Submitted:
11 August 2025
Posted:
12 August 2025
You are already at the latest version
Abstract
Incretins have a crucial role for postprandial glucose management residing in glucagon-like-peptide 1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP). The incretin role in glucose handling is estimated to 74%, and their absence or reduction being linked to severely impaired glucose control and diabetes. Dairy foods are crucial for adequate nutrition at young age, and with skim milk powder in infant formula, an optimal amount of nutrition is available. Skim milk powder after storage has however in experimental studies been found to reduce islet cell mass of pancreas with an unknown mechanism of action. The aim of this study is to investigate a potential effect of stored skim milk powder on incretin release from intestinal cells.
Intestinal endocrine cells (STC-1 cells) were incubated with fresh and stored skim milk powder, the latter containing 70-fold higher amounts of Maillard reactants. Viability, proliferation and release of incretins were measured after 24 hours.
Both viability and proliferation of the STC-1 cells decreased significantly during incubation with stored skim milk powder. The release of GLP-1 and GIP was suppressed in a dose-dependent way by stored skim milk powder compared to fresh milk powder.
Incretin release from intestinal cells was suppressed by stored skim milk powder containing high amount of Maillard reactants. The clinical significance of this for development of diabetes and obesity awaits further studies.
Keywords:
GLP-1
; GIP
; glucose management
; infant formula
; intestinal endocrine cells
; Maillard reactants
1. Introduction
Infant nutrition requires adequate amounts of fat, protein and carbohydrate as well as microelements. Human breast milk provides a highly nutritious source of food. Infant formula typically made from bovine milk, is an option for mothers that cannot provide breast milk. The production of formulas is highly regulated compared to other food products to ensure optimal quality for the infant nutrition [1]. Milk formula feeding is however at associated with an increased risk of developing necrotizing enterocolitis, respiratory infections, asthma, obesity, diabetes, and inflammatory bowel diseases compared to human breast milk feeding [2]. Therefore, there is a great interest to continuously improve the milk formula to resemble as much as possible human breast milk [3]. Improvements are mainly made by addition of various factors that are being identified in mother´s milk such as special oligosaccharides promoting a wholesome gut microbiota [4]. There are even studies showing that the content of breast milk is adapted to the child and that the child’s gender can play a role for nutrition during early lactation [5].
Our interest concerns the influence of diet composition on glucose management of the body. In this context, incretins are very important as they have a multifactorial effect including insulin secretion, gastric emptying, glucagon response and satiety. All these mechanisms work together, to maintain a normal glucose level.
Infant formula in contrast to fresh breast milk has been demonstrated to contain Maillard reactants, in particular after storage in high temperature and humidity, rendering potential health problems to the infant [6]. In recent studies, we have demonstrated a disruption in growth of the newborn in experimental animal models, who were fed with stored skim milk powder compared to fresh skim milk powder [7]. An increased level of inflammatory markers together with this temporary inhibition of body growth after exposure to stored skim milk powder containing high levels of Maillard reactants were observed [7]. In a following study we found reduced pancreatic beta-cell mass in newborn rats fed with milk powder rich in Maillard reactants compared to those exposed to fresh milk powder [8].
Post-prandial increase of glucose levels are mainly handled by the immediate release of incretins, the two hormones, glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) being responsible for 29% and 45%, respectively of insulin secretion, with glucose itself accounting for only 26% [9]. The question arising is if skim milk powder after storage disturbs the release of these incretins.
In this study we have therefore investigated the release of incretins from an intestinal endocrine cell line following treatment with stored skim milk powder containing elevated amounts of Maillard reactants.
2. Materials and Methods
2.1. Preparation of Milk Powder
Dried skim milk powder was obtained from Semper (Hero Group, Lenzburg, Switzerland). The milk powder was either used in its original form or after treatment for 28 days in a humid atmosphere at 45°C. For experiments, milk was prepared from milk powder according to the recipe on the package i.e. 0.96 g of milk powder was dissolved in 9.5 ml of cell culture medium, aliquoted and stored in -20°C until use.
For measurement of CML, milk was dissolved in PBS, supplemented with 0.1% BSA, and centrifugated for 5 min at 10 000 x g. The supernatant was used for measurements of N-epsilon-carboxymethyl-lysine content using a commercial ELISA kit (STA-816, Cell Biolabs) according to the instructions of the manufacturer.
2.2. Cell Lines and Cell Culturing
STC-1, a murine intestinal neuroendocrine cell line, was obtained from the American Tissue Culture Company (ATCC-CRL-3254). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 4.5 g/L D-glucose, L-glutamine, sodium pyruvate (#6429, Sigma-Aldrich) supplemented with 1% (v/v) non-essential amino acids (NEAA) (Gibco, #11140-035), 1% (v/v) glutagro (Corning, #25-015-Cl), penicillin 10,000 U/mL and streptomycin 10,000 U/mL (Cytiva, #SV30010), and 10% (v/v) heat-inactivated fetal bovine serum (Sigma-Aldrich, #7524) in a humidified atmosphere of 5% CO2 at 37°C. For experiments, cells were seeded in 24-well or 96-well plates (Sarstedt) at a density of 7x105 cells/well and 3 x104 cells/well, respectively. Medium was changed every third day and experiments were performed 21 days after seeding.
Caco-2, a human cell line used as model of the intestinal epithelial barrier, was obtained from the European Collection of Authenticated Cell Cultures (ECACC 86010202) through Sigma-Aldrich (Stockholm, Sweden). Cells were cultured as above. For experiments, cells were seeded in Corning Transwell 12-well inserts with a polyester membrane (0.4 μm pore size, 1.12 cm2 surface area) at a density of 5x105 cells/well.
INS-1E, a rat insulinoma cell line, obtained from Prof. Claes Wollheim, University of Geneva [10] were cultured in RPMI 1640 Medium with 11.1 mM glucose (Cytiva), supplemented with 10 mM Hepes (Cytiva), 5% heat-inactivated fetal bovine serum (Sigma-Aldrich, #7524), 1 mM sodium pyruvate (Sigma), 50 µM β-mercaptoethanol (Gibco) and penicillin 10,000 U/mL and streptomycin 10,000 U/mL (Cytiva, #SV30010) in a humidified atmosphere of 5% CO2 at 37°C. For experiments, cells were seeded in 96-well plates 96-(Sarstedt) at a density of 3 x104 cells/well.
2.3. Viability Assay
For measurement of viability of STC-1 cells, these were seeded at a density of 3 x104 in 96-well plates and cultured for 24 h, after which the medium was changed and fresh or stored skim milk at different concentrations was added. Medium was used to adjust the final volume to 100 µl/well. Cells were exposed to the milk for 24 h, after which cell viability was measured using the MTS assay from Promega (G3582) according to the instructions from the manufacturers and a FLUOstar OPTIMA plate reader (BMG Labtech).
2.4. Proliferation Assay
For measurement of proliferation of STC-1 cells, these were seeded at a density of 7.5 x103 in 96-well plates and cultured for 24 h, after which the medium was changed and fresh or stored skim milk at different concentrations was added. Medium was used to adjust the final volume to 100 µl/well. Cells were exposed to the milk for 24 h, after which cell proliferation was measured using BrdU Cell Proliferation Assay kit from Novus Biologicals (NBP2-54888) according to instruction of the manufacturer and a FLUOstar OPTIMA plate reader (BMG Labtech).
2.5. Secretion of GLP-1 and GIP
STC-1 cells were seeded in 24-well plates and cultured for 24 h, after which the medium was discarded and replaced with serum-free medium. Different concentrations of fresh and stored milk were then added to the plate and medium was used to correct the volume to 500 µl/well. After incubation for 24 h, the medium was discarded, and plates were washed twice with PBS. Serum-free medium, supplemented with 20 nM DPP-IV inhibitor (Biotechne) was added and plates were incubated for another 4 h. Medium was recovered and stored at -80°C until further analyses. GLP-1 was measured by ELISA from Invitrogen (BMS2194) and GIP by ELISA from Millipore (EZRMGIP-55K) according to the instructions of the manufacturers.
2.6. Protein Content in the Cell Cultures
Cells were harvested by adding 100 µl of lysis buffer containing 50 mM TES, pH 7.4, 250 mM sucrose, 1 mM EDTA, 2 mM EGTA, 40 mM phenyl-phosphate, 5 mM sodium fluoride, 1 mM dithiothreitol, 50 mM sodium vanadate, Pefabloc (Sigma), Complete (Roche, protease inhibitors) and 1% NP40 to each well and scraping the cells off the plate. Samples were transferred to tubes and incubated on ice for 10 min. After incubation, lysates were sonicated (10 short pulses, Sonifier 250 from Branson) and centrifuged for 5 min at 5000 x g at 4°C. The supernatant was used to analyze the protein content using the Bradford method.
2.7. Exposure of INS-1E Cells to Basolateral Medium from Milk-Exposed Caco-2 Cells
To study the absorption trough Caco-2 cells, these were incubated with fresh or stored skim milk for 24 hours to allow absorption. Thereafter the absorbed nutrients, in the form of basolateral transudate was collected. The transudate was then added to insulin producing cells, INS-1E cells and their viability studied.
Caco-2 cells were seeded at a density of 5 x105 cells/well in the apical compartment of transwell plates Following 21 days of culture, the medium in both compartments were replaced with serum-free medium without phenol red. Different concentrations of milk was added to the apical compartments, and medium was added to adjust the final volume to 500 µl/well. The plate was then incubated for 24 h. Medium from the basolateral compartment was thereafter collected and stored at -80°C. INS-1E cells were seeded at a density of 3 x104 cells/well in a 96-well plate and incubated for 24 h. Basolateral medium from the milk exposed Caco-2 cells was added in different concentrations to the wells and the plate was incubated for 24 h. Viability of the INS-1E cells was measured using the MTS assay from Promega (G3582) according to the instructions from the manufacturers.
2.8. Statistics
Viability in STC-1 cells exposed for fresh and stored milk in different concentrations of milk powder is compared in a paired Student´s T-test. Two-sided p-values are used. ANOVA and unpaired Student´s T-tests are used to test the dose dependent impact of milk powder.
Proliferation for fresh and stored milk for every concentration of milk powder are compared in a paired Student´s T-test. Two-sided p-values are used.
The amount of released GLP-1 and GIP after exposure for fresh and stored milk in stepwise increasing concentration are compared in a paired Student´s T-test. Two-sided p-values are used. ANOVA and unpaired T-tests is used to test the dose dependent impact of milk powder.
2.9. Ethical Consideration
This study involves cell lines, and all experiments are done in vitro. By law, no ethical approval is required for this study.
3. Results
3.1. Measurement of Maillard Reactants
As a representative of Maillard reactants, the carboxy methyl lysine (CML) levels were measured. CML concentration was 8.6 ng/ml in fresh milk and 601.3 ng/ml in stored milk powder. This means that the concentration of CML has increased 70 times.
3.2. Viability of STC-1 Cells Exposed to Skim Milk Powder
Exposure to skim milk powder decreased the viability of intestinal neuroendocrine cells (STC-1-cells) in a dose-dependent way. For fresh skim milk powder there was a decrease in viability between 5 and 25 µl added milk (p=0.006). For stored milk powder there was a decreased viability with 20 µl added milk compared to 5 ul (p=0.019) and with 25 µl compared to 5 µl added milk (p<0.001). For added milk volumes ≥15 µl, stored milk decreased viability more than fresh milk (p<0.001) (Figure 1).
3.3. Proliferation of STC-1 Cells Exposed to Skim Milk Powder
Exposure to stored milk powder inhibited the proliferation of STC-1 cells in a dose-dependent way (Figure 2). For fresh milk powder there was no significant change in proliferation in relation to increased exposition of milk powder. For stored milk powder there was a decrease in proliferation with 25 µl added milk compared to 5 µl (p<0.002). For added milk volumes ≥20 µl, stored skim milk powder decreased proliferation more than fresh skim milk powder (p<0.001). For higher volumes, ie ≥20 µl of added skim milk powder solution, the proliferation of STC-1 cells was significantly lower for cells exposed to stored milk powder compared to cells exposed to fresh milk powder.
3.4. Release of GLP-1 from STC-1 Cells
Fresh skim milk powder induced a release of GLP-1 from STC-1 cells in a dose-dependent way whereas stored skim milk powder failed to stimulate GLP-1 release from STC-1 cells. For added skim milk volumes ≥250 µl, the release of GLP-1 was significantly lower with stored skim milk powder compared to fresh (Figure 3).
3.5. Release of GIP from STC-1 Cells
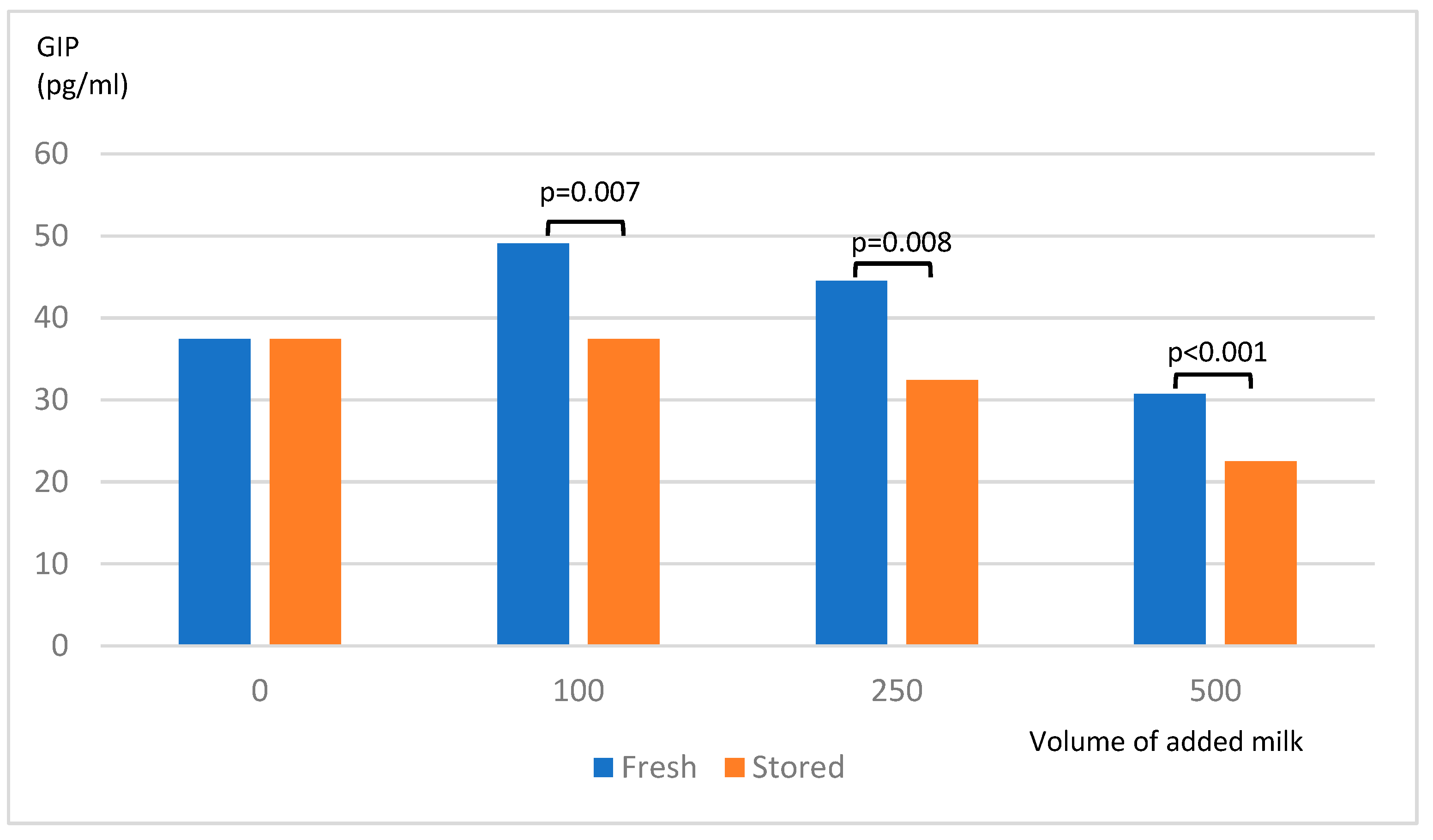
After incubation of STC-1 cells with stored skim milk powder ≥100 µl, the release of GIP was significantly lower compared to fresh skim milk powder (Figure 4). The protein content in the cell cultures tested with Bradford method showed very divergent results. No conclusion could be drawn whether the lower incretin secretion was due to fewer STC-1 or poorer function cells.
3.6. Viability of INS-1 Cells
The CML concentration in medium from the basolateral compartment of Caco-2 cells was very similar, i.e. 3.5 ng/ml in the presence of fresh skim milk powder and 3.15 ng/ml, in the presence of stored skim milk powder. This suggests that Maillard products do not pass the barrier of Caco-2 cells. In agreement with this, the viability of INS-1 cells incubated with basolateral medium from cells exposed to either fresh or stored milk powder, did not differ (data not shown).
3.7. Figures, Tables and Schemes
To test for viability, six independent experiments were done with three replicates for every concentration of milk in each assay. This will give 18 results for every concentration of both fresh and stored milk powder. Ratio of viability is calculated as measured absorbance in the trial sample divided by absorbance in a sample without added milk.
To test for proliferation, four different experiments were done with triplets. This will give 12 results for every concentration of both fresh and stored milk powder. Ratio of proliferation was calculated as measured absorbance in the trial sample divided by absorbance in a sample without added milk.
GLP-1 release from STC-1 cells were tested in duplicate in three different assays. This gives six results for every concentration of both fresh and stored milk powder. The concentrations of GLP-1 in pg/ml were obtained from a calibration curve.
GIP release from STC-1 cells were tested in duplicate in three different assays. This gives six results for every concentration of both fresh and stored milk powder. The concentrations of GIP in pg/ml were obtained from a calibration curve.
4. Discussion
In this study, we found that stored skim milk powder containing high amounts of Maillard reactants significantly decreased the release of GLP-1 and GIP from intestinal neuroendocrine STC-1-cells (Figure 3 and Figure 4). This is the first time incretins have been found to be suppressed by any dietary component. This emphasizes the role of dietary components in the regulation of glucose hemostasis.
STC-1-cells are derived from a neuroendocrine cell line from intestine releasing GLP-1 and GIP. During normal conditions, the release of these hormones occurs in response to intraluminal nutrients. Both GLP-1 and GIP are released after carbohydrate ingestion [11], but also in response to dietary fat and dietary proteins [12]. The release of GLP-1 and GIP from STC-1 cells treated with fresh skim milk powder, as demonstrated in this study, substantiates the ability of STC-1 cells to respond to normal dietary milk powder (Figure 3 and Figure 4). However, the conversion of skim milk powder to a powder enriched in Maillard reactants not only blocks the release of both incretins but also suppresses the release. High levels of Maillard reactants have for long time been considered harmful to health, especially affecting vascular inflammation [13], premature aging, and faster progression of diabetic complications [14]. The association between high consumption of highly processed foods, the so-called Western diet, and negative health effects such as diabetes, obesity and metabolic syndrome has long been known. The underlying mechanism for these negative effects has however not been fully described but an influence of a disturbed intestinal microbiota it proposed as an explanation [15].
In this study, by using Caco-2 cells as a model for the gastrointestinal barrier, we found no evidence of passage of Maillard reactants over the cellular barrier of the intestine. We therefore assume that Maillard reactants do not occur in significant amounts in the circulation and therefore do not have a direct effect on insulin secretion. We instead envisage a direct interference on the cellular level with the expected incretin release. This is in contrasts to other studies, where high levels of CML have been found in both plasma and urine in infants who received formula compared to breast milk [16].
In our previous studies, increase of inflammatory markers were found after treatment with stored skim milk powder for four weeks in newborn rats [7]. The same observation has been found in preterm pigs fed with high temperature treated formula and stored formula [17]. In our latter studies we also found a decrease in the number of GLP-1 expressing cells in the intestine and a decrease in beta cell mass in pancreas in rats treated with stored skim milk powder [8]. These initially difficult-to-interpret results can now be easily explained by our new findings of lack of secretion of incretins from the intestine. The suppression of GLP-1 release from the intestinal cells was hence supported by our present findings, whereas the suppressed release of GIP is a novel finding.
In conjunction to meals, incretins are important for insulin secretion to 75% [9] and maintenance of normal blood glucose levels [9,18]. The reduced secretion of incretins that we observed after exposure to stored milk compared to fresh, could either be due to a reduced cell mass or to a poorer response from the remaining cells. We attempted to calculate incretin secretion in relation to cell mass by determining the protein content in the wells and relate it to incretin level. However, the results of the protein determination showed divergent results so that no conclusions could be drawn. Since we also observed a decreased viability and a decreased proliferation of STC-1 cells after treatment with stored skim milk powder (Figure 1 and Figure 2), the reduced incretin levels are probably the result of a reduced number of viable cells.
The importance of incretins to keep a normal metabolism homeostasis is indisputable, as demonstrated by the use of new incretin drugs such as semaglutide and tirzepadide. Exogenous administration of GLP-1 in pharmacological doses is now an established drug treatment of both diabetes and obesity [19]. Addition of GIP has emerged in recent years as an even more effective drug for weight loss [20]. Here, only the release of GLP-1 and GIP have been studied, but other gastrointestinal hormones such as gastrin and cholecystokinin (CCK) could be of interest in this context.
5. Conclusions
In conclusion, the stored skim milk powder containing high amounts of Maillard reactants, gave a dose dependent inhibition of the incretin release from STC-1 cells, a cell model of intestinal endocrine cells. The eventual implication of these findings in humans for glucose regulation, diabetes, obesity and general human health must be further studied.
Author Contributions
All authors have contributed to the study in the following way: Mona Landin-Olsson posed the clinical question, formed the hypothesis, design the experiments and had main responsibility for the manuscript. Birgitte Ekholm conducted and interpreted the experimental trials. Charlotte Erlanson-Albertsson interpreted experimental results, wrote and revised the manuscript. Cecilia Holm designed experimental models, contributed with essential cell lines, had main responsibility for the trials, wrote and revised the manuscript.
Funding
The study was funded by ALF-grants, Lund University and grants from Region Skane.
Institutional Review Board Statement
This study involves cell lines, and all experiments are done in vitro. By law, no ethical approval is required for this study.
Data Availability Statement
The original contributions presented in this study are included in the article material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
None or the authors have any financial disclosure or any other conflict of interest concerning this manuscript.
Abbreviations
The following abbreviations are used in this manuscript:
| AGE | Advanced Glycated Endproducts |
| Caco-2 | human intestinal epithelial cell line |
| CCK CML |
Cholecystokinin N-epsilon-carboxymetyllysine |
| GIP GLP-1 INS-1 STC-1 |
Glucose-dependent insulinotropic peptide Glucagon-like peptide 1 Rat insulinoma cell line Murine intestinal neuroendocrine cell line |
References
- Wang, X.; Esquerre, C.; Downey, G.; Henihan, L.; O’Callaghan, D.; O’Donnell, C. Assessment of infant formula quality and composition using Vis-NIR, MIR and Raman process analytical technologies. Talanta 2018, 183, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Chong, H. Y.; Tan, L. T.; Law, J. W.; Hong, K. W.; Ratnasingam, V.; Ab Mutalib, N. S.; Lee, L. H.; Letchumanan, V. Exploring the Potential of Human Milk and Formula Milk on Infants’ Gut and Health. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Muts, J.; van Keulen, B. J.; van Goudoever, J. B.; van den Akker, C. H. P. Formula protein versus human milk protein and the effects on growth in preterm born infants. Curr Opin Clin Nutr Metab Care 2025, 28, 33–38. [Google Scholar] [CrossRef]
- Walsh, C.; Lane, J. A.; van Sinderen, D.; Hickey, R. M. Human milk oligosaccharides: Shaping the infant gut microbiota and supporting health. J Funct Foods 2020, 72, 104074. [Google Scholar] [CrossRef]
- Lithoxopoulou, M.; Karastogiannidou, C.; Karagkiozi, A.; Zafeiriadou, I. E.; Pilati, E.; Diamanti, E.; Kalogiannis, S.; Vassilopoulou, E. From Mother-Fetus Dyad to Mother-Milk-Infant Triad: Sex Differences in Macronutrient Composition of Breast Milk. Nutrients 2025, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, M.; Wang, Y.; Wu, Z.; Yang, J.; Li, H.; Li, H.; Yu, J. Control of the Maillard reaction and secondary shelf-life prediction of infant formula during domestic use. J Food Sci 2023, 88, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Hillman, M.; Weström, B.; Aalaei, K.; Erlanson-Albertsson, C.; Wolinski, J.; Lozinska, L.; Sjöholm, I.; Rayner, M.; Landin-Olsson, M. Skim milk powder with high content of Maillard reaction products affect weight gain, organ development and intestinal inflammation in early life in rats. Food Chem Toxicol 2019, 125, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Dereke, J.; Ekblad, E.; Westrom, B.; Erlanson-Albertsson, C.; Landin-Olsson, M.; Sjoholm, I.; Hillman, M. Structural and immunoendocrine remodeling in gut, pancreas and thymus in weaning rats fed powdered milk diets rich in Maillard reactants. Sci Rep 2022, 12, 4039. [Google Scholar] [CrossRef] [PubMed]
- Holst, J. J.; Gasbjerg, L. S.; Rosenkilde, M. M. The Role of Incretins on Insulin Function and Glucose Homeostasis. Endocrinology 2021, 162. [Google Scholar] [CrossRef] [PubMed]
- Merglen, A.; Theander, S.; Rubi, B.; Chaffard, G.; Wollheim, C. B.; Maechler, P. Glucose sensitivity and metabolism-secretion coupling studied during two-year continuous culture in INS-1E insulinoma cells. Endocrinology 2004, 145, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Vilsbøll, T.; Krarup, T.; Madsbad, S.; Holst, J. J. Both GLP-1 and GIP are insulinotropic at basal and postprandial glucose levels and contribute nearly equally to the incretin effect of a meal in healthy subjects. Regul Pept 2003, 114, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Carr, R. D.; Larsen, M. O.; Winzell, M. S.; Jelic, K.; Lindgren, O.; Deacon, C. F.; Ahrén, B. Incretin and islet hormonal responses to fat and protein ingestion in healthy men. Am J Physiol Endocrinol Metab 2008, 295, E779–784. [Google Scholar] [CrossRef] [PubMed]
- Bettiga, A.; Fiorio, F.; Di Marco, F.; Trevisani, F.; Romani, A.; Porrini, E.; Salonia, A.; Montorsi, F.; Vago, R. The Modern Western Diet Rich in Advanced Glycation End-Products (AGEs): An Overview of Its Impact on Obesity and Early Progression of Renal Pathology. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Gill, V.; Kumar, V.; Singh, K.; Kumar, A.; Kim, J. J. Advanced Glycation End Products (AGEs) May Be a Striking Link Between Modern Diet and Health. Biomolecules 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- García-Montero, C.; Fraile-Martínez, O.; Gómez-Lahoz, A. M.; Pekarek, L.; Castellanos, A. J.; Noguerales-Fraguas, F.; Coca, S.; Guijarro, L. G.; García-Honduvilla, N.; Asúnsolo, A.; et al. Nutritional Components in Western Diet Versus Mediterranean Diet at the Gut Microbiota-Immune System Interplay. Implications for Health and Disease. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Sebeková, K.; Saavedra, G.; Zumpe, C.; Somoza, V.; Klenovicsová, K.; Birlouez-Aragon, I. Plasma concentration and urinary excretion of N epsilon-(carboxymethyl)lysine in breast milk- and formula-fed infants. Ann N Y Acad Sci 2008, 1126, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Akıllıoğlu, H. G.; Aasmul-Olsen, K.; Ye, Y.; Lund, P.; Zhao, X.; Brunse, A.; Nielsen, C. F.; Chatterton, D. E. W.; Sangild, P. T.; et al. Ultra-High Temperature Treatment and Storage of Infant Formula Induces Dietary Protein Modifications, Gut Dysfunction, and Inflammation in Preterm Pigs. Mol Nutr Food Res 2022, 66, e2200132. [Google Scholar] [CrossRef] [PubMed]
- Alsalim, W.; Lindgren, O.; Ahrén, B. Glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1 secretion in humans: Characteristics and regulation. J Diabetes Investig 2023, 14, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Frías, J. P.; Auerbach, P.; Bajaj, H. S.; Fukushima, Y.; Lingvay, I.; Macura, S.; Søndergaard, A. L.; Tankova, T. I.; Tentolouris, N.; Buse, J. B. Efficacy and safety of once-weekly semaglutide 2·0 mg versus 1·0 mg in patients with type 2 diabetes (SUSTAIN FORTE): a double-blind, randomised, phase 3B trial. Lancet Diabetes Endocrinol 2021, 9, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Jastreboff, A. M.; le Roux, C. W.; Stefanski, A.; Aronne, L. J.; Halpern, B.; Wharton, S.; Wilding, J. P. H.; Perreault, L.; Zhang, S.; Battula, R.; et al. Tirzepatide for Obesity Treatment and Diabetes Prevention. N Engl J Med 2025, 392, 958–971. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Viability of STC-1-cells after exposure to milk powder.
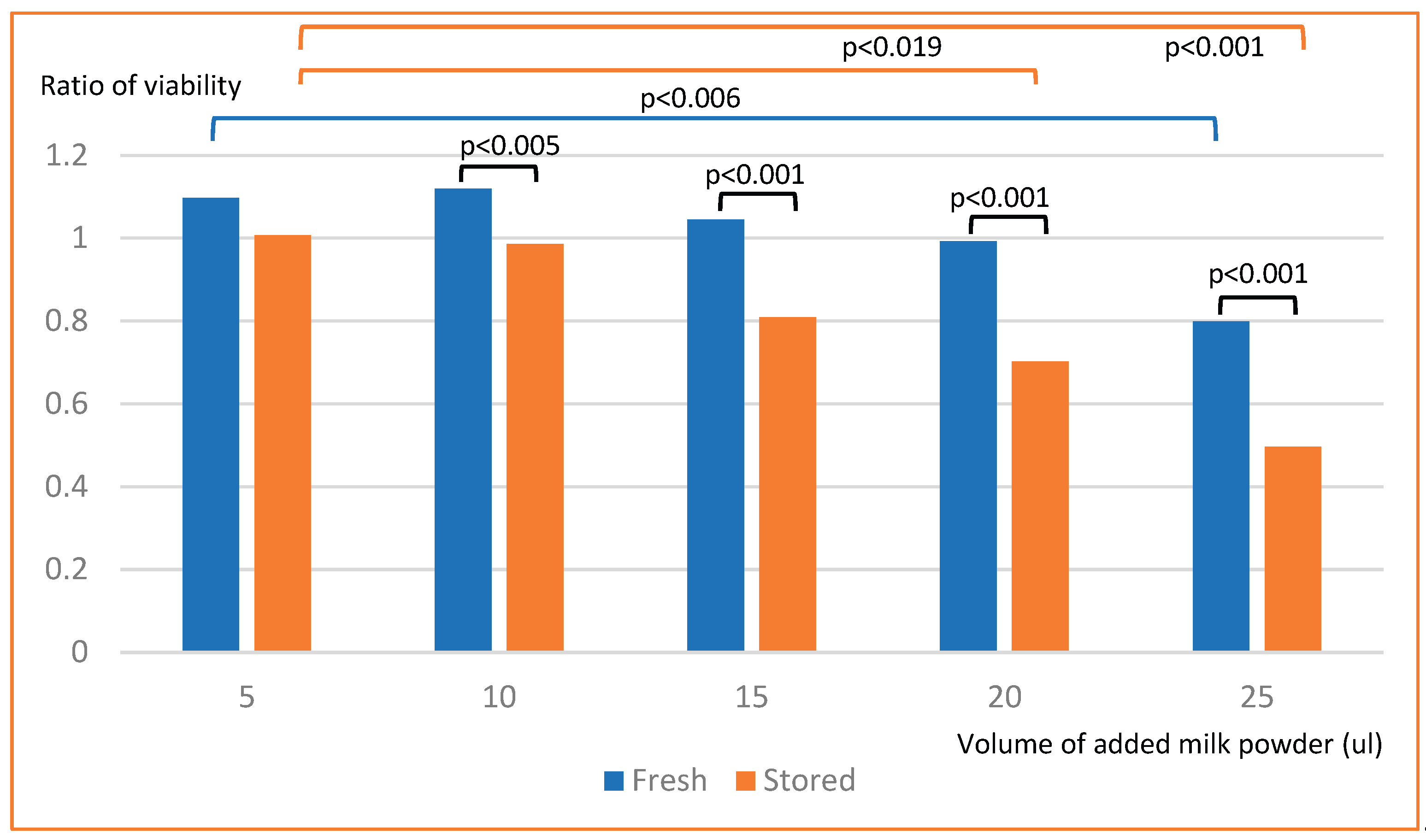
Figure 2.
Proliferation of STC-1-cells after exposure to milk powder.
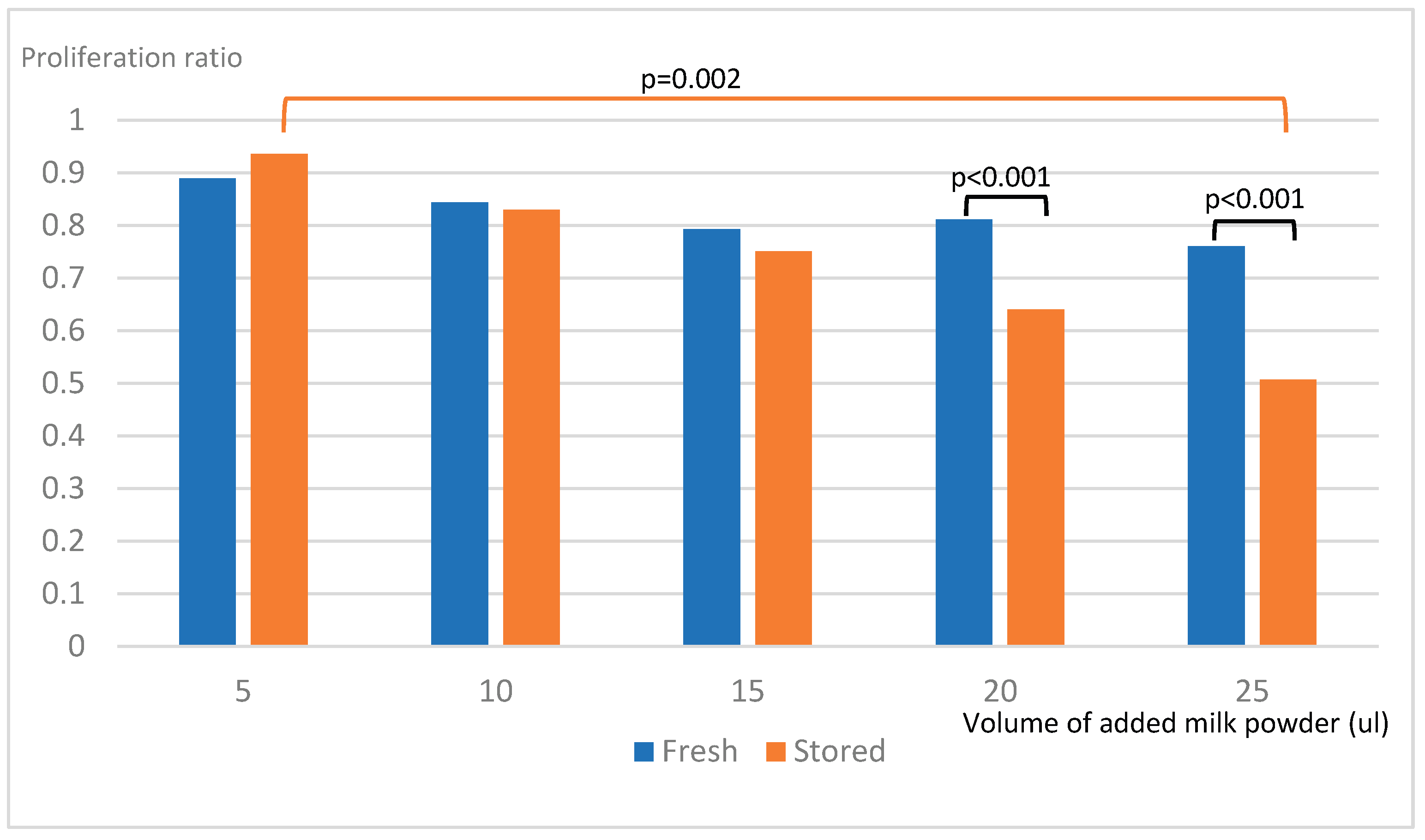
Figure 3.
Release of GLP-1 from STC-1-cells after exposure to milk powder.
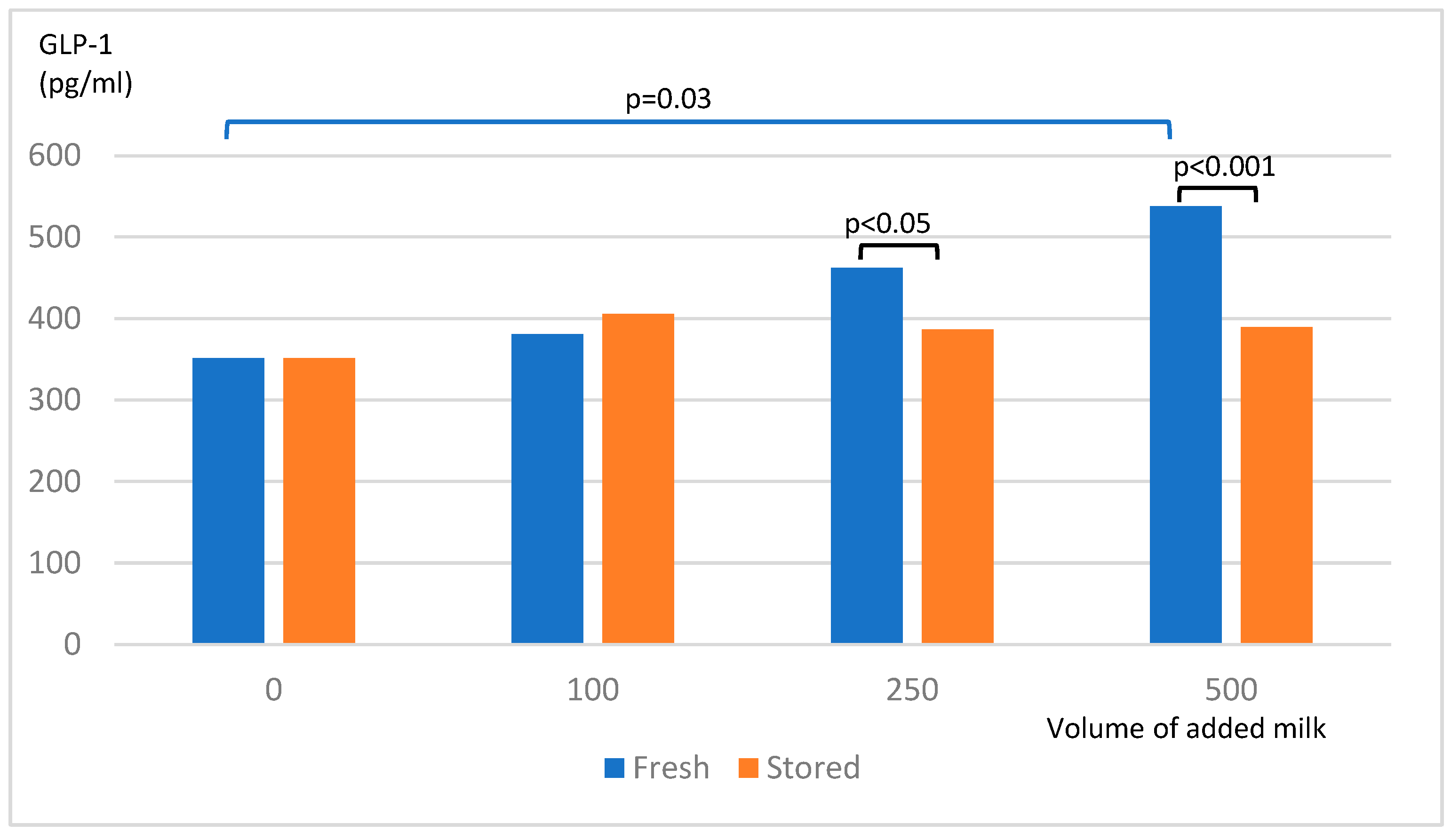
Figure 4.
Release of GIP from STC-1-cells after exposure to milk powder.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



