
Submitted:
22 July 2025
Posted:
22 July 2025
You are already at the latest version
Abstract
Background/Objectives: Previous research has demonstrated that the perception of self-motion, as signaled by cervical proprioception, is significantly altered during neck muscle fatigue, while no similar effects are observed when self-motion is signaled by the vestibular system. Given that in typical natural movements, both proprioceptive and vestibular signals are activated simultaneously, this study sought to investigate whether the misperception of motion persists during neck muscle fatigue when both proprioceptive and vestibular stimulation are present.
Methods: The study evaluated the gain of the perceptual responses to symmetric yaw sinusoidal head rotations on a stationary trunk during visual target localization tasks across different rotational frequencies. In addition, the final localization error of the visual target was assessed following asymmetric sinusoidal head rotations with differing half-cycle velocities. Results: The findings indicated that even with combined proprioceptive and vestibular stimulation, self-motion perceptual responses under neck muscle fatigue showed a pronounced reduction in the gain at low-frequency stimuli and a notable increase in localization error following asymmetric rotations. Notably, spatial localization error was observed to persist after asymmetric stimulation conditioning in the light. Additionally, even moderate levels of muscle fatigue were found to result in increased self-motion misperception. Conclusions: This study suggests that neck muscle fatigue can disrupt spatial orientation, even when the vestibular system is activated, so that slow movements are inaccurately perceived. This highlights the potential risks associated with neck muscle fatigue in daily activities that demand precise spatial perception.
Keywords:
Motion perception
; muscle fatigue
; neck muscle proprioception
; Balance
; vestibular
1. Introduction
Neck muscle fatigue occurs when the cervical muscles become overworked and exhausted, resulting in stiffness and a sensation of weakness or heaviness. Due to the crucial role of neck proprioception in regulating body position and motion detection [1,2,3,4,5,6,7] the effect of neck muscle fatigue may compromise beyond the motor performance of the muscles also balance [8], subjective vertical and horizontal [9], upper limb proprioception [10], posture [11,12], spatial orientation [13], and whole body pointing [14]. Recent studies [15,16] suggest that fatigue of the cervical muscles leads to altered perception of slow movements, resulting in inaccurate awareness of body position. This effect is suggested to arise from reduced proprioceptive signaling, as demonstrated in both animal [17,18] and human studies [19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36]. Specifically, ergoreceptive afferents are proposed to modulate proprioceptive input by inhibiting movement-related signals. This mechanism is further supported by findings that neck muscle vibration, which enhances neuromuscular spindle afferent discharge, can counteract the effects of fatigue [16].
Experimental evidence suggests that the erroneous self-motion perception induced by neck muscle fatigue may compromise balance and body positioning during physical activity, potentially increasing the risk of falls. However, this proprioceptive deficit can be mitigated when other sensory systems - such as the vestibular and visual systems - remain functional and can compensate.
Indeed, previous studies [15,16] have shown that the effects of neck muscle fatigue become apparent when self-motion perception is assessed through trunk rotation with a stationary head, which specifically engages neck proprioception. In contrast, when the entire body is rotated - stimulating only the vestibular system - no perceptual deficits have been observed, suggesting that vestibular-based self-motion perception remains unaffected by fatigue.
Since in everyday contexts, self-motion perception results from the integration of vestibular, visual, and proprioceptive cues, the proprioceptive deficit can be masked. Therefore, it may be important to examine how vestibular and visual signals could interfere with proprioceptive perceptual responses during fatigue. Furthermore, proprioceptive deficits have been observed only under conditions of intense fatigue, which may not reflect common real-world experiences. To better understand the practical relevance of fatigue-induced perceptual changes, we propose a study that incorporates vestibular stimulation through external perturbations, reduces fatigue intensity, and examines multisensory interactions.
To determine whether neck muscle fatigue affects motion perception in conditions involving both vestibular and proprioceptive input, we assessed self-motion perception during head rotations with a stationary trunk. Symmetric rotation trials were used to evaluate the gain of self-motion perception, while asymmetric rotations assessed self-positioning errors. These experiments were also conducted under illuminated conditions to test whether visual input mitigates the effects of fatigue. While most previous research has focused on fatigue induced by exhaustive effort, our study investigates whether even mild levels of fatigue can negatively influence motion perception, highlighting the broader implications of fatigue under less extreme conditions.
2. Method
2.1. Subjects
Ten right-handed subjects (7 men, 3 women; age range: 19–60 years; mean age: 38.35 ± 12.45) participated in the study. All the subjects had no history of neurological, visual or labyrinthine dysfunction and provided informed consent prior to their inclusion in the study. The experimental protocol was performed in accordance with the Declaration of Helsinki (1964) and was approved by the local ethical committee (University of Perugia, nr 2018-06R).
2.2. Experimental Setting
2.2.1. Neck Proprioceptive and Vestibular Stimulation
The subjects were seated on a rotating chair inside an acoustically isolated cabin. The chair rotated in the horizontal plane driven by a DC motor (Powertron, Contraves, Charlotte, NC), which was servo-controlled using an angular velocity encoder (0.01–1 Hz, 1% accuracy) (Figure 1). A head holder was used to tilt the head 30° downwards to align the horizontal semicircular canals with the rotational plane of the platform [37]. This holder maintained stationary the while rotating the trunk for stimulating neck proprioception (1° experimental setup, Figure 1 a). In a second experimental set up whole body (trunk and head) was rotated for stimulating vestibular receptors alone (2° experimental setup, Figure 1 b). In addition, a third experimental setup (3° experimental setup, Figure 1c) was employed, in which the chair was held stationary on the ground while the subject's head was rotated to stimulate both neck proprioception and the vestibular system. A DC motor (Powertron, Contraves, Charlotte, NC) was mounted on the ceiling, with its rotating axis connected to a helmet that constrained the subject's head.
The rotations activated mostly neck proprioceptors or/and vestibular receptors, but slight activation of skin receptors was also possible in all three experimental conditions. The trunk was tightly fastened to the chair through bands that kept the participant strapped to the chair. Symmetric and asymmetric sinusoidal yaw were administered in the dark (40° amplitude, 0.05. 0.1, 0.2, 0.5 Hz) starting from the center towards one side. The symmetric stimulus was used for the evaluation of the gain of perceptual responses as a relationship between the amplitudes of the tracking and the stimulus, while the asymmetrical stimulus was used for the evaluation of the perceptual responses in presence of rotations with contrast velocity in their half cycles (Pettorossi et al. 2013).
2.2.2. Asymmetric Rotation Protocol
In the case of asymmetric rotation [38], the stimulus profile was created by combining two sinusoidal half-cycles of equal amplitude (40°) but different frequencies: a fast half-cycle (Fast HC) at 0.39 Hz to one side, and a slow half-cycle (Slow HC) at 0.09 Hz to the opposite side. The fast rotation reached a peak acceleration of 120°/s² and a peak velocity of 47°/s, while the slow rotation had a peak acceleration of 7°/s² and a peak velocity of 11°/s. Both acceleration values exceeded the vestibular receptor activation thresholds (approximately 0.5–0.8°/s² for the vestibulo-ocular reflex (VOR) and 1.1–1.5°/s² for perception) (Seemungal et al., 2004).
Asymmetric rotation was implemented by introducing an 80% asymmetry into a 0.15 Hz sinusoidal waveform. This was accomplished using custom software developed in Labview (National Instruments), which controlled the DC motor of the rotating chair. Head and trunk positions were recorded using an infrared video camera mounted on the cabin ceiling. The camera tracked two pairs of reflective infrared markers: one pair placed on the head (at the bregma and 3 cm posterior to it) and the other on the shoulders (at the left and right acromion).
2.2.3. Self-Motion Perception Recordings
Self-motion perception was assessed using a psychophysical tracking procedure [39,40,41,43]. Prior to rotation, subjects fixated on a visual target—a light spot with a 1 cm diameter—projected onto the cabin wall 1.5 m in front of them. The target was extinguished immediately before rotation onset and re-illuminated at the end of the stimulus. During the rotation, performed in complete darkness, participants were instructed to continue imagining and mentally tracking the fixed target with their eyes closed.
The tracking task was executed using a pointer mounted on a support fixed to the rotating platform, located 25 cm from the subject's body axis and 100 cm above the platform surface. The pointer was connected to a precision potentiometer to measure movements.
We evaluated perceptual responses during both symmetric and asymmetric sinusoidal rotations. The symmetric stimulus was used to determine perceptual gain, calculated as the ratio between the amplitude of the pointer tracking and that of the chair rotation. In contrast, the asymmetric stimulus was used to assess adaptive responses to motion stimuli with velocity contrasts and their associated fatigue effects.
During asymmetric stimulation, we analyzed the final position error (FPE), defined as the discrepancy between the perceived and actual target location after four full cycles. This error arose from differential perception of fast and slow half-cycles: participants consistently perceived Fast HCs more vividly than Slow HCs [38]. Consequently, the perceived final target position was biased in the direction of the Slow HC. The magnitude of the error reflected the sensory system’s differential responsiveness to motion velocity, as well as the influence of adaptive mechanisms that enhance responses to faster stimuli while diminishing responses to slower ones.
2.2.4. Neck Extensor Muscle Fatigue
Neck extensor muscle fatigue was induced through isometric muscle contraction. Participants were positioned prone on a table, with their heads protruding and extending against gravity. An additional load, equivalent to 40% of the maximal voluntary contraction (MVC) of the neck extensor muscles, was applied to the head. The MVC was determined by measuring the maximum extension force exerted against a strain gauge during head extension. Participants maintained head extension at this load until voluntary exhaustion, as indicated by their reports. The time to exhaustion generally ranged from 8 to 10 minutes. Following the fatigue protocol, participants performed a neck extension stretch by flexing the head downward to mitigate potential thixotropic effects on the musculature and neuromuscular spindles [23].
Three minutes after the conclusion of the fatigue procedure, the individuals were tested to evaluate the FPE. In separate experiments, to examine the effect of a minor level of fatigue on FPE, tests were performed at 1, 3, and 6 minutes after the fatigue protocol.
Fatigue was assessed by recording the EMG activity of the posterior neck extensor muscles and by calculating a fatigue index using the following formula:
Fatigue index = (Amplitude ratio-AMP) / (Mean frequency-MF ratio), where: AMP ratio = AMP during the 10-second period every minute following load application/AMP during the 10-second baseline period (measure immediately after load application); MF ratio = MF during the 10-second period every minutes following load application/MF during the 10-second baseline period.
2.2.5. Experimental Protocol
The subjects were blinded to the protocol. They were told that they would be experiencing rotations. Prior to the beginning of the experiment, the subjects were instructed to use the earth-fixed visual target placed in front of them as a reference point. Before the test, they fixed their gaze on the visible target for at least 1 min. We tested the perceptual responses before neck muscle fatiguing and 5 min after the fatiguing procedure in the following separate ( at least 1 day interval ) randomized experimental conditions: (i) 4 cycles of symmetric and 4 cycles of asymmetric sinusoidal rotations of the trunk with the stationary head starting with the head and trunk aligned with the earth-fixed target for neck proprioception; (ii) 4 cycles of symmetric and 4 cycles of asymmetric sinusoidal whole body rotations starting with the head and trunk aligned in a straight position for vestibular stimulation; (iii) 4 cycles of symmetric and 4 cycles of asymmetric sinusoidal head rotation with the stationary trunk starting with the head and trunk aligned in a straight position and the visual target. All these tests were carried out before and after maximal fatigue (up to exhaustion) of the neck muscles, while other tests were carried out after partial fatigue (1, 3, 6 min). In the end, sequences of asymmetric stimulations of 4 cycles with buoyancy were carried out to test proprioceptive and vestibular stimulation together (3rd setup) both in light and in the dark. The same sequence was then repeated 3 minutes later in the dark to see if the light was able to cancel out the conditioning effects of the asymmetrical stimulations.
2.2.6. Acquisition and Analysis of the Pointer Data
The potentiometer and the chair signals were fed to the computer (sampling rate 50 Hz) for display and storage. The data were analyzed offline to evaluate the gain (amplitude of the perceptual responses/amplitude of the stimulus) for showing the efficacy of the proprioceptive and vestibular system in the self-motion perception and the tracking final position error (FPE) for showing the efficacy of these sensory signals in response to asymmetric rotation, during contrasting velocity stimulation. The responses were statistically analyzed using generalized mixed model analysis (GLM). The analysis included perceptual responses as the dependent variables; groups of tests before and under fatigue, the frequency of stimulation and interactions as the fixed effects of main interest; and a random effect for the repeated measures. This analysis allowed us to establish the statistical significance of the perceptual difference observed before and during neck muscle fatigue. Statistical post hoc analysis was performed with Bonferroni post hoc correction for multiple comparisons. The eta-square (Ƞ) was computed to assess effect size. The level of significance was set at p < 0.05 for both the GLM values and post hoc comparisons. Prior to GLM, the Shapiro‒Wilk test was used to assess normality, and Levene’s test was used to assess homogeneity of variance.
3. Results
Effect of neck muscle fatigue on self-motion perceptual gain in response to head and trunk symmetric sinusoidal rotation.
Figure 2.
Self-motion perception gain in response to symmetric sinusoidal rotation of the trunk on stationary head (A), whole body (B) and head on stationary trunk (C). Means and stand and deviations (SDs) before (filled symbol) and after (open symbol) fatigue induction are reported. The statistically significant differences of values between the two conditions are indicated with *p <0.05,**p<0.01, ***p<0.001.
Figure 2.
Self-motion perception gain in response to symmetric sinusoidal rotation of the trunk on stationary head (A), whole body (B) and head on stationary trunk (C). Means and stand and deviations (SDs) before (filled symbol) and after (open symbol) fatigue induction are reported. The statistically significant differences of values between the two conditions are indicated with *p <0.05,**p<0.01, ***p<0.001.
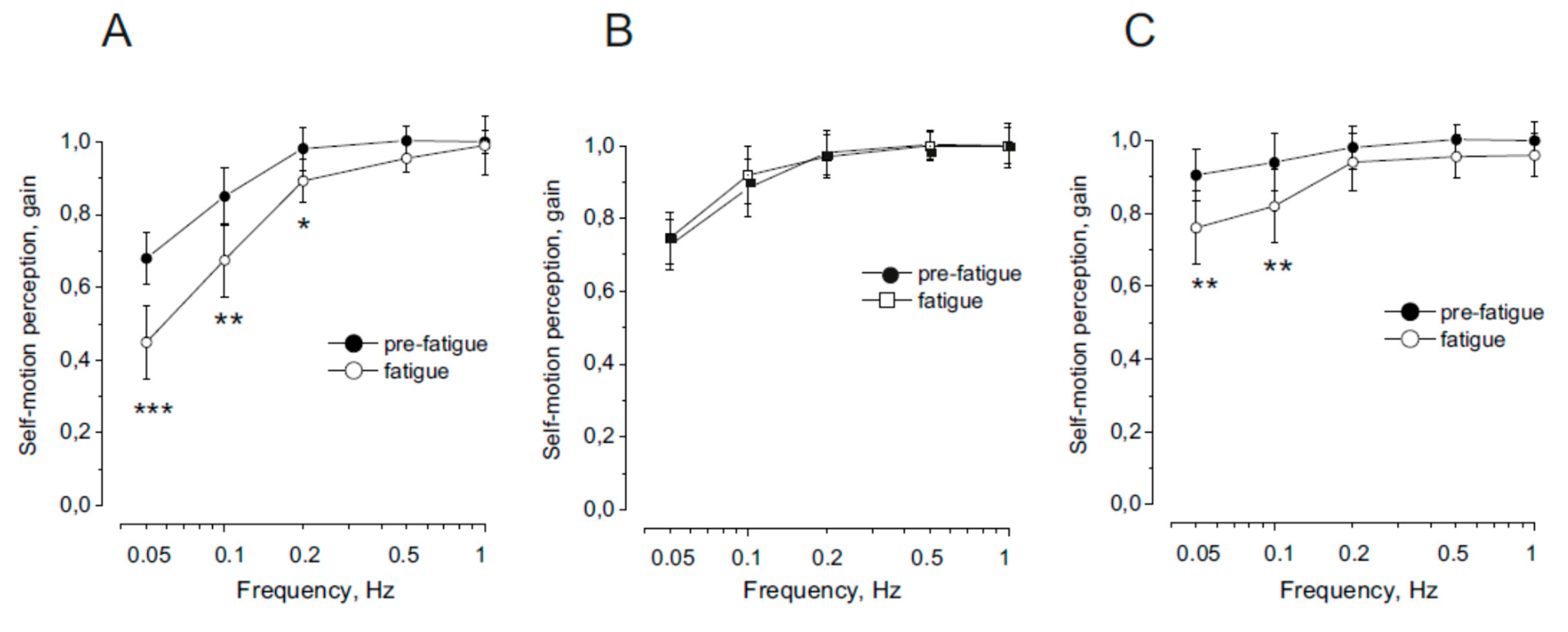
The effect of neck muscle fatigue on the gain of the self-motion perception in response to trunk rotation on stationary head before and after fatiguing neck muscle procedure is reported in Figure 1 A. In normal condition, as previously shown [15,45,46] the signal from the proprioception of the neck allowed to obtain an excellent response to high rotation frequencies, while to rotations with lower frequency the response tends to be reduced. Fatigue reduced the amplitude of the motion perception during rotation at 0.05 and 0.1 Hz. At higher frequency of rotation no significant change was observed.
Compared with the pre-fatigue condition, the neck muscle fatigue condition significantly reduced the gain (F (1,36) = 29.15, p < 0.001, Ƞ = 0.88), but the difference in gain was a function of rotation frequency. In fact, the gain decreased significantly at lower oscillation frequencies (0.05 and 0.1 Hz, p < 0.001; 0.2 Hz, p < 0.05) but remained unchanged at higher frequencies (0.5–1 Hz, p = 0.36–0.55).
Figure 1 b shows the effect of neck muscle fatigue on the gain of the self-motion perception in response to whole body rotation before and after fatiguing neck muscle procedure. In normal condition the gain profile was similar to that of the proprioception [15,45,46]. Fatigue did not affect the motion perception, as the amplitude before and after were not significantly different.
For all rotation frequencies, no difference in gain was noted between pre-fatigue and fatigue conditions (F (1,36) = 0.52, p = 0.76, Ƞ = 0.79).
Figure 1c shows the effect of neck muscle fatigue on the gain of the self-motion perception in response to head rotation on stationary trunk before and after fatiguing neck muscle procedure. In normal condition, it has been found, as previously reported [45,46], that when the stimulus combines cervical proprioception with vestibular signals the gain of perceptual response remained higher at the slow as well as higher frequencies of rotation. However, after fatigue procedure the gain significantly decreased (F (1,36) = 12.15, p < 0.01, Ƞ = 0.88). The difference in gain was a function of rotation frequency In fact, the gain decreased significantly at lower oscillation frequencies (0.05-0.1 Hz, p<001), but remained unchanged at higher frequencies (0.2-1 Hz, p = 0.27–0.45).
Effect of neck muscle fatigue on self-motion perceptual final position in responses to head and trunk asymmetric sinusoidal rotation
Figure 3.
Final position error (FPE) in response to asymmetric rotation before and after fatiguing procedure. In C the trunk was rotated in the stationary head for cervical proprioceptive stimulation, in V the whole body was rotated for stimulating the vestibular system and in V+C the head was rotated in the stationary trunk for stimulating both the cervical and vestibular receptors. Note that in C and V+C condition the fatigue increases significantly the FPE (***p<0.001,**p<0.01), while not significant change (ns) was observed in V condition.
Figure 3.
Final position error (FPE) in response to asymmetric rotation before and after fatiguing procedure. In C the trunk was rotated in the stationary head for cervical proprioceptive stimulation, in V the whole body was rotated for stimulating the vestibular system and in V+C the head was rotated in the stationary trunk for stimulating both the cervical and vestibular receptors. Note that in C and V+C condition the fatigue increases significantly the FPE (***p<0.001,**p<0.01), while not significant change (ns) was observed in V condition.
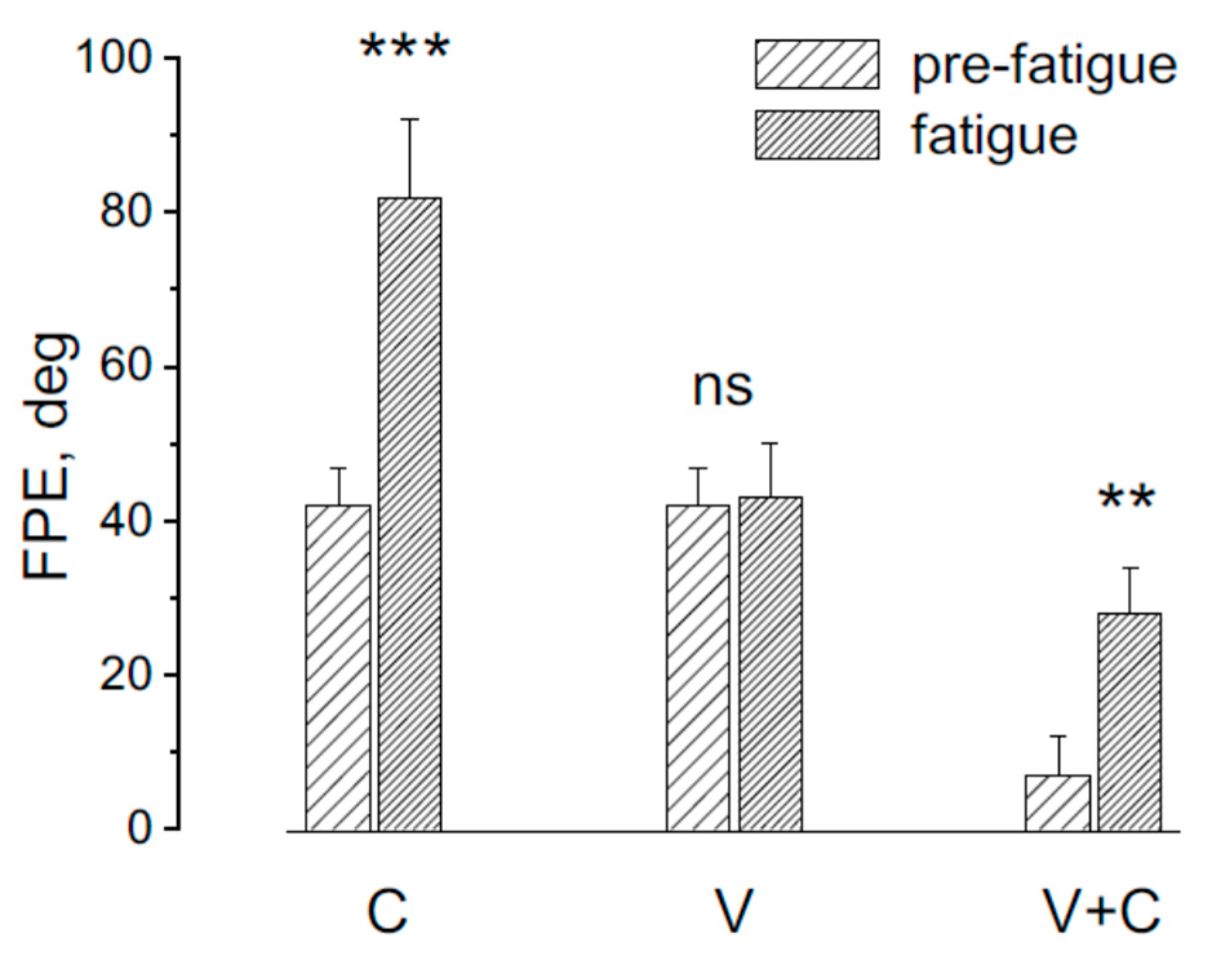
The effect of the asymmetric rotation in the FPE was examined before and after fatiguing procedure in three different conditions: 1) rotation activating neck proprioception alone (trunk rotation on stationary head; 2) rotation activating vestibular receptors (whole body rotation); 3) rotation activating both neck proprioceptors and vestibular receptors (Head rotation on stationary trunk) (Figure 4). After fatiguing procedure the FPE significantly changed (F(2,18) = 13.15, p < 0.01, Ƞ = 0.78 ), specifically in response to trunk rotation (p<0.001) and head rotation (p<0.001), while remained unchanged in response to whole body rotation (P = 0.21).
Effect of light in the induction of FPE in response to asymmetric rotation.
Cycles of asymmetric head rotation on a stationary trunk were performed under both dark and light conditions. The first three cycles served as a conditioning procedure and were conducted either in complete darkness or in the light, while the fourth cycle functioned as a testing cycle performed exclusively in the dark. This sequence of asymmetric rotations followed a neck muscle fatigue protocol.
In the dark condition, the error in position representation increased progressively across cycles. The final position error (FPE) during the first cycle was 3 ± 5°, rising to 6 ± 7° in the second cycle, 13 ± 7° in the third, and 17 ± 5° in the fourth cycle. Overall, the FPE at the end of all cycles was 39 ± 9°. This gradual escalation was due to the compounding of previous errors and the adaptation to the contrast velocity of the asymmetric rotations, likely resulting from the conditioning effects of prior cycles [15,38].
Consistent with previous findings, adaptation appeared to diminish the perception of slower motion over time. In contrast, during the light condition, observed error in motion perception was negligible across the first three cycles (error ranging from 0 to 1°). However, following the final cycle in the dark, the FPE increased to 15 ± 9°.
Statistical analysis revealed significant differences in FPE across the four cycles (F(1,27) = 19.11, p < 0.01, η² = 0.82). A comparison of the positional errors induced by the last (fourth) cycle in light versus dark conditions showed no significant difference (p = 0.43), suggesting that the conditioning effects induced by previous asymmetric rotations in darkness also influenced head perception during subsequent light cycles, despite the lack of observable error in the latter.
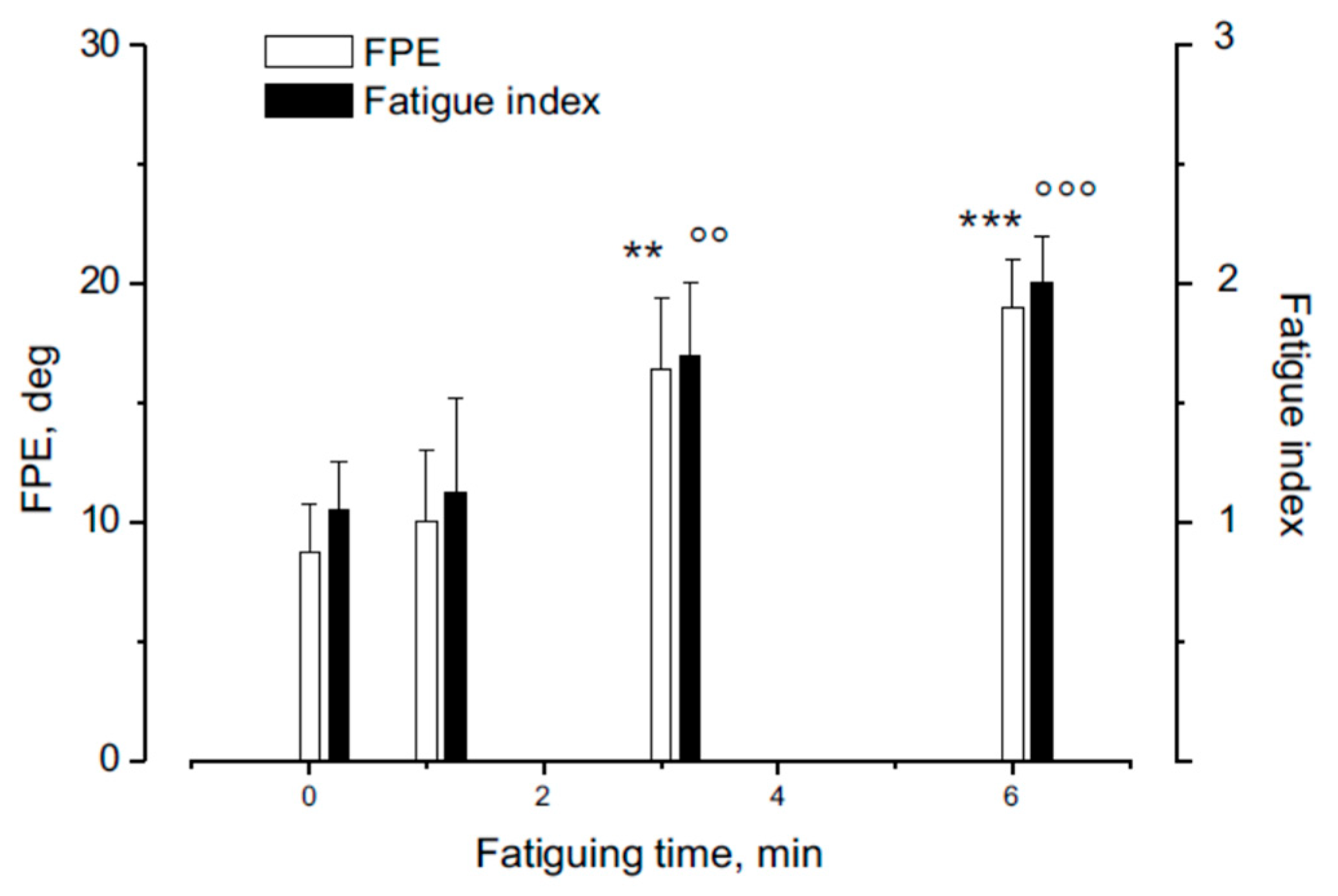
The FPE induced by head rotation on a stationary trunk was examined after varying levels of fatigue across different experimental sessions. The assessments were conducted before and at 1, 3, 6 minutes following the fatiguing procedure. Results showed that FPE significantly increased after fatigue of different durations (see Figure 5) (F(3,36) = 19.32, p < 0.001, Ƞ = 0.88). Specifically, FPE values and the fatigue index was not significantly increased after 1 min of fatigue while both showed a significant enhancement after 3 and 6 minutes of fatiguing procedure (see Figure 5).
Effect of different level of fatigue in the FPE effect.
4. Discussion
Previous [15,16] and current findings indicate that neck muscle fatigue impairs the ability to accurately signal trunk displacement relative to the head, particularly during slower body movements. In response to a sequence of combining fast and slow rotation, the final sense of position becomes almost double than that induced normally. These effects are likely due to the influences of fatigue-related afferent signals on the proprioceptive system, reducing the transmission of movement information to the central nervous system [19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36]. Supporting this hypothesis, it has been shown that vibration—capable of activating muscle spindles [47,48] —can reduce or eliminate the negative effects of muscle fatigue on movement perception [16]. At the same time, the detrimental effect on self-motion perception does not occur when vestibular signals without proprioceptive activation are involved in detecting self-motion. The finding that vestibular information remains unchanged by fatigue of neck muscle might suggest that motion perception deficit may not emerge when, as in most natural condition, proprioceptive and vestibular system are both activated, in case that vestibular signal can supply and compensate for the reduction of the proprioceptive information. This study provided an answer to this problem demonstrating that even in the presence of combined vestibular and neck proprioceptive stimulation the perceptual responses to symmetric rotation showed a decrease during slower movements after inducing neck muscle fatigue and the final position error in response to asymmetric stimulation that was almost null in normal condition, occurs and begin remarkable. This can be explained by the fact that fatigue reduces sensitivity to slow movements due to a weakened proprioceptive signal [15] This reduced the gain to slow rotation that mostly depends on neck proprioceptive stimulation and induced a different responsivity to fast and slow rotation. This likely causes the occurrence of large FPE.
Furthermore, the effect of fatigue in the position perception is induced even when rotation was performed in the light, in more common natural condition. In fact, while the effects of asymmetric stimuli are normally fully abolished by visual input, experiments in which head-on-trunk rotations were performed first in the light and then in darkness show that, after fatigue, the central nervous system retains the effects of asymmetric stimulation. This is demonstrated by the fact that the effect of the asymmetric rotation under neck muscle fatigue is not cancelled in the central nervous system, but persists, so that the positional error shows a further increased when the rotation is repeated in the dark, as the adaptive effect induced by asymmetric rotation is built up during rotation in the light, similarly to what happens in the dark. This effect has been observed also during asymmetric rotation activating the vestibular receptors alone [42].
However, a possible limitation concerns the generalization of these effects to real-life scenarios of neck muscle fatigue. The previous study demonstrating these effects induced fatigue up to the point of exhaustion—a condition that rarely occurs in daily life. The present study addresses this issue by showing that perceptual errors also occur under less intense fatigue, even although the magnitude of the errors is smaller.
In conclusion, this study demonstrates that neck muscle fatigue has a distinctly detrimental impact on self-motion and position perception. This impairment persists even in the presence of intact input from other sensory systems, thereby posing a risk to postural stability, balance, and motor performance. As a result, individuals are more likely to experience disorientation and reduced motor control.
Neck muscle fatigue is a common occurrence, as it can be induced by various factors such as bad posture, all day computer use, long drive, employees without ergonomic workstation / and athletic activities. It is particularly relevant in scenarios requiring sustained neck extension to maintain upward gaze (e.g., rock climbers), support of external head-borne loads (e.g., helicopter pilots wearing heavy helmets), or maintenance of rotated or asymmetric head positions (e.g., athletes engaged in asymmetrical sports). In all these situations, once a critical threshold of muscle fatigue is reached, individuals may experience significant disorientation and a diminished capacity to maintain balance.
References
- Schweigart G., Heimbrand S., Mergner T., Becker W Role of neck input for the perception of horizontal head and trunk rotation in patients with loss of vestibular function. Exp. Brain. Res. 1993, 95, 533–546.
- Schweigart G., Chien R.D., Mergner T. Neck proprioception compensates for age-related deterioration of vestibular self-motion perception. Exp. Brain Res. 2002, 147, 89–97. [CrossRef]
- Bottini G., Karnath H.O., Vallar G., Sterzi R., Frith C.D., Frackowiak R.S., Paulesu E. Cerebral representations for egocentric space: functional anatomical evidence from caloric vestibular stimulation and neck vibration. Brain 2001, 124, 1182–1196. [CrossRef]
- Bove M., Courtine G., Schieppati M. Neck muscle vibration and spatial orientation during stepping in place in humans. J Neurophysiol. 2002, 88, 2232–2241.
- Pettorossi V.E., Schieppati M. Neck proprioception shapes body orientation and perception of motion. Front. Hum. Neurosci. 2014, 4(8), 895. [CrossRef]
- Cullen K.E. The neural encoding of self-generated and externally applied movement: implications for the perception of self-motion and spatial memory. Front. Integr. Neurosci. 2014, 7, 108. [CrossRef]
- Jamal K., Leplaideur S., Leblanche F., Moulinet Raillon A., Honoré T., Bonan I. The effects of neck muscle vibration on postural orientation and spatial perception: a systematic review. Neuro physiol. Clin. 2020, 50, 227–267. [CrossRef]
- Gosselin G., Rassoulian H., Brown I. Effects of neck extensor muscles fatigue on balance. Clin. Biomech. 2004, 19, 473–479. [CrossRef]
- Gosselin G., Fagan M.J. Effects of cervical muscle fatigue on the perception of the subjective vertical and horizontal. Springerplus 2014, 8(3),78. [CrossRef]
- Zabihhosseinian M., Holmes M.W., Murphy B Neck muscle fatigue alters upper limb proprioception. Exp. Brain Res. 2015, 233, 1663–1675. [CrossRef]
- Schieppati M., Nardone A., Schmid M. Neck muscle fatigue affects postural control in man. Neurosci. 2003, 121, 277–285.
- Baccini, M., Risaliti, I., Rinaldi, L.A., Paci, M. Head position and neck muscle fatigue: effects on postural stability. Gait Posture 2006, 24, S9–S10. [CrossRef]
- Schmid M., Schieppati M. Neck muscle fatigue and spatial orientation during stepping in place in humans. J. Appl. Physiol.2005, 99, 141–153. [CrossRef]
- Schmid M., Schieppati M., Pozzo T. Effect of fatigue on the precision of a whole-body pointing task. Neurosci. 2006, 139, 909–920.
- Botti, F.M., Guardabassi, M., Ferraresi, A., Faralli, M., Filippi, G.M., Marcelli, V., Occhigrossi, C., Pettorossi, V.E. Neck muscle fatigue disrupts self-motion perception. 2025, Exp. Brain Res. 233, 1663–1675. [CrossRef]
- Guardabassi M., Botti F.M., Rodio A., Fattorini L.,Filippi G.M., Ferraresi A., Occhigrossi C., Pettorossi V.E. Prolonged neck proprioceptive vibratory stimulation prevents the self-motion misperception induced by neck muscle fatigue: immediate and sustained effects. Exp. Brain. Res. 2025, 243, 162. [CrossRef]
- Brunetti O., Della Torre G., Lucchi M.L., Chiocchetti R., Bortolami R., Pettorossi V.E. Inhibition of muscle spindle afferent activity during masseter muscle fatigue in the rat. Exp. Brain Res. 2003, 152, 251 262. [CrossRef]
- Pettorossi V.E., Della Torre G., Bortolami R., Brunetti O. The role of capsaicin-sensitive muscle afferents in fatigue-induced modulation of the monosynaptic reflex in the rat. J. Physiol. 2004, 515, 599–607. [CrossRef]
- Skinner H.B., Wyatt M.P., Hodgdon J.A. Effects of fatigue on joint position sense of the knee. J. Orthop. Res. 1986, 4, 112–118.
- Shape M.H., Miles T.S. Position sense at the elbow after fatiguing contractions. Exp. Brain Res. 1993, 94, 179–182.
- Garland S.J., Kaufman M.P. Role of muscle afferents in the inhibition of motoneurons during fatigue. In:. Fatigue neural and muscular mechanisms. Adv. in Exp. Med. and Biol.; Gandevia S.C., Enoka R.M., McComas A.J., Stuart D.G., Thomas C.K. Eds.; Plenum Press, New York and London, 1995,Volume 384, pp 271–278.
- Gandevia S.C., Enoka R.M., McComas A.J., Stuart D.G., Thomas C.K. Fatigue neural and muscular mechanisms. Adv. in Exp. Med. and Biol., vol 384. Plenum Press, New York and London, 1995.
- Hagbarth K.E., Macefield V.G. The fusimotor system. Its role in fatigue. In:. Fatigue neural and muscular mechanisms. Adv. in Exp. Med. and Biol.; Gandevia S.C., Enoka R.M., McComas A.J., Stuart D.G., Thomas C.K. Eds.; Plenum Press, New York and London, 1995,Volume 384, pp 259–270 pp 259–270.
- Lattanzio P.J., Petrella R.J., Sproule J.R., Fowler P.J. Effects of fatigue on knee proprioception. Clin. J. Sports Med. 1997, 7, 22–27.
- Pedersen J.M., Ljubislavlevic M., Bergenheim M., Johansson H. Alterations in information transmission in ensemble of primary muscle spindle afferents after muscle fatigue in heteronymous muscle. Neurosci. 1998, 84, 953–959.
- Pedersen J.M., Lonn J., Hellstrom F., Djupsjobacka M., Johansson H. Localised muscle fatigue decreases the acuity of the movement sense in the human shoulder. Med. Sci. Sports Exerc. 1999, 31, 1047–1052.
- Forestier N., Teasdale N., Nougier V. Alteration of the position sense at the ankle induced by muscular fatigue in humans. Med. Sci. Sports Exerc. 2002, 34, 117–122. [CrossRef]
- Hunter S.K., Duchateau J., Enoka R.M. Muscle fatigue and the mechanisms of task failure. Exerc. Sport Sci. Rev. 2004, 32, 44–49.
- Miura K., Ishibashi Y., Tsuda E., Okamura Y., Otsuka H., Toh S. The effect of local and general fatigue on knee proprioception. J. Arthrosc. Relat. Surg. 2004, 20, 414–418.
- Proske U., Gandevia S.C. Kinaesthetic Senses. J. Physiol. 2009, 587, 4139–4146.
- Proske U., Gandevia S.C. The proprioceptive senses: their roles in signaling body shape, body position and movement, and muscular force. Physiol. Rev. 2012, 92(4), 1651–1697.
- Abd-Elfattah, H.M., Abdelazeim, F.H., Elshennawy, S. Physical and cognitive consequences of fatigue: a review. J. Adv. Res. 2015, 6, 351–358.
- Proske U. Exercise, fatigue and proprioception: a retrospective. Exp. Brain. Res. 2019, 237, 2447–2459. [CrossRef]
- Proske U., Allen T.J. The neural basis of the senses of effort, force and heaviness. Exp. Brain Res. 2019, 237, 589–599.
- Karagiannopoulos C., Watson J., Kahan S., Lawler D. The effect of muscle fatigue on wrist joint position sense in healthy adults. J. Hand Ther. 2020, 33, 329–338.
- Sayyadi P., Minoonejad H., Seifi F., Sheikhhoseini R., Arghadeh R. The effectiveness of fatigue on repositioning sense of lower extremities: systematic review and meta-analysis. BMC Sports Sci. Med. Rehabil. 2024, 16, 35. [CrossRef]
- Blank, R.H., Curthoys, I.S., Markham, C.H. Planar relationships of the semicircular canals in man. Acta Otolaryngol. 1975, 80:185–196. [CrossRef]
- Pettorossi V.E., Panichi R., Botti F.M., Kyriakareli A., Ferraresi A., Faralli M., Schieppati M., Bronstein A.M. Prolonged asymmetric vestibular stimulation induces opposite, long-term effects on self-motion perception and ocular responses. J. Physiol. 2013, 591, 1907 1920. [CrossRef]
- Seemungal B.M., Gunaratne I.A., Fleming I.O., Gresty M.A., Bronstein A.M. Perceptual and nystagmic thresholds of vestibular function in yaw. J. Vestib. Res. 2004, 14, 461–466.
- Panichi R., Botti F.M., Ferraresi A., Faralli M., Kyriakareli A., Schieppati M., Pettorossi V.E. Self-motion perception and vestibulo ocular reflex during whole body yaw rotation in standing subjects: the role of head position and neck proprioception. Hum. Mov. Sci. 2011, 30, 314–332. [CrossRef]
- Pettorossi V.E., Panichi R., Botti F.M., Biscarini A., Filippi G.M., Schieppati M. Long-lasting effects of neck muscle vibration and contraction on self-motion perception of vestibular origin. Clin. Neurophysiol. 2015, 126, 1886–1900. [CrossRef]
- Pettorossi V.E., Occhigrossi C., Panichi R., Botti F.M., Ferraresi A., Ricci G., Faralli M. Induction and Cancellation of Self-Motion Misperception by Asymmetric Rotation in the Light. Audiol. Res. 2023, 13, 196-206. [CrossRef]
- Siegle J.H., Campos J.L., Mohler B.J., Loomis J.M., Bülthoff H.H. Measurement of instantaneous perceived self-motion using continuous pointing. Exp. Brain Res. 2009, 195, 429–444. [CrossRef]
- Merletti R., Lo Conte L.R. Surface EMG signal processing during isometric contractions. J. Electromyogr. Kinesiol. 1997, 7, 241-250.
- Mergner T., Siebold C., Schweigart G., Becker W. Human perception of horizontal trunk and head rotation in space during vestibular and neck stimulation. Exp. Brain Res. 1991, 85, 389–404. [CrossRef]
- Mergner T., Rosemeier T. Interaction of vestibular, somatosensory and visual signals for postural control and motion perception under terrestrial and microgravity conditions—a conceptual model. Brain Res. Rev. 1998, 28, 118–135. [CrossRef]
- Filippi G.M., Rodio A., Fattorini L., Faralli M., Ricci G., Pettorossi V.E. Plastic changes induced by muscle focal vibration: a possible mechanism for long-term motor improvements. Front. Neurosci. [CrossRef]
- Souron R., Besson T., Millet G., Lapole T. Acute and chronic neuromuscular adaptations to local vibration training. Eur. J. Appl. Physiol. 2017, 117, 1939–1964. [CrossRef]
Figure 1.
Schematic drawing of the experimental set. The individual was placed in the dark cabin and rotated sinusoidally on the chair. The velocity of rotation was symmetric or asymmetric (fast rotation of half cycle followed by slow rotation half cycle). The individual tracked the remembered visual target (T) with the pointer (P). The head holder (H) was fixed to the chair for stimulating the vestibular receptors during chair rotation, to the floor for stimulating the neck proprioceptive system during rotation. The chair was maintained fixed and the head was rotated by a motor (M) placed above the head for stimulating both the vestibular and the neck proprioceptive receptors.
Figure 1.
Schematic drawing of the experimental set. The individual was placed in the dark cabin and rotated sinusoidally on the chair. The velocity of rotation was symmetric or asymmetric (fast rotation of half cycle followed by slow rotation half cycle). The individual tracked the remembered visual target (T) with the pointer (P). The head holder (H) was fixed to the chair for stimulating the vestibular receptors during chair rotation, to the floor for stimulating the neck proprioceptive system during rotation. The chair was maintained fixed and the head was rotated by a motor (M) placed above the head for stimulating both the vestibular and the neck proprioceptive receptors.
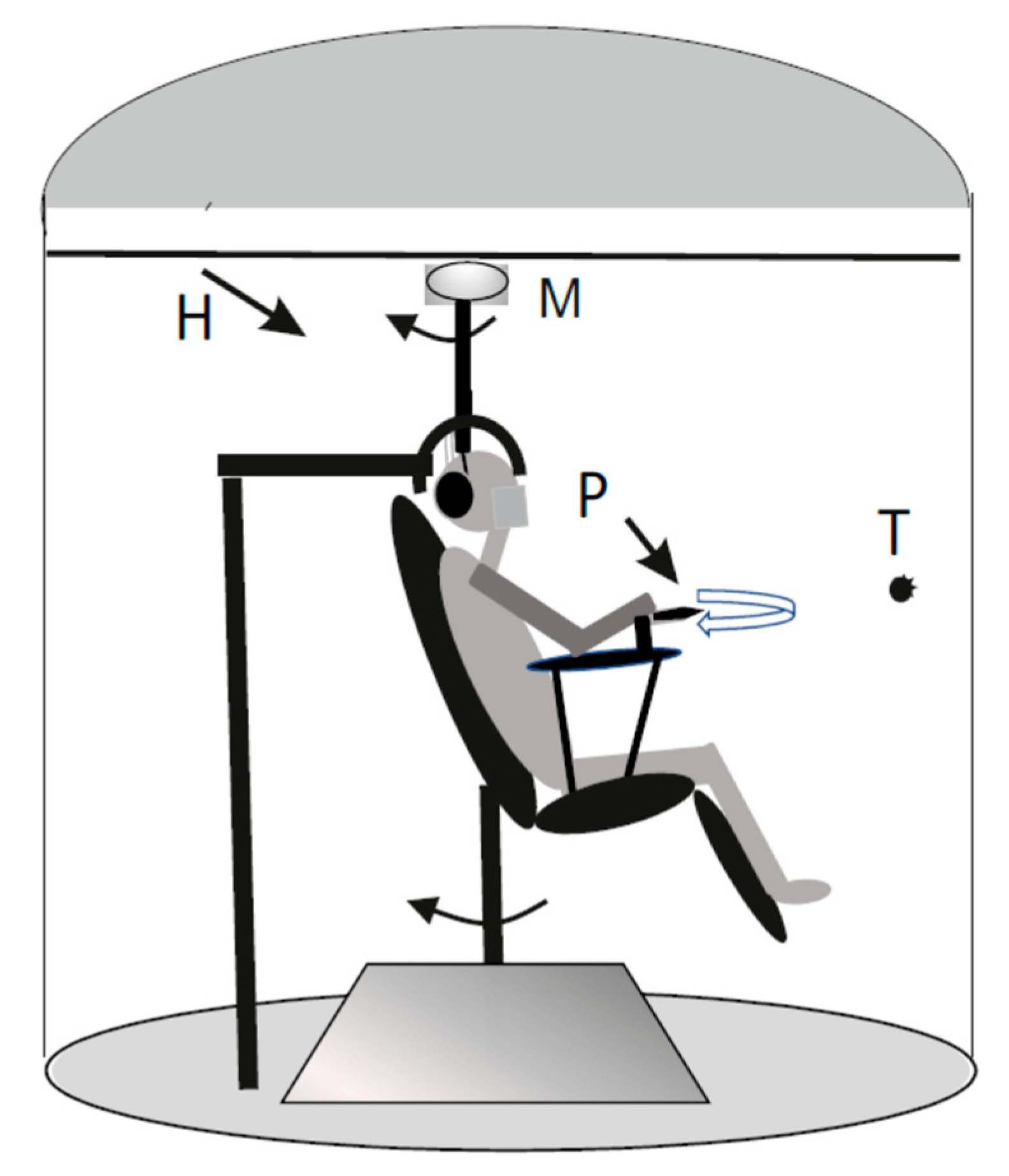
Figure 4.
Effect of the 4th cycle FPE following conditioning induced by three cycles of asymmetrical rotation in light (filled symbols and black bars) and dark (open symbols and white bars). Data represent the mean ± SD of FPE measured at the end of the cycles. Notably, the final cycle after light conditioning results in an increase in FPE (indicated by the bracket), like that observed following dark conditioning. No significant difference was found between the 4th cycle after light and dark conditioning (ns).
Figure 4.
Effect of the 4th cycle FPE following conditioning induced by three cycles of asymmetrical rotation in light (filled symbols and black bars) and dark (open symbols and white bars). Data represent the mean ± SD of FPE measured at the end of the cycles. Notably, the final cycle after light conditioning results in an increase in FPE (indicated by the bracket), like that observed following dark conditioning. No significant difference was found between the 4th cycle after light and dark conditioning (ns).
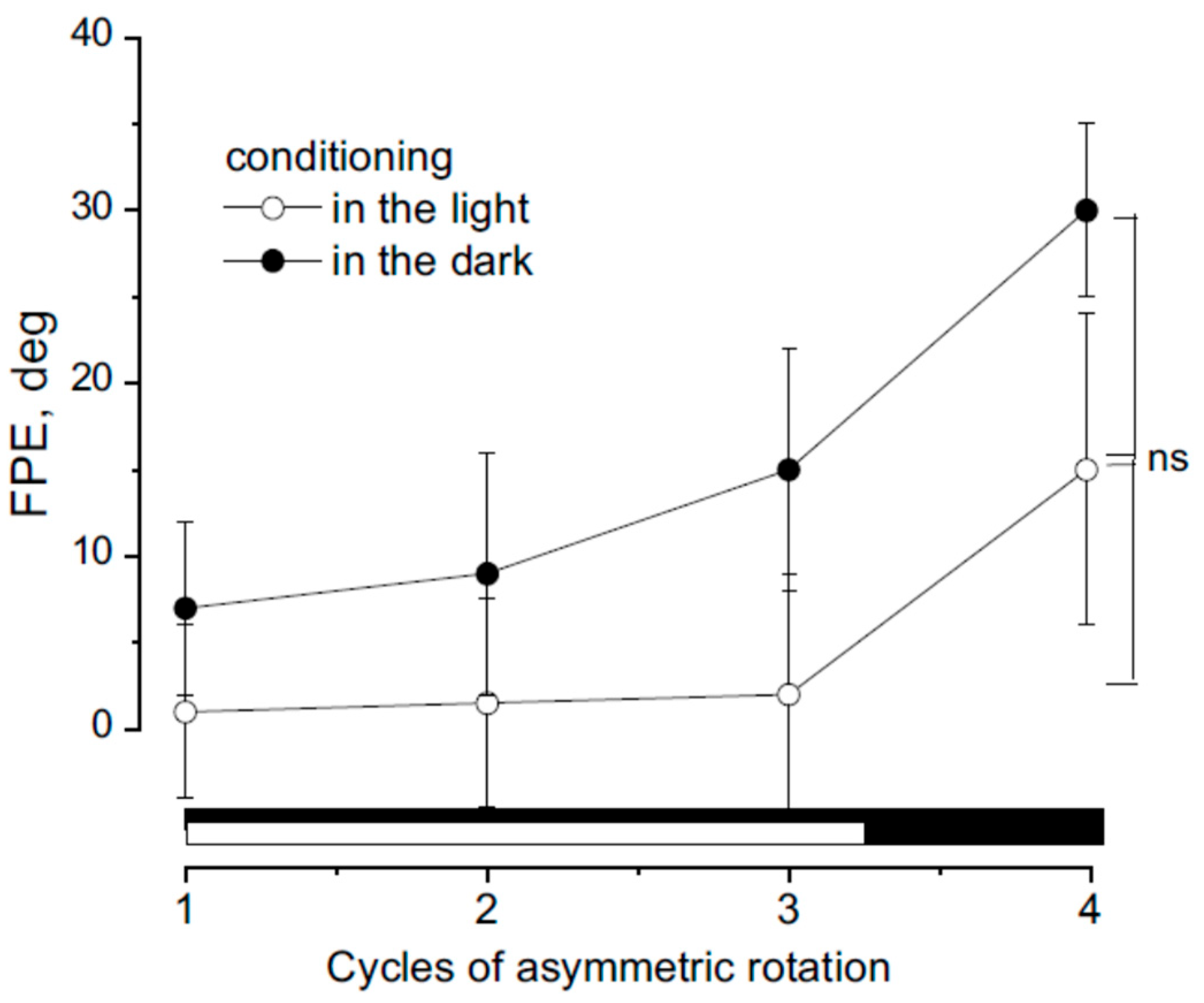
Figure 5.
Effect of different duration of neck muscle fatigue on FPE (1 ,3,6 min). The effect is compared with fatigue index from the EMG of the dorsal neck muscles. The FPE after fatiguing procedure was compared with the pre-fatigue. Note that the FPE was higher than the normal after a fatiguing procedure lasting 3 min (p< 0.01 **,°°) and 6 min (p< 0.001 ***,°°°).
Figure 5.
Effect of different duration of neck muscle fatigue on FPE (1 ,3,6 min). The effect is compared with fatigue index from the EMG of the dorsal neck muscles. The FPE after fatiguing procedure was compared with the pre-fatigue. Note that the FPE was higher than the normal after a fatiguing procedure lasting 3 min (p< 0.01 **,°°) and 6 min (p< 0.001 ***,°°°).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



