
Submitted:
21 July 2025
Posted:
22 July 2025
You are already at the latest version
Abstract
Conclusions: Our results indicate that polymorphism of these genes might play a role in the development of osteoporosis in Korean pre-menopausal women.
Keywords:
osteoporosis
; GWAS
; polymorphism
; menopause
; bone mineral density
1. Introduction
Osteoporosis is one of the most common age-related diseases and is characterized by a reduced bone mineral density (BMD). It is the most common cause of fractures in the elderly, and it can also lead to serious complications and even death. [1]. According to the ‘Sixth Korea National Health and Nutrition Examination Survey (KNHANES Ⅶ-1), 2016, Korea Centers for Disease Control and Prevention’, osteoporosis generally occurs in the elderly (42.6% in women > 65 years), although it can also occur in younger people (5.3% in women < 65 years).
The most common causes of osteoporosis are old age and menopause. It may be caused by systemic or genetic diseases, and it is also known to be related to family history, race, nutrition, and smoking and drinking habits [2]. Previous studies have reported that body weight, nutrition, and the genotypes of VDR and ER gene affect BMD in young women [3]. Hyperparathyroidism [4], hyperthyroidism [5], excessive drinking [6], glucocorticoid [7], and so on can also lead to osteoporosis in young women. Genetic factors, such as familial history also play a role in the value of BMD in young women [8]. There are many studies exploring the genetic causes of low BMD osteoporosis in post-menopausal women, and many genetic polymorphisms have been identified in individuals with different ethnicities. Single nucleotide polymorphisms (SNPs) of VDR and OPG gene were found to be associated with osteoporosis in Chinese post-menopausal women [9]. SNP of ESR1 gene has been also found to reduce BMD in post-menopausal women of southern Slovakia [10]. The polymorphism of the RANKL gene related to bone metabolism is also associated with osteoporosis in post-menopausal women [11]. However, there are few studies evaluating the association of genetic causes and osteoporosis in pre-menopausal women.
The Illumina Infinium HumanExome BeadChip targets approximately 240,000 coding variants, allowing for intensive detection of missense and nonsense variants that are critical for protein function. However, its coverage is limited to coding regions, and it exhibits relatively low statistical power for detecting rare or low-frequency variants [12]. The Affymetrix Axiom Exome Array demonstrates a very high positive predictive value (PPV) for most variants, offering high accuracy and reproducibility. However, it has a limitation: the PPV for heterozygous calls tends to be lower for exremely rare variants with a minor allele frequency (MAF) of less than 0.01% [13]. The advantages of these two technologies can complement each other's weaknesses. As they have different design strategies and signal readout mechanisms, they could complement platform-specific technical biases and variant calling errors when used together.
Therefore, in this study, we conducted a large-scale genetic analysis using data from both the Illumina Infinium HumanExome BeadChip and the Affymetrix Axiom Exome Array, based on participants from the Anseong and Ansan cohorts of the Korean Genome and Epidemiology Study (KoGES). The aim of this study was to identify genetic variants associated with the development of osteoporosis in Korean premenopausal women.
2. Results
The demographic characteristics of the subjects participating in the study are shown in Table 1.
A total of 304 subjects (57 osteoporosis patients and 247 healthy controls) were included in the analysis. Age, alcohol consumption, and calcium consumption did not significantly differ between the osteoporosis patients and healthy controls. There was no statistically significant difference between the two groups because the subjects who had a medical history of fracture or arthritis, smoking habit, long term steroid intake, and hormone therapy were excluded. Although previous studies have demonstrated that weight and BMI could be risk factors of osteoporosis in young women [14,15], these parameters were significantly higher in osteoporosis patient group in this study. The BMD values, distal radius speed of sound (DR-SOS), distal radius T-score (DR-T), distal radius Z-score (DR-Z), mid-shaft tibia speed of sound (MT-SOS), mid-shaft tibia T-score (MT-T), and mid-shaft tibia Z-score (MT-Z) were significantly higher in the control group.
Figure 1 is a Manhattan plot and Q-Q plot showing the results of the SNP analysis in the Illumina Infinium HumanExome BeadChip and Affymetrix Axiom exome arrays. 1,253 SNPs in the Illumina Infinium HumanExome BeadChip and 15,874 SNPs in the Affymetrix Axiom exome array were found to be statistically significant with a P value of < 0.05.
Panel A presents the analysis results from the Illumina Infinium HumanExome BeadChip, while Panel B shows the results from the Affymetrix Axiom Exome Array. In the Manhattan plot of Panel A, significant signals are generally well distributed across chromosomes, with notable peaks in −log10(P) values observed on certain chromosomes (e.g., chromosomes 6 and 11). The Q-Q plot reveals that most SNPs lie close to the diagonal line representing the expected and observed −log10(P) values, indicating that the statistical tests were performed appropriately. However, a number of points in the upper right corner deviate from this line, suggesting the presence of SNPs with higher-than-expected significance. Similarly, the Manhattan plot in Panel B demonstrates a widespread distribution of significant signals, with some SNPs reaching −log10(P) values as high as 5.8. The corresponding Q-Q plot also aligns closely with the diagonal under the null hypothesis, though several SNPs display clear deviations, further supporting the presence of statistically significant variants.
Figure 2 shows the chromosomal locations of the statistically significant SNPs in both the Illumina Infinium HumanExome BeadChip and the Affymetrix Axiom exome array. As shown in Figure 1, 113 SNPs were found to be significant in 69 genes (P < 0.05). Among them, 41 SNPs with no information of genes were excluded and the locations of the 72 SNPs were plotted in Figure 3. The red-marked locations are SNPs with P value < 0.01 in both analyses.
Figure 3 shows the protein-protein interaction (PPI) network of genes that were commonly significant (P-value < 0.05) in the Illumina BeadChip and Affymetrix Axiom exome arrays generated using the STRING database. Six clusters were distinguished through K-means clustering (K = 6). Genes within each of these clusters are likely to share similar biological functions. Among these genes, NCAM1, PECAM1, GNL3, and PTPRD appear to be hub genes with multiple interactions within the network.
The prediction of protein damage using PolyPhen-2, SIFT, and PROVEAN revealed that 6 SNPs (rs1799852 SNP in TF, rs11917356 SNP in COL6A5, rs2276360 in NADSYN1, rs1128431 SNP in EFTUD1, and rs7232237 and rs2282632 SNPs in ASXL3) were associated with damage of the protein structure. Table 2 shows the SNPs that could affect the structure of the proteins. Most of them were predicted to be benign; however, the rs1128431 SNP in the EFTUD1 gene was predicted to have a possible functional damage according to both PolyPhen-2 and SIFT.
Table 3 shows the list of statistically significant SNPs associated with osteoporosis in pre-menopausal women in both the analyses (P < 0.01). A total of 18 SNPs were found to be statistically significant in both the analyses: rs783540 SNP in CPEB1, rs3731646 SNP in SH3BP4, rs10506525 SNP in MSRB3, rs2110871 SNP in MAGI2, rs2172802 SNP in LPHN3, rs6895902 SNP in MAML1, rs2020945 SNP in PWP2, rs3756987 SNP in RSPH3, rs2286550 SNP in CATSPERG, rs4729759 SNP in CUX1, rs10513680 SNP in SAMD7, rs1052053 SNP in PMF1-BFLAP, rs2764020 SNP in STARD13, rs7088318 SNP in PIP4K2A, rs151719 SNP in HLA-DMB, rs2302234 SNP in FAM20A, rs16990991 SNP in EFCAB6 and rs12757165 SNP in ESRRG gene. In particular, rs783540 exhibited the strongest association with osteoporosis, showing a P-value of 0.000 in both analyses. This cross-platform consistency may strengthen the reliability of these SNPs as potential candidate markers.
3. Discussion
Previous studies have shown the close relationship between menopause and the development of osteoporosis. Estrogen deficiency due to menopause accelerates the induction of M-CSF, RANKL, and TNF-α, which promote a blood calcium concentration and bone resorption [16]. Estrogen deficiency also affects the production of parathyroid hormone and results in decreased intestinal calcium absorption and renal calcium conservation [17]. Parathyroid hormone is associated with vitamin D, which also regulates calcium absorption in the intestine [18,19]. Increased blood calcium level and decreased intestinal calcium absorption leads to decreased bone formation [20]. Estrogen is directly related to bone metabolism. Estrogen promotes the apoptosis of osteoclasts [21], and its deficiency increases osteoclastogenesis [22] as well as apoptosis of osteocytes and osteoblasts [23].
Our results showed that many genes are associated with osteoporosis in pre-menopausal women. In the pre-menopausal women, BMD could be reduced even before menopause. This implies that a mechanism other than menopause might play a role in the development of osteoporosis. Although not all the genes in our results were found to be associated with bone metabolism, several potential genes were identified.
In our results, a number of genes were found to be significantly associated with BMD value or bone formation. ESRRG gene encodes a member of the estrogen receptor-related receptor family. The polymorphism of ESRRG gene is associated with the determination of bone density in European women [24]. The protein encoded by the gene is a sex- and RUNX2- dependent negative regulator of postnatal bone formation, as demonstrated in mice [25]. NTN4 gene encodes netrin-4 which promotes the differentiation and migration of osteoblasts and inhibits differentiation of osteoclast [26,27]. TF gene encodes transferrin, which is related to BMD [28]. The polymorphism of this gene is associated with an increased risk of osteonecrosis of the femoral head [29]. The CLEC gene is closely related to adaptive immunity. The polymorphism of the CLEC16A gene could cause an alteration in the leukocyte count and lead to reduced BMD and fractures in elderly women [30]. In our results, the genes encoding adhesion molecules were also found to be involved in bone formation and resorption. PECAM1 gene encodes the platelet and endothelial cell adhesion molecule 1, and the protein encoded by this gene is a negative regulator of monocyte-derived osteoclastogenesis. Loss of this gene increases osteoclastogenesis and leads to bone loss [31]. NCAM1 gene encodes a cell adhesion molecule in the immunoglobulin superfamily and it plays various roles in cell differentiation including osteogenesis [32]. The roles of GGT1 and COL6A5 genes in bone metabolism are well studied in rats but less in humans. The function of GGT1 gene in human is not well understood, but mutation of GGT1 gene in rats has been found to promote the development of osteoclasts and increase bone resorption resulting in osteoporosis [33]. GGT1 is an activator of TLR4-mediated osteoclastogenesis and GGT1 overexpressed transgenic mice exhibited symptoms of osteoporosis [34]. COL6A5 is present in almost all the tissues in mice, but only in the skin, lung, testis, colon, and small bowel in humans. Therefore, studies in human generally focus on the skin and the small intestine [34]. However, another study reported that the polymorphism in COL6A5 gene could be linked to variations in BMD in both mouse and human [35]. STARD13 gene encodes a protein that may be involved in the regulation of cytoskeletal reorganization, cell proliferation, and so on. Some recent microRNA-based studies have shown that miR-125 is up-regulated in patients with osteoporosis, and STARD13 is the target of it [36,37].
It is well known that vitamin D plays an important role in BMD and development of osteoporosis. Our results demonstrated the association between SNPs in several genes associated with vitamin D metabolism and the development of osteoporosis in pre-menopausal women. NADSYN1 gene encodes NAD synthetase 1 which plays an important role in the synthesis of nicotinamide adenine dinucleotide. The polymorphism of NADSYN1 gene is associated with vitamin D level and metabolic profile [38]. NADSYN1 gene is one of the vitamin D pathway gene, the polymorphism of which affects the level of vitamin D in pregnant women [39]. EFTUD1 gene is a target gene for vitamin D in the mammary cells [40], and it also plays an important role in mediating the pro-apoptotic effects of vitamin D in breast cancer [41]. Table 2 shows that rs1128431 SNP of EFTUD1 gene can have a severe effect on the protein structure. This means that rs1129431 SNP can affect the function of the EFTUD1 protein.
The relationship between muscle and osteoporosis is well known. Decreased muscle strength is related to decreased BMD [42], and a decline in the function of skeletal muscle results in osteoporosis [43]. It is reported that the PACSIN2 and ESRRG genes, which were found to be significant in our results, are associated with skeletal muscle exercise. Differential changes in the expression were observed in transcriptome analysis during aerobic exercise [44].
The genes associated with the reproductive system were found to be significantly associated with osteoporosis in our results. Androgen deficiency is a common cause for the development of osteoporosis and androgen is used for the treatment of osteoporosis [45]. The EFCAB6 gene was called DJBP and is mainly expressed in the testis. The protein encoded by this gene plays a role in the negative regulation of the androgen receptor [46]. ASXL3 is mostly expressed in the bone [47]. It is reported that mutations in ASXL3 gene are associated with sporadic primary hyperparathyroidism and recurrently mutated in sporadic parathyroid adenomas [48]. ASXL3 is a gene associated with the androgen pathway and is regulated by androgen in the human neural cells [49]. ESRRG gene takes part in one of the estrogen pathways and may affect estrogenic response related to female [50]. NTN4 encodes netrin-4 which is required for the development of the mammary gland and is expressed in the normal breast epithelium. It plays an important role in the prognosis of ER-positive breast cancer [51]. As mentioned above, EFTUD1 is also associated with breast cancer. CPEB1 gene is another interesting gene and the rs783540 SNP of CPEB1 gene has shown very high significance in both the analyses in our study (P < 0.001). Some previous studies have shown the relationship between CPEB1 and reproductive system. A previous microRNA study reported that the CPEB1 gene is a target of miR-3646 and is downregulated in breast and ovarian cancers [52]. CPEB1 gene plays an important role in oocyte maturation. It regulates mRNA translation during oocyte maturation [53]. The polymorphism of CPEB1 gene might result in premature ovarian insufficiency [54]. A recent study has reported that oocytes maturation is associated with BMD, and superovulation decreases BMD [55].
Several genes were studied to be associated with the development of osteoporosis in pre-menopausal women. It is well known that menopause and aging are the major causes of osteoporosis. However, other causes might be also involved in the development of osteoporosis as it can also develop in the younger women. Other causes of osteoporosis include smoking, drinking, history of fracture, steroid and hormone therapy, systemic diseases, genetic factors and so on. Therefore, the other factors were matched in this study as much as possible, and the subjects who were young age and pre-menopausal women were involved in this analysis.
Table 4 summarizes the significant genes found to be associated with the development of osteoporosis in pre-menopausal women in this study. The genes that have been reported to be associated with osteogenesis, osteoclastogenesis, and BMD value were shown to be associated with osteoporosis in pre-menopausal women. Additionally, the genes involved in vitamin D metabolism, vitamin D pathway, and skeletal muscle exercise were shown to be significantly associated with osteoporosis. Finally, the genes associated with the reproductive process were significantly associated with osteoporosis. It is well known that menopause and estrogen deficiency are associated with the development of osteoporosis. It is important to note that the polymorphisms of genes involved in the reproductive processes or sex hormones are also associated with the development of osteoporosis in pre-menopausal women.
In this study, we identified various biological pathways associated with osteoporosis based on SNPs that were significantly detected using both platforms. Genetic pathways related to estrogen signaling and the reproductive system were observed in some variants, suggesting a potential association between hormonal changes and bone density loss in premenopausal women. Additionally, genes involved in ovarian function, germ cell maturation, androgen receptor regulation, and post-transcriptional regulation were also implicated in bone density regulation, further supporting the influence of hormonal mechanisms on early bone loss. Moreover, gene functions related to osteogenesis, differentiation of osteoblasts and osteoclasts, microRNA regulation, extracellular matrix organization, vitamin D metabolism, and muscle–bone interactions were identified. These findings indicate that osteoporosis is likely influenced by complex interactions among hormonal, metabolic, immune, and inter-tissue signaling pathways, rather than by a single molecular mechanism.
The main limitation of this study is the relatively small sample size. However, the incidence of osteoporosis in premenopausal women is low, and when excluding individuals with known risk factors such as alcohol use, steroid medication, or fracture history, the rate becomes even lower [58]. In this study, we sought to minimize the influence of external risk factors in order to more clearly evaluate the role of genetic predisposition. Additionally, the use of two independent genotyping platforms applied to the same population enhanced the reliability of our findings. The consistently detected variants may play a significant role in the development of osteoporosis in young Korean women and could serve as foundational data for future functional studies and the development of predictive genetic models.
4. Materials and Methods
4.1. Study Subjects
The subjects analyzed in this study were obtained from the Korean Genome and Epidemiology Study cohort. A total of 7,077 subjects from three phases of the Anseong and Ansan Cohort Study were screened and finally 304 subjects (57 osteoporosis patients and 247 healthy controls) were selected. Figure 4 shows the process of selection of 304 subjects from a pool of 7,077 subjects. First, men and post-menopausal women were excluded. Subsequently, T-scores and Z-scores were analyzed. Since the study focused exclusively on premenopausal women, the Z-score was also included as a criterion for assessment. Thus, the subjects with T-score < -2.5 and Z-score < -2.0 were classified as osteoporosis patients and, the subjects with T-score > -1.0 were classified as controls. The subjects with -2.5 < T-score < -1.0 were excluded from the analysis. To analyze the influence of the genetic risk factors on osteoporosis, other risk factors were controlled, such as environmental factors, general factors, habits, medical history and so on [12]. The subjects with a body weight < 58 kg, body mass index (BMI) < 19, alcohol consumption > 10 ml/day, smoking history, medication history of long-term steroid or hormone therapy, thyroid disorder, fracture, or arthritis were excluded. There were no data on parathyroid disorder or vitamin D consumption. As few subjects consumed calcium supplements more than the recommended dosage, they were excluded, and the subjects who consume less than the recommended dosage of calcium supplements were included in the study. Finally, 57 osteoporosis patients and 247 healthy controls were included in the study. The demographic characteristics of the subjects are shown in Table 1. This study was approved by the Institutional Review Board of the Dankook University (IRB no. 2018-08-004).
4.2. Statistical Analysis
To compare the osteoporosis patients and healthy controls, independent Student’s t-test was performed using PAWS 18.0 (SPSS, Chicago, IL, USA). The results of 30,538 SNPs for each subject were obtained using Illumina Infinium HumanExome BeadChip and the results of 242,901 SNPs for each subject were obtained using an Affymetrix Axiom exome array (Affymetrix, Santa Clara, California, USA). A logistic regression analysis was performed to examine the association between the development of osteoporosis and genetic polymorphisms using SNP & Variation Suite (Golden Helix, Bozeman, MT, USA) and PLINK software [59]. SNPs with MAF ≥ 0.05 and HWE P-value ≥ 0.01 were removed. Manhattan plots and QQ plots were plotted using SNPEVG program [60]. PhenoGram was used for the scheme of the location of the SNPs in chromosomes (http://visualization.ritchielab.org). The PolyPhen-2, SIFT, and PROVEAN were used for predicting protein damage by the SNPs [61,62,63].
5. Conclusions
Our results showed that the genes associated with bone metabolism, vitamin D metabolism, skeletal muscle exercise and reproductive processes were closely associated with the development of osteoporosis in pre-menopausal women. Among these, the CPEB1 gene, which was associated with oocyte maturation, had the most statistically significant association in both the analyses (P < 0.001). Our results showed that the development of osteoporosis in pre-menopausal women might have a relationship with genetic factors involved in various mechanisms. Further studies on these genes should be conducted in the future.
Author Contributions
SWK established a study plan, provided an idea of the mechanism of the osteoporosis and received funding. SKK analyzed the data, performed the statistics and wrote this manuscript. BJY, SJH and GK analyzed clinical information and analyzed genes for osteoporosis.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (No. NRF-2017R1C1B2012084). This study was conducted with bioresources from National Biobank of Korea, the Centers for Disease Control and Prevention, Republic of Korea (KBN-2018-058).
Institutional Review Board Statement
This study was approved by the Institutional Review Board of the Dankook University (IRB no. 2018-08-004).
Acknowledgments
We used ChatGPT (OpenAI, 2025) to assist in the English translation and to enhance the clarity of the manuscript. All conceptual content and data analysis were performed by the authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Adachi, J. D.; Ioannidis, G.; Pickard, L.; Berger, C.; Prior, J. C.; Joseph, L.; Hanley, D. A.; Olszynski, W. P.; Murray, T. M.; Anastassiades, T.; Hopman, W.; Brown, J. P.; Kirkland, S.; Joyce, C.; Papaioannou, A.; Poliquin, S.; Tenenhouse, A.; Papadimitropoulos, E. A. , The association between osteoporotic fractures and health-related quality of life as measured by the Health Utilities Index in the Canadian Multicentre Osteoporosis Study (CaMos). Osteoporos Int 2003, 14, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Smith, R. , Osteoporosis: cause and management. Br Med J (Clin Res Ed) 1987, 294, 329–332. [Google Scholar] [CrossRef]
- McGuigan, F. E.; Murray, L.; Gallagher, A.; Davey-Smith, G.; Neville, C. E.; Van't Hof, R.; Boreham, C.; Ralston, S. H. , Genetic and environmental determinants of peak bone mass in young men and women. J Bone Miner Res 2002, 17, 1273–1279. [Google Scholar] [CrossRef]
- Chen, K. Y.; Wang, C. H.; Lin, T. Y.; Chang, C. Y.; Liu, C. L.; Hsiao, Y. C.; Hung, C. C.; Wang, N. C. , Monitoring early developed low bone mineral density in HIV-infected patients by intact parathyroid hormone and circulating fibroblast growth factor 23. J Microbiol Immunol Infect 2019, 52, 693–699. [Google Scholar] [CrossRef]
- Sato, K. , [Graves' disease and bone metabolism]. Nihon Rinsho 2006, 64, 2317–2322. [Google Scholar]
- Seo, S.; Chun, S.; Newell, M. A.; Yun, M. , Association between alcohol consumption and Korean young women's bone health: a cross sectional study from the 2008 to 2011 Korea National Health and Nutrition Examination Survey. BMJ Open 2015, 5, e007914. [Google Scholar] [CrossRef] [PubMed]
- Briot, K. , Bone and glucocorticoids. Ann Endocrinol (Paris) 2018, 79, 115–118. [Google Scholar] [CrossRef]
- Jouanny, P.; Guillemin, F.; Kuntz, C.; Jeandel, C.; Pourel, J. , Environmental and genetic factors affecting bone mass. Similarity of bone density among members of healthy families. Arthritis Rheum 1995, 38, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, D.; Shen, G.; Cui, Y.; Lv, Q.; Wei, F. , Association of VDR and OPG gene polymorphism with osteoporosis risk in Chinese postmenopausal women. Climacteric 2019, 22, 208–212. [Google Scholar] [CrossRef]
- Mondockova, V.; Adamkovicova, M.; Lukacova, M.; Grosskopf, B.; Babosova, R.; Galbavy, D.; Martiniakova, M.; Omelka, R. , The estrogen receptor 1 gene affects bone mineral density and osteoporosis treatment efficiency in Slovak postmenopausal women. BMC Med Genet 2018, 19, 174. [Google Scholar] [CrossRef]
- Wolski, H.; Drews, K.; Bogacz, A.; Kaminski, A.; Barlik, M.; Bartkowiak-Wieczorek, J.; Klejewski, A.; Ozarowski, M.; Majchrzycki, M.; Seremak-Mrozikiewicz, A. , The RANKL/RANK/OPG signal trail: significance of genetic polymorphisms in the etiology of postmenopausal osteoporosis. Ginekol Pol 2016, 87, 347–352. [Google Scholar] [CrossRef]
- Igo, R. P., Jr.; Cooke Bailey, J. N.; Romm, J.; Haines, J. L.; Wiggs, J. L. , Quality Control for the Illumina HumanExome BeadChip. Curr Protoc Hum Genet 2016, 90, 2–14. [Google Scholar] [CrossRef]
- Mizrahi-Man, O.; Woehrmann, M. H.; Webster, T. A.; Gollub, J.; Bivol, A.; Keeble, S. M.; Aull, K. H.; Mittal, A.; Roter, A. H.; Wong, B. A.; Schmidt, J. P. , Novel genotyping algorithms for rare variants significantly improve the accuracy of Applied Biosystems Axiom array genotyping calls: Retrospective evaluation of UK Biobank array data. PLoS One 2022, 17, e0277680. [Google Scholar] [CrossRef]
- Shin, C. S.; Choi, H. J.; Kim, M. J.; Kim, J. T.; Yu, S. H.; Koo, B. K.; Cho, H. Y.; Cho, S. W.; Kim, S. W.; Park, Y. J.; Jang, H. C.; Kim, S. Y.; Cho, N. H. , Prevalence and risk factors of osteoporosis in Korea: a community-based cohort study with lumbar spine and hip bone mineral density. Bone 2010, 47, 378–387. [Google Scholar] [CrossRef]
- Tatsumi, Y.; Higashiyama, A.; Kubota, Y.; Sugiyama, D.; Nishida, Y.; Hirata, T.; Kadota, A.; Nishimura, K.; Imano, H.; Miyamatsu, N.; Miyamoto, Y.; Okamura, T. , Underweight Young Women Without Later Weight Gain Are at High Risk for Osteopenia After Midlife: The KOBE Study. J Epidemiol 2016, 26, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Cenci, S.; Weitzmann, M. N.; Roggia, C.; Namba, N.; Novack, D.; Woodring, J.; Pacifici, R. , Estrogen deficiency induces bone loss by enhancing T-cell production of TNF-alpha. J Clin Invest 2000, 106, 1229–1237. [Google Scholar] [CrossRef]
- Khosla, S.; Atkinson, E. J.; Melton, L. J., 3rd; Riggs, B. L. , Effects of age and estrogen status on serum parathyroid hormone levels and biochemical markers of bone turnover in women: a population-based study. J Clin Endocrinol Metab 1997, 82, 1522–1527. [Google Scholar] [CrossRef]
- Ebeling, P. R.; Sandgren, M. E.; DiMagno, E. P.; Lane, A. W.; DeLuca, H. F.; Riggs, B. L. , Evidence of an age-related decrease in intestinal responsiveness to vitamin D: relationship between serum 1,25-dihydroxyvitamin D3 and intestinal vitamin D receptor concentrations in normal women. J Clin Endocrinol Metab 1992, 75, 176–182. [Google Scholar] [PubMed]
- Lips, P.; Wiersinga, A.; van Ginkel, F. C.; Jongen, M. J.; Netelenbos, J. C.; Hackeng, W. H.; Delmas, P. D.; van der Vijgh, W. J. , The effect of vitamin D supplementation on vitamin D status and parathyroid function in elderly subjects. J Clin Endocrinol Metab 1988, 67, 644–650. [Google Scholar] [CrossRef]
- Holick, M. F. , Perspective on the impact of weightlessness on calcium and bone metabolism. Bone 1998, (5 Suppl), 105S–111S. [Google Scholar] [CrossRef]
- Hughes, D. E.; Dai, A.; Tiffee, J. C.; Li, H. H.; Mundy, G. R.; Boyce, B. F. , Estrogen promotes apoptosis of murine osteoclasts mediated by TGF-beta. Nat Med 1996, 2, 1132–1136. [Google Scholar] [CrossRef]
- D'Amelio, P.; Grimaldi, A.; Di Bella, S.; Brianza, S. Z. M.; Cristofaro, M. A.; Tamone, C.; Giribaldi, G.; Ulliers, D.; Pescarmona, G. P.; Isaia, G. , Estrogen deficiency increases osteoclastogenesis up-regulating T cells activity: a key mechanism in osteoporosis. Bone 2008, 43, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M. A.; Haugh, M. G.; O'Brien, F. J.; McNamara, L. M. , Estrogen withdrawal from osteoblasts and osteocytes causes increased mineralization and apoptosis. Horm Metab Res 2014, 46, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Elfassihi, L.; Giroux, S.; Bureau, A.; Laflamme, N.; Cole, D. E.; Rousseau, F. , Association with replication between estrogen-related receptor gamma (ESRRgamma) polymorphisms and bone phenotypes in women of European ancestry. Journal of bone and mineral research : the official journal of the American Society for Bone and Mineral Research 2010, 25, 901–911. [Google Scholar] [CrossRef]
- Cardelli, M.; Aubin, J. E. , ERRgamma is not required for skeletal development but is a RUNX2-dependent negative regulator of postnatal bone formation in male mice. PLoS One 2014, 9, e109592. [Google Scholar] [CrossRef] [PubMed]
- Enoki, Y.; Sato, T.; Kokabu, S.; Hayashi, N.; Iwata, T.; Yamato, M.; Usui, M.; Matsumoto, M.; Tomoda, T.; Ariyoshi, W.; Nishihara, T.; Yoda, T. , Netrin-4 Promotes Differentiation and Migration of Osteoblasts. In vivo 2017, 31, 793–799. [Google Scholar]
- Enoki, Y.; Sato, T.; Tanaka, S.; Iwata, T.; Usui, M.; Takeda, S.; Kokabu, S.; Matsumoto, M.; Okubo, M.; Nakashima, K.; Yamato, M.; Okano, T.; Fukuda, T.; Chida, D.; Imai, Y.; Yasuda, H.; Nishihara, T.; Akita, M.; Oda, H.; Okazaki, Y.; Suda, T.; Yoda, T. , Netrin-4 derived from murine vascular endothelial cells inhibits osteoclast differentiation in vitro and prevents bone loss in vivo. FEBS letters 2014, 588, 2262–2269. [Google Scholar] [CrossRef]
- D'Amelio, P.; Cristofaro, M. A.; Tamone, C.; Morra, E.; Di Bella, S.; Isaia, G.; Grimaldi, A.; Gennero, L.; Gariboldi, A.; Ponzetto, A.; Pescarmona, G. P.; Isaia, G. C. , Role of iron metabolism and oxidative damage in postmenopausal bone loss. Bone 2008, 43, 1010–1015. [Google Scholar] [CrossRef]
- Hong, J. M.; Kim, T. H.; Kim, H. J.; Park, E. K.; Yang, E. K.; Kim, S. Y. , Genetic association of angiogenesis- and hypoxia-related gene polymorphisms with osteonecrosis of the femoral head. Exp Mol Med 2010, 42, 376–385. [Google Scholar] [CrossRef]
- Swanberg, M.; McGuigan, F. E.; Ivaska, K. K.; Gerdhem, P.; Akesson, K. , Polymorphisms in the inflammatory genes CIITA, CLEC16A and IFNG influence BMD, bone loss and fracture in elderly women. PLoS One 2012, 7, e47964. [Google Scholar] [CrossRef]
- Wu, Y.; Tworkoski, K.; Michaud, M.; Madri, J. A. , Bone marrow monocyte PECAM-1 deficiency elicits increased osteoclastogenesis resulting in trabecular bone loss. Journal of immunology 2009, 182, 2672–2679. [Google Scholar] [CrossRef]
- Fang, J.; Hall, B. K. , N-CAM is not required for initiation of secondary chondrogenesis: the role of N-CAM in skeletal condensation and differentiation. The International journal of developmental biology 1999, 43, 335–342. [Google Scholar]
- Hiramatsu, K.; Asaba, Y.; Takeshita, S.; Nimura, Y.; Tatsumi, S.; Katagiri, N.; Niida, S.; Nakajima, T.; Tanaka, S.; Ito, M.; Karsenty, G.; Ikeda, K. , Overexpression of gamma-glutamyltransferase in transgenic mice accelerates bone resorption and causes osteoporosis. Endocrinology 2007, 148, 2708–2715. [Google Scholar] [CrossRef]
- Moriwaki, S.; Into, T.; Suzuki, K.; Miyauchi, M.; Takata, T.; Shibayama, K.; Niida, S. , gamma-Glutamyltranspeptidase is an endogenous activator of Toll-like receptor 4-mediated osteoclastogenesis. Sci Rep 2016, 6, 35930. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pandey, A. K.; Mulligan, M. K.; Williams, E. G.; Mozhui, K.; Li, Z.; Jovaisaite, V.; Quarles, L. D.; Xiao, Z.; Huang, J.; Capra, J. A.; Chen, Z.; Taylor, W. L.; Bastarache, L.; Niu, X.; Pollard, K. S.; Ciobanu, D. C.; Reznik, A. O.; Tishkov, A. V.; Zhulin, I. B.; Peng, J.; Nelson, S. F.; Denny, J. C.; Auwerx, J.; Lu, L.; Williams, R. W. , Joint mouse-human phenome-wide association to test gene function and disease risk. Nature communications 2016, 7, 10464. [Google Scholar] [CrossRef]
- Chen, R.; Liao, X.; Chen, F.; Wang, B.; Huang, J.; Jian, G.; Huang, Z.; Yin, G.; Liu, H.; Jin, D. , Circulating microRNAs, miR-10b-5p, miR-328-3p, miR-100 and let-7, are associated with osteoblast differentiation in osteoporosis. Int J Clin Exp Pathol 2018, 11, 1383–1390. [Google Scholar] [PubMed]
- Tang, F.; Zhang, R.; He, Y.; Zou, M.; Guo, L.; Xi, T. , MicroRNA-125b induces metastasis by targeting STARD13 in MCF-7 and MDA-MB-231 breast cancer cells. PLoS One 2012, 7, e35435. [Google Scholar] [CrossRef]
- Foucan, L.; Velayoudom-Cephise, F. L.; Larifla, L.; Armand, C.; Deloumeaux, J.; Fagour, C.; Plumasseau, J.; Portlis, M. L.; Liu, L.; Bonnet, F.; Ducros, J. , Polymorphisms in GC and NADSYN1 Genes are associated with vitamin D status and metabolic profile in Non-diabetic adults. BMC endocrine disorders 2013, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.; Jiang, S.; Muyiduli, X.; Wang, S.; Mo, M.; Li, M.; Wang, Z.; Yu, Y. , Vitamin D pathway gene polymorphisms influenced vitamin D level among pregnant women. Clin Nutr 2018, (6 Pt A), 2230–2237. [Google Scholar] [CrossRef]
- Simmons, K. M.; Beaudin, S. G.; Narvaez, C. J.; Welsh, J. , Gene Signatures of 1,25-Dihydroxyvitamin D3 Exposure in Normal and Transformed Mammary Cells. Journal of cellular biochemistry 2015, 116, 1693–1711. [Google Scholar] [CrossRef]
- Sheng, L.; Anderson, P. H.; Turner, A. G.; Pishas, K. I.; Dhatrak, D. J.; Gill, P. G.; Morris, H. A.; Callen, D. F. , Identification of vitamin D(3) target genes in human breast cancer tissue. J Steroid Biochem Mol Biol 2016, 164, 90–97. [Google Scholar] [CrossRef]
- Locquet, M.; Beaudart, C.; Reginster, J. Y.; Bruyere, O. , Association Between the Decline in Muscle Health and the Decline in Bone Health in Older Individuals from the SarcoPhAge Cohort. Calcif Tissue Int 2019, 104, 273–284. [Google Scholar] [CrossRef]
- Yoo, S. Z.; No, M. H.; Heo, J. W.; Park, D. H.; Kang, J. H.; Kim, S. H.; Kwak, H. B. , Role of exercise in age-related sarcopenia. J Exerc Rehabil 2018, 14, 551–558. [Google Scholar] [CrossRef]
- Popov, D. V.; Makhnovskii, P. A.; Kurochkina, N. S.; Lysenko, E. A.; Vepkhvadze, T. F.; Vinogradova, O. L. , Intensity-dependent gene expression after aerobic exercise in endurance-trained skeletal muscle. Biol Sport 2018, 35, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Negro-Vilar, A. , Selective androgen receptor modulators (SARMs): a novel approach to androgen therapy for the new millennium. J Clin Endocrinol Metab 1999, 84, 3459–3462. [Google Scholar] [CrossRef]
- Niki, T.; Takahashi-Niki, K.; Taira, T.; Iguchi-Ariga, S. M.; Ariga, H. , DJBP: a novel DJ-1-binding protein, negatively regulates the androgen receptor by recruiting histone deacetylase complex, and DJ-1 antagonizes this inhibition by abrogation of this complex. Mol Cancer Res 2003, 1, 247–261. [Google Scholar] [PubMed]
- Fishilevich, S.; Zimmerman, S.; Kohn, A.; Iny Stein, T.; Olender, T.; Kolker, E.; Safran, M.; Lancet, D. , Genic insights from integrated human proteomics in GeneCards. Database (Oxford) 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Sun, B.; Wang, Z. P.; He, J. W.; Fu, W. Z.; Fan, Y. B.; Zhang, Z. L. , Whole-Exome Sequencing Identifies Novel Recurrent Somatic Mutations in Sporadic Parathyroid Adenomas. Endocrinology 2018, 159, 3061–3068. [Google Scholar] [CrossRef]
- Quartier, A.; Chatrousse, L.; Redin, C.; Keime, C.; Haumesser, N.; Maglott-Roth, A.; Brino, L.; Le Gras, S.; Benchoua, A.; Mandel, J. L.; Piton, A. , Genes and Pathways Regulated by Androgens in Human Neural Cells, Potential Candidates for the Male Excess in Autism Spectrum Disorder. Biol Psychiatry 2018, 84, 239–252. [Google Scholar] [CrossRef]
- Giguere, V. , To ERR in the estrogen pathway. Trends in endocrinology and metabolism: TEM 2002, 13, 220–225. [Google Scholar] [CrossRef]
- Esseghir, S.; Kennedy, A.; Seedhar, P.; Nerurkar, A.; Poulsom, R.; Reis-Filho, J. S.; Isacke, C. M. , Identification of NTN4, TRA1, and STC2 as prognostic markers in breast cancer in a screen for signal sequence encoding proteins. Clinical cancer research : an official journal of the American Association for Cancer Research 2007, 13, 3164–3173. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C. N.; Ketabi, Z.; Rosenstierne, M. W.; Palle, C.; Boesen, H. C.; Norrild, B. , Expression of CPEB, GAPDH and U6snRNA in cervical and ovarian tissue during cancer development. APMIS 2009, 117, 53–59. [Google Scholar] [CrossRef]
- Sousa Martins, J. P.; Liu, X.; Oke, A.; Arora, R.; Franciosi, F.; Viville, S.; Laird, D. J.; Fung, J. C.; Conti, M. , DAZL and CPEB1 regulate mRNA translation synergistically during oocyte maturation. J Cell Sci 2016, 129, 1271–1282. [Google Scholar] [CrossRef]
- Hyon, C.; Mansour-Hendili, L.; Chantot-Bastaraud, S.; Donadille, B.; Kerlan, V.; Dode, C.; Jonard, S.; Delemer, B.; Gompel, A.; Reznik, Y.; Touraine, P.; Siffroi, J. P.; Christin-Maitre, S. , Deletion of CPEB1 Gene: A Rare but Recurrent Cause of Premature Ovarian Insufficiency. J Clin Endocrinol Metab 2016, 101, 2099–2104. [Google Scholar] [CrossRef]
- Zhang, J.; Lai, Z.; Shi, L.; Tian, Y.; Luo, A.; Xu, Z.; Ma, X.; Wang, S. , Repeated superovulation increases the risk of osteoporosis and cardiovascular diseases by accelerating ovarian aging in mice. Aging (Albany NY) 2018, 10, 1089–1102. [Google Scholar] [CrossRef]
- Chen, R. S.; Liao, X.; Chen, F. R.; Wang, B. W.; Huang, J. M.; Jian, G. J.; Huang, Z. Y.; Yin, G. H.; Liu, H. Y.; Jin, D. D. , Circulating microRNAs, miR-10b-5p, miR-328-3p, miR-100 and let-7, are associated with osteoblast differentiation in osteoporosis. Int J Clin Exp Patho 2018, 11, 1383. [Google Scholar]
- Sheng, L.; Anderson, P. H.; Turner, A. G.; Pishas, K. I.; Dhatrak, D. J.; Gill, P. G.; Morris, H. A.; Callen, D. F. , Identification of vitamin D3 target genes in human breast cancer tissue. The Journal of steroid biochemistry and molecular biology 2016, 164, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Vondracek, S. F.; Hansen, L. B.; McDermott, M. T. , Osteoporosis risk in premenopausal women. Pharmacotherapy 2009, 29, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Chang, C. C.; Chow, C. C.; Tellier, L. C.; Vattikuti, S.; Purcell, S. M.; Lee, J. J. , Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Wang, S.; Dvorkin, D.; Da, Y. , SNPEVG: a graphical tool for GWAS graphing with mouse clicks. BMC Bioinformatics 2012, 13, 319. [Google Scholar] [CrossRef]
- Adzhubei, I. A.; Schmidt, S.; Peshkin, L.; Ramensky, V. E.; Gerasimova, A.; Bork, P.; Kondrashov, A. S.; Sunyaev, S. R. , A method and server for predicting damaging missense mutations. Nat Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [PubMed]
- Vaser, R.; Adusumalli, S.; Leng, S. N.; Sikic, M.; Ng, P. C. , SIFT missense predictions for genomes. Nat Protoc 2016, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Chan, A. P. , PROVEAN web server: a tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Manhattan plot and Q-Q plot of GWAS. A: Illumina Infinium HumanExome BeadChip. B: Affymetrix Axiom exome array (P value of the threshold line = 0.01).
Figure 1.
Manhattan plot and Q-Q plot of GWAS. A: Illumina Infinium HumanExome BeadChip. B: Affymetrix Axiom exome array (P value of the threshold line = 0.01).
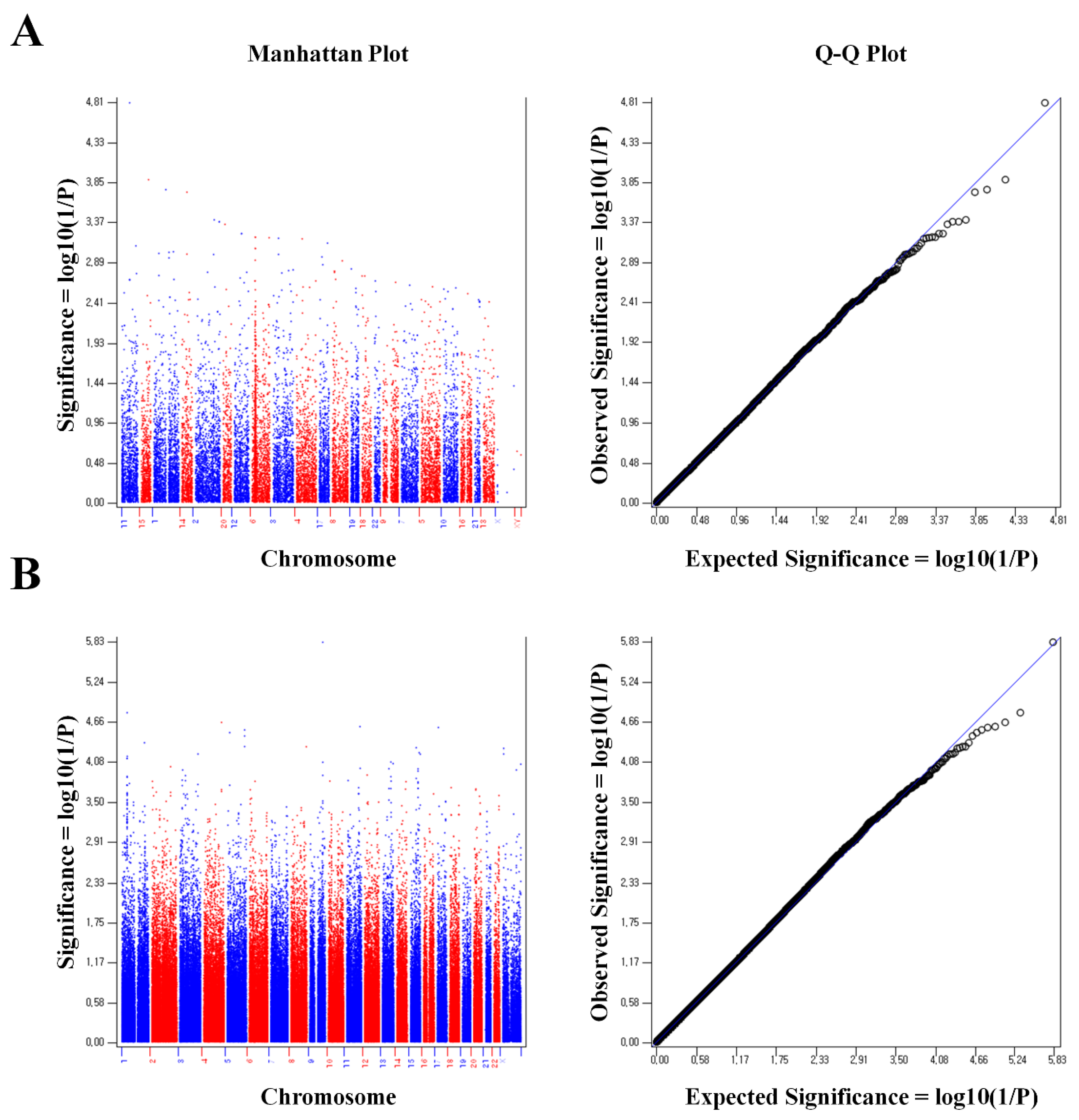
Figure 2.
Significant gene areas where linkage with osteoporosis in the pre-menopausal women are represented using phenogram (P < 0.05 in both Illumina Infinium HumanExome BeadChip and Affymetrix Axiom exome array. Red; P < 0.01).
Figure 2.
Significant gene areas where linkage with osteoporosis in the pre-menopausal women are represented using phenogram (P < 0.05 in both Illumina Infinium HumanExome BeadChip and Affymetrix Axiom exome array. Red; P < 0.01).
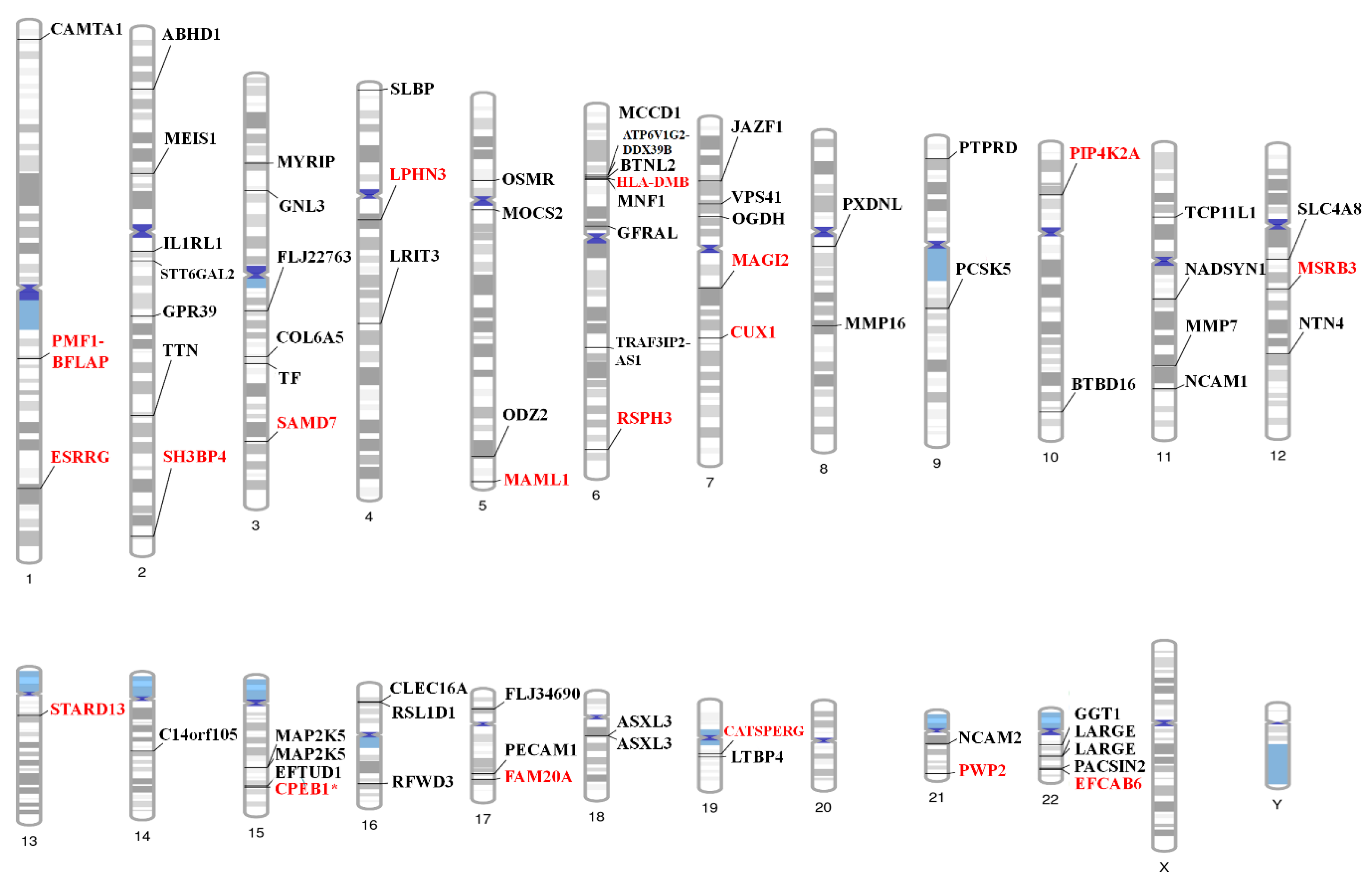
Figure 3.
Protein-Protein Interaction Network.
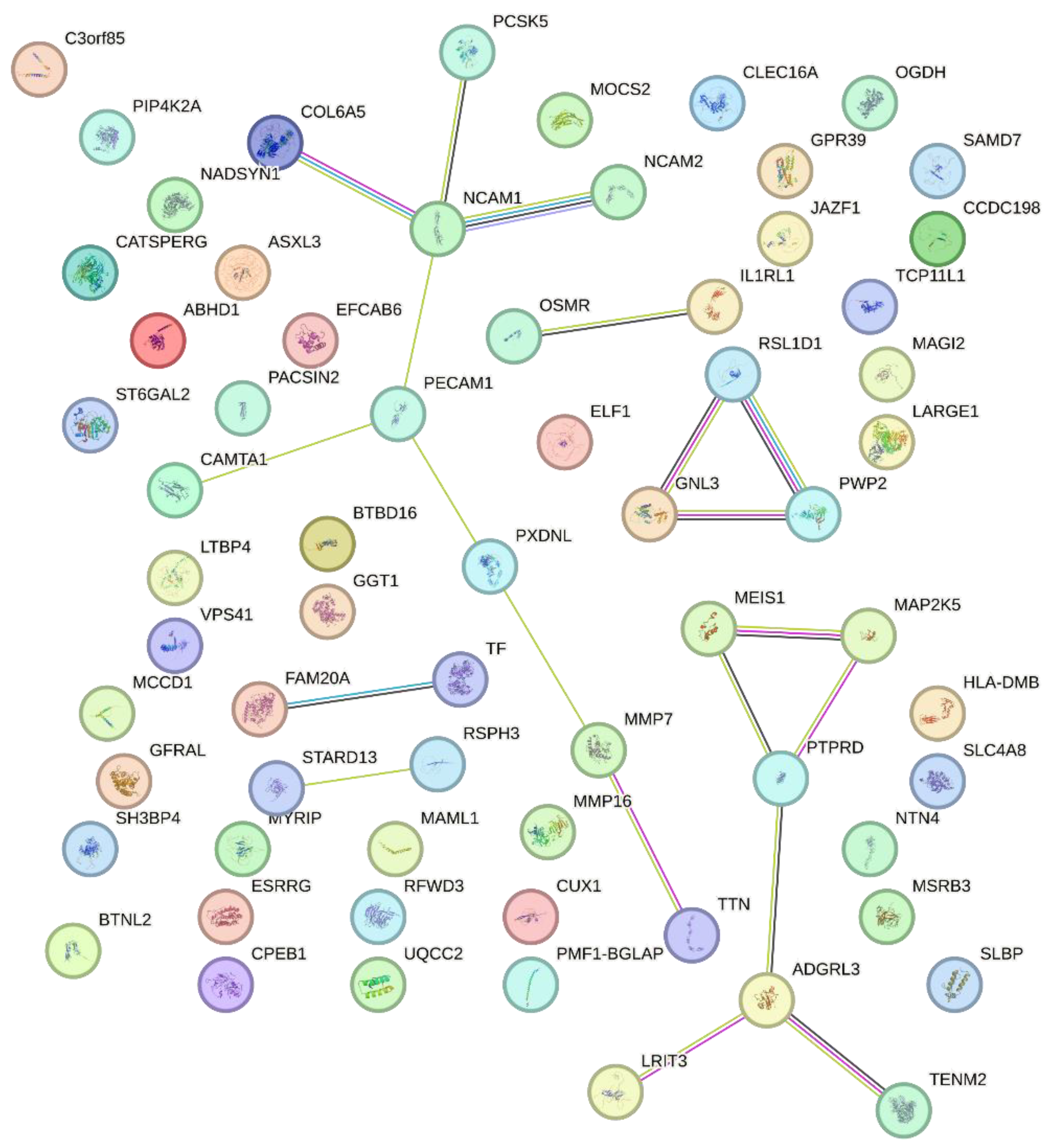
Figure 4.
Flow chart of selection of the study subjects.
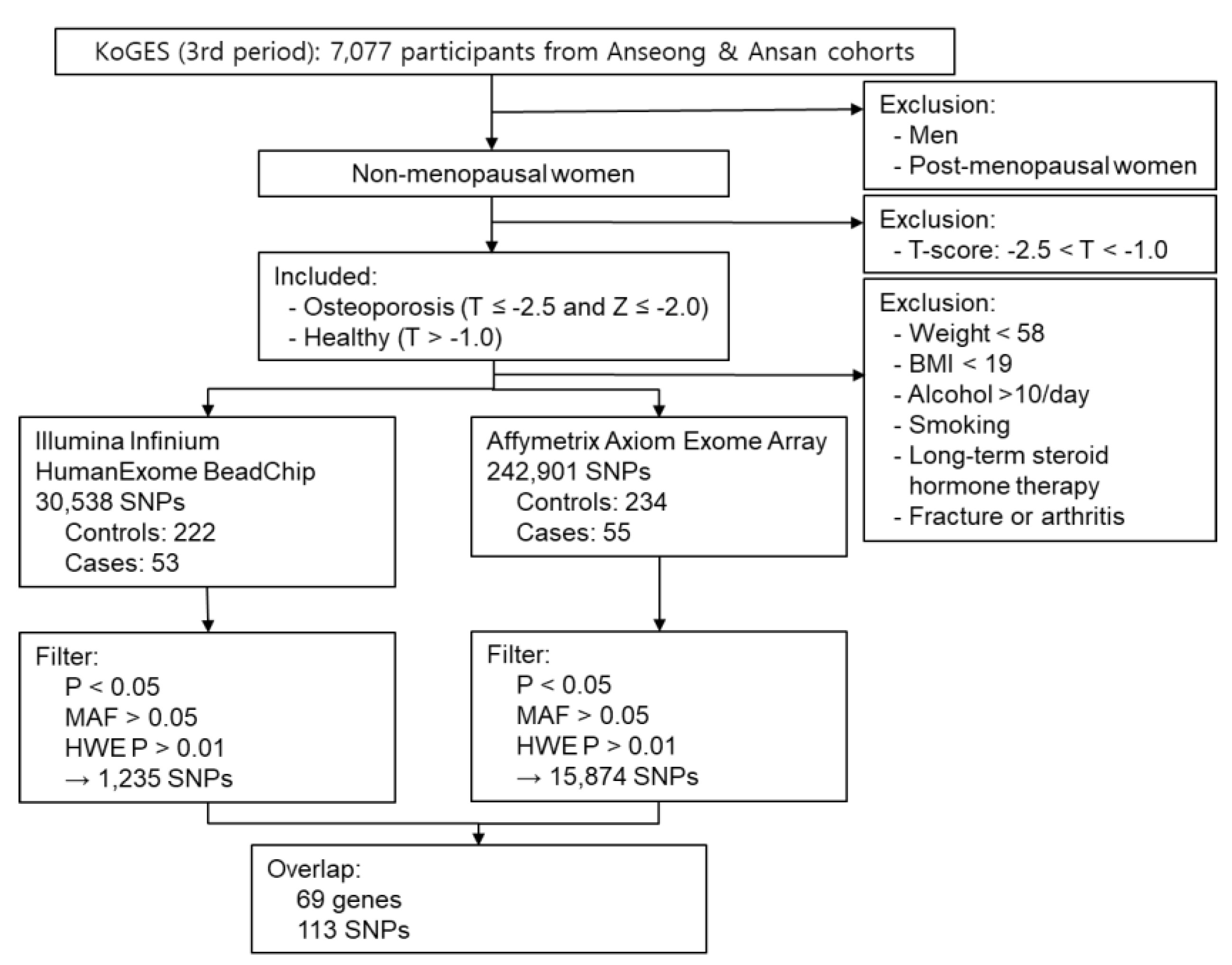

Table 1.
Demographic characteristics of the subjects in the study.
| Control | Osteoporosis | P value | |
|---|---|---|---|
| Age (years) | 47.08 ± 2.57 | 47.54 ± 2.46 | 0.218 |
| Weight (kg) | 63.94 ± 5.13 | 66.62 ± 7.98 | 0.018 |
| BMI (kg/m2) | 25.71 ± 2.35 | 27.23 ± 3.22 | 0.001 |
| Alcohol consumption | 1.06 ± 2.03 | 0.66 ± 1.35 | 0.072 |
| Calcium consumption | 435.88 ± 196.07 | 485.01 ± 205.45 | 0.092 |
| Medical history of fracture | none | none | |
| Medical history of arthritis | none | none | |
| Smoking | none | none | |
| Long term steroid | none | none | |
| Hormone therapy | none | none | |
| DR-SOS | 4269.38 ± 123.94 | 4107.46 ± 152.21 | 0.000* |
| DR-T | 0.8 ± 0.99 | -0.45 ± 1.19 | 0.000* |
| DR-Z | 0.92 ± 1.02 | -0.29 ± 1.22 | 0.000* |
| MT-SOS | 3959.12 ± 65.93 | 3608.74 ± 90.86 | 0.000* |
| MT-T | 0.001 ± 0.63 | -3.4 ± 0.89 | 0.000* |
| MT-Z | 0.2 ± 0.63 | -3.2 ± 0.92 | 0.000* |
BMI: body mass index; DR: distal radius; SOS: speed of sound; MT: mid-shaft tibia. * P < 1.0 x10-12
Table 2.
SNPs that could affect the structure of proteins.
| SNP | Chromosome | Position | Reference Allele | Alternate Allele | Gene | PolyPhen-2 | SIFT | PROVEAN | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| score | prediction | score | prediction | score | prediction | ||||||
| rs1799852 | 3 | 133475722 | C | T | TF | - | - | 0.333 | tolerated | 0.00 | neutral |
| rs11917356 | 3 | 130110550 | A | G | COL6A5 | 0.093 | benign | 0.717 | tolerated | -1.79 | neutral |
| rs2276360 | 11 | 71169547 | G | C | NADSYN1 | 0.000 | benign | 1.000 | tolerated | 2.56 | neutral |
| rs1128431 | 15 | 82456227 | T | C | EFTUD1 | 0.791 | possibly damaging | 0.010 | deleterious | -1.00 | neutral |
| rs7232237 | 18 | 31324934 | A | G | ASXL3 | 0.000 | benign | 0.668 | tolerated | -0.84 | neutral |
| rs2282632 | 18 | 31320229 | A | G | ASXL3 | 0.003 | benign | 0.744 | tolerated | -0.73 | neutral |
Table 3.
A list of the overlapped SNPs associated with osteoporosis between the statistically significant SNPs in Illumina Infinium HumanExome BeadChip (P < 0.01) and Affymetrix Axiom array (P < 0.01) in the Korean pre-menopausal women (logistic regression analysis).
Table 3.
A list of the overlapped SNPs associated with osteoporosis between the statistically significant SNPs in Illumina Infinium HumanExome BeadChip (P < 0.01) and Affymetrix Axiom array (P < 0.01) in the Korean pre-menopausal women (logistic regression analysis).
| SNP | Gene | Chromosome | Position |
P value (exome) |
P value (Affymetrix) |
|---|---|---|---|---|---|
| rs783540 | CPEB1 | 15 | 83254708 | 0.000 | 0.000 |
| rs3731646 | SH3BP4 | 2 | 235950002 | 0.000 | 0.003 |
| rs10506525 | MSRB3 | 12 | 65783378 | 0.001 | 0.003 |
| rs2110871 | MAGI2 | 7 | 78080548 | 0.002 | 0.002 |
| rs2172802 | LPHN3 | 4 | 62453209 | 0.003 | 0.001 |
| rs6895902 | MAML1 | 5 | 179201847 | 0.004 | 0.001 |
| rs2020945 | PWP2 | 21 | 45528919 | 0.004 | 0.003 |
| rs3756987 | RSPH3 | 6 | 159398700 | 0.004 | 0.010 |
| rs2286550 | CATSPERG | 19 | 38861362 | 0.004 | 0.008 |
| rs4729759 | CUX1 | 7 | 101536886 | 0.005 | 0.004 |
| rs10513680 | SAMD7 | 3 | 169644710 | 0.005 | 0.000 |
| rs1052053 | PMF1-BFLAP | 1 | 156202173 | 0.005 | 0.008 |
| rs2764020 | STARD13 | 13 | 34234642 | 0.006 | 0.003 |
| rs7088318 | PIP4K2A | 10 | 22852948 | 0.007 | 0.001 |
| rs151719 | HLA-DMB | 6 | 32903900 | 0.007 | 0.005 |
| rs2302234 | FAM20A | 17 | 66538239 | 0.007 | 0.008 |
| rs16990991 | EFCAB6 | 22 | 44167684 | 0.008 | 0.003 |
| rs12757165 | ESRRG | 1 | 216716537 | 0.009 | 0.003 |
Table 4.
List of the significant genes with possible mechanisms of the development of osteoporosis (P < 0.05).
Table 4.
List of the significant genes with possible mechanisms of the development of osteoporosis (P < 0.05).
| SNP | Chromo-some | Position | Gene | Functional Consequence |
P value (exome) |
P value (Affy-metrix) |
Possible mechanism in the development of osteoporosis |
Function | Reference |
|---|---|---|---|---|---|---|---|---|---|
| rs12757165 | 1 | 216716537 | ESRRG | intron | 0.009 | 0.003 | Bone mineral density, | Determination of bone density | [24] |
| rs1799852 | 3 | 133475722 | TF | synonymous | 0.029 | 0.005 | Osteoclastogenesis | Bone mineral density | [28] |
| rs1436109 | 11 | 112991618 | NCAM1 | intron | 0.012 | 0.001 | Osteogenesis | Osteogenesis | [32] |
| rs4341610 | 12 | 96149288 | NTN4 | intron | 0.029 | 0.027 | To promote osteoblasts and inhibit osteoclast | [26,27] | |
| rs6498142 | 16 | 11081249 | CLECL16A | intron | 0.046 | 0.030 | Bone mineral density | [30] | |
| rs11917356 | 3 | 130110550 | COL6A5 | missense | 0.014 | 0.005 | Variation in bone mineral density | [35] | |
| rs2812 | 17 | 62401118 | PECAM1 | UTR 3' | 0.016 | 0.027 | Negative regulator of Osteoclastogenesis | [31] | |
| rs4820599 | 22 | 24990213 | GGT1 | intron | 0.003 | 0.041 | Osteoclastogenesis | [33] | |
| rs2764020 | 13 | 34234642 | STARD13 | intron | 0.006 | 0.003 | Target of miR-125, which is up-regulated in Osteoporosis | [56] | |
| rs2276360 | 11 | 71169547 | NADSYN1 | missense | 0.038 | 0.027 | Vitamin D | Vitamin D status and metabolic profile | [38,39] |
| rs1128431 | 15 | 82456227 | EFTUD1 | missense | 0.025 | 0.032 | Target gene for vitamin D | [40,57] | |
| rs12757165 | 1 | 216716537 | ESRRG | intron | 0.009 | 0.003 | Skeletal muscle | Skeletal muscle exercise | [44] |
| rs11090122 | 22 | 43308475 | PACSIN2 | intron | 0.045 | 0.028 | Skeletal muscle exercise | [44] | |
| rs12757165 | 1 | 216716537 | ESRRG | intron | 0.009 | 0.003 | Reproductive system | Estrogen pathways | [24,50] |
| rs16990991 | 22 | 44167684 | EFCAB6 | intron | 0.008 | 0.003 | Regulation of androgen receptor | [46] | |
| rs4341610 | 12 | 96149288 | NTN4 | intron | 0.029 | 0.027 | Prognosis of ER-positive breast cancer | [51] | |
| rs783540 | 15 | 83254708 | CPEB1 | intron | 0.000 | 0.000 | Oocyte maturation | [53,54] | |
| rs7232237 | 18 | 31324934 | ASXL3 | missense | 0.015 | 0.011 | Androgen pathway | [49] | |
| rs2282632 | 18 | 31320229 | ASXL3 | missense | 0.019 | 0.038 | Androgen pathway | [49] | |
| rs1128431 | 15 | 82456227 | EFTUD1 | missense | 0.025 | 0.032 | Breast cancer | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



