Submitted:
08 August 2025
Posted:
11 August 2025
You are already at the latest version
Abstract
Iodine is a crucial component of thyroid hormones (THs), specifically triiodothyronine (T3) and thyroxine (T4) and is vital for normal thyroid function. During pregnancy, a deficiency in iodine can result in inadequate production of THs, leading to hypothyroidism. This con-dition can negatively impact the intellectual development of the newborn. Numerous stud-ies have demonstrated that iodine deficiency during pregnancy correlates with lower IQ levels in children. This deficiency can hinder critical processes such as neurogenesis, neu-ronal migration, and myelination, all of which are essential for proper cognitive develop-ment. Research indicates that children born to iodine-deficient mothers are at a higher risk of having lower IQs compared to those whose mothers received iodine supplements during pregnancy. Therefore, ensuring sufficient iodine levels and optimal TH function during pregnancy is crucial for the cognitive development of children. This highlights the im-portance of nutrition and regular monitoring of thyroid health in pregnant women. This re-view focuses on the key mechanisms of thyroid hormone homeostasis and discusses the implications of iodine deficiency during pregnancy.
Keywords:
thyroid hormones
; iodine
; pregnancy
; placenta
; hypothyroidism
1. Introduction
Iodine is an essential trace element required in small amounts for the synthesis of the THs ,T3 and T4 [1]. Its role during pregnancy is particularly important as it contributes to the development of the central nervous system (CNS). Due to physiological changes during pregnancy, iodine requirements significantly increase from 150 μg per day in non-pregnant adult women to 250 μg per day in pregnant women [2]. Epidemiological studies have shown that inadequate iodine consumption during pregnancy is linked to reduced cognitive function in children, highlighting the importance of monitoring thyroid health in pregnant women [3,4,5]
Thyroid Hormones Synthesis
THs are synthesized and released by the thyroid gland [6,7]. Anatomically, the thyroid gland is one of the largest endocrine glands in the neck [8], composed of two lobes joined together by an intermediate structure known as the isthmus [9]. The hormones produced by the thyroid gland are 3,5,3′,5’- tetraiodothyronine, or thyroxine (T4), and 3,5,3’- triiodothyronine (T3) [10]. THs biosynthesis requires iodine (I) which is an integral part of both T3 and T4 and supplied in the daily diet in the form of iodide (I-) [11]. Iodide is absorbed in the small intestine, and its absorption is facilitated by the sodium/iodide symporter (NIS), which is expressed on the apical membrane of intestinal cells [12]. In contrast, iodide is secreted into the lumen of the stomach and salivary glands through NIS located on the basolateral membrane. Once in the small intestine, iodide is reabsorbed into the bloodstream via NIS on the apical membrane, completing its circulation back into the body's systemic circulation [13]. Circulating iodide is either absorbed by the thyroid gland via the NIS located on the basolateral plasma membrane of thyrocytes or excreted from the body through urine [1].
The synthesis and release of THs are initially stimulated by the production of thyrotropin-releasing hormone (TRH) in the hypothalamus, which stimulates the synthesis and secretion of thyroid-stimulating hormone (TSH) in the anterior pituitary [14,15]. Subsequently, TSH binds to its receptor on the thyroid gland, stimulating the biosynthesis and secretion of THs [15]. This regulation axis is part of the neuroendocrine system known as the hypothalamic-pituitary-thyroid axis (HPT axis) [16]. Furthermore, THs regulate the secretion of TRH and TSH via a negative feedback loop mechanism [14]. This mechanism occurs because THs inhibit the transcription of TRH and TSH subunit genes, as well as posttranslational modification and release of TSH [17]. The biosynthesis process of THs involves several proteins and occurs within the thyroid follicles, which are the functional units of the thyroid. Thyroid follicles are composed of thyrocytes that form well-defined anatomical structures; at the apical side of thyrocytes is the lumen, and at the basolateral side of thyrocytes there are blood capillaries [9,18]. Iodide (I-) is transported from the bloodstream into the thyrocytes through NIS [19]. Subsequently, the efflux from the apical membrane towards the lumen is mediated by ionic channels such as pendrins [20] and anoctamin [21]. Thyrocytes also produce and secrete thyroglobulin (TG) into the lumen [22]. Then, the thyroperoxidase (TPO) catalyses the oxidation of I- for its subsequent incorporation into the tyrosine residues of thyroglobulin (TG) to produce iodinated TG [18]. This enzymatic oxidation reaction occurs in the presence of hydrogen peroxide (H2O2) [23], produced by the thyroid NADPH oxidase called dual oxidase 2 (DUOX2) [24,25]. Iodinated TG forms 3-iodotyrosine or monoiodothyronine (MIT) and 3,5-diiodotyrosine or diiodotyrosine (DIT) [26]. T3 and T4 are formed by the coupling reaction of MIT and DIT [27]. In this reaction, the oxidations of iodotyrosines are also catalysed by TPO [10]. Finally, thyrocytes endocyte THs from the follicular lumen through endocytic vesicles. Here, THs are proteolysed from iodinated TG, in a process mediated by an iodotyrosine dehalogenase (DEHAL1) [28], liberating intrathyroidal iodide. Subsequently, at the basolateral membrane, THs are released into the bloodstream through monocarboxylate transporters (MCT) 8 and 10 [29]. The mechanism of THs biosynthesis can be summarised in four stages: oxidation of the TPO by H2O2, iodination reaction, coupling reaction, and iodide recycling and release of THs from follicles [18] (Figure 1). T4, also considered a prohormone o reserve hormone, is the most abundantly secreted hormone by the gland [30]. Serum transport of T4 is mediated by their strong binding to the principal plasma thyroid hormone-binding proteins, thyroxine-binding globulin (TBG), transthyretin (TTR), and albumin [31]. Although in human TBG is the protein with the highest affinity for THs, TTR also plays a significant role in TH transport in specific tissues like placenta [32,33]. These proteins could change in several conditions like gestational stage and hyperthyroidism [34]. In peripheral tissues, the deiodination process also occurs, leading to the activation of THs and regulating their biological activities. This process is mediated by tissue enzymes with differential expression that provide dynamic control of THs signaling [35]. These enzymes, which catalyze deiodination, are known as type 1 (D1), type 2 (D2), and type 3 (D3) deiodinases [36]. T3 which is considered the biologically active hormone is produced by T4 deiodination in peripheral tissues, primarily by D1 and D2 [25,26]. Conversely, D3 degrades T4 to the inactive form known as reverse T3 (rT3) [36]. In this regard, it is proposed that the activating deiodinase D1 and D2 and the inactivating deiodinase D3 are capable of modifying THs action independently from THs serum concentrations changes based on the requirements of the specific tissue [35,37].
Within the mechanisms of action of THs, two effects are known that these hormones exert. The genomic effect, which is described by a direct action of THs on gene transcription, and, on the other hand, the non-genomic effects.
The genomic effect begins with the entry of thyroid hormones into the cell, in which T4, after its entry, is converted into T3 by D2. Once converted into T3, this hormone binds to the THs nuclear receptors (TR), which together bind to the promoter region of target genes [38,39,40,41,42,43]. For example, TRs may bind to genes necessary for the production of myelin basic protein in the CNS [44], or genes such as Endothelin-1 and fibronectin, in umbilical cord vein endothelial cells (HUVEC) [42]. In the other hand, non-genomic effects begin with the binding of thyroid hormones to their membrane receptor, integrin ɑvβ3. From which occurs the activation of the signaling cascade that results in the activation of cell proliferation signals, angiogenesis, secretion of growth factors in the membrane of erythrocytes, leukocytes or hepatocytes, intracellular trafficking of proteins, among other functions [45,46,47]. It has been described that both T3 and T4 can bind to the ɑvβ3 receptor, but this binding will take place in different domains, activating different signaling pathways [48]. T3 binds to the S1 domain, which activates phosphatidylinositol 3 kinase (PI3K) promoting via TRɑ the transcription of the hypoxia inducible factor-1ɑ (HIF-1ɑ). While in the S2 domain both T3 and T4 would bind, and both would activate the extracellular signaling regulatory kinase (ERK1/2), which via TRβ1 would promote cell proliferation [49]. It has also been described that the receptor's own ɑv subunit in response to T4, would enter the cell nucleus to bind to the promoter region of the HIF-1ɑ, TR-β1 and COX-2 genes and promote their transcription [47].
Thyroid Hormones and Placenta
The placenta is a transient, highly specialized organ which allows gas and nutrient exchange, while acting as a selective barrier. It also plays a crucial role in regulate THs concentration between maternal and fetal circulation, ensuring fetal development [33]. Since the fetus initially depends entirely on maternal THs, any disruption in their production or regulation may lead to complications such as pre-eclampsia or miscarriage [50].
The biological activity and cellular effects of THs depend on their interaction with membrane transporters, specific receptor proteins and their intracellular metabolism mainly mediated by iodothyronine deiodinases (D) [51]. During pregnancy, THs cross the placenta via thyroid hormone transporters (THT) including monocarboxylate transporters (MCT8 and MCT10), L-type amino acid transporters (LAT1 and LAT2), and organic anion transporters (OATP1A2 and OATP4A1) [33]. These transporters are differentially expressed in different placental cell types [52]. MCTs, OATPs and LAT1 localize to villous syncytiotrophoblasts while MCTs and OATP1A2 are also found in cytotrophoblasts and extravillous trophoblasts. Additionally, MCTs are expressed in the stroma [39].
Thyroid hormone receptors (THR) are widely expressed in placental structures, with THRα1 being the predominant isoform in the human term placenta. Strong THR expression has been detected in syncytiotrophoblast, villous, and extravillous cytotrophoblasts, and the chorionic villi [53]. In the decidua, stromal and endothelial cells express TRα1, TRα2 and TRβ1, with expression pattern changing through gestation [53,54]
D3 is the most highly expressed deiodinase in the placenta [55]. Placental D expression is dynamic throughout pregnancy: D2 is more abundant on early gestation but declines toward term, whereas D3 expression increases. Immunohistochemical studies show a predominant D3 expression in syncytiotrophoblast during the first trimester, while D2 becomes more prominent in late pregnancy [56]. These findings suggest that D3 may play a key role in regulating maternal T3 transfer to the fetus.
Also, T4 can be transferred from maternal to fetal circulation through the chaperone action of TTR, which facilitates the transport of TH across the placental barrier to the fetal capillaries [57].
The Critical Role of Thyroid Hormones for Fetal Development
Fetal development is complex and highly regulated by different factors. This process begins with gastrulation and the generation of the 3 germ layers known as Ectoderm, Mesoderm, and Endoderm [58]. The thyroid gland arises from the endoderm after 4 weeks of gestation and is located on the base of the primitive pharynx. In this place a thickening called thyroid diverticulum is formed. Then, the thyroid diverticulum will lead to the formation of thyroid follicles after 8 weeks of the gestation. Finally, once the thyroid follicle matures, it will form the thyroid gland, approximately the six weeks of gestation. It is worth to note that in the thyroid gland there are parafollicular cells (also known as C cells), which have neuroendocrine nature and synthesize mainly calcitonin, which is involved in calcium and phosphate balance [59]. On the other hand, the CNS is formed from the Ectoderm, which approximately occurs 20 days after the gastrulation process. Specifically, the Ectoderm begins to thicken in the dorsal region of the embryo to form the neural plate, which will give rise to the neural folds. These neural structures will shape the neural tube that will end up giving rise to the CNS [60]. The fetal brain is extremely plastic and vulnerable to environmental and nutritional influences and, alterations in any of these conditions could have an impact on the health and development of the fetus.
One of the factors involved in the development and maturation of the fetus is the thyroid gland, which is responsible for secreting and producing the body's THs, triiodothyronine (T3) and thyroxine (T4). These THs play multifaceted roles in the development of the organism and in the homeostatic control of fundamental physiological mechanisms such as body growth and energy expenditure in all vertebrates, and neurodevelopment [61].
Thyroid cells will not begin to secrete T3 and T4 until after 12 weeks of gestation, reaching significant concentrations around 18 to 20 weeks [62]. At of birth, T3 and T4 concentration increase rapidly [63]. The secretion of THs by the thyroid gland is highly controlled. The thyroid-stimulating hormone (TSH), secreted by the pituitary gland, is the main control factor of THs secretion. Premature babies have been reported to have lower concentrations of THs. This condition may be due to a problem in the hypothalamic-pituitary-thyroid (HPT) axis. Likewise, in the case of babies from mothers with severe autoimmune hypothyroidism, lower concentration of THs is a consequence of placental transfer of maternal anti-thyroid antibodies [64,65,66,67].
As mentioned above, THs are fundamental for multiple processes in organisms, especially in fetal neurological development. It has been reported that THs are crucial for migration, differentiation, myelination, and cell signaling [68]. THs deficiency in the early stages of gestation, can cause critical damage to embryonic development both physiologically and morphologically, as they are responsible for the differentiation and maturation of key brain regions such as the cerebral cortex, hippocampus and cerebellum [67,69]. Both congenital hypothyroidism and maternal hypothyroidism can lead to cretinism, which produces intellectual retardation, deaf-muteness, gait disturbances, and several cognitive problems. Indeed, it has been reported that low concentration of THs can affect gray matter volume [69]. As reported, maternal THs transferred during the gestation (particularly during the first trimester) are involved in the maturation and functional formation of the brain, proliferation and migration of neurons, differentiation of neuronal and glial cells and synaptogenesis, among others [65]. Moreover, alterations in THs concentration can also alter the brain biochemistry. Here, a population of neurons that is strongly affected by THs deficiency is γ-aminobutyric acid (GABA)-ergic interneurons in the neocortex, hippocampus and cerebellum [70]. The decrease of γ-aminobutyric acid (GABA) levels reduces the cell migration and myelination of axons in the CNS [71]. Other abnormalities that have been observed and that can be attributed to the low concentration of maternal THs include limited growth of axons and dendrites; decreased neuronal connectivity; myelin deficits; and reduced synaptic densities [72]. On the contrary, elevated concentrations of maternal THs accelerate neurological development with greater cell proliferation and differentiation, resulting in decreased brain and body tissue weight and brain degeneration [73]. Experimental hyperthyroidism in wistar rats during gestation leads to morphological changes of the placenta, displaying an altered basal zone and decidua, consistent with a reduction of the thickness in the basal decidua and an increase in the basal zone thickness [74]. Moreover, the authors showed an increased fetal number, weight, and placental weight on day 19 of gestation. These data suggests that hyperthyroidism impacts placental function and fetal development, both could be related to preterm labor observed in pregnant hyperthyroid women [75].
Iodine Metabolism During Pregnancy
Dietary iodide and recycled iodide from thyroid hormone (TH) metabolism are absorbed in the small intestine, aided by the expression of the sodium-iodide symporter (NIS) on the apical side of enterocytes [12]. This process allows iodide to enter the bloodstream. The recommended dietary iodine intake for adults is 150 μg per day. During pregnancy, the need for iodine increases due to higher production of THs, increased iodide clearance and transfer to the fetus, and the rise in type III deiodinase activity [12]. When iodide supply is insufficient, the thyroid gland adapts by increasing iodide uptake [76].
Deiodination is the first step in the activation/inactivation process of THs and involves the removal of one iodine atom from the outer tyrosyl ring of T4 to produce T3, which is considered the biologically active form of THs.
D1 and D2 catalyze the outer ring deiodination of T4 and its conversion to active hormone T3 [77]. D3 converts T4 to reverse T3 and T3 to T2 and is highly expressed in the pregnant uterus, placenta, and fetal and neonatal tissues [77].
Finally, T4 interacts with plasma membrane receptors, specifically integrin ανβ3. This interaction triggers non-genomic action, such as promoting angiogenesis and the proliferation of tumoral cells. Additionally, the binding of T4 to integrin ανβ3 affects the activity of specific membrane ion pumps and stimulates the trafficking of protein within the cell [47,49,78].
Iodide Requeriment During Pregnancy
The increase in iodine requirement due to urinary losses [79,80] increased TH production, and iodine transfer to the fetus is compensate by increasing thyroid iodide uptake [81,82,83,84,85].
The recommended adequate iodine intake in adults is 150 μg/day. The World Health Organization recommends an iodine intake of 250 μg/day in pregnant women [86]. The urinary iodine concentration (UIC) is a sensitive indicator for current iodine intake because roughly 90% of all iodine consumed is excreted in the urine [87].
The median Urinary Iodine Concentration (UIC) indicative of optimal iodine intake for pregnant women is between 150 and 249 µg/L [88]. In pregnant women with adequate iodine levels, the increase in iodide requirements due to renal loss and fetal iodide transfer often goes unnoticed. However, for those with borderline low iodine intake, pregnancy can worsen iodine deficiency, putting stress on the homeostatic mechanisms responsible for maintaining a euthyroid state [5]. A healthy, non-pregnant woman typically requires about 80 µg of iodide per day to synthesize THs. When iodide intake is optimal at 150 µg/d, a thyroid uptake of 35% is sufficient to achieve balance in the system and maintain optimal iodine stores, which range from 10 to 20 mg (Figure 2A). Out of the 80 µg of iodide consumed, 15 µg is lost through feces, while the remaining 65 µg is divided between the thyroid and kidney losses [89]. In situations where iodide intake is low, the thyroid compensates by increasing iodide uptake to meet the daily requirement of 80 µg for TH production. Despite this increase, a daily deficit of about 12 µg occurs, leading to a gradual depletion of thyroid iodine reserves from the gland (Figure 2B). In iodine-deficient pregnant women, the iodide requirement rises from 80 to 120 µg/d due to a 50% increase in TH production (Figure 2C). Although there is an increase in iodide uptake by the thyroid, the total iodide intake is insufficient to meet the heightened requirement for hormone production and the daily iodide deficit further increase to about 20 µg/d [89,90].
Physiological Adaptation of Thyroid Function During Pregnancy
The risk of iodine deficiency seems higher in women than men [91], predisposing women to iodine deficiency. A combination of dietary, physiological, and lifestyle factors may explain why women have lower iodine intake than men (Figure 3). Dietary preferences are one of them. Women are more likely to follow plant-based diets poor in iodine. Physiological conditions like pregnancy and menstrual blood loss may also play a role in iodine deficiency. Pregnancy may increase the risk of iodine deficiency by a double mechanism. First pregnancy increases iodine requirements, and the effect of multiple pregnancies on iodine storage depletion is cumulative. Secondly, pregnancies and menstrual blood loss are often associated with iron deficiency. Iron deficiency and iron-deficient anemia are widespread in women and may interfere with thyroid function [92,93,94]. Iron deficiency decreases TPO iodination activity, a heme-containing enzyme, impairing iodine organification and uptake. The combined iodine and iron deficiency may also explain the higher risk of iodine deficiency and prevalence of thyroid diseases in women than in men [95].
The definition of iodine status uses the population's UIC. It does not reflect the situation of a given subject or subgroup of the population. The drawback of using UIC to assess iodine status at the individual level stems from the fact that UIC is highly variable from day to day depending on other factors like the hydration status of the subject affecting urine dilution [96].
During pregnancy, iodine requirements increase, first because of increased THs production and iodine transfer to the fetus and second because of increased renal iodine clearance [97]. Iodine-sufficient pregnant women can adapt to the increased iodine requirements without visible consequences on the thyroid gland. However, in conditions of iodine deficiency, the thyroid capacity to maintain optimal thyroid function may be impaired, leading to functional and structural alterations in maternal and newborn thyroid glands.
Pregnancy triggers several physiological modifications of thyroid function caused by increased thyroid hormone-binding globulin concentrations (TBG) due to estrogen stimulation of hepatocytes protein synthesis by week 7 of gestation. Compared with preconception concentrations, TBG concentrations are 2.5-fold higher than in non-pregnant women [98,99]. The increase in serum TBG leads to a rise in total T4 and T3, associated with a slight decrease in free THs [91,100]. The synthesis of T4 increases during pregnancy to compensate for the increase in the hormone-binding capacity of TBG [7] to maintain optimal free hormone concentrations [101]. By the end of the third trimester of gestation, the stimulatory effect of the peak of human chorionic gonadotropin (hCG) induced a transient increase in free T4 and a transient decrease of TSH. In normal conditions, in the second half of gestation, TSH values are similar to concentrations of non-pregnant women. On the contrary, TSH levels may increase above preconception reference ranges in women with low iodine intake (Figure 4) [102].
Clinical and Biological Consequences of Mild Iodine Deficiency in Pregnant Women and Newborns
Mild iodine deficiency [50-99 µg/L) frequently occurs in pregnant women in many countries, including the European region [86]. Mild iodine deficiency may lead to goiter formation and an increased thyroid volume in the newborn. However, THs concentrations remain within the normal ranges in pregnant women and newborns [83,103] (Figure 5).
Unlike other nutritional deficiencies, where a single blood test is sufficient to determine whether a subject is deficient, no individual laboratory test can diagnose iodine deficiency. The lack of an individual marker of iodine status represents an obstacle to diagnosing iodine deficiency in a given subject and for including truly iodine-deficient subjects in clinical trials. Most clinical trials use the median UIC of the pregnant women in a given region, assuming that the median UIC represents the iodine status of each pregnant woman included in a specific clinical trial. It is probable that as many as 90 % of pregnant women in mildly iodine-deficient regions, based on the population median UIC, have no biological signs of iodine deficiency [103]. The uncertain iodine status of pregnant women is a clear bias in the design and the interpretation of outcomes of iodine supplementation clinical trials concerning the benefit of iodine supplementation, particularly in mildly iodine-deficient pregnant women [79,80].
Although there is not an individual parameter of iodine deficiency, a combination of different thyroid parameters may be helpful to detect excessive thyroid hyperstimulation in otherwise euthyroid pregnant women among these parameters: a low free T4, increased T3/T4 ratio, and elevated TG concentrations.
Free T4 concentrations decrease slightly by the end of gestation, even in pregnant women with an adequate iodine supply. In iodine-deficient women, however, the free T4 decrease is more pronounced, although free T4 concentrations remained generally within the normal ranges. In conditions of mild iodine deficiency, up to one-third of pregnant women may have free T4 concentrations near or below the normal limit [81,101]. Interestingly, this is not the case for newborns with free T4 concentrations higher than their respective mothers, suggesting that the fetus is protected from hypothyroxinemia [83]. Iodine supplementation administered early in pregnancy prevents the occurrence of hypothyroxinemia [81,82,104].
When iodine intake is low there is a preferential secretion of T3, leading to an elevated T3/T4 ratio [77]. Under conditions of normal iodine intake, the T3/T4 ratio ranges between 10 –22 x 10-3 in pregnant women [105,106]. Again, iodine supplements can partially blunt the T3/T4 rise [103]. After delivery in non-supplemented pregnant women, recovery of the T3/T4 ratio to normal may take several months [107], indicating that thyroidal stimulation may have long-lasting and cumulative effects on the thyroid gland. Consequently, pregnancy is a goitrogenic determinant for pregnant women with marginally low iodine intake, and this effect may be cumulative, as shown by the positive association between parity and thyroid volume [84,85,108].
Thyroglobulin (TG) concentration has been correlated with thyroid volume and is increased in regions of iodine deficiency [26,102,109,110]. Thyroglobulin seems more sensitive to variations of iodine status than TSH, as it is often the only parameter associated with UIC in mildly deficient pregnant women [102]. Therefore, TSH may be less useful to diagnose mild iodine deficiency early in pregnancy. In addition to elevated TG concentrations, the presence of increased diffuse thyroid volume (>18 mL) in otherwise euthyroid pregnant women may also be indicative of iodine deficiency [103].
Therefore, it is feasible to use routine laboratory tests and standardized criteria to identify iodine-deficient pregnant women by detecting excessive thyroidal stimulation. In practical terms, a euthyroid pregnant woman with a Free T4 below or equal to the lower reference value of iodine sufficient healthy pregnant women population, a T3/T4 ratio > 25 x 10-3, and a TG value above or equal to the higher reference value would have a higher risk of iodine deficiency (Figure 6). It may be helpful to use these surrogate markers of iodine status to select pregnant women who will benefit most from iodine supplements.
In summary, the thyroid gland adapts to mild iodine deficiency by augmenting thyroid function to maintain the euthyroid status during pregnancy. The primary step of this homeostatic mechanism is the enhanced iodine uptake triggered by TSH, secondary to the slight decrease in free T4. Although this compensatory mechanism succeeds in its objective, there is a price to pay: chronic hyperstimulation, which leads to increased thyroid volume not only in pregnant women and newborns but also to a higher risk of thyroid nodules later in life.
Meanwhile, the iodine uptake and synthesis of THs by the fetal thyroid starts at 10 –12 weeks of gestation, and thyroid secretion becomes effective by midgestation [111]. The fetal thyroid function depends entirely on the iodine transferred from the pregnant women [111,112]. In conditions of mild iodine deficiency, despite relative pregnant women’s hypothyroxinemia, the newborn is protected from hypothyroxinemia [83]. Similarly to pregnant women, iodine deficiency stimulated thyroid activity of fetal thyroid, increasing TG concentrations in cord blood and thyroid volume compared to iodine-sufficient newborns [83].
Two randomized iodine supplementation trials of pregnant women in Belgium and Denmark with mild iodine deficiency reported similar findings for cord TSH and TG concentration [81,83]. Cord TSH concentrations were not different in the iodine-supplemented and non-supplemented pregnant women. On the contrary, in the Danish and Belgian trials, iodine supplementation resulted in a significant decrease in cord TG concentrations compared with newborns from non-iodine-supplemented pregnant women. Interestingly, these results suggest that cord TG would be more sensitive to variation in iodine intake than TSH, as previously shown in an iodine-deficient region of Italy [113].
In addition, thyroid volume in newborns in the Brussels trial was significantly larger in newborns from non-supplemented pregnant women than newborns from supplemented pregnant women.
Clinical and Biological Consequences of Severe Iodine Deficiency In pregnant Women and Newborns
The most dramatic consequences of low iodine intake occur if iodine deficiency is severe. When the UIC of school-age children is below 20 μg/L, a population is considered severely iodine deficient [86]. In severe iodine-deficient populations, goiter, cretinism, and hypothyroidism are endemic [114]. Hypothyroidism during pregnancy may result in both maternal and fetal hypothyroidism with severe neurologic and cognitive deficits in the offspring. Unlike mild iodine deficiency, when iodine deficiency is severe, the thyroid gland can no longer compensate to maintain euthyroid, and hypothyroidism may occur during pregnancy or even before pregnancy. A double-blind controlled trial conducted between 1966 and 1982 in an iodine-deficient region of the Western Province of Papua New Guinea evaluated the efficacy of intramuscular administration of iodized oil as a prophylactic measure against endemic cretinism, compared with saline as a control. The study found that pre-conception supplementation with iodized oil prevented endemic cretinism. In addition, the cumulative survival rate at 15 years was higher in the treated group than in the control group. Children whose mothers received supplementation showed significantly better intellectual and motor performance [115,116,117]. A recent study conducted in Greece in 2024 investigated the effects of iodine intake on the motor and cognitive development of children aged 4 and 6 years. The findings reveal that low iodine intake during childhood (UIC 100 μg/L) is linked to motor impairments in 4-year-olds and decreased intelligence in 6-year-olds when compared to a control group [118].These studies confirm that iodine deficiency during pregnancy leads to severe cognitive and motor deficits and that iodine supplementation effectively prevents these deficits.
In some iodine-deficient regions, goitrogens in the staple food may aggravate even more iodine deficiency during pregnancy by inhibiting iodine uptake [119,120]. Thiocyanate present in cassava and the smoke of tobacco competitively inhibit iodine uptake. Flavonoids present in millet impaired iodide organification by inhibiting TPO iodination activity [121]. In addition, other trace elements may interact with thyroid metabolism as selenium and iron deficiency, modulating the adaptation of the thyroid to iodine deficiency [92,94,122,123].
Endocrine disruptors (EDCs) are exogenous chemicals that interfere with hormone synthesis, metabolism, or action, increasing the risk of adverse health outcomes [124]. A number of EDCs impede iodine uptake by the thyroid gland, including perchlorate, nitrate and thiocyanate [125]. EDCs may also disrupt the thyroid during pregnancy [126,127,128]. Has been reported that high perchlorate exposure in pregnant women is associated with low IQ in their 3-year-old children. This study was part of the Controlled Antenatal Thyroid Study (CATS), in which involved over 21,000 pregnant women in the United Kingdom and Italy. The researchers found that pregnant women with the highest levels of perchlorate in their urine were the most likely to have children with the lowest IQ scores. This finding remained true even after looking at whether or not hypothyroidism was corrected for, if present, beginning in the first trimester of pregnancy, thus suggesting that perchlorate may have adverse effects independent of TH status. Pregnant women, in general, had low urinary iodine levels in this study, consistent with inadequate iodine intake [129]. Meanwhile, the impacts of maternal exposure to perchlorate, thiocyanate, and nitrate on offspring neurodevelopment was explored on a birth cohort in China. This study showed that double increment of thiocyanate and nitrate in urine samples during the first trimester was associated with 1.56 [95% CI: -2.82, -0.30) and 1.22 (-2.40, -0.03) point decreases in the offspring mental development index (MDI) respectively [130]. These findings suggested that prenatal exposure to the three chemicals (at current levels), especially thiocyanate and nitrate, may impair neurodevelopment.
Iodine Excess
Acute high iodine intake can cause transient inhibition of TH synthesis by a mechanism known as the acute Wolff-Chaikoff effect [131]. This inhibition is transient, and the thyroid “escapes” from the Wolff-Chaikoff effect within a few days by downregulating the iodide thyroid uptake with the restoration of normal TH synthesis [131]. Failure of the “escape” can result in iodine-induced hypothyroidism [132]. The fetal thyroid gland is not fully able to escape from the Wolff-Chaikoff effect until late in gestation, and fetal hypothyroidism may occur in case of acute iodine excess despite thyroid function remaining normal in pregnant women [133].
The Wolff-Chaikoff effect generally occurs with acute iodine excess during medical or diagnostic procedures, which are relatively infrequent during pregnancy. However, the use of antiseptics containing iodine, mainly for cesarean section, has been associated with higher neonatal TSH, increasing the recall rate for congenital hypothyroidism [134].
Iodine excess due to dietary or environmental sources is a more frequent condition associated with increased prevalence of thyroid autoimmunity [81,135]. In countries exposed to iodine excess, like Chile, the prevalence of hypothyroidism in the adult population, including pregnant women, is highly elevated [136]. A cross-sectional study in China showed a U-shaped relation between thyroid function and UIC among pregnant women in the first trimester of pregnancy; pregnant women within the group with UICs of 150–249 µg/L had the lowest serum TSH and TG concentrations [137]. The WHO recommended safe upper iodine intake for pregnant women is 500 µg/d [86].
2. Discussion
Iodine and THs play an essential role in pregnancy, ensuring proper fetal development and maternal health [138]. THs are involved in cellular development, growth, lipids and carbohydrates metabolisms, cellular respiration, total energy expenditure, growth, tissue maturation brain development [10]. Since iodine is an essential trace element of these hormones, adequate intake is necessary to support both the mother and the brain and CNS development of fetus [139,140].
During pregnancy, more iodine should be consumed than at any stage of life since its renal excretion is increased and part of it is diverted to the placenta for its passage to the fetus. The ideal dietary allowance of iodine recommended by the World Health Organization (WHO) is 150 μg iodine per day [114]. The physiological adjustments during pregnancy iodine requirements increase significantly from 150 μg per day in non-pregnant adult women to 250 μg per day in pregnant woman [2]. An iodine deficiency can have serious consequences that include impaired cognitive development, leading to reduce IQ in the newborn [3,141,142].
Research has consistently shown that iodine deficiency during pregnancy is associated with lower IQ scores in children. Studies indicate that even mild iodine deficiency can result in an average IQ reduction of 8 to15 points, meanwhile chronic moderate to severe iodine deficiency can reduced expected average IQ by about 13.5 points [4,143].
Iodine deficiency during pregnancy is a major public health concern due to its irreversible effects on brain development and intelligence in children. Ensuring adequate iodine intake through diet and supplementation for the THs production is crucial to preventing cognitive impairments and supporting optimal neurodevelopment. Therefore, it is recommended that pregnant, lactating, or women planning a pregnancy take a daily supplement of 150 μg of iodine, begin supplementation three months before a planned pregnancy, and avoid exposure to excessive doses of iodine, as it can cause fetal hypothyroidism.
3. Conclusions
The primary physiological adaptation of the thyroid during pregnancy is an increase in TH production, which begins in the first trimester. This heightened production of THs requires adequate iodide intake. When dietary iodide is insufficient, this physiological adaptation can become overwhelmed, resulting in thyroid overstimulation and impaired thyroid function. Pregnant women and newborns are particularly vulnerable to even slight deficiencies in iodide intake, making them a key group for monitoring iodine staus in the population.
Severe iodine deficiency impaired neurodevelopment in offspring. However, even mild iodine deficiency may affect thyroid function, which is often characterized by an increase in thyroid volume due to overstimulation. It remains uncertain whether mild iodine deficiency impacts the cognitive development of offspring, as both maternal and newborn THs concentrations typically remain within normal ranges.
The risk of iodine deficiency appears to be higher in women than in men. Pregnant women experience increased iodine requirements, and the cumulative effect of multiple pregnancies can lead to further a depletion of thyoid iodine stores. Additionally, menstrual blood loss during pregnancies is often linked to iron deficiency, which can interfere with thyroid function. The combination of iodine and iron deficiency may contribute to the higher incidence of thyroid diseases in women compared to men.
Starting iodine supplements during pregnancy may not fully replenish thyroid iodine stores. Therefore, it is essential for women to begin their pregnancy with adequate iodine reserves to meet the increased demands. This can be achieved by improving access to food products made with iodized salt, which can help fill the nutritional gap not met by diet alone.
Funding
This research was funded by FOVI230114 and National Research and Development Agency (ANID), and VRID-UdeC 2021000369MUL.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed, or endorsed by the publisher.
References
- Sorrenti, S.; Baldini, E.; Pironi, D.; Lauro, A.; D'Orazi, V.; Tartaglia, F.; et al. Iodine: Its Role in Thyroid Hormone Biosynthesis and Beyond. Nutrients. 2021, 13. [Google Scholar] [CrossRef]
- Stagnaro-Green, A.; Abalovich, M.; Alexander, E.; Azizi, F.; Mestman, J.; Negro, R.; et al. Guidelines of the American Thyroid Association for the diagnosis and management of thyroid disease during pregnancy and postpartum. Thyroid. 2011, 21, 1081–125. [Google Scholar] [CrossRef]
- Zimmermann, M.B. The effects of iodine deficiency in pregnancy and infancy. Paediatr Perinat Epidemiol. 2012, 26 (Suppl 1), 108–17. [Google Scholar] [CrossRef]
- Levie, D.; Korevaar, T.I.M.; Bath, S.C.; Murcia, M.; Dineva, M.; Llop, S.; et al. Association of Maternal Iodine Status With Child IQ: A Meta-Analysis of Individual Participant Data. J Clin Endocrinol Metab. 2019, 104, 5957–67. [Google Scholar] [CrossRef]
- Pearce, E.N.; Lazarus, J.H.; Moreno-Reyes, R.; Zimmermann, M.B. Consequences of iodine deficiency and excess in pregnant women: an overview of current knowns and unknowns. Am J Clin Nutr. 2016, 104 (Suppl 3), 918S–23S. [Google Scholar] [CrossRef]
- Armstrong, M.; Asuka, E.; Fingeret, A. Physiology, Thyroid Function. StatPearls. Treasure Island (FL): StatPearls Publishing; 2024.
- Oppenheimer, J.H. Role of plasma proteins in the binding, distribution and metabolism of the thyroid hormones. N Engl J Med. 1968, 278, 1153–62. [Google Scholar] [CrossRef]
- Singla, R.; Sharma, R.; Kaur, K. An unusual 'w' shaped thyroid gland with absence of isthmus - a case report. J Clin Diagn Res. 2014, 8, AD03–4. [Google Scholar]
- Khan, Y.S.; Farhana, A. Histology, Thyroid Gland. StatPearls. Treasure Island (FL): StatPearls Publishing; 2024.
- Shahid, M.A.; Ashraf, M.A.; Sharma, S. Physiology, Thyroid Hormone. StatPearls. Treasure Island (FL): StatPearls Publishing; 2024.
- Köhrle, J. Selenium, Iodine and Iron-Essential Trace Elements for Thyroid Hormone Synthesis and Metabolism. Int J Mol Sci. 2023, 24, 3393. [Google Scholar] [CrossRef]
- Nicola, J.P.; Carrasco, N.; Masini-Repiso, A.M. Dietary I(-) absorption: expression and regulation of the Na(+)/I(-) symporter in the intestine. Vitam Horm. 2015, 98, 1–31. [Google Scholar]
- Levay, B.; Lantos, A.; Sinkovics, I.; Slezak, A.; Toth, E.; Dohan, O. The master role of polarized NIS expression in regulating iodine metabolism in the human body. Arch Endocrinol Metab. 2023, 67, 256–61. [Google Scholar] [CrossRef]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol Rev. 2014, 94, 355–82. [Google Scholar] [CrossRef]
- Ortiga-Carvalho, T.M.; Chiamolera, M.I.; Pazos-Moura, C.C.; Wondisford, F.E. Hypothalamus-Pituitary-Thyroid Axis. Compr Physiol. 2016, 6, 1387–428. [Google Scholar] [CrossRef] [PubMed]
- Costa-e-Sousa, R.H.; Hollenberg, A.N. Minireview: The neural regulation of the hypothalamic-pituitary-thyroid axis. Endocrinology. 2012, 153, 4128–35. [Google Scholar] [CrossRef] [PubMed]
- Chiamolera, M.I.; Wondisford, F.E. Minireview: Thyrotropin-releasing hormone and the thyroid hormone feedback mechanism. Endocrinology. 2009, 150, 1091–6. [Google Scholar] [CrossRef]
- Carvalho, D.P.; Dupuy, C. Thyroid hormone biosynthesis and release. Mol Cell Endocrinol. 2017, 458, 6–15. [Google Scholar] [CrossRef]
- Carrasco, N. Iodide transport in the thyroid gland. Biochim Biophys Acta. 1993, 1154, 65–82. [Google Scholar] [CrossRef]
- Royaux, I.E.; Suzuki, K.; Mori, A.; Katoh, R.; Everett, L.A.; Kohn, L.D.; Green, E.D. Pendrin, the protein encoded by the Pendred syndrome gene (PDS), is an apical porter of iodide in the thyroid and is regulated by thyroglobulin in FRTL-5 cells. Endocrinology. 2000, 141, 839–45. [Google Scholar] [CrossRef]
- Twyffels, L.; Strickaert, A.; Virreira, M.; Massart, C.; Van Sande, J.; Wauquier, C.; et al. Anoctamin-1/TMEM16A is the major apical iodide channel of the thyrocyte. Am J Physiol Cell Physiol. 2014, 307, C1102–12. [Google Scholar] [CrossRef]
- Di Jeso, B.; Arvan, P. Thyroglobulin From Molecular and Cellular Biology to Clinical Endocrinology. Endocr Rev. 2016, 37, 2–36. [Google Scholar] [CrossRef]
- Szanto, I.; Pusztaszeri, M.; Mavromati, M. H2O2 Metabolism in Normal Thyroid Cells and in Thyroid Tumorigenesis: Focus on NADPH Oxidases. Antioxidants (Basel). 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Ameziane-El-Hassani, R.; Morand, S.; Boucher, J.-L.; Frapart, Y.-M.; Apostolou, D.; Agnandji, D.; et al. Dual oxidase-2 has an intrinsic Ca2+-dependent H2O2-generating activity. J Biol Chem. 2005, 280, 30046–54. [Google Scholar] [CrossRef]
- Song, Y.; Driessens, N.; Costa, M.; De Deken, X.; Detours, V.; Corvilain, B.; et al. Roles of hydrogen peroxide in thyroid physiology and disease. J Clin Endocrinol Metab. 2007, 92, 3764–73. [Google Scholar] [CrossRef] [PubMed]
- Citterio, C.E.; Targovnik, H.M.; Arvan, P. The role of thyroglobulin in thyroid hormonogenesis. Nat Rev Endocrinol. 2019, 15, 323–38. [Google Scholar] [CrossRef] [PubMed]
- Mansourian, A.R. Metabolic pathways of tetraidothyronine and triidothyronine production by thyroid gland: a review of articles. Pak J Biol Sci. 2011, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gnidehou, S.; Caillou, B.; Talbot, M.; Ohayon, R.; Kaniewski, J.; Noël-Hudson, M.-S.; et al. Iodotyrosine dehalogenase 1 (DEHAL1) is a transmembrane protein involved in the recycling of iodide close to the thyroglobulin iodination site. FASEB J. 2004, 18, 1574–6. [Google Scholar] [CrossRef]
- Weber, J.; McInnes, J.; Kizilirmak, C.; Rehders, M.; Qatato, M.; Wirth, E.K.; et al. Interdependence of thyroglobulin processing and thyroid hormone export in the mouse thyroid gland. Eur J Cell Biol. 2017, 96, 440–56. [Google Scholar] [CrossRef]
- Schroeder, A.C.; Privalsky, M.L. Thyroid hormones, t3 and t4, in the brain. Front Endocrinol (Lausanne). 2014, 5, 40. [Google Scholar] [CrossRef]
- Refetoff, S. Thyroid Hormone Serum Transport Proteins. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E, et al., editors. Endotext. South Dartmouth (MA): MDText.com, Inc.
- Copyright © 2000-2025, MDText.com, Inc.; 2000.
- Sanguinetti, C.; Minniti, M.; Susini, V.; Caponi, L.; Panichella, G.; Castiglione, V.; et al. The Journey of Human Transthyretin: Synthesis, Structure Stability, and Catabolism. Biomedicines. 2022;10(8).
- Landers, K.; Richard, K. Traversing barriers - How thyroid hormones pass placental, blood-brain and blood-cerebrospinal fluid barriers. Mol Cell Endocrinol. 2017, 458, 22–8. [Google Scholar] [CrossRef]
- Lisowska-Myjak, B.; Strawa, A.; Zborowska, H.; Jakimiuk, A.; Skarzynska, E. Associations between the thyroid panel and serum protein concentrations across pregnancy. Sci Rep. 2021, 11, 15970. [Google Scholar] [CrossRef]
- Peeters, R.P.; Visser, T.J. Metabolism of Thyroid Hormone. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E, et al., editors. Endotext. South Dartmouth (MA): MDText.com, Inc.; 2000.
- Russo, S.C.; Salas-Lucia, F.; Bianco, A.C. Deiodinases and the Metabolic Code for Thyroid Hormone Action. Endocrinology. 2021, 162, bqab059. [Google Scholar] [CrossRef]
- Gereben, B.; Zavacki, A.M.; Ribich, S.; Kim, B.W.; Huang, S.A.; Simonides, W.S.; et al. Cellular and molecular basis of deiodinase-regulated thyroid hormone signaling. Endocr Rev. 2008, 29, 898–938. [Google Scholar] [CrossRef]
- Patel, J.; Landers, K.; Li, H.; Mortimer, R.H.; Richard, K. Delivery of maternal thyroid hormones to the fetus. Trends Endocrinol Metab. 2011, 22, 164–70. [Google Scholar] [CrossRef]
- Loubiere, L.S.; Vasilopoulou, E.; Bulmer, J.N.; Taylor, P.M.; Stieger, B.; Verrey, F.; et al. Expression of thyroid hormone transporters in the human placenta and changes associated with intrauterine growth restriction. Placenta. 2010, 31, 295–304. [Google Scholar] [CrossRef]
- Adu-Gyamfi, E.A.; Wang, Y.X.; Ding, Y.B. The interplay between thyroid hormones and the placenta: a comprehensive reviewdagger. Biol Reprod. 2020, 102, 8–17. [Google Scholar] [CrossRef]
- Gutierrez-Vega, S.; Armella, A.; Mennickent, D.; Loyola, M.; Covarrubias, A.; Ortega-Contreras, B.; et al. High levels of maternal total tri-iodothyronine, and low levels of fetal free L-thyroxine and total tri-iodothyronine, are associated with altered deiodinase expression and activity in placenta with gestational diabetes mellitus. PLoS One. 2020, 15, e0242743. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Gutierrez, E.; Veas, C.; Leiva, A.; Escudero, C.; Sobrevia, L. Is a low level of free thyroxine in the maternal circulation associated with altered endothelial function in gestational diabetes? Front Pharmacol. 2014, 5, 136. [Google Scholar]
- Brent, G.A. Mechanisms of thyroid hormone action. J Clin Invest. 2012, 122, 3035–43. [Google Scholar] [CrossRef] [PubMed]
- Pagnin, M.; Kondos-Devcic, D.; Chincarini, G.; Cumberland, A.; Richardson, S.J.; Tolcos, M. Role of thyroid hormones in normal and abnormal central nervous system myelination in humans and rodents. Front Neuroendocrinol. 2021, 61, 100901. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Leonard, J.L.; Davis, P.J. Molecular aspects of thyroid hormone actions. Endocr Rev. 2010, 31, 139–70. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Goglia, F.; Leonard, J.L. Nongenomic actions of thyroid hormone. Nat Rev Endocrinol. 2016, 12, 111–21. [Google Scholar] [CrossRef]
- Lin, H.Y.; Su, Y.F.; Hsieh, M.T.; Lin, S.; Meng, R.; London, D.; et al. Nuclear monomeric integrin alphav in cancer cells is a coactivator regulated by thyroid hormone. FASEB J. 2013, 27, 3209–16. [Google Scholar] [CrossRef]
- Lin, H.Y.; Sun, M.; Tang, H.Y.; Lin, C.; Luidens, M.K.; Mousa, S.A.; et al. L-Thyroxine vs. 3,5,3'-triiodo-L-thyronine and cell proliferation: activation of mitogen-activated protein kinase and phosphatidylinositol 3-kinase. Am J Physiol Cell Physiol. 2009, 296, C980–91. [Google Scholar]
- Davis, P.J.; Davis, F.B.; Mousa, S.A.; Luidens, M.K.; Lin, H.Y. Membrane receptor for thyroid hormone: physiologic and pharmacologic implications. Annu Rev Pharmacol Toxicol. 2011, 51, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Feng, X.; Lin, Z.; Li, X.; Su, S.; Cheng, H.; et al. Thyroid hormone transport and metabolism are disturbed in the placental villi of miscarriage. Reprod Biol Endocrinol. 2023, 21, 108. [Google Scholar] [CrossRef]
- Anyetei-Anum, C.S.; Roggero, V.R.; Allison, L.A. Thyroid hormone receptor localization in target tissues. J Endocrinol. 2018, 237, R19–R34. [Google Scholar] [CrossRef]
- Chen, Z.; van der Sman, A.S.E.; Groeneweg, S.; de Rooij, L.J.; Visser, W.E.; Peeters, R.P.; Meima, M.E. Thyroid Hormone Transporters in a Human Placental Cell Model. Thyroid. 2022, 32, 1129–37. [Google Scholar] [CrossRef]
- Knabl, J.; de Maiziere, L.; Huttenbrenner, R.; Hutter, S.; Juckstock, J.; Mahner, S.; et al. Cell Type- and Sex-Specific Dysregulation of Thyroid Hormone Receptors in Placentas in Gestational Diabetes Mellitus. Int J Mol Sci. 2020;21(11).
- Barber, K.J.; Franklyn, J.A.; McCabe, C.J.; Khanim, F.L.; Bulmer, J.N.; Whitley, G.S.; Kilby, M.D. The in vitro effects of triiodothyronine on epidermal growth factor-induced trophoblast function. J Clin Endocrinol Metab. 2005, 90, 1655–61. [Google Scholar] [CrossRef]
- Huang, S.A.; Dorfman, D.M.; Genest, D.R.; Salvatore, D.; Larsen, P.R. Type 3 iodothyronine deiodinase is highly expressed in the human uteroplacental unit and in fetal epithelium. J Clin Endocrinol Metab. 2003, 88, 1384–8. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.; Kachilele, S.; Hobbs, E.; Bulmer, J.N.; Boelaert, K.; McCabe, C.J.; et al. Placental iodothyronine deiodinase expression in normal and growth-restricted human pregnancies. J Clin Endocrinol Metab. 2003, 88, 4488–95. [Google Scholar] [CrossRef]
- Landers, K.A.; Li, H.; Subramaniam, V.N.; Mortimer, R.H.; Richard, K. Transthyretin-thyroid hormone internalization by trophoblasts. Placenta. 2013, 34, 716–8. [Google Scholar] [CrossRef] [PubMed]
- Muhr, J.; Arbor, T.C.; Ackerman, K.M. Embryology, Gastrulation. StatPearls. Treasure Island (FL) 2024.
- Visciano, C.; Prevete, N.; Liotti, F.; Marone, G. Tumor-Associated Mast Cells in Thyroid Cancer. International Journal of Endocrinology. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Stiles, J.; Jernigan, T.L. The basics of brain development. Neuropsychol Rev. 2010, 20, 327–48. [Google Scholar] [CrossRef]
- Maenhaut, C.; Christophe, D.; Vassart, G.; Dumont, J.; Roger, P.P.; Opitz, R. Ontogeny, Anatomy, Metabolism and Physiology of the Thyroid. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E, et al., editors. Endotext. South Dartmouth (MA): MDText.com, Inc.; 2000.
- Huget-Penner, S.; Feig, D.S. Maternal thyroid disease and its effects on the fetus and perinatal outcomes. Prenatal Diagnosis. 2020, 40, 1077–84. [Google Scholar] [CrossRef] [PubMed]
- Rousset, B.; Dupuy, C.; Miot, F.; Dumont, J. Chapter 2 Thyroid Hormone Synthesis And Secretion. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E, et al., editors. Endotext. South Dartmouth (MA): MDText.com, Inc.; 2000.
- Goldsmit, G.S.; Valdes, M.; Herzovich, V.; Rodriguez, S.; Chaler, E.; Golombek, S.G.; Iorcansky, S. Evaluation and clinical application of changes in thyroid hormone and TSH levels in critically ill full-term newborns. Journal of Perinatal Medicine. 2011;39(1).
- La Gamma, E.F.; Paneth, N. Clinical importance of hypothyroxinemia in the preterm infant and a discussion of treatment concerns. Current Opinion in Pediatrics. 2012, 24, 172–80. [Google Scholar] [CrossRef]
- Ballabio, M.; Nicolini, U.; Jowett, T.; De Elvira, M.C.R.; Ekins, R.P.; Rodeck, C.H. MATURATION OF THYROID FUNCTION IN NORMAL HUMAN FOETUSES. Clinical Endocrinology. 1989, 31, 565–72. [Google Scholar] [CrossRef]
- Simpson, J.; Williams, F.L.R.; Delahunty, C.; Van Toor, H.; Wu, S.Y.; Ogston, S.A.; et al. Serum Thyroid Hormones in Preterm Infants and Relationships to Indices of Severity of Intercurrent Illness. The Journal of Clinical Endocrinology & Metabolism. 2005, 90, 1271–9. [Google Scholar]
- Bernal, J. Thyroid Hormones and Brain Development. Vitamins & Hormones. 71: Elsevier; 2005. p. 95-122.
- Rovet, J.F. The Role of Thyroid Hormones for Brain Development and Cognitive Function. In: Szinnai G, editor. Endocrine Development. 26: S. Karger AG; 2014. p. 26-43.
- Moog, N.K.; Entringer, S.; Heim, C.; Wadhwa, P.D.; Kathmann, N.; Buss, C. Influence of maternal thyroid hormones during gestation on fetal brain development. Neuroscience. 2017, 342, 68–100. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Suzuki, M. Congenital Hypothyroidism and Brain Development: Association With Other Psychiatric Disorders. Front Neurosci. 2021, 15, 772382. [Google Scholar] [CrossRef]
- Opazo, M.C.; Gianini, A.; Pancetti, F.; Azkcona, G.; Alarcón, L.; Lizana, R.; et al. Maternal Hypothyroxinemia Impairs Spatial Learning and Synaptic Nature and Function in the Offspring. Endocrinology. 2008, 149, 5097–106. [Google Scholar] [CrossRef]
- Shallie, P.D.; Naicker, T. The placenta as a window to the brain: A review on the role of placental markers in prenatal programming of neurodevelopment. Intl J of Devlp Neuroscience. 2019, 73, 41–9. [Google Scholar] [CrossRef]
- Sánchez, M.B.; Neira, F.J.; Moreno-Sosa, T.; Michel Lara, M.C.; Viruel, L.B.; Germanó, M.J.; et al. Placental leukocyte infiltration accompanies gestational changes induced by hyperthyroidism. Reproduction. 2023, 165, 235–48. [Google Scholar] [CrossRef]
- Marx, H.; Amin, P.; Lazarus, J.H. Hyperthyroidism and pregnancy. BMJ. 2008, 336, 663–7. [Google Scholar] [CrossRef]
- Delange, F. The disorders induced by iodine deficiency. Thyroid: Official Journal of the American Thyroid Association. 1994, 4, 107–28. [Google Scholar] [CrossRef] [PubMed]
- Delange, F.; Camus, M.; Ermans, A.M. Circulating thyroid hormones in endemic goiter. J Clin Endocrinol Metab. 1972, 34, 891–5. [Google Scholar] [CrossRef]
- Hercbergs, A. Clinical Implications and Impact of Discovery of the Thyroid Hormone Receptor on Integrin alphavbeta3-A Review. Front Endocrinol (Lausanne). 2019, 10, 565. [Google Scholar] [CrossRef] [PubMed]
- Gowachirapant, S.; Jaiswal, N.; Melse-Boonstra, A.; Galetti, V.; Stinca, S.; Mackenzie, I.; et al. Effect of iodine supplementation in pregnant women on child neurodevelopment: a randomised, double-blind, placebo-controlled trial. The Lancet Diabetes & Endocrinology. 2017, 5, 853–63. [Google Scholar]
- Dineva, M.; Fishpool, H.; Rayman, M.P.; Mendis, J.; Bath, S.C. Systematic review and meta-analysis of the effects of iodine supplementation on thyroid function and child neurodevelopment in mildly-to-moderately iodine-deficient pregnant women. Am J Clin Nutr. 2020, 112, 389–412. [Google Scholar] [CrossRef]
- Pedersen, K.M.; Laurberg, P.; Iversen, E.; Knudsen, P.R.; Gregersen, H.E.; Rasmussen, O.S.; et al. Amelioration of some pregnancy-associated variations in thyroid function by iodine supplementation. J Clin Endocrinol Metab. 1993, 77, 1078–83. [Google Scholar]
- Silva, J.E.; Silva, S. Interrelationships among serum thyroxine, triiodothyronine, reverse triiodothyronine, and thyroid-stimulating hormone in iodine-deficient pregnant women and their offspring: effects of iodine supplementation. J Clin Endocrinol Metab. 1981, 52, 671–7. [Google Scholar] [CrossRef]
- Glinoer, D.; Delange, F.; Laboureur, I.; de Nayer, P.; Lejeune, B.; Kinthaert, J.; Bourdoux, P. Maternal and neonatal thyroid function at birth in an area of marginally low iodine intake. J Clin Endocrinol Metab. 1992, 75, 800–5. [Google Scholar]
- Knudsen, N.; Bülow, I.; Laurberg, P.; Ovesen, L.; Perrild, H.; Jørgensen, T. Parity is associated with increased thyroid volume solely among smokers in an area with moderate to mild iodine deficiency. Eur J Endocrinol. 2002, 146, 39–43. [Google Scholar] [CrossRef]
- Rotondi, M.; Sorvillo, F.; Mazziotti, G.; Balzano, S.; Iorio, S.; Savoia, A.; et al. The influence of parity on multinodular goiter prevalence in areas with moderate iodine deficiency. J Endocrinol Invest. 2002, 25, 442–6. [Google Scholar] [CrossRef]
- Andersson, M.; Bath, S.; Farrand, C.; Gerasimov, G.; Moreno-Reyes, R. Prevention and control of iodine deficiency in the WHO European Region: adapting to changes in diet and lifestyle. WHO Regional Office for Europe; 2024.
- Zimmermann, M.; Trumbo, P.R. Iodine. Adv Nutr. 2013, 4, 262–4. [Google Scholar] [CrossRef]
- Gyamfi, D.; Wiafe, Y.A.; Danquah, K.O.; Adankwah, E.; Amissah, G.A.; Odame, A. Urinary iodine concentration and thyroid volume of pregnant women attending antenatal care in two selected hospitals in Ashanti Region, Ghana: a comparative cross-sectional study. BMC Pregnancy Childbirth. 2018, 18, 166. [Google Scholar] [CrossRef]
- Lazarus, J. Thyroid Regulation and Dysfunction in the Pregnant Patient. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E, et al., editors. Endotext. South Dartmouth (MA)2000.
- Zimmermann, M.B.; Boelaert, K. Iodine deficiency and thyroid disorders. The Lancet Diabetes & Endocrinology. 2015, 3, 286–95. [Google Scholar]
- Visser, W.E.; Peeters, R.P. Interpretation of thyroid function tests during pregnancy. Best Pract Res Clin Endocrinol Metab. 2020, 34, 101431. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Burgi, H.; Hurrell, R.F. Iron deficiency predicts poor maternal thyroid status during pregnancy. J Clin Endocrinol Metab. 2007, 92, 3436–40. [Google Scholar] [CrossRef]
- Zimmermann, M.; Adou, P.; Torresani, T.; Zeder, C.; Hurrell, R. Persistence of goiter despite oral iodine supplementation in goitrous children with iron deficiency anemia in Côte d'Ivoire. Am J Clin Nutr. 2000, 71, 88–93. [Google Scholar] [CrossRef]
- Moreno-Reyes, R.; Corvilain, B.; Daelemans, C.; Wolff, F.; Fuentes Peña, C.; Vandevijvere, S. Iron Deficiency Is a Risk Factor for Thyroid Dysfunction During Pregnancy: A Population-Based Study in Belgium. Thyroid: Official Journal of the American Thyroid Association. 2021, 31, 1868–77. [Google Scholar] [CrossRef]
- Hess, S.Y.; Zimmermann, M.B.; Arnold, M.; Langhans, W.; Hurrell, R.F. Iron deficiency anemia reduces thyroid peroxidase activity in rats. J Nutr. 2002, 132, 1951–5. [Google Scholar] [CrossRef]
- Vejbjerg, P.; Knudsen, N.; Perrild, H.; Laurberg, P.; Andersen, S.; Rasmussen, L.B.; et al. Estimation of iodine intake from various urinary iodine measurements in population studies. Thyroid: Official Journal of the American Thyroid Association. 2009, 19, 1281–6. [Google Scholar] [CrossRef]
- Glinoer, D. The regulation of thyroid function in pregnancy: pathways of endocrine adaptation from physiology to pathology. Endocr Rev. 1997, 18, 404–33. [Google Scholar] [CrossRef]
- Sparre, L.S.; Brundin, J.; Carlström, K.; Carlström, A. Oestrogen and thyroxine-binding globulin levels in early normal pregnancy. Acta Endocrinol (Copenh). 1987, 114, 298–304. [Google Scholar] [CrossRef]
- Skjöldebrand, L.; Brundin, J.; Carlström, A.; Pettersson, T. Thyroid associated components in serum during normal pregnancy. Acta Endocrinol (Copenh). 1982, 100, 504–11. [Google Scholar] [CrossRef]
- Korevaar, T.I.M.; Medici, M.; Visser, T.J.; Peeters, R.P. Thyroid disease in pregnancy: new insights in diagnosis and clinical management. Nat Rev Endocrinol. 2017, 13, 610–22. [Google Scholar] [CrossRef]
- Glinoer, D.; de Nayer, P.; Bourdoux, P.; Lemone, M.; Robyn, C.; van Steirteghem, A.; et al. Regulation of maternal thyroid during pregnancy. J Clin Endocrinol Metab. 1990, 71, 276–87. [Google Scholar] [CrossRef]
- Moreno-Reyes, R.; Glinoer, D.; Van Oyen, H.; Vandevijvere, S. High prevalence of thyroid disorders in pregnant women in a mildly iodine-deficient country: a population-based study. J Clin Endocrinol Metab. 2013, 98, 3694–701. [Google Scholar] [CrossRef]
- Glinoer, D.; De Nayer, P.; Delange, F.; Lemone, M.; Toppet, V.; Spehl, M.; et al. A randomized trial for the treatment of mild iodine deficiency during pregnancy: maternal and neonatal effects. J Clin Endocrinol Metab. 1995, 80, 258–69. [Google Scholar]
- Thilly, C.H.; Vanderpas, J.B.; Bebe, N.; Ntambue, K.; Contempre, B.; Swennen, B.; et al. Iodine deficiency, other trace elements, and goitrogenic factors in the etiopathogeny of iodine deficiency disorders (IDD). Biol Trace Elem Res. 1992, 32, 229–43. [Google Scholar] [CrossRef]
- Meinhold, H.; Dudenhausen, J.W.; Wenzel, K.W.; Saling, E. Amniotic fluid concentrations of 3,3',5'-tri-iodothyronine (reverse T3), 3,3'-di-iodothyronine, 3,5,3'-tri-iodothyronine (T3) and thyroxine (T4) in normal and complicated pregnancy. Clinical Endocrinology. 1979, 10, 355–65. [Google Scholar] [CrossRef]
- Weeke, J.; Dybkjaer, L.; Granlie, K.; Eskjaer Jensen, S.; Kjaerulff, E.; Laurberg, P.; Magnusson, B. A longitudinal study of serum TSH, and total and free iodothyronines during normal pregnancy. Acta Endocrinol (Copenh). 1982, 101, 531–7. [Google Scholar] [CrossRef]
- Glinoer, D.; Lemone, M.; Bourdoux, P.; De Nayer, P.; DeLange, F.; Kinthaert, J.; LeJeune, B. Partial reversibility during late postpartum of thyroid abnormalities associated with pregnancy. J Clin Endocrinol Metab. 1992, 74, 453–7. [Google Scholar]
- Rotondi, M.; Amato, G.; Biondi, B.; Mazziotti, G.; Del Buono, A.; Rotonda Nicchio, M.; et al. Parity as a thyroid size-determining factor in areas with moderate iodine deficiency. J Clin Endocrinol Metab. 2000, 85, 4534–7. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Aeberli, I.; Andersson, M.; Assey, V.; Yorg, J.A.J.; Jooste, P.; et al. Thyroglobulin is a sensitive measure of both deficient and excess iodine intakes in children and indicates no adverse effects on thyroid function in the UIC range of 100-299 μg/L: a UNICEF/ICCIDD study group report. J Clin Endocrinol Metab. 2013, 98, 1271–80. [Google Scholar] [CrossRef] [PubMed]
- Mullan, K.; McMullan, P.; Kayes, L.; McCance, D.; Hunter, A.; Woodside, J.V. Thyroglobulin levels among iodine deficient pregnant women living in Northern Ireland. Eur J Clin Nutr. 2022, 76, 1542–7. [Google Scholar] [CrossRef] [PubMed]
- Morreale de Escobar, G.; Pastor, R.; Obregon, M.J.; Escobar del Rey, F. Effects of maternal hypothyroidism on the weight and thyroid hormone content of rat embryonic tissues, before and after onset of fetal thyroid function. Endocrinology. 1985, 117, 1890–900. [Google Scholar] [CrossRef]
- Contempré, B.; Jauniaux, E.; Calvo, R.; Jurkovic, D.; Campbell, S.; de Escobar, G.M. Detection of thyroid hormones in human embryonic cavities during the first trimester of pregnancy. J Clin Endocrinol Metab. 1993, 77, 1719–22. [Google Scholar] [PubMed]
- Sava, L.; Tomaselli, L.; Runello, F.; Belfiore, A.; Vigneri, R. Serum thyroglobulin levels are elevated in newborns from iodine-deficient areas. J Clin Endocrinol Metab. 1986, 62, 429–32. [Google Scholar] [CrossRef]
- Zimmermann, M.B. Iodine deficiency. Endocr Rev. 2009, 30, 376–408. [Google Scholar] [CrossRef]
- PHAROAHPOD; CONNOLLYKJ; EKINSRP; HARDINGAG MATERNAL THYROID HORMONE LEVELS IN PREGNANCY AND THE SUBSEQUENT COGNITIVE AND MOTOR PERFORMANCE OF THE CHILDREN. Clinical Endocrinology. 1984, 21, 265–70. [CrossRef]
- PHAROAHPOD; CONNOLLYKJ A Controlled Trial of lodinated Oil for the Prevention of Endemic Cretinism: A Long-Term Follow-Up. International Journal of Epidemiology. 1987, 16, 68–73. [CrossRef]
- Pharoah, P.O.; Connolly, K.J. Effects of maternal iodine supplementation during pregnancy. Archives of Disease in Childhood. 1991, 66, 145–7. [Google Scholar] [CrossRef]
- Kampouri, M.; Margetaki, K.; Koutra, K.; Kyriklaki, A.; Daraki, V.; Roumeliotaki, T.; et al. Urinary iodine concentrations in preschoolers and cognitive development at 4 and 6 years of age, the Rhea mother-child cohort on Crete, Greece. Journal of Trace Elements in Medicine and Biology. 2024, 85, 127486. [Google Scholar] [CrossRef]
- Thilly, C.H.; Swennen, B.; Bourdoux, P.; Ntambue, K.; Moreno-Reyes, R.; Gillies, J.; Vanderpas, J.B. The epidemiology of iodine-deficiency disorders in relation to goitrogenic factors and thyroid-stimulating-hormone regulation. Am J Clin Nutr. 1993;57(2 Suppl):267S-70S.
- Moreno-Reyes, R.; Boelaert, M.; el Badawi, S.; Eltom, M.; Vanderpas, J.B. Endemic juvenile hypothyroidism in a severe endemic goitre area of Sudan. Clinical Endocrinology. 1993, 38, 19–24. [Google Scholar] [CrossRef]
- de Souza Dos Santos, M.C.; Gonçalves, C.F.L.; Vaisman, M.; Ferreira, A.C.F.; de, C.a.r.v.a.l.h.o.DP. Impact of flavonoids on thyroid function. Food Chem Toxicol. 2011, 49, 2495–502. [Google Scholar] [CrossRef] [PubMed]
- Corvilain, B.; Contempré, B.; Longombé, A.O.; Goyens, P.; Gervy-Decoster, C.; Lamy, F.; et al. Selenium and the thyroid: how the relationship was established. Am J Clin Nutr. 1993;57(2 Suppl):244S-8S.
- Contempré, B.; Duale, N.L.; Dumont, J.E.; Ngo, B.; Diplock, A.T.; Vanderpas, J. Effect of selenium supplementation on thyroid hormone metabolism in an iodine and selenium deficient population. Clinical Endocrinology. 1992, 36, 579–83. [Google Scholar] [CrossRef]
- La Merrill, M.A.; Vandenberg, L.N.; Smith, M.T.; Goodson, W.; Browne, P.; Patisaul, H.B.; et al. Consensus on the key characteristics of endocrine-disrupting chemicals as a basis for hazard identification. Nat Rev Endocrinol. 2020, 16, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Nascimento, C.; Nunes, M.T. Perchlorate, nitrate, and thiocyanate: Environmental relevant NIS-inhibitors pollutants and their impact on thyroid function and human health. Front Endocrinol (Lausanne). 2022, 13, 995503. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.M. Early-life exposure to EDCs: role in childhood obesity and neurodevelopment. Nat Rev Endocrinol. 2017, 13, 161–73. [Google Scholar] [CrossRef]
- Toledano, J.M.; Puche-Juarez, M.; Moreno-Fernandez, J.; Gonzalez-Palacios, P.; Rivas, A.; Ochoa, J.J.; Diaz-Castro, J. Implications of Prenatal Exposure to Endocrine-Disrupting Chemicals in Offspring Development: A Narrative Review. Nutrients. 2024;16(11).
- Brambilla, M.M.; Perrone, S.; Shulhai, A.M.; Ponzi, D.; Paterlini, S.; Pisani, F.; et al. Systematic review on Endocrine Disrupting Chemicals in breastmilk and neuro-behavioral development: Insight into the early ages of life. Neurosci Biobehav Rev. 2025, 169, 106028. [Google Scholar] [CrossRef]
- Taylor, P.N.; Okosieme, O.E.; Murphy, R.; Hales, C.; Chiusano, E.; Maina, A.; et al. Maternal perchlorate levels in women with borderline thyroid function during pregnancy and the cognitive development of their offspring: data from the Controlled Antenatal Thyroid Study. J Clin Endocrinol Metab. 2014, 99, 4291–8. [Google Scholar] [CrossRef]
- Li, J.; Tu, F.; Wan, Y.; Qian, X.; Mahai, G.; Wang, A.; et al. Associations of Trimester-Specific Exposure to Perchlorate, Thiocyanate, and Nitrate with Childhood Neurodevelopment: A Birth Cohort Study in China. Environ Sci Technol. 2023, 57, 20480–93. [Google Scholar] [CrossRef]
- Eng, P.H.; Cardona, G.R.; Fang, S.L.; Previti, M.; Alex, S.; Carrasco, N.; et al. Escape from the acute Wolff-Chaikoff effect is associated with a decrease in thyroid sodium/iodide symporter messenger ribonucleic acid and protein. Endocrinology. 1999, 140, 3404–10. [Google Scholar] [CrossRef]
- Markou, K.; Georgopoulos, N.; Kyriazopoulou, V.; Vagenakis, A.G. Iodine-Induced hypothyroidism. Thyroid: Official Journal of the American Thyroid Association. 2001, 11, 501–10. [Google Scholar] [CrossRef]
- Connelly, K.J.; Boston, B.A.; Pearce, E.N.; Sesser, D.; Snyder, D.; Braverman, L.E.; et al. Congenital hypothyroidism caused by excess prenatal maternal iodine ingestion. J Pediatr. 2012, 161, 760–2. [Google Scholar] [CrossRef]
- Chanoine, J.P.; Pardou, A.; Bourdoux, P.; Delange, F. Withdrawal of iodinated disinfectants at delivery decreases the recall rate at neonatal screening for congenital hypothyroidism. Arch Dis Child. 1988, 63, 1297–8. [Google Scholar] [CrossRef]
- Pedersen, I.B.; Knudsen, N.; Jørgensen, T.; Perrild, H.; Ovesen, L.; Laurberg, P. Thyroid peroxidase and thyroglobulin autoantibodies in a large survey of populations with mild and moderate iodine deficiency. Clinical Endocrinology. 2003, 58, 36–42. [Google Scholar] [CrossRef]
- Mosso, L.; Margozzini, P.; Trejo, P.; Domínguez, A.; Solari, S.; Valdivia, G.; Arteaga, E. [Thyroid stimulating hormone reference values derived from the 2009-2010 Chilean National Health Survey]. Rev Med Chil. 2013, 141, 95–103. [Google Scholar] [CrossRef]
- Shi, X.; Han, C.; Li, C.; Mao, J.; Wang, W.; Xie, X.; et al. Optimal and safe upper limits of iodine intake for early pregnancy in iodine-sufficient regions: a cross-sectional study of 7190 pregnant women in China. J Clin Endocrinol Metab. 2015, 100, 1630–8. [Google Scholar] [CrossRef]
- Mégier, C.; Dumery, G.; Luton, D. Iodine and Thyroid Maternal and Fetal Metabolism during Pregnancy. Metabolites. 2023, 13, 633. [Google Scholar] [CrossRef]
- Grossklaus, R.; Liesenkotter, K.P.; Doubek, K.; Volzke, H.; Gaertner, R. Iodine Deficiency, Maternal Hypothyroxinemia and Endocrine Disrupters Affecting Fetal Brain Development: A Scoping Review. Nutrients. 2023;15(10).
- Abel, M.H.; Caspersen, I.H.; Sengpiel, V.; Jacobsson, B.; Meltzer, H.M.; Magnus, P.; et al. Insufficient maternal iodine intake is associated with subfecundity, reduced foetal growth, and adverse pregnancy outcomes in the Norwegian Mother, Father and Child Cohort Study. BMC Med. 2020, 18, 211. [Google Scholar] [CrossRef]
- Skeaff, S.A. Iodine deficiency in pregnancy: the effect on neurodevelopment in the child. Nutrients. 2011, 3, 265–73. [Google Scholar] [CrossRef]
- Trumpff, C.; De Schepper, J.; Tafforeau, J.; Van Oyen, H.; Vanderfaeillie, J.; Vandevijvere, S. Mild iodine deficiency in pregnancy in Europe and its consequences for cognitive and psychomotor development of children: a review. J Trace Elem Med Biol. 2013, 27, 174–83. [Google Scholar] [CrossRef] [PubMed]
- Bougma, K.; Aboud, F.E.; Harding, K.B.; Marquis, G.S. Iodine and mental development of children 5 years old and under: a systematic review and meta-analysis. Nutrients. 2013, 5, 1384–416. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Synthesis of thyroid hormones. The stimulus starts from the hypothalamus with the production of TRH which promotes the production of TSH in the anterior pituitary. The thyroid is stimulated by TSH to synthesize T3 and T4 in the thyroid follicles and release it to circundant blood vessels. Created in BioRender.com.
Figure 1.
Synthesis of thyroid hormones. The stimulus starts from the hypothalamus with the production of TRH which promotes the production of TSH in the anterior pituitary. The thyroid is stimulated by TSH to synthesize T3 and T4 in the thyroid follicles and release it to circundant blood vessels. Created in BioRender.com.
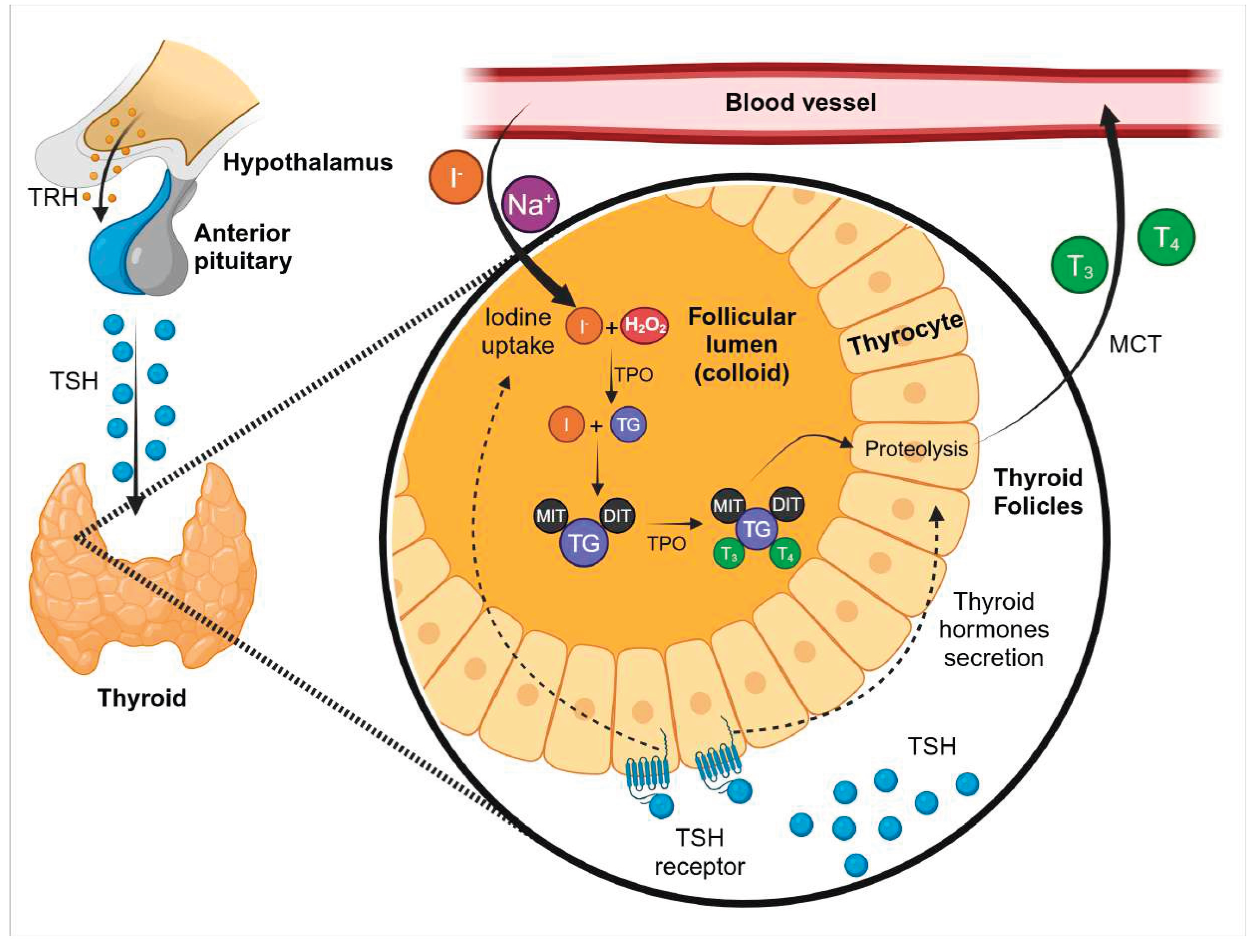
Figure 2.
Iodide metabolism in healthy non pregnant and pregnant women. (A) Non-pregnant woman with optimal iodine intake. (B) Non-pregnant with low iodine intake. (C) Pregnant women with low iodide intake.
Figure 2.
Iodide metabolism in healthy non pregnant and pregnant women. (A) Non-pregnant woman with optimal iodine intake. (B) Non-pregnant with low iodine intake. (C) Pregnant women with low iodide intake.
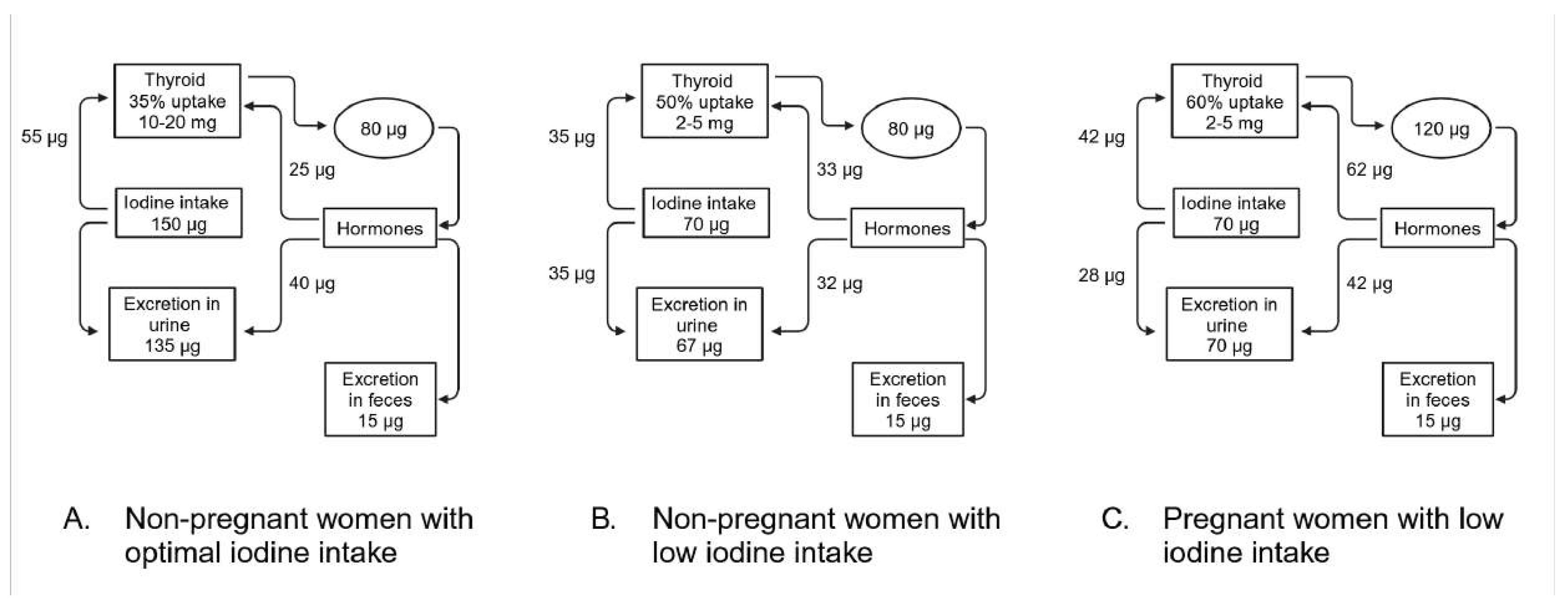
Figure 3.
Determinant of the risk of iodine deficiency in women.
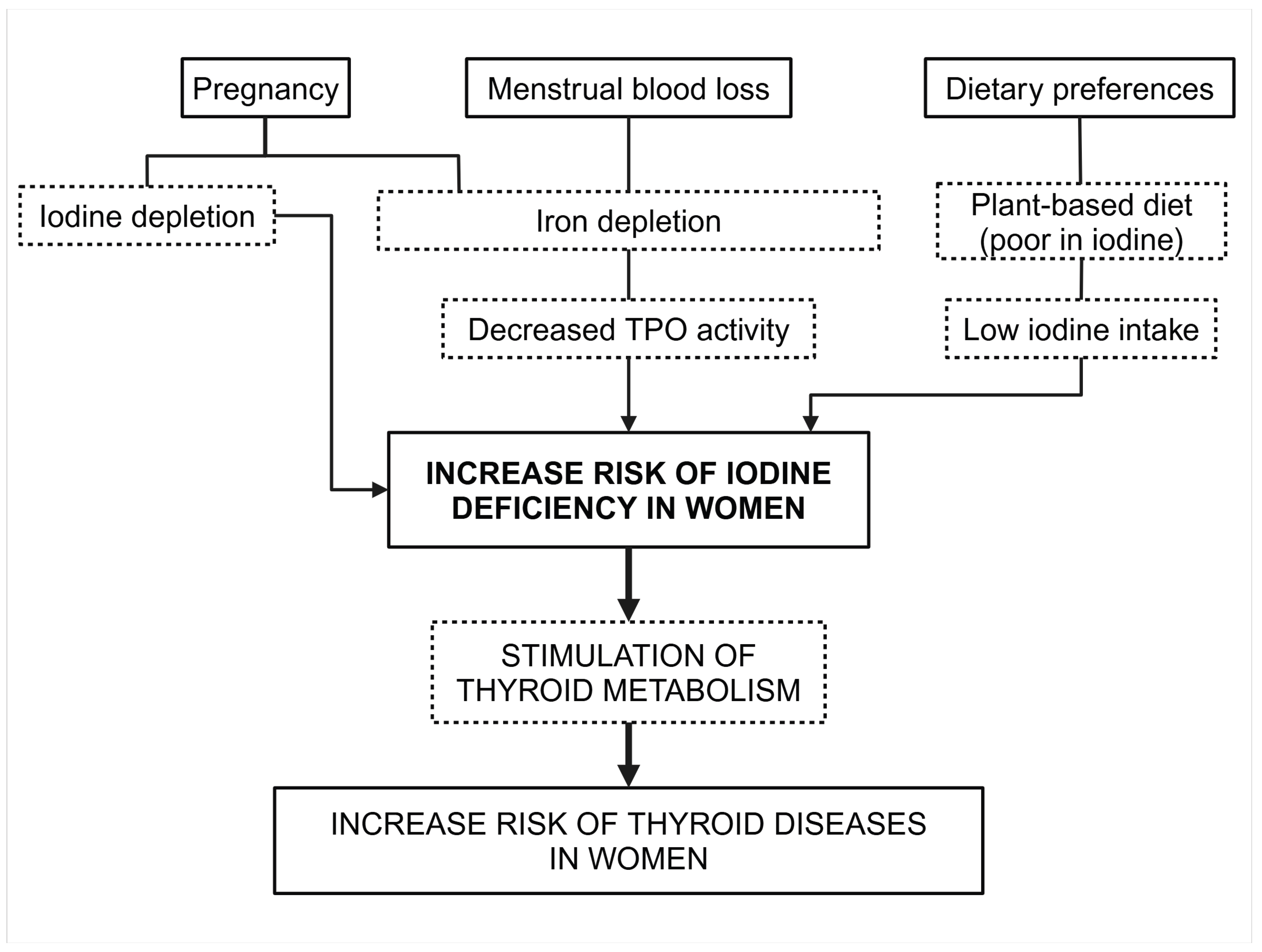
Figure 4.
Changes in thyroid physiology during pregnancy. The increase in hCG in the first week triggers an upregulation of TBG and free T4 and a decrease of TSH accordingly. The amount of free T4 decreases as the degradation by placental type 3 deiodinase increases. As the offspring thyroid is developed enough to produce it´s own hormones around week 20, the dependency on maternal thyroxine decreases at the same time.
Figure 4.
Changes in thyroid physiology during pregnancy. The increase in hCG in the first week triggers an upregulation of TBG and free T4 and a decrease of TSH accordingly. The amount of free T4 decreases as the degradation by placental type 3 deiodinase increases. As the offspring thyroid is developed enough to produce it´s own hormones around week 20, the dependency on maternal thyroxine decreases at the same time.
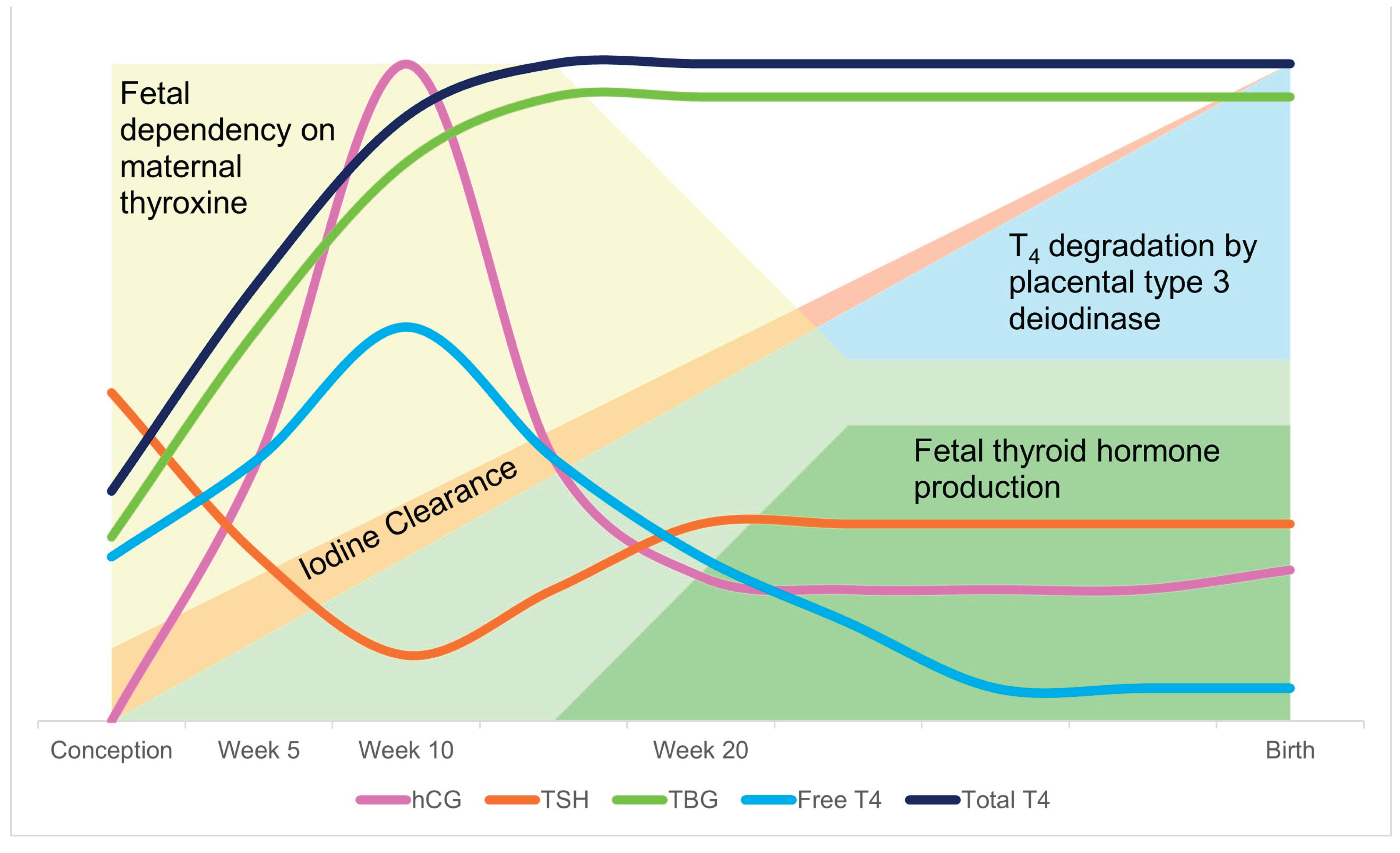
Figure 5.
Adaptation of the thyroid function during pregnancy in iodine sufficient and deficient conditions.
Figure 5.
Adaptation of the thyroid function during pregnancy in iodine sufficient and deficient conditions.
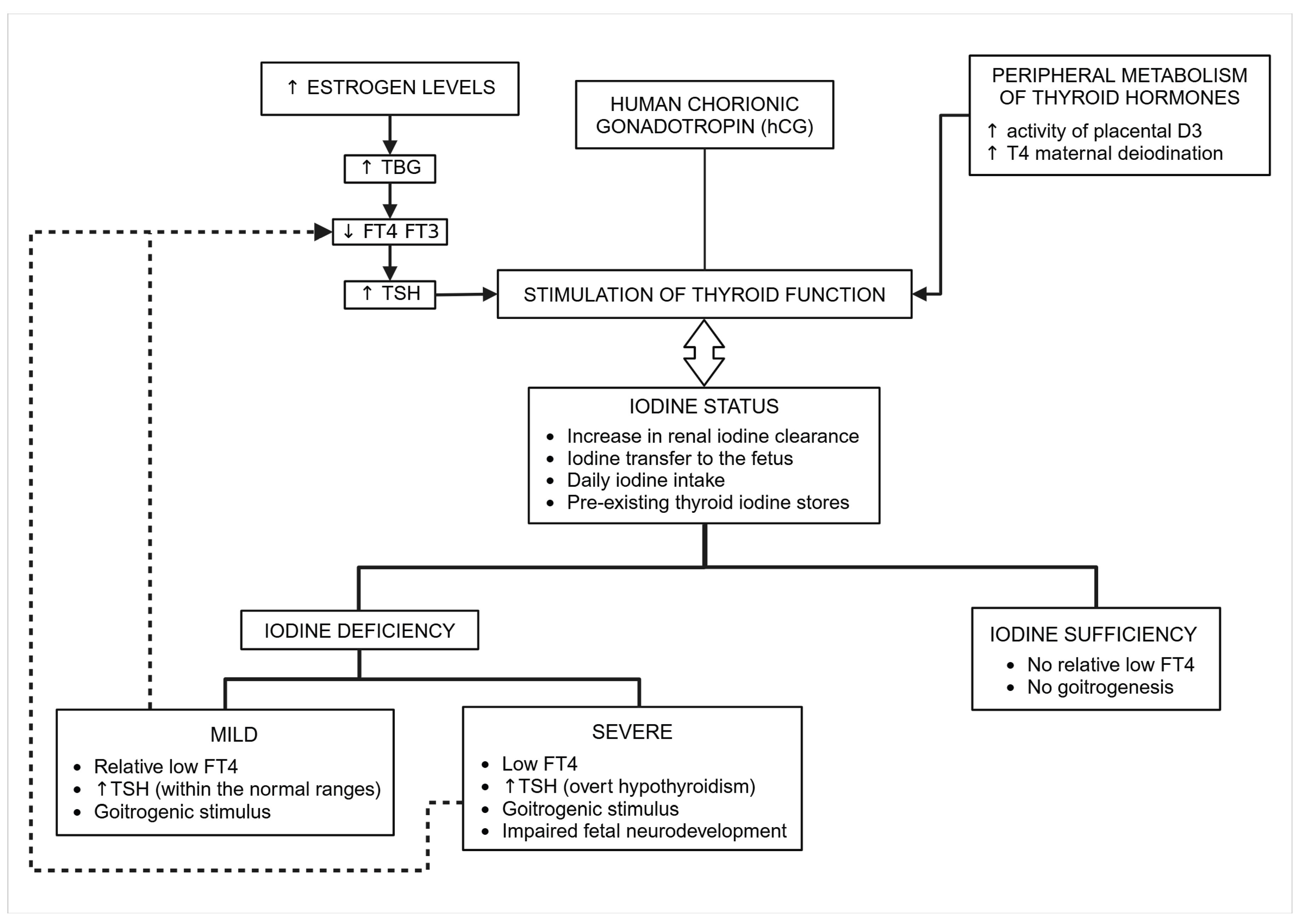
Figure 6.
Assessing iodine adequacy in pregnant women at an individual level.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



