Submitted:
09 July 2025
Posted:
09 July 2025
You are already at the latest version
Abstract
The [2Fe-2S] transcription activator SoxR, a member of the MerR family, functions as a bacterial stress-response sensor. The response governed by SoxR is mediated either directly or indirectly by superoxide (O2-). In this review, we describe functional differences between Escherichia coli SoxR (EcSoxR) and Pseudomonas aeruginosa SoxR (PaSoxR). Pulse radiolysis demonstrated that the reduced form of EcSoxR reacts directly with O2- with a second-order rate constant of 5.0 × 108 M-1s-1. PaSoxR was found to undergo a similar reaction, although with 10-fold smaller rate constant (4.0 × 107 M-1s-1). This difference in rate constants reflects distinct regulatory features of EcSoxR and PaSoxR. Specifically, mutagenesis studies, have shown that Lysine residues, located close to [2Fe-2S] clusters, in EcSoxR―which are not conserved in PaSoxR―are essential for EcSoxR activation. Supporting the model of indirect activation by O2-, both EcSoxR and PaSoxR were found to react with various redox-active compounds (RACs), including viologens, phenazines, and quinones, with no apparent differences in the kinetic behavior or specificity of the two proteins.
Keywords:
iron-sulfur cluster
; superoxide response
; redox active compound
; oxidative stress
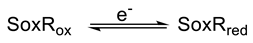
1. Introduction
Iron-sulfur clusters, composed of iron and inorganic sulfur, are universal cofactors that play many important roles in biological processes [1,2,3,4,5]. Iron-sulfur clusters in signal-transducing sensor proteins are functionally distinct from most iron-sulfur clusters, which act as redox centers for electron transfer reactions. In sensor proteins, iron-sulfur clusters respond to small signaling molecules in the environment [4,6,7,8], resulting in changes in the cluster type, oxidation state, or occupancy, that may alter gene expression in cells. The [2Fe-2S] cluster of SoxR, one example of such iron-cluster-containing sensor proteins, undergoes reversible one-electron oxidation and reduction between SoxRox and SoxRred, according to the equation.
Because SoxRred is paramagnetic, its redox changes can be quantitatively assessed by EPR spectroscopy. In vivo EPR spectra show that SoxR in E. coli (EcSoxR) is maintained in a reduced, inactive form during aerobic growth [8]. Exposure of E. coli cells to O2- generating compounds such as metyl viologen (MV2+) causes EPR signals to disappear rapidly, indicating oxidation of the EcSoxR cluster. Thus SoxR senses oxidative stress. The resulting activated redox state is transmitted to the SoxR DNA-binding domain, resulting in activation of target gene promoters [9,10,11,12,13].
The target of activated SoxR is a single adjacent soxS gene, whose product induces transcription of more than 100 genes in the soxRS regulon [14]. This two-step soxRS-based regulatory mechanism controls the expression of genes involved in antioxidant defense proteins such as Mn-containing superoxide dismutase (SOD), glucose-6-phosphatedehydrogenase, DNA repair nucleobase, and outer membrane drug efflux pump, as shown in Figure 1.
A number of SoxR homologs have been identified in various bacterial species [15], all of which contain a SoxR-specific cysteine motif required for binding of the [2Fe-2S] cluster. Unlike those in E. coli and related enteric bacteria, however, the majority of SoxR regulons in non-enteric bacteria such as P. aeruginosa and Streptomyces coelicolor lack genes typically involved in O2- resistance and detoxification [15]. Many of these bacteria produce redox-active compounds (RACs) independent of oxidative stress [15,16]. PaSoxR is activated by endogenous, RAC redox-antibiotic compounds such as pyocyanin (Pyo) and actinorhodin (Act) [15,16,17,18], which are the physiological signals for upregulation of quorum-sensing-regulated genes. These SoxR target genes regulate genes involved in transport,-(via the mexGHI-opmD RND efflux pump [19])-, and the metabolism of small molecules [19], including RAC antibiotics and endogenous pigments (Figure 2). In these species, SoxR appears to function as a defense against the toxicity of RACs.
2. Direct Reaction of O2- with [2Fe-2S] Cluster in SoxR
An important question is the nature of the signal sensed by SoxR. The first candidate SoxR-activating signal identified was O2- [20], but the mechanism by which O2- activates SoxR remains unclear [21]. In fact, Gu and Imaly [22] reported that the soxRS response of E. coli is directly activated by RACs rather than by O2-. To investigate the reaction of O2- with EcSoxR, we monitored the reactions that occurred following pulse radiolysis of oxygen-saturated solutions in the presence of EcSoxRox [13]. Under these experimental conditions, most of the hydrated electrons (eaq-) generated reacted with O2 to form O2-, while a part of the eaq- population rapidly reduced the [2Fe-2S] cluster. The initial changes in absorption subsequently partially reversed on a millisecond time scale, a. recovery that was inhibited by the addition of Cu/Zn-SOD. On the basis of these pulse radiolysis results, we propose the reaction schemes summarized by reactions (1) and (2):
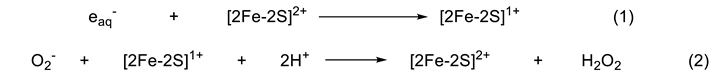 where [2Fe-2S]2+ and [2Fe-2S]1+ are oxidized and reduced forms of SoxR, respectively.
where [2Fe-2S]2+ and [2Fe-2S]1+ are oxidized and reduced forms of SoxR, respectively.
Pulse radiolysis revealed that a similar process was at work in PaSoxR. To compare the reactions of EcSoxR and PaSoxR, we studied the effect of SOD on the oxidation of SoxRred (Figure 3). The oxidation of PaSoxR was completely inhibited by 0.5 µM SOD (Figure 3B), whereas the same concentration of SOD inhibited EcSoxR by only 20 % (Figure 3A). This suggests that the reactivity of O2- toward EcSoxR is much greater than toward PaSoxR. The SOD dependence of absorption changes at 420 nm for EcSoxR is presented in Figure 3C. Based on the concentration of SOD needed to half-maximally inhibit oxidation of SoxRred and the known rate constant for the reaction of O2- with SOD, we obtained a second-order rate constant of 5.0 × 108 M-1s-1 for the reaction of O2- with EcSoxRred. In contrast, the second-order rate constant for PaSoxRred with O2- was calculated to be 4.0 × 107 M-1s-1. The difference in rate constant reflects the distinct regulatory roles for EcSoxR and PaSoxR in response to their physiological activation by O2-. The mechanism underlying the different sensitivities of various SoxRs to O2- remains an important question.
These data clearly suggest that small alternation in structure between highly homologous SoxR proteins can lead to functional differences. Figure 4A shows structure-based sequence alignments and secondary structure assignments for SoxR proteins from different enteric and non-enteric bacteria. SoxR homologs from E. coli, enteric species contain two Lysine residues, which are replaced by Alanic residues in SoxR of almost all non-enteric species [23]. The crystal structure of EcSoxR [24] shows that Lys89 and Lys92, located immediately upstream of dimerization helix 5, are adjacent to the [2Fe-2S] cluster (Figure 4B). In addition, SoxRs from most species of enteric bacteria contain a conserved three-residue hydrophilic motif (R127S128D129) in the vicinity of [2Fe-2S] clusters that affects SoxR sensitivity to RACs [23] and is not conserved in SoxR from other non-enteric bacteria [23].
Mutagenesis experiments have investigated determinants of SoxR sensitivity to O2-, focusing on several amino acids that are not conserved among SoxR homologues. Specifically, the EcSoxR mutants (K89A, K92A, K89AK92A, D129A, and R127LS128QD129A) and PaSoxR mutants (A87K, A90K, and L125RQ126SA127D) were prepared and their O2- sensitivity was determined by examining the concentration-dependence of SOD inhibition [23]. Table 1 shows the rate constants for EcSoxR and PaSoxR mutants. Although the rate constants for K89A (3.8 ×108 M-1 s-1), and K92A (2.2 × 108 M-1s-1) mutants were only slightly affected by the indicated substitutions, the rate constant for the double-Lys, K89AK92A mutant (3.3 x 107 M-1s-1) dramatically affected. The rate constant of 3.3 × 107 M-1s-1 was 10 times smaller than that of wild-type SoxR. Conversely, the corresponding substitution of K to A in PaSoxR at residues (K87A) or 90 (K90A) increased rate constants approximately 10-fold- to 2.1 × 108 and 5.4 × 108 M-1s-1, respectively (Table 1). Therefore, substituting an Alanine residue for a Lysine residue in the [2Fe-2S] region of EcSoxR is sufficient to transform the O2--sensitivity of EcSoxR such that it mimics that of PaSoxR. The effects of the electrostatic charge of amino acid residues were further analyzed by replacing both positive charged both Lys89 and Lys92 residues in EcSoxR with negatively charged glutamate (K89EK92E). The resulting charge reversal from positive to negative decreased the rate constant; in contrast substituting arginine, another positively charged amino acid, for these Lysine (K89RK92R) did not change rate constants (Table 1). Thus, these Lysine residues are responsible for the high diffusion-limited rate constant. Collectively, these results demonstrate that these electrostatically positive Lysine residues are critical for the reaction of O2- with the [2Fe-2S] cluster in EcSoxR and suggest that they guide O2- into the active site on the surface of SoxR. These findings are consistent with the previously reported involvement of Lysine residues in enzymatic reactions with O2- [25,26,27].
SoxR from most species of enteric bacteria contains a three-residue hydrophilic motif (R127S128D129) in the vicinity of [2Fe-2S] clusters. These residues are not conserved in SoxR of other bacteria. It has been proposed that the presence of this conserved motif affects the sensitivity of SoxR to RACs [18,28] and is responsible for the observed species-specific differences in RAC sensitivity. However, unlike the above Lys substitutions, the RSD→LQA substitution in EcSoxR and LQA→RSD substitution in PaSoxR did not affect O2- reaction rates (Table 1).
3. Response of the SoxR [2Fe-2S] Cluster to RACs
EcSoxR has been shown respond to endogenous, as well as various synthetic RACs [17,18,28,29], suggesting that EcSoxRred is oxidized directly, but nonspecifically, by RACs. In contrast, the non-enteric PaSoxR is activated by endogenous small-molecule RACs and appears to sense and respond to a more restricted range of molecules. The sensitivity and specificity of various RACs were examined by investigating the kinetics of RAC-mediated oxidation of the [2Fe-2S] cluster in SoxR using pulse radiolysis [30]. Pulse radiolysis enables reduction of [2Fe-2S]2+ in SoxRox by a hydrated electron (eaq-) within 1 µs (reaction 1) , which permits subsequent oxidation of [2Fe-2S]red by RACs such as MV2+ (reaction 3).
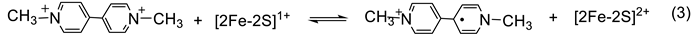
Figure 5 shows the changes in absorption after pulse radiolysis of PaSoxR in the absence and presence of MV2+, demonstrating reduction of [2Fe-2S] by MV2+, as reflected in a rapid decrease in absorbance at 420 nm. After initially decreasing rapidly in the presence of MV2+, partially reversed on a millisecond time scale. From these findings, it can be inferred that the [2Fe-2S] cluster of SoxRox is initially reduced by eaq-, and subsequently becomes partially reoxidized, implying that the recovery process reflects the oxidation of SoxRred by MV2+ (reaction 3). Similar absorption changes were observed in EcSoxR.
To determine the second-order rate constants in the reaction of MV2+ with PaSoxRred, we used conditions in which the concentrations of the RAC (10-25 µM) was much higher than that of SoxRred (0.5-1 µM). Under these conditions, the oxidation of SoxRred obeyed pseudo-first-order kinetics, with the observed rate constant increasing linearly with RAC concentration. The rate constant of the reaction, obtained from the slope in Figure 5B was determined to be 3.0 × 108 M-1s-1. Similar absorption changes were observed in the presence of other RACs, including pyocyanin (Pyo), phenazine methosulfate (PMS), duroquinone (Dur), and diquat (DQ). The second-order rate constants increased in the order Dur < MV2+ < DQ < Pyo < PMS (Table 2) and were found to correlate with the electrode potential of the RAC. In all cases, a rapid decrease phase was followed by a slower recovery phase. Notably, absorbance changes during the slower reoxidation phase varied depending on the specific RACs. The incomplete reoxidation of the [2Fe-2S] cluster may be explained by a rapid equilibrium for electron transfer between the RAC and the [2Fe-2S] cluster (reaction 3).
The pulse radiolysis results presented here show that the [2Fe-2S]red cluster of EcSoxR and PaSoxR react with RAC compounds with electrode potentials in the range of -440 mV (MV2+) [31] to + 80 mV (PMS) [33]. Importantly, these results are consistent with the responsiveness of SoxR to RACs observed in vivo [18,22,29]. In comparison of the sensitivity profiles of SoxR towards RACs, Singh et al. showed that both EcSoxR and PaSoxR responded to all RACs, except Act [18].
SoxR homologs exhibit different selectivities toward RACs with each displaying distinctive effective concentrations and cellular response times. However, our study found that these differences are not reflected in the kinetics of the reactions. Specifically, pulse radiolysis reveled no difference in the reactivity of the [2Fe-2S] cluster between EcSoxR and PaSoxR, despite their differing cellular response times to MV2+ [22].
4. Physiological Significance of SoxR Response to O2- and RACs
In this review, we demonstrated that O2- itself is the signal that activates SoxR [11,13,23]. The functional differences between the SoxR proteins, EcSoxR and PaSoxR, revealed by pulse radiolysis, reflect differing responses to O2-. Mutagenesis studies have identified residues that are essential for the ability of EcSoxR to respond O2-. In addition, it is noteworthy that reaction (2) observed in SoxR represents the first reported example of a reaction between a reduced form of iron-sulfur protein and O2-. Whether other [2Fe-2S] proteins show similar reactivity was tested by examining their reactions with O2- using spinach ferredoxin and adrenodoxin as examples. No recovery of the absorption signal was observed following oxidation of the reduced forms of these [2Fe-2S] proteins. On the other hand, in the reaction of oxidized form of iron-sulfur proteins, it was reported that the reactions between iron-sulfur proteins and O2- with second-order rate constants ranging from 106 to 107 M-1 s-1 [34,35,36]. In these cases, the reactions were irreversible and were accompanied by with degradation of the iron-sulfur cluster. By contrast, reaction (2) in SoxR is reversible. Therefore, it can be said that reaction (2) of SoxR is a specific characteristic of SoxR protein.
At this point it is important to ask whether the oxidation of EcSoxR with O2- occurs in vivo. How O2- can interact with SoxR under the conditions of high levels of SOD and a low level of SoxR presented constitutively [37,38]. The extent to which EcSoxR transcriptional activity in vivo is correlated with the effects of mutation on O2- sensitivity observed by pulse radiolysis was investigated by β-galactosidase assays in an E. coli strain (ΔsoxR soxS::lacZ) expressing SoxR mutants [23]. The EcSoxR Lysine mutant, K89AK92A, exhibited defective in vivo transcriptional activity in response to MV2+, approximately 10 % that of the wild-type-as measured by β-galactosidase expression [23]. This in vivo data is consistent with the mutant protein’s low O2- sensitivity and supports the idea that the reaction of O2- with the [2Fe-2S]+ of SoxR is responsible for the transcription activity of EcSoxR in vivo.
The data presented here provide new insight into electron transfer between RACs and the [2Fe-2S] cluster of SoxR homologs. The observed oxidation of the [2Fe-2S] in SoxR proceeds via direct electron transfer from the [2Fe-2S] cluster to the RAC. It is likely that structurally distinct RAC compounds can gain access via the solvent-exposed surface area allowing rapid electron transfer between the [2Fe-2S] cluster and RAC. An X-ray analysis of DNA-free EcSoxR [24] showed that the upper sulfur atom (S2) and one of the Fe atoms are completely exposed to the solvent. A similar mechanism may take place at the SoxR active site in PaSoxR. The oxidizability of SoxRred is assisted by the low potential of its [2Fe-2S] cluster in EcSoxR (-285 mV) [39] and PaSoxR (-290 mV) [40] which provide a strong driving force for electron transfer to even the moderate oxidant (-440 mV), MV2+ [33].
5. Conclusion and Future Respective
In this review, we have identified reactions of [2Fe-2S] cluster with O2- and RACs using pulse radiolysis. The rate constants are extremely high and these reactions are specific character of SoxR. A major challenge now is to determine the factors that control cluster of SoxR. Future progress will continue at the interface of fields of physical chemistry, biological chemistry, and inorganic chemistry to understand how cells respond to their environment special and effective machinery to control and detoxify reactive radicals.
References
- E. Boncella, E. T. Sabo, R. M. Santore, J. Carter, J. Whalen, J. D. Hudspeth, and C. N. Morrison, “The expanding utility of iron-sulfur clusters: Their functional roles in biology, synthetic small molecules, maquettes and artificial proteins, biomimetic materials, and therapeutic strategies” Coordination Chem. Vol. 453, 214229 2022.
- M. Gupta and C. E. Outten, “Iron-Sulfur Cluster Signaling; The Common Thread in Fungal Iron Regulation,” Curr. Opin. Chem. Biol. Vol. 55, 189-201, 2020.
- J. C. Crack, J. Green, A. J. Thomson, and N. E. le Brun, “Iron-sulfur cluster sensor-regulators.” Curr. Opin. in Chem. Biol. Vol. 16, 35-44, 2012.
- H. Beinert, R. H. Holm, E. Münck, “Iron-sulfur clusters: Nature’s modular, multipurpose structures.” Science, Vol.277, 653-659, 1997.
- D. Read, R. ET. Bentrley, S. L. Archer, K. J. Dunham-Snary, “Mitochondrial iron-sulfur clusters: Structure, function, and an emerging role in vascular biology.” Redox Biology, Vol.47, 12164, 2021.
- S. M. Bian and J. A. Cowan, “Protein-bound iron-sulfur centers. Form, function, and assembly.” Coord. Chem. Rev. 1999; 190, 1049–1066.
- P. J. Kiley and H. Beinert, “The role of Fe-S proteins in sensing and regulation in bacteria,” Curr. Opin. Microbiol., Vol. 6, 181-185, 2003.
- H. Ding, B. Demple, “In vivo kinetics of a redox-regulated transcriptional switch,” Proc. Natl. Acad. Sci. U.S.A. Vol. 97, 5146-5150, 2000.
- F. Amábile-Cuevas, B. Demple, “Molecular characterization of the soxRS genes of Escherichia coli: Two genes control a superoxide stress regulon,” Nucleic Acids Res. Vol.19, 4479-4484, 1991.
- P. J. Pomposiello, M. H. J. Bennik, B. Demple, “Genome-wide transcriptional profiling of the Escherichia coli responses to superoxide stress and sodium salicylate.” J. Bacteriol. Vol.183, 3890-3892, 2001.
- K. Kobayashi, M. Fujikawa, and T. Kozawa, “Oxidative stress sensing by the iron-sulfur cluster in the transcription factor, SoxR” J. Inorg. Biochem, Vol.133, 87-91, 2014.
- P. Gaudu, and B. Weiss, “SoxR, a [2Fe-2S] transcription factor, is active only in its oxidized.” Proc. Natl. Acad. Sci. U. S. A. Vol. 93, 10094-10098, 1996.
- M. Fujikawa, K. Kobayashi, and T. Kozawa, “Direct Oxidation of the [2Fe-2S] Cluster in SoxR Protein by Superoxide. Distinct Differential Sensitivity to Superoxide-Mediated Signal Transduction.” J. Biol. Chem. Vol. 287, 35702-35708, 2012.
- J.L. Blanchard, W. Y. Wholey, E. M. Conlon, and P. J. Pomposiello, “Rapid changes in gene expression dynamics in response to superoxide reveal soxRS-dependent and -independent transcriptional networks.” PLoS ONE Vol.2 e1186, 2007.
- L. E. P. Dietrich, T. K. Teal, A. Price-Whelan, and A. D. K. Newman, “ Redox-Active Antibiotics Control Gene Expression and Community Behavior in Divergent Bacteria.” Science Vol. 321, 1203-1206, 2008.
- R. D. Cruz, Y. Gao, S. Penumetcha, R. Sheplock, K. Weng, and M. Chander, “Expression of the Streptomyces coelicolor SoxR Regulon Is Intimately Linked with Actinorhodin Production.” J. Bactriol. Vol. 192, 6428-6438, 2010.
- J.-H. Shin, A. K. Singh, D.-J. Cheon, and J.-H. Roe, J.-H. “Activation of the SoxR Regulon in Streptomyces coelicolor by the Extracellular Form of the Pigmented Antibiotic Actinorhodin.” J. Bacteriol. Vol.193, 75-81, 2011.
- K. Singh, J.-H. Shin, K.-L. Lee, J. A. Imlay, and J.-H. Roe, “Comparative study of SoxR activation by redox-active compounds” Mol. Microbiol. Vol. 90, 983–996, 2013.
- Z. Ma, F. E. Jacobsen, and D. P. Giedroc, “Coordination Chemistry of Bacterial Metal Transport and Sensing” Chem. Rev. Vol. 109, 4644-4681, 2009.
- S. I. Liochev, “Is superoxide able to induce SoxRS ?” Free Radic. Biol. Med Vol. 2011; 50, 1813.
- L. E. P. Dietrich and P. J. Kiley “A shared mechanism of SoxR activation by redox-cycling compounds” Mol. Microbiol. Vol. 2011; 79, 1119–1122.
- M. Gu and J. A. Imaly, “The SoxRS response of Escherichia coli is directly activated by redox-cycling drugs rather than by superoxide.” Mol. Microbiol. Vol. 79, 1136-1150, 2011.
- M. Fujikawa, K. Kobayashi, Y. Tsutsui, T. Tanaka, and T. Kozawa, “Rational Tuning of Superoxide Sensitivity in SoxR, the [2Fe-2S] Transcription Factor: Implication of Species-Specific Lysine Residues.” Biochemistry Vol. 56, 403-410, 2017.
- S. Watanabe, A. Kita, K. Kobayashi, and K. Miki, “Crystal structure of the [2Fe-2S] oxidative-stress sensor SoxR bound to DNA. Proc. Natl. Acad. Sci. U. S. A. Vol. 105, 4121-4126, 2008.
- E. D. Getzoff, J. A. Tainer, P. K.Weiner, P. A. Kollman, J. S. Richardson, and D. C. Richardson, “Electrostatic recognition between superoxide and copper, zinc superoxide dismutase.” Nature Vol. 306, 287-290, 1983.
- M. L. Ludwig, A. L. Metzger, K. A. Pattrige, and W. C. Stallings (1991) “Manganese superoxide dismutase from Thermus thermophilus: A structural model refined at 1.8 Å resolution.” J. Mol. Biol. Vol. 219, 335-358, 1991.
- Y. Sheng, I. A. Abreu, D. E. Cabelli, M. J. Maroney, A.-F. Miller, M. Teixeira, J. S.Valentine, “Superoxide Dismutases and Superoxide Reductases.” Chem. Rev. Vol. 114, 3854-3918, 2014.
- K. L. Lee, A. K. Singh, L. Heo, C. Seok, and J.-H. Roe “Factors affecting redox potential and differential sensitivity of SoxR to redox-active compounds.” Mol. Microbiol. Vol. 97, 808-821 2015.
- R. Sheplock, D. A. Recinos, N. Macknow, L. E. P. Dietrich, and M. Chander, Species-specific residues calibrate SoxR sensitivity to redox-active molecules. Mol. Microbiol. Vol. 87, 368-381. 2013.
- K. Kobayashi, T. Tanaka, T. Kozawa, “Kinetics of the Oxidation of the [2Fe-2S] Cluster in SoxR by Redox-Active Compounds as Studied by Pulse Radiolysis.” Biochemistry Vol. 64, 895-902, 2025.
- E. Steckhan, and T. Kuwana, “Spectrochemical study of mediators. I Bipyridyum salts and their electron transfer to cytochrome C.” Ber. Bunsengesellschaft Phys. Chem, Vol. 78, 253-258, 1974.
- P. Wardman, “Reduction Potentials of One-Electron Couples involving Free Radicals in Aqueous Solution.” J. Phys. Chem. Ref. Data Vol. 18, 1637-1655, 1989.
- A. Moffet, J. Foley, J., and M. H. Hecht “Midpoint reduction potentials and heme binding stoichiometries of de nova proteins from designed combinatorial libraries.” Biophys. Chem. Vol. 35105, 231-239, 2003.
- F. Kuo, T. Mashino and I. Fridovich, “α,β-Dihydroxyisovalerate Dehydratase.” J. Biol. Chem. Vol. 262, 4724-4727 1987.
- P. N. Gardner and I. Fridovich, I. “Superoxide Sensitivity of the Escherichia coli Aconitase.” J. Biol. Chem. Vol. 266, 19328-19333, 1991.
- P. N. Gardner, and I. Fridovich, “Superoxide Sensitivity of the Escherichia coli 6-Phosphogluconate Dehydratase.” J. Biol. Chem. Vol. 266, 1478-1483, 1991.
- P. J. Pomposiello, and B. Demple “ReDox-operated genetic switches: the SoxR and OxyR transcription factors.” Trends Biotechnol. Vol. 19, 109-114, 2001.
- J. A. Imaly and I. Fridovich “Superoxide sensitivity of the Escherichia coli 6-phosphogluconate dehydratase” J. Biol. Chem. Vol. 266, 1478-1483, 1991.
- H. Ding, E. Hidalgo, B. Demple “ The redox stateof the [2Fe-2S] clusters in SoxR protein regulates its activity as a transcription factor” J. Biol. Chem. Vol. 271, 33173-33175, 1996.
- K. Kobayashi, and S. Tagawa “ Activation of SoxR-dependent transcription in Pseudomonas aeruginosa. J. Biochem. Vol. 136, 607-615, 2004.
Figure 1.
The mechanism underlying transcription of the soxRS regulon in E. coli. SoxR is activated by exposure to O2--generating compounds, redox active compounds (RACs), or nitric oxide (NO). The target of activated SoxR is the soxS gene. Increased SoxS induces expression of various antioxidant proteins.
Figure 1.
The mechanism underlying transcription of the soxRS regulon in E. coli. SoxR is activated by exposure to O2--generating compounds, redox active compounds (RACs), or nitric oxide (NO). The target of activated SoxR is the soxS gene. Increased SoxS induces expression of various antioxidant proteins.
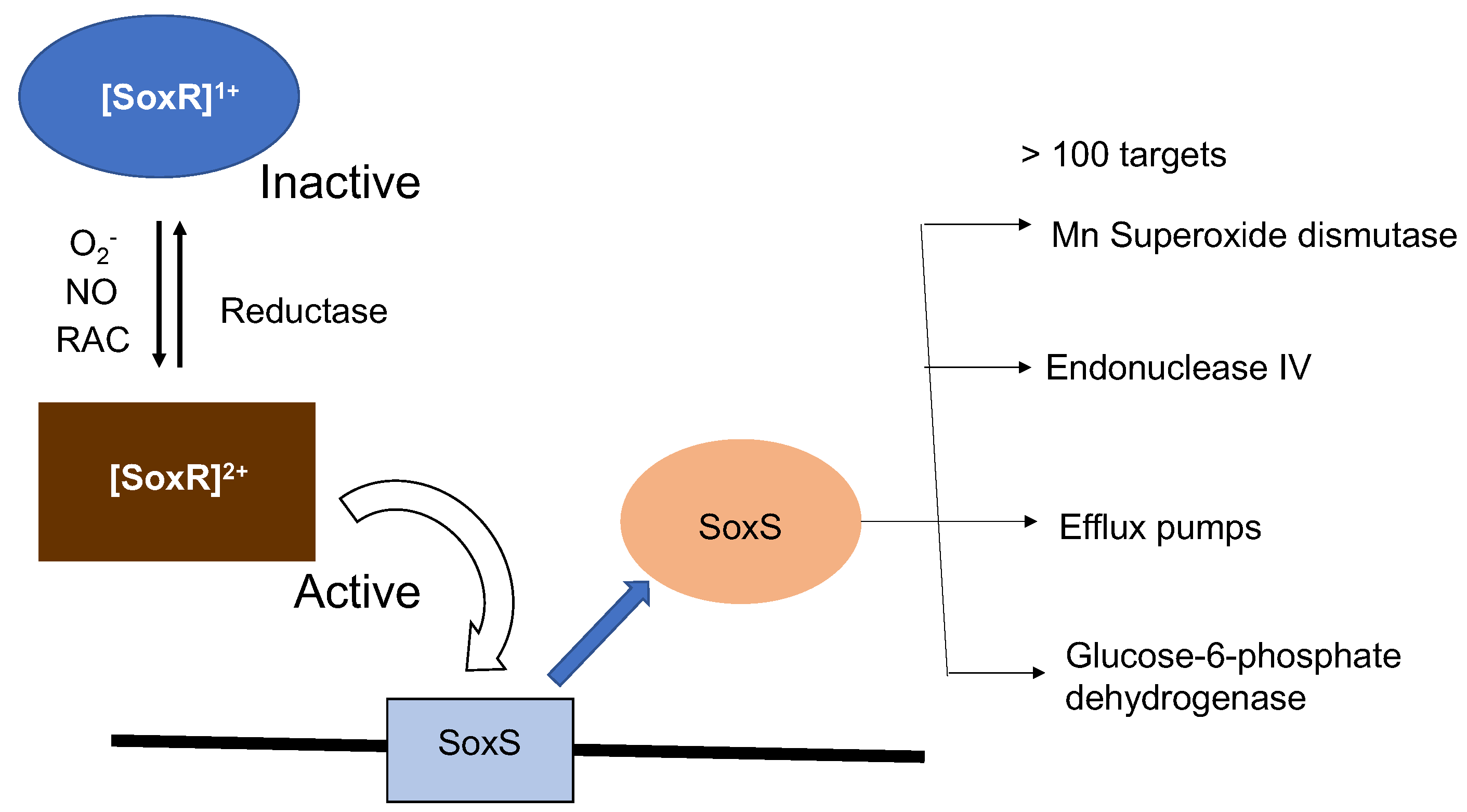
Figure 2.
SoxR response in non-enteric bacteria.
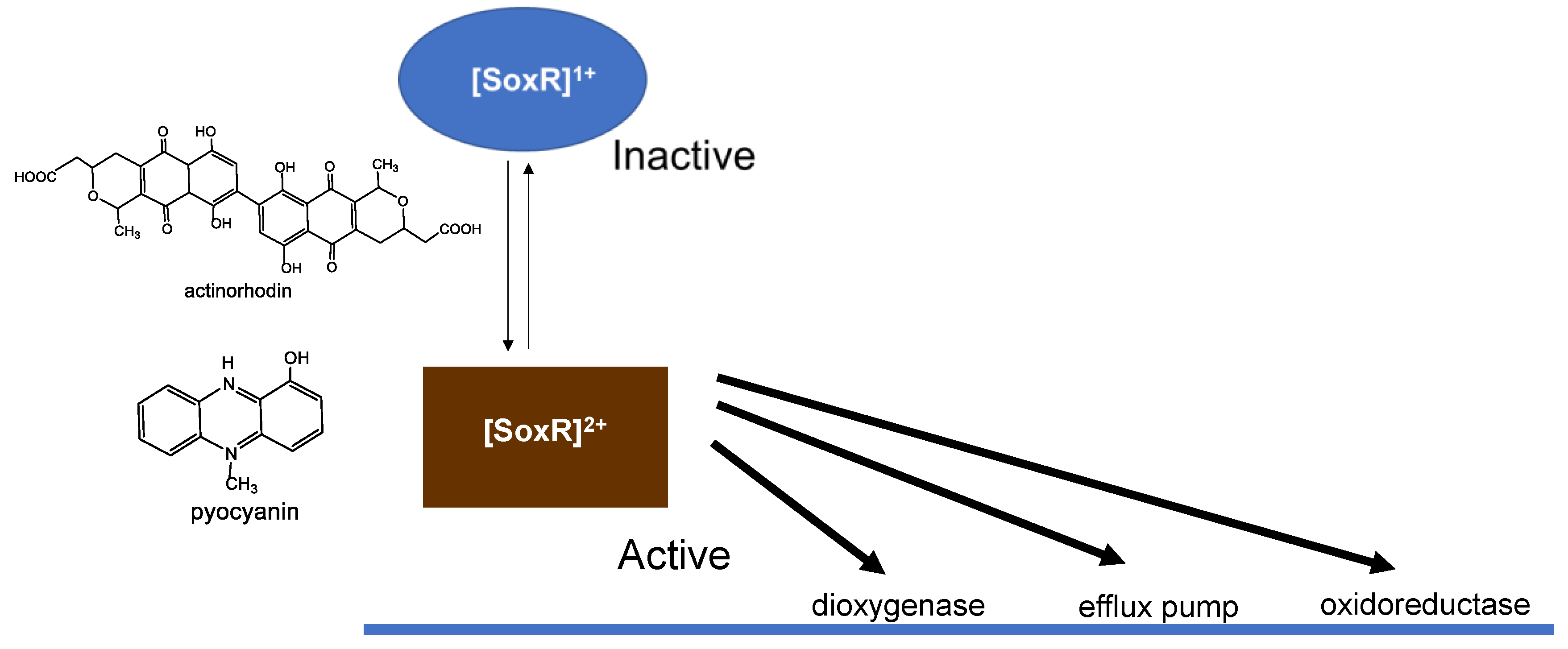
Figure 3.
SOD effects on the oxidation of SoxR. Changes in absorbance measured after pulse radiolysis of EcSoxR (A) and PaSoxR (B) in the absence (blue line) or presence (red line) of 0.5 µM SOD. (C) The absorbance change at time (ΔAt), normalized to the total absorbance change (ΔA0) plotted against the concentration of SOD. Reproduced from ref. [13]. Copyright 2012 J. Biol. Chem.
Figure 3.
SOD effects on the oxidation of SoxR. Changes in absorbance measured after pulse radiolysis of EcSoxR (A) and PaSoxR (B) in the absence (blue line) or presence (red line) of 0.5 µM SOD. (C) The absorbance change at time (ΔAt), normalized to the total absorbance change (ΔA0) plotted against the concentration of SOD. Reproduced from ref. [13]. Copyright 2012 J. Biol. Chem.
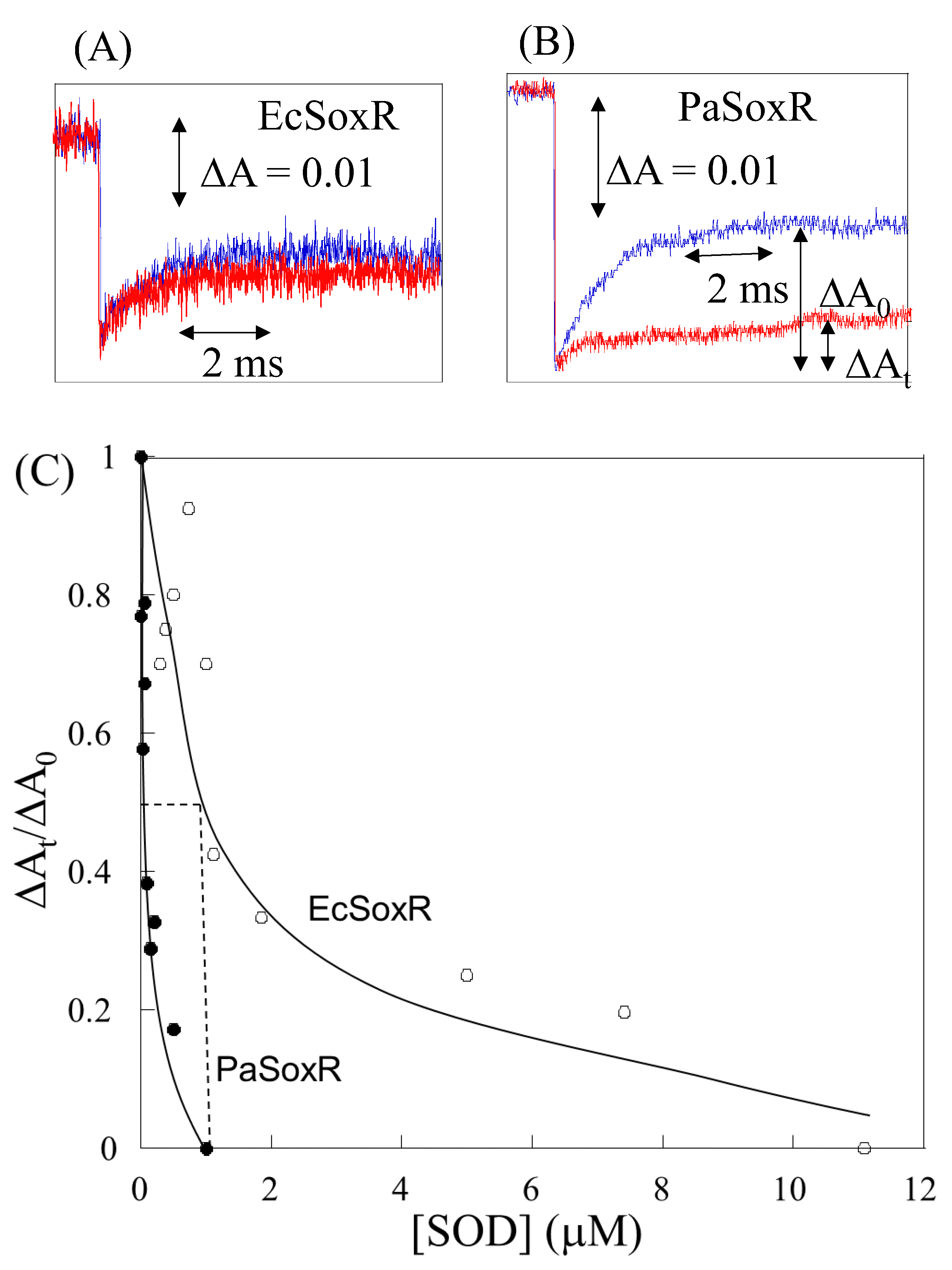
Figure 4.
(A) Sequence alignments of SoxR proteins from enterics E. coli, Sphingopyxis alaskensis, and Shigella flexneri and non-enteric (P. aeruginosa, Xanthomonas axonopodi, Vibrio cholerae) bacteria. Identical and conserved residues among SoxR proteins are denoted in red and green, respectively. Hypervariable residues and the hypervariable motif are highlighted in yellow. (B) structure of EcSoxR in the region near the [2Fe-2S] cluster. The structure was produced with PyMol using a structure from the Protein Data Bank (entry 2ZHG). Reproduced from ref. [23]. Copyright 2016 American Chemical Society.
Figure 4.
(A) Sequence alignments of SoxR proteins from enterics E. coli, Sphingopyxis alaskensis, and Shigella flexneri and non-enteric (P. aeruginosa, Xanthomonas axonopodi, Vibrio cholerae) bacteria. Identical and conserved residues among SoxR proteins are denoted in red and green, respectively. Hypervariable residues and the hypervariable motif are highlighted in yellow. (B) structure of EcSoxR in the region near the [2Fe-2S] cluster. The structure was produced with PyMol using a structure from the Protein Data Bank (entry 2ZHG). Reproduced from ref. [23]. Copyright 2016 American Chemical Society.
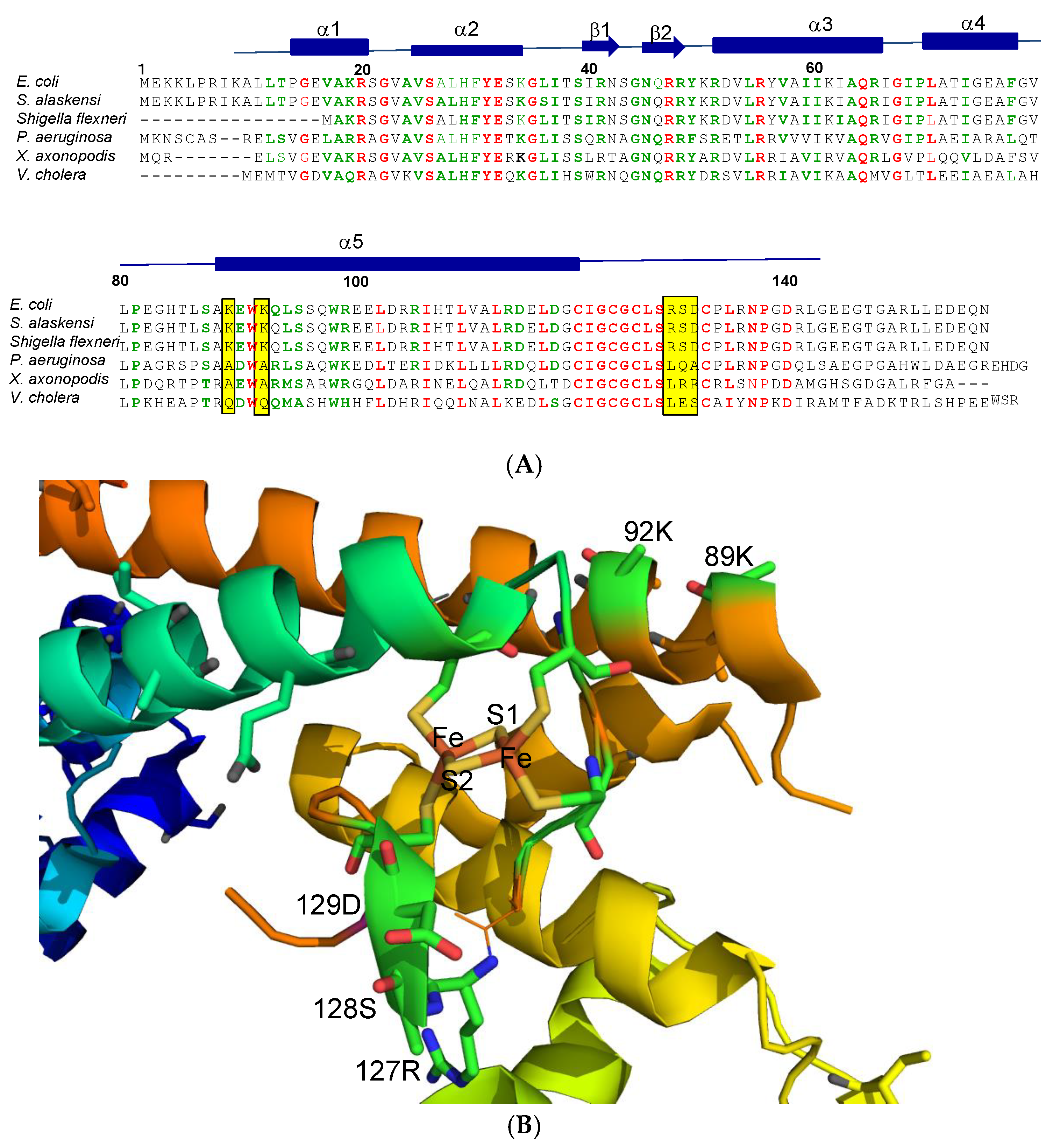
Figure 5.
(A) Changes in absorbance at 420 nm after pulse radiolysis of PaSoxR. Samples contained 70 µM PaSoxR, 20 mM potassium phosphate buffer (pH 7.0), 10 mM potassium/sodium tartrate, 0.5 M KCl, and 0.1 M t-butanol, to which was 20 µM MV2+ was further added. (B) Concentration dependence of MV2+ versus kapp in PaSoxR.
Figure 5.
(A) Changes in absorbance at 420 nm after pulse radiolysis of PaSoxR. Samples contained 70 µM PaSoxR, 20 mM potassium phosphate buffer (pH 7.0), 10 mM potassium/sodium tartrate, 0.5 M KCl, and 0.1 M t-butanol, to which was 20 µM MV2+ was further added. (B) Concentration dependence of MV2+ versus kapp in PaSoxR.

Table 1.
Second-Order Rate Constants in Reactions O2- with the Reduced Forms of Wild Type and Mutant SoxR Proteins.
Table 1.
Second-Order Rate Constants in Reactions O2- with the Reduced Forms of Wild Type and Mutant SoxR Proteins.
| E. coli | k × 108 (M-1 s-1) |
| WT | 5.0 |
| R127LS128QD129A | 4.8 |
| D129A | 5.0 |
| K89A | 3.8 |
| K92A | 2.2 |
| K89AK92A | 0.33 |
| K89RK92R | 4.7 |
| K89EK92E | 0.31 |
| P.aeruginosa | |
| WT | 0.4 |
| A87K | 2.1 |
| A90K | 5.4 |
| L125RQ126SA127D | 0.4 |
Table 2.
Rate constants of [2Fe-2S] cluster of EcSoxR, and PaSoxR, with RACs and the electrode potentials E0 (vs NHE) at pH 7.0.
Table 2.
Rate constants of [2Fe-2S] cluster of EcSoxR, and PaSoxR, with RACs and the electrode potentials E0 (vs NHE) at pH 7.0.
| RAC compounds | E0 (vs NHE) (mv) | EcSoxR k (× 108 M-1s-1) |
PaSoxR k (× 108 M-1s-1) |
|---|---|---|---|
| MV2+ Dq Dur Pyo PMS |
-440 31) -358 32) -260 32) -34 33) +8033) |
3.0 5.7 2.1 7.1 16.0 |
3.0 5.5 2.6 6.8 14.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



