomarkers, interventions, and challenges." Frontiers in Microbiology 16 (2025): 1559521.
- Musat, N.; Halm, H.; Winterholler, B.; Hoppe, P.; Peduzzi, S.; Hillion, F.; Horreard, F.; Amann, R.; Jørgensen, B.B.; Kuypers, M.M.M. A single-cell view on the ecophysiology of anaerobic phototrophic bacteria. Proc. Natl. Acad. Sci. 2008, 105, 17861–17866. [Google Scholar] [CrossRef] [PubMed]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Chen, Q.; Ma, X.; Guo, Z.; Zhang, P.; Li, B.; Wang, Z. Gut microbiota: A key role for human milk oligosaccharides in regulating host health early in life. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13431. [Google Scholar] [CrossRef]
- Luo, Z.; Chen, A.; Xie, A.; Liu, X.; Jiang, S.; Yu, R. Limosilactobacillus reuteri in immunomodulation: molecular mechanisms and potential applications. Front. Immunol. 2023, 14, 1228754. [Google Scholar] [CrossRef]
- Weber, Katherine T., Bernard J. Varian, and Susan E. Erdman. "The gut microbiome and sociability." Frontiers in Neuroscience 18 (2024): 1372274.
- Freedberg, D.E.; Lebwohl, B.; Abrams, J.A. The Impact of Proton Pump Inhibitors on the Human Gastrointestinal Microbiome. Clin. Lab. Med. 2014, 34, 771–785. [Google Scholar] [CrossRef]
- Beasley, D.E.; Koltz, A.M.; Lambert, J.E.; Fierer, N.; Dunn, R.R.; Li, X. The Evolution of Stomach Acidity and Its Relevance to the Human Microbiome. PLOS ONE 2015, 10, e0134116. [Google Scholar] [CrossRef]
- Walsh, J.; Griffin, B.T.; Clarke, G.; Hyland, N.P. Drug–gut microbiota interactions: implications for neuropharmacology. Br. J. Pharmacol. 2018, 175, 4415–4429. [Google Scholar] [CrossRef]
- Tasnim, N.; Abulizi, N.; Pither, J.; Hart, M.M.; Gibson, D.L. Linking the Gut Microbial Ecosystem with the Environment: Does Gut Health Depend on Where We Live? Front. Microbiol. 2017, 8, 1935–1935. [Google Scholar] [CrossRef]
- Camp, J.G.; Kanther, M.; Semova, I.; Rawls, J.F. Patterns and Scales in Gastrointestinal Microbial Ecology. Gastroenterology 2009, 136, 1989–2002. [Google Scholar] [CrossRef] [PubMed]
- Ramos Meyers, Guilherme, Hanen Samouda, and Torsten Bohn. "Short chain fatty acid metabolism in relation to gut microbiota and genetic variability." Nutrients 14, no. 24 (2022): 5361.
- Dargenio, V.N.; Cristofori, F.; Brindicci, V.F.; Schettini, F.; Dargenio, C.; Castellaneta, S.P.; Iannone, A.; Francavilla, R. Impact of Bifidobacterium longum Subspecies infantis on Pediatric Gut Health and Nutrition: Current Evidence and Future Directions. Nutrients 2024, 16, 3510. [Google Scholar] [CrossRef] [PubMed]
- Tudela, H.; Claus, S.P.; Saleh, M. Next Generation Microbiome Research: Identification of Keystone Species in the Metabolic Regulation of Host-Gut Microbiota Interplay. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.; Araújo, J.R.; Di Santo, J.P. A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 558–572. [Google Scholar] [CrossRef] [PubMed]
- Nogal, A.; Valdes, A.M.; Menni, C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut Microbes 2021, 13, 1897212. [Google Scholar] [CrossRef]
- Sanna, Serena, Natalie R. van Zuydam, Anubha Mahajan, Alexander Kurilshikov, Arnau Vich Vila, Urmo Võsa, Zlatan Mujagic et al. "Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases." Nature genetics 51, no. 4 (2019): 600-605.
- Arora, T.; Tremaroli, V. Therapeutic Potential of Butyrate for Treatment of Type 2 Diabetes. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Rinninella, Emanuele, Pauline Raoul, Marco Cintoni, Francesco Franceschi, Giacinto Abele Donato Miggiano, Antonio Gasbarrini, and Maria Cristina Mele. "What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases." Microorganisms 7, no. 1 (2019): 14.
- Bibbo, Stefano, Gianluca Ianiro, Valentina Giorgio, Franco Scaldaferri, Luca Masucci, Antonio Gasbarrini, and G. J. E. R. M. P. S. Cammarota. "The role of diet on gut microbiota composition." European Review for Medical & Pharmacological Sciences 20, no. 22 (2016).
- Morreale, C.; Giaroni, C.; Baj, A.; Folgori, L.; Barcellini, L.; Dhami, A.; Agosti, M.; Bresesti, I. Effects of Perinatal Antibiotic Exposure and Neonatal Gut Microbiota. Antibiotics 2023, 12, 258. [Google Scholar] [CrossRef]
- Rogers, M.A.M.; Aronoff, D.M. The influence of non-steroidal anti-inflammatory drugs on the gut microbiome. Clin. Microbiol. Infect. 2016, 22, 178.e1. [Google Scholar] [CrossRef]
- Maseda, Damian, and Emanuela Ricciotti. "NSAID–gut microbiota interactions." Frontiers in pharmacology 11 (2020): 558924.
- Washio, Ema, Motohiro Esaki, Yuji Maehata, Masashi Miyazaki, Hiroyuki Kobayashi, Hideki Ishikawa, Takanari Kitazono, and Takayuki Matsumoto. "Proton pump inhibitors increase incidence of nonsteroidal anti-inflammatory drug–induced small bowel injury: a randomized, placebo-controlled trial." Clinical Gastroenterology and Hepatology 14, no. 6 (2016): 809-815.
- Jeong, S. Factors influencing development of the infant microbiota: from prenatal period to early infancy. Clin. Exp. Pediatr. 2021, 65, 438–447. [Google Scholar] [CrossRef]
- Madan, J.C.; Hoen, A.G.; Lundgren, S.N.; Farzan, S.F.; Cottingham, K.L.; Morrison, H.G.; Sogin, M.L.; Li, H.; Moore, J.H.; Karagas, M.R. Association of Cesarean Delivery and Formula Supplementation With the Intestinal Microbiome of 6-Week-Old Infants. JAMA Pediatr. 2016, 170, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Villalba Gimenez, Jazmin. "Gut Microbiome Changes in Urbanizing Brazil and Their Implications for Metabolic Health." (2025).
- Rivera, K.; Gonzalez, L.; Bravo, L.; Manjarres, L.; Andia, M.E. The Gut–Heart Axis: Molecular Perspectives and Implications for Myocardial Infarction. Int. J. Mol. Sci. 2024, 25, 12465. [Google Scholar] [CrossRef] [PubMed]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: a systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [PubMed]
- Tu, P.; Chi, L.; Bodnar, W.; Zhang, Z.; Gao, B.; Bian, X.; Stewart, J.; Fry, R.; Lu, K. Gut Microbiome Toxicity: Connecting the Environment and Gut Microbiome-Associated Diseases. Toxics 2020, 8, 19. [Google Scholar] [CrossRef]
- Jin, Y.; Wu, S.; Zeng, Z.; Fu, Z. Effects of environmental pollutants on gut microbiota. Environ. Pollut. 2017, 222, 1–9. [Google Scholar] [CrossRef]
- Barra, N.G.; Fang, H.; Bhatwa, A.; Schmidt, A.M.; Syed, S.A.; Steinberg, G.R.; Morrison, K.M.; Surette, M.G.; Wade, M.G.; Holloway, A.C.; et al. Food supply toxicants and additives alter the gut microbiota and risk of metabolic disease. Am. J. Physiol. Metab. 2025, 328, E337–E353. [Google Scholar] [CrossRef]
- Imhann, F.; Bonder, M.J.; Vila, A.V.; Fu, J.; Mujagic, Z.; Vork, L.; Tigchelaar, E.F.; Jankipersadsing, S.A.; Cenit, M.C.; Harmsen, H.J.M.; et al. Proton pump inhibitors affect the gut microbiome. Gut 2016, 65, 740–748. [Google Scholar] [CrossRef]
- Jackson, M.A.; Goodrich, J.K.; Maxan, M.-E.; Freedberg, D.E.; Abrams, J.A.; Poole, A.C.; Sutter, J.L.; Welter, D.; Ley, R.E.; Bell, J.T.; et al. Proton pump inhibitors alter the composition of the gut microbiota. Gut 2016, 65, 749–756. [Google Scholar] [CrossRef]
- Rodenas, C.L.G.; Lepage, M.; Ngom-Bru, C.; Fotiou, A.; Papagaroufalis, K.; Berger, B. Effect of Formula Containing Lactobacillus reuteri DSM 17938 on Fecal Microbiota of Infants Born by Cesarean-Section. J. Pediatr. Gastroenterol. Nutr. 2016, 63, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Alam, M. D. , Mirza Islam, Ziaul MD, M. D. Tayab, Hossain Sahid Kamrul Alam, M. D. Kamrul, Salauddin Mahmood, and A. T. M. Haque. "Role of probiotic Lactobacillus reuteri in improving Gut Health and Immunity in infants and toddlers: a review." International Journal of Nutrition Sciences 7, no. 2 (2022): 75-80.
- Henrick, B.; Chew, S.; Mitchell, R.; Contreras, L.; Casaburi, G.; Frese, S.; Smilowitz, J.; Underwood, M. Restoring Bifidobacterium Infantis EVC001 to the Infant Gut Microbiome Significantly Reduces Intestinal Inflammation (OR12-01-19). Curr. Dev. Nutr. 2019, 3, nzz049–OR12. [Google Scholar] [CrossRef]
- Wong, Chyn Boon, Huidong Huang, Yibing Ning, and Jinzhong Xiao. "Probiotics in the new era of human milk oligosaccharides (HMOs): HMO utilization and beneficial effects of Bifidobacterium longum subsp. infantis M-63 on infant health." Microorganisms 12, no. 5 (2024): 1014.
- Gavzy, S.J.; Kensiski, A.; Lee, Z.L.; Mongodin, E.F.; Ma, B.; Bromberg, J.S. Bifidobacterium mechanisms of immune modulation and tolerance. Gut Microbes 2023, 15, 2291164. [Google Scholar] [CrossRef]
- Donald, K.; Finlay, B.B. Early-life interactions between the microbiota and immune system: impact on immune system development and atopic disease. Nat. Rev. Immunol. 2023, 23, 735–748. [Google Scholar] [CrossRef]
- Groeger, D.; O’mAhony, L.; Murphy, E.F.; Bourke, J.F.; Dinan, T.G.; Kiely, B.; Shanahan, F.; Quigley, E.M. Bifidobacterium infantis35624 modulates host inflammatory processes beyond the gut. Gut Microbes 2013, 4, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Asemi, Zatollah, Zohreh Zare, Hossein Shakeri, Sima-sadat Sabihi, and Ahmad Esmaillzadeh. "Effect of multispecies probiotic supplements on metabolic profiles, hs-CRP, and oxidative stress in patients with type 2 diabetes." Annals of nutrition and metabolism 63, no. 1-2 (2013): 1-9.
- Sheil, B.; MacSharry, J.; O'Callaghan, L.; O'Riordan, A.; Waters, A.; Morgan, J.; Collins, J.K.; O'Mahony, L.; Shanahan, F. Role of interleukin (IL-10) in probiotic-mediated immune modulation: an assessment in wild-type and IL-10 knock-out mice. Clin. Exp. Immunol. 2006, 144, 273–280. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, D.; Xie, Y.; Yao, X.; Li, Y. Bifidobacterium infantis Induces Protective Colonic PD-L1 and Foxp3 Regulatory T Cells in an Acute Murine Experimental Model of Inflammatory Bowel Disease. Gut Liver 2019, 13, 430–439. [Google Scholar] [CrossRef]
- Zuo, Li, Kai-Tao Yuan, Li Yu, Qing-Hong Meng, Peter Chee-Keung Chung, and Ding-Hua Yang. "Bifidobacterium infantis attenuates colitis by regulating T cell subset responses." World Journal of Gastroenterology: WJG 20, no. 48 (2014): 18316.
- O'MAhony, C.; Scully, P.; O'MAhony, D.; Murphy, S.; O'BRien, F.; Lyons, A.; Sherlock, G.; MacSharry, J.; Kiely, B.; Shanahan, F.; et al. Commensal-Induced Regulatory T Cells Mediate Protection against Pathogen-Stimulated NF-κB Activation. PLOS Pathog. 2008, 4, e1000112. [Google Scholar] [CrossRef]
- Xie, R.; Jiang, P.; Lin, L.; Jiang, J.; Yu, B.; Rao, J.; Liu, H.; Wei, W.; Qiao, Y. Oral treatment with Lactobacillus reuteri attenuates depressive-like behaviors and serotonin metabolism alterations induced by chronic social defeat stress. J. Psychiatr. Res. 2020, 122, 70–78. [Google Scholar] [CrossRef]
- Wang, C.; Chen, W.; Jiang, Y.; Xiao, X.; Zou, Q.; Liang, J.; Zhao, Y.; Wang, Q.; Yuan, T.; Guo, R.; et al. A synbiotic formulation of Lactobacillus reuteri and inulin alleviates ASD-like behaviors in a mouse model: the mediating role of the gut–brain axis. Food Funct. 2023, 15, 387–400. [Google Scholar] [CrossRef]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Kiely, B.; Cryan, J.F.; Dinan, T.G. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience 2010, 170, 1179–1188. [Google Scholar] [CrossRef]
- Radford-Smith, D.E.; Anthony, D.C. Prebiotic and Probiotic Modulation of the Microbiota–Gut–Brain Axis in Depression. Nutrients 2023, 15, 1880. [Google Scholar] [CrossRef] [PubMed]
- McLean, P.G.; Bergonzelli, G.E.; Collins, S.M.; Bercik, P. Targeting the microbiota–gut–brain axis to modulate behavior: Which bacterial strain will translate best to humans? Proc. Natl. Acad. Sci. 2012, 109, E174–E174. [Google Scholar] [CrossRef]
- Wang, H.; Lee, I.-S.; Braun, C.; Enck, P. Effect of Probiotics on Central Nervous System Functions in Animals and Humans: A Systematic Review. J. Neurogastroenterol. Motil. 2016, 22, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.K.; Oh, J.S. Interaction of the Vagus Nerve and Serotonin in the Gut–Brain Axis. Int. J. Mol. Sci. 2025, 26, 1160. [Google Scholar] [CrossRef]
- Park, J.C.; Im, S.-H. The gut-immune-brain axis in neurodevelopment and neurological disorders. Microbiome Res. Rep. 2022, 1, 23. [Google Scholar] [CrossRef]
- Sun, Yang, Xiujuan Li, Xiulian Li, Lun Liu, and Song Wei. "Lactobacillus yogurts display antidepressant-like effects in CUMS mice via inhibition of NF-κB pathway, activating CREB-BDNF pathway and regulating gut-brain axis." (2024).
- Borre, Yuliya E., Rachel D. Moloney, Gerard Clarke, Timothy G. Dinan, and John F. Cryan. "The impact of microbiota on brain and behavior: mechanisms & therapeutic potential." Microbial endocrinology: The microbiota-gut-brain axis in health and disease (2014): 373-403.
- Foster, J.A.; McVey Neufeld, K.-A. Gut–brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Salvatore, S.; Pensabene, L.; Borrelli, O.; Saps, M.; Thapar, N.; Concolino, D.; Staiano, A.; Vandenplas, Y. Mind the gut: probiotics in paediatric neurogastroenterology. Benef. Microbes 2018, 9, 883–898b. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Brain-Gut-Microbiota Axis and Mental Health. Psychosom. Med. 2017, 79, 920–926. [Google Scholar] [CrossRef]
- Mobini, R.; Tremaroli, V.; Ståhlman, M.; Karlsson, F.; Levin, M.; Ljungberg, M.; Sohlin, M.; Forslund, H.B.; Perkins, R.; Bäckhed, F.; et al. Metabolic effects of Lactobacillus reuteriDSM 17938 in people with type 2 diabetes: A randomized controlled trial. Diabetes, Obes. Metab. 2016, 19, 579–589. [Google Scholar] [CrossRef]
- Patro-Gołąb, B.; Szajewska, H. Systematic Review with Meta-Analysis: Lactobacillus reuteri DSM 17938 for Treating Acute Gastroenteritis in Children. An Update. Nutrients 2019, 11, 2762. [Google Scholar] [CrossRef] [PubMed]
- Dinleyici, Ener Cagri, PROBAGE Study Group, Yvan Vandenplas, Nazan Dalgic, Sirin Guven, Özge Metin, Makbule Eren et al. "Lactobacillus reuteri DSM 17938 effectively reduces the duration of acute diarrhoea in hospitalised children." Acta Paediatrica 103, no. 7 (2014): e300-e305.
- Wang, G.; Huang, S.; Cai, S.; Yu, H.; Wang, Y.; Zeng, X.; Qiao, S. Lactobacillus reuteri Ameliorates Intestinal Inflammation and Modulates Gut Microbiota and Metabolic Disorders in Dextran Sulfate Sodium-Induced Colitis in Mice. Nutrients 2020, 12, 2298. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zheng, F.; Stanton, C.; Ross, R.P.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus reuteri FYNLJ109L1 Attenuating Metabolic Syndrome in Mice via Gut Microbiota Modulation and Alleviating Inflammation. Foods 2021, 10, 2081. [Google Scholar] [CrossRef] [PubMed]
- Marras, L.; Caputo, M.; Bisicchia, S.; Soato, M.; Bertolino, G.; Vaccaro, S.; Inturri, R. The Role of Bifidobacteria in Predictive and Preventive Medicine: A Focus on Eczema and Hypercholesterolemia. Microorganisms 2021, 9, 836. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, Judith, Moritz V. Warmbrunn, Max Nieuwdorp, and Karine Clément. "Metabolism and metabolic disorders and the microbiome: the intestinal microbiota associated with obesity, lipid metabolism, and metabolic health—pathophysiology and therapeutic strategies." Gastroenterology 160, no. 2 (2021): 573-599.
- Fujimori, S.; Gudis, K.; Mitsui, K.; Seo, T.; Yonezawa, M.; Tanaka, S.; Tatsuguchi, A.; Sakamoto, C. A randomized controlled trial on the efficacy of synbiotic versus probiotic or prebiotic treatment to improve the quality of life in patients with ulcerative colitis. Nutrition 2009, 25, 520–525. [Google Scholar] [CrossRef]
- Vangay, P.; Johnson, A.J.; Ward, T.L.; Al-Ghalith, G.A.; Shields-Cutler, R.R.; Hillmann, B.M.; Lucas, S.K.; Beura, L.K.; Thompson, E.A.; Till, L.M.; et al. US Immigration Westernizes the Human Gut Microbiome. Cell 2018, 175, 962–972.e10. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate Improves Insulin Sensitivity and Increases Energy Expenditure in Mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- Sheng, K.; He, S.; Sun, M.; Zhang, G.; Kong, X.; Wang, J.; Wang, Y. Synbiotic supplementation containing Bifidobacterium infantis and xylooligosaccharides alleviates dextran sulfate sodium-induced ulcerative colitis. Food Funct. 2020, 11, 3964–3974. [Google Scholar] [CrossRef]
- Simon, M.-C.; Strassburger, K.; Nowotny, B.; Kolb, H.; Nowotny, P.; Burkart, V.; Zivehe, F.; Hwang, J.-H.; Stehle, P.; Pacini, G.; et al. Intake of Lactobacillus reuteri Improves Incretin and Insulin Secretion in Glucose-Tolerant Humans: A Proof of Concept. Diabetes Care 2015, 38, 1827–1834. [Google Scholar] [CrossRef]
- Urbańska, M.; Gieruszczak-Białek, D.; Szajewska, H. Systematic review with meta-analysis: Lactobacillus reuteri DSM 17938 for diarrhoeal diseases in children. Aliment. Pharmacol. Ther. 2016, 43, 1025–1034. [Google Scholar] [CrossRef]
- Jang, H.-M.; Lee, K.-E.; Kim, D.-H. The Preventive and Curative Effects of Lactobacillus reuteri NK33 and Bifidobacterium adolescentis NK98 on Immobilization Stress-Induced Anxiety/Depression and Colitis in Mice. Nutrients 2019, 11, 819. [Google Scholar] [CrossRef]
- Ahn, S.-I.; Cho, S.; Jeon, E.; Park, M.; Chae, B.; Ditengou, I.C.P.; Choi, N.-J. The Effect of Probiotics on Intestinal Tight Junction Protein Expression in Animal Models: A Meta-Analysis. Appl. Sci. 2022, 12, 4680. [Google Scholar] [CrossRef]
- Dooling, S.W.; Sgritta, M.; Wang, I.-C.; Duque, A.L.R.F.; Costa-Mattioli, M.; Yildirim, S. The Effect of Limosilactobacillus reuteri on Social Behavior Is Independent of the Adaptive Immune System. mSystems 2022, 7, e0035822. [Google Scholar] [CrossRef]
- Shi, Y.-C.; Cai, S.-T.; Tian, Y.-P.; Zhao, H.-J.; Zhang, Y.-B.; Chen, J.; Ren, R.-R.; Luo, X.; Peng, L.-H.; Sun, G.; et al. Effects of Proton Pump Inhibitors on the Gastrointestinal Microbiota in Gastroesophageal Reflux Disease. Genom. Proteom. Bioinform. 2019, 17, 52–63. [Google Scholar] [CrossRef]
Figure 1.
PRISMA Flow Diagram of Literature Search and Study Selection. This flow diagram details the systematic screening process for studies evaluating the therapeutic effects of Bifidobacterium infantis and Lactobacillus reuteri. The selection involved initial identification of 547 records (plus 30 from other sources), with 56 studies ultimately included in the qualitative synthesis after full-text assessment and duplicate removal. Notes and Abbreviations: The diagram corresponds to the Qualitative Systematic Review (QualSR) methodology. K = number of published records reviewed; n = number of studies included in the final synthesis. The review focuses on metabolic and gut–brain axis outcomes related to keystone species depletion and restoration.
Figure 1.
PRISMA Flow Diagram of Literature Search and Study Selection. This flow diagram details the systematic screening process for studies evaluating the therapeutic effects of Bifidobacterium infantis and Lactobacillus reuteri. The selection involved initial identification of 547 records (plus 30 from other sources), with 56 studies ultimately included in the qualitative synthesis after full-text assessment and duplicate removal. Notes and Abbreviations: The diagram corresponds to the Qualitative Systematic Review (QualSR) methodology. K = number of published records reviewed; n = number of studies included in the final synthesis. The review focuses on metabolic and gut–brain axis outcomes related to keystone species depletion and restoration.
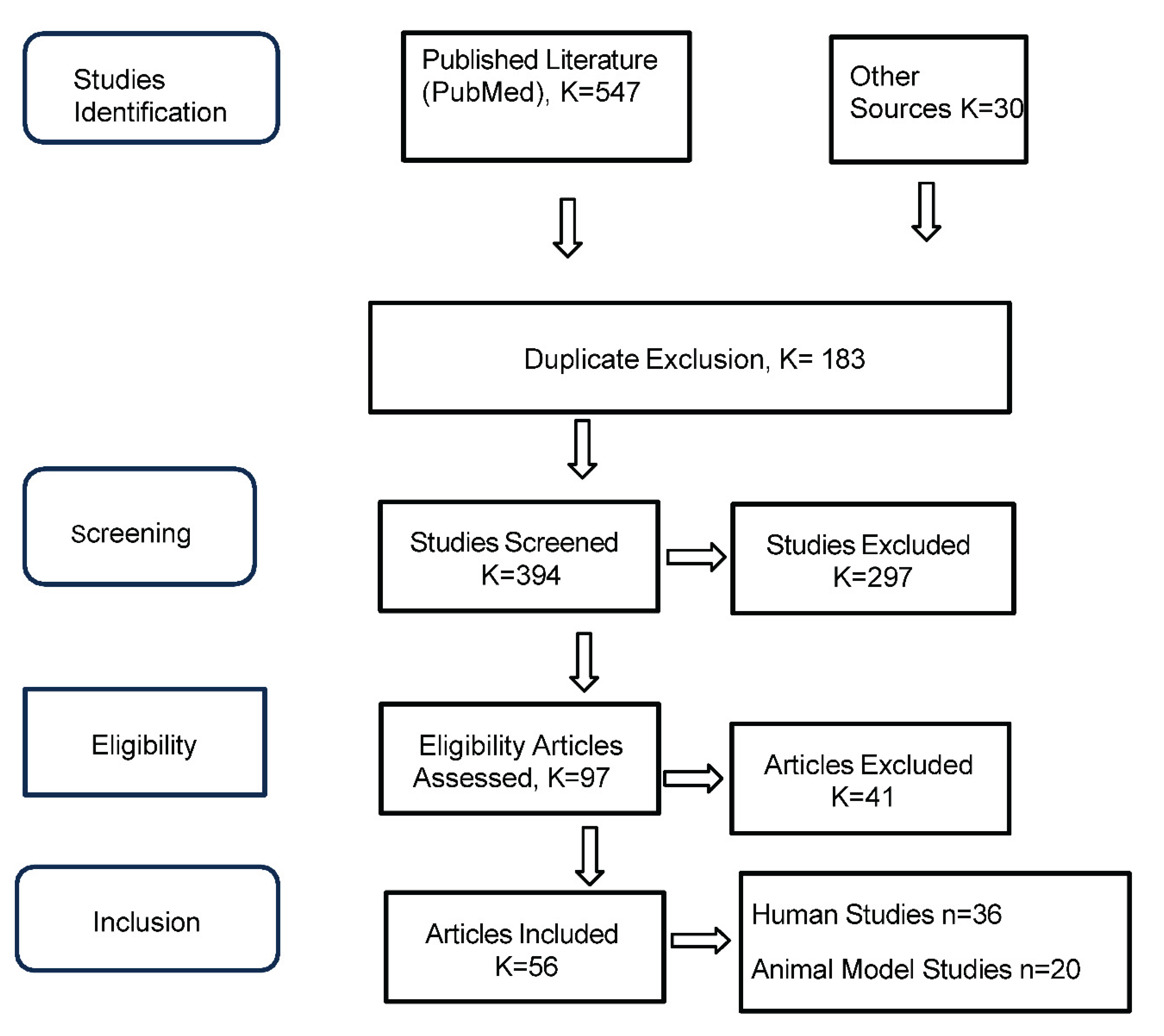
Figure 2.
Mechanistic Pathways of Action for Bifidobacterium infantis and Lactobacillus reuteri. This schematic illustrates the integrated biological pathways through which B. infantis and L. reuteri exert their effects across immune, metabolic, and neurobehavioral domains. Mechanisms include SCFA production, tight junction modulation, vagus nerve signaling, and neurotransmitter synthesis. The diagram also highlights strain-specific functions such as HMO fermentation by B. infantis and reuterin production by L. reuteri, along with their downstream impact on inflammation, glucose regulation, gut–brain axis communication, and clinical outcomes across conditions such as IBD, metabolic syndrome, type 2 diabetes, and mood disorders. Notes and Abbreviations: SCFA = short-chain fatty acids; HMO = human milk oligosaccharides; GLP-1 = glucagon-like peptide-1; Treg = regulatory T cell; sIgA = secretory immunoglobulin A; IL-10 = interleukin-10; TGF-β1 = transforming growth factor beta 1; TNF-α = tumor necrosis factor alpha; GABA = gamma-aminobutyric acid.
Figure 2.
Mechanistic Pathways of Action for Bifidobacterium infantis and Lactobacillus reuteri. This schematic illustrates the integrated biological pathways through which B. infantis and L. reuteri exert their effects across immune, metabolic, and neurobehavioral domains. Mechanisms include SCFA production, tight junction modulation, vagus nerve signaling, and neurotransmitter synthesis. The diagram also highlights strain-specific functions such as HMO fermentation by B. infantis and reuterin production by L. reuteri, along with their downstream impact on inflammation, glucose regulation, gut–brain axis communication, and clinical outcomes across conditions such as IBD, metabolic syndrome, type 2 diabetes, and mood disorders. Notes and Abbreviations: SCFA = short-chain fatty acids; HMO = human milk oligosaccharides; GLP-1 = glucagon-like peptide-1; Treg = regulatory T cell; sIgA = secretory immunoglobulin A; IL-10 = interleukin-10; TGF-β1 = transforming growth factor beta 1; TNF-α = tumor necrosis factor alpha; GABA = gamma-aminobutyric acid.
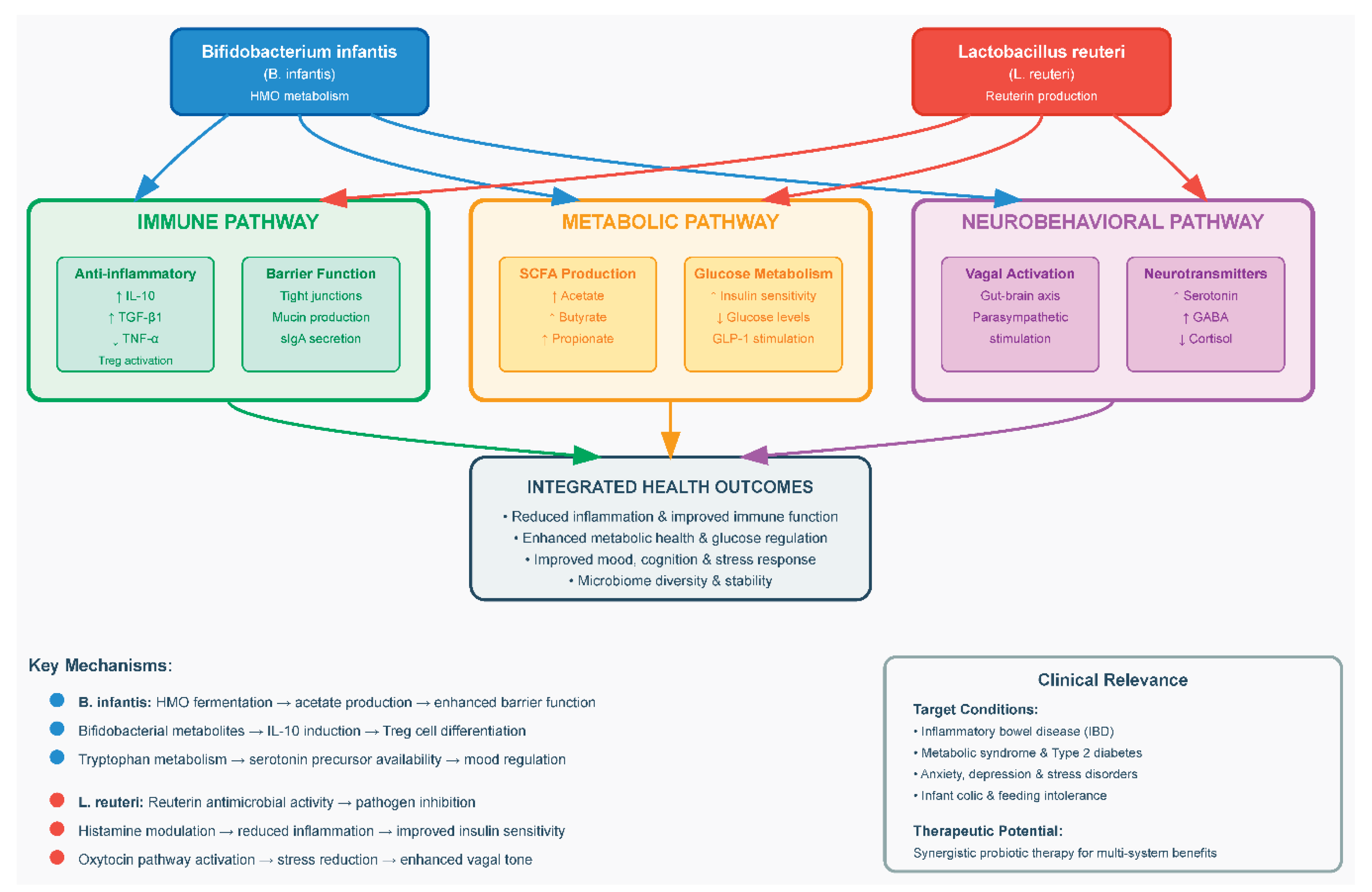
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



