Submitted:
16 July 2025
Posted:
17 July 2025
You are already at the latest version
Abstract
Allergic contact dermatitis is one of the most common adverse events associated with cosmetic use. Accordingly, assessment of skin sensitisation hazard is required for safety evaluation of cosmetic ingredients. Urged by the European cosmetics legislation ban on animal testing, skin sensitisation has been an intense field in the past two decades, focused on transitioning to the use of alternative methods for hazard prediction. The first alternative methods accepted for regulatory use have been in place for almost a decade, but none as stand-alone replacement for the reference animal method, the murine Local Lymph Node Assay (LLNA). While their strengths and limitations are being evaluated, and the search for new combinations of the adopted methods and other sources of information increase confidence, novel methods based on reconstructed human epidermis (RHE) have recently been validated and joined the available methods. Their unique characteristics provide important advantages that will potentially enhance non-animal testing for skin sensitisation in cosmetics and related industries. In this review, we recapitulate how reconstructed human skin models have been used as platforms for skin sensitisation testing, including the latest approaches using organ-on-a-chip and microfluidic technologies, aimed to develop next-generation organotypic skin models with increased complexity and monitoring capabilities.
Keywords:
skin sensitisation
; in vitro skin model
; allergic contact dermatitis
; adverse outcome pathway
; new alternative methods
1. Introduction
The skin is a barrier organ that is repeatedly exposed to a wide range of chemicals and inducing as a result an adverse outcome such as allergic contact dermatitis (ACD) in sensitized individuals. ACD is a T cell-mediated hypersensitivity reaction specific towards foreign low molecular weight chemical substances that can freely penetrate the skin, resulting in erythema and/or eczema [1,2]. The development of ACD is characterized by two temporally different stages: first the sensitisation or induction phase, characterized by the generation of allergen-specific immunological memory after the initial exposure and secondly, the elicitation of clinical allergy after a subsequent exposure to the allergen [3,4]. In European countries, approximately 20% of the population experiences contact sensitization [5], being a significant proportion caused by ingredients found in cosmetic products such as fragrances, preservatives and dyes [6,7,8].
Skin sensitisation is a term used in regulatory fields to identify the human being health hazards associated with ACD. It is a key factor that affects the cosmetic industry due to its implications in consumer health and safety, product manufacturing and regulatory fulfilment [9,10,11,12]. For this reason, the EU regulation on cosmetics requires the assessment of skin sensitisation hazard for every cosmetic ingredient. The European In-Vitro Toxicology Testing Market Size was valued at €6494.59 million in 2024. Despite this growing market, it is estimated that approximately 32,000 animals are still used annually in Europe for sensitisation testing, representing a total cost of around €9.45 million per year [13,14].
Traditionally, sensitisation tests were performed using in vivo methods, being the Local Lymph Node Assay (LLNA) in mice a reference method [15] that provided valuable and quantitative information for the classification of chemicals. However, the use of animal testing has become progressively controversial. In the European Union (EU), regulatory measures such as EU Cosmetics Regulation 1223/2009 banned the use of animal testing for cosmetic ingredients and finished products, thus forcing the use of alternative non-animal testing strategies. In addition to the EU, which has been a leader in the replacement of animal testing, there is a worldwide trend towards reducing or eliminating the testing of cosmetics on animals. Key global markets have adopted regulatory approaches in this direction, some of them have completely banned the use of animals while others are implementing the use and acceptance of alternative methods following the Replacement, Reduction, and Refinement principle (3Rs). This change in paradigm played a key role in the field of toxicology to move from the traditional use of animal testing for health hazard prediction to the development of approaches based on alternative non-animal methods (NAMs). Skin sensitisation has been an intense field over the last two decades, with joint efforts from governments, academia, and industry in basic research, the development of NAMs, and international acceptance through formal validation procedures [16,17,18]. As a result, several NAMs have been adopted by the Organisation for Economic Co-operation and Development (OECD) within the framework of the Adverse Outcome Pathway (AOP) for skin sensitisation [19]. This conceptual model describes the sensitisation process as a sequence of well-defined Key Events (KEs), each of which is addressed by validated NAMs. These tests provide valuable mechanistic information, however none of them alone can fully predict the sensitisation potential of a substance, due to, for example, the lack of complex crosstalk between the skin and immunological cells. For this reason, researchers and the cosmetic industry are making a great effort to overcome these limitations and improve current methods to provide in vitro models that better recapitulate human skin physiology. As in vitro models with unique characteristics, three-dimensional (3D) reconstructed human skin models have been explored for many years as potential tools for skin sensitisation testing. Research has focused on identifying appropriate cellular markers and endpoints to assess the response of keratinocytes or dendritic cells to sensitisers. However, only recently the first RHE-based test method, EpiSensA, has been officially adopted by the OECD and incorporated into the test guideline No.442D [20].
In summary, the necessity to ensure consumer safety while complying with strict bans on animal testing has triggered substantial advancements in skin sensitisation testing within the cosmetic industry. The shift from conventional in vivo assays to integrated strategies incorporating in chemico methods, in vitro cell-based assays, and organotypic human skin models represent a significant step toward more ethical, reproducible, and physiologically relevant approaches. These innovative methodologies not only align with current regulatory frameworks but also enhance the reliability of sensitisation risk assessment. This review offers a comprehensive overview of advances in skin sensitisation testing employing skin models, encompassing standardized methods based on RHE and approaches that utilize skin models to evaluate keratinocyte or dendritic cell responses. Recent developments in organ-on-a-chip technologies, in silico models, or omics technologies are also covered, as they aim to achieve greater significant biological responses, physiological relevance, or enhanced real-time monitoring capabilities.
2. Skin Sensitisation, the Adverse Outcome Pathway and Alternative Test Methods
2.1. Skin Sensitisation
The skin is the largest organ of the human body, accounting for approximately 15% of total body weight and it performs a broad variety of complex functions. Thanks to its structure composed by three layers (epidermis, dermis and hypodermis) it acts as a mechanical barrier regulating the passage of ions and molecules, limiting water loss and avoiding the entrance of potentially harmful elements from the environment, such as microorganisms or damaging substances [21,22]. In addition to this well-known barrier function role, the skin is an active immune organ. Along with keratinocytes and fibroblasts, which are the most abundant cells found in the epidermis and dermis respectively, the skin also contains resident antigen presenting cells (APC), such as Langerhans cells (LCs) and dendritic cells (DCs). These APC are responsible for capturing and processing foreign antigens and their subsequent presentation to naive T-cells in local lymph nodes, which initiate and regulate the adaptive immune response [23].
Contact sensitisers are low-molecular weight molecules that behave as haptens. They are not immunogenic by themselves but become immunoreactive after binding to self-proteins. Most sensitisers contain reactive electrophilic groups that can form covalent bonds with nucleophilic residues on skin proteins (such as cysteine or lysine side chains) [24]. Some contact sensitisers need abiotic activation (for example, by air oxidation) or metabolic transformation by host enzymes to become immunogenic. These molecules are called pre-haptens and pro-haptens, respectively [25,26]. Repeated topical exposure to sensitisers can trigger an inflammatory response in the skin known as ACD, which has two phases, induction (or afferent) and elicitation (or efferent) phase. The induction phase refers to the first time an allergen contacts the sensitive individual. During this phase, the hapten penetrates the epidermis and it binds to self-proteins. DCs capture and process hapten-protein complexes in the skin and then migrate to the regional lymph nodes, where they present the antigen via the Major Histocompatibility Complex (MHC) to naive T-cells, leading to the proliferation and differentiation of hapten-specific T cells. The elicitation phase starts upon re-exposure to the allergen, which leads to the recruitment and activation of hapten-specific memory T cells, leading to the symptoms of ACD characterized by inflammation, erythema and itching [2].
2.2. The Adverse Outcome Pathway and Alternative Methods
Alternative methods have been developed within the framework of the Adverse Outcome Pathway for skin sensitisation, initiated by covalent binding to proteins (AOP, OECD, 2014) (Figure 1). AOPs were initially developed as conceptual frameworks to represent existing knowledge about the linkage between a particular molecular initiating event (MIE) and a final adverse outcome at the organism or population levels, to aid in ecotoxicology risk assessment [27]. The AOP for skin sensitisation represents the process in 11 sequential steps from the initial penetration of the stratum corneum of the skin to the final adverse outcome manifested as ADC, with four steps in the induction phase defined as key events (KE) (Figure 1).
The first key event (KE1) represents the Molecular Initiating Event (MIE), which is the binding of haptens to self-proteins. As shown in Figure 1, several methods are used to assess this KE1, which are the Amino acid Derivative Reactivity Assay (ADRA), Direct Peptide Reactivity Assay (DPRA) and its modified version named kinetics DPRA (kDPRA). Briefly, these methods are in chemico assays that evaluate the ability of specific chemicals to react with synthetic peptides that mimic the nucleophilic groups of shelf proteins [28]. Consequently, keratinocytes are activated and release proinflammatory molecules such as interleukin 18 (IL-18) and interleukin 1α (IL-1α), which is considered the second key event (KE2). The production of these interleukins may be mediated by the activation of several pathways, including inflammasomes, multiprotein platforms that drive the caspase-dependent maturation and secretion of IL-1β and IL-18. Inflammasomes assembly can be initiated by a variety of damage-associated molecular patterns (DAMPs), extracellular ATP, and other cellular damage associated signals, like reactive oxygen species (ROS) [2,29]
KE2 also comprises changes in gene expression patterns that are linked to specific cell signalling pathways related to cyto-protective responses to oxidative and electrophilic stress, such as the antioxidant/electrophile response element (ARE)-dependent pathways. One of these pathways is the Nfr2/Keap1 pathway. In homeostatic situations, transcription factor Nfr2 is sequestered in the cytoplasm by Keap1 (Kelch-like ECH-associated protein 1), avoiding its function through ubiquitination and degradation in the proteasome. The exposure to haptens promotes their binding to the cysteines of Keap1 causing conformational changes in Keap1 that result in the release of Nfr2. Nfr2 travels to the nucleus and facilitates the transcription of antioxidant response element (ARE)-dependent genes, mostly detoxification enzymes like HMOX1 (hemo-oxygenase 1). While Nrf2 signaling occurs in many cell types, the regulatory assays for KE2 primarily focus on keratinocytes. In vitro methods, such as KeratinoSens and LuSens have been developed and achieved regulatory acceptance for assessing KE2 [20].
In response to cytokines secreted by keratinocytes and the uptake of hapten-protein complexes, APCs like DCs and LCs become activated and maturate. This process represents the KE3. As a result of this activation, APCs secrete proinflammatory molecules and upregulate specific markers (CD54, CD86). Several in vitro methods have been developed to address this activation, which are mainly based on the detection of expression changes on specific transcripts, surface markers (i.e. CD54, CD86), and chemokines and cytokines after the exposure of DCs to potential sensitisers. The OECD validated methods to address DCs activation include the Human Cell Line Activation Test (h-CLAT), the Interleukin-8 Reporter Gene Assay (IL-8 Luc assay) and the Genomic Allergen Rapid Detection (GARD) [30]. Straightaway, activated DCs migrate to lymph nodes, where they present the protein-hapten complexes to naive T cells via MHC molecules, leading to their clonal expansion and proliferation. This process is defined as KE4. The only validated method to assess this KE is the LLNA, which measures the proliferation of T lymphocytes in draining lymph nodes after topical application of the chemical in murine models [31]. It represents a refinement and reduction compared to the previous guinea pig methods (GPMT and Buehler test), which measure the elicitation phase of skin sensitisation (Figure 1). The LLNA also had the advantage of providing a quantitative estimation of the relative potency of contact sensitisers (the EC3 value) and became the reference method to be compared against the newly developed NAMs.
3. Regulatory Landscape and Advancements in Skin Sensitization Testing for Cosmetics
The established relation between ACD and cosmetic product use, with thousands of substances identified as potential skin sensitizers, has made skin sensitization a critical endpoint in cosmetic ingredient safety assessment [9,10,11,32]. Consequently, regulatory frameworks worldwide establish requirements for testing, risk assessment procedures, ingredient labelling, and concentration limits for sensitizing chemicals. These legislations, however, vary considerably across countries and regions [33,34]. While the European Union (EU) enforces a full ban on animal testing for cosmetics, other countries have adopted partial bans or allow testing under specific conditions. For example, the USA has no federal ban, although several states have prohibited the sale of animal-tested cosmetics. Japan supports the incorporation of alternative testing methods, and China officially eliminated animal testing for most cosmetics in May 2021, with some exceptions. In Brazil, several states have banned animal testing despite its allowance at the federal level. In contrast, a growing number of countries have implemented complete bans on animal testing for cosmetics, including the United Kingdom, India, Israel, Norway, New Zealand, Taiwan, South Korea, Switzerland, Guatemala, Australia, Mexico, Colombia, Iceland, Turkey, and Canada.
Within the EU, chemicals are regulated under the Classification, Labelling and Packaging Regulation [35], CLP) and the Registration, Evaluation, Authorisation and Restriction of Chemicals Regulation [36], REACH). For cosmetic-specific chemicals, the Cosmetics Regulations [37,38,39] impose strict controls via annexed lists of prohibited, restricted, and approved substances. To ensure cosmetic ingredient safety, hazard assessments must evaluate skin sensitization potential, potency, ingredient concentrations, and predicted human exposure levels. As animal testing for cosmetics is prohibited under the EU Cosmetics Regulation, alternative in vitro or in silico methods are essential. However, animal data generated under other regulatory frameworks like REACH remain acceptable in certain instances. This regulatory overlap can create a conflict, as REACH obligations for worker safety may still necessitate animal testing if no alternatives exist [40].
Historically, laboratory animals played a significant role in evaluating skin sensitization, with classical methods such as the GPMT and the Buehler Test [41] examining both induction and elicitation phases, and the LLNA [31] focusing on the induction response. The LLNA, which measures T lymphocyte proliferation in draining lymph nodes, has been the benchmark for assessing KE4 of the AOP for decades and was the first method to provide relative potency comparisons using the EC3 value. This value provides information about the concentration of chemicals required to induce a T-cell stimulation index of three compared to controls [42,43]. Although animal methods are not illegal and can be used for justified reasons when alternatives are unavailable, in vitro or in chemico methods are prioritized due to animal welfare concerns. The necessity of reducing animal use led to the design and adoption of in chemico and in vitro alternative test methods addressing the first three key events (KE1, KE2, KE3) of the AOP. Efforts continue to expand and refine these NAMs, with the ultimate goal of replacing the LLNA. Given the complexity of sensitization mechanisms and the limitations of individual NAMs, research has increasingly focused on integrating approaches that combine data from multiple key events to enhance predictive accuracy, particularly for determining sensitizing potency. It has been officially accepted three Defined Approaches (DAs) under OECD Guideline [44] that form part of Integrated Approaches to Testing and Assessment (IATA) for skin sensitization. These DAs combine data from the DPRA, KeratinoSens, and h-CLAT to provide information equivalent or superior to LLNA for hazard identification [45,46]. Furthermore, the Integrated Testing Strategy (ITS)[47,48] and the kDPRA [49] have been accepted by the OECD for discriminating Globally Harmonized System (GHS) potency classes 1A and 1B.
Beyond categorical classification, continuous quantitative data are crucial for determining the highest non-sensitizing dose, which is used to derive a Point of Departure (PoD) for quantitative risk assessment (QRA)[50]. KeratinoSens, h-CLAT, and kDPRA can yield such continuous quantitative data, and various models have been proposed for integrating them [51,52,53]. New methodologies like the GARDskin dose-response approach also show promise in quantitatively assessing sensitizing potency by measuring the GARDskin Genomic Prediction Signature [54] and determining the cDV₀ value, which correlates with LLNA EC3 and human PoDs, supporting its utility for QRA [55]. This test is currently under consideration within the OECD Test Guideline Program. Recent recommendations advocate for data integration strategies that incorporate all available data, including animal data, non-animal data, and in silico data, along with read-across predictions from similar chemicals, to build systematic models for comprehensive decision-making [56]. However, for new ingredients, the data used to train these models often come from NAMs with limitations in their applicability domains, such as challenges in testing hydrophobic ingredients, pre-pro haptens, and complex substances like natural extracts and fragrances [57]. These limitations can significantly constrain reliable risk assessment for new cosmetic products.
Among NAMs, three-dimensional reconstructed human epidermis (RHE) models are particularly noteworthy for their ability to replicate human skin physiology and address solubility restrictions. These models have been explored for skin sensitization testing, evaluating responses in keratinocytes or dendritic cells using various systems and markers. Notably, the EpiSensA, the first RHE-based method, was recently adopted by the OECD and incorporated into Test Guideline KE2 [20]. Subsequent sections will review work performed using skin models to assess skin sensitization responses in both keratinocytes and dendritic cells.
4. In Vitro Skin Models for Skin Sensitisation Testing
4.1. In Vitro Skin Models: Unique Tools for Dermatology Applications
Skin models are organotypic 3D cultures where isolated primary keratinocytes grow and fully differentiate when lifted to the air-liquid interface. They reconstitute the viable basal, spinosum and granular layers, and the non-viable apical multi-layered stratum corneum, closely resembling the native epidermis [58]. Basic skin models are broadly divided into two main types: reconstructed human epidermis (RHE), formed only by keratinocytes grown over organic or artificial supports, and reconstructed human skin (RHS), that have an additional dermal equivalent with fibroblasts embedded generally in a collagen or fibrin matrix. The similarity of skin models to native epidermis in histology, differentiation markers and lipid composition [59,60], and the unique feature of a stratum corneum in contact with the air that allows topical application of substances in a similar way to the in vivo situation, made skin models attractive in vitro tools for cutaneous pharmacology and toxicology applications [61,62,63,64,65,66]. To this end, the first highly standardized commercial RHE models EPISKIN [66], SkinEthics [67] and EpiDerm [68] were produced in the 1990s, and several other RHE and RHS models have been produced throughout the following years [61].
For regulatory toxicology applications, the primary focus of commercial RHE models were initially dermal absorption, phototoxicity and irritation testing [69,70]. RHE models have been successfully applied to build validated methods adopted by the OECD as replacement methods for skin corrosion, irritation and phototoxicity [71,72,73,74]. The establishment of methods using RHE for these tests was facilitated by the good performance of a single readout of cytotoxicity measured with the methyl thiazole tetrazolium (MTT) reduction assay. For skin sensitisation, the search for suitable cells, markers and prediction models to develop in vitro methods took longer. Although most of this research was done with conventional cultures using a variety of cells and cell lines, skin models have also been incorporated through these years as platforms with unique features to model keratinocyte and dendritic cell responses to contact sensitisers.
4.2. RHE-Based Approaches to Evaluate Keratinocyte Response to Sensitisers (KE2)
4.2.1. Markers for the Keratinocyte Response
Studies in murine models of contact hypersensitivity have shown that contact sensitisers and irritants trigger different patterns of cytokine mRNA expression shortly after skin exposure [75], suggesting that the expression of cytokines could be used to identify contact sensitisers [76]. Thus, the analysis of cytokine mRNA or protein expression was the first approach used by researchers to investigate markers in keratinocyte response to sensitisers. For instance, Corsini et al. used the EPISKIN RHE model to demonstrate the selective expression of IL-12 in response to the contact sensitiser dinitrochlorobenzene (DNCB) [77]. This response was not observed after exposure to the irritants sodium lauryl sulphate (SLS) and benzalkonium chloride (BZC). Similarly, Coquette et al. showed that changes in the ratio of IL-8 to IL-1A production enable discrimination among five sensitisers from three irritants using the RHE model SkinEthic [78]. Despite these promising initial results, similar attempts using either RHE models, monolayers of primary keratinocytes, or established cell lines during the same period failed to identify reliable cytokine markers for distinguishing contact sensitisers (reviewed in [79,80]).
In the following years, the urgent demand and increased investment in alternative skin sensitisation testing methods led to the identification of new markers and approaches. The first reliable marker used in keratinocytes to discriminate contact sensitisers was the ubiquitous Keap1/Nrf2 electrophile-sensing pathway [81]. Natsch and Emter found that this pathway was selectively triggered by cysteine-reactive contact sensitisers [82]. Eventually, this finding led to the development of the Keratinosens assay, the first OECD-adopted method for KE2 [83]. This test was based on a stable luciferase reporter introduced into the HaCaT cell line; however, it was not directly transferable to commercial RHE models. Another approach emerged from microarray technology, which allowed global transcriptomic profiling. Studies investigating genome-wide gene expression changes in both DCs [84,85] and keratinocytes [86,87] confirmed Keap1/Nrf2 as a primary pathway activated by sensitisers. This research also demonstrated its potential as a toxicogenomic approach for the development of cell-based test methods. Keratinocyte-derived IL-18 played a key role in allergen-induced LC mobilization in mice [88]. This, along with other approaches, has contributed to the development of RHE models designed to assess the keratinocyte response in skin sensitisation.
4.2.2. The RHE Methods for Keratinocyte Response
A total of five RHE-based methods developed in the past decade were subjected to the formal validation process (Table A1). Some of them had been adapted from methods initially developed in conventional cell cultures to be used on RHE models, while others were created from the start using RHE models.
Epidermal Equivalent (EE) potency and RHE/IL-18 assays were developed within the EU-funded Sens-it-iv research program [89,90]. The EE potency assay [91] originated from an RHE-based method developed for ranking the potency of irritant substances. This method employed the MTT assay as a measure of cytotoxicity to estimate the concentration for a 50 % reduction in cell viability (EC50) from dose-response curves. Based on an existing correlation between the irritant and sensitiser potencies, EE potency was intended to rank the potency of substances already classified as sensitisers. Also within the Sens-it-iv program, Corsini et al. developed an assay l using the NCTC2544 keratinocyte cell line to discriminate contact sensitisers based on the increase in intracellular production of the cytokine IL-18 [92]. Evaluation of secreted rather than intracellular IL-18 in a small subset of substances topically applied to Episkin RHE model also showed good results. The RHE/IL-18 assay was then developed, combining the hazard prediction capability of IL-18 determination with the potency estimation of the EE potency assay into a single method [93]. Studies on the transferability to external laboratories were published for EE potency [90] and the RHE/IL-18 assay [93]. However, these methods did not progress from the initial pre-submission step of the validation process (Table A1). Several prediction models for hazard and potency estimation have been published for the RHE/IL-18 method [94,95].
SensCeeTox method was initially developed using the HaCaT cell line as a two-test platform for the combined assessment of KE1 and KE2 [96]. In this assay, glutathione depletion was evaluated separately as a measure of KE1, and the keratinocyte response was assessed by RT-PCR analysis of gene expression changes in a selected panel of 11 genes under the control of the Keap1/Nrf2, ARNT/AhR, and Nrf1/MTF pathways. Levels of glutathione depletion, gene expression, and cytotoxicity were combined in a proprietary prediction model to provide an In Vitro Toxicity Index (IVTI) for hazard and potency determination. This method was later transferred to the EpiDerm and SkinEthic RHE models, and initial results in the RHE models relative to the HaCaT cell line were published [97]. SensCeeTox was the first RHE method submitted to ECVAM to start the validation process in 2011. Not long after, the results of an inter-laboratory study were published [98], but the approach did not progress further in the validation process.
EpiSensA (Kao Corporation, Japan) was initially developed using the EpiDerm RHE model [99] and later refined to its final version using the LabCyte EPI-MODEL 24 [100] In this assay, expression levels were measured by RT-PCR in a panel of four targets and one control gene. For hazard prediction, the test substance induced fold changes in target genes over vehicle control were calculated, and a positive result was obtained when any of the markers exceeded individually selected cut-off values [100]. Potency classification according to the GHS categories 1A or 1B can be determined using a predefined concentration cut-off, corresponding to the lowest concentration of test substance that yields a positive result. Different cut-offs were initially set for each gene marker [100] and later changed to a single cut-off for all markers [101]. The validation study for EpiSensA was coordinated by JACVAM [102,103], peer-reviewed by an international review panel [104], and the method has recently been adopted by the OECD and included in the test guideline for KE2 [20].
The SENS-IS assay was developed on the Episkin RHE model [105]. In this assay, gene expression levels were measured by RT-PCR in three separate gene panels, REDOX (17 genes), SENS-IS (21 genes), and IRRITATION (23 genes), along with three housekeeping genes. Genes in the REDOX panel were Nrf2/Keap1-dependent with ARE sequences in their promoter regions. The SENS-IS panel comprised non-ARE-containing genes with various functions, including inflammation, regulation of DCs activation and migration, and stress response. The IRRITATION panel contained genes related to inflammation and cell death. For hazard prediction, expression levels over vehicle control were calculated. A positive result was determined if 7 or more genes in the REDOX or SENS-IS panels showed expression above the defined cut-off value. In this assay, doses of 0.1%, 1%, 10%, 50%, and 100%, were used, matching the cut-off values in LLNA for extreme, strong, moderate and weak potency categories, and the same potency categories were assigned for test substances based on the lowest concentration that gives a valid positive result.
The validation study of SENS-IS was conducted by ImmunoSearch, a company specializing in innovative non-animal in vitro testing and human-reconstituted 3D tissue technology [106]. The SENS-IS progression to peer review was delayed until 2020 due to EURL-ECVAM's requests for additional validation data, partly because the assay was patented. The peer-review process was suspended in 2022 by the EURL ECVAM ESAC due to concerns about the quality of data[107]. To move the process on, ImmunoSearch assembled an independent peer-review panel that completed the process in June 2024. [108]. A draft test guideline was then published by the OECD [109].
4.2.3. Performance of EpiSensA and SENS-IS
The results of EpiSensA and SENS-IS on over 100 substances have been documented in the validation study reports as well as in several published works (Table A2). Compared with previously validated NAMs, their overall performance for hazard prediction was similar. However, some published works indicated that their particular characteristics could be advantageous for testing specific sets of substances that are problematic with NAMs based on conventional culture systems. For instance, the performance of EpiSensA for hazard prediction was shown to be better compared to the established methods DPRA, Keratinosens, and hCLAT in a subset of highly lipophilic substances and for detecting pre-/pro-haptens [100,101]. SENS-IS was the only RHE model included in phases 2 [110] and 3 [111] of the Long Range Science Strategy, carried out by Cosmetics Europe for the evaluation of NAMs [112,113]. In the extended substance panel analysed in phase 3 of the study, SENS-IS correctly predicted all pre-/pro-haptens, and contrary to several of the NAMs, its performance was independent of the lipophilicity of the substances [111]. Further studies have demonstrated the applicability of these RHE methods to specific substance groups. For instance, EpiSensA has been used with methacrylates [114,115], while SENS-IS has shown utility for polysiloxanes [116], botanical extracts, and polymers [117,118]. These works supported the wide applicability provided by the use of RHE models. This capacity includes the possibility of testing substances in their intended vehicles or finished formulations. An example has been described using SENS-IS [119], showing how this test method was able to capture vehicle effects and the impact of irritants in the mixture, both known to influence the potency of skin sensitisers.
The ability to provide a measure of relative potency has also been a major challenge for NAMs, still unmet for any single established method except for kDPRA, which was approved to distinguish GHS class 1A substances [28]. The prediction model for classification according to the GHS categories 1A and 1B was analyzed in the peer-review stage of the validation process of EpiSensA, and the conclusion was that the predictive capacity was insufficient due to a substantial rate of underprediction, particularly for class 1A substances [120]. Thus, EpiSensA was approved only for hazard prediction, and the current OECD test guideline for EpiSensA does not include potency determination [121].
The SENS-IS protocol was designed to provide potency categories (extreme, strong, moderate, and weak) according to the LLNA classification [122]. This method showed an impressive initial performance for potency categorization [122]. Subsequent investigations have demonstrated robust correlations with reference data based on the weight of evidence classification in a large dataset of fragrance ingredients [123] and with human reference data in the CE database [110]. The approval of SENS-IS as the first alternative method for potency classification is still pending.
4.2.4. EpiSensA and SENS-IS in the NAM Battery for Skin Sensitisation
Over the past years, considerable efforts have been devoted to the design of strategies termed defined approaches (DA) or integrated Testing Strategies (ITS). These strategies aim to combine the information provided by several NAMs, along with other sources of information, as physicochemical characteristics or QSAR models [47]. One of the main drivers for this work has been the need to provide potency estimations to perform risk assessment [124].
Most DAs and ITSs developed so far have not incorporated EpisSensA or SENS-IS. However, several established DAs have been developed using the new RHE methods. EpiSensA has been combined with the QSAR models, TIMES-SS, and OECD QSAR Toolbox to improve potency predictions, showing similar performance to the three OECD-adopted DAs, but with a higher rate of conclusive predictions [101]. This strategy has been extended through the application of read-across approaches [125,126].
SENS-IS was integrated into a testing strategy as the KE2 method, alongside DPRA (KE1) and hCLAT (KE3) [127]. A Sequential Testing Strategy with SENS-IS as the first method, followed by hCLAT and KeratinoSens was compared with the 2o3 DA on a panel of 25 cosmetic ingredients [128]. This STS was then used to test a panel of botanical extracts [117]. In the near future, EpiSensA and SENS-IS are expected to be gradually incorporated alongside other NAMs in DA/ITS to evaluate their contribution to hazard and potency prediction. Furthermore, they are expected to play a role in the ongoing work to assess their applicability to risk assessment following the NGRA framework [52], although their presence in such assessments has been largely exploratory to date [129,130].
One of the major limitations of EpiSensA and SENS-IS may be their dependence on the availability of a single commercial RHE model. The use of these methods with a different RHE model is foreseen in the OECD test guidelines, assuming that their performance in the new model is demonstrated in a panel of specified performance standards. Recently, SENS-IS was tested in the novel commercial RHE model Skin+ Sterlab, France), showing good concordance with the original results in the panel of reference substances [131].
As an alternative to commercial RHE models, some investigations have explored the performance of RHE models based on keratinocyte cell lines. They are under investigation as more scalable, cost-effective alternatives to primary-cell-based models. The N/TERT keratinocyte cell lines [132] have shown a strong differentiation capacity in a RHE [133]. Alloul-Ramdhani et al. [134] compared the response to the contact sensitiser DNCB between RHEs made with the N/TERT cell line and their in-house RHE (Leiden epidermal model) [135]. Both models showed a similar upregulation of Nrf2-dependent genes. Recently, a new assay for skin sensitisation testing has been proposed based on RHE generated with a patented subclone of the Ker-CT cell line transfected with a stable Keap1/Nrf2 reporter that expresses embryonic secreted alkaline phosphatase [136]. In an initial panel of 10 substances, the results obtained with this method were comparable to KeratinoSens assay. Further testing will show the potential of this new method.
4.3. Immunocompetent Skin Models for KE3
Prior to the extensive development of current NAMs, when Langerhans cells were considered the prime candidates for reliable cell-based in vitro methods to discriminate contact sensitisers, the first immunocompetent skin model was developed by L'Oréal researchers in 1997 [137,138]. In this model, LCs were differentiated in vitro from blood cord-derived CD34+ hematopoietic progenitor cells, co-cultured with primary keratinocytes and melanocytes over a de-epidermized acellular dermis. The keratinocytes formed a RHE with melanocytes in the basal layer and regularly spaced LCs situated suprabasally [137]. Unlike human skin explants, where LCs spontaneously migrated out of the epidermis after short-term culture [139], the LCs notably remained resident for a minimum of four weeks. The same group transferred the model to the Episkin RHE model in later years [140] and studied the response of the LCs to several topically applied contact sensitisers and irritants. Resembling the in vivo response, contact sensitisers, but not irritants, elicited changes in cell morphology and induced overexpression of IL-1β and CD86 mRNAs. This system was sensitive enough to detect weak sensitisers, unlike previous results in conventional DC cultures. However, a significant proportion of donor-derived LCs did not produce a response, a phenomenon already described [141]. Donor variability, together with the limited availability of CD34+ precursors and the complexity of the system, significantly limited the potential of this model for establishing a standardized test method.
In the following years, interest remained in assessing the response of LCs or DCs in the context of their interactions with keratinocytes and fibroblasts [16]. These interactions are crucial for mimicking the immunocompetence of skin, especially for modelling KE2 and KE3 in the AOP for skin sensitisation. Beyond basic in vitro co-culture systems, several laboratories have explored diverse skin model approaches for sensitisation testing, integrating monocyte-derived dendritic cells or cell lines within various system architectures—from LCs or DCs embedded in epidermal/dermal compartments to RHE models placed atop dendritic cell line cultures—and employing a variety of cell response markers, making each model unique (Table A2).
Examples of systems that used monocyte-derived DCs are the works by Chau et al. [142] and Uchino et al. [143]. Both systems shared a similar design, including a dedicated, separate layer of DCs placed in the middle of the epidermal compartment and a fibroblast-populated dermal compartment. Using their model, Uchino et al. tested a small panel of contact sensitisers and irritants, measuring IL-1A, IL-4, and IL-8 secreted into the culture medium. Surprisingly, the cytokine that provided the best discrimination was the Th2-type cytokine IL-4. Production of Th2-type cytokines, including IL-4, has been consistently demonstrated in the draining lymph nodes after the induction of skin sensitisation in mice [144]. However, IL-4 has not been described as one of the cytokines involved in the early response to contact sensitisers in the skin and is not a cytokine normally produced by LCs or DCs [3]. Unfortunately, Uchino et al. could not conclusively demonstrate that the incorporated DCs were the source of IL-4 produced in response to the contact sensitisers.
The MUTZ-3 cell line has been frequently used as an alternative to primary cell-derived LCs (Table A2). Gibbs et al, at the VUCM (The Netherlands), reported the first human RHS model containing LCs derived from a cell line. They incorporated MUTZ-3-derived LCs into their established full-thickness RHS model, composed of a dermal compartment consisting of a collagen gel populated with primary human fibroblasts, and an epidermal compartment made of primary human keratinocytes and melanocytes [145]. After topical exposure to contact sensitisers, MUTZ-3 LCs migrated from the epidermis to the dermal compartment. Interestingly, they later demonstrated that this sensitiser-induced migration was dependent on CXCL12 produced by dermal fibroblasts, while irritant-induced migration relied on CCL5, replicating observations from human skin explants [146]. Although this model was not further developed as a standardized test method, it was more recently used to show an irritant-type migration evoked by titanium salts.
A more simplified model utilizing MUTZ-3-derived LCs was developed by Lee et al. Their system involved placing a RealSkin Human Skin Equivalent (HSE) model (Episkin) over a culture plate containing MUTZ-3 LCs. They investigated the response of the system to topical application of two sensitisers (isoeugenol and p-Phenylenediamine) and one irritant (salicylic acid), measuring a comprehensive panel of secreted cytokines. A support vector machine (SVM) analysis revealed that the system containing MUTZ-3-LCs, unlike the RHS model alone, allowed for the discrimination of the two sensitisers and even distinguished between their different potencies. Nevertheless, the group subsequently continued their work using a simplified co-culture system instead of the RHS model [147].
Similar to the examples mentioned, none of the existing skin models for evaluating the response of LCs or DCs to contact sensitisers have been standardized for testing purposes. The high complexity of immunocompetent skin models, along with the lack of clear added value over recently validated KE3 methods based on submerged cell cultures, has probably discouraged further efforts in this direction. However, the ongoing goal of creating in vitro models that more closely replicate the in vivo counterparts in structure, cell composition, and function will persist. Furthermore, the evolution of knowledge and technology, such as advances in organotypic cultures, rapid prototyping, and microfluidic chip fabrication for organ-on-a-chip, will open additional possibilities. Altogether, these advances will eventually enable the standardization and cost-effective production of complex models that could provide a more precise estimation of the in vivo response to contact sensitisers.
5. Emerging Technologies to Improve Predictability in Skin Sensitisation
Emerging technologies for evaluating skin sensitisation offer innovative alternatives to conventional animal testing. Significant advances include skin-on-a-chip (SoC) technology, omics approaches and in silico methods, each of which provides unique insights into the mechanisms of skin sensitisation and improves predictive accuracy.
Despite considerable progress, no standalone in vitro assay has yet succeeded in covering all four KEs of the AOP, particularly KE4, which involves the activation of T-cells. Among the emerging tools, only SoC models currently offer the potential to integrate the complete sensitisation cascade. Their ability to incorporate immune components and mimic dynamic physiological processes uniquely positions them to simulate all four KEs in a single system, making them a promising platform for comprehensive, mechanistically driven safety assessments.
5.1. Skin-on-a-Chip (SoC)
In recent years, organ-on-a-chip technology has emerged as a promising alternative to animal testing, aligning with the ethical and scientific goals of the 3Rs. Among these platforms, SoC systems have been specifically developed to replicate not only the structural complexity of human skin but also its dynamic physiological processes, such as nutrient and signal molecule exchange, and even aspects of the immune response, crucial for evaluating mechanisms like skin sensitisation [21,148,149] Advanced SoC models typically incorporate multiple skin layers - including the epidermis, dermis, and occasionally a vascularized component [150,151] to mimic systemic drug delivery and immune cell migration. These layers are often separated by porous membranes that support interlayer communication while preserving tissue integrity [152]. This multi-layered architecture enables more accurate modelling of physiological and pathological processes, improving the system’s ability to mimic native human skin. The inclusion of a vascular compartment further enhances the relevance of these models by allowing dynamic perfusion and immune cell trafficking under controlled conditions.
A key advancement in SoC technology is the incorporation of immune cells components, such as dendritic cells or T-lymphocytes, which are essential for studying skin sensitisation pathways. For instance,[153], developed 3D co-culture system that integrates immortalized human keratinocytes (HaCaT) with U937 cells, representing the epidermal barrier and dendritic cells, respectively. This model, implemented within a microfluidic environment, promotes dynamic perfusion that enhances cell viability and tight junction integrity, as evidenced by increased trans-epithelial electrical resistance (TEER) values. The research aims to create an innovative in vitro microphysiological model, including immunology, to understand the mechanisms underlying skin sensitisation. Similar models have demonstrated the ability to replicate immune cell migration and inflammatory responses by dynamic immune responses, providing insights into conditions like atopic dermatitis and psoriasis [154].
In this context, research by [151] demonstrates the power of microfluidic SoC systems to study dynamic immune responses at single-cell resolution. By integrating a vascular microchannel within a full-thickness skin construct, the model allows for immune cell delivery and migration that closely mirrors physiological immune trafficking. Upon keratinocyte-driven inflammation, monocytes migrate from the vascular channel into the dermal and epidermal layers. Single-cell transcriptomics revealed differentiation trajectories of monocytes into mature dermal macrophages, effectively simulating in vivo immune responses. These platforms provide valuable insight into inflammatory skin diseases atopic dermatitis and psoriasis, and they hold potential for studying age-related immune dysfunction and developing targeted therapies.
L'Oréal, in collaboration with HμREL, has developed a pioneering device known as the Allergy Test-on-a-Chip, which aims to replicate human cutaneous allergic reactions more accurately [155]. The system comprises two compartments: one simulating a lymph node and the other representing skin tissue. These are connected by a network of microchannels that maintain a chemical gradient. This gradient enables the observation of immune cell migration, including DCs and T-lymphocytes, from the 'lymph node' to the 'skin', thereby simulating the immune process that occurs during an allergic response. This configuration enables the model to replicate not only allergen exposure, but also the elicitation phase. Consequently, it offers a highly relevant tool with which to assess skin sensitisation in non-animal cosmetic studies [156]. The microfluidic design SoC devices and their ability to replicate key inflammatory processes, such as immune cell recruitment and cytokine release, make them ideal for rapidly identifying potential irritants in cosmetics and pharmaceuticals [157,158]. This, along with precise monitoring of skin barrier integrity through TEER measurements and dye permeation assays, offers valuable insights into allergic reactions and the evaluation of anti-inflammatory compounds [159]. Importantly, this capacity for real-time, dynamic monitoring is one of the most distinctive advantages of SoC systems, allowing researchers to capture transient immune events, track cytokine fluctuations, and assess tissue responses under physiologically relevant conditions.
Despite their growing potential, SoC technologies still face challenges in fully recapitulating the complexity of human skin, particularly in terms of immune system diversity and sensory neuron integration [160]. Standardized protocols and broader regulatory acceptance are also needed to validate and implement SoC systems in allergen testing and pharmaceutical development [161]. Nevertheless, continued advancements in tissue engineering, immunointegration, and high-throughput analysis are steadily improving the physiological relevance and predictive capabilities of these platforms. As these challenges are progressively addressed, SoC systems are poised to become essential tools in the next generation of animal-free safety and efficacy assessments.
5.2. Integration of Omics Approaches for Mechanistic and Predictive Insight
Recent advancements in omics technologies—such as genomics, proteomics, and metabolomics—have significantly enhanced comprehensive characterization of biological responses at the molecular level, providing detailed information on the pathways involved in skin sensitisation. These approaches provide detailed insights into the cellular pathways involved in skin sensitisation and enable the identification of specific biomarkers, thereby contributing to a deeper understanding of inter-individual variability in allergen responses. The integration of omics data with physicochemical parameters has allowed the development of highly predictive multivariate models. These models, which often combine results from validated in vitro assays such as the DPRA, h-CLAT, and KeratinoSens, have achieved predictive accuracies of up to 92% in human skin sensitisation studies [56]. Notably, these integrated approaches have outperformed both individual in vitro assays and the aforementioned traditional in vivo LLNA. Moreover, omics-based analyses have played a pivotal role in identifying key events within AOPs, thereby reinforcing the mechanistic underpinnings of alternative testing strategies. This not only enhances the scientific credibility of non-animal methods but also supports their regulatory acceptance and applicability in risk assessment frameworks [162].
5.3. In Silico Methods
While skin-on-a-chip platforms and omics data offer unprecedented physiological and mechanistic insight, the integration of computational models adds another essential layer of efficiency, scalability, and predictive power to non-animal skin sensitisation testing strategies. In silico tools complement experimental systems by enabling rapid screening of large chemical libraries, help bridge data gaps and support regulatory decisions with reproducible and standardized outputs. These models utilize computational algorithms to simulate biological responses, offering a rapid, ethical and cost-effective alternative to traditional animal-based methods. Machine learning algorithms and Quantitative Structure–Activity Relationship (QSAR) models are among the most widely used in silico approaches [163,164]. By analysing large datasets—including those from historical in vivo tests like the LLNA and GPMT—these models identify correlations between chemical structure and sensitisation potential. Several have demonstrated classification accuracies above 88%, surpassing traditional methods in some cases [163] Platforms like the OECD QSAR Toolbox, which uses read-across and mechanistic alerts, have achieved up to 77% accuracy [56], reinforcing their relevance for regulatory purposes.
Among these, the SkinSensPred platform illustrates the potential of integrating computational models with laboratory-derived data to enhance predictive performance. Using a transfer learning algorithm, SkinSensPred incorporates information from in chemico and in vitro assays—aligned with AOPs—and has shown improved accuracy in both hazard identification and potency classification compared to earlier ITS models [165,166]. The tool is compliant with OECD recommendations and supports regulatory decision-making. However, like other machine learning-based systems, its performance may be influenced by the diversity and quality of training datasets, particularly for less well-represented chemical groups such as agrochemicals [167].
Beyond individual models, machine learning approaches are increasingly employed to develop robust predictive frameworks for skin sensitisation. One such example is the Skin Doctor tool developed by [168], which combines random forest and support vector machine (SVM) classifiers trained on large LLNA datasets. The model achieved accuracies up to 76% and AUC values of 0.83 and notably incorporates a clearly defined applicability domain and reliability indicators for each prediction, enhancing its practical usability in regulatory contexts.
Similarly, the HuSSPred model introduced by [169] represents a significant step forward by basing its predictions on human data from the Human Predictive Patch Test (HPPT). Using Bayesian-optimized QSAR models, HuSSPred demonstrated strong performance across multiple metrics, with correct classification rates ranging from 55% to 88% and high positive predictive values (up to 97%). Its open-access platform offers a user-friendly, ethical alternative for assessing human skin sensitisation potential, further aligning with the 3Rs principles and regulatory needs. A particularly promising approach to improving prediction reliability involves the integration of multiple models through the Dempster-Shafer Theory of evidence. As presented by [163] this method merges predictions from diverse QSAR models trained on assays such as GPMT, LLNA, DPRA, KeratinoSens™ and h-CLAT, resulting in a combined correct classification rate of 88%. The study explored various evidence fusion strategies, with the custom-defined Q3 rule yielding the best balance of performance and coverage (0.893). In addition, the use of SHAP (Shapley Additive Explanations) enhanced interpretability by identifying key molecular features contributing to sensitisation outcomes. This evidence-based framework was ultimately packaged into the HSkinSensDS tool, providing both high predictive power and accessibility for end users.
In silico models play a crucial role in advancing non-animal approaches to skin sensitisation testing by offering fast, cost-effective, and reliable predictions. Tools such as SkinSensPred, Skin Doctor and HuSSPred demonstrate how machine learning and QSAR methods can effectively complement experimental data, while evidence integration strategies like the Dempster-Shafer Theory further enhance prediction robustness.
While these emerging technologies represent a major step forward, challenges remain in terms of validation, standardization, and regulatory acceptance. Successfully integrating these tools into routine safety assessment will require continued interdisciplinary collaboration and refinement to ensure both scientific credibility and practical applicability.
6. Conclusions
In the European Union, the risk assessment of cosmetic ingredients used exclusively in cosmetics must be evaluated without using data from animal testing, as the testing and marketing bans introduced in the Cosmetic Regulation were definitively implemented in 2013, regardless of the availability of alternative methods. The impact on the cosmetic industry is reflected by the fact that no new ingredients of exclusive use in cosmetics have been introduced since then [170]. At the same time, this law provided an important stimulus for accelerating the development of alternative methods, a tendency that is undoubtedly irreversible. Accordingly, regulations worldwide have been increasingly adapting to favour the acceptance of NAM data, incorporating bans on animal testing in most cases [33].
Skin sensitisation has been a paradigmatic example in the development of alternative methods, which have nevertheless faced significant challenges. Limitations in the capacity of established NAMs to determine potency and their applicability to poorly water-soluble substances have been and remain major obstacles. This is evident from the accumulated experience in the use of NAM data for regulatory requirements in risk assessment [171]. Many efforts to overcome these challenges have focused on designing DAs or ITSs that combine information from several NAMs and other sources to maximize predictive capacity and obtain potency estimations. In parallel, new methods have been developed and validated, such as new RHE methods that showed unparalleled capabilities for testing lipophilic substances. Furthermore, due to their capacity to provide an estimation of potency, they could lead to the design of new integrated strategies that need fewer NAM data. An example of this possibility is the similar performance shown by the combination of EpiSensA with QSARs compared to the ITS included in the OECD test guideline for Defined Approaches [125]. Although SENS-IS has not yet been definitively adopted by the OECD, this method has shown unprecedented stand-alone performance in potency prediction. These emerging RHE methods could make a substantial contribution towards the non-animal risk assessment of cosmetic ingredients. The development of immunocompetent in vitro skin models, which integrate immune cells like LCs or DCs into RHE to more closely mimic in vivo skin physiology and immune responses, represents a crucial and ongoing area of research in non-animal sensitisation testing, despite the challenges in standardization encountered by early models.
The continued evolution of organ-on-a-chip and other engineering technologies is poised to further revolutionize the field of in vitro skin models. While still in an incipient stage within the field of organotypic skin models, these approaches are expected to advance steadily. They have great potential to assist the development of more complex skin models by combining the ability to incorporate time-resolved monitoring systems with the possibility of cost-effective, high-throughput fabrication, thus offering even more sophisticated tools for non-animal risk assessment.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors would like to express their gratitude for the financial support received through the First Animal Rights Prize for the Most Innovative Project in Alternative Experimentation 2024, awarded by the Ministry of Social Rights, Consumer Affairs and 2030 Agenda of the Government of Spain. he authors acknowledge the partially financial support for this work received from la Fundación Lair and “Modelos in vitro de piel: desarrollo y aplicaciones” project (Bioengineering Department, 2012/00053/008).This support has contributed to the development of the ideas discussed in this review.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| RHE | Reconstructed Human Epidermis |
| LLNA | Local Lymph Node Assay |
| ACD | Allergic Contact Dermatitis |
| GPMT | Guinea Pig Maximization Test |
| 3Rs | Replacement, Reduction, and Refinement |
| NAMs | New Approach Methods |
| OECD | Organisation for Economic Co-operation and Development |
| AOP | Adverse Outcome Pathway |
| KEs | Key Events |
| 3D | Three-dimensional |
| APC | Antigen Presenting Cells |
| DCs | Dendritic Cells |
| LCs | Langerhans Cells |
| MIE | Molecular Initiating Event |
| ADRA | Amino acid Derivative Reactivity Assay |
| DPRA | Direct Peptide Reactivity Assay |
| kDPRA | kinetics Direct Peptide Reactivity Assay |
| IL | Interleukin |
| DAMPs | Damage-Associated Molecular Patterns |
| ROS | Reactive Oxygen Species |
| ARE | Antioxidant Response Element |
| Keap1 | Kelch-like ECH-associated protein 1 |
| HMOX1 | Heme oxygenase1 |
| h-CLAT | Human Cell Line Activation Test |
| GARD | Genomic Allergen Rapid Detection |
| MHC | Major Histocompatibility Complex |
| CLP | Classification, Labelling and Packaging |
| REACH | Registration, Evaluation, Authorisation and Restriction of Chemicals |
| DAs | Defined Approaches |
| IATA | Integrated Approaches to Testing and Assessment |
| ITS | Integrated Testing Strategy |
| GHS | Globally Harmonized System |
| PoD | Point of Departure |
| QRA | Quantitative Risk Assessment |
| RHS | Reconstructed Human Skin |
| MTT | Methyl Thiazole Tetrazolium |
| DNCB | Dinitrochlorobenzene |
| SLS | Sodium Lauryl Sulphate |
| BZC | Benzalkonium Chloride |
| EE | Epidermal Equivalent |
| IVTI | In Vitro Toxicity Index |
| HSE | Human Skin Equivalent |
| SVM | Support Vector Machine |
| SoC | Skin-on-a-chip |
| HaCaT | Spontaneously Transformed Human Keratinocyte Cell Culture |
| TEER | Trans-epithelial electrical resistance |
| QSAR | Quantitative Structure-Activity Relationship |
| SVM | Support Vector Machine |
| HPPT | Human Predictive Patch Test |
| SHAP | Shapley Additive Explanations |
Appendix A

Table A1.
Summary of RHE models subjected to the formal validation process.
| EE potency assay | RHE-IL--18 | SensCeeTox | Episens A | Sens-IS | |
| Test developer | VUMC (The Netherlands) | Università degli Studi di Mano (Italy) | CeeTox (USA) | Kao Corporation (Japan) | ImmunoSearch (France) |
| RHE models used | EpiCS® (CellSystems) SkinEthic™ (L'Oréal) |
In-house RHE (VUMC-EE), EpiCS® (CellSystems), EpiDerm™ (MatTek), SkinEthic™ (L’oreal) |
SkinEthic™ (L'Oréal); EpiDerm (MatTek) |
LabCyte EPI-MODEL 24 (J-Tec) |
EpiSkin™ (L’oreal) SkinEthic™ (L’oreal) |
| Pre-submission (TSAR ID)* |
to ECVAM in 2011 (TM2011-12) |
To EVCAM in 2012 (TM2012-05) |
to ECVAM in 2011 (TM2011-02) |
to JaCVAM in 2018 (TM2018-01) |
to ECVAM in 2011 (TM2011-11) |
| Formal validation | NO |
NO |
NO | Peer-review completed in 2023 |
Peer-review completed in 2024 |
| OEDC adoption | NO | NO | NO | Test Guideline 442D (june 2024) Test No 442D, 2024 | NO |
| Exposure time | 24 h | 24 h | 24 h | 6 h | 15 min (6h incubation) |
| Read out | 1. Cytotoxicity (MTT) |
1. IL-18 release by keratinocytes 2. Cytotoxicity (MTT) |
1. Glutathione (GSH) Depletion. 2. Gene expression of 7 genes controlled by the Nrf2/Keap1/ARE or AhR/ARNT/XRE signaling pathways: NADPH-quinone oxidoreductase 1 (NQO1) Aldoketoreductase 1C2 (AKR1C2) Interleukin 8 (IL-8) Cytochrome P450 1A1 (CYP1A1) Aldehyde dehydrogenase 3A1 (ALDH3A) Heme-oxygenase 1 (HMOX1) Glutamate cysteine ligase catalytic subunit C (GCLC). 3. Cytotoxicity (LDH) | 1. Gene Expression Analysis activating transcription factor 3 (ATF3); glutamate-cysteine ligase, modifier subunit (GCLM); DnaJ (Hsp40) homolog, subfamily B, member 4 (DNAJB4); and interleukin-8 (IL-8) 2.Cytotoxicity |
1. Gene Expression Analysis of 64 genes biomarkers in 3 groups: skin irritation (23), antioxidant pathways: ARE genes (17) SENS-IS genes (21) and housekeeping (3) Which vehicles are used in Episens A assay |
| Hazard prediction | NO | Several prediction models based on thresholds for IL-18 secretion and viability | Proprietary algorithm with data from GSH depletion, viability and marker gene expression | Positive if any marker gene expressed above individual thresholds values | Positive if expression of 7 or more marker genes in REDOX or SENS-IS panels above threshold value |
| Potency prediction Approach | Concentration for 50% reduction in viability (EC50) interpolated in a regression curve of reference substances | Concentration for 50% reduction in viability (EC50) or stimulation of IL-18 secretion (SI2) interpolated in a regression curve of reference substances. | Proprietary algorithm with data from GSH depletion, viability and gene expression (In Vitro Toxicity Index, IVTI) |
Cut-off value of the lowest positive concentration determines GHS potency categories | Lowest positive concentration determines potency according to LLNA categories |
| References | [90,91,172] | [93,94,95,173,174,175] | [96,176] | [99,100,101,114,115,125,177,178] | [105,110,113,116,117,118,122,123,127,128,179] |
Table A2.
Summary of immune competent skin models for KE3 assessment.
| Immune cells incorporated | Differentiation conditions | Skin equivalent | Immune cell incorporation | Exposure to sensitisers | Read-out | Ref |
| CD34-derived Langerhans cells | CD34+ hematopoietic progenitor cells differentiated to LCs after 6 days in the presence of 200 ng/ml GM-CSF and 2.5 ng/ml TNF-a. | RHE-LCs | a) Co-seeding CD34-derived Langerhans cells with keratinocytes onto the Episkin™ support. b) CD34+ hematopoietic progenitor cells, not exposed to GM-CSF and TNF-a, co-seeded with keratinocytes and melanocytes onto dermal equivalents. |
No | Immunohistology staining, migration of CD1a+, Lag+ cells | [137] |
| CD34-derived Langerhans cells | Differentiated into DCs for 7 days in a medium with 2000 U/ml GM-CSF, 20 U/ml TNF-a, 20 ng/ml SCF. | RHE-LCs | CD34-derived LCs and keratinocytes were co-seeded onto the Episkin™ support. | 24 h topical application or solar simulated radiation. Cytokines: TNF-a and IL-1b. Sensitisers: dinitrofluorobenzene (DNFB), oxazolone, p-phenylenediamine (pPD), NiSO4, eugenol, benzocaine. Irritants: sodium lauryl sulphate, benzalkonium chloride, eugenol. |
Immunohistology staining: loss of dendricity. IL-1b, CD86 mRNA expression by RT-PCR |
[180] |
| Monocyte derived DCs (MoDCs) |
MoDCs were derived from peripheral blood CD14+ cells cultured for 6 days in the presence of, 250 U/ml IL-4 and 50 ng/ml GM-CSF. |
RHS-DCs | Layer of agarose–fibronectin gel containing immature MoDCs placed between a bottom fibroblast containing layer and a top keratinocyte layer | 24 h topical application sensitisers: dinitrochlorobenzene (DNCB). Irritant: sodium dodecyl sulphate (SDS). |
CD86 and HLA-DR expression by flow cytometry. IL-1α, IL-6 and IL-8 secretion by ELISA. |
[142] |
| Monocytes | CD14+ cells differentiated into dendritic cells when incorporated into this 3D skin model | RHS-DCs | For the RHS construct, keratinocytes and freshly isolated CD14+ cells were seeded on a fibrin-based dermal compartment populated by fibroblasts. | 24 h topical application f Sensitisers: Formaldehyde and Manganese (II) Chloride Tetrahydrate (MnCl2·4H2O). Irritant: sodium dodecyl sulphate (SDS) |
Immunohistology staining: Migration of CD1a+, Langerin+ cells. | [181] |
| DCs | Commercial normal human dendritic cells | RHS-DCs | RHS constructs were generated by preparing a collagen vitrigel membrane (VG-KDF-Skin) populated with fibroblasts, followed by normal human dendritic cells in collagen and then keratinocytes seeded on top | 1h topical application Sensitisers: Cobalt chloride (CoCl2), 2,4-dinitrofluorobenzene (DNFB), Formaldehyde (HCHO) and glutaraldehyde (GA), m-amino-phenol (m-AP), cinnamaldehyde (CA), DNCB, α-hexyl cinnamic aldehyde (HCA), isoeugenol (IE) . Non-sensitisers: dimethyl sulfoxide (DMSO), isopropanol (IP), lactic acid (LA), sodium dodecyl sulphate (SDS), Tween 80 |
IL-1α and IL-4 release by ELISA | [143] |
| MUTZ-3-LCs | cells were differentiated into MUTZ-3-LCs for for 7 days in the presence of 100 ng/ml GM-CSF, 10 ng/ml TGF-b and 2.5 ng/ml TNF-a. | RHS-LCs | For RHS generation keratinocytes and MUTZ-3-LCs were seeded on top of a dermal equivalent based on fibroblasts seeded onto a collagen matrix. | No | Immunohistology staining, Langerin+ cells. | [182] |
| MUTZ-3-LCs | MUTZ-3-LCs were derived in the presence of 100 ng/mL GM-CSF, 10 ng/mL, TGF-b, and 2.5 ng/mL TNF-a for 7 days. | RHS-LCs | full-thickness skin equivalent was made by co-culture MUTZ-3--LC with keratinocytes onto fibroblast-populated collagen gels. | 16 h topical application: sensitisers: NiSO4, resorcinol |
Immunohistology staining, migration of CD1a+, Langerin+ cells. IL-1b, CCR7 mRNA expression by RT-PCR. |
[145] |
| MUTZ-3-LCs | MUTZ-3 cells were differentiated into MUTZ-3-LCs for 7 days by treatment with 100 ng/ml GM-CSF, 10 ng/ml TGF-b1 and 2.5 ng/ml TNF-a |
RHS-LCs | SE containing MUTZ-3-LC was achieved by co-seeding CFSE labelled MUTZ-3-LC with Keratinocytes onto fibroblast-populated collagen gels. |
16 h Topical exposure Sensitisers: nickel sulphate, resorcinol, cinnamaldehyde Irritants: Triton X-100, SDS, Tween 80 |
Immunohistology staining and flow cytometry: migration of CD1a+, Langerin+ cells. CD68 mRNA expression by RT-PCR |
[146] |
| MUTZ-3-LCs | Not indicated | Co-culture MUTZ-3-LCs with RHEs | Dermal equivalent with a lattice of collagen and fibroblasts overlaid by a stratified epidermis. RealSkin was used either as a stand-alone assay or co-cultured with MUTZ-3-LCs |
48 h topical exposure: Sensitisers: isoeugenol, and a stron p-phenylenediamine (PPD). Irritant: salicylic acid |
Release of 27 cytokines panel using multiplex bead-based immunoassay. Transwell chemotactic assay to CCL19. |
[183] |
| MUTZ-3-LCs and MoLCsMUTZ-3-LCs | MUTZ-3 cells were differentiated into MUTZ-3-LCs for 10 days by treatment with 10 ng/ml TGF-b1, 100 ng/ml GM-CSF, 2.5 ng/ml TNF-a. MoLCs were obtained after 7 days of monocyte cultivation with 100 ng/mlGM-CSF, 20 ng/ml interleukin IL-4 and 20 ng/ml TGF-b1 MUTZ-3 cells were differentiated into MUTZ-3-LCs for 7 days by treatment with 100 ng/ml GM-CSF, 10 ng/ml TGF-b1 and 2.5 ng/ml TNF-a |
RHS-LCs | Full-thickness skin equivalents prepared by seeding normal human keratinocytes and MUTZ-LCs or MoLCs, respectively, onto the dermal compartment populated with fibroblast on collagen I gel. | 24h topical application: Sensitisers: 2,4-dinitrochlorobenzene (DNCB), isoeugenol. Irritant: sodium dodecyl sulphate (SDS)16 h Topical exposure: Sensitisers: cinnamaldehyde, resorcinol or nickel (II) sulphate hexahydrate (NiSO4) |
Immunohistology staining and flow cytometry: migration CD1a+, Langerin+ cells. IL-6-, IL-8- and IL-18 releases quantified by ELISA ATF3, CD83, CXCR4, IL-1b, PD-L1 mRNA expression by RT-PCR |
[184] |
| MUTZ-3-LCsMUTZ-3-LCs and MoLCs | MUTZ-3 cells were differentiated into MUTZ-3-LCs for 7 days by treatment with 100 ng/ml GM-CSF , 10 ng/ml TGF-b1 and 2.5 ng/ml TNF-a. |
RHS-LCs RHS-LCs/ and MoLCs | RHS-LCs were constructed by preparing a fibroblast populated collagen I hydrogel and coculture Keratinocytes and CFSE-labelled MUTZ-LCs on top of the hydrogel. | 24h topical application: Sensitisers: TiO2, CaO3Ti, C12H28O4Ti, TiALH, nickel sulphate. |
Immunohistology staining CD1a+, Langerin+ cells. CXCL12 vs CCL5-dependent migration of MUTZ-3—LCs. Increase in CD83/CD86 expression by flow cytometry. CXCL8 release quantified by ELISA. IL-1b, CCR7, IL-10 mRNA expression by RT-PCR |
[185] |
| THP-1 MUTZ-3-LCs | THP-1 in RPMI 10% FBS. (Non-differentiated) |
Co-culture of THP-1 with RHE | THP-1 cells were seeded in the basolateral compartment underneath the RHE models (OS-REp, SkinEthic™ RHE).. |
24h topical application. Sensitisers: eugenol, coumarin Irritant: Lactic acid. |
Increase in CD86, CD54, CD40 and HLA-DR expression by flow cytometry | [186] |
| THP1-DCsTHP-1 | THP1 cells were differentiated to DCs for 5 days by treatment with 1500 IU/mL rhGM-CSF and 1500 IU/ml rhIL-4.THP-1 in RPMI 10% FBS. |
RHS-DCs | RHS-DCs were constructed by seeding Keratinocytes with THP-1-derived iDCs onto dermis models based on a solid and porous collagen matrix and primary human foreskin fibroblasts. | 24h topical application: Sensitisers: 1-chloro-2,4-dinitrobenzene (DNCB), nickel sulphate (NiSO4). |
Increase in CD86, CD54, expression by flow cytometry IkBa degradation and phosphorylation of p38 MAPK by western blot. IL-6, IL-8, IL-1b, TNFa and protein secretion by ELISA mRNA expression by RT-PCR |
[187] |
| THP1-DCs and MUTZ-3-LCs |
THP-1 cells were differentiated to DCs for 5 days by treatment with 1500 IU/mL rhGM-CSF and 1500 IU/ml rhIL-4. MUTZ-3 cells were differentiated to LCs for 9 days by treatment with 1000 U/ml rhGM-CSF, 400 U/mL TGF-b and 100 U/ml TNF-a. |
RHS-DCs/LCs | RHS-DCs were constructed by seeding Keratinocytes with MUTZ-3_LCs and THP-1-DCs onto the dermis models. After that freshly detached keratinocytes were seeded on top of the MUTZ-3-LCs +THP1-DCs models. RHS-DCs were constructed by seeding Keratinocytes with THP-1-DCs onto dermis models based on a solid and porous collagen matrix and primary human foreskin fibroblasts. | 6-24h topical application Sensitisers: DNCB, NiSO4 |
Immunohistology staining, migration CD1a+ cells. IkBa degradation, and phosphorylation of p38 MAPK by western blot CD86, CD83, CD54, CXCR4, CCR7, IL-6, IL-8, TNFa, IL-1a IL-1b and IL-12p40 mRNA expression by qPCR.Increase in CD86, CD54, expression by flow cytometry IkBa degradation and phosphorylation of p38 MAPK by western blot. mRNA expression by RT-PCR IL-6, IL-8, IL-1b, TNFa and protein secretion by ELISA |
[188] |
References
- Corsini, E.; Engin, A.B.; Neagu, M.; Galbiati, V.; Nikitovic, D.; Tzanakakis, G.; Tsatsakis, A.M. Chemical-Induced Contact Allergy: From Mechanistic Understanding to Risk Prevention. Arch Toxicol 2018, 92, 3031–3050. [CrossRef]
- Kaplan, D.H.; Igyártó, B.Z.; Gaspari, A.A. Early Events in the Induction of Allergic Contact Dermatitis. Nature reviews. Immunology 2012, 12, 114. [CrossRef]
- Rustemeyer, T. Immunological Mechanisms in Allergic Contact Dermatitis. Curr Treat Options Allergy 2022, 9, 67–75. [CrossRef]
- Yamaguchi, H.; Y, Y.; E, P. Role of Innate Immunity in Allergic Contact Dermatitis: An Update. International journal of molecular sciences 2023, 24. [CrossRef]
- Alinaghi, F.; Bennike, N.H.; Egeberg, A.; Thyssen, J.P.; Johansen, J.D. Prevalence of Contact Allergy in the General Population: A Systematic Review and Meta-Analysis. Contact Dermatitis 2019, 80, 77–85. [CrossRef]
- Park, M.E.; Zippin, J.H. Allergic Contact Dermatitis to Cosmetics. 2014. [CrossRef]
- Diepgen, T.L.; Ofenloch, R.F.; Bruze, M.; Bertuccio, P.; Cazzaniga, S.; Coenraads, P.-J.; Elsner, P.; Goncalo, M.; Svensson, Å.; Naldi, L. Prevalence of Contact Allergy in the General Population in Different European Regions. Br J Dermatol 2016, 174, 319–329. [CrossRef]
- Alani, J.I.; Davis, M.D.P.; Yiannias, J.A. Allergy to Cosmetics: A Literature Review. Dermatitis 2013, 24, 283–290. [CrossRef]
- Uter, W.; Werfel, T.; Lepoittevin, J.-P.; White, I.R. Contact Allergy—Emerging Allergens and Public Health Impact. International Journal of Environmental Research and Public Health 2020, 17, 2404. [CrossRef]
- Martin, S.F. The Role of the Innate Immune System in Allergic Contact Dermatitis*. Allergol Select 2017, 1, 39–43. [CrossRef]
- Uter, W.; Werfel, T.; White, I.R.; Johansen, J.D. Contact Allergy: A Review of Current Problems from a Clinical Perspective. Int J Environ Res Public Health 2018, 15, 1108. [CrossRef]
- de Groot, A.C. Contact Allergy to Cosmetics: Causative Ingredients. Contact Dermatitis 1987, 17, 26–34. [CrossRef]
- Meigs, L.; Smirnova, L.; Rovida, C.; Leist, M.; Hartung, T. Animal Testing and Its Alternatives - the Most Important Omics Is Economics. ALTEX 2018, 35, 275–305. [CrossRef]
- Circabc Available online: https://circabc.europa.eu/ui/group/8ee3c69a-bccb-4f22-89ca-277e35de7c63/library/051e5787-7746-46cf-8a0d-310f84fd1900/details (accessed on 8 July 2025).
- Basketter, D.; Ball, N.; Cagen, S.; Carrillo, J.-C.; Certa, H.; Eigler, D.; Garcia, C.; Esch, H.; Graham, C.; Haux, C.; et al. Application of a Weight of Evidence Approach to Assessing Discordant Sensitisation Datasets: Implications for REACH. Regulatory Toxicology and Pharmacology 2009, 55, 90–96. [CrossRef]
- Aeby, P.; Ashikaga, T.; Bessou-Touya, S.; Schepky, A.; Gerberick, F.; Kern, P.; Marrec-Fairley, M.; Maxwell, G.; Ovigne, J.-M.; Sakaguchi, H.; et al. Identifying and Characterizing Chemical Skin Sensitizers without Animal Testing: Colipa’s Research and Method Development Program. Toxicol In Vitro 2010, 24, 1465–1473. [CrossRef]
- Worth, A.P.; Balls, M. The Principles of Validation and the ECVAM Validation Process. Altern Lab Anim 2004, 32 Suppl 1B, 623–629. [CrossRef]
- Maxwell, G.; MacKay, C.; Cubberley, R.; Davies, M.; Gellatly, N.; Glavin, S.; Gouin, T.; Jacquoilleot, S.; Moore, C.; Pendlington, R.; et al. Applying the Skin Sensitisation Adverse Outcome Pathway (AOP) to Quantitative Risk Assessment. Toxicol In Vitro 2014, 28, 8–12. [CrossRef]
- OECD The Adverse Outcome Pathway for Skin Sensitisation Initiated by Covalent Binding to Proteins; OECD Series on Testing and Assessment; OECD: Paris, France, 2014;
- OECD Test No. 442D: In Vitro Skin Sensitisation: ARE-Nrf2 Luciferase Test Method; OECD Publishing: Paris, 2024;
- Risueño, I.; Valencia, L.; Jorcano, J.L.; Velasco, D. Skin-on-a-Chip Models: General Overview and Future Perspectives. APL Bioengineering 2021, 5, 030901. [CrossRef]
- Footner, E.; Firipis, K.; Liu, E.; Baker, C.; Foley, P.; Kapsa, R.M.I.; Pirogova, E.; O’Connell, C.; Quigley, A. Layer-by-Layer Analysis of In Vitro Skin Models. ACS Biomater Sci Eng 2023, 9, 5933–5952. [CrossRef]
- Kashem, S.W.; Haniffa, M.; Kaplan, D.H. Antigen-Presenting Cells in the Skin. Annu. Rev. Immunol. 2017, 35, 469–499. [CrossRef]
- Goodwin, B.F.; Roberts, D.W. Structure-Activity Relationships in Allergic Contact Dermatitis. Food Chem Toxicol 1986, 24, 795–798. [CrossRef]
- Lepoittevin, J.-P. Metabolism versus Chemical Transformation or Pro- versus Prehaptens? Contact Dermatitis 2006, 54, 73–74. [CrossRef]
- Aptula, A.O.; Roberts, D.W.; Pease, C.K. Haptens, Prohaptens and Prehaptens, or Electrophiles and Proelectrophiles. Contact Dermatitis 2007, 56, 54–56. [CrossRef]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse Outcome Pathways: A Conceptual Framework to Support Ecotoxicology Research and Risk Assessment. Environ Toxicol Chem 2010, 29, 730–741. [CrossRef]
- OECD Test Guideline No. 442C In Chemico Skin Sensitisation_KE1 ASSESSMENT; OECD Guidelines for the Testing of Chemicals, Section 4; OECD: Paris, France, 2023;
- Weber, F.C.; Esser, P.R.; Müller, T.; Ganesan, J.; Pellegatti, P.; Simon, M.M.; Zeiser, R.; Idzko, M.; Jakob, T.; Martin, S.F. Lack of the Purinergic Receptor P2X7 Results in Resistance to Contact Hypersensitivity. J Exp Med 2010, 207, 2609–2619. [CrossRef]
- OECD Test Guideline No. 442E In Vitro Skin Sensitisation_KE3 ASSESSMENT; OECD Guidelines for the Testing of Chemicals, Section 4; OECD: Paris, France, 2023;
- OECD Test No. 429: Skin Sensitisation: Local Lymph Node Assay; OECD Guidelines for the Testing of Chemicals, Section 4; París, 2010;
- de Groot, A.C. Contact Allergy to Cosmetics: Causative Ingredients. Contact Dermatitis 1987, 17, 26–34. [CrossRef]
- Daniel, A.B.; Strickland, J.; Allen, D.; Casati, S.; Zuang, V.; Barroso, J.; Whelan, M.; Régimbald-Krnel, M.J.; Kojima, H.; Nishikawa, A.; et al. International Regulatory Requirements for Skin Sensitization Testing. Regulatory Toxicology and Pharmacology 2018, 95, 52–65. [CrossRef]
- Ferreira, M.; Matos, A.; Couras, A.; Marto, J.; Ribeiro, H. Overview of Cosmetic Regulatory Frameworks around the World. Cosmetics 2022, 9, 72. [CrossRef]
- Regulation (EC) No 1272/2008 on Classification, Labelling and Packaging of Substances and Mixtures (CLP Regulation); 2008; pp. 1–1355;.
- Commission Regulation (EU) 2017/706 of 19 April 2017 Amending Annex VII to Regulation (EC) No 1907/2006 of the European Parliament and of the Council on the Registration, Evaluation, Authorisation and Restriction of Chemicals (REACH) as Regards Skin Sensitisation and Repealing Commission Regulation (EU) 2016/1688 (Text with EEA Relevance. ); 2017; Vol. 104;.
- Council Directive 76/768/EEC of 27 July 1976 on the Approximation of the Laws of the Member States Relating to Cosmetic Products; 1976; Vol. 262;.
- Regulation (EC) No 1907/2006 Concerning the Registration, Evaluation, Authorisation and Restriction of Chemicals (REACH); 2006; pp. 1–849;.
- Commission Regulation (EU) 2023/… of 26 July 2023 Amending Regulation (EC) No 1223/2009 of the European Parliament and of the Council as Regards Labelling of Fragrance Allergens in Cosmetic Products; 2023;
- Knight, J.; Rovida, C.; Kreiling, R.; Zhu, C.; Knudsen, M.; Hartung, T. Continuing Animal Tests on Cosmetic Ingredients for REACH in the EU. ALTEX 2021, 38, 653–668. [CrossRef]
- OECD Test No. 406: Skin Sensitisation; Skin Sensitisation, OECD Guidelines for the Testing of Chemicals; París, 1992;
- Gerberick, G.F.; Ryan, C.A.; Dearman, R.J.; Kimber, I. Local Lymph Node Assay (LLNA) for Detection of Sensitization Capacity of Chemicals. Methods 2007, 41, 54–60. [CrossRef]
- Chayawan; Selvestrel, G.; Baderna, D.; Toma, C.; Caballero Alfonso, A.Y.; Gamba, A.; Benfenati, E. Skin Sensitization Quantitative QSAR Models Based on Mechanistic Structural Alerts. Toxicology 2022, 468, 153111. [CrossRef]
- OECD Test Guideline No. 497: Defined Approaches on Skin Sensitisation; OECD Guidelines for the Testing of Chemicals, Section 4; OECD: Paris, France, 2023;
- Kleinstreuer, N.C.; Hoffmann, S.; Alépée, N.; Allen, D.; Ashikaga, T.; Casey, W.; Clouet, E.; Cluzel, M.; Desprez, B.; Gellatly, N.; et al. Non-Animal Methods to Predict Skin Sensitization (II): An Assessment of Defined Approaches *. Crit Rev Toxicol 2018, 48, 359–374. [CrossRef]
- Assaf Vandecasteele, H.; Gautier, F.; Tourneix, F.; Vliet, E. van; Bury, D.; Alépée, N. Next Generation Risk Assessment for Skin Sensitisation: A Case Study with Propyl Paraben. Regul Toxicol Pharmacol 2021, 123, 104936. [CrossRef]
- OECD Guidance Document on the Reporting of Defined Approaches to Be Used Within Integrated Approaches to Testing and Assessment; OECD Series on Testing and Assessment, No. 256; OECD (Organisation for Economic Co-operation and Development): Paris, 2016;
- Takenouchi, O.; Fukui, S.; Okamoto, K.; Kurotani, S.; Imai, N.; Fujishiro, M.; Kyotani, D.; Kato, Y.; Kasahara, T.; Fujita, M.; et al. Test Battery with the Human Cell Line Activation Test, Direct Peptide Reactivity Assay and DEREK Based on a 139 Chemical Data Set for Predicting Skin Sensitizing Potential and Potency of Chemicals. J Appl Toxicol 2015, 35, 1318–1332. [CrossRef]
- Natsch, A.; Haupt, T.; Wareing, B.; Landsiedel, R.; Kolle, S.N. Predictivity of the Kinetic Direct Peptide Reactivity Assay (kDPRA) for Sensitizer Potency Assessment and GHS Subclassification. ALTEX - Alternatives to animal experimentation 2020, 37, 652–664. [CrossRef]
- Api, A.M.; Basketter, D.A.; Cadby, P.A.; Cano, M.-F.; Ellis, G.; Gerberick, G.F.; Griem, P.; McNamee, P.M.; Ryan, C.A.; Safford, R. Dermal Sensitization Quantitative Risk Assessment (QRA) for Fragrance Ingredients. Regulatory Toxicology and Pharmacology 2008, 52, 3–23. [CrossRef]
- Reynolds, J.; Gilmour, N.; Baltazar, M.T.; Reynolds, G.; Windebank, S.; Maxwell, G. Decision Making in next Generation Risk Assessment for Skin Allergy: Using Historical Clinical Experience to Benchmark Risk. Regul Toxicol Pharmacol 2022, 134, 105219. [CrossRef]
- Gilmour, N.; Kern, P.S.; Alépée, N.; Boislève, F.; Bury, D.; Clouet, E.; Hirota, M.; Hoffmann, S.; Kühnl, J.; Lalko, J.F.; et al. Development of a next Generation Risk Assessment Framework for the Evaluation of Skin Sensitisation of Cosmetic Ingredients. Regul Toxicol Pharmacol 2020, 116, 104721. [CrossRef]
- Natsch, A. Integrated Skin Sensitization Assessment Based on OECD Methods (III): Adding Human Data to the Assessment. ALTEX 2023, 571–583. [CrossRef]
- Zeller, K.S.; Forreryd, A.; Lindberg, T.; Gradin, R.; Chawade, A.; Lindstedt, M. The GARD Platform for Potency Assessment of Skin Sensitizing Chemicals. ALTEX - Alternatives to animal experimentation 2017, 34, 539–559. [CrossRef]
- Gradin, R.; Forreryd, A.; Mattson, U.; Jerre, A.; Johansson, H. Quantitative Assessment of Sensitizing Potency Using a Dose-Response Adaptation of GARDskin. Sci Rep 2021, 11, 18904. [CrossRef]
- Strickland, J.; Zang, Q.; Paris, M.; Lehmann, D.M.; Allen, D.; Choksi, N.; Matheson, J.; Jacobs, A.; Casey, W.; Kleinstreuer, N. Multivariate Models for Prediction of Human Skin Sensitization Hazard. Journal of Applied Toxicology 2017, 37, 347–360. [CrossRef]
- Giménez-Arnau, E. Chemical Compounds Responsible for Skin Allergy to Complex Mixtures: How to Identify Them? Cosmetics 2019, 6, 71. [CrossRef]
- Brohem, C.A.; da Silva Cardeal, L.B.; Tiago, M.; Soengas, M.S.; de Moraes Barros, S.B.; Maria-Engler, S.S. Artificial Skin in Perspective: Concepts and Applications. Pigment Cell Melanoma Res 2011, 24, 35–50. [CrossRef]
- Ponec, M.; Boelsma, E.; Weerheim, A.; Mulder, A.; Bouwstra, J.; Mommaas, M. Lipid and Ultrastructural Characterization of Reconstructed Skin Models. Int J Pharm 2000, 203, 211–225. [CrossRef]
- Ponec, M.; Boelsma, E.; Gibbs, S.; Mommaas, M. Characterization of Reconstructed Skin Models. Skin Pharmacology and Applied Skin Physiology 2004, 15, 4–17. [CrossRef]
- Wang, M.; Zhang, L.; Hao, H.; Yan, M.; Zhu, Z. Applications of Engineered Skin Tissue for Cosmetic Component and Toxicology Detection. Cell Transplant 2024, 33, 9636897241235464. [CrossRef]
- Roguet, R.; Régnier, M.; Cohen, C.; Dossou, K.G.; Rougier, A. The Use of in Vitro Reconstituted Human Skin in Dermotoxicity Testing. Toxicol In Vitro 1994, 8, 635–639. [CrossRef]
- Gay, R.; Swiderek, M.; Nelson, D.; Ernesti, A. The Living Skin Equivalent as a Model in Vitro for Ranking the Toxic Potential of Dermal Irritants. Toxicology in Vitro 1992, 6, 303–315. [CrossRef]
- Ponec, M.; Kempenaar, J. Use of Human Skin Recombinants as an in Vitro Model for Testing the Irritation Potential of Cutaneous Irritants. Skin Pharmacol 1995, 8, 49–59. [CrossRef]
- Bernard, F.X.; Barrault, C.; Deguercy, A.; De Wever, B.; Rosdy, M. Development of a Highly Sensitive in Vitro Phototoxicity Assay Using the SkinEthic Reconstructed Human Epidermis. Cell Biol Toxicol 2000, 16, 391–400. [CrossRef]
- Roguet, R.; Cohen, C.; Robles, C.; Courtellemont, P.; Tolle, M.; Guillot, J.P.; Pouradier Duteil, X. An Interlaboratory Study of the Reproducibility and Relevance of Episkin, a Reconstructed Human Epidermis, in the Assessment of Cosmetics Irritancy. Toxicol In Vitro 1998, 12, 295–304. [CrossRef]
- de Brugerolle de Fraissinette, null; Picarles, V.; Chibout, S.; Kolopp, M.; Medina, J.; Burtin, P.; Ebelin, M.E.; Osborne, S.; Mayer, F.K.; Spake, A.; et al. Predictivity of an in Vitro Model for Acute and Chronic Skin Irritation (SkinEthic) Applied to the Testing of Topical Vehicles. Cell Biol Toxicol 1999, 15, 121–135. [CrossRef]
- Monteiro-Riviere, N.A.; Inman, A.O.; Snider, T.H.; Blank, J.A.; Hobson, D.W. Comparison of an in Vitro Skin Model to Normal Human Skin for Dermatological Research. Microsc Res Tech 1997, 37, 172–179. [CrossRef]
- Netzlaff, F.; Lehr, C.-M.; Wertz, P.W.; Schaefer, U.F. The Human Epidermis Models EpiSkin, SkinEthic and EpiDerm: An Evaluation of Morphology and Their Suitability for Testing Phototoxicity, Irritancy, Corrosivity, and Substance Transport. Eur J Pharm Biopharm 2005, 60, 167–178. [CrossRef]
- Bouwstra, J.A.; Helder, R.W.J.; El Ghalbzouri, A. Human Skin Equivalents: Impaired Barrier Function in Relation to the Lipid and Protein Properties of the Stratum Corneum. Advanced Drug Delivery Reviews 2021, 175, 113802. [CrossRef]
- OECD Test Guideline No. 439: In Vitro Skin Irritation: Reconstructed Human Epidermis Test Methods; OECD Guidelines for the Testing of Chemicals; OECD: Paris, France, 2021;
- OECD Test No. 431: In Vitro Skin Corrosion: Reconstructed Human Epidermis (RHE) Test Method; OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing: Paris, 2019;
- Caloni, F.; De Angelis, I.; Hartung, T. Replacement of Animal Testing by Integrated Approaches to Testing and Assessment (IATA): A Call for in Vivitrosi. Arch Toxicol 2022, 96, 1935–1950. [CrossRef]
- OECD Test No. 498: In Vitro Phototoxicity - Reconstructed Human Epidermis Phototoxicity Test Method; París, 2023;
- Enk, A.H.; Katz, S.I. Early Molecular Events in the Induction Phase of Contact Sensitivity. Proc. Natl. Acad. Sci. U.S.A. 1992, 89, 1398–1402. [CrossRef]
- Müller, G.; Knop, J.; Enk, A.H. Is Cytokine Expression Responsible for Differences between Allergens and Irritants? American Journal of Contact Dermatitis 1996, 7, 177–184. [CrossRef]
- Corsini, E.; Limiroli, E.; Marinovich, M.; Cohen, C.; Roguet, R.; Galli, C.L. Selective Induction of Interleukin-12 in Reconstructed Human Epidermis by Chemical Allergens. Altern Lab Anim 1999, 27, 261–269. [CrossRef]
- Coquette, A.; Berna, N.; Vandenbosch, A.; Rosdy, M.; Poumay, Y. Differential Expression and Release of Cytokines by an in Vitro Reconstructed Human Epidermis Following Exposure to Skin Irritant and Sensitizing Chemicals. Toxicol In Vitro 1999, 13, 867–877. [CrossRef]
- Gerberick, G.F.; Sikorski, E.E. In Vitro and in Vivo Testing Techniques for Allergic Contact Dermatitis. Am J Contact Dermat 1998, 9, 111–118.
- Ryan, C.A.; Hulette, B.C.; Gerberick, G.F. Approaches for the Development of Cell-Based in Vitro Methods for Contact Sensitization. Toxicol In Vitro 2001, 15, 43–55. [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V.; Canning, P. Keap1, the Cysteine-Based Mammalian Intracellular Sensor for Electrophiles and Oxidants. Arch Biochem Biophys 2017, 617, 84–93. [CrossRef]
- Natsch, A.; Emter, R. Skin Sensitizers Induce Antioxidant Response Element Dependent Genes: Application to the in Vitro Testing of the Sensitization Potential of Chemicals. Toxicol Sci 2008, 102, 110–119. [CrossRef]
- Emter, R.; Ellis, G.; Natsch, A. Performance of a Novel Keratinocyte-Based Reporter Cell Line to Screen Skin Sensitizers in Vitro. Toxicol Appl Pharmacol 2010, 245, 281–290. [CrossRef]
- Ryan, C.A.; Gildea, L.A.; Hulette, B.C.; Dearman, R.J.; Kimber, I.; Gerberick, G.F. Gene Expression Changes in Peripheral Blood-Derived Dendritic Cells Following Exposure to a Contact Allergen. Toxicology Letters 2004, 150, 301–316. [CrossRef]
- Gildea, L.A.; Ryan, C.A.; Foertsch, L.M.; Kennedy, J.M.; Dearman, R.J.; Kimber, I.; Gerberick, G.F. Identification of Gene Expression Changes Induced by Chemical Allergens in Dendritic Cells: Opportunities for Skin Sensitization Testing. Journal of Investigative Dermatology 2006, 126, 1813–1822. [CrossRef]
- Van Der Veen, J.W.; Soeteman-Hernández, L.G.; Ezendam, J.; Stierum, R.; Kuper, F.C.; Van Loveren, H. Anchoring Molecular Mechanisms to the Adverse Outcome Pathway for Skin Sensitization: Analysis of Existing Data. Critical Reviews in Toxicology 2014, 44, 590–599. [CrossRef]
- Vandebriel, R.J.; Pennings, J.L.A.; Baken, K.A.; Pronk, T.E.; Boorsma, A.; Gottschalk, R.; Van Loveren, H. Keratinocyte Gene Expression Profiles Discriminate Sensitizing and Irritating Compounds. Toxicol Sci 2010, 117, 81–89. [CrossRef]
- Antonopoulos, C.; Cumberbatch, M.; Mee, J.B.; Dearman, R.J.; Wei, X.-Q.; Liew, F.Y.; Kimber, I.; Groves, R.W. IL-18 Is a Key Proximal Mediator of Contact Hypersensitivity and Allergen-Induced Langerhans Cell Migration in Murine Epidermis. J Leukoc Biol 2008, 83, 361–367. [CrossRef]
- Roggen, E.L. Application of the Acquired Knowledge and Implementation of the Sens-It-Iv Toolbox for Identification and Classification of Skin and Respiratory Sensitizers. Toxicology in Vitro 2013, 27, 1122–1126. [CrossRef]
- Teunis, M.A.T.; Spiekstra, S.W.; Smits, M.; Adriaens, E.; Eltze, T.; Galbiati, V.; Krul, C.; Landsiedel, R.; Pieters, R.; Reinders, J.; et al. International Ring Trial of the Epidermal Equivalent Sensitizer Potency Assay: Reproducibility and Predictive-Capacity. ALTEX 2014, 31, 251–268. [CrossRef]
- dos Santos, G.G.; Spiekstra, S.W.; Sampat-Sardjoepersad, S.C.; Reinders, J.; Scheper, R.J.; Gibbs, S. A Potential in Vitro Epidermal Equivalent Assay to Determine Sensitizer Potency. Toxicol In Vitro 2011, 25, 347–357. [CrossRef]
- Corsini, E.; Galbiati, V.; Mitjans, M.; Galli, C.L.; Marinovich, M. NCTC 2544 and IL-18 Production: A Tool for the Identification of Contact Allergens. Toxicology in Vitro 2013, 27, 1127–1134. [CrossRef]
- Gibbs, S.; Corsini, E.; Spiekstra, S.W.; Galbiati, V.; Fuchs, H.W.; DeGeorge, G.; Troese, M.; Hayden, P.; Deng, W.; Roggen, E. An Epidermal Equivalent Assay for Identification and Ranking Potency of Contact Sensitizers. Toxicology and Applied Pharmacology 2013, 272, 529–541. [CrossRef]
- Galbiati, V.; Papale, A.; Marinovich, M.; Gibbs, S.; Roggen, E.; Corsini, E. Development of an in Vitro Method to Estimate the Sensitization Induction Level of Contact Allergens. Toxicol Lett 2017, 271, 1–11. [CrossRef]
- Andres, E. A New Prediction Model for Distinguishing Skin Sensitisers Based on IL-18 Release from Reconstructed Epidermis: Enhancing the Assessment of a Key Event in the Skin Sensitisation Adverse Outcome Pathway. JDC 2020, 4, 123–137. [CrossRef]
- McKim Jr, J.M.; Keller III, D.J.; Gorski, J.R. An in Vitro Method for Detecting Chemical Sensitization Using Human Reconstructed Skin Models and Its Applicability to Cosmetic, Pharmaceutical, and Medical Device Safety Testing. Cutaneous and Ocular Toxicology 2012, 31, 292–305. [CrossRef]
- McKim, J.M.; Keller, D.J.; Gorski, J.R. A New in Vitro Method for Identifying Chemical Sensitizers Combining Peptide Binding with ARE/EpRE-Mediated Gene Expression in Human Skin Cells. Cutan Ocul Toxicol 2010, 29, 171–192. [CrossRef]
- Clippinger, A.J.; Keller, D.; McKim, J.M.; Witters, H. Inter-Laboratory Validation of an In Vitro Method to Classify Skin Sensitizers. In Proceedings of the PETA Science consortium; 2014.
- Saito, K.; Nukada, Y.; Takenouchi, O.; Miyazawa, M.; Sakaguchi, H.; Nishiyama, N. Development of a New in Vitro Skin Sensitization Assay (Epidermal Sensitization Assay; EpiSensA) Using Reconstructed Human Epidermis. Toxicology in Vitro 2013, 27, 2213–2224. [CrossRef]
- Saito, K.; Takenouchi, O.; Nukada, Y.; Miyazawa, M.; Sakaguchi, H. An in Vitro Skin Sensitization Assay Termed EpiSensA for Broad Sets of Chemicals Including Lipophilic Chemicals and Pre/pro-Haptens. Toxicology in Vitro 2017, 40, 11–25. [CrossRef]
- Mizumachi, H.; Suzuki, S.; Sakuma, M.; Natsui, M.; Imai, N.; Miyazawa, M. Reconstructed Human Epidermis-Based Testing Strategy of Skin Sensitization Potential and Potency Classification Using Epidermal Sensitization Assay and in Silico Data. Journal of Applied Toxicology 2024, 44, 415–427. [CrossRef]
- Epidermal Sensitisation Assay (EpiSensA) Available online: https://tsar.jrc.ec.europa.eu/test-method/tm2018-01-0.
- JaCVAM (Japanese Center for the Validation of Alternative Methods) Epidermal Sensitisation Assay (EpiSensA) Validation Study Report; National Institute of Health Sciences: Japan, 2023;
- OECD Epidermal Sensitisation Assay (EpiSensA) Peer Review Report; Series on Testing and Assesment; OECD Publishing: Paris, France, 2023;
- Cottrez, F.; Boitel, E.; Auriault, C.; Aeby, P.; Groux, H. Genes Specifically Modulated in Sensitized Skins Allow the Detection of Sensitizers in a Reconstructed Human Skin Model. Development of the SENS-IS Assay. Toxicology in Vitro 2015, 29, 787–802. [CrossRef]
- OECD Draft Validation Report of the SENS-IS Assay; Organisation for Economic Co-operation and Development: Paris, France, 2024;
- SENS-IS | EURL ECVAM - TSAR Available online: https://tsar.jrc.ec.europa.eu/test-method/tm2011-11 (accessed on 9 July 2025).
- OECD Peer Review Report of the Epidermal Sensitisation Assay (EpiSensA); Organisation for Economic Co-operation and Development, 2023;
- OECD Draft Test Guideline: Sens-IS Assay; 2024;
- Hoffmann, S.; Kleinstreuer, N.; Alépée, N.; Allen, D.; Api, A.M.; Ashikaga, T.; Clouet, E.; Cluzel, M.; Desprez, B.; Gellatly, N.; et al. Non-Animal Methods to Predict Skin Sensitization (I): The Cosmetics Europe Database<sup/>. Crit Rev Toxicol 2018, 48, 344–358. [CrossRef]
- Hoffmann, S.; Alépée, N.; Gilmour, N.; Kern, P.S.; van Vliet, E.; Boislève, F.; Bury, D.; Cloudet, E.; Klaric, M.; Kühnl, J.; et al. Expansion of the Cosmetics Europe Skin Sensitisation Database with New Substances and PPRA Data. Regul Toxicol Pharmacol 2022, 131, 105169. [CrossRef]
- Desprez, B.; Dent, M.; Keller, D.; Klaric, M.; Ouédraogo, G.; Cubberley, R.; Duplan, H.; Eilstein, J.; Ellison, C.; Grégoire, S.; et al. A Strategy for Systemic Toxicity Assessment Based on Non-Animal Approaches: The Cosmetics Europe Long Range Science Strategy Programme. Toxicology in Vitro 2018, 50, 137–146. [CrossRef]
- Reisinger, K.; Hoffmann, S.; Alépée, N.; Ashikaga, T.; Barroso, J.; Elcombe, C.; Gellatly, N.; Galbiati, V.; Gibbs, S.; Groux, H.; et al. Systematic Evaluation of Non-Animal Test Methods for Skin Sensitisation Safety Assessment. Toxicol In Vitro 2015, 29, 259–270. [CrossRef]
- Mizumachi, H.; LeBaron, M.J.; Settivari, R.S.; Miyazawa, M.; Marty, M.S.; Sakaguchi, H. Characterization of Dermal Sensitization Potential for Industrial or Agricultural Chemicals with EpiSensA. J Appl Toxicol 2021, 41, 915–927. [CrossRef]
- Kimber, I. The Activity of Methacrylate Esters in Skin Sensitisation Test Methods II. A Review of Complementary and Additional Analyses. Regul Toxicol Pharmacol 2021, 119, 104821. [CrossRef]
- Petry, T.; Bosch, A.; Koraïchi-Emeriau, F.; Eigler, D.; Germain, P.; Seidel, S. Assessment of the Skin Sensitisation Hazard of Functional Polysiloxanes and Silanes in the SENS-IS Assay. Regulatory Toxicology and Pharmacology 2018, 98, 209–214. [CrossRef]
- Puginier, M.; Roso, A.; Groux, H.; Gerbeix, C.; Cottrez, F. Strategy to Avoid Skin Sensitization: Application to Botanical Cosmetic Ingredients. Cosmetics 2022, 9, 40. [CrossRef]
- Kolle, S.N.; Flach, M.; Kleber, M.; Basketter, D.A.; Wareing, B.; Mehling, A.; Hareng, L.; Watzek, N.; Bade, S.; Funk-Weyer, D.; et al. Plant Extracts, Polymers and New Approach Methods: Practical Experience with Skin Sensitization Assessment. Regul Toxicol Pharmacol 2023, 138, 105330. [CrossRef]
- Cottrez, F.; Boitel, E.; Berrada-Gomez, M.-P.; Dalhuchyts, H.; Bidan, C.; Rattier, S.; Ferret, P.-J.; Groux, H. In Vitro Measurement of Skin Sensitization Hazard of Mixtures and Finished Products: Results Obtained with the SENS-IS Assays. Toxicology in Vitro 2020, 62, 104644. [CrossRef]
- Reinke, E.N.; Corsini, E.; Ono, A.; Fukuyama, T.; Ashikaga, T.; Gerberick, G.F. Peer Review Report of the EpiSensA Skin Sensitization Assay. 2023.
- OECD Test Guideline No. 442D In Vitro Skin Sensitisation_KE2_ARE-Nrf2 Luciferase Test Method; OECD Guidelines for the Testing of Chemicals, Section 4; OECD: Paris, France, 2022;
- Cottrez, F.; Boitel, E.; Ourlin, J.-C.; Peiffer, J.-L.; Fabre, I.; Henaoui, I.-S.; Mari, B.; Vallauri, A.; Paquet, A.; Barbry, P.; et al. SENS-IS, a 3D Reconstituted Epidermis Based Model for Quantifying Chemical Sensitization Potency: Reproducibility and Predictivity Results from an Inter-Laboratory Study. Toxicology in Vitro 2016, 32, 248–260. [CrossRef]
- Na, M.; O’Brien, D.; Lavelle, M.; Lee, I.; Gerberick, G.F.; Api, A.M. Weight of Evidence Approach for Skin Sensitization Potency Categorization of Fragrance Ingredients. Dermatitis 2022, 33, 161–175. [CrossRef]
- Basketter, D.; Ashikaga, T.; Casati, S.; Hubesch, B.; Jaworska, J.; de Knecht, J.; Landsiedel, R.; Manou, I.; Mehling, A.; Petersohn, D.; et al. Alternatives for Skin Sensitisation: Hazard Identification and Potency Categorisation: Report from an EPAA/CEFIC LRI/Cosmetics Europe Cross Sector Workshop, ECHA Helsinki, April 23rd and 24th 2015. Regulatory Toxicology and Pharmacology 2015, 73, 660–666. [CrossRef]
- Suzuki, S.; Mizumachi, H.; Miyazawa, M. Skin Sensitization Potency Prediction Based on Read-Across (RAx) Incorporating RhE-Based Testing Strategy (RTSv1)-Defined Approach: RTSv1-Based RAx. J Appl Toxicol 2024. [CrossRef]
- Suzuki, S.; Mizumachi, H.; Miyazawa, M. Skin Sensitization Potency Prediction Based on Read-Across (RAx) Incorporating RhE-Based Testing Strategy (RTSv1)-Defined Approach: RTSv1-Based RAx. J Appl Toxicol 2025, 45, 620–635. [CrossRef]
- Clouet, E.; Kerdine-Römer, S.; Ferret, P.-J. Comparison and Validation of an in Vitro Skin Sensitization Strategy Using a Data Set of 33 Chemical References. Toxicol In Vitro 2017, 45, 374–385. [CrossRef]
- Bergal, M.; Puginier, M.; Gerbeix, C.; Groux, H.; Roso, A.; Cottrez, F.; Milius, A. In Vitro Testing Strategy for Assessing the Skin Sensitizing Potential of “Difficult to Test” Cosmetic Ingredients. Toxicology in Vitro 2020, 65, 104781. [CrossRef]
- Gautier, F.; Tourneix, F.; Assaf Vandecasteele, H.; van Vliet, E.; Bury, D.; Alépée, N. Read-across Can Increase Confidence in the Next Generation Risk Assessment for Skin Sensitisation: A Case Study with Resorcinol. Regulatory Toxicology and Pharmacology 2020, 117, 104755. [CrossRef]
- Gilmour, N.; Alépée, N.; Hoffmann, S.; Kern, P.; Vliet, E.V.; Bury, D.; Miyazawa, M.; Nishida, H.; Europe, C. Applying a next Generation Risk Assessment Framework for Skin Sensitisation to Inconsistent New Approach Methodology Information. ALTEX - Alternatives to animal experimentation 2023, 40, 439–451. [CrossRef]
- Cottrez, F.; Boitel, E.; Sahli, E.; Groux, H. Validation of a New 3D Epidermis Model for the SENS-IS Assay to Evaluate Skin Sensitization Potency of Chemicals. Toxicol In Vitro 2025, 106, 106039. [CrossRef]
- Dickson, M.A.; Hahn, W.C.; Ino, Y.; Ronfard, V.; Wu, J.Y.; Weinberg, R.A.; Louis, D.N.; Li, F.P.; Rheinwald, J.G. Human Keratinocytes That Express hTERT and Also Bypass a P16(INK4a)-Enforced Mechanism That Limits Life Span Become Immortal yet Retain Normal Growth and Differentiation Characteristics. Mol Cell Biol 2000, 20, 1436–1447. [CrossRef]
- Smits, J.P.H.; Niehues, H.; Rikken, G.; van Vlijmen-Willems, I.M.J.J.; van de Zande, G.W.H.J.F.; Zeeuwen, P.L.J.M.; Schalkwijk, J.; van den Bogaard, E.H. Immortalized N/TERT Keratinocytes as an Alternative Cell Source in 3D Human Epidermal Models. Sci Rep 2017, 7, 11838. [CrossRef]
- Alloul-Ramdhani, M.; Tensen, C.P.; El Ghalbzouri, A. Performance of the N/TERT Epidermal Model for Skin Sensitizer Identification via Nrf2-Keap1-ARE Pathway Activation. Toxicol In Vitro 2014, 28, 982–989. [CrossRef]
- El Ghalbzouri, A.; Siamari, R.; Willemze, R.; Ponec, M. Leiden Reconstructed Human Epidermal Model as a Tool for the Evaluation of the Skin Corrosion and Irritation Potential According to the ECVAM Guidelines. Toxicol In Vitro 2008, 22, 1311–1320. [CrossRef]
- Brandmair, K.; Dising, D.; Finkelmeier, D.; Schepky, A.; Kuehnl, J.; Ebmeyer, J.; Burger-Kentischer, A. A Novel Three-Dimensional Nrf2 Reporter Epidermis Model for Skin Sensitization Assessment. Toxicology 2024, 503, 153743. [CrossRef]
- Régnier, M.; Staquet, M.J.; Schmitt, D.; Schmidt, R. Integration of Langerhans Cells into a Pigmented Reconstructed Human Epidermis. J Invest Dermatol 1997, 109, 510–512. [CrossRef]
- LeClaire, J.; de Silva, O. Industry Experience with Alternative Methods. Toxicol Lett 1998, 102–103, 575–579. [CrossRef]
- Lukas, M.; Stössel, H.; Hefel, L.; Imamura, S.; Fritsch, P.; Sepp, N.T.; Schuler, G.; Romani, N. Human Cutaneous Dendritic Cells Migrate through Dermal Lymphatic Vessels in a Skin Organ Culture Model. J Invest Dermatol 1996, 106, 1293–1299. [CrossRef]
- Facy, V.; Flouret, V.; Régnier, M.; Schmidt, R. Langerhans Cells Integrated into Human Reconstructed Epidermis Respond to Known Sensitizers and Ultraviolet Exposure. J Invest Dermatol 2004, 122, 552–553. [CrossRef]
- Pichowski, J.S.; Cumberbatch, M.; Dearman, R.J.; Basketter, D.A.; Kimber, I. Allergen-Induced Changes in Interleukin 1 Beta (IL-1 Beta) mRNA Expression by Human Blood-Derived Dendritic Cells: Inter-Individual Differences and Relevance for Sensitization Testing. J Appl Toxicol 2001, 21, 115–121. [CrossRef]
- Chau, D.Y.S.; Johnson, C.; MacNeil, S.; Haycock, J.W.; Ghaemmaghami, A.M. The Development of a 3D Immunocompetent Model of Human Skin. Biofabrication 2013, 5, 035011. [CrossRef]
- Uchino, T.; Takezawa, T.; Ikarashi, Y.; Nishimura, T. Development of an Alternative Test for Skin Sensitization Using a Three-Dimensional Human Skin Model Consisting of Dendritic Cells, Keratinocytes and Fibroblasts. AATEX 2011, 16, 1–8. [CrossRef]
- Xu, H.; DiIulio, N.A.; Fairchild, R.L. T Cell Populations Primed by Hapten Sensitization in Contact Sensitivity Are Distinguished by Polarized Patterns of Cytokine Production: Interferon Gamma-Producing (Tc1) Effector CD8+ T Cells and Interleukin (Il) 4/Il-10-Producing (Th2) Negative Regulatory CD4+ T Cells. J Exp Med 1996, 183, 1001–1012. [CrossRef]
- Ouwehand, K.; Spiekstra, S.W.; Waaijman, T.; Scheper, R.J.; de Gruijl, T.D.; Gibbs, S. Technical Advance: Langerhans Cells Derived from a Human Cell Line in a Full-Thickness Skin Equivalent Undergo Allergen-Induced Maturation and Migration. Journal of Leukocyte Biology 2011, 90, 1027–1033. [CrossRef]
- Kosten, I.J.; Spiekstra, S.W.; de Gruijl, T.D.; Gibbs, S. MUTZ-3 Derived Langerhans Cells in Human Skin Equivalents Show Differential Migration and Phenotypic Plasticity after Allergen or Irritant Exposure. Toxicol Appl Pharmacol 2015, 287, 35–42. [CrossRef]
- Lee, S.; Greenstein, T.; Shi, L.; Maguire, T.; Schloss, R.; Yarmush, M. Tri-Culture System for pro-Hapten Sensitizer Identification and Potency Classification. Technology (Singap World Sci) 2018, 6, 67–74. [CrossRef]
- Zoio, P.; Oliva, A. Skin-on-a-Chip Technology: Microengineering Physiologically Relevant In Vitro Skin Models. Pharmaceutics 2022, 14, 682. [CrossRef]
- Fernandez-Carro, E.; Angenent, M.; Gracia-Cazaña, T.; Gilaberte, Y.; Alcaine, C.; Ciriza, J. Modeling an Optimal 3D Skin-on-Chip within Microfluidic Devices for Pharmacological Studies. Pharmaceutics 2022, 14, 1417. [CrossRef]
- Jones, C.F.E.; Di Cio, S.; Connelly, J.T.; Gautrot, J.E. Design of an Integrated Microvascularized Human Skin-on-a-Chip Tissue Equivalent Model. Front. Bioeng. Biotechnol. 2022, 10. [CrossRef]
- Hindle, S.A.; Bachas Brook, H.; Chrysanthou, A.; Chambers, E.S.; Caley, M.P.; Connelly, J.T. Replicating Dynamic Immune Responses at Single-Cell Resolution within a Microfluidic Human Skin Equivalent. Adv Sci (Weinh) 2025, 12, e2415717. [CrossRef]
- Wufuer, M.; Lee, G.; Hur, W.; Jeon, B.; Kim, B.J.; Choi, T.H.; Lee, S. Skin-on-a-Chip Model Simulating Inflammation, Edema and Drug-Based Treatment. Sci Rep 2016, 6, 37471. [CrossRef]
- Ramadan, Q.; Ting, F.C.W. In Vitro Micro-Physiological Immune-Competent Model of the Human Skin. Lab Chip 2016, 16, 1899–1908. [CrossRef]
- Moon, S.; Kim, D.H.; Shin, J.U. In Vitro Models Mimicking Immune Response in the Skin. Yonsei Med J 2021, 62, 969–980. [CrossRef]
- Yarmush, M.L.; Freedman, R.; Bufalo, A.D.; Teissier, S.; Meunier, J.-R. Immune System Modeling Devices and Methods 2017.
- Zhang, Q.; Sito, L.; Mao, M.; He, J.; Zhang, Y.S.; Zhao, X. Current Advances in Skin-on-a-Chip Models for Drug Testing. Microphysiological Systems 2018, 2. [CrossRef]
- Ren, X.; Getschman, A.E.; Hwang, S.; Volkman, B.F.; Klonisch, T.; Levin, D.; Zhao, M.; Santos, S.; Liu, S.; Cheng, J.; et al. Investigations on T Cell Transmigration in a Human Skin-on-Chip (SoC) Model. Lab Chip 2021, 21, 1527–1539. [CrossRef]
- Quan, Q.; Weng, D.; Li, X.; An, Q.; Yang, Y.; Yu, B.; Ma, Y.; Wang, J. Analysis of Drug Efficacy for Inflammatory Skin on an Organ-Chip System. Frontiers in Bioengineering and Biotechnology 2022, 10.
- Sasaki, N.; Tsuchiya, K.; Kobayashi, H. Photolithography-free Skin-on-a-chip for Parallel Permeation Assays. Sensors and Materials 2019, 31, 107–115. [CrossRef]
- Cho, S.W.; Malick, H.; Kim, S.J.; Grattoni, A. Advances in Skin-on-a-Chip Technologies for Dermatological Disease Modeling. J Invest Dermatol 2024, 144, 1707–1715. [CrossRef]
- Govey-Scotland, J.; Johnstone, L.; Myant, C.; Friddin, M.S. Towards Skin-on-a-Chip for Screening the Dermal Absorption of Cosmetics. Lab Chip 2023, 23, 5068–5080. [CrossRef]
- Ta, G.H.; Weng, C.-F.; Leong, M.K. In Silico Prediction of Skin Sensitization: Quo Vadis? Front Pharmacol 2021, 12, 655771. [CrossRef]
- Wang, H.; Huang, Z.; Lou, S.; Li, W.; Liu, G.; Tang, Y. In Silico Prediction of Skin Sensitization for Compounds via Flexible Evidence Combination Based on Machine Learning and Dempster–Shafer Theory. Chem. Res. Toxicol. 2024, 37, 894–909. [CrossRef]
- Wilm, A.; Kühnl, J.; Kirchmair, J. Computational Approaches for Skin Sensitization Prediction. Critical Reviews in Toxicology 2018, 48, 738–760. [CrossRef]
- Wang, S.-S.; Wang, C.-C.; Tung, C.-W. SkinSensPred as a Promising in Silico Tool for Integrated Testing Strategy on Skin Sensitization. International Journal of Environmental Research and Public Health 2022, 19, 12856. [CrossRef]
- Kan, H.-L.; Wang, S.-S.; Liao, C.-L.; Tsai, W.-R.; Wang, C.-C.; Tung, C.-W. An Integrated Testing Strategy and Online Tool for Assessing Skin Sensitization of Agrochemical Formulations. Toxics 2024, 12, 936. [CrossRef]
- Asai, T.; Umeshita, K.; Sakurai, M.; Sakane, S. Development of an in Silico Evaluation System That Quantitatively Predicts Skin Sensitization Using OECD Guideline No. 497 ITSv2 Defined Approach for Skin Sensitization Classification. Food and Chemical Toxicology 2024, 185, 114444. [CrossRef]
- Wilm, A.; Stork, C.; Bauer, C.; Schepky, A.; Kühnl, J.; Kirchmair, J. Skin Doctor: Machine Learning Models for Skin Sensitization Prediction That Provide Estimates and Indicators of Prediction Reliability. International Journal of Molecular Sciences 2019, 20, 4833. [CrossRef]
- Tieghi, R.S.; Moreira-Filho, J.T.; Martin, H.-J.; Wellnitz, J.; Otoch, M.C.; Rath, M.; Tropsha, A.; Muratov, E.N.; Kleinstreuer, N. A Novel Machine Learning Model and a Web Portal for Predicting the Human Skin Sensitization Effects of Chemical Agents. Toxics 2024, 12, 803. [CrossRef]
- Rogiers, V.; Benfenati, E.; Bernauer, U.; Bodin, L.; Carmichael, P.; Chaudhry, Q.; Coenraads, P.J.; Cronin, M.T.D.; Dent, M.; Dusinska, M.; et al. The Way Forward for Assessing the Human Health Safety of Cosmetics in the EU - Workshop Proceedings. Toxicology 2020, 436, 152421. [CrossRef]
- Basketter, D.; Safford, B. Skin Sensitization Quantitative Risk Assessment: A Review of Underlying Assumptions. Regul Toxicol Pharmacol 2016, 74, 105–116. [CrossRef]
- Teunis, M.; Corsini, E.; Smits, M.; Madsen, C.B.; Eltze, T.; Ezendam, J.; Galbiati, V.; Gremmer, E.; Krul, C.; Landin, A.; et al. Transfer of a Two-Tiered Keratinocyte Assay: IL-18 Production by NCTC2544 to Determine the Skin Sensitizing Capacity and Epidermal Equivalent Assay to Determine Sensitizer Potency. Toxicology in Vitro 2013, 27, 1135–1150. [CrossRef]
- Corsini, E.; Gibbs, S.; Roggen, E.; Kimber, I.; Basketter, D.A. Skin Sensitization Tests: The LLNA and the RhE IL-18 Potency Assay. Methods Mol Biol 2021, 2240, 13–29. [CrossRef]
- Andres, E.; Barry, M.; Hundt, A.; Dini, C.; Corsini, E.; Gibbs, S.; Roggen, E.L.; Ferret, P.-J. Preliminary Performance Data of the RHE/IL-18 Assay Performed on SkinEthicTM RHE for the Identification of Contact Sensitizers. Int J Cosmet Sci 2017, 39, 121–132. [CrossRef]
- de Souza, I.R.; Iulini, M.; Galbiati, V.; Silva, E.Z.M.; Sivek, T.W.; Rodrigues, A.C.; Gradia, D.F.; Pestana, C.B.; Leme, D.M.; Corsini, E. An Integrated in Silico-in Vitro Investigation to Assess the Skin Sensitization Potential of 4-Octylphenol. Toxicology 2023, 493, 153548. [CrossRef]
- Mehling, A.; Adriaens, E.; Casati, S.; Hubesch, B.; Irizar, A.; Klaric, M.; Letasiova, S.; Manou, I.; Müller, B.P.; Roggen, E.; et al. In Vitro RHE Skin Sensitisation Assays: Applicability to Challenging Substances. Regul Toxicol Pharmacol 2019, 108, 104473. [CrossRef]
- Reinke, E. N; Hines, R. N; Strickland, J; Casey, W. Integrated Approaches for Skin Sensitization: Regulatory Status and Future Opportunities.; NTP / ICCVAM: Nashville, TN, USA, 2023.
- Kasahara, T.; Yamamoto, Y.; Nakashima, N.; Imamura, M.; Mizumachi, H.; Suzuki, S.; Aiba, S.; Kimura, Y.; Ashikaga, T.; Kojima, H.; et al. Borderline Range Determined Using Data From Validation Study of Alternative Methods for Skin Sensitization: ADRA, IL-8 Luc Assay, and EpiSensA. J Appl Toxicol 2025, 45, 432–439. [CrossRef]
- Pellevoisin, C.; Cottrez, F.; Johansson, J.; Pedersen, E.; Coleman, K.; Groux, H. Pre-Validation of SENS-IS Assay for in Vitro Skin Sensitization of Medical Devices. Toxicol In Vitro 2021, 71, 105068. [CrossRef]
- Facy, V.; Flouret, V.; Régnier, M.; Schmidt, R. Reactivity of Langerhans Cells in Human Reconstructed Epidermis to Known Allergens and UV Radiation. Toxicology in Vitro 2005, 19, 787–795. [CrossRef]
- Gaviria Agudelo, C. Modelling Human Skin: Organotypic Cultures for Applications in Toxicology, Immunity, and Cancer:, University of Groningen, 2022.
- Laubach, V.; Zöller, N.; Rossberg, M.; Görg, K.; Kippenberger, S.; Bereiter-Hahn, J.; Kaufmann, R.; Bernd, A. Integration of Langerhans-like Cells into a Human Skin Equivalent. Arch Dermatol Res 2011, 303, 135–139. [CrossRef]
- Lee, S.; Dong, D.X.; Jindal, R.; Maguire, T.; Mitra, B.; Schloss, R.; Yarmush, M. Predicting Full Thickness Skin Sensitization Using a Support Vector Machine. Toxicol In Vitro 2014, 28, 1413–1423. [CrossRef]
- Bock, S.; Said, A.; Müller, G.; Schäfer-Korting, M.; Zoschke, C.; Weindl, G. Characterization of Reconstructed Human Skin Containing Langerhans Cells to Monitor Molecular Events in Skin Sensitization. Toxicol In Vitro 2018, 46, 77–85. [CrossRef]
- Rodrigues Neves, C.; Gibbs, S. Progress on Reconstructed Human Skin Models for Allergy Research and Identifying Contact Sensitizers. In Three Dimensional Human Organotypic Models for Biomedical Research; Bagnoli, F., Rappuoli, R., Eds.; Current Topics in Microbiology and Immunology; Springer International Publishing: Cham, 2021; pp. 103–129 ISBN 978-3-030-62452-1.
- Schellenberger, M.T.; Bock, U.; Hennen, J.; Groeber-Becker, F.; Walles, H.; Blömeke, B. A Coculture System Composed of THP-1 Cells and 3D Reconstructed Human Epidermis to Assess Activation of Dendritic Cells by Sensitizing Chemicals after Topical Exposure. Toxicology in Vitro 2019, 57, 62–66. [CrossRef]
- Hölken, J.M.; Friedrich, K.; Merkel, M.; Blasius, N.; Engels, U.; Buhl, T.; Mewes, K.R.; Vierkotten, L.; Teusch, N.E. A Human 3D Immune Competent Full-Thickness Skin Model Mimicking Dermal Dendritic Cell Activation. Front. Immunol. 2023, 14. [CrossRef]
- Hölken, J.M.; Wurz, A.-L.; Friedrich, K.; Böttcher, P.; Asskali, D.; Stark, H.; Breitkreutz, J.; Buhl, T.; Vierkotten, L.; Mewes, K.R.; et al. Incorporating Immune Cell Surrogates into a Full-Thickness Tissue Equivalent of Human Skin to Characterize Dendritic Cell Activation. Sci Rep 2024, 14, 30158. [CrossRef]
Figure 1.
Skin sensitisation AOP and validated assays. (a) During the afferent phase, haptens penetrate through the stratum corneum of the epidermis and bind covalently to nucleophilic residues of self-proteins (e. g. cysteines and lysines) becoming immune-reactive (darker colours) (KE1) (pre-haptens require abiotic activation, and pro-haptens are metabolically transformed in the epidermis). (b) Hapten exposure leads to activation of keratinocytes, which respond to the chemical stress by releasing pro-inflammatory cytokines such as IL-1α and IL-18 (KE2). These danger signals activate resident antigen-presenting cells (APCs), including Langerhans cells and dermal dendritic cells, which internalize and process hapten–protein complexes. (c) As a result, APCs get activated and mature (KE3), secreting proinflammatory molecules and upregulating specific markers (CD54, CD86). APCs then migrate to lymph nodes through lymphatic vessels, where they present the hapten-peptide complexes via Major Histocompatibility Complex to specific naive T cells (d) Naive T cells undergo clonal expansion and differentiate into effector and memory T cells (KE4). Re-exposure to the hapten triggers the activation of memory T cells, cytokine release, recruitment of inflammatory cells, and the clinical manifestations of allergic contact dermatitis (ACD), such as erythema, itching, and eczema. At the bottom, validated NAMs methods (in black) to measure each KE and the number of the OECD Test Guidelines. In red, in vivo methods. (Image created in https://BioRender.com).
Figure 1.
Skin sensitisation AOP and validated assays. (a) During the afferent phase, haptens penetrate through the stratum corneum of the epidermis and bind covalently to nucleophilic residues of self-proteins (e. g. cysteines and lysines) becoming immune-reactive (darker colours) (KE1) (pre-haptens require abiotic activation, and pro-haptens are metabolically transformed in the epidermis). (b) Hapten exposure leads to activation of keratinocytes, which respond to the chemical stress by releasing pro-inflammatory cytokines such as IL-1α and IL-18 (KE2). These danger signals activate resident antigen-presenting cells (APCs), including Langerhans cells and dermal dendritic cells, which internalize and process hapten–protein complexes. (c) As a result, APCs get activated and mature (KE3), secreting proinflammatory molecules and upregulating specific markers (CD54, CD86). APCs then migrate to lymph nodes through lymphatic vessels, where they present the hapten-peptide complexes via Major Histocompatibility Complex to specific naive T cells (d) Naive T cells undergo clonal expansion and differentiate into effector and memory T cells (KE4). Re-exposure to the hapten triggers the activation of memory T cells, cytokine release, recruitment of inflammatory cells, and the clinical manifestations of allergic contact dermatitis (ACD), such as erythema, itching, and eczema. At the bottom, validated NAMs methods (in black) to measure each KE and the number of the OECD Test Guidelines. In red, in vivo methods. (Image created in https://BioRender.com).
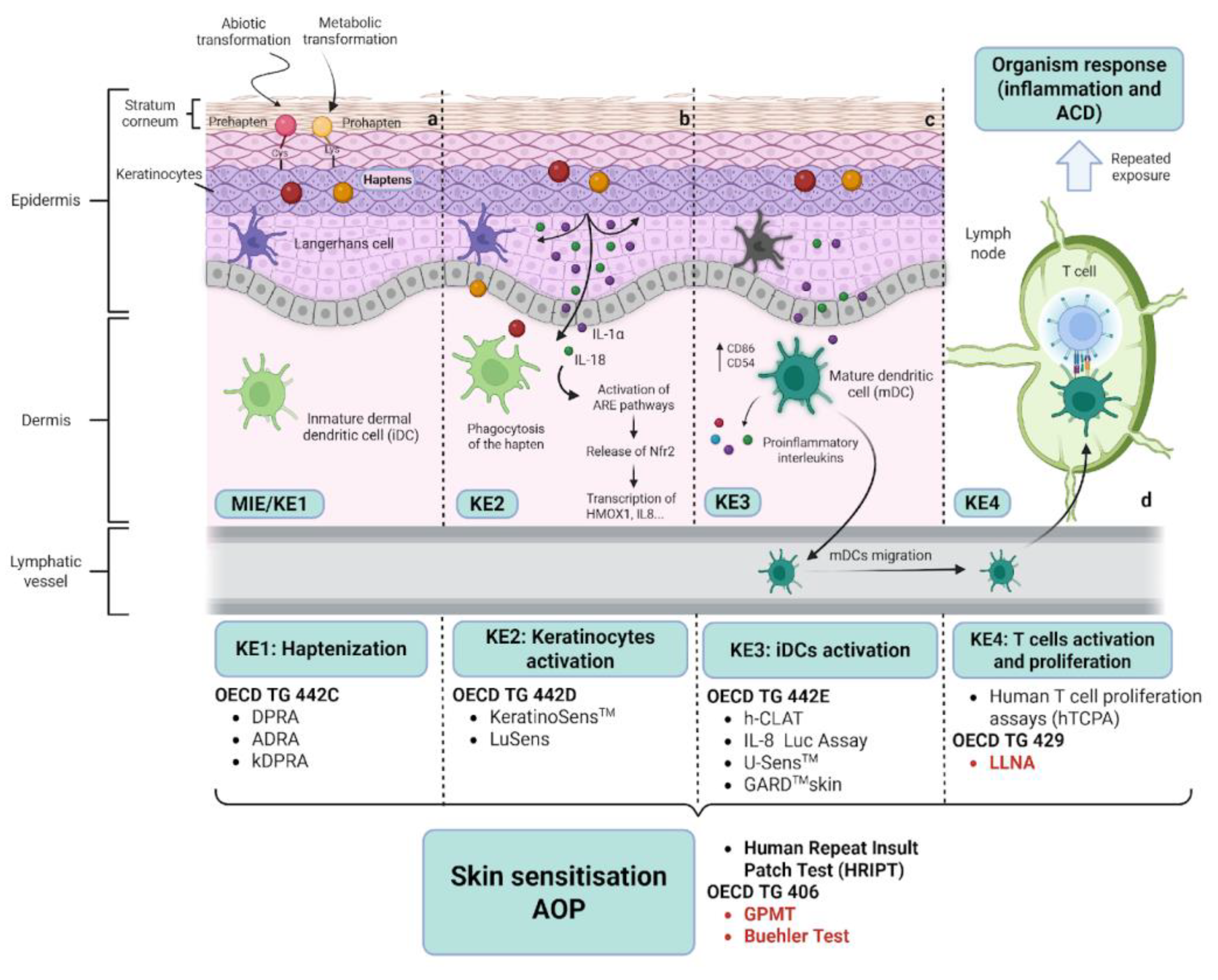
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



