Submitted:
23 July 2025
Posted:
23 July 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The Varicella attenuated virus vaccine, developed in Japan in the 1970s, has dramatically reduced the number of pediatric chickenpox cases over the past 30 years due to its widespread use. However, a small number of cases of chickenpox, singles, and aseptic meningitis caused by vaccine strains have been reported. There are also issues that need to be addressed, such as breakthrough infections and the persistence of the preventive effect of vaccination. In addition, there is the possibility of emergence of revertants or mutations in the vaccine strain. In recent years, subunit vaccines have been developed, their immune-stimulating effects have been demonstrated, and they are being applied clinically. In addition, development of an mRNA varicella vaccine is underway. In this review, the history and impact of the varicella vaccine, are overviewed as well as its future challenges.
Keywords:
varicella zoster virus
; chickenpox
; singles
; attenuated live virus vaccine
; Oka strain
; COVID-19 pandemic
; latent infection
Introduction
Chickenpox is an acute contagious disease caused by varicella zoster virus (VZV). Until the end of the 19th century, chickenpox and smallpox were not clearly distinguished. In 1875 Steiner showed that chickenpox could be caused by inoculation with the blister contents of a chickenpox patient, and in 1888 von Bokay confirmed that a child susceptible to chickenpox could develop chickenpox from contact with a shingles patient. In 1954, Thomas Weller, Nobel Prize winner with John Enders and Frederick Robbins for the isolation of the poliovirus in 1954, confirmed that VZV could be isolated from the blisters of both chickenpox and shingles patients.[1,2] Varicella-zoster virus is a DNA virus belonging to the alpha subfamily of the herpesviridae family, which latently infects sensory ganglia and reactivates to cause shingles. In the 1970s, the attenuated live varicella vaccine was developed for the first time in Japan, and its widespread use has led to a dramatic decrease in the number of chickenpox cases in childhood.
A decrease in various virus infections has been reported during the COVID-19 pandemic due to social behavior restrictions, social distancing, and the promotion of non-pharmaceutical interventions (NPIs).[3,4,5] On the other hand, recovery of virus infections varies depending on the biological characteristics of each virus and differs between regions and countries, probably due to differences in culture, lifestyle, and infection control mitigation policies by country or region.[6]
A review of the trends in fixed-point outbreaks of chickenpox cases in Tokyo, Japan since 2012 shows that, although a prominent downward trend in outbreaks was observed after the introduction of routine two-dose varicella vaccination from 2014, a slight upward trend was observed from 2024 (Figure 1-A). A look at trends by age shows that vaccination has a significant impact on children under 5 years of age, while children 6 years of age and older are almost completely unaffected, and the impact of the COVID-19 pandemic is observed across all age groups (Figure 1-B, C).[7]
Against this background, this review will outline the history and impact of the varicella vaccine, as well as future challenges, especially focusing on childhood. In the text, the term “chickenpox” will be used to refer to disease caused by initial infection with the varicella zoster virus, “shingles” will be used to refer to disease with skin eruptions caused by reactivation after infection or attenuated live vaccination of varicella zoster virus, and “varicella” will be used to refer to infection with the varicella zoster virus in general.
Mechanism of Varicella Zoster Virus (VZV) Infection
Infectious VZV particles adsorbed on airway mucosal epithelium infect T cells or dendritic cells.[8,9,10] These cells are transported to regional lymph nodes, where virus production occurs and infected T cells are transported into the bloodstream to the rest of the body (primary viremia). Infected T cells infiltrate the dermis from blood vessels, migrate to the epithelium while producing virus, and infect epithelial cells with VZV.[11] Infected epithelial cells contribute to the formation of a bullous-pustular rash with virus production. Large amounts of infectious particles are contained within the rash and serve as a source of human-to-human transmission. During this process, infected epithelial cells simultaneously transmit the infection to uninfected T cells, resulting in systemic spread of the infection (secondary viremia). Through Langerhans cells or T cells, VZV eventually reaches the nerve fiber terminals and is transported retrogradely along the axon to the cell body, where it establishes a latent infection that lasts for life.[12,13,14,15]
Glycoprotein B (gB), gE, gH, and gL are thought to be essential for VZV infection of cells.[16] However, the common receptors by which the VZV virus infects a variety of cells, including T cells, monocytes, dendritic cells, and epithelial cells, are not yet known. Cation-independent mannose-6-phospahe receptor (MPRci) has been reported to promote VZV infection [17], but it is not clear which molecules on VZV is associates with. It was also reported that gE associates with insulin degrading enzyme (IDE) and promotes VZV infection.[18] However, IDE and MPRci are not considered sufficient to be called entry receptors in VZV infection, since IDE and MPRci cannot trigger membrane fusion. On the other hand, as for latent infection of neurons, in the search for molecules with homology to paired immunoglobulin-like type 2 receptor α (PILRα), which was identified as a cellular receptor for HSV-1 belonging to the same α-herpes subfamily, myelin-associated glycoprotein (MAG), which is specifically expressed in neurons, was identified.[19] By binding to gangliosides such as NgR1 (Nogo Receptor 1), NgR2, paired Ig-like receptor B (PIR-B), GD1a, and GT1b, MAG inhibits the growth of neuroaxons and is involved in the construction of neural tissue.[20] MAG was found to be an entry receptor for VZV that can associate with VZV gB and trigger membrane fusion.[19]
Genes for Latent Infection
Molecular biological analysis has shown that transcripts such as open reading frame 4 (ORF4), ORF21, ORF29, ORF62, ORF63, ORF66 and ORF66 are expressed in a latently infected state.[21,22] In a rat infection model, ORF4 deletion reduces the frequency of latent infection [23], but ORF21 is not essential for the establishment of latent infection.[24] In VZV-infected rats lacking ORF29, the establishment of latent infection in the trigeminal ganglion was significantly reduced, although the infection rate did not change, and the establishment of latent infection was also reduced when ORF29 was overexpressed, suggesting that the expression level of ORF29 regulates the establishment of latent infection.[25] ORF63 is not essential for growth in cell culture, but has been shown to contribute to the establishment of latent infection in rats.[26]
Recently, a novel viral gene expressed in multiple human trigeminal ganglia was discovered and named VLT (Varicella Zoster Virus Latency-associated Transcript). In an experimental system using cultured cells, this VLT was found to specifically repress the expression of a gene called ORF61, which plays an important role in the very early stage of infection spread during VZV reactivation, suggesting that the VLT may work to maintain latent infection in the human body.[27]
Immune Response to Varicella Zoster Virus (VZV)
From the moment of adsorption to the mucosal epithelium, VZV is exposed to the host immune response. First, the innate immune system triggers the activation of Natural Killer (NK) cells and the production of type I (alpha, beta) and type II (gamma) interferons (IFN). In vitro, both types of IFN inhibit VZV replication. [28,29] It is shown that IFN-α can relieve symptoms in immunocompromised children with chickenpox.[30] With the activation of innate immunity, the production of specific antibodies and T-cell immunity are also induced, and VZV-specific T cells are thought to be important in controlling viral replication both during early infection and during reactivation, especially during recovery after chickenpox onset.[31,32] Reactivation of VZV is associated with a decrease in immunity with aging, particularly with a decrease in cellular mediated immunity (CMI) via VZV-specific T cells.[33] Because CMI can be stimulated by additional varicella vaccine immunization to suppress the onset of singles [34], additional vaccination of the elderly is recommended recently as discussed later.
Attenuated Live Vaccine Against Varicella Zoster Virus (VZV)
The varicella vaccine strain was isolated in 1971 from a Japanese patient named Oka as the parent strain (wild strain), and after 11 passages in human cells at the low temperature of 34°C, and then 12 passages were made in fetal guinea pig cells to establish vaccine strain (vaccine Oka strain: vOka-strain). Then, in 1974, Japan reported that attenuated live vaccine was effective in preventing varicella infection in pediatric patients with secondary immunodeficiency.[35] Between vOka and wild-Oka strains there are 42 nucleotide sequence substitutions and 20 amino acid substitutions, 8 of which are concentrated in the immediately early gene62 (IE62gene: the transactivator that is first initiated by intracellular growth). [36]
The mechanism of attenuated Oka vaccine strains (vOka strain) is still largely unknown. It is known that the wild strain blocks the production of Th1 cytokines IFN-γ and IL-12 by inhibiting the innate immune pathway by TLR2, thereby inhibiting induction of Th1 by DC, while the vaccine strain rather promotes IL-12 production.[37] In terms of infection kinetics, the vaccine strain infects T cells and proliferates as well as the wild type [8,38], but has been shown in the SCID-hu(skin) model to have reduced proliferative potential in the skin.[39] At the gene level, there are differences at 42 base sites between parental and vaccine strains[36], and differential activation of viral genes by ORF62 protein (IE62) has been reported [40] and the presence of attenuated markers of proliferation in the skin in the region of ORF30-55 [41], suggesting that multiple gene products are likely to be involved in attenuated viral growth.
Are Varicella Vaccinees Infectious?
A relatively large randomized control study was conducted in the early stage of vaccine introduction. For sibling examples, one patient was inoculated with attenuated live vaccine and another with placebo, and the results were observed. 6/493 (1%) in the placebo group showed increased antibody titer (infection) (no appearance of rash), and no rash was observed in the siblings of those with increased antibody titer. Based on the pattern of antibody titer elevation, 3 of the 6 patients may have been inoculated with attenuated live vaccine by mistake. The possibility of “false positives” remains for the positive antibody titers. They concluded that the infectivity of the vaccinated is almost negligible.[42]
On the other hand, eight cases (aged 4 months to 39 years) who developed chickenpox after a family member of 6 cases (2 adults and 4 children) developed a rash after varicella vaccination. From all eight patients with chickenpox, vaccine strain was isolated. All vaccinees have rash appearance 12₋24 days after vaccination, and infected patients have onset of disease 16-21 days after rash appearance in vaccinees. None of the infected patients were immunocompromised, and all were mildly ill. Furthermore, five cases (two to 35 years old) with chickenpox have been reported who were infected from the vaccinees who developed singles after 8months to 2 years after vaccination. From all five patients, vaccine strain was isolated. Four of the infected patients were within the family and one was within the school. All had mild rashes, and one had mild meningitis.[43]
Varicella vaccination was given to 575 children with ALL (acute lymphocytic leukemia) who were in remission after treatment (10%) or during maintenance treatment (90%). In ALL patients under maintenance treatment, treatment was discontinued before one week and after two weeks of vaccination. Rash appeared in 40% of the patients, on average 30 days (7-41 days) after vaccination. 21 siblings (23%) of the 93 patients who had rash after vaccination were infected. 16 siblings had rash after 14-22 days and vaccine strain was isolated from 4 siblings. 5 patients were asymptomatic but antibody titers increased.[44] Infectivity after vaccination with attenuated live varicella vaccine is almost negligible, but if the vaccinees present with a rash or blistering rash after vaccination, they are considered to be infectious.
The results of a 10-year post-marketing surveillance indicate that adverse reactions to vaccination are few (<5%) and mild. Among the adverse reactions, skin rashes other than at the vaccination site were reported to include rash (4.2%), erythema (1.4%), and blistering (0.63%), with an estimated total frequency of skin rash of less than 0.5%.[45]
Efficacy of Attenuated Live Varicella Vaccine in Preventing Chickenpox
A study in European countries reported that more than 90% of children under 10 years of age had chickenpox.[46] A survey conducted before vaccine dissemination in Japan reported that 96% had chickenpox by the age of 7 years.[47]
A double-blind randomized study in children has shown that a single dose of attenuated live vaccine provides 90-100% varicella protection for several years.[42,48] The safety and efficacy of attenuated live vaccine for uninfected adults was also confirmed, albeit on a small scale.[49] Based on the above results, the varicella attenuated live vaccine was approved in the United States in 1995. Between 1995-2006, the number of pediatric chickenpox cases decreased by 70%, hospitalizations for varicella decreased by 80%, and deaths from varicella decreased by 97% in the United States. However, antibody testing using the FAMA (Fluorescent Antibody to Membrane Antigen) assay showed that a single vaccination did not produce sufficient antibodies in 24% of cases.[50] In 2007, two-dose vaccination was recommended, and the number of chickenpox cases further decreased.[51]
In Japan, which was the first country in the world to develop attenuated live varicella vaccine, the attenuated live varicella vaccine was approved in 1986 and vaccination began the following year. For a while, voluntary inoculation was performed by self-pay, and one-dose inoculation was the mainstream. Subsequently, routine vaccination with two doses at public expense was introduced in 2014. A similar downward trend in chickenpox cases has been reported from Japan.[52]
Childhood Shingles
A study of residents in Rochester, Minnesota, reviewing medical records from 1960-1981, reported a 2.8 to 20.9-fold higher incidence of shingles under age 20 years if the patient had chickenpox within the first year of life.[53] It is considered that this is due to the fact that VZV spreads over a wide area of the body due to immature immunity, and that sufficient acquired immunity cannot be obtained in children under one year of age. Similarly, it has been reported that chickenpox incidence before the age of 1 year is a risk factor for childhood shingles.[54,55] Childhood shingles is less common than in adults and the elderly, mostly in secondary immunocompromised patients with childhood cancer or immunosuppressive drugs.[56] Before and after the introduction of varicella vaccine, childhood shingles was reported to have decreased from 20-60/100,000 person-years to 14/100,000 person-years.[53,57]
Fewer cases of shingles are reported with the varicella vaccine strain, with a 79% reduction in shingles in vaccinated individuals compared to natural infection.[58] A population-based active surveillance study in Antelope Valley, California, USA, found a 55% reduction in the incidence of shingles among those under 10 years of age in 2006 compared to 2000. It also found that the incidence of shingles was 4 to 12 times lower in vaccinated individuals than in individuals infected with wild varicella strain virus.[59] A single-center retrospective observational study in Japan also reported a decrease in shingles cases among healthy children from 5.0/year to 3.3/year between 1990-2000 and 2001-2017 after the introduction of varicella vaccine.[60] The lower incidence of shingles in varicella vaccinees may be due to the lower toxicity of the vaccine strain compared to the wild strain.
There is a difference in the site of onset (dermadrome) between singles after chickenpox and singles after vaccination. A report summarizes 11 cases of childhood shingles in which the vaccine strain was identified, ranging in age from 1 year and 7 months to 14 years, and one case had a history of chickenpox, as shown in the Table 1. Ten cases were after one dose of vaccine and one after two doses; three cases developed meningitis and one encephalitis. The most common sites of shingles are in the cervical and lumbar areas. [61] This is markedly different from the incidence of singles in adults and elderly after chickenpox, which is reported to occur in 17.6% for head to face, 14.5% for neck to upper extremities, 31.2% for upper extremities to back of chest, 19.6% for abdominal back, and 17.2% for buttocks to lower extremities.[62]
In the case of chickenpox, viremia occurs in 100% of cases, the virus can spread throughout the body, and the virus migrates retrogradely from the sensory nerve fibers at the blister site to the sensory ganglia, where it causes latent infection. In the case of vaccination of the upper extremities, latent infection occurs in the cervical spine area, and in the case of vaccination of the thighs and buttocks, latent infection occurs in the lumbar spine area, resulting in the onset of shingles from these areas. In addition, reports from Japan [63] and the United States [64], which examined the incidence of singles after vaccination of pediatric patients with leukemia, both found that the frequency of shingles was 10 times higher in cases in which a rash appeared after vaccination than in those in which it did not. On the other hand, studies from the United States [65], Europe [66], and Japan [67] both found that shingles after vaccination of healthy individuals tends to occur at a younger age and the time between vaccination and shingles is shorter compared to that between chickenpox and singles. In cases where the rash appeared after vaccination, it is possible that sufficient immunity could not be acquired.
The incidence of postherpetic neuralgia is reported to be much less common in child- to adolescent-onset singles than in adult- or elderly-onset singles [68].
Meningitis Due to Reactivation of Varicella Virus
A small number of cases of meningitis have been reported in healthy individuals with the varicella vaccine strain. Most cases are meningitis associated with the onset of singles. Cases of meningitis due to reactivation of wild strain not presenting singles have been reported.[69,70] There is one case report of vaccine strain meningitis due to reactivation not presenting shingles from a vaccinee.[71] A healthy 15-year-old female with a chief complaint of fever and headache was found to have varicella zoster virus of the vaccine strain in her spinal fluid; she had received one dose of live attenuated varicella vaccine at age 1 and had no history of chickenpox.
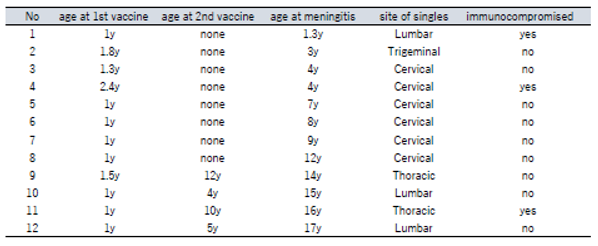
Reports of 12 cases of childhood varicella meningitis in which vaccine strains were identified are summarized (Table 2).[72] Eight cases were after 1 vaccination (2 had carcinoma, 6 were healthy) and 4 after 2 vaccinations (3 were healthy). All cases developed meningitis after shingles (0.3 to 11 years after vaccination). Six cases presented singles in the cervical area, one in the trigeminal area, and one in the lumbar area (infant immunized in the thigh). A report comparing childhood meningitis caused by vaccine strain reactivation versus wild strain reactivation found that vaccine strain cases were younger (7 ± 3.4 years vs. 11.9 ± 3.6 years, P = 0.0038) and were more related with singles (100% vs. 55%, P = 0.04).[69] In cases of chickenpox, the varicella virus latently infects the trigeminal ganglia due to the appearance of a varicella rash on the head and face and viremia. If reactivation occurs there, the infection may spread directly to the meninges even if shingles is not seen in the trigeminal area. Meningitis from singles after vaccination is thought to progress from the sensory ganglia to the meninges via the meningeal cavity [72].
Maintenance of Immunity to VZV
The National Institute of Health Risk Management in Japan has been investigating trends in varicella antibody titers by age as part of its “Survey on the Prediction of Infectious Disease Epidemics”. The graph of varicella antibody titers by age (2020-2024) (Figure 2) shows that more than half of the generation within a few years of receiving two varicella vaccines have EIA (enzyme immunoassay) antibody titers <4 [73](antibody titers in children under 5 months are thought to indicate transferred antibodies from the mother). Referring to data on varicella vaccination coverage since 2016 from the public website [74], around 95% for one dose of varicella vaccine and 90% for two doses of varicella vaccine in Japan are maintained. In other words, more than half of infants have EIA antibody titers <4 even after two doses of varicella vaccine (In fact, trends in antibody titers show a transient increase in the ratio of EIA antibody titers >4 at 2 years of age immediately after the two vaccinations. In Japan, two varicella vaccinations are given between the ages of 1 and 2 years).
Why is the increase in antibody titers not sufficient in infants even after two doses of vaccine, and why are antibody titers elevated in adults? It is antigen stimulation (i.e., exposure to the varicella virus) that drives the rise in antibody titers. There are two possible explanations for this: external exposure and internal exposure. Before the varicella vaccine was introduced, in environments where sufficient chickenpox cases were occurring, adults who already had varicella immunity would have had the opportunity to have their immunity stimulated by exposure to VZV in their daily lives. It has been shown that contact of a person with a history of varicella infection with a varicella patient stimulates immunity and increases immunocompetence.[75] However, when the varicella vaccine is introduced and the number of chickenpox cases in children declines by more than 90%, external exposure decreases dramatically. However, surveys of trends in anti-varicella antibody titers in adults show no significant change since 2014.
The other is internal exposure (stimulation). As you know, the varicella zoster virus, a double-stranded DNA virus, like the herpes virus of the same genus, once infected, latently infects ganglion cells and other organs in the body. Reactivation and proliferation in the skin along the nerves results in singles. The internal stimulation theory suggests that even if singles does not develop, mild reactivation may occur asymptomatically, which may provide antigenic stimulation and boost immunocompetence. This was first advocated by Hope-Simpson in 1952.[76] Subsequently, searches for the virus in the saliva of astronauts returning from space travel have confirmed the reactivation of varicella zoster virus asymptomatically.[77] In addition, the possibility of asymptomatic reactivation of varicella zoster virus in children and adults in general has been reported.[78] Another epidemiological study found no significant difference in the incidence of singles between populations with no contact with children and those with contact with children.[79] This indicates that latent infection plays an important role in maintaining anti-varicella virus immunity over the long term.
As mentioned previously, the development of shingles due to reactivation of the varicella zoster virus is associated with a decrease in the function of varicella-specific T cells.[33] Concerns were raised that the introduction of the varicella vaccine would lead to an increase in shingles in adults and the elderly because of a drastic decrease in the number of chickenpox patients, which would reduce the external stimuli to maintain varicella-specific T cells.[80] While shingles in adults has continued to increase as chickenpox cases in children have decreased dramatically with the introduction of the varicella vaccine, the incidence of shingles has shown an increasing trend since the 1950s, long before the vaccine was introduced.[81] Multiple factors are thought to contribute to the increase in shingles, including increased stress in society, medically-induced immunosuppression, an aging population, and increased diagnostic rates.
To reduce the increasing incidence of shingles, varicella-specific T cells must be stimulated and augmented.[33] Additional vaccination with attenuated live attenuated vaccine has been shown to enhance varicella-specific T cells and contribute to the prevention of shingles development. Immunization with attenuated live attenuated vaccine reduced the incidence of singles by 51.3% (315/19,254 vs. 642/19,247) and postherpetic neuralgia by 66.5% (27/19,254 vs. 80/19,247) in persons over 50 years old.[82,83,84]
If the patient is already immune through natural infection or attenuated live vaccination, a subunit vaccine has been shown to be effective enough to stimulate varicella-specific T cells. In a study of subjects aged 50 years and older, the subunit vaccination group showed an 89.8% reduction in incidence versus the placebo group over a mean 3.7-year observation period (23/6,950 vs. 223/6,950). There was also a 90.0% reduction in the 70-79 age group and an 89.1% reduction in the 80+ age group, independent of age.[85,86] It has attracted attention as an immune-boosting strategy that can be safely used in immunocompromised individuals because there is no risk of developing chickenpox or shingles due to subunit vaccine compared to attenuated live vaccine.[87] Following the subunit vaccine, an mRNA vaccine that attracted attention as a vaccine against SARS-CoV-2 has been developed as a varicella vaccine and its efficacy in animal models has been reported.[88]
Problems and Future Challenges with Varicella Attenuated Live Vaccine
<Breakthrough Infection>
Breakthrough infections were 3-5% per year over a 3-year observation period (431 patients aged 3-15 years). However, all of them were reported to have a milder disease than infection by wild strains.[89] A 7-year observational study found that annual breakthrough infections were 0.2-2.3%, cumulative breakthrough infections were less than 6.5%, and symptoms were all mild.[90] A report from Japan reported that in a follow-up study of 593 persons who received the attenuated live vaccine in 1987-1989, a cumulative total of 157/459 (34.2%) had breakthrough infections by 1996, all of which were mild cases.[91] In a 10-year observational study of approximately 2,000 children aged 1 to 12 years, the authors found 94.4% protection with one dose of vaccine and 98.3% with two doses. It also states that antibodies were detected in all vaccinated individuals over a 9-year period.[92]
<Persistence of Immunity>
Although various observational and epidemiological studies suggest that the vaccine’s efficacy in preventing varicella persists for 10-20 years or longer, there are variable factors such as the boosting effect due to the prevalence of chickenpox, and some points are still unclear. With the widespread use of the attenuated live varicella vaccine, the number of wild-type chickenpox cases has been drastically decreasing, especially in developed countries. It may be necessary to examine and verify whether the current vaccine program is sufficient to prevent sudden outbreaks of wild-type varicella or novel mutated varicella virus.
<Trends In Shingles Caused by Vaccine Strains>
As mentioned previously, the incidence of shingles caused by vaccine strains has been reported to be lower than that of wild strains. However, even in developed countries, only 30-40 years have passed since the vaccine was introduced. Advances in medicine and changes in social structure, such as an aging population, may lead to an increase in the number of people with secondary immunosuppressive conditions, which could significantly alter the population immunity situation. Long-term effects of vaccines are still unknown.
<Emergence of Mutant Vaccine Strains>
Quinlivan et al. compared the sequences of viruses isolated from skin eruptions caused by vaccine strains with those of vaccine strains and reported that several SNPs were found. However, these SNPs were not associated with increased viral virulence. [93,94] There have been no reports so far of reversion of vaccine strains to the parent strain or mutation to a highly virulent strain. However, monitoring by surveillance systems remains important.
Conclusions
The attenuated live varicella vaccine developed in Japan in the 1970s dramatically reduced the number of chickenpox cases in children over a period of about 30 years. It is regarded as a highly safe vaccine with very few adverse reactions. On the other hand, it is necessary to consider whether the current vaccine program for varicella infection is sufficient in an era when the number of vaccine strain infections exceeds the number of wild strain infections. Subunit and mRNA vaccines are promising new approaches to maintaining herd anti-varicella immunity. In addition, careful monitoring is needed to ensure that possible emergence of varicella infections caused by newly mutated wild strains or vaccine strains could not pose a new threat.
Author Contributions
Conceptualization, methodology, formal analysis, review, and editing, M.N.
Funding
This research received no external funding.
Data Availability Statement
Data available on request from the authors. The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
I would like to thank the staff of the Infection Control Office, the Department of Infectious Diseases, and the Department of Pediatrics at the Musashino Red Cross Hospital for their constructive exchange of ideas in the preparation of this review.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Weller TH. Serial propagation in vitro of agents producing inclusion bodies derived from varicella and herpes zoster. Proc Soc Exp Biol Med 1953;83:340-6. [CrossRef]
- Ligon BL. Thomas Huckle Weller MD: Nobel Laureate and research pioneer in poliomyelitis, varicella-zoster virus, cytomegalovirus, rubella, and other infectious diseases. Semin Pediatr Infect Dis 2002;13:55-63.
- Cohen R, Ashman M, Taha MK, Varon E, Angoulvant F, Levy C, Rybak A, Ouldali N, Guiso N, Grimprel E. Pediatric Infectious Disease Group (GPIP) position paper on the immune debt of the COVID-19 pandemic in childhood, how can we fill the immunity gap? Infect Dis Now 2021;51:418-23. [CrossRef]
- Principi N, Autore G, Ramundo G, Esposito S. Epidemiology of Respiratory Infections during the COVID-19 Pandemic. Viruses 2023;15:1160. [CrossRef]
- Yang MC, Su YT, Chen PH, Tsai CC, Lin TI, Wu JR. Changing patterns of infectious diseases in children during the COVID-19 pandemic. Front Cell Infect Microbiol 2023;13:1200617. [CrossRef]
- Nagasawa M. Verification of Immune Debts in Children Caused by the COVID-19 Pandemic from an Epidemiological and Clinical Perspective. Immuno 2025;5:5. [CrossRef]
- https://survey.tmiph.metro.tokyo.lg.jp/epidinfo/epimenu.do. accessed on June 8, 2025.
- Soong W, Schultz JC, Patera AC, Sommer MH, Cohen JI. Infection of human T lymphocytes with varicella-zoster virus: an analysis with viral mutants and clinical isolates. J Virol 2000;74:1864-70.
- Ku CC, Padilla JA, Grose C, Butcher EC, Arvin AM. Tropism of varicella-zoster virus for human tonsillar CD4(+) T lymphocytes that express activation, memory, and skin homing markers. J Virol 2002;76:11425-33.
- Abendroth A, Morrow G, Cunningham AL, Slobedman B. Varicella-zoster virus infection of human dendritic cells and transmission to T cells: implications for virus dissemination in the host. J Virol 2001;75:6183-92.
- Ku CC, Zerboni L, Ito H, Graham BS, Wallace M, Arvin AM. Varicella-zoster virus transfer to skin by T Cells and modulation of viral replication by epidermal cell interferon-alpha. J Exp Med 2004;200:917-25. [CrossRef]
- Levin MJ, Cai GY, Manchak MD, Pizer LI. Varicella-zoster virus DNA in cells isolated from human trigeminal ganglia. J Virol 2003;77:6979-87. [CrossRef]
- Annunziato PW, Lungu O, Panagiotidis C, Zhang JH, Silvers DN, Gershon AA, Silverstein SJ. Varicella-zoster virus proteins in skin lesions: implications for a novel role of ORF29p in chickenpox. J Virol 2000;74:2005-10. [CrossRef]
- Nikkels AF, Debrus S, Sadzot-Delvaux C, Piette J, Rentier B, Pierard GE. Localization of varicella-zoster virus nucleic acids and proteins in human skin. Neurology 1995;45:S47-S9.
- Weigle KA, Grose C. Common expression of varicella-zoster viral glycoprotein antigens in vitro and in chickenpox and zoster vesicles. J Infect Dis 1983;148:630-8. [CrossRef]
- Cole NL, Grose C. Membrane fusion mediated by herpesvirus glycoproteins: the paradigm of varicella-zoster virus. Rev Med Virol 2003;13:207-22.
- Chen JJ, Zhu Z, Gershon AA, Gershon MD. Mannose 6-phosphate receptor dependence of varicella zoster virus infection in vitro and in the epidermis during varicella and zoster. Cell 2004;119:915-26. [CrossRef]
- Li Q, Ali MA, Cohen JI. Insulin degrading enzyme is a cellular receptor mediating varicella-zoster virus infection and cell-to-cell spread. Cell 2006;127:305-16.
- Suenaga T, Satoh T, Somboonthum P, Kawaguchi Y, Mori Y, Arase H. Myelin-associated glycoprotein mediates membrane fusion and entry of neurotropic herpesviruses. Proc Natl Acad Sci U S A 2010;107:866-71. [CrossRef]
- Quarles RH. A hypothesis about the relationship of myelin-associated glycoprotein’s function in myelinated axons to its capacity to inhibit neurite outgrowth. Neurochem Res 2009;34:79-86.
- Cohrs RJ, Barbour M, Gilden DH. Varicella-zoster virus (VZV) transcription during latency in human ganglia: detection of transcripts mapping to genes 21, 29, 62, and 63 in a cDNA library enriched for VZV RNA. J Virol 1996;70:2789-96. [CrossRef]
- Kennedy PG, Grinfeld E, Bell JE. Varicella-zoster virus gene expression in latently infected and explanted human ganglia. J Virol 2000;74:11893-8.
- Cohen JI, Krogmann T, Ross JP, Pesnicak L, Prikhod’ko EA. Varicella-zoster virus ORF4 latency-associated protein is important for establishment of latency. J Virol 2005;79:6969-75. [CrossRef]
- Xia D, Srinivas S, Sato H, Pesnicak L, Straus SE, Cohen JI. Varicella-zoster virus open reading frame 21, which is expressed during latency, is essential for virus replication but dispensable for establishment of latency. J Virol 2003;77:1211-8.
- Cohen JI, Krogmann T, Pesnicak L, Ali MA. Absence or overexpression of the Varicella-Zoster Virus (VZV) ORF29 latency-associated protein impairs late gene expression and reduces VZV latency in a rodent model. J Virol 2007;81:1586-91. [CrossRef]
- Cohen JI, Krogmann T, Bontems S, Sadzot-Delvaux C, Pesnicak L. Regions of the varicella-zoster virus open reading frame 63 latency-associated protein important for replication in vitro are also critical for efficient establishment of latency. J Virol 2005;79:5069-77.
- Depledge DP, Ouwendijk WJD, Sadaoka T, Braspenning SE, Mori Y, Cohrs RJ, Verjans G, Breuer J. A spliced latency-associated VZV transcript maps antisense to the viral transactivator gene 61. Nat Commun 2018;9:1167. [CrossRef]
- Balachandra K, Thawaranantha D, Ayuthaya PI, Bhumisawasdi J, Shiraki K, Yamanishi K. Effects of human alpha, beta and gamma interferons on varicella zoster virus in vitro. Southeast Asian J Trop Med Public Health 1994;25:252-7.
- Desloges N, Rahaus M, Wolff MH. Role of the protein kinase PKR in the inhibition of varicella-zoster virus replication by beta interferon and gamma interferon. J Gen Virol 2005;86:1-6.
- Arvin AM, Kushner JH, Feldman S, Baehner RL, Hammond D, Merigan TC. Human leukocyte interferon for the treatment of varicella in children with cancer. N Engl J Med 1982;306:761-5. [CrossRef]
- Arvin AM, Koropchak CM, Williams BR, Grumet FC, Foung SK. Early immune response in healthy and immunocompromised subjects with primary varicella-zoster virus infection. J Infect Dis 1986;154:422-9.
- Kumagai T, Chiba Y, Wataya Y, Hanazono H, Chiba S, Nakao T. Development and characteristics of the cellular immune response to infection with varicella-zoster virus. J Infect Dis 1980;141:7-13. [CrossRef]
- Burke BL, Steele RW, Beard OW, Wood JS, Cain TD, Marmer DJ. Immune responses to varicella-zoster in the aged. Arch Intern Med 1982;142:291-3.
- Oxman MN, Levin MJ. Vaccination against Herpes Zoster and Postherpetic Neuralgia. J Infect Dis 2008;197 Suppl 2:S228-36.
- Takahashi M, Otsuka T, Okuno Y, Asano Y, Yazaki T. Live vaccine used to prevent the spread of varicella in children in hospital. Lancet 1974;2:1288-90.
- Gomi Y, Sunamachi H, Mori Y, Nagaike K, Takahashi M, Yamanishi K. Comparison of the complete DNA sequences of the Oka varicella vaccine and its parental virus. J Virol 2002;76:11447-59. [CrossRef]
- Gutzeit C, Raftery MJ, Peiser M, Tischer KB, Ulrich M, Eberhardt M, Stockfleth E, Giese T, Sauerbrei A, Morita CT, et al. Identification of an important immunological difference between virulent varicella-zoster virus and its avirulent vaccine: viral disruption of dendritic cell instruction. J Immunol 2010;185:488-97.
- Moffat JF, Stein MD, Kaneshima H, Arvin AM. Tropism of varicella-zoster virus for human CD4+ and CD8+ T lymphocytes and epidermal cells in SCID-hu mice. J Virol 1995;69:5236-42. [CrossRef]
- Moffat JF, Zerboni L, Kinchington PR, Grose C, Kaneshima H, Arvin AM. Attenuation of the vaccine Oka strain of varicella-zoster virus and role of glycoprotein C in alphaherpesvirus virulence demonstrated in the SCID-hu mouse. J Virol 1998;72:965-74.
- Gomi Y, Imagawa T, Takahashi M, Yamanishi K. Oka varicella vaccine is distinguishable from its parental virus in DNA sequence of open reading frame 62 and its transactivation activity. J Med Virol 2000;61:497-503.
- Zerboni L, Hinchliffe S, Sommer MH, Ito H, Besser J, Stamatis S, Cheng J, Distefano D, Kraiouchkine N, Shaw A, et al. Analysis of varicella zoster virus attenuation by evaluation of chimeric parent Oka/vaccine Oka recombinant viruses in skin xenografts in the SCIDhu mouse model. Virology 2005;332:337-46.
- Weibel RE, Neff BJ, Kuter BJ, Guess HA, Rothenberger CA, Fitzgerald AJ, Connor KA, McLean AA, Hilleman MR, Buynak EB, et al. Live attenuated varicella virus vaccine. Efficacy trial in healthy children. N Engl J Med 1984;310:1409-15. [CrossRef]
- Marin M, Leung J, Gershon AA. Transmission of Vaccine-Strain Varicella-Zoster Virus: A Systematic Review. Pediatrics 2019;144(3):e20191305.
- Gershon AA, LaRussa P, Steinberg S. The varicella vaccine. Clinical trials in immunocompromised individuals. Infect Dis Clin North Am 1996;10:583-94. [CrossRef]
- Willis ED, Woodward M, Brown E, Popmihajlov Z, Saddier P, Annunziato PW, Halsey NA, Gershon AA. Herpes zoster vaccine live: A 10 year review of post-marketing safety experience. Vaccine 2017;35:7231-9.
- Bollaerts K, Riera-Montes M, Heininger U, Hens N, Souverain A, Verstraeten T, Hartwig S. A systematic review of varicella seroprevalence in European countries before universal childhood immunization: deriving incidence from seroprevalence data. Epidemiol Infect 2017;145:2666-77.
- Saito M, Haruyama C, Ohba H, Wada A, Takeuchi Y. [A seroepidemiological study of varicella]. Kansenshogaku Zasshi 1987;61:783-8.
- Varis T, Vesikari T. Efficacy of high-titer live attenuated varicella vaccine in healthy young children. J Infect Dis 1996;174 Suppl 3:S330-4. [CrossRef]
- Gershon AA, Steinberg SP, LaRussa P, Ferrara A, Hammerschlag M, Gelb L. Immunization of healthy adults with live attenuated varicella vaccine. J Infect Dis 1988;158:132-7.
- Shapiro ED, Vazquez M, Esposito D, Holabird N, Steinberg SP, Dziura J, LaRussa PS, Gershon AA. Effectiveness of 2 doses of varicella vaccine in children. J Infect Dis 2011;203:312-5.
- Gershon AA, Gershon MD, Shapiro ED. Live Attenuated Varicella Vaccine: Prevention of Varicella and of Zoster. J Infect Dis 2021;224:S387-S97. [CrossRef]
- Uda K, Okubo Y, Tsuge M, Tsukahara H, Miyairi I. Impacts of routine varicella vaccination program and COVID-19 pandemic on varicella and herpes zoster incidence and health resource use among children in Japan. Vaccine 2023;41:4958-66.
- Guess HA, Broughton DD, Melton LJ, 3rd, Kurland LT. Epidemiology of herpes zoster in children and adolescents: a population-based study. Pediatrics 1985;76:512-7.
- Baba K, Yabuuchi H, Takahashi M, Ogra PL. Increased incidence of herpes zoster in normal children infected with varicella zoster virus during infancy: community-based follow-up study. J Pediatr 1986;108:372-7. [CrossRef]
- Terada K, Kawano S, Yoshihiro K, Miyashima H, Morita T. Characteristics of herpes zoster in otherwise normal children. Pediatr Infect Dis J 1993;12:960-1.
- Lin HC, Chao YH, Wu KH, Yen TY, Hsu YL, Hsieh TH, Wei HM, Wu JL, Muo CH, Hwang KP, et al. Increased risk of herpes zoster in children with cancer: A nationwide population-based cohort study. Medicine (Baltimore) 2016;95:e4037.
- Leung AK, Robson WL, Leong AG. Herpes zoster in childhood. J Pediatr Health Care 2006;20:300-3.
- Weinmann S, Chun C, Schmid DS, Roberts M, Vandermeer M, Riedlinger K, Bialek SR, Marin M. Incidence and clinical characteristics of herpes zoster among children in the varicella vaccine era, 2005-2009. J Infect Dis 2013;208:1859-68. [CrossRef]
- Harpaz R, Leung JW. The Epidemiology of Herpes Zoster in the United States During the Era of Varicella and Herpes Zoster Vaccines: Changing Patterns Among Children. Clin Infect Dis 2019;69:345-7.
- Terada K, Wakabayashi S, Ono S, Tanaka Y, Kato A, Teranishi H, Miyata I, Ogita S, Oishi T, Ohno N, et al. Characteristics of zoster in otherwise healthy children‒ a comparative study between 55 zoster patients between 1990 and 2000.
- (11 years) and 56 from 2001 to 2017 (17 years) ‒. Pediatric Infectious Immunity 2019;31:3-6.
- Shang BS, Hung CJ, Lue KH. Herpes Zoster in an Immunocompetent Child without a History of Varicella. Pediatr Rep 2021;13:162-7.
- Ishikawa H, Tamai K, Mibou K, Tunoda T, Sawamura D, Umeki K, Sugawara T, Yajima H, Sasaki C, Kumano T, et al. A multicenter, joint annual statistical analysis of herpes zoster (April 2000-March 2001). The Japanese Journal of Dermatology 2003;113:1229-39.
- Takahashi M, Baba K, Horiuchi K, Kamiya H, Asano Y. A live varicella vaccine. Adv Exp Med Biol 1990;278:49-58.
- Hardy I, Gershon AA, Steinberg SP, LaRussa P. The incidence of zoster after immunization with live attenuated varicella vaccine. A study in children with leukemia. Varicella Vaccine Collaborative Study Group. N Engl J Med 1991;325:1545-50. [CrossRef]
- Galea SA, Sweet A, Beninger P, Steinberg SP, Larussa PS, Gershon AA, Sharrar RG. The safety profile of varicella vaccine: a 10-year review. J Infect Dis 2008;197 Suppl 2:S165-9.
- Goulleret N, Mauvisseau E, Essevaz-Roulet M, Quinlivan M, Breuer J. Safety profile of live varicella virus vaccine (Oka/Merck): five-year results of the European Varicella Zoster Virus Identification Program (EU VZVIP). Vaccine 2010;28:5878-82.
- Yoshikawa T, Ando Y, Nakagawa T, Gomi Y. Safety profile of the varicella vaccine (Oka vaccine strain) based on reported cases from 2005 to 2015 in Japan. Vaccine 2016;34:4943-7. [CrossRef]
- Petursson G, Helgason S, Gudmundsson S, Sigurdsson JA. Herpes zoster in children and adolescents. Pediatr Infect Dis J 1998;17:905-8.
- Amaral V, Shi JZ, Tsang AM, Chiu SS. Primary varicella zoster infection compared to varicella vaccine reactivation associated meningitis in immunocompetent children. J Paediatr Child Health 2021;57:19-25.
- Barry R, Prentice M, Costello D, O’Mahony O, DeGascun C, Felsenstein S. Varicella Zoster Reactivation Causing Aseptic Meningitis in Healthy Adolescents: A Case Series And Review Of The Literature. Pediatr Infect Dis J 2020;39:e278-e82.
- Kawamura Y, Suzuki D, Kono T, Miura H, Kozawa K, Mizuno H, Yoshikawa T. A Case of Aseptic Meningitis Without Skin Rash Caused by Oka Varicella Vaccine. Pediatr Infect Dis J 2022;41:78-9. [CrossRef]
- Heusel EH, Grose C. Twelve Children with Varicella Vaccine Meningitis: Neuropathogenesis of Reactivated Live Attenuated Varicella Vaccine Virus. Viruses 2020;12(10):1078.
- https://id-info.jihs.go.jp/surveillance/nesvpd/graph/YearComparison/vzv2024/2024/20250602154218.html. accessed on June 8, 2025.
- https://www.mhlw.go.jp/topics/bcg/other/5.html. accessed on June 8, 2025.
- Gershon AA, LaRussa P, Steinberg S, Mervish N, Lo SH, Meier P. The protective effect of immunologic boosting against zoster: an analysis in leukemic children who were vaccinated against chickenpox. J Infect Dis 1996;173:450-3. [CrossRef]
- Hope-Simpson RE. THE NATURE OF HERPES ZOSTER: A LONG-TERM STUDY AND A NEW HYPOTHESIS. Proc R Soc Med 1965;58:9-20.
- Mehta SK, Cohrs RJ, Forghani B, Zerbe G, Gilden DH, Pierson DL. Stress-induced subclinical reactivation of varicella zoster virus in astronauts. J Med Virol 2004;72:174-9.
- White CJ. Clinical trials of varicella vaccine in healthy children. Infect Dis Clin North Am 1996;10:595-608. [CrossRef]
- Gaillat J, Gajdos V, Launay O, Malvy D, Demoures B, Lewden L, Pinchinat S, Derrough T, Sana C, Caulin E, et al. Does monastic life predispose to the risk of Saint Anthony’s fire (herpes zoster)? Clin Infect Dis 2011;53:405-10.
- Brisson M, Gay NJ, Edmunds WJ, Andrews NJ. Exposure to varicella boosts immunity to herpes-zoster: implications for mass vaccination against chickenpox. Vaccine 2002;20:2500-7.
- Harpaz R. Do varicella vaccination programs change the epidemiology of herpes zoster? A comprehensive review, with focus on the United States. Expert Rev Vaccines 2019;18:793-811.
- Oxman MN, Levin MJ, Johnson GR, Schmader KE, Straus SE, Gelb LD, Arbeit RD, Simberkoff MS, Gershon AA, Davis LE, et al. A vaccine to prevent herpes zoster and postherpetic neuralgia in older adults. N Engl J Med 2005;352:2271-84. [CrossRef]
- Zussman J, Young L. Zoster vaccine live for the prevention of shingles in the elderly patient. Clin Interv Aging 2008;3:241-50.
- Mbinta JF, Nguyen BP, Awuni PMA, Paynter J, Simpson CR. Post-licensure zoster vaccine effectiveness against herpes zoster and postherpetic neuralgia in older adults: a systematic review and meta-analysis. Lancet Healthy Longev 2022;3:e263-e75.
- Lal H, Cunningham AL, Godeaux O, Chlibek R, Diez-Domingo J, Hwang SJ, Levin MJ, McElhaney JE, Poder A, Puig-Barberà J, et al. Efficacy of an adjuvanted herpes zoster subunit vaccine in older adults. N Engl J Med 2015;372:2087-96.
- Cunningham AL, Lal H, Kovac M, Chlibek R, Hwang SJ, Díez-Domingo J, Godeaux O, Levin MJ, McElhaney JE, Puig-Barberà J, et al. Efficacy of the Herpes Zoster Subunit Vaccine in Adults 70 Years of Age or Older. N Engl J Med 2016;375:1019-32. [CrossRef]
- Racine É, Gilca V, Amini R, Tunis M, Ismail S, Sauvageau C. A systematic literature review of the recombinant subunit herpes zoster vaccine use in immunocompromised 18-49 year old patients. Vaccine 2020;38:6205-14.
- Huang L, Zhao T, Zhao W, Shao A, Zhao H, Ma W, Gong Y, Zeng X, Weng C, Bu L, et al. Herpes zoster mRNA vaccine induces superior vaccine immunity over licensed vaccine in mice and rhesus macaques. Emerg Microbes Infect 2024;13:2309985. [CrossRef]
- Scheifele DW, Halperin SA, Diaz-Mitoma F. Three-year follow-up of protection rates in children given varicella vaccine. Can J Infect Dis 2002;13:382-6.
- Vessey SJ, Chan CY, Kuter BJ, Kaplan KM, Waters M, Kutzler DP, Carfagno PA, Sadoff JC, Heyse JF, Matthews H, et al. Childhood vaccination against varicella: persistence of antibody, duration of protection, and vaccine efficacy. J Pediatr 2001;139:297-304. [CrossRef]
- Takayama N, Minamitani M, Takayama M. High incidence of breakthrough varicella observed in healthy Japanese children immunized with live attenuated varicella vaccine (Oka strain). Acta Paediatr Jpn 1997;39:663-8.
- Kuter B, Matthews H, Shinefield H, Black S, Dennehy P, Watson B, Reisinger K, Kim LL, Lupinacci L, Hartzel J, et al. Ten year follow-up of healthy children who received one or two injections of varicella vaccine. Pediatr Infect Dis J 2004;23:132-7. [CrossRef]
- Cohen JI. Varicella-zoster vaccine virus: evolution in action. Proc Natl Acad Sci U S A 2007;104:7-8.
- Quinlivan ML, Gershon AA, Al Bassam MM, Steinberg SP, LaRussa P, Nichols RA, Breuer J. Natural selection for rash-forming genotypes of the varicella-zoster vaccine virus detected within immunized human hosts. Proc Natl Acad Sci U S A 2007;104:208-12. [CrossRef]
Figure 1.
Outbreak Trends of Chikenpox Patients According to Fixed-Point Reporting in Tokyo, Japan. (A): Figure A shows the total number of reported chickenpox cases (bar graph; right axis) and the number of reported cases by age ((line graph; left axis) from week 1 to week 21 of 2012-2025. (B, C): The relative number of reported outbreaks by age group is shown in the line graph, with the number of reported outbreaks in 2012 as 1.
Figure 1.
Outbreak Trends of Chikenpox Patients According to Fixed-Point Reporting in Tokyo, Japan. (A): Figure A shows the total number of reported chickenpox cases (bar graph; right axis) and the number of reported cases by age ((line graph; left axis) from week 1 to week 21 of 2012-2025. (B, C): The relative number of reported outbreaks by age group is shown in the line graph, with the number of reported outbreaks in 2012 as 1.
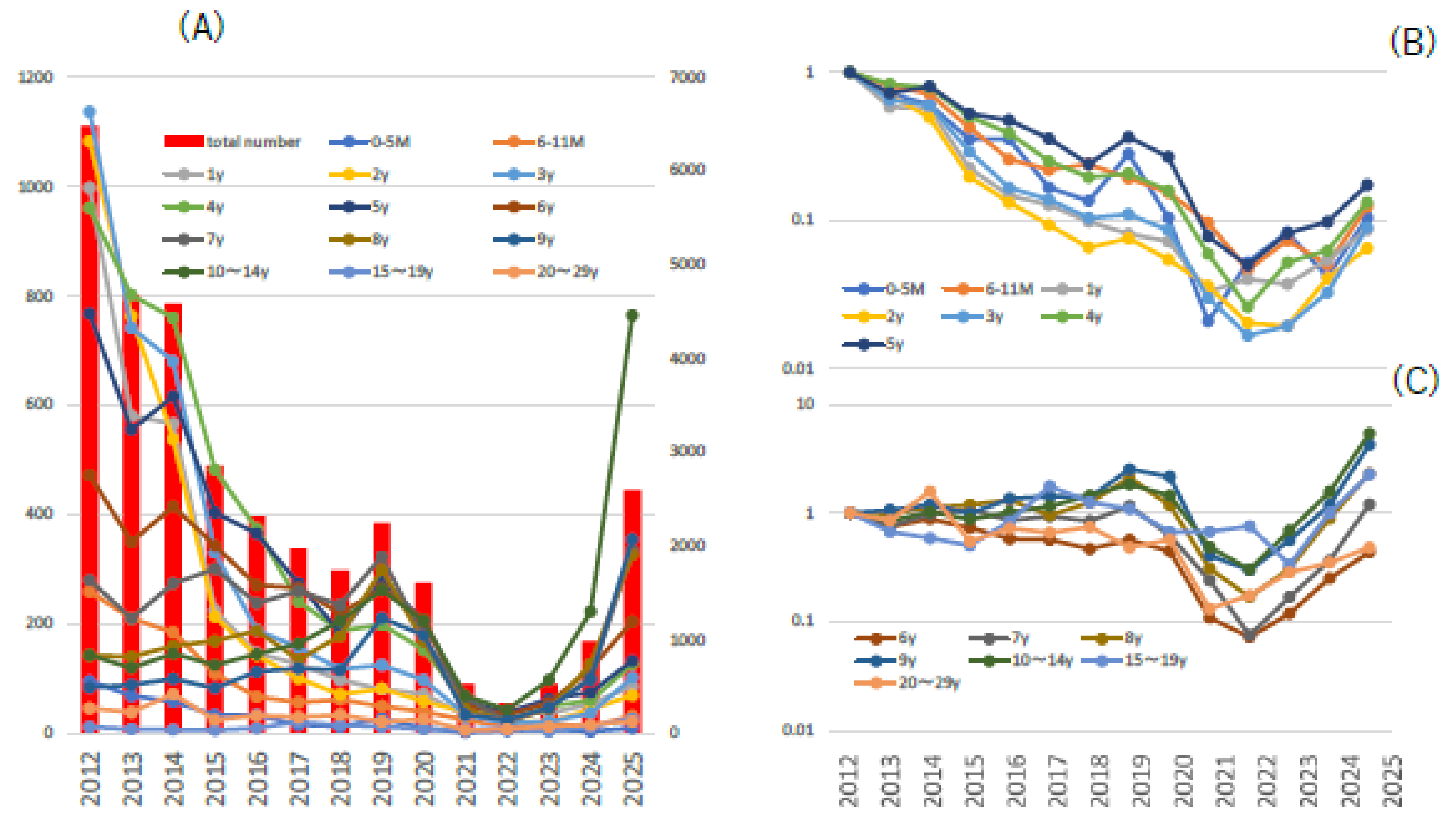
Figure 2.
Trends in anti-varicella antibody titer (EIA≥4) carriage rates by age in Japan are shown. The introduction of routine vaccination with two doses of varicella vaccine in Japan began in October 2014. Approximately 1,400 people were tested for antibody titers in each year. EIA: enzyme immunoassay.
Figure 2.
Trends in anti-varicella antibody titer (EIA≥4) carriage rates by age in Japan are shown. The introduction of routine vaccination with two doses of varicella vaccine in Japan began in October 2014. Approximately 1,400 people were tested for antibody titers in each year. EIA: enzyme immunoassay.
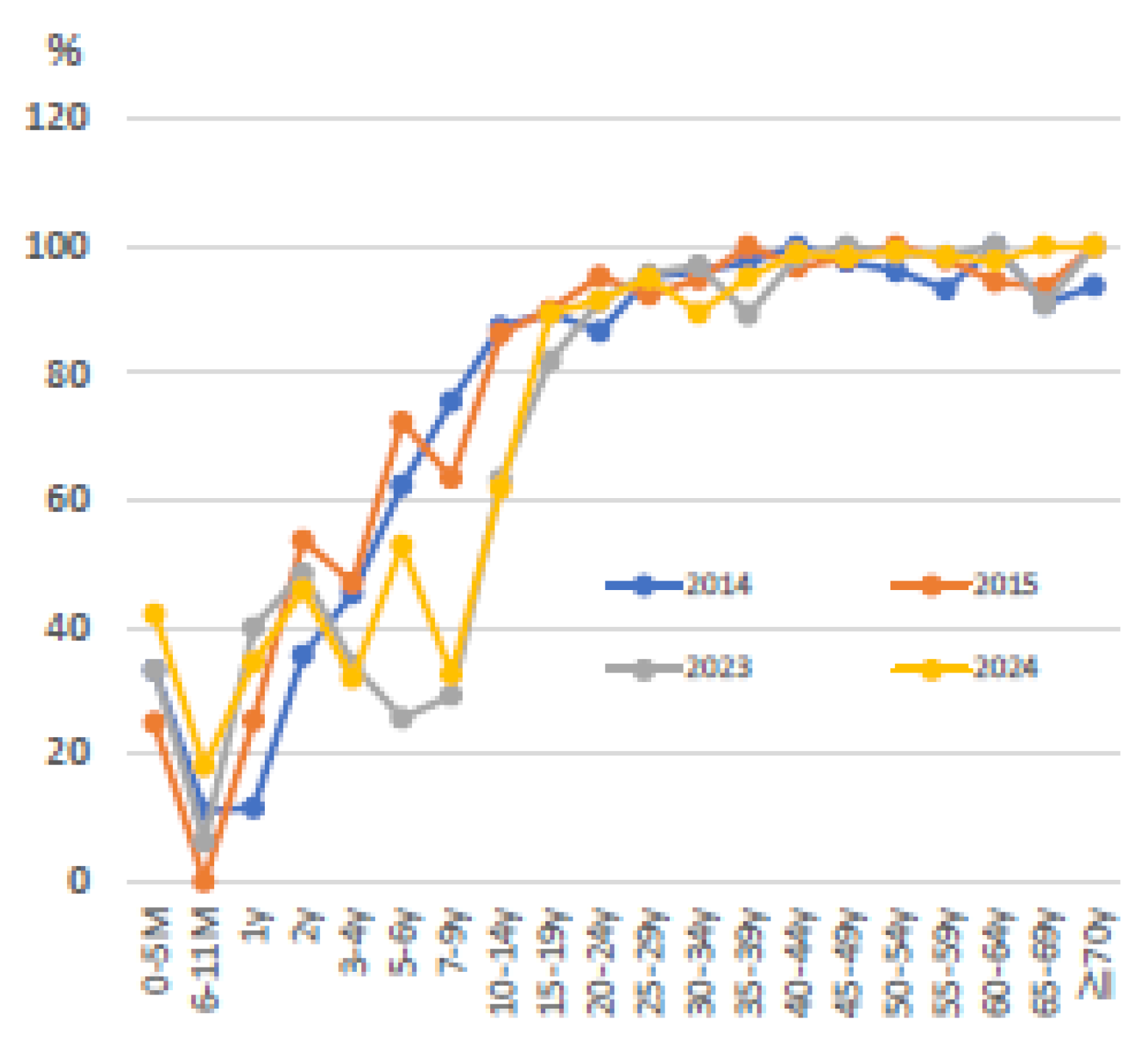

Table 1.
Pediatric cases of shingles in children with identified and confirmed vaccine strains. This table is an edited and revised version from the literature0 by the author.
Table 1.
Pediatric cases of shingles in children with identified and confirmed vaccine strains. This table is an edited and revised version from the literature0 by the author.
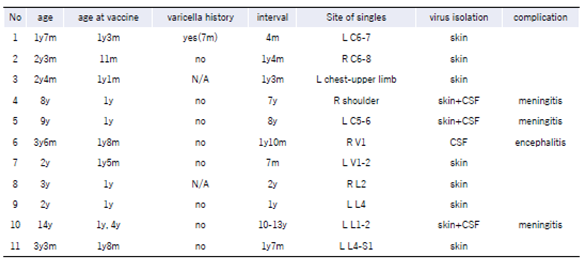 |
Table 2.
Varicella meningitis pediatric cases in which the vaccine strain was identified and confirmed. All cases of meningitis presented secondary to singles. This table is an edited and revised version from the literature0 by the author.
Table 2.
Varicella meningitis pediatric cases in which the vaccine strain was identified and confirmed. All cases of meningitis presented secondary to singles. This table is an edited and revised version from the literature0 by the author.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



