
Submitted:
01 August 2025
Posted:
04 August 2025
You are already at the latest version
Abstract
CRABS CLAW (CRC) is a member of the plant-specific YABBY transcription factor family, defined by the presence of a C2C2 zinc-finger domain and a C-terminal YABBY domain. CRC is essential for proper floral development, functioning in the termination of the floral meristem, maintenance of adaxial–abaxial polarity within the gynoecium, and regulation of nectary and leaf morphogenesis.
CRC orchestrates its diverse regulatory functions through interaction networks comprising other transcription factors and plant developmental regulators, including chromatin-modifying enzymes and proteins involved in auxin bio-synthesis, transport, and signaling. The roles of genes and proteins interacting with CRC have been characterized in several model plant species, and the number of identified CRC-associated interactions continues to expand, revealing both species-specific and conserved functional roles across angiosperms.
Many functions of CRC and its interacting partners have been elucidated through the analysis of anatomical and phy-siological phenotypes associated with specific gene mutations. The functional roles of CRC in plant development ap-pear to have been acquired progressively through evolutionary diversification. These evolutionary changes have been associated with the relative conservation of CRC gene copy number and a predominant role of mutations occurring in non-coding regulatory regions. These properties are attributed to the relatively limited number of genes comprising the CRC regulatory network and the capacity to induce dosage-dependent effects via the emergence of novel proteins with overlapping or analogous functions.
The identification and functional characterization of CRC transcription factors across diverse plant species has advanced rapidly in recent years, yet a comprehensive synthesis of these findings has not been presented in a dedicated article. Therefore, this study reviews the current knowledge on CRC transcription factors, with a focus on their iden-tification, expression patterns, and functional roles in plant development.
Keywords:
CRABS CLAW
; gynoecium
; floral meristem
; gene and protein interactions
1. Introduction
The CRABS CLAW (CRC) gene belongs to the YABBY transcription factor family and plays a pivotal role in meristem termination, carpel development, and nectary formation in plants [1,2,3,4,5,6]. Its ortholog in the monocot Oryza sativa, known as DROOPING LEAF (DL), regulates not only carpel identity but also the development of the leaf midrib [7,8,9].
The termination of plant meristem activity requires repression of the WUSCHEL (WUS) gene, which is achieved through two feedback loop systems [10,11]. In one of these loops, WUS is initially repressed either directly by AGAMOUS (AG) or indirectly via the AG-dependent activation of KNUCKLES (KNU) during early stages of floral development [10,11]. This repression is subsequently enhanced by the action of additional regulatory proteins, including CRC and TORNADO2 (TRN2), which contribute to the maintenance of auxin maxima necessary for proper meristem termination and gynoecium development [12,13,14,15]. Simultaneously, auxin represses cytokinin biosynthesis and signaling, a regulatory interaction essential for proper floral meristem determinacy [16].
CRABS CLAW (CRC) and AG regulate the expression of YUC4, a gene involved in the two-step conversion of tryptophan to indole-3-acetic acid (IAA), a key process in auxin biosynthesis [12,13,17,18]. Elevated local auxin concentrations resulting from YUC4 activation promote cell wall loosening and cell proliferation, particularly within the medial domain of the developing gynoecium [12,13,17,18].
The regulation of auxin biosynthesis and very-long-chain fatty acid (VLCFA) metabolism by Arabidopsis thaliana CRABS CLAW (AtCRC) appears to be functionally interconnected, although the precise molecular mechanisms governing both processes remain incompletely understood [12,19]. Key genes associated with auxin metabolism have been identified in A. thalianaas critical components of CRC and AG regulatory networks involved in floral meristem termination [12,14]. These findings emerged also from transcriptomic analyses conducted in crc-1, knu-1, ag-12, and wild-type (WT) plants [13].
The scope of novel genes putatively affected by CRCa allele overexpression in tomato (Solanum lycopersicum) has been expanded to include 356 upregulated and 1,222 downregulated genes [20]. Comparable transcriptomic analyses in other plant species could further broaden the catalog of CRC-regulated targets, offering valuable insights into the diverse and evolutionarily conserved functions of CRC in plant development [21,22,23,24].
The mechanisms underlying meristem termination induced by high temperature appear to be species-specific [25]. In Arabidopsis thaliana, repression of WUS is mediated through auxin signaling and does not involve direct binding of AtCRC to the WUS gene [25,26]. In contrast, in Solanum lycopersicum (tomato), both SlCRCa and SlCRCb directly bind to the SlWUS promoter. Notably, SlCRCa recruits the SlIMA chromatin-remodeling complex to achieve complete inactivation of SlWUS expression, highlighting a mechanistic divergence in thermal response between species [25,26].
The observed results support the continued search for CRC homologs in other plant species and their functional characterization, with the aim of uncovering novel specificities and evolutionary diversification in CRC-mediated developmental processes [27,28,29,30].
The understanding of CRC functions has been broadened by studies on gene polymorphisms and spontaneous mutations [31,32]. Notably, a 365-bp insertion of a putative transposable element within the intron between exons 4 and 5 of the SlCRCa gene was identified as the molecular basis for aberrant carpel development, resulting in iterative fruit growth characteristic of the fig mutant phenotype in tomato [3].
Additionally, the rare CsCRCA allele, associated with shortened fruit length in cucumber (Cucumis sativus), was identified through quantitative trait loci (QTL) analysis and map-based cloning [27,31]. Furthermore, mutagenesis studies conducted on the crc-1 mutant background in Arabidopsis led to the discovery of novel genetic modifiers of the CRC mutant phenotype, including rebelote-1 (rbl-1), squint-4 (sqn-4), and ultrapetala1-4 (ult1-4) [33].
Due to the pivotal role of CRC in carpel formation and floral meristem termination, its expression is tightly regulated, predominantly by other transcription factors [30,34,35]. Yeast one-hybrid (Y1H) screening in Arabidopsis thaliana identified 140 proteins capable of binding to the CRC promoter [35]. While only 48 of these were annotated as transcription factors in the PlantPAN3 database, the remaining proteins may represent novel regulators potentially involved in modulating AtCRC expression [35,36,37].
Analysis of phylogenetic studies on CRC genes suggests their numerical stability across angiosperms [36]. This feature is relatively uncommon among evolutionarily younger genes and may be attributed to the compact size of the gene regulatory network involved in carpel development [38,39]. The introduction of novel genes with overlapping functions could disrupt this delicate balance, potentially causing dosage imbalances—particularly detrimental in processes such as the termination of reproductive meristems [33,38].
Ancestral CRC genes likely contributed to the polarity of the carpel—an organ unique to angiosperms—through subfunctionalization of the broader role of YABBY genes in establishing lateral organ polarity [1,40,41,42]. During angiosperm evolution, CRC expression and function underwent significant diversification, including the acquisition of roles in specifying carpel identity, the loss of polarity-regulating function in grass carpels, and the evolution of novel functions such as midrib development in grass leaves and nectary formation in core eudicots [1,4,7,8,9,34,42,43].
This review aims to synthesize current knowledge on the role of the CRC transcription factor in floral meristem termination and the development of carpels and nectaries. Particular emphasis is placed on the identification and characterization of CRC-associated gene and protein partners, highlighting their contributions to the regulatory networks that coordinate plant growth, organ specification, and reproductive development.
2. Results
2.1. Structural Features of the CRC Protein
The CRABS CLAW (CRC) transcription factor belongs to the YABBY family and exhibits characteristic structural elements, including a C2C2-type zinc finger motif spanning amino acid residues 26–53, which confers DNA, RNA, and protein-binding capabilities [4,44]. The CRC protein consists of 181 amino acids, with an intermediate segment located between residues 54–108 that likely functions as a flexible linker. This region contains a serine- and proline-rich domain at its center—a typical feature of transcription factor activation domains [45,46].
The YABBY domain of CRC is located between amino acid residues 109–155 and is composed of two short and two long α-helices. This domain shares structural similarity with helix–loop–helix motifs found in HMG-box-containing proteins and plays a critical role in DNA binding [4,44]. Proteins exhibiting this domain organization were classified in the late 1990s as members of a newly defined YABBY transcription factor family. The family was named after the Australian freshwater crayfish “yabby,” in reference to its founding member, CRABS CLAW (CRC) [4,47,48].
Subcellular localization studies using CRC deletion mutants suggest the presence of a putative nuclear localization signal (NLS) within the YABBY domain, specifically spanning amino acid residues 110–117 (Lys-Pro-Pro-Lys-Glu-Lys-Lys-Gln-Arg; KPPEKKQR). Despite the basic residue composition characteristic of classical NLSs, site-directed mutagenesis of this sequence does not abolish nuclear import, indicating that the NLS may exhibit non-canonical features [44,49]. This behavior is consistent with NLS motifs that interact with the minor binding site of importin α, as described for other atypical nuclear import signals [49,50,51].
The functional activity of transcription factors is frequently mediated not by monomeric forms but through the formation of homo- or heterodimers, which may assemble in the cytoplasm prior to nuclear translocation or upon binding to closely spaced cis-regulatory motifs within evolutionarily conserved promoter regions [52,53]. Dimerization is a well-documented feature among members of the YABBY transcription factor family [44,48]. In the case of CRC, bimolecular fluorescence complementation (BiFC) assays suggest that dimer formation is dependent on the presence of the YABBY domain [44].
However, these CRC–CRC or CRC–YABBY protein interactions appear to be relatively weak and could not be confirmed using yeast two-hybrid (Y2H) assays [44]. Interestingly, Y2H analyses have indicated a weak interaction between the transactivation domain of CRC and the YABBY domain of the INNER NO OUTER (INO) protein, suggesting possible functional crosstalk between these two transcription factors. CRC-interacting partners may exert either activating or repressive effects on CRC-mediated transcriptional regulation [44]. Further evidence from Y2H and bimolecular fluorescence complementation (BiFC) assays in Punica granatum (pomegranate) demonstrated that both CRC and INO interact with a common partner, PgBEL1, implicated in the maintenance of ovule identity [54].
The cooperative function of the YABBY domain and the zinc finger motif in DNA binding is supported by Y1H assays. These experiments demonstrated that neither the zinc finger domain nor the YABBY domain of CRC alone was sufficient to strongly activate reporter gene expression when targeting the promoter region of the KCS15 gene in yeast. However, Y1H screens using isolated or mutated CRC domains revealed that the YABBY domain plays a predominant role in DNA binding, while the zinc finger contributes a weaker, auxiliary interaction with the target DNA sequence [44].
In silico analysis of the regulatory mechanisms governing CRC expression suggests a predominant role for transcription factors, as indicated by the presence of only two CHH-context DNA methylation sites located approximately 0.3 to 3 kb upstream of the transcription start site (TSS). Furthermore, the CRC genomic locus appears to be largely unaffected by microRNA-mediated regulation and exhibits only moderate epigenetic control. This is supported by the presence of the repressive histone mark H3K27me3, which spans most of the CRC locus, including the promoter region, and by H2AK121ub enrichment within the transcribed region [35].
To identify putative transcription factors interacting with the CRC promoter, yeast Y1H screening was performed, yielding 140 candidate proteins. Of these, only 48 were annotated in the PlantPAN3 database and exhibited predicted DNA-binding motifs matching sequences within the CRC promoter region [35]. However, the inherently flexible nature of DNA-binding specificity in plant transcription factors allows them to recognize suboptimal or imperfectly matched cis-regulatory motifs, particularly within the context of chromatin architecture [55,56,57,58,59].
Therefore, among the remaining 92 proteins identified as interacting with the CRC promoter, it is likely that several represent biologically relevant DNA–protein interactions, potentially associated with authentic yet uncharacterized regulatory mechanisms governing CRC expression.
The majority of observed transcription factor binding sites were located within CRC promoter regions conserved among Brassicaceae species, suggesting their functional relevance [35]. Gene Ontology (GO) terms overexpressed among the putative CRC regulators were associated with known CRC functions, including abaxial cell fate specification, carpel development, meristem maintenance, and the regulation of flower development [1,3,5,6,35].
Among the proteins identified as binding to the CRC promoter in yeast, several transcription factors with established roles in carpel development were detected, including FRUITFUL (FUL), HALF FILLED (HAF), ETTIN (ETT), and AUXIN RESPONSE FACTOR 8 (ARF8). Additionally, transcription factors not previously associated with gynoecium development, such as WRKY41 and REVEILLE4, were also identified [35,36,37].
Co-expression analysis of CRC with other genes identified 555 TFs that are either positively or negatively correlated with CRC expression. Among these, 32 TFs were also found to bind the CRC promoter in Y1H assays, including KNOTTED-LIKE FROM ARABIDOPSIS THALIANA 1 (KNAT1), ETT, BHL9 (RPL), ULTRPETALA1 (ULT1), JAG, ARF8, FUL, NGA2, SEP3, HAF, INO, and CAL, thereby supporting the reliability of the Y1H screen [35]. Phenotypic analyses of single mutants for these genes, as well as their combinations with the crc background in Arabidopsis, may help to validate their proposed roles in meristem maintenance and carpel development.
2.2. Regulatory Roles of WUSCHEL, AGAMOUS, and CRABS CLAW in Floral Meristem Termination
Meristematic cells of the floral meristem (FM) originate from the shoot apical meristem (SAM), which remains active throughout the plant's life cycle and maintains a population of stem cells in its apex, referred to as the central zone (CZ) [60,61]. Cells in the CZ undergo slow divisions and are subsequently displaced laterally into the peripheral zone, developing further into lateral organs as flowers or leaves [60,61].
In Arabidopsis thaliana, the timing of floral meristem (FM) termination is primarily regulated by AGAMOUS (AG), which controls the expression of WUSCHEL (WUS), KNUCKLES (KNU), and CRABS CRC, through different feedback loop systems [25,62,63,64,65,66].
The first regulatory feedback loop involves WUS, which activates the expression of CLAVATA3 (CLV3) in CZ [59,65,66]. The activation of CLV3 transcription requires the physical interaction between WUS and members of the HAIRY MERISTEM (HAM) protein family [69]. Additionally, WUS binds to the CLV3 promoter as a heterodimer in association with SHOOT MERISTEMLESS (STM) [70].
In turn, CLV3 is perceived by receptor complexes composed of CLAVATA1 (CLV1), CLAVATA2 (CLV2), RECEPTOR-LIKE PROTEIN KINASE 2 (RPK2), CLAVATA3 INSENSITIVE RECEPTOR KINASES (CIKs), CORYNE (CRN), and BARELY ANY MERISTEMS (BAMs) [71]. These complexes mediate the repression of WUSCHEL (WUS) expression, thereby regulating stem cell maintenance in the shoot apical meristem [60,71,72].
The functions of CLV1and CLV3 are modulated by the heterotrimeric G-protein α-subunit CsGPA1 in cucumber (Cucumis sativus). CsGPA1 physically interacts with the kinase domain of CLV1, as demonstrated by Y2H and bimolecular fluorescence assays [73]. This interaction appears to be critical for the post-translational modification of CsGPA1 by CLV1 and for the regulation of WUS function. CRISPR/Cas9-generated mutants of CsGPA1 and CLV1exhibit increased floral organ number and reduced fruit length, indicating a disruption in meristem determinacy and floral development. Reverse transcription PCR (RT-PCR) analyses in these mutants suggest that the CsCLV1–CsGPA1 signaling pathway inhibits CsWUS expression while promoting the transcription of CsCRC, thereby coordinating meristem termination and carpel development in cucumber [73].
CLV1 and CLV3 mutants exhibit significantly enlarged floral meristems (FMs), characterized by the formation of one or two additional whorls of carpels at the flower center—nested within the primary carpels—as well as an increased number of primary floral organs across all whorls compared to wild-type plants [74,75]. These phenotypic alterations result from the failure to repress WUS expression, leading to an expanded WUS expression domain and delayed downregulation of WUS, which in turn promotes continued meristem activity and the formation of supernumerary carpels [71,75,76]. A related but independent feedback loop involves the CLE40 peptide, which is closely related to CLV3 and promotes WUS expression through the CLV1-family receptor BARELY ANY MERISTEM1 (BAM1) [77]. In turn, CLE40 expression is repressed in a WUS-dependent manner, establishing a regulatory circuit that contributes to meristem homeostasis [77].
Yeast two-hybrid (Y2H) screens, bimolecular fluorescence complementation (BiFC), and co-immunoprecipitation (Co-IP) assays using full-length coding sequences demonstrated that KNU physically interacts with full-length WUS, whereas no interaction was detected between KNU and CLV3 [63]. Analysis of protein deletion mutants demonstrated that the N-terminal region of KNU (amino acids 1–100), which includes the C2H2 zinc finger domain, is essential for its interaction with WUS. In WUSCHEL, the interaction is mediated by the N-terminal segment (amino acids 1–236), encompassing the homeodomain (HD), homeodomain adjacent domain (HOD), and the HAM-binding domain (HBD) [63].
Each of the three WUS domains—the HD,HOD, and HBD—is individually sufficient to mediate interaction with KNU [63]. The binding of KNU to WUS interferes with WUS dimerization, a process essential for repression of CLV3 expression. WUS monomers lack this repressive capacity. WUS dimerization occurs through interactions between its N-terminal regions, specifically involving the HD and HOD domains [63]. Additionally, HBD domain (amino acids 203–236) facilitates interaction with HAM proteins, a critical component for meristem maintenance. This WUS–HAM interaction is also inhibited by KNU. Conversely, the interaction between WUS and SHOOTMERISTEMLESS (STM), which reinforces CLV3 expression in SAM, occurs via the acidic domain of WUS (amino acids 229–249) and is not disrupted by KNU binding [63,78].
In both Arabidopsis thaliana and tomato (Solanum lycopersicum), KNU proteins are recruited to the WUS locus by adapter proteins—INHIBITOR OF MERISTEM ACTIVITY (SlIMA) in tomato and MINI ZINC FINGER2 (AtMIF2) in A. thaliana [26]. The C-terminal region of AtKNU contains a repressive EAR (Ethylene-Responsive Element Binding Factor-Associated Amphiphilic Repression) motif, which mediates the recruitment of the co-repressor TOPLESS (TPL) and HISTONE DEACETYLASE19 (HDA19). Together, these components form a transcriptional repressor complex that silences WUS expression [26]. It is postulated that both WUSCHEL (WUS) and CLAVATA3 (CLV3) are simultaneously repressed via KNUCKLES (KNU)-mediated H3K27me3 modification, as indicated by the enhanced meristem defects observed in transgenic plants harboring pCLV3:KNU and pWUS:KNU constructs, where KNU expression is driven by the CLV3 and WUS promoters, respectively [10].
Although a weak interaction between AtKNU and TOPLESS was confirmed by Y2H assays, the stabilization of this complex appears to require additional interaction with adapter proteins such as AtMIF2 in Arabidopsis or SlIMA in tomato [26,79]. This requirement is supported by yeast three-hybrid (Y3H) analyses, which suggest that these adapters enhance the AtKNU–TOPLESS association [26,80].
AGAMOUS (AG) plays a central role in the second feedback loop that regulates the termination of WUS expression and is overlayed on the CLV-WUS pathway [81]. Mutations in AG disrupt this regulation, leading to the failure of WUS repression and resulting in the formation of supernumerary floral whorls from the floral center. This produces a characteristic 'flower-within-a-flower' phenotype, also referred to as the Russian doll phenotype [82].
The mechanisms by which AG regulates WUS expression are dependent on the developmental stage of the floral bud. During floral stage 3, WUS promotes the expression of the C-class gene AG and LEAFY (LFY) [83]. Subsequently, AG represses WUS by recruiting the TERMINAL FLOWER 2 (TFL2) protein, a component of the Polycomb Repressive Complex 1 (PRC1) [10,84]. In a later developmental stage (stage 6), AG directly enhances the expression of the C2H2-type zinc finger protein KNU, initiating a two-step repression mechanism of WUS expression [11,26,85]. Initially, WUS expression is repressed through the association of KNU with a histone deacetylase complex [26]. This initial repression is subsequently stabilized by KNU-mediated recruitment of the Polycomb Repressive Complex 2 (PRC2), which catalyzes the trimethylation of lysine 27 on histone H3 (H3K27me3), a hallmark of stable gene silencing [11,86]. The recruitment of PRC2 by KNU occurs via direct interaction with FERTILIZATION-INDEPENDENT ENDOSPERM (FIE), a core component of PRC2 responsible for the enzymatic activity leading to H3K27me3 deposition [10].
The findings of Gómez-Mena et al. suggest that AG and its interacting partners—SEPALLATA3 (SEP3), APETALA3 (AP3), and PISTILLATA (PI)—are regulated in a coordinated positive feedback loop to sustain their own expression [87]. Within this model, AG promotes gibberellin biosynthesis, which may facilitate the transition from meristem identity to organ differentiation [87]. Another gene involved in the AG regulatory network is ULTRAPETALA1 (ULT1), which contributes to FM termination independently of LEAFY by regulating floral meristem identity and determinacy [88]. ULT1 functions to prevent the formation of supernumerary primary floral organs and additional carpel whorls, thereby contributing to the proper determinacy of the floral meristem [89].
SUPERMAN (SUP) also contributes to FM termination, acting synergistically with AG, although the precise nature of their functional interaction remains unclear [90]. It has been hypothesized that SUP promotes FM determinacy by regulating auxin biosynthesis [91]. According to Sakai et al., SUP expression is essential for maintaining the boundary between whorl 3 and whorl 4 [92].
It is plausible that CRC alone is not sufficient to modulate WUS activity, as suggested by analyses of CRC mutants across multiple species, including Arabidopsis thaliana, pomegranate, and rice, which exhibit no overt floral meristem (FM) defects [7,43,54]. In A. thaliana, CRC appears to play an ancillary role to AGAMOUS (AG) in FM development [34,43,87,93]. However, more direct and independent roles of CRC have been observed in tomato, where it contributes to high temperature-induced meristem termination [25].
In tomato, SlCRCa and SlCRCb directly bind to the SlWUS promoter, with SlCRCb also interacting with the second intron of SlWUS [25]. Interestingly, AtCRC does not bind to the AtWUS promoter (pAtWUS) or its genomic locus (gAtWUS), suggesting a species-specific regulatory mechanism [25].
Under heat shock conditions, a reduction in brassinosteroid concentrations within the floral meristem results in the persistent downregulation of SlCRCa [25]. This failure to terminate SlWUS expression and ultimately leads to the development of malformed fruits [25]. Although previous studies have demonstrated the critical role of tomato AG1 (TAG1) in activating SlKNU and SlIMA, leading to repression of WUS expression, overexpression of TAG1 alone is insufficient to activate the promoter activity of SlKNU and SlIMA. Therefore, in tomato, SlCRCa is necessary for TAG1-mediated activation of SlIMA and SlKNU [26].
Dual-luciferase assays suggest that this missing transcriptional activation may be provided by SlCRCa, but not by SlCRCb, indicating functional divergence between the two CRC homologs in tomato [26]. Moreover, SlCRCa directly binds to the SlWUS locus, where it recruits the SlIMA complex, leading to complete repression of SlWUS expression during the establishment of floral meristem determinacy [3].
2.3. Regulation of Floral Meristem Termination by AG and CRC via Auxin Metabolism Control
Further studies have demonstrated that the pivotal role of AG in FM termination and gynoecium development is mediated through its regulation of the crosstalk between cytokinin and auxin signaling pathways [16,94,95]. Similarly, findings by Yamaguchiet al. highlight the essential function of CRC in FM termination and gynoecium formation via the fine-tuning of auxin homeostasis [12,13,96]. Moreover, the the transcriptional repressor KNU integrates and modulates the activities of auxin and cytokinin, thus securing the timed FM termination [97].
The histone acetyltransferase (HAT) GENERAL CONTROL NON-REPRESSIBLE 5 (GCN5) participates in the activation of WUS expression in response to cytokinin signaling. Additionally, GCN5 is involved in the transcriptional activation of AGAMOUS (AG), KNUCKLES (KNU), and CRABS CLAW (CRC), suggesting its broader role in regulating key genes involved in floral meristem determinacy [98].
Although AG represses WUS directly or via KNU during floral stages 3–5, this repression is further reinforced at stage 6 by additional regulatory factors, including CRC and TORNADO2 (TRN2). CRC represses TRN2, leading to the formation of local auxin maxima [12,13]. Concurrently, AG and auxin synergistically activate ETT, which in turn downregulates the expression of ISOPENTENYL TRANSFERASES (IPTs), LONELY GUY (LOG), and ARABIDOPSIS HISTIDINE KINASE 4 (AHK4), thereby attenuating cytokinin signaling activity [16].
The pivotal role of auxin in the regulatory functions of CRC and AG during floral meristem termination has been elucidated through transcriptomic analyses of relevant mutants, including crc-1,knu-1 versus knu-1, and ag-12 versus wild-type plants. These comparisons led to the identification of a core set of 53 genes that are co-regulated by both CRC and AG [12]. Among the 53 genes co-regulated by AG and CRC, a subset of nine genes were identified as direct AG targets based on publicly available AG ChIP-seq datasets. These include: ARABIDOPSIS THALIANA HOMEOBOX PROTEIN 2 (ATHB2), MITOGEN-ACTIVATED PROTEIN KINASE 1(At1g10210), HALF FILLED (HAF), a wound-responsive family protein (At4g28240), SHATTERPROOF 2 (SHP2), a Sec14p-like phosphatidylinositol transfer family protein (At1g05370), GLABROUS INFLORESCENCE STEMS (GIS), TARGET OF EAT 3 (TOE3), and YUCCA4 (YUC4). Among these, YUC4 is an auxin-related target of CRC, acting downstream and synergistically with TRN2 to promote floral meristem termination through the indirect repression of WUS [12].
YUC4, in conjunction with TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS (TAA) family enzymes, catalyzes a two-step conversion oftryptophan to indole-3-acetic acid (IAA), representing a critical phase in the auxin biosynthesis pathway [17,99].
Putative synergistic and positive regulation of YUC4 by CRC and AG was supported by qRT-PCR analysis in crc-1 and ag-1-/+ crc-1 double mutants. Notably, YUC4 reporter (GUS) expression in the abaxial carpels at floral stage 6 was markedly reduced in the ag-1-/+ crc-1 double mutants compared to the crc-1 single mutant [12]. Induced expression of AG and CRC, either individually or in combination, supported a synergistic role in the activation of YUC4 transcription. This conclusion is further substantiated by the spatial overlap of CRC mRNA, CRC protein, and YUC4 reporter (GUS) activity observed in the abaxial carpels at floral stage 6 in wild-type plants [12].
The regulation of YUC4 by CRC involves its binding to YABBY-binding motifs (GA[A/G]AGAAA) located within conserved regulatory modules 1–4 (CRMs 1–4) of the YUC4 promoter. Chromatin immunoprecipitation (ChIP) assays using a CRC-myc fusion protein confirmed a direct interaction between CRC and the evolutionarily conserved YABBY-binding site within CRM4 [12].
The synergistic regulation of YUC4 by CRC and AG does not involve direct protein–protein interaction, as excluded by Y2H assays. Instead, the proposed mechanism suggests that AG facilitates the recruitment of its interacting chromatin remodeling factors, CHROMATIN REMODELING 11 (CHR11) and CHR17, to the YUC4 promoter. This recruitment promotes a nucleosome-free chromatin state, thereby enabling CRC to bind to DNA [12]
Increased expression of YUC4 augments local auxin concentration, resulting in cell wall loosening and cell proliferation, which may be supported by activity in the medial region of the developing gynoecium [13,18]. The dynamic distribution of the plant morphogen auxin is pivotal for the bilateral-to-radial symmetry transition, which is required for proper formation of the apical style in the Arabidopsis gynoecium [100].
AtCRC and AtAP1 may participate in the regulation of very long-chain fatty acid (VLCFA) biosynthesis. Co-expression analysis revealed that six out of eight genes co-expressed with AtCRC and AtAP1 encode enzymes involved in the VLCFA biosynthetic pathway. However, Y1H and electrophoretic mobility shift assays (EMSAs) demonstrated that only AtCRC directly interacts with the promoters of KCS7 and KCS15, indicating a direct regulatory role for AtCRC in VLCFA biosynthesis [101].
The KCS genes are involved in the response to salt and drought stress [102]. The biosynthesis of very long-chain fatty acids (VLCFAs) and auxin-related gene expression appear functionally linked, as suggested by studies on loss-of-function mutants of ONION1 (ONI1), which encodes a fatty acid elongase (β-ketoacyl CoA synthase) in rice [103]. In Arabidopsis, VLCFAs also regulate polar auxin transport and lateral root organogenesis [19].
2.4. Molecular Network and Gene Interactions Involving CRC
Mutations in the CRC gene in Arabidopsis thaliana lead to a distinct floral phenotype, characterized by a wider and shorter gynoecium, unfused carpels at the apex, reduced ovule number, and a complete absence of nectary development at all stages of floral maturation [4]. The gene was mapped between CLV2 and CLV1 on chromosome 1, subsequently positionally cloned, and effectively used to complement the crc mutation, thereby suppressing early radial expansion of the gynoecium and promoting its elongation at later developmental stages [4,43].
The inhibition of radial growth in the developing gynoecium by CRC begins in the lateral regions of the gynoecial primordium and is later reinforced within the epidermal layer. Although CRC also functions in the early stages of nectary development, its regulatory interactions differ between the gynoecium and nectary tissues. In the gynoecium, CRC expression is negatively regulated by A-class genes, including APETALA2 (AP2) and LEUNIG (LUG) [4]. The same holds true for the B-class genes in Arabidopsis, PISTILLATA (PI) and APETALA3 (AP3), which negatively regulate CRC expression in the third whorl [4]. Since mutations in the C-class gene AGAMOUS (AG) disrupt carpel development, the effect of AG loss-of-function was assessed in the first whorl. These studies showed that CRC expression persists in the absence of AG, but is significantly reduced and displays altered spatial distribution. Notably, CRC expression in nectaries remains unaffected by mutations in A-, B-, or C-class genes [4].
In Arabidopsis thaliana, the gynoecium is composed of two congenitally fused carpels, forming a single-chambered ovary (Figure 1) [104,105].
Abaxial–adaxial polarity within the gynoecium is evident from distinct epidermal cell morphologies on the abaxial (outer) and adaxial (inner) surfaces of the ovary valves, which contribute to proper organ development (Figure 1) [106].
A novel mutant phenotype in Arabidopsis carpels, characterized by the duplication of adaxial tissues—such as placentae and ovules—at abaxial positions, in addition to their normal localization on the adaxial (internal) side, was observed in double mutants where the function of CRC and either GYMNOS (GYM) or KANADI (KAN) is compromised [106]. The ectopic development of adaxial tissues is normally suppressed by CRC and KAN, which act through independent pathways and provide positional information to specify abaxial cell fate in the developing carpel. GYM functions as part of a chromatin-remodeling complex that represses transcription via the activity of the histone deacetylase Mi2 [106]. The targets of this complex are determined by specific transcription factors that recruit the GYM-containing complex. Consequently, GYM restricts primordia formation at the margins of postgenitally fused carpels by repressing genes that promote meristematic activity. The distinct genetic interactions between CRC and GYM suggest a lack of molecular redundancy. It is proposed that CRABS CLAW regulates transcription in a spatial manner, whereas GYMNOS acts temporally on downstream targets to ensure proper carpel differentiation [106].
The patterning events in the developing gynoecium depend on the activity of SEUSS and AINTEGUMENTA genes [107]. Phenotypic analyses of mutants for both genes suggest their synergistic interaction in the development of ovules and the medial domain of the gynoecium. Identified downstream targets of SEUSS and AINTEGUMENTA include PHABULOSA (PHB), REVOLUTA (REV), and CRABS CLAW (CRC) genes [106]. Analysis of the expression of two gynoecial adaxial fate regulators, PHB and REV, revealed that in seu ant double mutants, PHB expression was reduced within the adaxial core of the gynoecium at stages 5 and 6. This reduction was more pronounced compared to the ant single mutant [107]. Putatively, SEU and ANT regulate the expression of PHB during the transition from the floral meristem to the gynoecial primordia and continue to do so during later stages of gynoecial development. Moreover, in the seu ant double mutant, REV expression is still detected in the adaxial core of the gynoecium at stages 6 and 7, at levels comparable to the wild type [107]. However, at stages 8 and 9, expression within the adaxial gynoecium was reduced compared to the wild type. The seu ant double mutants exhibited defects in the expression of the abaxial fate regulator CRC, which became evident after stage 6. At stages 7 and 8, CRC expression within internal ovarian domains was either strongly reduced or absent, while expression in the abaxial epidermis remained clearly detectable [108]. Therefore, SEU and ANT promote the expression of CRC in the internal domains of the gynoecial ovary. However, in the apical region of the gynoecium, SEU and ANT act to repress CRC expression within the medial ridge, which correlates with an increased incidence of carpel separation spanning approximately 90% of the apical–basal axis of the gynoecium [107].
JAIBA (HAT1), a member of the class II HD-ZIP transcription factor subfamily, is implicated in both male and female reproductive development. In jab homozygous mutants, defects include a reduced number of mature pollen grains and ovules lacking a functional embryo sac [108,109]. Double jaiba crc-1 mutants exhibit severe defects in floral meristem determinacy and development of the gynoecium medial domain, indicating a functional relationship with CRC [108]. The name JAIBA was thus coined after the large blue swimming crab species (Callinectes sapidus), native to the Gulf of Mexico [108,109].
The jab crc-1 double mutant produced flowers with up to six sepals, up to six petals, and between five and seven stamens [108]. Additionally, the gynoecia of these flowers were composed of two to four carpels, which at later developmental stages transformed into carpeloid structures within the fruit. These observations indicate that JAB and CRC function cooperatively in the regulation of floral meristem development and its determinacy [109]. On the other hand, examination of the medial domain in jab crc-1 double mutant fruits revealed that although medial tissues are formed, they remain unfused (108].
Gynoecia lacking functional STYLISH1 (STY1) expression due to a transposon insertion exhibit aberrant style morphology. These defects are further enhanced in sty1 sty2 double mutants, which show a marked reduction in stylar and stigmatic tissues as well as decreased proliferation of stylar xylem [110]. CRC is proposed to interact with STY1, as sty1-1 crc-1 double mutants display a more severe reduction in stylar, stigmatic, septal, and medial xylem tissues compared to either single mutant [110]. The observed changes are too substantial to be accounted for by simple additive effects and are likely related to direct protein–protein interactions, as suggested by the overlapping expression of STY1 in the apical region of the gynoecium with the CRC expression domain in the abaxial carpel epidermis, which persists until approximately stage 10 of flower development [110]. STY1 functions as a transcriptional activator of the gene encoding the flavin-containing monooxygenase THREAD/YUCCA4, a key enzyme in the auxin biosynthesis pathway, ultimately resulting in altered auxin homeostasis [111].
2.5. Polymorphisms and Functional Mutations in the CRC Gene
T-DNA and ethyl methanesulfonate (EMS) mutagenesis screens performed in the crc-1 background led to the identification of novel genetic modifiers of the crc-1 phenotype. These were cloned and characterized as rebelote-1 (rbl-1, meaning "once again" in French), squint-4 (sqn-4), and ultrapetala1-4 (ult1-4). In all three resulting double mutants, the floral meristem (FM) remains indeterminate, with ectopic stamens and carpels arising from the central region of the FM, between the primary carpels, and along an elongated floral axis [33].
In the double mutants crc-1 rbl-1, crc-1 sqn-4, and crc-1 ult1-4, the expression of WUS and CLV3 persists in the center of FM at least until stage 6, indicating sustained FM activity and a failure of its determinacy [33]. RBL, SQN, and ULT1 function redundantly in the regulation of FM termination, likely through modulation of WUS activity [33].
Moreover, the rbl-1, sqn-4, and ult1-4 mutations enhance the ag-4 phenotype, suggesting that RBL, SQN, and ULT1 play partially redundant roles in supporting AG function [33]. Additionally, genetic crosses with the ag-6 mutant indicate that RBL, SQN, and, to a lesser extent, ULT1 contribute to floral meristem determinacy via an alternative pathway, potentially involving SUPERMAN (SUP), as the phenotypes of ag-6 rbl-1 and ag-6 sqn-4 resemble those of the ag-1 sup-1 double mutant [33,112].
Mutations in the tomato (Solanum lycopersicum) CRCa gene (SlCRCa) lead to the development of abnormal carpels that grow concentrically, one inside another, resulting in iterative fruit morphologies characteristic of the fig mutant phenotype [3]. In addition, affected fruits frequently develop secondary fruit structures that initiate internally and progressively expand to emerge on the fruit surface [3]. The molecular basis of the fig mutation is a 365-bp insertion of a putative transposable element within the intron between exons 4 and 5 of the SlCRCa gene [3]. The functional connection between the fig mutation in the SlCRCa gene and defects in carpel development was confirmed through the generation of SlCRCa knockdown (RNAi) and knockout (CRISPR/Cas9) lines, which exhibited phenotypes resembling those of fig mutants [3]. However, microsynteny analyses of genomic regions surrounding CRC genes in Arabidopsis and tomato indicate a greater similarity to the second tomato allele, SlCRCb, than to the initially identified SlCRCa. Although knockout mutations of both SlCRCa and SlCRCb result in similar phenotypic alterations, and their spatial expression patterns largely overlap, the temporal expression profile of SlCRCb differs: SlCRCb transcripts are detected throughout all stages of floral development—from early floral buds at stages 0–6 to flowers 10 days post-anthesis [3].
The molecular function of SlCRCa and SlCRCb involves repression of the WUSCHEL (WUS) locus, a key regulator of floral meristem determinacy, consistent with prior findings in both Arabidopsis and tomato [25]
Although SlWUS expression is terminated from stage 6 onward in wild-type plants, SlCRCa and SlCRCb mutants maintain SlWUS expression beyond stage 6, resulting in defects in floral determinacy [3]. Similar to Arabidopsis thaliana, the SlWUS locus in tomato is associated with a chromatin remodeling complex exhibiting histone deacetylase activity, as demonstrated by bimolecular fluorescence complementation and co-immunoprecipitation assays. More specifically, SlCRCa and SlCRCb proteins can interact with one another, and each can also interact with SlKNU, SlIMA, and SlHDA1. Notably, only SlCRCb, and not SlCRCa, interacts with SlTPL1 [3]
Analysis of quantitative trait loci (QTL), in combination with map-based cloning, led to the identification of a nonsynonymous polymorphism (G to A) in the CRABS CLAW gene of cucumber (Cucumis sativus, CsCRC) as the causal factor underlying variations in fruit size and shape. RT-PCR analyses revealed that CsCRC expression is predominantly confined to reproductive organs, including flowers and fruits, with no detectable expression in vegetative tissues such as stems and leaves. Among reproductive organs, the highest CsCRC expression was observed in the nectary, followed by female flower buds and young ovaries [31].
The presence of a rare CsCRC^A allele is associated with reduced fruit length in Cucumis sativus, whereas the CsCRC^G allele shows a positive correlation with increased fruit size. Knockdown of CsCRC^G leads to shorter fruits with reduced cell size, while introgression of the CsCRC^G allele into a near-isogenic line homozygous for CsCRC^A restored the elongated fruit phenotype [31]. The causal variant is a nonsynonymous SNP (G-to-A) in CsGy5G023910, a homolog of ArabidopsisCRABS CLAW (CRC), which leads to an amino acid substitution from arginine (R) to glutamine (Q) within the conserved YABBY DNA-binding domain [31]. RNA-seq analyses of CsCRC and auxin-responsive protein 1 (CsARP1) in different cucumber lines revealed that CsARP1 transcript levels are significantly reduced in CsCRC-RNAi plants and elevated in CsCRC^G-overexpressing lines [31]. The arginine-to-glutamine (R→Q) substitution within the YABBY DNA-binding domain of CsCRC markedly diminishes its binding affinity to [A/T]ATCAT[A/T] and [T/A]ATGAT[T/A] cis-regulatory motifs in the CsARP1 promoter, thereby reducing CsARP1 expression [31].
The functional significance of the DNA–protein interaction in regulating CsARP1 transcription was validated through Y1H assays, chromatin immunoprecipitation, electrophoretic mobility shift assays (EMSA), and transient expression assays in Nicotiana benthamiana leaves [31]. Putatively, the upregulated CsARP1 transcription enhances cell expansion in the pulp cells of cucumber fruit mesocarp, potentially via transmembrane electron transport and cell wall modification, as implied by the presence of a conserved cytochrome b561 domain and an additional DOMON domain in the encoded protein [31].
These findings support a critical link between CRC function and auxin-regulated pathways, consistent with earlier studies [12,13,96]. In tomato, changes in fruit size are inversely correlated with SlCRCa expression levels [20]. Light microscopy analysis demonstrated that the reduction in fruit size caused by SlCRCa overexpression is associated with a decreased number of cell layers and reduced cell size in the fruit pericarp [20].
These phenotypic changes were reversed in tomato plants with reduced SlCRCa expression via RNA interference (RNAi). Furthermore, transcriptomic analysis of SlCRCa-overexpressing fruits identified 356 upregulated and 1,222 downregulated genes. Among these were numerous genes involved in cell division, including cyclins, cyclin-dependent kinases (CDKs), and expansins, as well as genes implicated in cell expansion and stress responses, such as two RADICAL-INDUCED CELL DEATH1 (RCD1) genes [20]. The regulation of fruit size changes induced by SlCRCa may also involve YABBY5a and genes of the gibberellin biosynthesis pathway, both of which are generally downregulated in response to SlCRCa overexpression [20].
2.6. Phylogenetic Insights into CRC Gene Lineage
The origin of CRC coincided with the emergence of angiosperms, as CRC plays a pivotal role in the development of the carpel—an organ unique to flowering plants [38,113]. Although closely related CRC paralogs are present in gymnosperm genomes, it remains unclear how CRC was integrated into gene regulatory networks to control carpel-specific functions during the evolution of angiosperms [38,114].
Phylogenetic analyses indicate that CRC genes involved in floral meristem termination, carpel organ identity, and abaxial–adaxial polarity in Arabidopsis thaliana have remained numerically stable and have not undergone significant expansion across plant lineages [38,114].
The CRC gene is maintained as a single-copy gene in most angiosperms, which is atypical for evolutionarily recent genes [38]. It is hypothesized that the evolution of a relatively small and tightly regulated carpel gene regulatory network (GRN)—comprising genes such as ALCATRAZ (ALC), CRABS CLAW (CRC), HALF FILLED (HAF), HECATE (HEC), INDEHISCENT (IND), NGATHA (NGA), and SPATULA (SPT)—does not tolerate dosage imbalances or the expansion of gene families encoding proteins with redundant functions [38].
The termination of reproductive meristems appears particularly sensitive to dosage imbalance, as demonstrated by the effects of increased CRC expression. Artificial elevation of CRC levels can lead to premature termination of reproductive meristems—an outcome that is strongly selected against in natural populations [1,33].
Although CRC is typically maintained as a single-copy gene in most plant species, an exception is observed in the Solanaceae family, where it occurs as paralogous pairs—CRCa and CRCb in Solanum lycopersicum, and CRC1 and CRC2 in Petunia hybrida [115]. These paralogs are believed to have arisen from a large segmental duplication event that likely occurred in a common ancestor of the Solanaceae [115].
The CRABS CLAW (CRC) ortholog in the monocot Oryza sativa, known as DROOPING LEAF (DL), exhibits differences in both spatial expression and function compared to CRC [7,8,9]. In Arabidopsis thaliana, CRC is expressed in the abaxial region of carpel primordia and in floral nectaries, where it plays a key role in regulating carpel morphology and nectary development [1,3,5,6,7,35]. In Oryza sativa, DROOPING LEAF (DL) is expressed throughout the entire carpel primordium and in the central undifferentiated cells of developing leaves, where it regulates both carpel identity and midrib development [7].
Analysis of CRC orthologs in two Fabaceae species, Pisum sativum and Medicago truncatula, revealed not only an atypical absence of abaxial expression in the carpel, but also an unusual expression pattern associated with the medial vein of the ovary. This suggests a potential role in vascular development—previously thought to be specific to DL in monocotyledons [116].
The temporal and spatial expression patterns of DL orthologs in three grass species—maize, wheat, and sorghum—closely mirror those of Oryza sativa DL, during both floral and leaf development. These findings suggest a high degree of functional conservation of DL-related genes within the Poaceae (grass) family [117].
Analysis of the spatial expression patternsofCRC/DL orthologs in eudicots and basal eudicots—such as Petunia hybrida, Gossypium hirsutum, Eschscholzia californica (basal eudicot), and Aquilegia formosa (basal eudicot)—reveals that the abaxial specificity and absence of expression in leaves, characteristic of Arabidopsis thaliana CRC, are highly conserved among these species. This conservation suggests a stable and ancestral regulatory role for CRC/DL orthologs in gynoecial development across eudicot lineages [1,34].
The ancestral role of CRC in carpel development was confirmed in the basal angiosperm Amborella trichopoda, whose CRC orthologue partially complemented the crc-1 mutant phenotype in carpels but not in nectaries [118]. This partial complementation suggests that novel functions of CRC orthologues, such as nectary development, may be mediated by evolutionary changes in non-coding regulatory sequences [119].
CRC also regulates the formation of nectaries, which are present not only in the floral nectaries of Arabidopsis but also in the extrafloral nectaries of Gossypium hirsutum and Capparis flexuosa [4,120]. However, in basal eudicots such as Aquilegia formosa, the CRC orthologue is not expressed in the nectary, suggesting that CRC orthologues were recruited for nectary development at the base of the core eudicots lineage [119,120,121].
Studies of CRC/DL expression across major angiosperm clades demonstrate the stepwise acquisition of novel functions without gene duplication, exemplifying the role of regulatory elements—such as promoters and enhancers—in broadening gene function [1,42,119,120]. Alternatively, the novel function of CRC in nectary development may have arisen through one or more changes affecting the composition of downstream target genes, the availability or identity of protein interaction partners, and/or the spatiotemporal expression of CRC in nectary tissues [119].
Analysis of the functional diversification of DROOPING LEAF (DL) in Asparagus asparagoides revealed that the evolution of CRC/DL genes from their ancestral role in specifying abaxial cell fate during carpel development to acquiring expression in leaves occurred prior to the divergence of the order Asparagales. However, traits such as expression throughout the entire carpel primordium, in the central region of leaves, and involvement in carpel organ identity were gained after the divergence of Asparagales [42].
In the orchid Phalaenopsis equestris, two DL/CRC paralogs, PeDL1 and PeDL2, were identified as products of a whole-genome duplication (WGD) event in the last common ancestor of orchids. Both genes are expressed in the floral meristem and carpel tissue, supporting the ancestral role of DL/CRC genes in floral meristem determinacy and carpel specification [41].
Moreover, expression of both PeDL genes was observed in the placenta and ovule primordia during early stages of ovary development in Phalaenopsis equestris [121]. Similarly, in California poppy (Eschscholzia californica), the CRC ortholog EcCRC has also been co-opted for additional roles in ovule initiation [1].
3. Discussion
Studies on the function of CRC in meristem termination and gynoecium or nectary development were initially conducted in the model plant Arabidopsis thaliana [4,28,34,35,44,60,108,120]. The phenotypic consequences of CRC gene mutations, together with the availability of the A. thaliana genome sequence and transcriptomic datasets, provided a foundation for the identification of novel genes that are either co-expressed with CRC or modulate its role in meristem termination and gynoecium development [23,33,35,39,101,106].
Furthermore, Y2H and bimolecular fluorescence complementation (BiFC) assays have yielded critical insights into proteins that interact with CRC or AG, and directly contribute to the termination of WUS expression [14,44,54].
Beyond Arabidopsis thaliana, CRC paralogs have been relatively well studied in tomato, providing insights into species- or developmental stage-specific variations in the composition of protein complexes involved in WUS repression [3,25,26]. However, studies investigating CRC function in other plant species remain relatively limited [1,24,30,32,42,123,124].
Consequently, future research should focus on the isolation of CRC orthologs and paralogs in a broader range of plant species. Generating transgenic lines with CRC overexpression, gene silencing, or targeted mutations would provide a basis for analyzing phenotypic and physiological alterations associated with CRC function.
These effects could be compared with observations from Arabidopsis thaliana, rice and tomato to elucidate both conserved and novel roles of the CRC gene. In newly studied plant species, CRC gene mutations could be combined with chemical mutagenesis or transcriptomic analyses to identify previously uncharacterized genes that are co-expressed with CRC and modulate phenotypic or physiological traits associated with meristem termination and gynoecium development. In this way the already known and novel functionalites of CRC could be revealed.
The application of Y1H assays or chromatin immunoprecipitation (ChIP) techniques could facilitate the identification of trans-acting factors regulating CRC gene expression in novel, often non-model, plant species. Similarly, Y2H, Y3H, and BiFC assays could be employed to identify components of protein complexes centered around CRC that are directly involved in executing its function. The functional relevance of these interactions could be validated by analyzing phenotypic alterations in plants harboring mutations in CRC-associated proteins.
Y2H screening often yields a substantial number of putative interacting proteins, frequently exceeding one hundred, many of which are false positives or are partially annotated in current databases [126,127,128,129,130,131]. Moreover, future progress in identifying cis-elements or trans-acting factors regulating CRC-dependent gene expression will rely upon advancements in the functionality and comprehensiveness of plant genomic and proteomic databases and improvements of Y1H methodology [125,127,128,129,132].
Author Contributions
Conceptualization, writing—original draft preparation, P.S.; English correction, writing—review and editing J.N.; Visualization, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by statutory funds of the Department of Biology and Pharmaceutical Botany at the Medical University of Łódź (503/3-012-01/503-31-001-19-00).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Orashakova, S.; Lange, M.; Lange, S.; Wege, S.; Becker, A. The CRABS CLAW ortholog from California poppy (Eschscholzia californica, Papaveraceae), EcCRC, is involved in floral meristem termination, gynoecium differentiation and ovule initiation. Plant J. 2009, 58, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Thong, Z.; Yu, H. Coming into bloom: the specification of floral meristems. Development. 2009, 136, 3379–3391. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, L.; Giménez, E.; Pineda, B.; García-Sogo, B.; Ortiz-Atienza, A.; Micol-Ponce, R.; Angosto, T.; Capel, J.; Moreno, V.; Yuste-Lisbona, F.J.; Lozano, R. Tomato CRABS CLAW paralogues interact with chromatin remodelling factors to mediate carpel development and floral determinacy. New Phytol. 2022, 234, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Smyth, D.R. CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development. 1999, 126, 2387–2396. [Google Scholar] [CrossRef]
- Golz, J.F.; Hudson, A. Plant development: YABBYs claw to the fore. Curr Biol. 1999, 9, R861–R863. [Google Scholar] [CrossRef]
- Han, K.; Lai, M.; Zhao, T.; Yang, X.; An, X.; Chen, Z. Plant YABBY transcription factors: a review of gene expression, biological functions, and prospects. Crit Rev Biotechnol. 2025, 45, 214–235. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell. 2004, 16, 500–509. [Google Scholar] [CrossRef]
- Ohmori, Y.; Toriba, T.; Nakamura, H.; Ichikawa, H.; Hirano, H.Y. Temporal and spatial regulation of DROOPING LEAF gene expression that promotes midrib formation in rice. Plant J. 2011, 65, 77–86. [Google Scholar] [CrossRef]
- Abiko, M.; Ohmori, Y.; Hirano, H.Y. Genome-wide expression profiling and identification of genes under the control of the DROOPING LEAF gene during midrib development in rice. Genes Genet Syst. 2008, 83, 237–244. [Google Scholar] [CrossRef]
- Shang, E.; Wang, X.; Li, T.; Guo, F.; Ito, T.; Sun, B. Robust control of floral meristem determinacy by position-specific multifunctions of KNUCKLES. Proc. Natl. Acad. Sci. U S A. 2021, 118, e2102826118. [Google Scholar] [CrossRef]
- Sun, B.; Zhou, Y.; Cai, J.; Shang, E.; Yamaguchi, N.; Xiao, J.; Looi, L.S.; Wee, W.Y.; Gao, X.; Wagner, D.; Ito, T. Integration of Transcriptional Repression and Polycomb-Mediated Silencing of WUSCHEL in Floral Meristems. Plant Cell. 2019, 31, 1488–1505. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Huang, J.; Tatsumi, Y.; Abe, M.; Sugano, S.S.; Kojima, M.; Takebayashi, Y.; Kiba, T.; Yokoyama, R.; Nishitani, K.; Sakakibara, H.; Ito, T. Chromatin-mediated feed-forward auxin biosynthesis in floral meristem determinacy. Nat Commun. 2018, 9, 5290. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Huang, J.; Xu, Y.; Tanoi, K.; Ito, T. Fine-tuning of auxin homeostasis governs the transition from floral stem cell maintenance to gynoecium formation. Nat Commun. 2017, 8, 1125. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Ubaldo, H.; Campos, S.E.; López-Gómez, P.; Luna-García, V.; Zúñiga-Mayo, V.M.; Armas-Caballero, G.E.; González-Aguilera, K.L.; DeLuna, A.; Marsch-Martínez, N.; Espinosa-Soto, C.; de Folter, S. The protein-protein interaction landscape of transcription factors during gynoecium development in Arabidopsis. Mol Plant. 2023, 16, 260–278. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.J.; Larsson, E.; Spíchal, L.; Sundberg, E. Cytokinin-Auxin Crosstalk in the Gynoecial Primordium Ensures Correct Domain Patterning. Plant Physiol. 2017, 175, 1144–1157. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, R.; Zi, H.; Li, Y.; Cao, X.; Li, D.; Guo, L.; Tong, J.; Pan, Y.; Jiao, Y.; Liu, R.; Xiao, L.; Liu, X. AUXIN RESPONSE FACTOR3 Regulates Floral Meristem Determinacy by Repressing Cytokinin Biosynthesis and Signaling. Plant Cell. 2018, 30, 324–346. [Google Scholar] [CrossRef]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef]
- Majda, M.; Robert, S. The Role of Auxin in Cell Wall Expansion. Int J Mol Sci. 2018, 19, 951. [Google Scholar] [CrossRef]
- Roudier, F.; Gissot, L.; Beaudoin, F.; Haslam, R.; Michaelson, L.; Marion, J.; Molino, D.; Lima, A.; Bach, L.; Morin, H.; Tellier, F.; Palauqui, J.C.; Bellec, Y.; Renne, C.; Miquel, M.; Dacosta, M.; Vignard, J.; Rochat, C.; Markham, J.E.; Moreau, P. ; Napier, J; Faure, J. D. Very-long-chain fatty acids are involved in polar auxin transport and developmental patterning in Arabidopsis. Plant Cell. 2010, 22, 364–375. [Google Scholar] [CrossRef]
- Yang, T.; He, Y.; Niu, S.; Zhang, Y. A YABBY gene CRABS CLAW a (CRCa) negatively regulates flower and fruit sizes in tomato. Plant Sci. 2022, 320, 111285. [Google Scholar] [CrossRef]
- Xu, G.; Huang, J.; Lei, S.K.; Sun, X.G.; Li, X. Comparative gene expression profile analysis of ovules provides insights into Jatropha curcas L. ovule development. Sci Rep. 2019, 9, 15973. [Google Scholar] [CrossRef]
- Fu, Q.; Niu, L.; Chen, M.S.; Tao, Y.B.; Wang, X.; He, H.; Pan, B.Z.; Xu, Z.F. De novo transcriptome assembly and comparative analysis between male and benzyladenine-induced female inflorescence buds of Plukenetia volubilis. J Plant Physiol. 2018, 221, 107–118. [Google Scholar] [CrossRef]
- Deng, M.H.; Zhao, K.; Lv, J.H.; Huo, J.L.; Zhang, Z.Q.; Zhu, H.S.; Zou, X.X.; Wen, J.F. Flower transcriptome dynamics during nectary development in pepper (Capsicum annuum L.). Genet Mol Biol. 2020, 43, e20180267. [Google Scholar] [CrossRef] [PubMed]
- Katzer, A.M.; Wessinger, C.A.; Anaya, B.M.; Hileman, L.C. CRABS CLAW-independent floral nectary development in Penstemon barbatus. Am J Bot. 2025, 112, e70058. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, P.; Li, M.; Zhu, D.; Ma, H.; Xu, H.; Li, S.; Wei, J.; Bian, X.; Wang, M.; Lai, Y.; Peng, Y.; Li, H.; Rahman, A.; Wu, S. Heat stress impairs floral meristem termination and fruit development by affecting the BR-SlCRCa cascade in tomato. Plant Commun. 2024, 5, 100790. [Google Scholar] [CrossRef]
- Bollier, N.; Sicard, A.; Leblond, J.; Latrasse, D.; Gonzalez, N.; Gévaudant, F.; Benhamed, M.; Raynaud, C.; Lenhard, M.; Chevalier, C.; Hernould, M.; Delmas, F. At-MINI ZINC FINGER2 and Sl-INHIBITOR OF MERISTEM ACTIVITY, a Conserved Missing Link in the Regulation of Floral Meristem Termination in Arabidopsis and Tomato. Plant Cell. 2018, 30, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Chen, B.; Qiao, L.; Chen, F.; Zhao, J.; Cheng, Z.; Weng, Y. Phenotypic Characterization and Fine Mapping of a Major-Effect Fruit Shape QTL FS5. 2 in Cucumber, Cucumis sativus L., with Near-Isogenic Line-Derived Segregating Populations. Int J Mol Sci. 2022, 23, 13384. [Google Scholar] [CrossRef]
- Morel, P.; Heijmans, K.; Ament, K.; Chopy, M.; Trehin, C.; Chambrier, P.; Rodrigues Bento, S.; Bimbo, A.; Vandenbussche, M. The Floral C-Lineage Genes Trigger Nectary Development in Petunia and Arabidopsis. Plant Cell. 2018, 30, 2020–2037. [Google Scholar] [CrossRef]
- Peréz-Mesa, P.; Ortíz-Ramírez, C.I.; González, F.; Ferrándiz, C.; Pabón-Mora, N. Expression of gynoecium patterning transcription factors in Aristolochia fimbriata (Aristolochiaceae) and their contribution to gynostemium development. Evodevo. 2020, 11, 4. [Google Scholar] [CrossRef]
- Duan, X.; Xie, W.; Chen, X.; Zhang, H.; Zhao, T.; Huang, J.; Zhang, R.; Li, X. Morphological and molecular mechanisms of floral nectary development in Chinese Jujube. BMC Plant Biol. 2024, 24, 1041. [Google Scholar] [CrossRef]
- Che, G.; Pan, Y.; Liu, X.; Li, M.; Zhao, J.; Yan, S.; He, Y.; Wang, Z.; Cheng, Z.; Song, W.; Zhou, Z.; Wu, T.; Weng, Y.; Zhang, X. Natural variation in CRABS CLAW contributes to fruit length divergence in cucumber. Plant Cell. 2023, 35, 738–755. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Li, C.; Duan, Y.; Qu, W.; Wang, H.; Miao, H.; Zhang, H. A SNP Mutation of SiCRC Regulates Seed Number Per Capsule and Capsule Length of cs1 Mutant in Sesame. Int J Mol Sci. 2019, 20, 4056. [Google Scholar] [CrossRef] [PubMed]
- Prunet, N.; Morel, P.; Thierry, A.M.; Eshed, Y.; Bowman, J.L.; Negrutiu, I.; Trehin, C. REBELOTE, SQUINT, and ULTRAPETALA1 function redundantly in the temporal regulation of floral meristem termination in Arabidopsis thaliana. Plant Cell. 2008, 20, 901–919. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Baum, S.F.; Alvarez, J.; Patel, A.; Chitwood, D.H.; Bowman, J.L. Activation of CRABS CLAW in the Nectaries and Carpels of Arabidopsis. Plant Cell. 2005, 17, 25–36. [Google Scholar] [CrossRef]
- Gross, T.; Becker, A. Transcription Factor Action Orchestrates the Complex Expression Pattern of CRABS CLAW in Arabidopsis. Genes (Basel). 2021, 12, 1663. [Google Scholar] [CrossRef]
- Kidokoro, S.; Konoura, I.; Soma, F.; Suzuki, T.; Miyakawa, T.; Tanokura, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Clock-regulated coactivators selectively control gene expression in response to different temperature stress conditions in Arabidopsis. Proc Natl Acad Sci U S A. 2023, 120, e2216183120. [Google Scholar] [CrossRef]
- Ding, Z.J.; Yan, J.Y.; Li, G.X.; Wu, Z.C.; Zhang, S.Q.; Zheng, S.J. WRKY41 controls Arabidopsis seed dormancy via direct regulation of ABI3 transcript levels not downstream of ABA. Plant J. 2014, 79, 810–823. [Google Scholar] [CrossRef]
- Pfannebecker, K.C.; Lange, M.; Rupp, O.; Becker, A. Seed Plant-Specific Gene Lineages Involved in Carpel Development. Mol Biol Evol. 2017, 34, 925–942. [Google Scholar] [CrossRef]
- Zúñiga-Mayo, V.M.; Gómez-Felipe, A.; Herrera-Ubaldo, H.; de Folter, S. Gynoecium development: networks in Arabidopsis and beyond. J Exp Bot. 2019, 70, 1447–1460. [Google Scholar] [CrossRef]
- Scutt, C.P.; Vinauger-Douard, M.; Fourquin, C.; Finet, C.; Dumas, C. An evolutionary perspective on the regulation of carpel development. J Exp Bot. 2006; 57, 2143–2152. [CrossRef]
- Yamada, T.; Yokota, S.; Hirayama, Y.; Imaichi, R.; Kato, M.; Gasser, C.S. Ancestral expression patterns and evolutionary diversification of YABBY genes in angiosperms. Plant J. 2011, 67, 26–36. [Google Scholar] [CrossRef]
- Nakayama, H.; Yamaguchi, T.; Tsukaya, H. Expression patterns of AaDL, a CRABS CLAW ortholog in Asparagus asparagoides (Asparagaceae), demonstrate a stepwise evolution of CRC/DL subfamily of YABBY genes. Am J Bot. 2010, 97, 591–600. [Google Scholar] [CrossRef]
- Alvarez, J.; Smyth, D.R. CRABS CLAW and SPATULA, two Arabidopsis genes that control carpel development in parallel with AGAMOUS. Development. 1999, 126, 2377–2386. [Google Scholar] [CrossRef] [PubMed]
- Gross, T.; Broholm, S.; Becker, A. CRABS CLAW Acts as a Bifunctional Transcription Factor in Flower Development. Front Plant Sci. 2018, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- Bobb, A.J.; Eiben, H.G.; Bustos, M.M. PvAlf, an embryo-specific acidic transcriptional activator enhances gene expression from phaseolin and phytohemagglutinin promoters. Plant J. 1995, 8, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; Tjian, R. Transcriptional regulation in mammalian cells by sequence-specific DNA binding proteins. Science. 1989, 245, 371–378. [Google Scholar] [CrossRef]
- Romanova, M.A.; Maksimova, A.I.; Pawlowski, K.; Voitsekhovskaja, O.V. YABBY Genes in the Development and Evolution of Land Plants. Int J Mol Sci. 2021, 22, 4139. [Google Scholar] [CrossRef]
- Zhang, T.; Li, C.; Li, D.; Liu, Y.; Yang, X. Roles of YABBY transcription factors in the modulation of morphogenesis, development, and phytohormone and stress responses in plants. J Plant Res. 2020, 133, 751–763. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six classes of nuclear localization signals specific to different binding grooves of importin alpha. J Biol Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef]
- Cross, E.M.; Akbari, N.; Ghassabian, H.; Hoad, M.; Pavan, S.; Ariawan, D.; Donnelly, C.M.; Lavezzo, E.; Petersen, G.F.; Forwood, J.K.; Alvisi, G. A functional and structural comparative analysis of large tumor antigens reveals evolution of different importin α-dependent nuclear localization signals. Protein Sci. 2024, 33, e4876. [Google Scholar] [CrossRef]
- Chang, C.W.; Couñago, R.M.; Williams, S.J.; Bodén, M.; Kobe, B. Distinctive conformation of minor site-specific nuclear localization signals bound to importin-α. Traffic. 2013, 14, 1144–1154. [Google Scholar] [CrossRef]
- Strader, L.; Weijers, D.; Wagner, D. Plant transcription factors—Being in the right place with the right company. Curr. Op. Plant Biol. 2022, 65, 102136. [Google Scholar] [CrossRef]
- Amoutzias, G.D.; Robertson, D.L.; Van de Peer, Y.; Oliver, S.G. Choose your partners: Dimerization in eukaryotic transcription factors. Trends Biochem. Sci. 2008, 33, 220–229. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Yan, M.; Liu, C.; Yuan, Z. BELL1 interacts with CRABS CLAW and INNER NO OUTER to regulate ovule and seed development in pomegranate. Plant Physiol. 2023, 191, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- de Boer, C.G.; Vaishnav, E.D.; Sadeh, R.; Abeyta, E.L.; Friedman, N.; Regev, A. Deciphering eukaryotic gene-regulatory logic with 100 million random promoters. Nat. Biotechnol. 2020, 38, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Louphrasitthiphol, P.; Siddaway, R.; Loffreda, A.; Pogenberg, V.; Friedrichsen, H.; Schepsky, A.; Zeng, Z.; Lu, M.; Strub, T.; Freter, R.; et al. Tuning transcription factor availability through acetylation-mediated genomic redistribution. Mol. Cell 2020, 79, 472–487.e10. [Google Scholar] [CrossRef]
- Grau, J.; Schmidt, F.; Schulz, M.H. Widespread effects of DNA methylation and intra-motif dependencies revealed by novel transcription factor binding models. Nucleic Acids Res. 2023, 51, e95. [Google Scholar] [CrossRef]
- Shahein, A.; López-Malo, M.; Istomin, I.; Olson, E.J.; Cheng, S.; Maerkl, S.J. Systematic analysis of low-affinity transcription factor binding site clusters in vitro and in vivo establishes their functional relevance. Nat. Commun. 2022, 13, 5273. [Google Scholar] [CrossRef]
- Fontana, M.; Roosjen, M.; Crespo García, I.; van den Berg, W.; Malfois, M.; Boer, R.; Weijers, D.; Hohlbein, J. Cooperative action of separate interaction domains promotes high-affinity DNA binding of Arabidopsis thaliana ARF transcription factors. Proc. Natl. Acad. Sci. USA 2023, 120, e2219916120. [Google Scholar] [CrossRef]
- Shang, E.; Ito, T.; Sun, B. Control of floral stem cell activity in Arabidopsis. Plant Signal Behav. 2019;14(11):1659706. [CrossRef]
- Yadav, S.; Kumar, H.; Mahajan, M.; Sahu, S.K.; Singh, S.K.; Yadav, R.K. Local auxin biosynthesis promotes shoot patterning and stem cell differentiation in Arabidopsis shoot apex. Development. 2023, 150, dev202014. [Google Scholar] [CrossRef]
- Kwaśniewska, K.; Breathnach, C.; Fitzsimons, C.; Goslin, K.; Thomson, B.; Beegan, J.; Finocchio, A.; Prunet, N.; Ó'Maoiléidigh, D.S.; Wellmer, F. Expression of KNUCKLES in the Stem Cell Domain Is Required for Its Function in the Control of Floral Meristem Activity in Arabidopsis. Front Plant Sci. 2021, 12, 704351. [Google Scholar] [CrossRef]
- Guo, L.; Cao, X.; Liu, Y.; Li, J.; Li, Y.; Li, D.; Zhang, K.; Gao, C.; Dong, A.; Liu, X. A chromatin loop represses WUSCHEL expression in Arabidopsis. Plant J. 2018, 94, 1083–1097. [Google Scholar] [CrossRef]
- ÓMaoiléidigh, D.S.; Wuest, S.E.; Rae, L.; Raganelli, A.; Ryan, P.T.; Kwasniewska, K.; Das, P.; Lohan, A.J.; Loftus, B.; Graciet, E.; Wellmer, F. Control of reproductive floral organ identity specification in Arabidopsis by the C function regulator AGAMOUS. Plant Cell. 2013, 25, 2482–2503. [Google Scholar] [CrossRef]
- Liu, Z.; Shpak, E.D.; Hong, T. A mathematical model for understanding synergistic regulations and paradoxical feedbacks in the shoot apical meristem. Comput Struct Biotechnol J. 2020, 18, 3877–3889. [Google Scholar] [CrossRef]
- Han, H.; Liu, X.; Zhou, Y. Transcriptional circuits in control of shoot stem cell homeostasis. Curr Opin Plant Biol. 2020, 53, 50–56. [Google Scholar] [CrossRef]
- Daum, G.; Medzihradszky, A.; Suzaki, T.; Lohmann, J.U. A mechanistic framework for noncell autonomous stem cell induction in Arabidopsis. Proc Natl Acad Sci U S A. 2014, 111, 14619–14624. [Google Scholar] [CrossRef]
- Zhang, L.; DeGennaro, D.; Lin, G.; Chai, J.; Shpak, E.D. ERECTA family signaling constrains CLAVATA3 and WUSCHEL to the center of the shoot apical meristem. Development. 2021, 148, dev189753. [Google Scholar] [CrossRef]
- Rodriguez, K.; Kao, L.; Cerbantez-Bueno, V.E.; Delgadillo, C.; Nguyen, D.; Ullah, S.; Delgadillo, C.; Reddy, G.V. HAIRY MERISTEM proteins regulate the WUSCHEL protein levels in mediating CLAVATA3 expression. Physiol Plant. 2024, 176, e14505. [Google Scholar] [CrossRef]
- Su, Y.H.; Zhou, C.; Li, Y.J.; Yu, Y. Tang, L. P, Zhang WJ, Yao WJ, Huang R, Laux T, Zhang XS. Integration of pluripotency pathways regulates stem cell maintenance in the Arabidopsis shoot meristem. Proc Natl Acad Sci U S A. 2020, 117, 22561–22571. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zhu, Y.; Cui, Y.; Cheng, K.; Liang, W.; Wei, Z.; Zhu, M.; Yin, H.; Zeng, L.; Xiao, Y.; Lv, M.; Yi, J.; Hou, S.; He, K.; Li, J.; Gou, X. A group of receptor kinases are essential for CLAVATA signalling to maintain stem cell homeostasis. Nat Plants. 2018, 4, 205–211. [Google Scholar] [CrossRef] [PubMed]
- 72 Wang, Y.; Jiao, Y. Cell signaling in the shoot apical meristem. Plant Physiol. 2023, 193, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.E.; Running, M. P, Meyerowitz EM. CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development. 1993, 119, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.E.; Running, M.P.; Meyerowitz, E.M. CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development. 1995, 121: 2057–2067.
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.; Jürgens, G.; Laux, T. The stem cell population of Arabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell. 2000, 100, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Brand, U.; Fletcher, J.C.; Hobe, M.; Meyerowitz, E.M.; Simon, R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science. 2000, 289, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, J.; Denay, G.; Wink, R.; Pinto, K.G.; Stahl, Y.; Schmid, J.; Blümke, P.; Simon, R.G. Control of Arabidopsis shoot stem cell homeostasis by two antagonistic CLE peptide signalling pathways. Elife. 2021, 10, 70934. [Google Scholar] [CrossRef]
- Lenhard, M.; Jürgens, G.; Laux, T. The WUSCHEL and SHOOTMERISTEMLESS genes fulfil complementary roles in Arabidopsis shoot meristem regulation. Development. 2002, 129, 3195–3206. [Google Scholar] [CrossRef]
- Causier, B.; Ashworth, M.; Guo, W.; Davies, B. The TOPLESS interactome: a framework for gene repression in Arabidopsis. Plant Physiol. 2012, 158, 423–438. [Google Scholar] [CrossRef]
- Kagale, S.; Rozwadowski, K. EAR motif-mediated transcriptional repression in plants: an underlying mechanism for epigenetic regulation of gene expression. Epigenetics. 2011, 6, 141–146. [Google Scholar] [CrossRef]
- Lenhard, M.; Bohnert, A.; Jürgens, G.; Laux, T. Termination of stem cell maintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS. Cell. 2001, 105, 805–814. [Google Scholar] [CrossRef]
- Sieburth, L.E.; Running, M.P.; Meyerowitz, E.M. Genetic separation of third and fourth whorl functions of AGAMOUS. Plant Cell. 1995, 7, 1249–1258. [Google Scholar] [CrossRef]
- Lohmann, J.U.; Hong, R.L.; Hobe, M.; Busch, M.A.; Parcy, F.; Simon, R.; Weigel, D. A molecular link between stem cell regulation and floral patterning in Arabidopsis. Cell. 2001, 105, 793–803. [Google Scholar] [CrossRef]
- Liu, X.; Kim, Y.J.; Müller, R.; Yumul, R.E.; Liu, C.; Pan, Y.; Cao, X.; Goodrich, J.; Chen, X. AGAMOUS terminates floral stem cell maintenance in Arabidopsis by directly repressing WUSCHEL through recruitment of Polycomb Group proteins. Plant Cell. 2011, 23, 3654–3670. [Google Scholar] [CrossRef]
- Sun, B.; Xu, Y.; Ng, K.H.; Ito, T. A timing mechanism for stem cell maintenance and differentiation in the Arabidopsis floral meristem. Genes Dev. 2009, 23, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Liu, X.; Zhou, C.; Zhou, Q.; Zhao, Y.; Li, G.; Zhou, D.X. Cooperation between the H3K27me3 Chromatin Mark and Non-CG Methylation in Epigenetic Regulation. Plant Physiol. 2016, 172, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mena, C.; de Folter, S.; Costa, M.M.; Angenent, G.C.; Sablowski, R. Transcriptional program controlled by the floral homeotic gene AGAMOUS during early organogenesis. Development. 2005, 132, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Engelhorn, J.; Moreau, F.; Fletcher, J.C.; Carles, C.C. ULTRAPETALA1 and LEAFY pathways function independently in specifying identity and determinacy at the Arabidopsis floral meristem. Ann Bot. 2014, 114, 1497–1505. [Google Scholar] [CrossRef]
- Carles, C.C.; Choffnes-Inada, D.; Reville, K.; Lertpiriyapong, K.; Fletcher, J.C. ULTRAPETALA1 encodes a SAND domain putative transcriptional regulator that controls shoot and floral meristem activity in Arabidopsis. Development. 2005, 132, 897–911. [Google Scholar] [CrossRef]
- Uemura, A.; Yamaguchi, N.; Xu, Y.; Wee, W.; Ichihashi, Y.; Suzuki, T.; Shibata, A.; Shirasu, K.; Ito, T. Regulation of floral meristem activity through the interaction of AGAMOUS, SUPERMAN, and CLAVATA3 in Arabidopsis. Plant Reprod. 2018, 31, 89–105. [Google Scholar] [CrossRef]
- Xu, Y.; Prunet, N.; Gan, E.S.; Wang, Y.; Stewart, D.; Wellmer, F.; Huang, J.; Yamaguchi, N.; Tatsumi, Y.; Kojima, M.; Kiba, T.; Sakakibara, H.; Jack, T.P.; Meyerowitz, E.M.; Ito, T. SUPERMAN regulates floral whorl boundaries through control of auxin biosynthesis. EMBO J. 2018, 37, e97499. [Google Scholar] [CrossRef]
- Sakai, H. ; Krizek, B,A. ; Jacobsen, S.E.; Meyerowitz, E.M. Regulation of SUP expression identifies multiple regulators involved in arabidopsis floral meristem development. Plant Cell. 2000, 12, 1607–1618. [Google Scholar] [CrossRef]
- Pelayo, M.A.; Yamaguchi, N.; Ito, T. One factor, many systems: the floral homeotic protein AGAMOUS and its epigenetic regulatory mechanisms. Curr Opin Plant Biol. 2021, 61, 102009. [Google Scholar] [CrossRef]
- Zúñiga-Mayo, V.M.; Reyes-Olalde, J.I.; Marsch-Martinez, N.; de Folter, S. Cytokinin treatments affect the apical-basal patterning of the Arabidopsis gynoecium and resemble the effects of polar auxin transport inhibition. Front Plant Sci. 2014, 5, 191. [Google Scholar] [CrossRef]
- Cerbantez-Bueno, V.E.; Zúñiga-Mayo, V.M.; Reyes-Olalde, J.I.; Lozano-Sotomayor, P.; Herrera-Ubaldo, H.; Marsch-Martinez, N.; de Folter, S. Redundant and Non-redundant Functions of the AHK Cytokinin Receptors During Gynoecium Development. Front Plant Sci. 2020, 11, 568277. [Google Scholar] [CrossRef]
- Han, N.; Zhu, H.; Li, F.; Wang, M.; Tian, Z.; Wei, J.; Zhang, Z. Genome-wide identification of YABBY genes and functional characterization of CRABS CLAW (AktCRC) in flower development of Akebia trifoliata. Int J Biol Macromol. 2025, 311, 143892. [Google Scholar] [CrossRef]
- Wang, G.; Wu, Z.; Sun, B. KNUCKLES regulates floral meristem termination by controlling auxin distribution and cytokinin activity. Plant Cell. 2024, 37, koae312. [Google Scholar] [CrossRef] [PubMed]
- Hawar, A.; Chen, W.; Zhu, T.; Wang, X.; Liu, J.; Xiong, S.; Ito, T.; Chen, D.; Sun, B. The histone acetyltransferase GCN5 regulates floral meristem activity and flower development in Arabidopsis. Plant Cell. 2025, 37, koaf135. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants. Int J Mol Sci. 2019, 20, 6343. [Google Scholar] [CrossRef]
- Carabelli, M.; Turchi, L.; Morelli, G.; Østergaard, L.; Ruberti, I.; Moubayidin, L. Coordination of biradial-to-radial symmetry and tissue polarity by HD-ZIP II proteins. Nat Commun. 2021, 12, 4321. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yin, L.; Xue, H. Co-expression analysis identifies CRC and AP1 the regulator of Arabidopsis fatty acid biosynthesis. J Integr Plant Biol. 2012, 54, 486–499. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, A.; Liu, C.; Saeed, M.; Li, J.; Wu, Y.; Wu, Y.; Gu, H.; Yuan, J.; Wang, B.; Li, P.; Fang, H. A Comprehensive Analysis In Silico of KCS Genes in Maize Revealed Their Potential Role in Response to Abiotic Stress. Plants (Basel). 2024, 13, 3507. [Google Scholar] [CrossRef]
- Takasugi, T.; Ito, Y. Altered expression of auxin-related genes in the fatty acid elongase mutant oni1 of rice. Plant Signal Behav. 2011, 6, 887–888. [Google Scholar] [CrossRef]
- Reyes-Olalde, J.I.; Zúñiga-Mayo, V.M.; Serwatowska, J.; Chavez Montes, R.A.; Lozano-Sotomayor, P.; Herrera-Ubaldo, H.; Gonzalez-Aguilera, K.L.; Ballester, P.; Ripoll, J.J.; Ezquer, I.; Paolo, D.; Heyl, A.; Colombo, L.; Yanofsky, M.F.; Ferrandiz, C.; Marsch-Martínez, N.; de Folter, S. The bHLH transcription factor SPATULA enables cytokinin signaling, and both activate auxin biosynthesis and transport genes at the medial domain of the gynoecium. PLoS Genet. 2017, 13, e1006726. [Google Scholar] [CrossRef] [PubMed]
- Jamil, I.; Giri, A.; Moubayidin, L. Organ symmetry establishment during gynoecium development. Curr Opin Plant Biol. 2025, 85, 102732. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Baum, S.F.; Bowman, J.L. Distinct mechanisms promote polarity establishment in carpels of Arabidopsis. Cell. 1999, 99, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Azhakanandam, S.; Nole-Wilson, S.; Bao, F.; Franks, R.G. SEUSS and AINTEGUMENTA mediate patterning and ovule initiation during gynoecium medial domain development. Plant Physiol. 2008, 146, 1165–1181. [Google Scholar] [CrossRef]
- Zúñiga-Mayo, V.M.; Marsch-Martínez, N.; de Folter, S. JAIBA, a class-II HD-ZIP transcription factor involved in the regulation of meristematic activity, and important for correct gynoecium and fruit development in Arabidopsis. Plant J. 2012, 71, 314–326. [Google Scholar] [CrossRef]
- Zúñiga-Mayo, V.M.; Marsch-Martínez, N.; de Folter, S. The class II HD-ZIP JAIBA gene is involved in meristematic activity and important for gynoecium and fruit development in Arabidopsis. Plant Signal Behav. 2012, 11, 1501–1503. [Google Scholar] [CrossRef]
- Kuusk, S.; Sohlberg, J.J.; Long, J.A.; Fridborg, I.; Sundberg, E. STY1 and STY2 promote the formation of apical tissues during Arabidopsis gynoecium development. Development. 2002, 129, 4707–4717. [Google Scholar] [CrossRef]
- Sohlberg, J.J.; Myrenås, M.; Kuusk, S.; Lagercrantz, U.; Kowalczyk, M.; Sandberg, G.; Sundberg, E. STY1 regulates auxin homeostasis and affects apical-basal patterning of the Arabidopsis gynoecium. Plant J. 2006, 47, 112–123. [Google Scholar] [CrossRef]
- Bowman, J.L.; Sakai, H.; Jack, T.; Weigel, D.; Mayer, U.; Meyerowitz, E.M. SUPERMAN, a regulator of floral homeotic genes in Arabidopsis. Development. 1992, 114, 599–615. [Google Scholar] [CrossRef]
- Pfannebecker, K.C.; Lange, M.; Rupp, O.; Becker, A. An Evolutionary Framework for Carpel Developmental Control Genes. Mol Biol Evol. 2017, 34, 330–348. [Google Scholar] [CrossRef]
- Dreni, L.; Kater, M.M. MADS reloaded: evolution of the AGAMOUS subfamily genes. New Phytol. 2014, 201, 717–732. [Google Scholar] [CrossRef]
- Phukela, B.; Geeta, R.; Das, S.; Tandon, R. Ancestral segmental duplication in Solanaceae is responsible for the origin of CRCa-CRCb paralogues in the family. Mol Genet Genomics. 2020, 295, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Fourquin, C.; Primo, A.; Martínez-Fernández, I.; Huet-Trujillo, E.; Ferrándiz, C. The CRC orthologue from Pisum sativum shows conserved functions in carpel morphogenesis and vascular development. Ann Bot. 2014, 114, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Ohmori, Y.; Tanaka, W.; Hirabayashi, C.; Murai, K.; Ogihara, Y.; Yamaguchi, T.; Hirano, H.Y. The spatial expression patterns of DROOPING LEAF orthologs suggest a conserved function in grasses. Genes Genet Syst. 2009, 84, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Fourquin, C.; Vinauger-Douard, M.; Fogliani, B.; Dumas, C.; Scutt, C.P. Evidence that CRABS CLAW and TOUSLED have conserved their roles in carpel development since the ancestor of the extant angiosperms. Proc Natl Acad Sci U S A. 2005, 102, 4649–4654. [Google Scholar] [CrossRef]
- Fourquin, C.; Vinauger-Douard, M.; Chambrier, P.; Berne-Dedieu, A.; Scutt, C.P. Functional conservation between CRABS CLAW orthologues from widely diverged angiosperms. Ann Bot. 2007, 100, 651–657. [Google Scholar] [CrossRef]
- Lee, J.Y.; Baum, S.F.; Oh, S.H.; Jiang, C.Z.; Chen, J.C.; Bowman, J.L. Recruitment of CRABS CLAW to promote nectary development within the eudicot clade. Development. 2005, 132, 5021–5032. [Google Scholar] [CrossRef]
- Endress, P.K. The flowers in extant basal angiosperms and inferences on ancestral flowers. Int. J. Plant Sci. 2001, 162, 1111–1140. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Hsiao, Y.Y.; Li, C.I.; Yeh, C.M.; Mitsuda, N.; Yang, H.X.; Chiu, C.C.; Chang, S.B.; Liu, Z.J.; Tsai, W.C. The ancestral duplicated DL/CRC orthologs, PeDL1 and PeDL2, function in orchid reproductive organ innovation. J Exp Bot. 2021, 72, 5442–5461. [Google Scholar] [CrossRef]
- Gong, P.; Song, C.; Liu, H.; Li, P.; Zhang, M.; Zhang, J.; Zhang, S.; He, C. Physalis floridana CRABS CLAW mediates neofunctionalization of GLOBOSA genes in carpel development. J Exp Bot. 2021, 72, 6882–6903. [Google Scholar] [CrossRef]
- Qin, P.; Gao, J.; Shen, W.; Wu, Z.; Dai, C.; Wen, J.; Yi, B.; Ma, C.; Shen, J.; Fu, T.; Tu, J. BnaCRCs with domestication preference positively correlate with the seed-setting rate of canola. Plant J. 2022, 111, 1717–1731. [Google Scholar] [CrossRef]
- Wang, P.; Nolan, T.M.; Yin, Y.; Bassham, D.C. Identification of transcription factors that regulate ATG8 expression and autophagy in Arabidopsis. Autophagy. 2020, 16, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Shivhare, D.; Musialak-Lange, M.; Julca, I.; Gluza, P.; Mutwil, M. Removing auto-activators from yeast-two-hybrid assays by conditional negative selection. Sci Rep. 2021, 11, 5477. [Google Scholar] [CrossRef]
- Strickler, S.R.; Bombarely, A.; Mueller, L.A. Designing a transcriptome next-generation sequencing project for a nonmodel plant species. Am J Bot. 2012, 99, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Klepikova, A.V.; Penin, A.A. Gene Expression Maps in Plants: Current State and Prospects. Plants (Basel). 2019, 8, 309. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Qian, Q.; Lu, M.; Chen, M.; Fan, Z.; Shang, Y.; Bu, C.; Du, Z.; Song, S.; Zeng, J.; Xiao, J. PlantPan: A comprehensive multi-species plant pan-genome database. Plant J. 2025, 122, e70144. [Google Scholar] [CrossRef]
- Elmore, J.M.; Velásquez-Zapata, V.; Wise, R.P. Next-Generation Yeast Two-Hybrid Screening to Discover Protein-Protein Interactions. Methods Mol Biol. 2023, 2690, 205–222. [Google Scholar] [CrossRef]
- Velásquez-Zapata, V.; Elmore, J.M.; Banerjee, S.; Dorman, K.S.; Wise, R.P. Next-generation yeast-two-hybrid analysis with Y2H-SCORES identifies novel interactors of the MLA immune receptor. PLoS Comput Biol. 2021, 17, e1008890. [Google Scholar] [CrossRef]
- Jingwen, W.; Jingxin, W.; Ye, Z.; Yan, Z.; Caozhi, L.; Yanyu, C.; Fanli, Z.; Su, C.; Yucheng, W. Building an improved transcription factor-centered yeast one hybrid system to identify DNA motifs bound by protein comprehensively. BMC Plant Biol. 2023, 23, 236. [Google Scholar] [CrossRef]
Figure 1.
Structure and developmental stages of the gynoecium in Arabidopsis thaliana. Schematic models and transverse sections at stage 5, 8 and 13. Dashed lines indicate the positions of the cross sections. A – apical; B – basal; Ad – adaxial; Ab – abaxial [105].
Figure 1.
Structure and developmental stages of the gynoecium in Arabidopsis thaliana. Schematic models and transverse sections at stage 5, 8 and 13. Dashed lines indicate the positions of the cross sections. A – apical; B – basal; Ad – adaxial; Ab – abaxial [105].
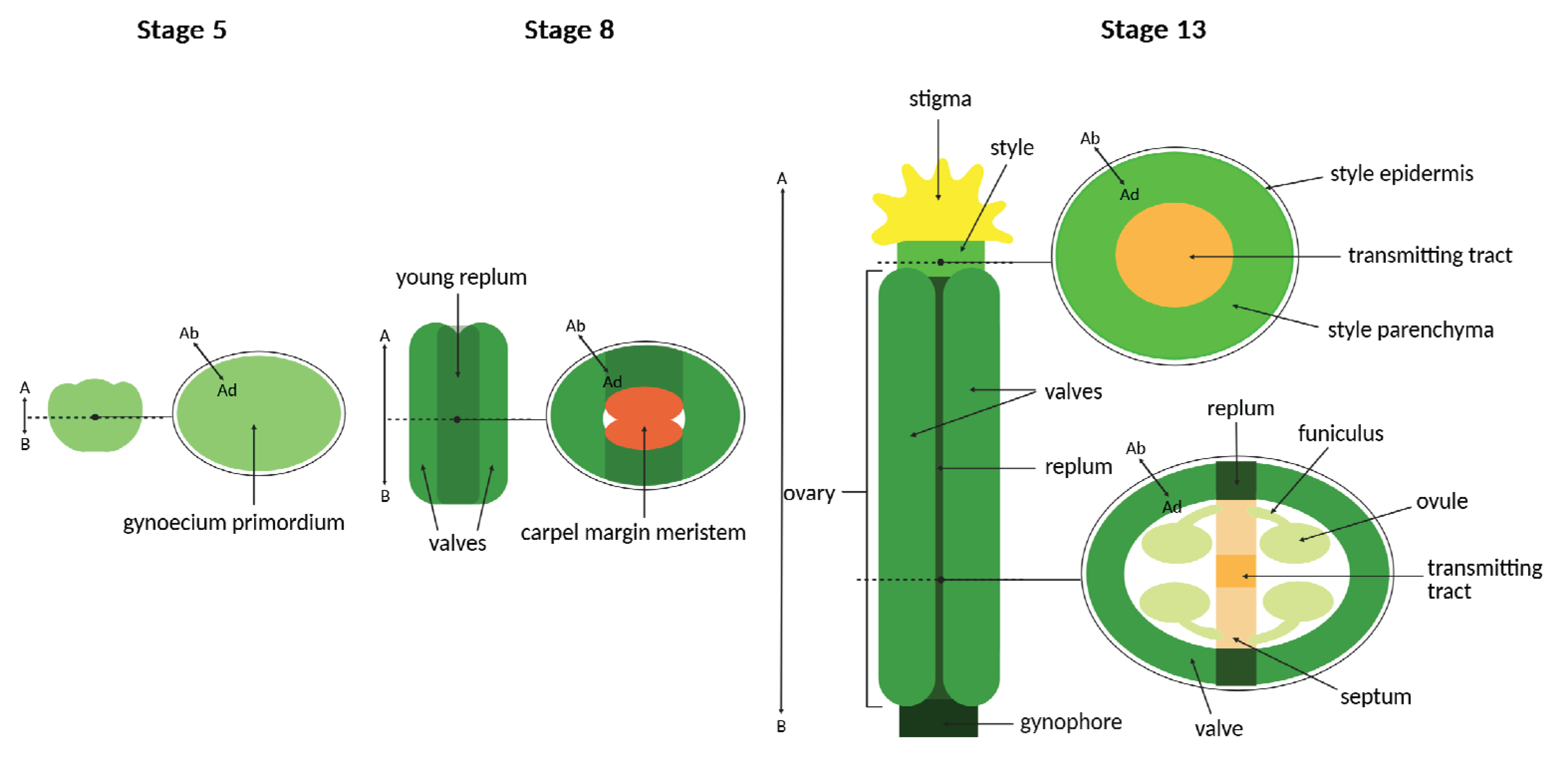
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



