Submitted:
12 August 2025
Posted:
12 August 2025
You are already at the latest version
Abstract
Four Botryosphaeria spp. isolated from apple with dieback symptoms: Diplodia mutila, Diplodia seriata, Neofusicoccum arbuti and Lasiodiplodia theobromae, were subjected to differ-ent conditions to induce sporulation, then freeze-dried and stored in glass vials and am-poules at a temperature of 4 °C using two protective media. Viability was assessed after storage periods of 1, 90, 180 and 365 days. Low-nutrient growth conditions on 2% water agar supplemented with pine needles, incubated under UV light (λ = 350 nm) and at 24 °C, effectively stimulated sporulation of all four Botryosphaeriaceae species. The survival rate of the isolates was similar. Overall viability showed slight but significant differences de-pending on the type of protective medium and storage container used for the freeze-dried cultures (p< 0.001). Among the tested media, the highest viability was maintained in vac-uum-sealed glass ampoules using either a medium containing 20% skimmed milk pow-der with 5% inositol or 20% skimmed milk powder alone.
Keywords:
Botryosphaeria spp.
; dieback
; sporulation stimulation
; freeze-drying
; conidium
; preservation
1. Introduction
Several species of Botryosphaeriaceae including Botryosphaeria, Diplodia, Lasiodiplodia, and Neofusicoccum have been reported to cause Botryosphaeria canker and dieback in apple trees worldwide [1,2,3,4,5,6,7,8,9]. In Chile, Diplodia mutila, D. seriata, Lasiodiplodia theobromae and Neofusicoccum arbuti are the most frequently reported species causing Botryosphaeria canker and dieback in apple orchards [10].
Light has been considered a key stimulus for conidial formation in fungi. Exposure of mycelia to ultraviolet (UV) light has been a widely used method [11,12,13,14]. UV light irradiation has successfully induced sporulation in Ascomycetes species [15], Basidiomycetes [16], Myxomycetes [17] and Zygomycetes [18]. Wavelengths between 350 and 500 nm have been reported to be the most effective for promoting sporulation [12]. However, different isolates may exhibit different threshold doses under specific wavelengths, while excessive exposure can inhibit sporulation [19,20]. Although the mechanism by which light induces fungal conidiation is not yet fully understood. Dahlberg and Etten el al. [12] suggested that photoreceptors might influence the electron transport system, affecting pH, cellular ionic balance, and glucose and carbohydrate metabolism. Several photosensitive proteins have been identified during fungal conidiation [18,21,22,23], for instance, White Collar-1 and Vivid proteins, have been recognized as blue light photoreceptors that mediate light induction of rhythmic conidiation in Neurospora crassa [24,25,26]. Furthermore, transcription factors such as BLR-1 and BLR-2 are essential in the photoconidiation process in Trichoderma atroviride [27].
Fungi are notoriously difficult to maintain under good conditions, as they are inherently variable [28]. Methods of preserving fungal strains as research tool require that stored cultures remain viable over time, without undergoing morphological or physiological changes. Among the various methods used for long-term storage of fungal cultures, some have been evaluated as more effective than others depending on the species or genus [29]. When preserving fungi, several factors must be considered, including economic aspects. Common methods include preservation under mineral oil or paraffin [30], in sterile water [30,31], in grain or soil at room temperature [30], in silica gel [30,32] and freezing at -20°C or -80°C [33]. However, it should be noted that cryopreservation at –20 °C is not recommended whereas, but preservation in liquid nitrogen (−196 °C) is considered an optimal method for long-term storage of fungal cultures due to the stability of secondary metabolite production and minimal genetic variations. Although, certain fungi have shown significant polymorphism after recovery [34,35], the cryopreservation is particularly suitable for the storage of valuable stock cultures, such as proprietary isolates [36].
Freeze-drying, or lyophilization, is a technique widely used by culture collections since it offers several advantages for the preservation of fungal cultures. For instance, it enables long-term storage stability, allowing samples to remain viable for decades, typically between 20 and 40 years, without significant viability loss. Freeze-dried materials can often be stored at room temperature, although at temperatures between 4 to 18 °C is more suitable. The removal of moisture limits the risk of contamination and inhibits metabolic activity that could compromise the integrity of the preserved organisms, at the same time, ensuring high viability and genetic stability upon rehydration. Another aspect to consider is that the resulting dried material is easy to handle and transport, which facilitates the exchange of cultures between laboratories and culture collections worldwide [36,37,38]. However, it has certain limitations, such as the time needed for recovery of the preserved isolate, to maintain optimal viability without sublethal damage, requires the implementation of rigorous and controlled protocols, so the selection of an appropriate protective suspension medium, the optimal cooling rate, the cooling method, the temperature and speed of drying and the final residual moisture content are critical to reduce damage to the fungal cell [28]. In this process, generally, only fungal conidia survive [28,35]. Skimmed milk is frequently used as a protective agent during freeze-drying as it helps preserving the structural integrity of cell walls. Components of skimmed milk, such as amino acid, disaccharides and calcium, create a protective layer around cells, which prevents damage caused by ice crystal formation during freezing and reduces stress during the drying phase. This protective effect is essential to maintain high cell viability, and the survival rate can be significantly improved compared to other protective agents [39,40,41]. Inositol is also used as a protective agent in mushroom freeze-drying due to its ability to stabilize cell walls during the freezing and drying process. This stabilization is crucial to preserve cell viability, as inositol acts to maintain the structural and functional integrity of cells by preventing ice crystal formation and other damage associated with the freeze-drying process [43]. Currently, freeze-drying remains one of the most effective methods for maintaining viable and stable fungal cultures for long periods. Numerous species of phytopathogenic fungi have been successfully preserved by freeze-drying [43,44,45]. It should be noted that no preservation method can be universally applied to all fungi and that intraspecific variability makes it impossible to apply standard protocols, even at the species level [28]. In particular, for Botryosphaeria spp., in addition to freeze-drying and cryopreservation in liquid nitrogen, the use of agar plug storage in mineral oil has been reported, which allowed the viability of unidentified Botryosphaeria spp. to be preserved for 31 years, Botryosphaeria obtusa for 20 years and Botryosphaeria ribis for 6 years [30] (M. Fletcher, ICMP Landcare Research, Auckland, personal communication). Culture viability is a key indicator of success in fungal preservation methods. Although preservation of some Botryosphaeriaceae spp. has been described, the preservation by freeze-drying of species isolated from apple trees with dieback symptoms such as D. mutila, D. seriata, N. arbuti and L. theobromae has not yet been reported.
This study evaluated the sporulation stimulation conditions and the viability of the freeze-drying conservation method over time, to preserve four Botryosphaeria spp. that affect apple trees, causing dieback and canker [10]. Maintaining the viability of fungal cultures is crucial to advance research on the mechanisms of action, fungicide resistance and pathogenicity of these phytopathogens.
2. Materials and Methods
2.1. Study Location
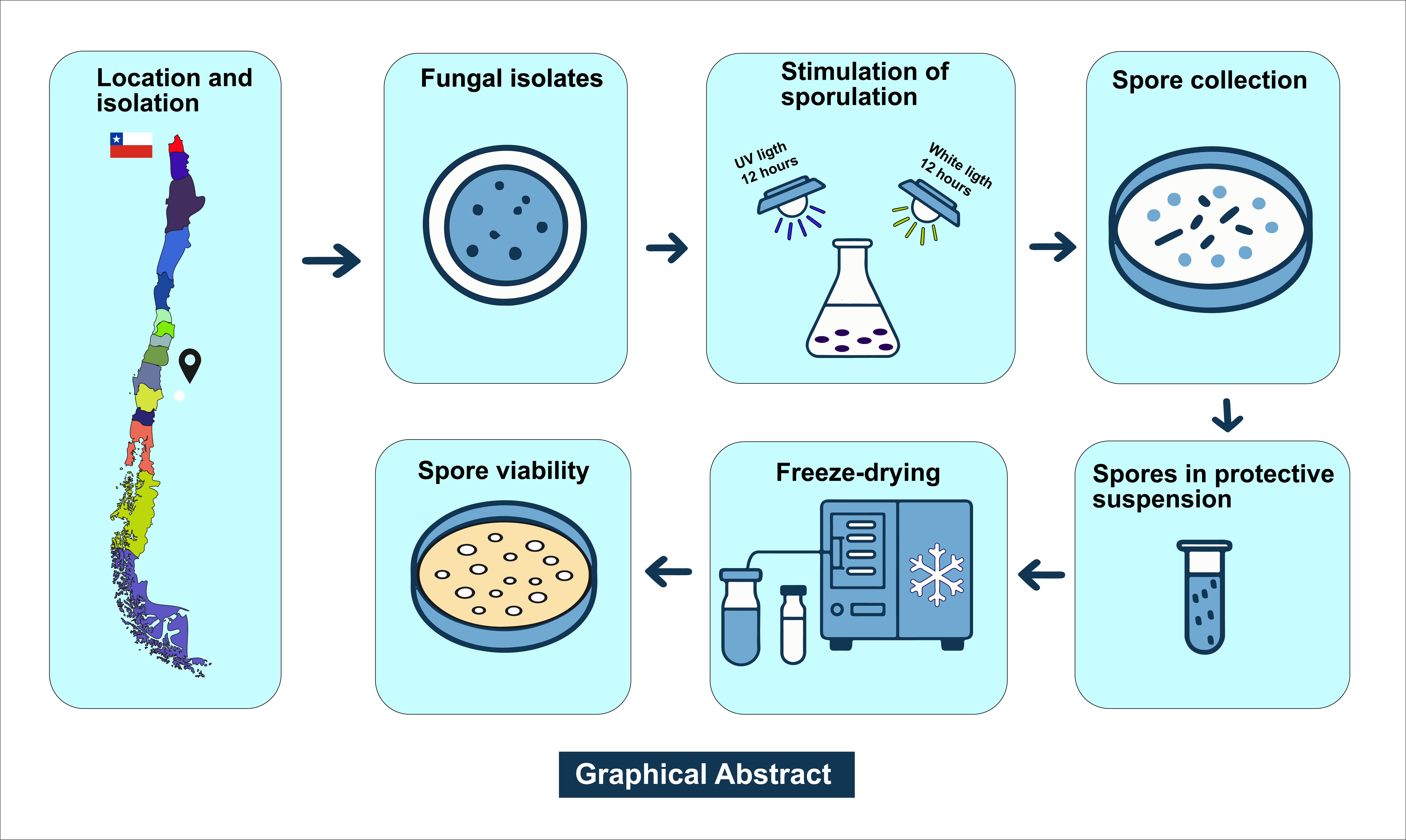
The experiments to stimulate conidial production were carried out at the Fruit Pathology Laboratory of the University of Talca, Chile during 2023. The freeze-drying tests were carried out at the Microbial Genetic Resources Bank (BRGM), located at the Agricultural Research Institute (INIA) Quilamapu, Chillán, Chile in the same year.
2.2. Isolation and Molecular Identification
In studies carried out by Díaz et al. [10], meticulous procedures for isolation, morphological characterization, molecular identification, and phylogenetic analysis of D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), L. theobromae (Bot-2018-LT45) and N. arbuti (Bot-2018-NA32) were performed. The isolation began with the collection of samples of infected plant tissue, which were subjected to a surface disinfection process and subsequent cultivation in selective media to obtain pure cultures [10]. The isolates obtained were morphologically characterized by observing the macroscopic characteristics of the colonies and the micromorphology of the conidia. For molecular identification, genomic DNA was extracted from the isolates using standardized commercial extraction protocols, and specific gene regions were amplified through polymerase chain reaction (PCR) [10]. In particular, molecular markers such as the internal transcribed spacer (ITS) region, translation elongation factor 1-α (tef1), and β-tubulin (tub2) were used. The amplified products were sequenced and the obtained sequences were compared by BLAST analysis against public databases, such as GenBank, for identification. Finally, phylogenetic analyses were performed to determine the evolutionary relationships between the isolates and other known members of the genus. These analyses combined molecular and morphological data, providing a robust and detailed identification of D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), L. theobromae (Bot-2018-LT45) and N. arbuti (Bot-2018-NA32) [10]. It was then maintained in Acidified Potato Dextrose Agar (APDA) medium and stored in refrigeration at 4 ◦C until their use in the fungal collection of the Fruit Growing Laboratory of the University of Talca. Each isolate was recovered by seeding a mycelial plug (5 mm diameter) in 0.1% PDA medium plus 0.01% tetracycline hydrochloride) incubated at 25 ± 2ºC for 10 days (Supplement Figures S1, S2). The isolates of D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), L. theobromae (Bot-2018-LT45), and N. arbuti (Bot-2018-NA32) were deposited as live cultures at the Chilean Collection of Microbial Genetic Resources in Chillán, Chile (www.cchrgm.cl), under the codes RGM 3110, RGM 3114, RGM 3136, and RGM 3138 respectively.
2.3. Conidial Production
Different growth conditions and media were evaluated to stimulate sporulation and conidial production in fungi of the Botryosphaeriaceae family, such as D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45), isolated from apple wood. Multiple growth conditions were considered, including exposure to temperature ranges between 5 and 24 ºC. The type of growth medium and exposure to ultraviolet (UV) light at λ = 350 nm were evaluated. The media evaluated included potato dextrose agar (PDA), 2% agar-water (A-A), PDA with the addition of pine needles, PDA with the addition of apple chips, PDA with the addition of grapevine chips, A-A with the addition of pine needles, A-A with the addition of apple chips, A-A with the addition of grapevine chips, A-A in slant and PDA in a 350 mL flask (Supplementary Table S1). UV light exposure was performed in a dark chamber using a UV lamp at λ = 350 nm for periods of 15 days continuous. Subsequently, the mature pycnidia from each treatment under a flow chamber were collected and crushed using sterile ceramic mortars in 20 mL of sterile water. The solution with conidia was then placed in 20 mL falcon tubes and subjected to a centrifugation cycle of 5 min at 6 ºC at 6000 rpm. The number of conidia was quantified in a Neubauer chamber.
2.4. Spore Collection, Protective Suspension Media, and Freeze-Drying
Conidia were obtained from cultures in 2% agar-water petri dishes where autoclaved pine needles were placed and this incubated at 24 °C under a 12 h/12 h light/dark cycle for 10 days for D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), L. theobromae (Bot-2018-LT45), and N. arbuti (Bot-2018-NA32). Conidia production was induced at 24 °C for 15 days under ultraviolet light (λ = 350 nm) (12 h light UV/12 h light white). A total of 20 petri dishes with 10, 60 mm pine needles per dish were used for each Botryosphaeriaceae spp. Mature pycnidia were collected under a laminar flow hood and ground in 50 mL of sterile water using sterile ceramic mortars. The solution with conidia was then sieved with a 100-micron filter and then subjected to a centrifugation cycle of 10 min at 6 ºC at 3000 rpm. The supernatant was removed and conidia concentrate were then resuspended with two protectives solutions. the first consisted of skimmed milk powder in water at 20% and the second solution was prepared with a mixture skimmed milk powder at 20% plus 5% inositol, both solutions were subsequently subjected to autoclave cycle at 120ºC for 5 min. Approximately 0.5 ml of each suspension was added into two types of sterile containers, 2 ml. glass vials and glass ampoules. The vial and glass ampoules were placed in a first cooling phase at a temperature of -20 ºC for two hours. They were then placed on the racks of a primary lyophilizer (Biobase BK-FD12PT) under operating conditions of -60 ºC, vacuum pressure of 0.04 bar. The suspensions were dried for 15 h which left a residual moisture content of the dried suspension at 1-2 %. The chamber was then brought to atmospheric pressure and the vials and ampoules were removed. The group of containers, which were made up of glass vials, were then vacuum-sealed immediately after the freeze-drying process. However, the ampoules were fitted with a cotton plug, followed by a silica bead suspended above the sterilized cotton plug. The glass ampoules were then contracted approximately 1 cm above the top of the cotton plug and silica bead using a hydrogen-oxygen torch. This stage was carried out as quick as possible to minimize the material to be exposed to atmospheric oxygen and water vapour from the atmosphere and these factors must be kept to a minimum; overexposure can cause deterioration. The glass ampoules were then placed in the manifold of a freeze-dryer (CRIST 1-4 LSC basic) for a secondary drying phase at working conditions of 0.04 bar and -55ºC for 8 hours. The ampoules were vacuum sealed using an air/gas torch [36]. Both vials and ampoules were stored at 4 ºC.
2.5. Spore Viability
The viability of vials and ampoules containing freeze-dried Botryosphaeriaceae spp was assessed after 1, 30, 180, and 365 days of preservation. The samples were placed at room temperature for two hours to assess their viability. Under a flow chamber, both types of containers were uncovered and reconstituted with 1000 µL of a 20% skimmed milk powder solution in each container, allowing 15-20 minutes for the spores to absorb moisture. The viability rate of freeze-dried Botryosphaeriaceae spp. was quantified by serial dilutions (10⁻¹ to 10⁻⁷) in PDA. The strains were incubated at 24 ºC for 24 hours. The colony-forming units (CFU) produced by viable conidia were counted using a Nikon electronic magnifying glass. The percentage reduction in viability (%RV) was calculated using the formula CFUt = CFU count at storage time t (days). CFU0 = initial colony forming unit (CFU) count on day 1 (control). The percentage viability (RV) was analyzed using multifactorial ANOVA (p=0.01) to identify significant effects of temperature, culture medium, supplements, UV, and their interactions. When overall differences were detected (p<0.0001), Scheffé's post hoc test (p<0.0001) was applied for multiple comparisons between all experimental groups (including combinations of conditions).
2.6. Data Availability
The complete nucleotide sequence sets of DNA sequence for the ITS, tef1-α, and tub2 regions was been deposited in GenBank under accession numbers: Diplodia mutila (MW560102, MW591887, MW574060), Diplodia Seriata (MH675471, MH908100, MH745086), Lasiodiplodia Theobromae (MW560112, MW591897, MW574070) and Neofusicoccum arbuti (MW560114, MW591899, MW574072) [10].
3. Results
3.1. Conidia Production
ANOVA analysis of variance showed that sporulation was significantly higher (p-value <0.0001) in the four Botryosphaeriaceae spp. in 2% agar-water medium (W-A) in Petri dishes (PP), with the addition of apple chips at a growth temperature of 24 ºC and with a 15-day exposure to light (UV) at λ = 350 nm, followed by (W-A) in petri dishes (PD) with the addition of grapevine chips under the same temperature and light (UV) conditions as the most significant treatment. (W-A) in petri dishes (PD) with the addition of pine needles did not show significant differences with the second-best treatment and under the same temperature and light (UV) conditions (Figure 1 and Figure 2) (Supplementary Table S1).
3.2. Lyophilization and Spore Viability
The freeze-drying process was evaluated using four Botryosphaeriaceae spp., D. mutila, D. seriata, L. theobromae and N. arbuti. The conidia of the described species were placed in two types of protective media before their freeze-drying process: the first, composed of a 20% skimmed milk powder solution in distilled water, and the second consisting of 20% skimmed milk powder in distilled water supplemented with 5% inositol. The survival and viability results were evaluated after 1, 30, 180 and 365 days after the freeze-drying process. Using the two types of protective media, it was observed that the four Botryosphaeriaceae spp. evaluated survived successfully. However, significant differences in long-term viability were observed between the two types of protective media used after an ANOVA analysis of variance (p-value <0.0001) and a Scheffé statistical test (Supplementary Table S2a, 2b). After 1 day of freeze-drying, the four Botryosphaeriaceae spp. freeze-dried using the two types of protective media described showed similar viability, indicating that both 20% skimmed milk powder alone and the combination with 5% inositol provided adequate initial protection. However, after 3 months, slight differences in the viability rate began to be observed. In the group treated with inositol, N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45) showed greater stability and a smaller decrease in viability compared to the group that only used 20% skimmed milk. This trend was maintained and was accentuated at 180 and 365 days, where the viability of N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45) in the group with inositol was slightly higher. D. mutila (Bot-2017-DM21) and D. seriata (Bot-2018-S3) showed similar viability values using the two types of protective media after 1, 30, 180 and 365 days after the freeze-drying process. In addition to the evaluation of the protective medium, two storage methods were compared: glass vials and glass ampoules. It was observed that the viability of Botryosphaeriaceae spp. was slightly lower in glass vials compared to glass ampoules, indicating that the fully airtight and vacuum sealing of the storage container also influenced the long-term survival of freeze-dried Botryosphaeriaceae spp. (Figure 3 and Figure 4) (Supplementary Table S2a, 2b, Supplementary Figures S3, S4).
4. Discussion
Fungi of the genus Botryosphaeria are important pathogens that affect a wide variety of fruit trees, causing diseases such as branch dieback and canker on trunks and main branches. These diseases are particularly devastating in high-value fruit trees, such as apple, pear, citrus, and grapevine, among others, generating significant losses in global fruit production [3,10]. The preservation of plant pathogenic fungi by freeze-drying is essential for scientific research and disease management. This method has allowed fungi to be stored in an inactive state, maintaining their viability and genetic characteristics in the long term without the need for continuous refrigeration, which compared to the cryopreservation method is less expensive and carries less risks of loss due to storage failures [36]. Freeze-drying can ensure the genetic purity of strains, which is essential for epidemiology studies, pathogenicity testing, and fungicide resistance evaluation. In addition, it facilitates the exchange of strains between institutions and laboratories, ensuring the availability of biological material for global research and the development of new disease control strategies. Considering the effectiveness of the freeze-drying method in the long-term preservation of plant-sporulating pathogenic fungi, the objective of this work was to establish the appropriate conditions to stimulate conidia production and evaluate the viability and survival for their conservation by the freeze-drying method in the species D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), N. arbuti (Bot-2018-NA32), and L. theobromae (Bot-2018-LT45) isolated from apple trees with dieback and canker symptoms in the Maule region, Chile.
The lack of sporulation of many fungi in culture is a problem frequently faced by mycologists and plant pathologists. This is particularly true in fungal identification where the difficulty of identifying non-sporulating colonies is a major obstacle to developing adequate routine detection procedures or failing that for their use in laboratory and field experiments and especially for their preservation by cryopreservation and freeze-drying. Our study revealed that exposure to UV light (λ = 350 nm) of cultures of Botryosphaeriaceae spp. in 2% water agar alone was not sufficient to induce sporulation of the fungal species tested. Similarly, the addition of wood (pine needles, apple and grape wood chips) to the culture medium without the presence of UV light did not produce significant sporulation. However, when both treatments were applied together (UV light (λ = 350 nm) and the addition of wood), robust sporulation was observed in D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45). This suggests that the lignin contained in wood could be acting as an inducer or sensitizer that, when interacting with UV radiation, activates certain metabolic or signalling pathways that are essential for cell differentiation and spore production [46,47]. Lignin is a complex and abundant polymer in nature, mainly in the cell walls of vascular plants such as apple, and its presence in the culture medium could be simulating natural environmental conditions that favour sporulation. In addition, UV radiation is known for its ability to generate oxidative stress, which can induce defence responses in fungi, including the production of reproductive structures such as conidia. The combination of both factors could be triggering an adaptive response in these fungal species, promoting sporulation as a survival strategy under adverse conditions [48,49]. Exposure to low temperatures did not influence sporulation or conidia production in D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45). This indicates that, unlike other fungi whose sporulation rates can be highly influenced by low temperatures [12,50,51], the Botryosphaeriaceae spp. evaluated in this study do not require a specific temperature to complete their sporulation cycle, as long as the other two key factors (UV light and lignin) are present. This observation may be related to the ability of these Botryosphaeriaceae spp. to adapt to diverse climates and environmental conditions, which reinforces their phytopathogenic character and their wide range of hosts in different geographical regions [52]. It should be noted that the sporulation of Botryosphaeriaceae spp. was higher when poor culture media, composed only of agar and water, were used compared to rich media such as PDA (Potato Dextrose Agar) medium. This observation suggests that a nutritious culture medium could be inhibiting or delaying the sporulation of these species. Under conditions of nutritional richness, fungi could prioritize vegetative growth over asexual reproduction, which would explain the lower production of conidia in media such as PDA. On the contrary, in a poor culture medium, the lack of essential nutrients could act as a stimulus that induces a stress response, promoting sporulation as a dispersal and survival strategy (Supplementary Table S1). Furthermore, our results highlight the importance of simulating minimal and non-enriched environmental conditions for sporulation studies, since fungi in their natural environment face nutritional limitation scenarios.
On the other hand, the use of 20% skimmed milk powder with the addition of 5% inositol as a protective medium in the freeze-drying process showed slight advantages in the survival rate and viability (RV) of conidia of the Botryosphaeriaceae spp. evaluated, compared to the protective medium containing only 20% skimmed milk powder, where a slightly lower viability and survival of conidia was observed after a storage period of 365 days after the freeze-drying process. However, both protective media showed a similar survival rate in all Botryosphaeriaceae spp. This could be because inositol is known for its role in stabilizing cell membranes and preserving the integrity of cellular structures during the freeze-drying process [53]. The percentage of viability reduction (RV) after the freeze-drying process showed that D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45) reduced their viability by 2.8, 2.5, 4.2 and 2.5 % respectively, when using the protective medium composed of 20% skimmed milk powder the addition plus 5% inositol, using glass ampoules as containers and after 365 days of storage. While the protective medium that contained only 20% skimmed milk powder yielded % (RV) of 5.9, 5.5, 5.9 and 3.5 % for D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45), respectively, using glass vials as containers after the same storage time. Overall, it was observed that the protective medium was more effective when conjugated with the addition of 5% inositol, along with the use of glass ampoules as containers, significantly helping to reduce the viability loss. The addition of 5% inositol appeared to improve viability in all four Botryosphaeriaceae spp. This suggests that the addition of additional protective compounds such as inositol may contribute to a better preservation of the Botryosphaeriaceae spp. evaluated by reducing cell damage during dehydration and storage. This improvement was reflected in the higher number of CFU observed in the group treated with the combined medium and in the reduction of viability (RV) (Supplementary Table S2a, 2b). Skimmed milk alone has been widely used as a protective medium due to its protein and carbohydrate content, which can help stabilize fungal cells during the freeze-drying process [28]. However, the results obtained in this study suggest that inositol increases viability when combined with skimmed milk, possibly due to its ability to interact with fungal cell lipids and proteins, minimizing damage caused by the freeze-drying process [54]. Skimmed milk is an ideal lioprotectant, which is inexpensive (standard skimmed milk, available at any grocery store, can also be used). The glass transition temperature (Tg) of frozen skimmed milk is –18 °C, which facilitates freeze-drying, and it contains a mixture of macromolecules (lactalbumin and casein) and a saccharide. The macromolecules serve as filling agents, the amino acids contained in the macromolecules help repair sublethal damage and provide energy during rehydration. The saccharide lactose helps to decrease membrane transitions during dehydration by replacing water dipoles [55,56].
Regarding the type of container used, when comparing glass vials and ampoules as storage methods for the lyophilized product, slight significant differences were observed in the viability of the Botryosphaeriaceae spp. evaluated. Glass ampoules proved to be more effective in preserving viability after 365 days of storage. This finding may be related to the lower permeability of glass ampoules to moisture and air compared to glass vials, which contributed to better protection of the fungi against adverse environmental conditions. Exposure to moisture and temperature fluctuations can negatively affect the stability of lyophilized organisms [57]. Glass ampoules, by offering a more airtight seal, may have reduced these risks, resulting in greater preservation of the viability of the Botryosphaeriaceae spp. evaluated. The use of glass ampoules as a storage method proved to be a more robust container with better sealing properties and provided better protection during long-term storage. In contrast, glass vials presented a less efficient seal, allowing the entry of moisture and air, which may have compromised the viability of the lyophilized Botryosphaeriaceae spp. in this study.
In summary, this study demonstrated that the most effective conditions to induce sporulation in the evaluated Botryosphaeriaceae spp. were through their development in a poor agar-water culture medium with the addition of wood, under exposure to ultraviolet (UV) light at a temperature of 24 ºC. It also demonstrated that during the freeze-drying process of Botryosphaeriaceae spp. the most effective protective medium was a skimmed milk solution supplemented with inositol, which was shown to maintain viability after long storage periods. This study is the first report describing specific methods to stimulate sporulation in Botryosphaeriaceae spp. and its subsequent preservation by freeze-drying process using isolates obtained from apple trees with dieback and canker symptoms in Chile and underlines the need for additional research to validate these results and explore the underlying mechanisms in future investigations.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Colony and conidial morphology of Botryosphaeriaceae species obtained from Botryosphaeria canker and dieback on apple trees in central Chile; S2: One of the seven most parsimonious trees inferred from maximum parsimony phylogeny of Botryosphaeriaceae species according to concatenated alignments of sequences of the internal transcribed spacer (ITS), portion of translation elongation factor 1-α (tef1) and portion of the beta tubulin (tub2) gene regions; S3: Glass ampoules (G.A.) vacuum sealed containing freeze-dried Botryosphaeriaceae spp.; S4: Glass vials (G.V.) containing lyophilized Botryosphaeriaceae spp. Table S1: . Culture media and stimulation conditions for conidia production in Botryosphaeriaceae spp. isolated from apple twigs with dieback and canker symptoms. Table S2a, S2b: Viability and survival of D. mutila (Bot-2017-DM21), D. seriata (Bot-2018-S3), N. arbuti (Bot-2018-NA32) and L. theobromae (Bot-2018-LT45) after the freeze-drying process over time.
Author Contributions
Conceptualization, A.V.-T., and G.A.D.; methodology, A.V.-T.; formal analysis, A.V.-T., M.E.G., M.G., J.F.C., S.A.C. and G.A.D.; investigation, A.V.-T., M.E.G., M.G., J.F.C., S.A.C. and G.A.D.; data curation, A.V.-T., M.E.G., M.G., J.F.C., S.A.C. and G.A.D.; writing: preparation of the original draft, A.V.-T.; writing: review and editing, A.V.-T., M.E.G., M.G., J.F.C., S.A.C. and G.A.D.; supervision A.V.-T.; project administration, A.V.-T., and G.A.D. All authors have read and agreed to the published version of the manuscript.
Funding
Work carried out as part of the PhD program in Agricultural Sciences at the Universidad de Talca, Chile, funded by the Faculty of Agricultural Sciences of the Universidad de Talca and the National Agency for Research and Development of Chile ANID, scholarship folio 21221384.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The complete nucleotide sequence sets of DNA sequence for the ITS, tef1-α, and tub2 regions was been deposited in GenBank under accession numbers: D. mutila (MW560102, MW591887, MW574060), D. Seriata (MH675471, MH908100, MH745086), L. Theobromae (MW560112, MW591897, MW574070) and N. arbuti (MW560114, MW591899, MW574072).
Acknowledgments
This study was funded by the UNIVERSIDAD DE TALCA and the FONDECYT number 1210109 of the National Agency for Research and Development (ANID), Government of Chile. Dr. Adrián Valdez-Tenezaca thanks the University of Talca for the doctoral scholarship, and the ANID for the doctoral scholarship folio 21221384 granted to carry out his postgraduate studies. We thank the Microbial Genetic Resources Bank (BRGM), located at the Institute of Agricultural Research (INIA) Quilamapu, Chillán, Chile. The authors acknowledge the FONDEQUIP Program from the Chilean National Agency for Research and Development (ANID) (grant: EQM200205) for funding a platform of equipment for the preservation of microbial genetic resources. We thank Prof. Manuel Pariguana of the Bio and Nanomaterials Laboratory, Faculty of Health Sciences (FCS), University of Talca for his unconditional support. We also thank the Fruit Pathology Laboratory of the University of Talca.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Latorre, B.A.; Toledo, M.V. Occurrence and relative susceptibility of apple cultivars to Botryosphaeria canker in Chile. Plant Dis. 1984, 68, 36–39. [Google Scholar] [CrossRef]
- Brown-Rytlewski, D.E.; McManus, P.S. Virulence of Botryosphaeria dothidea and Botryosphaeria obtusa on apple and management of stem cankers with fungicides. Plant Dis. 2000, 84, 1031–1037. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: diversity, ecology and impact. Fungal Biol. Rev. 2007, 212, 90–106. [Google Scholar] [CrossRef]
- Phillips, A. J. L.; Lopes, J.; Abdollahzadeh, J.; Bobev, S.; Alves, A. Resolving the Diplodia complex on apple and other Rosaceae hosts. Persoonia. 2012, 29, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Sutton, T. B.; Aldwinkle, H.S.; Agnello, A.M.; Walgenbach, J.F. Compendium of apple and pear diseases and pest. Second edition. American Phytopathological Society Press, 2014. [Google Scholar]
- Abdollahzadeh, J. Diplodia bulgarica, as a new pathogen and potential threat to the apple industry in Iran. Phytopathol. Mediterr. 2015, 54, 128–132. [Google Scholar]
- Delgado-Cerrone, L.; Mondino-Hintz, P.; Alaniz-Ferro, S. Botryosphaeriaceae species associated with stem canker, die-back and fruit rot on apple in Uruguay. Eur. J. Plant Pathol. 2016, 146, 637–655. [Google Scholar] [CrossRef]
- Havenga, M.; Gatsi, G.M.; Halleen, F.; Spies, C.F.J.; van der Merwe, R.; Mostert, L. Canker and Wood Rot Pathogens Present in Young Apple Trees and Propagation Material in the Western Cape of South Africa. Plant Dis. 2019, 103, 3129–3141. [Google Scholar] [CrossRef]
- Martino, I.; Agustí-Brisach, C.; Nari, L.; Gullino, M.L.; Guarnaccia, V. Characterization and pathogenicity of fungal species associated with dieback of apple trees in Northern Italy. Plant Dis. 2024, 108, 311–331. [Google Scholar] [CrossRef]
- Díaz, G.A.; Valdez, A.; Halleen, F.; Ferrada, E.; Lolas, M.; Latorre, B.A. Characterization and pathogenicity of Diplodia, Lasiodiplodia, and Neofusicoccum species causing Botryosphaeria canker and dieback of apple trees in central Chile. Plant Dis. 2022, 106, 925–937. [Google Scholar] [CrossRef]
- Marsh, P.B.; Taylor, E.E.; Bassler, L.M. A guide to the literature on certain effects of light on fungi: reproduction, morphology, pigmentation, and phototropic phenomena. CABI. 1959, 251–312. [Google Scholar]
- Dahlberg, K.R.; Etten, J.L.V. Physiology and biochemistry of fungal sporulation. Annu. Rev. Phytopatho. 1982, 1982. 20, 281–301. [Google Scholar] [CrossRef]
- Betina, V. Photoinduced conidiation in Trichoderma viride. Folia Microbiol. 1995, 40, 219–224. [Google Scholar] [CrossRef]
- Xu, L.L.; Li, F.; Xie, H.Y.; Liu, X.Z. A novel method for promoting conidial production by a nematophagous fungus, Pochonia chlamydosporia AS6.8. World J. Microbiol. Biotechnol. 2009, 25, 1989–1994. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.; Philips, A.J.; Groenewald, J. Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef]
- Miller Jr, O.K. The role of light in the fruiting of Panus fragilis. Can. J. Bot. 1967, 4511, 1939–1943. [Google Scholar] [CrossRef]
- Starostzik, C.; Marwan, W. A photoreceptor with characteristics of phytochrome triggers sporulation in the true slime mould Physarum polycephalum. FEBS Lett. 1995, 370, 146–148. [Google Scholar] [CrossRef]
- Idnurm, A.; Rodríguez-Romero, J.; Corrochano, L.M.; Sanz, C.; Iturriaga, E.A.; Eslava, A.P.; Heitman, J. The Phycomyces madA gene encodes a blue-light photoreceptor for phototropism and other light responses. Proc. Natl. Aca. Sci. 2006, 103, 4546–4551. [Google Scholar] [CrossRef]
- Rakoczy, L. Influence of monochromatic light on the fructification of Physarum nudum. Acta Soc Bot Pol. 1963, 11, 559–562. [Google Scholar]
- Rakoczy, L. Action spectrum in sporulation of slime-mold Physarum nudum Macbr. Acta Soc Bot Pol. 1965, 341, 97–112. [Google Scholar] [CrossRef]
- Corrochano, L.M. Fungal photoreceptors: sensory molecules for fungal development and behaviour. Photochem. Photobiol. Sci. 2007, 6, 725–736. [Google Scholar] [CrossRef]
- Rodriguez-Romero, J.; Hedtke, M.; Kastner, C.; Müller, S.; Fischer, R. Fungi, hidden in soil or up in the air: light makes a difference. Annu. Rev. Microbiol. 2010, 641, 585–610. [Google Scholar] [CrossRef]
- Ruger-Herreros, C.; Rodríguez-Romero, J.; Fernández-Barranco, R.; Olmedo, M.; Fischer, R.; Corrochano, L.M.; Canovas, D. Regulation of conidiation by light in Aspergillus nidulans. Genetics. 2011, 1884, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Froehlich, A.C.; Liu, Y.; Loros, J.J.; Dunlap, J.C. White Collar-1, a circadian blue light photoreceptor, binding to the frequency promoter. Science. 2002, 297, 815–819. [Google Scholar] [CrossRef]
- He, Q.; Cheng, P.; Yang, Y.; Wang, L.; Gardner, K.H.; Liu, Y. White collar-1, a DNA binding transcription factor and a light sensor. Science. 2002, 297, 840–843. [Google Scholar] [CrossRef] [PubMed]
- Schafmeier, T.; Diernfellner, A.C. Light input and processing in the circadian clock of Neurospora. FEBS Lett. 2011, 585, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Arreguín, A.; Pérez-Martínez, A.S.; Herrera-Estrella, A. Proteomic analysis of Trichoderma atroviride reveals independent roles for transcription factors BLR-1 and BLR-2 in light and darkness. Eukaryot. Cell. 2012, 111, 30–41. [Google Scholar] [CrossRef]
- Smith, D. Long-term preservation of test strains fungus. Int. Biodeterior. Biodegrad. 1993, 313, 227–230. [Google Scholar]
- Baskarathevan, J.; Jaspers, M.V.; Jones, E.E.; Ridgway, H.J. Evaluation of different storage methods for rapid and costeffective preservation of Botryosphaeria species. N. Z. Plant Prot. 2009, 62, 234–237. [Google Scholar] [CrossRef]
- Smith, D.; Onions, A.H.S. The preservation and maintenance of living fungi, 2nd edition. IMI Technical Handbook 2. CAB International, Wallingford, Oxon, United Kingdom.
- Burdsall Jr, H.H.; Dorworth, E.B. Preserving cultures of wood-decaying Basidiomycotina using sterile distilled water in cryovials. Mycologia. 1994, 86, 275–280. [Google Scholar] [CrossRef]
- Elliott, M.L. Survival, growth and pathogenicity of Gaeumannomyces graminis var. graminis with different methods of long-term storage. Mycologia.
- Bueno, C.J.; Ambrósio, M.M.; Souza, N.L. Storage of soilborne phytopathogenic fungi. Summa Phytopathol. 2006, 32, 42–50. [Google Scholar] [CrossRef]
- Ryan, M.J.; Jeffries, P.; Bridge, P.D.; Smith, D. Developing cryopreservation protocols to secure fungal gene function. Cryo Lett. 2001, 222, 115–124. [Google Scholar]
- Borman, A.M.; Szekely, A.; Campbell, C.K.; Johnson, E.M. Evaluation of the viability of pathogenic filamentous fungi after prolonged storage in sterile water and review of recent published studies on storage methods. Mycopathologia. 2006, 161, 361–368. [Google Scholar] [CrossRef]
- Smith, D.; Onions, A.H. A comparison of some preservation techniques for fungi. Trans. Br. Mycol. Soc. 1983, 813, 535–540. [Google Scholar] [CrossRef]
- Gallo, M.B.; Guimarães, D.O.; Momesso, L.D.; Pupo, M.T. Natural products from endophytic fungi. Microbiol. Biotechnol. 2008, 4, 39–147. [Google Scholar]
- Ward, K.R.; Matejtschuk, P. Lyophilization of Pharmaceuticals and Biologicals. 2019. Springer New York, New York, NY.
- Nasran, H.S.; Mohd Yusof, H.; Halim, M.; Abdul Rahman, N.A. Optimization of protective agents for the freeze-drying of Paenibacillus polymyxa Kp10 as a potential biofungicide. Molecules. 2020, 2511, 2618. [Google Scholar] [CrossRef] [PubMed]
- Olukotun, G.B.; Adamu, B.B.; Asake, O.J.; Zobeashia, S.S. Preservation of Indigenous Fungal Cultures by Freeze Drying Technique using Skim Milk and Honey as the Protectants. Asian J. Biotechnol. Bioresour. Technol. 2021, 73, 36–42. [Google Scholar] [CrossRef]
- Guerrero-Sanchez, M.; Passot, S.; Campoy, S.; Olivares, M.; Fonseca, F. Effect of protective agents on the storage stability of freeze-dried Ligilactobacillus salivarius CECT5713. Appl. Microbiol. Biotechnol. 2022, 106, 7235–7249. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.S.; Van Ingen, C.W.; Talsma, H.; Van Miltenburg, J.C.; Steffensen, C.L.; Vlug, I.A.; Stalpers, J.A. Freeze-drying of fungi: influence of composition and glass transition temperature of the protectant. Cryobiology. 1995, 32, 60–67. [Google Scholar] [CrossRef]
- Bunse, T.; Steigleder, G.K. The preservation of fungal cultures by lyophilization: Die Konservierung von Pilzkulturen mittels Gefriertrocknung. Mycoses. 1991, 34, 173–176. [Google Scholar] [CrossRef]
- Larena, I.; Melgarejo, P.; De Cal, A. Drying of conidia of Penicillium oxalicum, a biological control agent against Fusarium wilt of tomato. J. Phytopathol. 2003, 151, 600–606. [Google Scholar] [CrossRef]
- Milošević, M.; Medić-Pap, S.; Ignjatov, M.; Milošević, D. Lyophilization as a method for pathogens long term preservation. Zb. Mat. Srp. Prirod. Nauke. 2007, 113, 203–210. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Messner, R.; Gruber, F.; Mach, R.L.; Kubicek-Pranz, E.M. The Trichoderma cellulase regulatory puzzle: from the interior life of a secretory fungus. Enzyme Microb. Technol. 1993, 15, 90–99. [Google Scholar] [CrossRef]
- Hardham, A.R. Cell biology of fungal and Oomycete infection of plants. In Biology of the Fungal Cell. Berlin, Heidelberg: Springer Berlin Heidelberg. 2007, pp. 251-289.
- Leach, C.M. Sporulation of diverse species of fungi under near-ultraviolet radiation. Can. J. Bot. 1962, 401, 151–161. [Google Scholar] [CrossRef]
- Su, Y.Y.; Qi, Y.L.; Cai, L. Induction of sporulation in plant pathogenic fungi. Mycology. 2012, 33, 195–200. [Google Scholar] [CrossRef]
- Cruickshank, I.A.M. Environment and Sporulation in Phytopathogenic Fungi Li. Conidia Formation in Peronospora tabacina Adam as a Function of Temperature. Aust. J. Biol. Sci. 1961, 14, 198–207. [Google Scholar] [CrossRef]
- Zhao, Q.; Shi, Y.; Wang, Y.; Xie, X.; Li, L.; Fan, T.; Li, B. Temperature and Humidity Regulate Sporulation of Corynespora cassiicola That Is Associated with Pathogenicity in Cucumber Cucumis sativus L. Biology. Biology. 2022, 11, 1675. [Google Scholar] [CrossRef] [PubMed]
- Batista, E.; Lopes, A.; Alves, A. What do we know about Botryosphaeriaceae? An overview of a worldwide cured dataset. Forests. 2021, 12, 313. [Google Scholar] [CrossRef]
- Rockinger, U.; Funk, M.; Winter, G. Current approaches of preservation of cells during freeze- drying. J. Pharm. Sci. 2021, 1108, 2873–2893. [Google Scholar] [CrossRef]
- Balla, T. Inositol-lipid binding motifs: signal integrators through protein-lipid and protein-protein interactions. J. Cell Sci. 2005, 118, 2093–2104. [Google Scholar] [CrossRef]
- Crowe, J.H.; Crowe, L.M.; Chapman, D. Preservation of membranes in anhydrobiotic organisms: the role of trehalose. Science. 1984, 223, 701–703. [Google Scholar] [CrossRef]
- Crowe, J.H.; Carpenter, J.F.; Crowe, L.M.; Anchordoguy, T.J. Are freezing and dehydration similar stress vectors? A comparison of modes of interaction of stabilizing solutes with biomolecules. Cryobiology. 1990, 27, 219–231. [Google Scholar] [CrossRef]
- Sinskey, T.J.; Silverman, G.J.; Goldblith, S.A. Influence of Platen Temperatures and Storage Conditions on the Survival of Freeze-dried Salmonella typhimurium. Appl. Microbiol. 1967, 151, 22–30. [Google Scholar] [CrossRef]
Figure 1.
Average conidia production in D. mutila, D. seriata, N. arbuti, and L. theobromae under different substrates and growth conditions. Bars show culture media based on: Water agar (WA) with the addition of apple chips, grape chips, pine needles y PDA medium with pine needles, and Water agar (WA) without supplements. Development temperature was 24 °C and 15 days of exposure to UV light (λ = 350 nm) under a dark chamber. Values with the same letters are not significantly different according to the Tukey test (P value <0.0001).
Figure 1.
Average conidia production in D. mutila, D. seriata, N. arbuti, and L. theobromae under different substrates and growth conditions. Bars show culture media based on: Water agar (WA) with the addition of apple chips, grape chips, pine needles y PDA medium with pine needles, and Water agar (WA) without supplements. Development temperature was 24 °C and 15 days of exposure to UV light (λ = 350 nm) under a dark chamber. Values with the same letters are not significantly different according to the Tukey test (P value <0.0001).
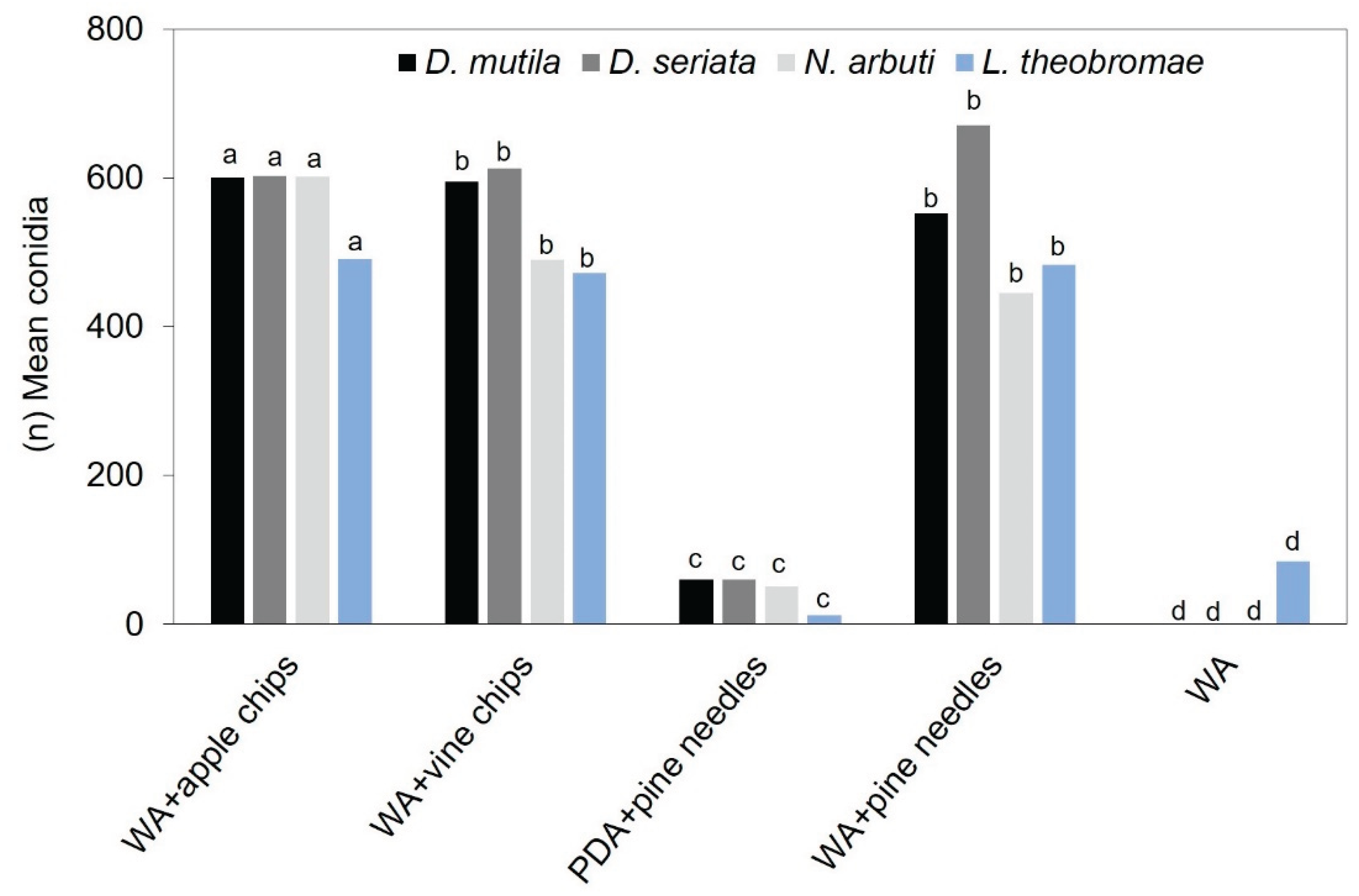
Figure 2.
Conidia produced under different evaluated stimulation parameters. Agar-water medium (W-A) at 2% with the addition of pine needles in petri dishes (P.D), development temperature 24 ºC and 15 days of exposure to light (UV) λ = 350 nm under a dark camera. Microscope (Nikon), 100 and 40× magnifications. (A) D. mutila (Bot-2017-DM21), (B) D. seriata (Bot-2018-S3), (C) N. arbuti (Bot-2018-NA32) and (D) L. theobromae (Bot -2018-LT45). Scale bar = 10 µm.
Figure 2.
Conidia produced under different evaluated stimulation parameters. Agar-water medium (W-A) at 2% with the addition of pine needles in petri dishes (P.D), development temperature 24 ºC and 15 days of exposure to light (UV) λ = 350 nm under a dark camera. Microscope (Nikon), 100 and 40× magnifications. (A) D. mutila (Bot-2017-DM21), (B) D. seriata (Bot-2018-S3), (C) N. arbuti (Bot-2018-NA32) and (D) L. theobromae (Bot -2018-LT45). Scale bar = 10 µm.
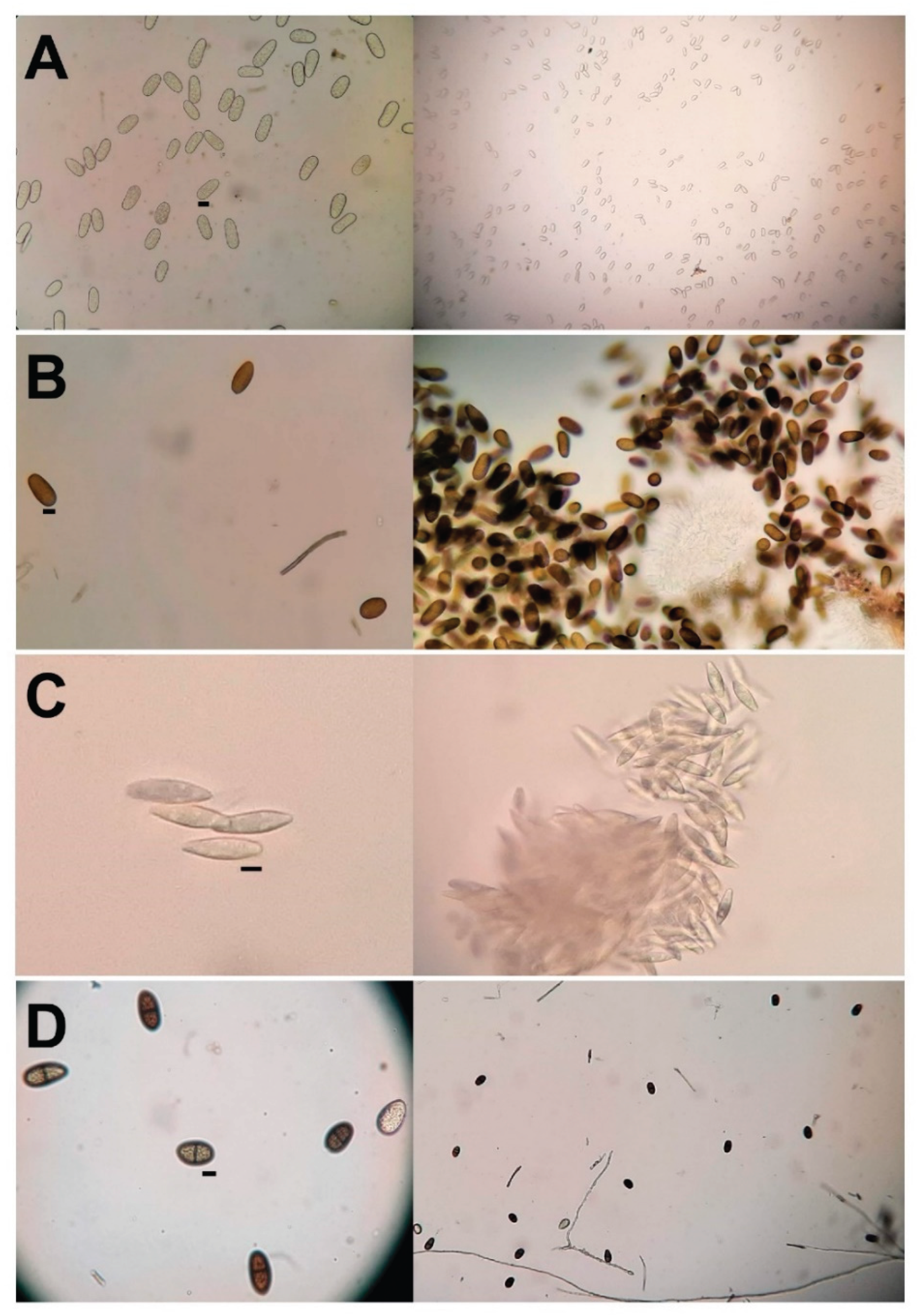
Figure 3.
Viability and survival of D. mutila, D. seriata, N. arbuti and L. theobromae after freeze-drying. The number of colony-forming units (CFU) counted at a 10-4 post-freeze-drying dilution is shown. Type of container used for the freeze-drying process: G.V.= glass vials and G.A.= glass ampoules. Protective medium used for the freeze-drying process. (smp) 20% skimmed milk powder. (smp + inositol) 20% evaporated skimmed milk plus the addition of 5% inositol. Percentage reduction (RV) in viability after the freeze-drying process, over time (after 365 days of storage).
Figure 3.
Viability and survival of D. mutila, D. seriata, N. arbuti and L. theobromae after freeze-drying. The number of colony-forming units (CFU) counted at a 10-4 post-freeze-drying dilution is shown. Type of container used for the freeze-drying process: G.V.= glass vials and G.A.= glass ampoules. Protective medium used for the freeze-drying process. (smp) 20% skimmed milk powder. (smp + inositol) 20% evaporated skimmed milk plus the addition of 5% inositol. Percentage reduction (RV) in viability after the freeze-drying process, over time (after 365 days of storage).
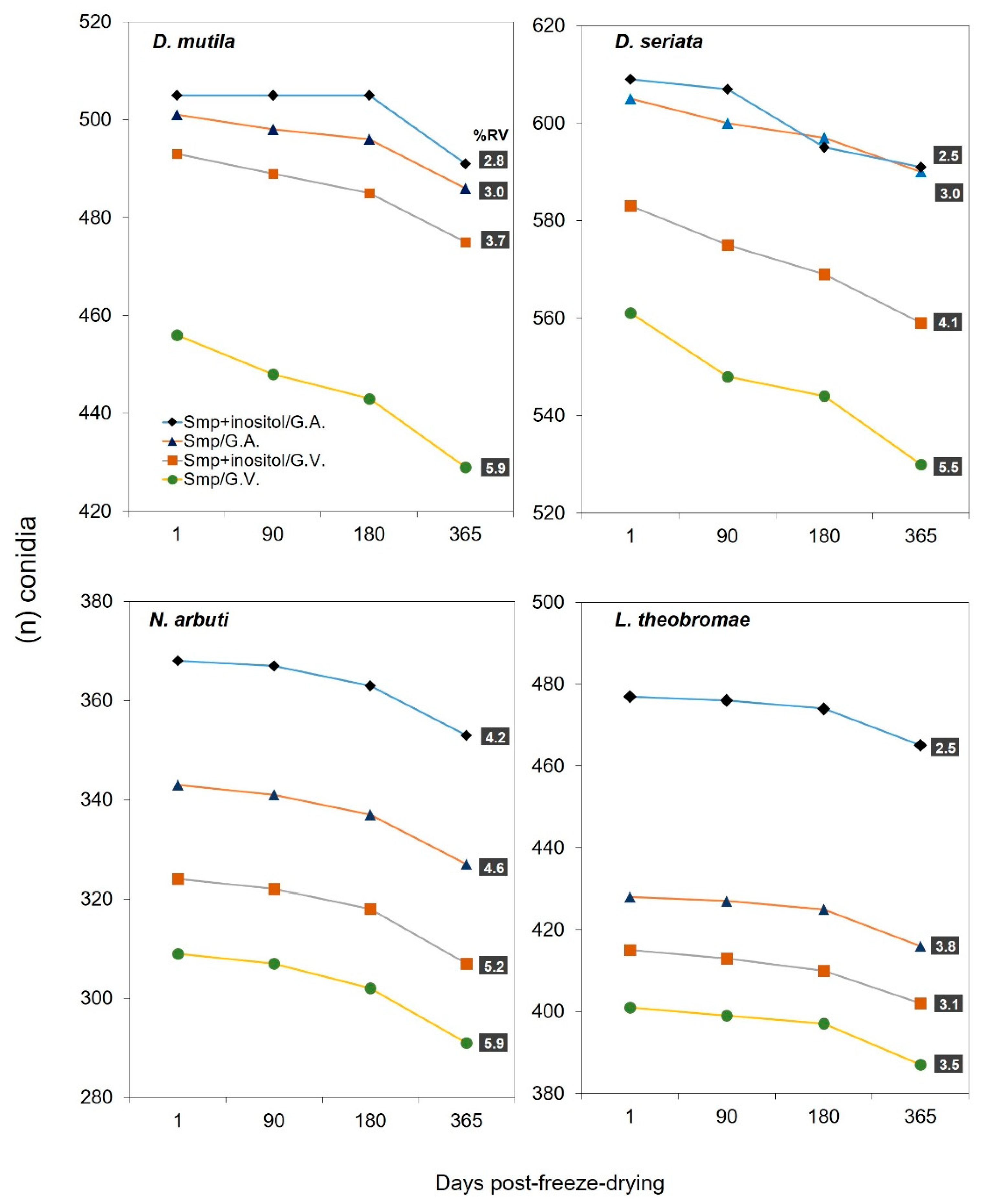
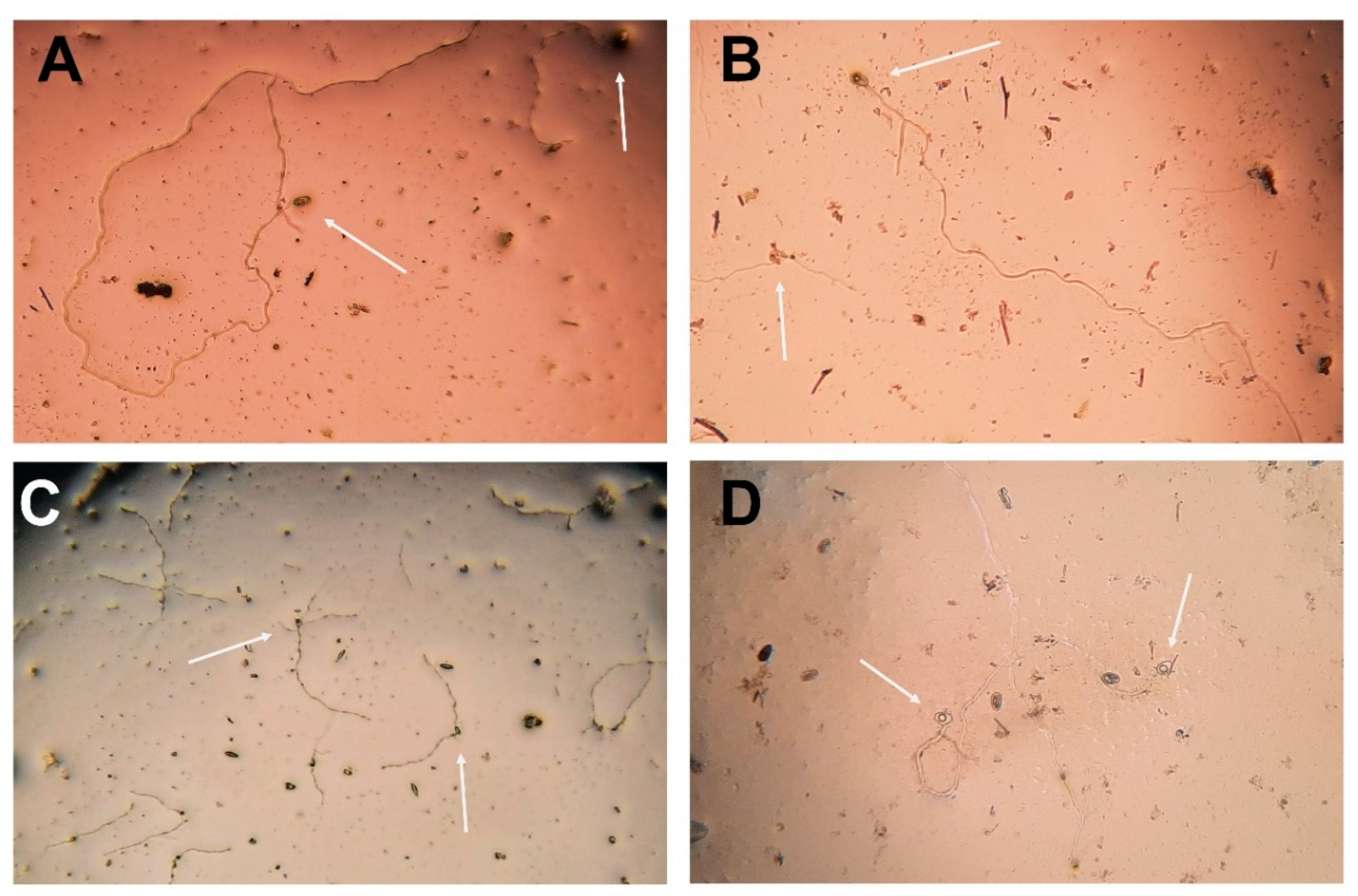
Figure 4.
Conidial germination after 365 days of the freeze-drying process. Conidial viability was assessed in 0.1% PDA medium plus 0.01% tetracycline hydrochloride incubated at 24 ± 2ºC after 24 hours. Arrows indicate conidia, germ tube development, and viability of: (A) D. mutila (Bot-2017-DM21), (B) D. seriata (Bot-2018-S3), (C) N. arbuti (Bot-2018-NA32) and (D) L. theobromae (Bot-2018-LT45). Microscope (Nikon), magnification 40×. .
Figure 4.
Conidial germination after 365 days of the freeze-drying process. Conidial viability was assessed in 0.1% PDA medium plus 0.01% tetracycline hydrochloride incubated at 24 ± 2ºC after 24 hours. Arrows indicate conidia, germ tube development, and viability of: (A) D. mutila (Bot-2017-DM21), (B) D. seriata (Bot-2018-S3), (C) N. arbuti (Bot-2018-NA32) and (D) L. theobromae (Bot-2018-LT45). Microscope (Nikon), magnification 40×. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



