Submitted:
07 July 2025
Posted:
08 July 2025
You are already at the latest version
Abstract
This review establishes a robust conceptual and mechanistic framework for rationally designing advanced nanobiotechnological systems explicitly tailored for efficient blood–brain barrier (BBB) translocation, addressing one of the most critical challenges in neurological drug delivery. We systematically dissect the mechanisms governing nanoparticle (NP)-BBB interactions, focusing on receptor-mediated transcytosis, adsorptive-mediated transcytosis, cell-mediated transport, and transient barrier modulation, based on recent cellular and preclinical pharmacokinetic studies. Key NP physicochemical parameters, including particle size, surface chemistry, shape, mechanical stiffness, and biofunctionalization strategies, are critically evaluated for their roles in pharmacological efficacy and precision targeting of neurological disorders. Moreover, validated preclinical models and cutting-edge imaging techniques, essential for assessing nanocarrier delivery efficiency and therapeutic performance, are comprehensively discussed. Ultimately, this review provides pharmaceutical researchers and formulation scientists with actionable insights and evidence-based guidelines for designing next-generation nanoformulations that can effectively deliver therapeutics to the brain, thereby significantly enhancing treatment outcomes for neurological diseases.
Keywords:
blood–brain barrier
; nanomedicine
; CNS drug delivery
; nanoparticle
; neurotherapeutics
1. Introduction
Neurological disorders and brain tumors remain among the most devastating and hard-to-treat health challenges worldwide. Neurodegenerative diseases like Alzheimer’s disease and Parkinson’s disease affect tens of millions (over 57 million people lived with dementia globally in 2021) [1], incurring enormous social and economic costs (~$1.3 trillion annual global cost for dementia in 2019) [2]. Malignant CNS cancers such as glioblastoma multiforme (GBM) are relatively rarer but extremely lethal, with a median survival of only ~12–15 months even with aggressive therapy. GBM is the most deadly brain tumor, causing about 200,000 deaths per year worldwide [3]. A unifying obstacle in these conditions, from neurodegeneration to brain cancer, is the difficulty of delivering therapeutic agents into the brain. The culprit is the blood–brain barrier (BBB), a specialized network of cerebral microvascular endothelium that usually preserves the neural microenvironment but also strictly limits drug access to the central nervous system (CNS).
The BBB shields the brain from toxins, pathogens, and fluctuations in blood composition through tight junctions between endothelial cells, pericytes, and astrocytic end-feet, forming a highly selective barrier [4]. Its tight junctions (~1.4 nm pore size) and efflux transporters block nearly all large molecules and ~98% of small-molecule drugs (F. Wang et al., 2024). Consequently, systemic therapeutics rarely penetrate effectively; monoclonal antibodies typically achieve <0.1% brain uptake, and even FDA-approved anti-amyloid antibodies for AD reach just ~0.01–0.1% [5]. Similarly, most small-molecule neurotherapeutics require high lipophilicity, yet remain limited by efflux pumps, size, and polarity. Thus, the BBB remains a significant obstacle for CNS drug delivery [6].
This barrier-imposed delivery problem leads to several critical challenges in current therapy. First, many efficacious drugs in vitro or in peripheral tissues fail in CNS trials because adequate concentrations in the brain cannot be attained. For instance, the chemotherapy paclitaxel is potent against glioma cells but is a substrate of P-glycoprotein; when given systemically, it accumulates to only a minute fraction of plasma levels in the brain, rendering it ineffective unless the BBB is modulated [7]. Second, limited BBB permeability forces clinicians to use extreme measures for CNS delivery. High systemic doses are often required to push a small amount of drug across the BBB, which can cause significant off-target toxicity in the rest of the body (e.g., dose-limiting cardiotoxicity of doxorubicin or peripheral immunosuppression by high-dose cytokines). In other cases, drugs must be delivered directly or locally to the CNS, for example, intrathecal chemotherapy or convection-enhanced delivery in brain tumors to bypass the BBB. Such invasive approaches are risky, impractical for chronic conditions, and poorly tolerated by patients [8]. Similarly, methods to transiently open the BBB (such as focused ultrasound with microbubbles) can enable higher drug entry but also carry risks of tissue damage or infection by allowing toxins or pathogens to breach. Owing to these limitations, many CNS diseases remain inadequately treated. In GBM, for instance, standard chemoradiation yields a two-year survival under 20% in part because drugs like temozolomide, while somewhat BBB-permeable, still do not uniformly reach all tumor cells behind an intact barrier. There is an urgent need for new delivery strategies that can safely and effectively ferry therapeutic molecules across the BBB [9].
Nanotechnology has emerged as a promising avenue to overcome the BBB’s filtering mechanism and improve drug delivery to the brain [10,11,12]. Nanoscale drug carriers, such as polymeric NPs, liposomes, micelles, dendrimers, and protein-based nanovectors, can be engineered to evade or exploit the BBB’s defenses in ways conventional drug formulations cannot. A key advantage is that nanocarriers can be “rationally” designed with specific physicochemical properties (size, shape, surface charge) and functional ligands to engage transport pathways of the BBB actively. In parallel, advances in materials science have led to “smarter” nanobiomaterials that respond to the brain microenvironment. For instance, stimuli-responsive nanocarriers that release their drug cargo upon sensing pH or enzymatic triggers in the brain, or neutrophil-mimetic NPs that can migrate across an inflamed BBB. These approaches collectively aim to maximize CNS delivery while minimizing systemic exposure. Indeed, targeted nano-delivery can reduce off-target toxicity. In one study, an intranasally administered ferritin NP was shown to concentrate in glioma tissue while largely sparing healthy organs, suggesting a safer profile than intravenous chemotherapy [13,14].
This review provides a robust conceptual and mechanistic framework for rationally designing nanobiotechnological systems capable of effectively crossing the BBB. Rather than merely listing existing delivery methods, we emphasize understanding nanoparticle (NP)-barrier interactions and identifying critical factors underlying successful translocation. We systematically examine key NP transport mechanisms, drawing from recent cellular and in vivo studies, and detail essential physicochemical parameters such as NP size, shape, surface chemistry, targeting ligands, and stimulus-responsive properties. Exemplary systems illustrate successful strategies, including functionalized polymeric NPs, engineered exosomes, and BBB-targeted lipid nanoparticles. Therapeutic applications across various CNS disorders are critically evaluated, showcasing preclinical data demonstrating improved drug delivery, enhanced brain bioavailability, extended survival, and neurological recovery. This integrative review serves as a practical guide for researchers developing NP-based therapies, assisting in the informed selection of design parameters and identifying appropriate validation models. Our framework supports the clinical translation of CNS nanotherapeutics by linking fundamental BBB biology to advanced nanotechnological engineering and addressing essential regulatory considerations. Ultimately, strategic and evidence-based NP design represents a powerful approach to effectively overcome the BBB, enabling targeted and efficacious brain therapies.
2. The BBB: Physiology and Restrictive Mechanisms
2.1. Composition and Function of the BBB
The BBB's composition and function comprise brain endothelial cells sealed by tight junctions, supported by pericytes, astrocytic end-feet, and a basement membrane. This neurovascular unit provides selective permeability, maintaining CNS homeostasis. Human brain capillaries extend extensively (~20 m² surface area), ensuring close neuronal proximity to the blood supply. BBB endothelial cells lack fenestrations and exhibit minimal vesicular transport, restricting transport pathways [15,16,17].
Pericytes, which are abundant in CNS vessels (covering more than 90% of vessels in mice), structurally support the blood-brain barrier and induce endothelial tightness. Astrocytic end-feet regulate BBB function, while the basement membrane (50–100 nm thick) restricts permeability and anchors cells [18,19]. These features yield high transendothelial electrical resistance (TEER) and low permeability, protecting the CNS from blood-derived fluctuations and toxins.
2.2. Tight Junctions and Paracellular Restriction
Tight junctions between endothelial cells, composed primarily of claudin-5, occludin, and junctional adhesion molecules linked to actin via ZO-1, create the primary physical barrier. These junctions minimize the paracellular space, allowing only tiny hydrophilic molecules (<1.8 nm diameter) to enter with negligible entry [6,20]. Claudin-5 deletion demonstrates selective permeability to molecules <800 Da, reinforcing the tight junctions’ role in size-dependent exclusion [21,22,23]. Consequently, more than 98% of small-molecule drugs fail to cross the intact blood-brain barrier (BBB) passively [4,24]. The integrity of tight junctions can be disrupted by inflammation or hypoxia, transiently increasing permeability, whereas the tightening of tight junctions correlates with BBB maturation [25]. Conversely, in development, the tightening of tight junctions (especially claudin-5 enrichment) correlates with the maturation of BBB function [23,26]. Overall, the tight junctions network endows the BBB with a restrictive paracellular gate that, in healthy states, admits to virtually no polar molecules larger than a few hundred daltons.
The BBB also features biochemical defenses: efflux transporters and metabolic enzymes. P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP), abundant on endothelial membranes (P-gp: ~4–7 pmol/g, BCRP: ~8–16 pmol/g), actively extrude xenobiotics, significantly restricting CNS entry (Storelli et al., 2021). Impaired efflux activity in Alzheimer’s disease reduces clearance of neurotoxic peptides despite normal transporter levels [27]. Brain endothelial cells also express enzymes (γ-glutamyl transpeptidase, alkaline phosphatase, MAO, and cytochrome P450s) that degrade circulating molecules, thereby preventing CNS entry. Thus, tight junctions, efflux transporters, and enzymatic activity collectively ensure the BBB effectively excludes nearly all drugs and biomolecules from passive CNS entry [28].
2.3. Major Transport Mechanisms Across the BBB
Despite its stringent selectivity, the BBB allows regulated molecular transport via several distinct mechanisms. Passive diffusion enables the transport of a limited range of small (<400–500 Da), lipophilic molecules, including gases (O₂, CO₂) and certain drugs (nicotine, ethanol). However, only approximately 2–6% of small-molecule therapeutics penetrate the BBB passively due to stringent physicochemical constraints [28,29]. Tight junctions (~1–2 nm pore size) severely restrict paracellular transport between endothelial cells, permitting negligible passage of water and small ions. Pathological disruption of tight junction integrity can temporarily increase permeability to molecules that are otherwise restricted [6,21,22]. Carrier-mediated transport (CMT) involves specialized solute transporters that facilitate the uptake of essential nutrients, such as glucose (GLUT1, ~0.5 μmol/g/min), amino acids (LAT1), and vitamins. Notably, alterations in transporter expression occur in diseases such as Alzheimer's [27]. Receptor-mediated transcytosis (RMT) utilizes specific endothelial receptors (e.g., TfR, LDLR, LRP-1, insulin receptor) to shuttle essential peptides, proteins, and nutrients across the BBB. Although effective, RMT typically delivers limited amounts (~0.1–2% of the injected dose for therapeutic antibodies), sufficient for potent drugs [30]. Adsorptive-mediated transcytosis (AMT) involves non-specific electrostatic interactions with cationic or amphipathic molecules, significantly enhancing uptake (e.g., ~10–20-fold increase for cationic albumin). Despite its non-selectivity, AMT is exploited therapeutically, though potential cytotoxicity warrants careful control [31]. Table 1 summarizes the key features of each central transport mechanism across the BBB, highlighting representative molecules, size limitations, typical efficiencies, and relevant examples.
Given the complexity and strictness of BBB physiology, selecting appropriate experimental models is critical for evaluating nanocarrier transport, biodistribution, and therapeutic efficacy. Table 2 provides a comparative overview of in vitro and in vivo models for studying nanotechnology transport across the BBB. Each model offers physiological relevance, experimental control, and throughput balance. In vitro models range from simple immortalized cell monolayers with high scalability but low barrier tightness to advanced hiPSC-derived BBB-on-chip systems that closely recapitulate human BBB properties (with TEER values approaching in vivo levels) [32,33]. Incorporating astrocytes, pericytes, or flow typically enhances barrier integrity and transporter functionality, albeit with increased complexity. In vivo rodent models remain indispensable for demonstrating NP delivery in an intact organism; they provide the full neurovascular unit context and pharmacokinetic realism. However, species differences (e.g., rodent vs. human P-gp expression) must be accounted for [34]. Non-human primates offer the most human-like BBB for late-stage validation, although practical and ethical constraints limit their use. Notably, emerging zebrafish models can expedite early screening, but results should be extrapolated to mammals with caution. In summary, no single model is “best” for all purposes – researchers should choose the model that best fits their experimental goals. Early high-throughput screens can be done in simpler systems (cell lines or zebrafish), medium-complexity models (primary co-cultures, iPSC chips) are ideal for mechanistic insights, and critical translational studies are reserved for rodent and primate models. This strategic, multi-model approach maximizes discovery efficiency and clinical relevance, guiding nanomedicine development across the BBB toward successful human applications.
2.4. Key NP Properties for Crossing the BBB
Delivering therapeutics across the BBB requires NPs with finely tuned physicochemical properties. Size, surface charge, shape, and composition determine a nanocarrier’s ability to traverse the brain endothelium while evading clearance [14,49]. Recent preclinical studies (2020–2025) provide quantitative insights that guide the rational design of BBB-penetrant nanotechnologies, as summarized in Figure 2.
- a)
- Size
NP size profoundly influences BBB transport. Optimal diameters are generally in the tens of nanometers, balancing efficient transcytosis with minimal sequestration by clearance organs. In a mouse model, 20 nm insulin-coated gold NPs (AuNPs) achieved the highest brain accumulation (at 2 hours post-injection) compared to identical 50 nm and 70 nm formulations. The 20 nm AuNPs showed the widest biodistribution in brain tissue. In contrast, larger AuNPs were less effective [50]. Similarly, an in vitro BBB model found that 25 nm PEGylated silica NPs had superior transport efficiency relative to 50 nm and 100 nm particles [51]. In vivo, 25 nm silica NPs (with slight positive surface charge) remained in circulation for over 24 hours and yielded a 6-fold higher brain drug concentration than free drug administration [52]. These small NPs likely exploit transcellular pathways and narrow paracellular gaps between endothelial cells, whereas tight junctions mostly exclude larger particles [53,54].
There are practical size limits at both extremes. Particles <10 nm are rapidly filtered by the kidneys and can diffuse out of the brain almost as quickly as they enter. For instance, ultrasmall ~2 nm gold nanoclusters readily cross the BBB but show non-negligible neurotoxicity and rapid clearance [55,56]. On the other hand, NPs >200 nm are prone to opsonization and sequestration by the reticuloendothelial system (RES) in the liver and spleen, leaving an insufficient fraction to reach the brain. Indeed, 200–250 nm gold NPs were found to accumulate less in the brain (and in systemic organs) than 10–15 nm gold NPs, which distributed more broadly (albeit with higher potential toxicity) [56,57]. These studies indicate an optimal NP size window (roughly 10–100 nm) that maximizes BBB penetration. Within this window, 50–150 nm particles often achieve longer plasma half-lives than tiny ones yet are still small enough to extravasate; for example, ~50 nm polymeric NPs showed deeper brain tissue penetration in a brain injury model than 200 or 800 nm analogues.
Additionally, particle size strongly influences the protein corona composition in circulation, impacting BBB receptor-mediated uptake. For instance, lipid NPs (~30 nm) preferentially bind apolipoprotein E, enhancing LDLR- and LRP-1-mediated transcytosis compared to larger counterparts. Notably, optimal size windows may shift under pathological conditions; ~100 nm polymeric NPs demonstrated superior accumulation in GBM models versus smaller or larger particles [58,59]. Thus, the ideal size range of ~10–100 nm balances efficient receptor targeting, prolonged systemic circulation, and enhanced brain delivery.
- b)
- Surface charge (zeta potential)
The BBB endothelium carries a negatively charged glycocalyx, meaning NP surface charge critically mediates electrostatic interactions with the barrier. Mildly cationic NPs can undergo enhanced adsorptive uptake by BBB cells, but strongly positive charge also risks disrupting the tight junction integrity. Quantitative perfusion studies have shown that neutral or slightly anionic surface charges are optimal for safe BBB transit. Lockman et al. found that neutral NPs and low-concentration anionic formulations preserved rat BBB integrity and achieved brain delivery, whereas highly cationic NPs caused immediate BBB leakage and toxicity. Notably, the brain uptake rate of anionic NPs (at non-disruptive concentrations) exceeded that of neutral or cationic versions in the same model [60]. This suggests a trade-off: positively charged NPs bind avidly to the negatively charged cell membranes, promoting internalization, but excessive positive charge triggers unwanted barrier opening and clearance by proteoglycan-rich regions of the endothelium.
Recent mechanistic studies support a moderate cationic charge for BBB crossing. A 2020 mathematical model incorporating endothelial surface charge predicted that a positively charged NP experiences enhanced transcellular permeability by electrostatic attraction to the negatively charged membrane [61]. Experimentally, cationic liposomes (+30 mV ζ-potential) show higher uptake by brain endothelial cells than equivalent neutral or −30 mV liposomes [62,63]. In one comparative study across eight cell lines, positively charged NPs had the fastest uptake rates vs. negative or neutral NPs [64]. However, high uptake does not guarantee deep brain delivery. Strong cationic particles may stick to the luminal surface or become entrapped in endothelial lysosomes. In contrast, negatively charged NPs (e.g. ~−35 mV) diffuse more freely and can penetrate further into brain parenchyma [62,65]. For example, one report noted that although +30 mV gold NPs enter BBB cells efficiently, -36 mV gold NPs penetrated brain tissue due to greater mobility through the endothelium [63]. The downside is that negative NPs (and uncharged PEGylated NPs) tend to be less readily internalized. In vivo, neutral or PEG-coated NPs often exhibit the longest circulation times; for instance, neutral PEGylated liposomes can circulate for hours. In contrast, highly cationic NPs are rapidly opsonized and cleared from the bloodstream [66]. Slightly negative or near-neutral surfaces thus appear to best balance BBB uptake and systemic stealth. In summary, a zeta potential in the range of –10 to +10 mV (or slightly beyond) is often favorable for BBB-targeted nanocarriers: it provides some affinity for the negatively charged endothelium without causing cytotoxicity or immediate clearance [60]. Fine-tuning surface charge (for example, by incorporating zwitterionic or charge-shielding coatings that become positive only near the membrane) is a strategic approach to maximize brain delivery.
- c)
- Shape
Whether spherical, rod-like, discoidal, or filamentous, NP shape has emerged as a critical design parameter for BBB transit. Shape affects how particles navigate blood flow, how they interface with cell membranes, and how phagocytes recognize them. Recent studies show that non-spherical shapes can improve brain delivery under certain conditions. For instance, polymeric nanorods targeted to transferrin receptors on BBB endothelium exhibited a seven-fold higher cellular uptake than equivalent spheres in an in vitro BBB model [67]. Although that dramatic 7-fold uptake was reported in an earlier study with functionalized rods, even untargeted rods demonstrate advantages in transcytosis efficiency. Nowak et al. (2020) used a human microfluidic BBB model to compare 200 nm spheres vs. rod-shaped particles: while spherical NPs adhered more to the endothelial surface, the rod-shaped NPs translocated across the endothelium about twice as efficiently per cell-bound particle. The authors hypothesized that rods enter endothelial cells via a distinct pathway or orientation that favors vesicular transport [57,68]. In essence, a rod or filamentous shape can “partition” into the cell membrane differently than a sphere, potentially exploiting elongated endocytic pits or aligning with membrane invaginations.
Particle flexibility often goes hand-in-hand with shape. Filamentous micelles and worm-like filomicelles (long and flexible) have demonstrated extraordinarily long circulation and increased accumulation in some tissues, likely by evading phagocytosis. In the brain context, one benefit of discoidal or rod-shaped NPs is improved margination in capillaries; these shapes drift toward the endothelial wall under flow more than spheres, increasing the odds of BBB contact. A 2024 study by Sierri et al. directly compared ~100 nm lipid NPs of identical chemistry but different shapes (spherical vs. discoid vs. deformable “soft” particles) in a human BBB model. Strikingly, discoidal NPs had about double the endothelial permeability of spheres (permeability coefficient ~1.3×10−5 cm/min for discoids vs ~6–7×10−6 for spheres). The deformable (ellipsoidal) NPs were intermediate in BBB crossing. The discoids also traversed intercellular tunneling nanotubes more efficiently, suggesting that shape influences initial BBB crossing and subsequent spread in brain tissue. The superior BBB transit of discoids was attributed to their larger surface area in contact with the cell membrane and favorable orientation during trafficking [69]. Consistent with this, Fu et al. (2022) found that rod-shaped polymeric NPs outperformed spherical NPs in crossing an in vitro brain microvascular model [70]. In summary, elongated or flattened shapes (rods, disks) promote BBB interaction and transcytosis, whereas spherical NPs often remain in circulation or are taken up by the RES. That said, extremes of shape can affect biodistribution: high aspect-ratio filaments might avoid uptake by liver/spleen but could be too large to traverse tight endothelial junctions if very long. Thus, an aspect ratio of roughly 2–5 (standard for short rods or discoids) appears optimal for BBB delivery [57]. By tailoring NP shape, researchers can influence circulation half-life (with filaments lasting longer in blood and enhance margination to brain capillaries, thereby improving the chances of BBB penetration.
- d)
- Stiffness
NP stiffness significantly influences BBB permeability and transcellular transport efficiency. Generally, stiffer NPs exhibit greater endothelial uptake and enhanced BBB penetration compared to softer particles, as evidenced by rigid polystyrene spheres (bulk modulus ~3000 MPa) demonstrating approximately tenfold higher translocation than softer PEG-based hydrogels (~3 MPa) [57,71,72,73].
In vivo GBM models confirm greater accumulation of stiffer NPs within brain tumors than their deformable counterparts [63]. Mechanistically, stiffer particles are more readily endocytosed due to lower membrane deformation requirements, although softer NPs may undergo more efficient intracellular trafficking and exocytosis, facilitating transcytosis. Stiffness also influences vascular margination: rigid particles migrate toward vessel walls, increasing endothelial interactions without disrupting tight junction integrity [57]. Thus, NP stiffness modulation represents a promising strategy for optimizing drug delivery across the BBB.
- e)
- Composition and material class
The composition of NPs (lipidic, polymeric, protein-based, inorganic) significantly influences their BBB permeability and biocompatibility.
Lipid-based NPs: Liposomes and solid lipid NPs (SLNPs) effectively cross the BBB, particularly when functionalized. PEGylated liposomes (~100 nm) have limited penetration alone, but ligand decoration markedly improves uptake. For instance, VCAM-1 peptide-targeted liposomes achieved ~6% injected dose/g brain versus ~1.2% untargeted [74]. Thiamine-functionalized SLNs also increased brain drug concentrations 1.4-fold [75]. Lipid NPs excel in genetic cargo delivery, showing efficient mRNA transfection with reduced off-target accumulation [66].
Polymeric NPs: Biodegradable polymers (PLGA, PLA, dendrimers) commonly enhance BBB transport via PEGylation or targeting ligands. LDL receptor-targeted PLGA NPs achieved ~4% ID/g brain compared to ~1% untargeted [18,76]. Polymeric nanocarriers offer tunable release kinetics and flexible shapes for enhanced penetration and tumor suppression [57].
Protein-based NPs: Ferritin and virus-like particles (VLPs) naturally exploit receptor-mediated pathways. Human H-ferritin effectively delivers antibodies into brain tumors [77]. Peptide-functionalized protein NPs (e.g., RVG29, transferrin, angiopep-2) further enhance receptor-specific uptake [78].
Inorganic NPs: Gold, iron oxide, and silica NPs provide stable cores suitable for functionalization and imaging. Targeted gold NPs (~50 nm) significantly outperform untargeted ones (0.23% vs. 0.04% ID/g) [56]. Iron oxide nanoparticles (SPIONs, <20 nm) achieve brain accumulation suitable for MRI imaging when suitably coated [18]. Moreover, an “optimal” iron oxide size of around 10–50 nm has been identified to maximize circulation time and minimize phagocytic uptake [79]. Silica NPs (~25 nm) form beneficial protein coronas, enhancing BBB penetration [52].
Table 3 presents an overview of recent experimental findings from original research on NP performance in brain delivery. It includes key physicochemical attributes (size, charge, composition), quantitative metrics on brain accumulation (% ID/g), circulation half-life, and therapeutic outcomes across both healthy and pathological BBB models. By contrasting NP types, lipid-based, polymeric, protein-based, and inorganic, under varied surface modifications and targeting strategies, this table is a critical reference for guiding the rational design of nanocarriers optimized for BBB penetration. Overall conclusions highlight that NP functionalization using BBB receptor-specific antibodies or appropriate polymer coatings significantly enhances brain uptake compared to untargeted particles. Optimal performance is typically achieved with NP sizes below 100 nm, carrying neutral or slightly negative surface charges, which facilitates longer circulation half-lives and improved brain retention. However, functionalization alone does not universally guarantee therapeutic efficacy; thus, surface modifications and particle composition must be carefully tailored to maximize clinical outcomes.
The BBB severely restricts brain uptake of systemically administered therapeutics. Innovative nanotechnologies are being developed to ferry drugs across the BBB in meaningful quantities (Figure 3). Recent studies emphasize human-relevant models to assess translational potential, from induced pluripotent stem cell (iPSC)-derived BBB cultures to non-human primates and early clinical trials. Below, we review key nanotechnological strategies for BBB crossing, highlighting quantitative brain delivery metrics (e.g., percentage of injected dose per gram brain, brain:plasma ratios) and therapeutic outcomes (tumor suppression, cognitive improvements) where available.
1. Receptor-Mediated Transcytosis (RMT)
RMT exploits endogenous nutrient/carrier receptors on brain endothelium to shuttle NPs or biologics into the brain. Binding to receptors such as the transferrin receptor (TfR), insulin receptor (INSR), low-density lipoprotein receptor-related protein 1 (LRP1), lactoferrin receptor, or folate receptor triggers internalization and vesicular transport across endothelial cells. This strategy has dramatically improved BBB delivery in multiple models:
1a. Transferrin Receptor (TfR)
The TfR is abundantly expressed on brain microvessels (especially during development) and is a prime RMT target. Classical studies in rats showed that an anti-TfR antibody (OX26) achieved ~0.3% of the injected dose (ID) in the brain, over 10-fold higher than a non-specific IgG (0.03% ID). Recent work has refined TfR-targeting by adjusting antibody affinity/valency. Johnsen et al. demonstrated that gold NPs conjugated with a low-affinity, monovalent anti-TfR delivered 0.23% ID/g into brain parenchyma, versus only 0.04–0.08% ID/g with high-affinity bivalent variants [18]. Likewise, Denali Therapeutics engineered a TfR-binding “Enzyme Transport Vehicle” (ETV) with moderate affinity; in mice, an ETV–IDS (iduronate-2-sulfatase) fusion achieved broad CNS distribution and enzyme uptake by neurons, lowering pathological glycosaminoglycans in brain by 49–76%, compared to only 25–43% reduction using a high-affinity bivalent IgG format. Notably, monovalent TfR carriers avoid “trapping” in endothelial endosomes, yielding higher transcytosis efficiency [91]. In non-human primates, systemically delivered TfR-targeted biologics have demonstrated distributed brain delivery without significant peripheral side effects, validating TfR RMT as a clinically viable route. For example, a bispecific antibody with one TfR-binding arm and one therapeutic arm reached ~0.5–1.1% ID/g in capillaries and significantly enhanced parenchymal delivery [18]. These advances underscore that binding affinity and avidity are critical: too high affinity can impede release into brain parenchyma, whereas optimized moderate affinity promotes deeper delivery.
1b. Insulin receptor (INSR)
The INSR is widely expressed on the BBB endothelium and can mediate brain uptake of insulin and analogues [92]. High-affinity human INSR antibodies (e.g., 83-14) have been used as “Trojan horses” to ferry drugs across the BBB. In a clinical trial for Hunter syndrome, an INSR-targeted enzyme (valanafusp alpha) showed CNS activity, indicating successful BBB penetration [82]. NPs have also leveraged INSR: for instance, HSA (human serum albumin) NPs conjugated with an insulin mimetic (29B4 antibody) penetrated the BBB and elicited CNS therapeutic effects in rodents. Although INSR targeting can risk hypoglycemic signaling, careful engineering (e.g., insulin agonist antibodies that activate transport without strong insulin-like effects) has enabled significant brain delivery [93,94].
1c. LRP1 (Low-Density Lipoprotein Receptor-Related Protein 1)
LRP1 is highly expressed on brain endothelium and shuttles ligands like ApoE and lactoferrin. The peptide Angiopep-2 (Ang-2) binds LRP1 and has been widely used to decorate NPs. Angiopep-functionalization triggers robust transcytosis; an Ang–2–drug conjugate crossed the BBB and accumulated in brain tumors in clinical trials (ANG1005 for glioma). Recent docking studies have optimized Ang-2 analogues for tighter LRP1 binding [95,96]; an Ang-2–drug conjugate crossed the BBB and accumulated in brain tumors in clinical trials (ANG1005 for glioma). Recent docking studies have optimized Ang-2 analogues for tighter LRP1 binding [97]. In one study, an artificial LRP1-binding peptide “L57” showed efficient uptake in primary human brain microvascular endothelial cells, indicating it may serve as a new BBB shuttle [98]. Tubular transcytosis pathways have been observed for LRP1, suggesting unique mechanisms that can be modulated by peptide “shuttles” [99].
1d. Lactoferrin receptor
Lactoferrin (Lf), an iron-binding glycoprotein, crosses the BBB via its receptor on brain endothelium [100,101]. NPs coated with lactoferrin exploit this route. For example, Youssef et al. (2025) coated lipid nanocapsules with Lf to deliver the antioxidant drug apocynin (APO) and lavender oil in a rat epilepsy model. The Lf-coated nanocapsules achieved greater brain accumulation of APO and significantly reduced seizure severity, yielding a Racine score of ~0.67 (near absence of seizures) versus much higher scores in uncoated NP or free drug groups. Lf-NPs also prolonged seizure latency and lowered neuroinflammatory markers, indicating effective therapeutic delivery to the brain [101]. Similarly, oral Lf-decorated gold NPs have been studied for GBM, given that LfR is upregulated in many tumors [102]. These results highlight lactoferrin’s potential as an endogenous targeting ligand to improve drug BBB permeability.
1e. Folate receptor
Folate receptor-α is minimally expressed on the intact BBB, but it is present in the choroid plexus and often overexpressed in brain tumor cells (gliomas) [103]. Folic acid is a popular ligand targeting NPs (Y. Wu et al., 2023), especially targeting metastatic or leptomeningeal tumor deposits. In glioma-bearing models, folate-targeted NPs accumulate in tumors via folate receptor-mediated endocytosis [104]. For instance, folate-coated gold NPs loaded with indomethacin showed enhanced uptake in FR-positive glioma cells and extended survival in tumor-bearing mice [105]. While folate RMT is not a significant route into the normal brain, it provides a tumor-specific BBB bypass, improving drug delivery to FR-rich brain tumors without affecting normal brain tissue.
To maximize RMT delivery, researchers combine ligands (e.g., TfR antibody plus a cell-penetrating peptide or RVG peptide) on the same NP [78]. These multi-target NPs aim to engage multiple uptake pathways synergistically. Overall, RMT-based nanocarriers have achieved measurable brain uptake on the order of 0.1–0.5% ID/g in rodents [106], which, while modest in absolute terms, represents an order-of-magnitude improvement over untargeted delivery. More importantly, these carriers have shown therapeutic efficacy in disease models, from enzyme replacement in lysosomal disease (reducing CNS pathology) to drug delivery in brain tumors (inhibiting growth), underscoring the translational promise of RMT nanotechnologies [91].
2. Adsorptive-Mediated Transcytosis (AMT)
Adsorptive-mediated transcytosis relies on electrostatic attraction of cationic molecules to the negatively charged endothelial membrane, inducing nonspecific endocytosis. Unlike RMT, AMT does not require specific receptors, making it broadly applicable. Various cationic coatings and cell-penetrating peptides (CPPs) have been used to trigger AMT for brain delivery:
2a. Cationic cell-penetrating peptides
Poly-arginine peptides (such as R9), the TAT peptide (derived from the HIV transactivator protein), and penetratin (from Antennapedia) carry multiple positive charges that promote adsorption to the BBB surface. For example, NPs functionalized with PepH3, a 7-amino-acid cationic peptide derived from dengue virus capsid, showed greatly enhanced uptake in BBB models. PepH3-tagged NPs exhibited active transcytosis across both rat and human BBB cell monolayers [107,108]. In vivo, radiolabeled PepH3 derivatives achieved high brain uptake with low accumulation in liver, lung, and kidney, indicating a degree of selectivity [107,109]. Notably, making the endothelial surface less negative (by enzymatically removing glycocalyx sialic acids) reduced PepH3-NP uptake, confirming that electrostatic interactions are the primary driver. These findings show that cationic shuttles can exploit AMT to cross the BBB, and some (like PepH3) may do so with minimal off-target deposition [108]. Other CPPs, such as penetratin and poly(arginine) -8, have similarly increased NP translocation in vitro and in situ (e.g., brain perfusion models) by several-fold compared to unmodified NPs. However, in vivo quantitation is less common.
2b. Cationic polymers (e.g., chitosan)
Chitosan, a polycationic polysaccharide, has been widely used to decorate NPs for BBB delivery as recently reviewed by [110]. Its positive charge and mucoadhesive properties facilitate both AMT and modulation of tight junctions. Khan et al. (2023) demonstrated that DNA-loaded chitosan nanoparticles (NPs, ~260 nm, with a positive ζ potential) could effectively transfect brain cells in vivo after systemic administration. In their study, GFP-encoded chitosan NPs injected intraperitoneally in mice showed GFP expression in the brain, confirming that the NPs crossed the BBB. The brain delivery was achieved without the use of chemical targeting ligands; the chitosan’s inherent AMT property was sufficient. Notably, the chitosan vectors showed low cytotoxicity and immunogenicity in vitro (U87 glioma cells had ~85% viability) and no apparent toxicity in vivo [111]. These data position chitosan NPs as safe, efficient gene delivery vehicles to the CNS, leveraging adsorptive uptake. Other cationic polymers (e.g., polyethyleneimine, cationic dendrimers) also promote BBB transit, though toxicity must be carefully managed.
2c. Cationic surface coatings
Even without distinct peptides or polymers, tuning an NP’s surface charge can impact BBB uptake. Slightly positive or even “near-neutral” (mildly negative) zeta potentials favor BBB transcytosis [106]. For instance, one study found that rod-shaped polymeric NPs with a mildly negative surface had optimal uptake in brain endothelial cells (7-fold higher than neutral NPs) [67,112]. In practice, some researchers coat NPs with cell membranes (from e.g., leukocytes or platelets) to confer biological identity and charge that enhance BBB passage via a combination of AMT and other mechanisms. The general principle is that increasing NP affinity for the endothelial membrane (through charge or hydrophobic patches) can initiate vesicular transport across the BBB.
Overall, AMT-based approaches often lack the absolute specificity of RMT, but they offer versatility. They are beneficial for delivering large macromolecular complexes or gene vectors that might not fit into a single receptor pathway. By combining AMT peptides with targeting ligands (dual-function NPs), researchers aim to achieve both high uptake and specific delivery. The main quantitative limitation of AMT is potential sequestration in endothelial cells or perivascular spaces; however, evidence from studies like the chitosan NP study shows that a meaningful fraction of the dose can reach the parenchyma to exert biological effects.
3. Magnetically guided NPs
Magnetic targeting employs external magnetic fields to direct superparamagnetic iron oxide NPs (SPIONs) across the BBB, concentrating them in specific brain regions such as tumors. SPIONs coated with surfactants (e.g., Tween) have successfully crossed the intact BBB in rats when guided by magnets, with negligible uptake observed without magnetic fields [113,114,115].
3a. Magnetic liposomes
In glioma therapy, magnetic liposomes containing temozolomide and SPIONs demonstrated significant tumor accumulation, slowed tumor growth, and modestly improved survival under magnetic guidance compared to controls. These NPs also acted as MRI contrast agents without observed toxicity [114].
3b. Magnetic hyperthermia combo
Combining magnetic targeting with hyperthermia via alternating magnetic fields (AMF) further enhances therapeutic outcomes. A recent study utilized Fe₃O₄@Chitosan@ZIF-8@RVG29 nanoparticles, resulting in effective glioma cell killing and tumor apoptosis [116].
Magnetic targeting significantly increases drug concentration and spatial precision in brain tumors. Despite requiring specialized equipment and facing limitations in magnetic field penetration, ongoing clinical trials suggest this approach as a promising strategy for overcoming the BBB.
4. Virotechnological strategies
Viruses naturally evolved to penetrate biological barriers, including the BBB (some neurotropic viruses cross via receptor-mediated mechanisms). Engineered viral vectors and virus-like particles leverage this capability for drug and gene delivery:
4a. Adeno-Associated Viruses (AAV)
AAV vectors (∼25 nm) are leading vehicles for gene delivery. Specific serotypes (e.g., AAV9) can cross the BBB, especially in neonatal mice or when the BBB is mildly perturbed. However, BBB transduction by natural AAVs is very limited in adult primates. Breakthrough work from 2020 to 2023 has resulted in engineered AAV capsids with enhanced blood-brain barrier (BBB) penetration. Chuapoco and colleagues developed AAV.CAP-Mac was selected by iterative screening for marmosets and macaques. In adult rhesus monkeys, AAV.CAP-Mac achieved widespread brain transgene expression after IV injection, transducing ~1.3% of cortical neurons, compared to only 0.5% with conventional AAV9.CAP-Mac showed broad tropism (neurons and some glia) and delivered functional genes (e.g., GCaMP for neural imaging) across multiple brain areas [117]. Another group created AAV-F (AAV.PHP.eB variants)* that crosses the BBB in mice by interacting with LY6A on endothelial cells, though these do not translate to primates. In 2023, an AAV capsid was reported that binds human TfR1 to ferry genes across the BBB. Such TfR-targeted AAVs increased mouse CNS gene delivery and showed uptake in ex vivo human BBB models [118]. The quantitative gains are striking; one variant (AAV.CAP-B10) gave 3–4 times higher neuronal transduction in macaques than AAV9, with much lower liver off-targeting. These advances suggest that IV AAV gene therapy for the brain is becoming feasible. Indeed, systemic AAV9 is already used in infants (e.g., onasemnogene for SMA) to deliver genes to spinal motor neurons, leveraging a partially immature BBB. Ongoing clinical studies are exploring AAV capsids for adults with Alzheimer’s and Parkinson’s, aiming to achieve a few percentage points of CNS targeting, enough for therapeutic benefit in many cases [117].
4b. Lentiviral and other viral vectors
Lentiviruses (e.g., modified HIV-1) and adenoviruses can transduce brain cells but generally do not cross the BBB efficiently when given IV. Hence, they have been used via direct injection into the brain or CSF. Recent innovations involve coating or retargeting these larger viruses. For example, a lentivirus pseudotyped with rabies glycoprotein (RVG) was shown to reach CNS neurons from the bloodstream in mice, using RVG’s ability to interact with nicotinic acetylcholine receptors on BBB endothelium [116]. Another approach uses exosomes to “piggyback” lentiviral vectors (see hybrid exosome section). While systemic lentiviral delivery to the brain isn’t yet routine, the field is moving toward producing virus-like particles decorated with BBB-targeting ligands.
4c. Virus-Like Particles (VLPs)
These are self-assembled protein cages derived from viruses, but without any viral genome, essentially “nanocontainers” that can carry drugs or genes. VLPs from various sources have been tested for brain delivery, including bacteriophage Qβ and MS2, plant viruses like Tobacco Mosaic Virus (TMV) and Cowpea Chlorotic Mottle Virus (CCMV), and human virus capsids (e.g., hepatitis B core). VLPs typically range 20–150 nm, a size amenable to crossing fenestrated barriers and, with modifications, potentially the BBB. For instance, TMV nanorods (300×18 nm) were albumin-coated to prolong circulation and successfully used to image and treat brain tumors in mice [68,119]. The filamentous shape of TMV may aid transport along endothelial cells. Qβ VLPs, which are ~30 nm icosahedra, have been functionalized with peptides such as angiopep or transferrin to engage RMT, effectively combining VLPs with the RMT approach. One study reports that exosomes loaded with a VLP carrying rhodamine dye crossed the BBB and increased brain drug delivery [120], illustrating a hybrid of VLP and exosome strategies. While still preclinical, VLPs offer high customizability (both genetic and chemical) to display targeting ligands and can be produced in high yields from plants or bacteria. They also tend to be biodegradable and less immunogenic than whole viruses. The challenge is achieving efficient BBB traversal; current VLPs require surface functionalization. Nonetheless, VLPs are a versatile platform, such as a ~120 nm hepatitis B VLP, which has been used to carry siRNA and target brain metastases, showing improved survival in mice (by homing to a tumor antigen and releasing siRNA upon endocytosis).
In summary, viral vectors have the advantage of active transport mechanisms (receptor- or cell fusion-mediated) that can be highly potent; for example, a single dose of AAV can transduce millions of neurons if it crosses the blood-brain barrier (BBB). The quantitative goal often cited is achieving a few percent of the total brain cell population transduced or a brain-to-serum drug ratio approaching 0.1; recent viral innovations are closing in on these targets [117]. Meanwhile, non-pathogenic VLPs offer a safer alternative, such as nanocarriers, that can utilize viral entry mechanisms without replicating them. Both are promising for diseases requiring gene therapy, enzyme replacement, or widespread neuromodulation.
5. Exosomes and extracellular vesicles
Exosomes (30–150 nm extracellular vesicles) naturally cross biological barriers and mediate intercellular communication, making them attractive endogenous delivery vehicles for the brain [121]. They possess cell-derived membrane proteins that can confer innate brain tropism; for instance, exosomes from neurons or macrophages may preferentially home to the brain. Key features of exosomes include biocompatibility, low immunogenicity, the ability to be loaded with drugs or RNA, and the flexibility to engineer their surface. Recent developments include:
5a. Intrinsic brain tropism
Certain exosomes inherently migrate to the brain. MSC (mesenchymal stem cell)-derived exosomes have been shown to cross the BBB in models of stroke and neuroinflammation. In a mouse stroke model, IV MSC exosomes enhanced functional recovery by improving neurogenesis and synaptic remodeling in the brain [122]. This therapeutic benefit implies significant vesicle delivery to brain tissue. Tissue distribution studies often find exosome uptake in the brain 2–5 fold higher than equivalent doses of free drug [120]. Moreover, exosomes can cross the BBB without disrupting it; they likely use native endocytosis/exocytosis pathways.
5b. Surface-engineered exosomes
To enhance targeting, researchers decorate exosome membranes with peptides or antibodies. A prominent example is the exosome displaying the RVG peptide (a rabies virus glycoprotein fragment that binds to nicotinic receptors). Cui et al. (2019) report that RVG-tagged exosomes, loaded with therapeutic cargo, showed enriched delivery to the cortex and hippocampus after IV injection, significantly improving learning and memory in Alzheimer’s mice. In that study, the RVG-exosomes (carrying siRNA to knockdown BACE1) restored cognitive function in a murine Alzheimer’s model, whereas non-targeted exosomes had minimal effect. Another group used vesicular stomatitis virus G protein (VSV-G) on exosomes to target brain endothelium, achieving successful gene delivery across the BBB in mice (pseudotyping exosomes with viral fusogens can create a hybrid between viral vector and natural vesicle) [123]. These approaches highlight that by displaying targeting ligands (such as RVG, RGD, and antibodies), exosomes can be directed to specific brain regions or cell types, much like synthetic nanoparticles, but with a biological cloak.
5c. Drug delivery and hybrids
Exosomes have been loaded with chemotherapeutics, proteins, or siRNA for brain delivery. Haney et al. demonstrated that exosomes can transport the anti-cancer drug doxorubicin and even large antibodies across the blood-brain barrier (BBB). For glioblastoma, exosomes carrying the antibody cetuximab (targeting EGFR) plus doxorubicin significantly increased antibody brain accumulation and inhibited tumor growth compared to free cetuximab plus the drug. Exosomes delivered more than double the amount of antibody to brain tissue compared to direct antibody injection in that study, due to protected transport and possibly transferrable membrane proteins aiding in BBB transit [124]. Beyond using natural exosomes, “hybrid” NPs are being designed, for example, by coating polymeric NPs with exosome membranes (sometimes referred to as cell-derived nanovesicles) to endow them with exosomal homing abilities [120]. These hybrids have shown increased brain uptake and reduced clearance by the mononuclear phagocyte system. One study mentions glioma-targeted exosomes delivering siRNA to suppress oncogenes, crossing the BBB, and prolonging animal survival [125].
Exosome-based delivery is already moving to clinical testing. A first-in-human trial used autologous exosomes loaded with curcumin to treat patients with brain cancer and showed good safety at high doses (though efficacy is still under evaluation). The primary challenges are scalable production and efficient cargo loading. However, since exosomes can be derived from patient cells, they offer a personalized and highly biocompatible mode of therapy. They naturally avoid P-glycoprotein efflux and can even regulate BBB permeability (some brain endothelial uptake of exosomes triggers signaling that temporarily loosens tight junctions) [121] with their ability to cross the BBB and deliver functional payloads (e.g., restoring memory in AD models [123], exosomes are poised to become a powerful tool in the neurotherapeutic arsenal.
Active transport nanomotors represent an emerging approach for BBB traversal, with early evidence suggesting significantly improved drug penetration compared to passive methods. The continued development of these active, targeted, and responsive systems holds substantial promise for advancing BBB drug delivery.
Table 4 summarizes nanotechnological strategies for crossing the BBB, detailing delivery vehicles, targeting mechanisms, experimental models, precise quantitative brain uptake metrics, and therapeutic outcomes. Receptor-mediated transcytosis (RMT) is a clinically validated strategy, demonstrating robust brain delivery efficiencies (~0.1–1% ID/g) and significant therapeutic efficacy. Notably, transferrin receptor (TfR)-targeted NPs, including T7-functionalized PLGA NPs, achieve approximately sixfold increases in brain delivery, with measurable improvements in therapeutic outcomes in stroke and glioma models. Angiopep-2-functionalized lipid–silica NPs similarly doubled drug concentrations in the brain, highlighting the advantage of moderate receptor affinity ligands such as LRP1 for brain-targeted therapies.
2.5. Toxicological and Regulatory Considerations for Translational Advancement of BBB-Crossing Nanobiomaterials
A significant limitation in the clinical translation of BBB-crossing nanobiomaterials is the scarcity of robust clinical data. Translational medicine faces substantial barriers, primarily due to inadequate characterization of safety and efficacy profiles. A critical step forward involves implementing well-designed preclinical studies following a Quality by Design (QbD) approach, ensuring rigorous evaluation of critical attributes before clinical trials.
Before progressing to clinical application, CNS-targeted nanomedicines must undergo rigorous safety and regulatory assessments. To facilitate this essential stage, Table 5 provides an organized overview of critical pre-in vivo toxicological evaluations, including hemocompatibility, neurotoxicity, and long-term accumulation, along with specific regulatory references and recommended methodologies. This structured approach ensures comprehensive characterization and regulatory compliance for NPs designed to cross the BBB.
Hemocompatibility assessments (hemolysis <5%, minimal platelet activation, and low complement activation) ensure intravenous safety, mitigating risks of thrombotic and immune-mediated adverse events [152]. For neurotoxicity, CNS-targeted NPs should demonstrate no significant behavioral impairments or histopathological abnormalities at therapeutic doses, aligning with safety pharmacology guidelines [153,154]. Moreover, long-term accumulation studies must confirm that NPs are cleared from the CNS over time, ideally exhibiting >90% elimination within weeks post-administration, thereby minimizing delayed neuroinflammation or chronic toxicity [155].
Regulatory bodies (FDA and EMA) mandate comprehensive biodistribution and pharmacokinetic profiling, necessitating quantitative in vivo analyses using labeled nanomaterials to map NP fate, organ retention, and BBB crossing efficiency [156]. Likewise, immunotoxicity evaluations must preclude excessive immune activation or complement-mediated reactions (CARPA), verifying that anti-NP antibodies or cytokine storms are absent or manageable [157].
Finally, achieving manufacturing reproducibility during scale-up is pivotal. Establishing and maintaining Critical Quality Attributes (CQAs), such as particle size, polydispersity (PDI), zeta potential, and drug loading, across batches ensures consistent therapeutic outcomes and regulatory compliance [158]. Nanomedicine developers should proactively define and validate manufacturing processes to control batch-to-batch variability, leveraging GMP practices and possibly advanced techniques like microfluidics to maintain quality during scale transitions. Collectively, these data-driven recommendations form a robust foundation for safely advancing CNS-targeted NPs from preclinical validation towards clinical implementation.
3. Conclusions
This comprehensive guidance establishes a practical and rigorous framework for rationally designing and validating nanobiotechnological systems capable of effectively traversing the BBB. The summarized tables delineate key parameters for optimal NP physicochemical properties, targeting mechanisms, and validated experimental models, guiding researchers through informed and methodical design decisions. By integrating detailed considerations of NP size, charge, composition, and regulatory and toxicological standards, this guide enables precision engineering of nanocarriers that demonstrate improved CNS targeting, therapeutic efficacy, and safety profiles.
However, significant translational hurdles remain despite these structured approaches and technical advances. Future research must systematically address regulatory compliance, scale-up reproducibility, and rigorous safety validation, explicitly highlighted through recommended toxicological assays, immunogenicity assessments, and pharmacokinetic evaluations in the tables. By proactively addressing these practical considerations and rigorously validating nanoparticle performance in biologically relevant models, nanomedicine can more effectively overcome current barriers, accelerating the clinical realization of innovative therapies for neurological disorders.
Author Contributions
LNM: Conceptualization, investigation, formal analysis, methodology, writing, and editing the original draft. MPG, EPC, AGO, IGA: Investigation, formal analysis, methodology, editing of the original draft. FVF: Conceptualization, writing and editing the original draft, funding acquisition, formal analysis, and methodology.
Funding
This work was made possible thanks to the financial support of the Instituto Politécnico Nacional [Projects SIP20242814, SIP20250306, and SIP20251096] and the Secretaría de Investigación y Posgrado. Convocatoria de Apoyos económicos para Publicaciones y Trámite de Invenciones. MPG, LNM, and EPC received scholarships from SECIHTI (2051579, 2046158, and 2048244, respectively).
Acknowledgments
We thank Rodolfo Guillermo Valle Altamirano, Luis Manuel Vázquez Sánchez, and Alan Fabricio Mendoza Peralta (CICATA Unidad Morelos, IPN) for their technical support. ChatGPT-4 was used exclusively to assist with language editing, grammar refinement, and clarity improvements during the preparation of this manuscript.
Conflicts of Interest
The authors declare that the research was conducted without commercial or financial relationships that could create a conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BBB | Blood–Brain Barrier |
| NP | Nanoparticle |
| CNS | Central Nervous System |
| RMT | Receptor-Mediated Transcytosis |
| AMT | Adsorptive-Mediated Transcytosis |
| TfR | Transferrin Receptor |
| LDLR | Low-Density Lipoprotein Receptor |
| LRP-1 | Low-Density Lipoprotein Receptor-Related Protein 1 |
| GLUT1 | Glucose Transporter 1 |
| LAT1 | Large Neutral Amino Acid Transporter 1 |
| TAT | Trans-Activator of Transcription |
| hCMEC/D3 | Human Cerebral Microvascular Endothelial Cell Line/D3 |
| hiPSC | Human Induced Pluripotent Stem Cell |
| SPION | Superparamagnetic Iron Oxide Nanoparticle |
| GBM | Glioblastoma Multiforme |
| TEER | Transendothelial Electrical Resistance |
| PEG | Polyethylene Glycol |
| RES | Reticuloendothelial System |
| VLP | Virus-Like Particle |
| AuNPs | Gold Nanoparticles |
| APO | Apocynin |
| AAV | Adeno-Associated Virus |
| RVG | Rabies Virus Glycoprotein |
| MSC | Mesenchymal Stem Cell |
| CPP | Cell-Penetrating Peptide |
| CNS | Central Nervous System |
| NHP | Non-Human Primate |
| CSF | Cerebrospinal Fluid |
| TEM | Transmission Electron Microscopy |
| MRI | Magnetic Resonance Imaging |
| PET | Positron Emission Tomography |
| EMA | European Medicines Agency |
| FDA | Food and Drug Administration |
| GMP | Good Manufacturing Practice |
| CQAs | Critical Quality Attributes |
| CARPA | Complement Activation-Related Pseudoallergy |
| PK | Pharmacokinetics |
| ADME | Absorption, Distribution, Metabolism, and Excretion |
| PBPK | Physiologically Based Pharmacokinetic |
| ISO | International Organization for Standardization |
| ASTM | American Society for Testing and Materials |
| ICH | International Council for Harmonisation |
| LAL | Limulus Amebocyte Lysate |
| CMC | Chemistry, Manufacturing and Controls |
| PDI | Polydispersity Index |
| ABC | Accelerated Blood Clearance |
References
- World Health Organization. Dementia. Available online: https://www.who.int/news-room/fact-sheets/detail/dementia (accessed on 26 Feburary 2020).
- Trinh, D.-L.; Kim, S.-H.; Yang, H.-J.; Lee, G.-S. The Efficacy of Shape Radiomics and Deep Features for Glioblastoma Survival Prediction by Deep Learning. Electronics 2022, 11, 1038. [Google Scholar] [CrossRef]
- Poursaeed, R.; Mohammadzadeh, M.; Safaei, A.A. Survival prediction of glioblastoma patients using machine learning and deep learning: a systematic review. BMC Cancer 2024, 24, 1581. [Google Scholar] [CrossRef]
- Pardridge, WM. The blood-brain barrier: Bottleneck in brain drug development. Neurotherapeutics. 2005 Jan;2(1):3–14.
- Chew, K.S.; Wells, R.C.; Moshkforoush, A.; Chan, D.; Lechtenberg, K.J.; Tran, H.L.; Chow, J.; Kim, D.J.; Robles-Colmenares, Y.; Srivastava, D.B.; et al. CD98hc is a target for brain delivery of biotherapeutics. Nat. Commun. 2023, 14, 5053. [Google Scholar] [CrossRef]
- Wang, F.; Qi, L.; Zhang, Z.; Duan, H.; Wang, Y.; Zhang, K.; Li, J. The Mechanism and Latest Research Progress of Blood–Brain Barrier Breakthrough. Biomedicines 2024, 12, 2302. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.-Y.; Kang, Y.-S. The decrease of paclitaxel efflux by pretreatment of interferon-γ and tumor necrosis factor-α after intracerebral microinjection. Brain Res. 2013, 1499, 158–162. [Google Scholar] [CrossRef]
- Terstappen, G.C.; Meyer, A.H.; Bell, R.D.; Zhang, W. Strategies for delivering therapeutics across the blood–brain barrier. Nat. Rev. Drug Discov. 2021, 20, 362–383. [Google Scholar] [CrossRef]
- Voth, B.; Nagasawa, D.T.; Pelargos, P.E.; Chung, L.K.; Ung, N.; Gopen, Q.; Tenn, S.; Kamei, D.T.; Yang, I. Transferrin receptors and glioblastoma multiforme: Current findings and potential for treatment. J. Clin. Neurosci. 2015, 22, 1071–1076. [Google Scholar] [CrossRef]
- Jiménez, A.; Estudillo, E.; Guzmán-Ruiz, M.A.; Herrera-Mundo, N.; Victoria-Acosta, G.; Cortés-Malagón, E.M.; López-Ornelas, A. Nanotechnology to Overcome Blood–Brain Barrier Permeability and Damage in Neurodegenerative Diseases. Pharmaceutics 2025, 17, 281. [Google Scholar] [CrossRef] [PubMed]
- Xie, A.; Cheng, G.; Wu, J.; Li, Z.; Yu, G.; Zhu, X.; Chen, T. Highly BBB-permeable nanomedicine reverses neuroapoptosis and neuroinflammation to treat Alzheimer's disease. Biomaterials 2025, 312, 122749. [Google Scholar] [CrossRef]
- Zha, S.; Liu, H.; Li, H.; Li, H.; Wong, K.-L.; All, A.H. Functionalized Nanomaterials Capable of Crossing the Blood–Brain Barrier. ACS Nano 2024, 18, 1820–1845. [Google Scholar] [CrossRef]
- Marrocco, F.; Falvo, E.; Mosca, L.; Tisci, G.; Arcovito, A.; Reccagni, A.; Limatola, C.; Bernardini, R.; Ceci, P.; D’aLessandro, G.; et al. Nose-to-brain selective drug delivery to glioma via ferritin-based nanovectors reduces tumor growth and improves survival rate. Cell Death Dis. 2024, 15, 26. [Google Scholar] [CrossRef]
- Villanueva-Flores, F.; Garcia-Atutxa, I.; Santos, A.; Armendariz-Borunda, J. Toward a New Generation of Bio-Scaffolds for Neural Tissue Engineering: Challenges and Perspectives. Pharmaceutics 2023, 15, 1750. [Google Scholar] [CrossRef] [PubMed]
- Kaya, S.; Callan, B.; Hawthorne, S. Non-Invasive, Targeted Nanoparticle-Mediated Drug Delivery across a Novel Human BBB Model. Pharmaceutics 2023, 15, 1382. [Google Scholar] [CrossRef]
- Pardridge, W.M. Blood–brain barrier endogenous transporters as therapeutic targets: a new model for small molecule CNS drug discovery. Expert Opin. Ther. Targets 2015, 19, 1059–1072. [Google Scholar] [CrossRef]
- Zlokovic, B.V. Neurovascular pathways to neurodegeneration in Alzheimer's disease and other disorders. Nat. Rev. Neurosci. 2011, 12, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.S.; Routhe, L.J.; Moos, T. The vascular basement membrane in the healthy and pathological brain. J. Cereb. Blood Flow Metab. 2017, 37, 3300–3317. [Google Scholar] [CrossRef]
- Uemura, M.T.; Maki, T.; Ihara, M.; Lee, V.M.Y.; Trojanowski, J.Q. Brain Microvascular Pericytes in Vascular Cognitive Impairment and Dementia. Front. Aging Neurosci. 2020, 12, 80. [Google Scholar] [CrossRef]
- Aird, WC. Phenotypic Heterogeneity of the Endothelium: I. Structure, Function, and Mechanisms. Circulation Research. 2007 Feb 2;100(2):158–73.
- Liu, C.; Wu, J.; Zou, M.-H. Activation of AMP-activated protein kinase alleviates High-glucose-induced dysfunction of brain microvascular endothelial cell tight-junction dynamics. Free. Radic. Biol. Med. 2012, 53, 1213–1221. [Google Scholar] [CrossRef]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Zhu, N.; Wei, M.; Yuan, L.; He, X.; Chen, C.; Ji, A.; Zhang, G. Claudin-5 relieves cognitive decline in Alzheimer’s disease mice through suppression of inhibitory GABAergic neurotransmission. Aging 2022, 14, 3554–3568. [Google Scholar] [CrossRef]
- Heffron, T.P.; Salphati, L.; Alicke, B.; Cheong, J.; Dotson, J.; Edgar, K.; Goldsmith, R.; Gould, S.E.; Lee, L.B.; Lesnick, J.D.; et al. The Design and Identification of Brain Penetrant Inhibitors of Phosphoinositide 3-Kinase α. J. Med. Chem. 2012, 55, 8007–8020. [Google Scholar] [CrossRef] [PubMed]
- Bicker, J.; Alves, G.; Fortuna, A.; Falcão, A. Blood–brain barrier models and their relevance for a successful development of CNS drug delivery systems: A review. Eur. J. Pharm. Biopharm. 2014, 87, 409–432. [Google Scholar] [CrossRef]
- Sasson, E.; Anzi, S.; Bell, B.; Yakovian, O.; Zorsky, M.; Deutsch, U.; Engelhardt, B.; Sherman, E.; Vatine, G.; Dzikowski, R.; et al. Nano-scale architecture of blood-brain barrier tight-junctions. eLife 2021, 10, e63253. [Google Scholar] [CrossRef]
- Storelli, F.; Billington, S.; Kumar, A.R.; Unadkat, J.D. Abundance of P-Glycoprotein and Other Drug Transporters at the Human Blood-Brain Barrier in Alzheimer’s Disease: A Quantitative Targeted Proteomic Study. Clin. Pharmacol. Ther. 2021, 109, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, F.M.; Markert, G.; Deutsch, G.; Antonara, M.; Faaij, N.; Bartelink, I.; Noske, D.; Vandertop, W.P.; Bender, A.; Westerman, B.A. Explaining Blood–Brain Barrier Permeability of Small Molecules by Integrated Analysis of Different Transport Mechanisms. J. Med. Chem. 2023, 66, 7253–7267. [Google Scholar] [CrossRef] [PubMed]
- Di L, Rong H, Feng B. Demystifying Brain Penetration in Central Nervous System Drug Discovery: Miniperspective. J Med Chem. 2013 Jan 10;56(1):2–12.
- Zhang, W.; Liu, Q.Y.; Haqqani, A.S.; Leclerc, S.; Liu, Z.; Fauteux, F.; Baumann, E.; Delaney, C.E.; Ly, D.; Star, A.T.; et al. Differential expression of receptors mediating receptor-mediated transcytosis (RMT) in brain microvessels, brain parenchyma and peripheral tissues of the mouse and the human. Fluids Barriers CNS 2020, 17, 47. [Google Scholar] [CrossRef]
- Olivieri Jr PH, Assis IF, Lima AF, Hassan SA, Torquato RJS, Hayashi JY, et al. Glycocalyx Interactions Modulate the Cellular Uptake of Albumin-Coated Nanoparticles. ACS Appl Bio Mater. 2024 Nov 18;7(11):7365–77.
- Ozgür, B.; Puris, E.; Brachner, A.; Appelt-Menzel, A.; Oerter, S.; Balzer, V.; Holst, M.R.; Christiansen, R.F.; Hyldig, K.; Buckley, S.T.; et al. Characterization of an iPSC-based barrier model for blood-brain barrier investigations using the SBAD0201 stem cell line. Fluids Barriers CNS 2023, 20, 96. [Google Scholar] [CrossRef]
- Park, T.-E.; Mustafaoglu, N.; Herland, A.; Hasselkus, R.; Mannix, R.; FitzGerald, E.A.; Prantil-Baun, R.; Watters, A.; Henry, O.; Benz, M.; et al. Hypoxia-enhanced Blood-Brain Barrier Chip recapitulates human barrier function and shuttling of drugs and antibodies. Nat. Commun. 2019, 10, 2621. [Google Scholar] [CrossRef]
- Verscheijden, L.F.M.; Koenderink, J.B.; de Wildt, S.N.; Russel, F.G.M. Differences in P-glycoprotein activity in human and rodent blood–brain barrier assessed by mechanistic modelling. Arch. Toxicol. 2021, 95, 3015–3029. [Google Scholar] [CrossRef]
- Kanjanasirirat, P.; Saengsawang, W.; Ketsawatsomkron, P.; Asavapanumas, N.; Borwornpinyo, S.; Soodvilai, S.; Hongeng, S.; Charoensutthivarakul, S. GDNF and cAMP significantly enhance in vitro blood-brain barrier integrity in a humanized tricellular transwell model. Heliyon 2024, 10, e39343. [Google Scholar] [CrossRef]
- Masuda T, Hoshiyama T, Uemura T, Hirayama-Kurogi M, Ogata S, Furukawa A, et al. Large-Scale Quantitative Comparison of Plasma Transmembrane Proteins between Two Human Blood–Brain Barrier Model Cell Lines, hCMEC/D3 and HBMEC/ciβ. Mol Pharmaceutics. 2019;16(5):2162–71.
- Monteiro AR, Barbosa DJ, Remião F, Silva R. Co-Culture Models: Key Players in In Vitro Neurotoxicity, Neurodegeneration and BBB Modeling Studies. Biomedicines. 2024 Mar 12;12(3):626.
- Weksler, B.; A Romero, I.; Couraud, P.-O. The hCMEC/D3 cell line as a model of the human blood brain barrier. Fluids Barriers CNS 2013, 10, 16. [Google Scholar] [CrossRef]
- Fujimoto, T.; Morofuji, Y.; Nakagawa, S.; Kovac, A.; Horie, N.; Izumo, T.; Niwa, M.; Matsuo, T.; Banks, W.A. Comparison of the rate of dedifferentiation with increasing passages among cell sources for an in vitro model of the blood–brain barrier. J. Neural Transm. 2020, 127, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Neuhoff, S.; Yeo, K.R.; Barter, Z.; Jamei, M.; Turner, D.B.; Rostami-Hodjegan, A. Application of permeability-limited physiologically-based pharmacokinetic models: Part I–digoxin pharmacokinetics incorporating P-glycoprotein-mediated efflux. J. Pharm. Sci. 2013, 102, 3145–3160. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.S.; Humle, N.; Hede, E.; Moos, T.; Burkhart, A.; Thomsen, L.B.; A Eugenin, E. The blood-brain barrier studied in vitro across species. PLOS ONE 2021, 16, e0236770. [Google Scholar] [CrossRef]
- Ribeiro, M.M.B.; Domingues, M.M.; Freire, J.M.; Santos, N.C.; Castanho, M.A.R.B. Translocating the blood-brain barrier using electrostatics. Front. Cell. Neurosci. 2012, 6, 27659. [Google Scholar] [CrossRef]
- Zidarič, T.; Gradišnik, L.; Velnar, T. Astrocytes and human artificial blood-brain barrier models. Bosn. J. Basic Med Sci. 2022, 22, 651–672. [Google Scholar] [CrossRef]
- Uchida, Y.; Wakayama, K.; Ohtsuki, S.; Chiba, M.; Ohe, T.; Ishii, Y.; Terasaki, T. Blood-Brain Barrier Pharmacoproteomics-Based Reconstruction of the In Vivo Brain Distribution of P-Glycoprotein Substrates in Cynomolgus Monkeys. J. Pharmacol. Exp. Ther. 2014, 350, 578–588. [Google Scholar] [CrossRef]
- Xia, C.Q.; Xiao, G.; Liu, N.; Pimprale, S.; Fox, L.; Patten, C.J.; Crespi, C.L.; Miwa, G.; Gan, L.-S. Comparison of Species Differences of P-Glycoproteins in Beagle Dog, Rhesus Monkey, and Human Using ATPase Activity Assays. Mol. Pharm. 2005, 3, 78–86. [Google Scholar] [CrossRef]
- O'BRown, N.M.; Megason, S.G.; Gu, C. Suppression of transcytosis regulates zebrafish blood-brain barrier function. eLife 2019, 8. [Google Scholar] [CrossRef]
- Saleem, S.; Kannan, R.R. Zebrafish: A Promising Real-Time Model System for Nanotechnology-Mediated Neurospecific Drug Delivery. Nanoscale Res. Lett. 2021, 16, 1–14. [Google Scholar] [CrossRef]
- Xie, J.; Farage, E.; Sugimoto, M.; Anand-Apte, B. A novel transgenic zebrafish model for blood-brain and blood-retinal barrier development. BMC Dev. Biol. 2010, 10, 76–76. [Google Scholar] [CrossRef]
- Villanueva-Flores, F.; Castro-Lugo, A.; Ramírez, O.T.; A Palomares, L. Understanding cellular interactions with nanomaterials: towards a rational design of medical nanodevices. Nanotechnology 2020, 31, 132002. [Google Scholar] [CrossRef] [PubMed]
- Betzer, O.; Shilo, M.; Opochinsky, R.; Barnoy, E.; Motiei, M.; Okun, E.; Yadid, G.; Popovtzer, R. The effect of nanoparticle size on the ability to cross the blood–brain barrier: an in vivo study. Nanomedicine (Lond) 2017, 12, 1533–1546. [Google Scholar] [CrossRef]
- Liu, D.; Lin, B.; Shao, W.; Zhu, Z.; Ji, T.; Yang, C. In Vitro and in Vivo Studies on the Transport of PEGylated Silica Nanoparticles across the Blood–Brain Barrier. ACS Appl. Mater. Interfaces 2014, 6, 2131–2136. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-A.; Wu, C.-H.; Wu, S.-H.; Huang, C.-Y.; Mou, C.-Y.; Wei, K.-C.; Yen, Y.; Chien, I.-T.; Runa, S.; Chen, Y.-P.; et al. Receptor Ligand-Free Mesoporous Silica Nanoparticles: A Streamlined Strategy for Targeted Drug Delivery across the Blood–Brain Barrier. ACS Nano 2024, 18, 12716–12736. [Google Scholar] [CrossRef]
- Etame, A.B.; Smith, C.A.; Chan, W.C.; Rutka, J.T. Design and potential application of PEGylated gold nanoparticles with size-dependent permeation through brain microvasculature. Nanomedicine: Nanotechnology, Biol. Med. 2011, 7, 992–1000. [Google Scholar] [CrossRef]
- Sonavane, G.; Tomoda, K.; Makino, K. Biodistribution of colloidal gold nanoparticles after intravenous administration: Effect of particle size. Colloids Surfaces B: Biointerfaces 2008, 66, 274–280. [Google Scholar] [CrossRef]
- Longmire, M.; Choyke, P.L.; Kobayashi, H. Clearance Properties of Nano-Sized Particles and Molecules as Imaging Agents: Considerations and Caveats. Nanomedicine 2008, 3, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.; Luo, Z.; Wu, Y.-L.; Huo, S.; Liang, X.-J. Gold-based nanomaterials for the treatment of brain cancer. Cancer Biol. Med. 2021, 18, 372–387. [Google Scholar] [CrossRef]
- Nowak, M.; Brown, T.D.; Graham, A.; Helgeson, M.E.; Mitragotri, S. Size, shape, and flexibility influence nanoparticle transport across brain endothelium under flow. Bioeng. Transl. Med. 2019, 5, e10153. [Google Scholar] [CrossRef]
- Galstyan, A.; Markman, J.L.; Shatalova, E.S.; Chiechi, A.; Korman, A.J.; Patil, R.; Klymyshyn, D.; Tourtellotte, W.G.; Israel, L.L.; Braubach, O.; et al. Blood–brain barrier permeable nano immunoconjugates induce local immune responses for glioma therapy. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Sykes, E.A.; Chen, J.; Zheng, G.; Chan, W.C. Investigating the Impact of Nanoparticle Size on Active and Passive Tumor Targeting Efficiency. ACS Nano 2014, 8, 5696–5706. [Google Scholar] [CrossRef]
- Lockman, P.R.; Koziara, J.M.; Mumper, R.J.; Allen, D.D. Nanoparticle Surface Charges Alter Blood–Brain Barrier Integrity and Permeability. J. Drug Target. 2004, 12, 635–641. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, J.; Li, G.; Yin, Z.; Fu, B.M. Transcellular Model for Neutral and Charged Nanoparticles Across an In Vitro Blood–Brain Barrier. Cardiovasc. Eng. Technol. 2020, 11, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Asimakidou E, Tan JKS, Zeng J, Lo CH. Blood-Brain Barrier-Targeting Nanoparticles: Biomaterial Properties and Biomedical Applications in Translational Neuroscience. Pharmaceuticals (Basel). 2024;17(5):612.
- Kim, B.; Han, G.; Toley, B.J.; Kim, C.-K.; Rotello, V.M.; Forbes, N.S. Tuning payload delivery in tumour cylindroids using gold nanoparticles. Nat. Nanotechnol. 2010, 5, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.-G.; Wei, W.; Lv, P.-P.; Yue, H.; Wang, L.-Y.; Su, Z.-G.; Ma, G.-H. Surface Charge Affects Cellular Uptake and Intracellular Trafficking of Chitosan-Based Nanoparticles. Biomacromolecules 2011, 12, 2440–2446. [Google Scholar] [CrossRef]
- S. Hersh D, S. S. Hersh D, S. Wadajkar A, B. Roberts N, G. Perez J, P. Connolly N, Frenkel V, et al. Evolving Drug Delivery Strategies to Overcome the Blood Brain Barrier. CPD. 2016 Mar 4;22(9):1177–93.
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2020, 20, 101–124. [Google Scholar] [CrossRef]
- Kolhar, P.; Anselmo, A.C.; Gupta, V.; Pant, K.; Prabhakarpandian, B.; Ruoslahti, E.; Mitragotri, S. Using shape effects to target antibody-coated nanoparticles to lung and brain endothelium. Proc. Natl. Acad. Sci. USA 2013, 110, 10753–10758. [Google Scholar] [CrossRef]
- Peralta-Cuevas, E.; Garcia-Atutxa, I.; Huerta-Saquero, A.; Villanueva-Flores, F. The Role of Plant Virus-like Particles in Advanced Drug Delivery and Vaccine Development: Structural Attributes and Application Potential. Viruses 2025, 17, 148. [Google Scholar] [CrossRef]
- Sierri, G.; Saenz-De-Santa-Maria, I.; Renda, A.; Koch, M.; Sommi, P.; Anselmi-Tamburini, U.; Mauri, M.; D'ALoia, A.; Ceriani, M.; Salerno, D.; et al. Nanoparticle shape is the game-changer for blood–brain barrier crossing and delivery through tunneling nanotubes among glioblastoma cells. Nanoscale 2024, 17, 992–1006. [Google Scholar] [CrossRef]
- Fu, L.; Shi, B.; Wen, S.; Morsch, M.; Wang, G.; Zhou, Z.; Mi, C.; Sadraeian, M.; Lin, G.; Lu, Y.; et al. Aspect Ratio of PEGylated Upconversion Nanocrystals Affects the Cellular Uptake In Vitro and In Vivo. Acta Biomater. 2022, 147, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Zhang, M.; Kumar, S.; Vogus, D.R.; Menegatti, S.; Helgeson, M.E.; Mitragotri, S. Elasticity of Nanoparticles Influences Their Blood Circulation, Phagocytosis, Endocytosis, and Targeting. ACS Nano 2015, 9, 3169–3177. [Google Scholar] [CrossRef]
- Key, J.; Palange, A.L.; Gentile, F.; Aryal, S.; Stigliano, C.; Di Mascolo, D.; De Rosa, E.; Cho, M.; Lee, Y.; Singh, J.; et al. Soft Discoidal Polymeric Nanoconstructs Resist Macrophage Uptake and Enhance Vascular Targeting in Tumors. ACS Nano 2015, 9, 11628–11641. [Google Scholar] [CrossRef]
- Merkel, T.J.; Jones, S.W.; Herlihy, K.P.; Kersey, F.R.; Shields, A.R.; Napier, M.; Luft, J.C.; Wu, H.; Zamboni, W.C.; Wang, A.Z.; et al. Using mechanobiological mimicry of red blood cells to extend circulation times of hydrogel microparticles. Proc. Natl. Acad. Sci. 2011, 108, 586–591. [Google Scholar] [CrossRef]
- Wu JR, Hernandez Y, Miyasaki KF, Kwon EJ. Engineered nanomaterials that exploit blood-brain barrier dysfunction for delivery to the brain. Advanced Drug Delivery Reviews. 2023 Jun;197:114820.
- Cai, X.; Drummond, C.J.; Zhai, J.; Tran, N. Lipid Nanoparticles: Versatile Drug Delivery Vehicles for Traversing the Blood Brain Barrier to Treat Brain Cancer. Adv. Funct. Mater. 2024, 34. [Google Scholar] [CrossRef]
- Johnsen, K.B.; Bak, M.; Kempen, P.J.; Melander, F.; Burkhart, A.; Thomsen, M.S.; Nielsen, M.S.; Moos, T.; Andresen, T.L. Antibody affinity and valency impact brain uptake of transferrin receptor-targeted gold nanoparticles. Theranostics 2018, 8, 3416–3436. [Google Scholar] [CrossRef] [PubMed]
- Rizzuto, M.A.; Magro, R.D.; Barbieri, L.; Pandolfi, L.; Sguazzini-Viscontini, A.; Truffi, M.; Salvioni, L.; Corsi, F.; Colombo, M.; Re, F.; et al. H-Ferritin nanoparticle-mediated delivery of antibodies across a BBB in vitro model for treatment of brain malignancies. Biomater. Sci. 2021, 9, 2032–2042. [Google Scholar] [CrossRef]
- Rodrigues, B.d.S.; Arora, S.; Kanekiyo, T.; Singh, J. Efficient neuronal targeting and transfection using RVG and transferrin-conjugated liposomes. Brain Res. 2020, 1734, 146738. [Google Scholar] [CrossRef]
- Wehn, A.C.; Krestel, E.; Harapan, B.N.; Klymchenko, A.; Plesnila, N.; Khalin, I. To see or not to see: In vivo nanocarrier detection methods in the brain and their challenges. J. Control. Release 2024, 371, 216–236. [Google Scholar] [CrossRef]
- Ye, Z.; Gastfriend, B.D.; Umlauf, B.J.; Lynn, D.M.; Shusta, E.V. Antibody-Targeted Liposomes for Enhanced Targeting of the Blood-Brain Barrier. Pharm. Res. 2022, 39, 1523–1534. [Google Scholar] [CrossRef]
- Orii, K.O.; Grubb, J.H.; Vogler, C.; Levy, B.; Tan, Y.; Markova, K.; Davidson, B.L.; Mao, Q.; Orii, T.; Kondo, N.; et al. Defining the Pathway for Tat-mediated Delivery of β-Glucuronidase in Cultured Cells and MPS VII Mice. Mol. Ther. 2005, 12, 345–352. [Google Scholar] [CrossRef]
- Pardridge, W.M. Brain Delivery of Nanomedicines: Trojan Horse Liposomes for Plasmid DNA Gene Therapy of the Brain. Front. Med Technol. 2020, 2, 602236. [Google Scholar] [CrossRef]
- Sarko D, Beijer B, Garcia Boy R, Nothelfer EM, Leotta K, Eisenhut M, et al. The Pharmacokinetics of Cell-Penetrating Peptides. Mol Pharmaceutics. 2010 Dec 6;7(6):2224–31.
- Madani, F.; Morovvati, H.; Webster, T.J.; Asaadi, S.N.; Rezayat, S.M.; Hadjighassem, M.; Khosravani, M.; Adabi, M. Combination chemotherapy via poloxamer 188 surface-modified PLGA nanoparticles that traverse the blood–brain–barrier in a glioblastoma model. Sci. Rep. 2024, 14, 1–22. [Google Scholar] [CrossRef]
- Li, J.; Sabliov, C. PLA/PLGA nanoparticles for delivery of drugs across the blood-brain barrier. 2016. [Google Scholar] [CrossRef]
- Liaw, K.; Zhang, F.; Mangraviti, A.; Kannan, S.; Tyler, B.; Kannan, R.M. Dendrimer size effects on the selective brain tumor targeting in orthotopic tumor models upon systemic administration. Bioeng. Transl. Med. 2020, 5, e10160. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, X.; Zhu, Y.; Qian, Y.; Feng, Y.; Li, H.; Hu, G. Preparation and brain targeting effects study of recombinant human ferritin nanoparticles. Biochem. Biophys. Res. Commun. 2024, 712-713, 149939. [Google Scholar] [CrossRef]
- Simon-Santamaria, J.; Rinaldo, C.H.; Kardas, P.; Li, R.; Malovic, I.; Elvevold, K.; McCourt, P.; Smedsrød, B.; Hirsch, H.H.; Sørensen, K.K.; et al. Efficient Uptake of Blood-Borne BK and JC Polyomavirus-Like Particles in Endothelial Cells of Liver Sinusoids and Renal Vasa Recta. PLOS ONE 2014, 9, e111762–e111762. [Google Scholar] [CrossRef]
- Ye, D.; Zimmermann, T.; Demina, V.; Sotnikov, S.; Ried, C.L.; Rahn, H.; Stapf, M.; Untucht, C.; Rohe, M.; Terstappen, G.C.; et al. Trafficking of JC virus-like particles across the blood–brain barrier. Nanoscale Adv. 2021, 3, 2488–2500. [Google Scholar] [CrossRef]
- Li, X.; Vemireddy, V.; Cai, Q.; Xiong, H.; Kang, P.; Li, X.; Giannotta, M.; Hayenga, H.N.; Pan, E.; Sirsi, S.R.; et al. Reversibly Modulating the Blood–Brain Barrier by Laser Stimulation of Molecular-Targeted Nanoparticles. Nano Lett. 2021, 21, 9805–9815. [Google Scholar] [CrossRef]
- Arguello, A.; Mahon, C.S.; Calvert, M.E.; Chan, D.; Dugas, J.C.; Pizzo, M.E.; Thomsen, E.R.; Chau, R.; Damo, L.A.; Duque, J.; et al. Molecular architecture determines brain delivery of a transferrin receptor–targeted lysosomal enzyme. J. Exp. Med. 2022, 219. [Google Scholar] [CrossRef]
- Wu D, Chen Q, Chen X, Han F, Chen Z, Wang Y. The blood–brain barrier: Structure, regulation and drug delivery. Sig Transduct Target Ther. 2023;8(1):217.
- Chu, C.; Zhang, Y.; Boado, R.J.; Pardridge, W.M. Decline in Exogenous Gene Expression in Primate Brain Following Intravenous Administration Is Due to Plasmid Degradation. Pharm. Res. 2006, 23, 1586–1590. [Google Scholar] [CrossRef]
- Ulbrich, K.; Knobloch, T.; Kreuter, J. Targeting the insulin receptor: nanoparticles for drug delivery across the blood–brain barrier (BBB). J. Drug Target. 2010, 19, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Li Y, Zheng X, Gong M, Zhang J. Delivery of a peptide-drug conjugate targeting the blood brain barrier improved the efficacy of paclitaxel against glioma. Oncotarget. 2016 Nov 29;7(48):79401–7.
- Zhang Z, Li J, Wang Y, Tang C, Zhou Y, Li J, et al. Angiopep-2 conjugated biomimetic nano-delivery system loaded with resveratrol for the treatment of methamphetamine addiction. International Journal of Pharmaceutics. 2024 Sep;663:124552.
- di Polidoro, A.C.; Cafarchio, A.; Vecchione, D.; Donato, P.; De Nola, F.; Torino, E. Revealing Angiopep-2/LRP1 Molecular Interaction for Optimal Delivery to Glioblastoma (GBM). Molecules 2022, 27, 6696. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.P.; Prajapati, N.; DeCoster, M.A.; Poh, S.; Murray, T.A. Efficient LRP1-Mediated Uptake and Low Cytotoxicity of Peptide L57 In Vitro Shows Its Promise as CNS Drug Delivery Vector. J. Pharm. Sci. 2021, 110, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Leite, D.M.; Scarpa, E.; Nyberg, S.; Fullstone, G.; Forth, J.; Matias, D.; Apriceno, A.; Poma, A.; Duro-Castano, A.; et al. On the shuttling across the blood-brain barrier via tubule formation: Mechanism and cargo avidity bias. Sci. Adv. 2020, 6, eabc4397. [Google Scholar] [CrossRef]
- Pereira, P.; Barreira, M.; Cruz, C.; Tomás, J.; Luís, Â.; Pedro, A.Q.; Queiroz, J.A.; Sousa, F. Brain-Targeted Delivery of Pre-miR-29b Using Lactoferrin-Stearic Acid-Modified-Chitosan/Polyethyleneimine Polyplexes. Pharmaceuticals 2020, 13, 314. [Google Scholar] [CrossRef]
- Youssef JR, Boraie NA, Ismail FA, Bakr BA, Allam EA, El-Moslemany RM. Brain targeted lactoferrin coated lipid nanocapsules for the combined effects of apocynin and lavender essential oil in PTZ induced seizures. Drug Deliv and Transl Res. 2025 Feb;15(2):534–55.
- Sharma, S.; Dang, S. Nanocarrier-Based Drug Delivery to Brain: Interventions of Surface Modification. Curr. Neuropharmacol. 2023, 21, 517–535. [Google Scholar] [CrossRef]
- McCord, E.; Pawar, S.; Koneru, T.; Tatiparti, K.; Sau, S.; Iyer, A.K. Folate Receptors’ Expression in Gliomas May Possess Potential Nanoparticle-Based Drug Delivery Opportunities. ACS Omega 2021, 6, 4111–4118. [Google Scholar] [CrossRef]
- Morshedi, B.; Esfandyari-Manesh, M.; Atyabi, F.; Ghahremani, M.H.; Dinarvand, R. Local delivery of ibrutinib by folate receptor-mediated targeting PLGA–PEG nanoparticles to glioblastoma multiform: in vitro and in vivo studies. J. Drug Target. 2025, 1–16. [Google Scholar] [CrossRef]
- Yücel, O.; Aksüt, Y.; Şengelen, A.; Yıldırım, E.; Emik, S.; Arda, N.; Gürdağ, G. Folate receptor-targeted indomethacin-loaded gold nanoparticles enhance drug chemotherapeutic efficacy in glioblastoma cells and spheroids. J. Drug Deliv. Sci. Technol. 2024, 100. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Johnsen, K.B.; Kucharz, K.; Lauritzen, M.; Moos, T. Blood–Brain Barrier Transport of Transferrin Receptor-Targeted Nanoparticles. Pharmaceutics 2022, 14, 2237. [Google Scholar] [CrossRef]
- Neves, V.; Aires-Da-Silva, F.; Morais, M.; Gano, L.; Ribeiro, E.; Pinto, A.; Aguiar, S.; Gaspar, D.; Fernandes, C.; Correia, J.D.G.; et al. Novel Peptides Derived from Dengue Virus Capsid Protein Translocate Reversibly the Blood–Brain Barrier through a Receptor-Free Mechanism. ACS Chem. Biol. 2017, 12, 1257–1268. [Google Scholar] [CrossRef] [PubMed]
- Szecskó, A.; Mészáros, M.; Simões, B.; Cavaco, M.; Chaparro, C.; Porkoláb, G.; Castanho, M.A.; Deli, M.A.; Neves, V.; Veszelka, S. PepH3-modified nanocarriers for delivery of therapeutics across the blood-brain barrier. Fluids Barriers CNS 2025, 22, 1–18. [Google Scholar] [CrossRef]
- Cavaco, M.; Fraga, P.; Valle, J.; Silva, R.D.M.; Gano, L.; Correia, J.D.G.; Andreu, D.; Castanho, M.A.R.B.; Neves, V. Molecular determinants for brain targeting by peptides: a meta-analysis approach with experimental validation. Fluids Barriers CNS 2024, 21, 1–13. [Google Scholar] [CrossRef]
- Ibrahim, R.M.; Teaima, M.; El-Nabarawi, M.; Badawi, N.M. Intranasal delivery of chitosan-based nanoparticles as an innovative way for management of neurodegenerative disorders: A comprehensive review of advanced strategies for CNS targeting. J. Drug Deliv. Sci. Technol. 2024, 99. [Google Scholar] [CrossRef]
- Khan, I.N.; Navaid, S.; Waqar, W.; Hussein, D.; Ullah, N.; Khan, M.U.A.; Hussain, Z.; Javed, A. Chitosan-Based Polymeric Nanoparticles as an Efficient Gene Delivery System to Cross Blood Brain Barrier: In Vitro and In Vivo Evaluations. Pharmaceuticals 2024, 17, 169. [Google Scholar] [CrossRef] [PubMed]
- Sepand, M.R.; Ghavami, M.; Zanganeh, S.; Stacks, S.; Ghasemi, F.; Montazeri, H.; Corbo, C.; Derakhshankhah, H.; Ostad, S.N.; Ghahremani, M.H.; et al. Impact of plasma concentration of transferrin on targeting capacity of nanoparticles. Nanoscale 2020, 12, 4935–4944. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, B.; Xie, S.; Yang, B.; Xu, Q.; Tan, J. Superparamagnetic Iron Oxide Nanoparticles Modified with Tween 80 Pass through the Intact Blood–Brain Barrier in Rats under Magnetic Field. ACS Appl. Mater. Interfaces 2016, 8, 11336–11341. [Google Scholar] [CrossRef]
- Thomas, R.G.; Kim, S.; Tran, T.-A.; Kim, Y.H.; Nagareddy, R.; Jung, T.-Y.; Kim, S.K.; Jeong, Y.Y. Magnet-Guided Temozolomide and Ferucarbotran Loaded Nanoparticles to Enhance Therapeutic Efficacy in Glioma Model. Nanomaterials 2024, 14, 939. [Google Scholar] [CrossRef]
- Thomsen, L.B.; Thomsen, M.S.; Moos, T. Targeted Drug Delivery to the Brain Using Magnetic Nanoparticles. Ther. Deliv. 2015, 6, 1145–1155. [Google Scholar] [CrossRef]
- Savari, M.-N. Fe3O4@Chitosan@ZIF-8@RVG29, an anti-glioma nanoplatform guided by fixed and activated by alternating magnetic field. Sci. Rep. 2024, 14, 1–11. [Google Scholar] [CrossRef]
- Chuapoco, M.R.; Flytzanis, N.C.; Goeden, N.; Octeau, J.C.; Roxas, K.M.; Chan, K.Y.; Scherrer, J.; Winchester, J.; Blackburn, R.J.; Campos, L.J.; et al. Adeno-associated viral vectors for functional intravenous gene transfer throughout the non-human primate brain. Nat. Nanotechnol. 2023, 18, 1241–1251. [Google Scholar] [CrossRef]
- Huang, Q.; Chan, K.Y.; Wu, J.; Botticello-Romero, N.R.; Zheng, Q.; Lou, S.; Keyes, C.; Svanbergsson, A.; Johnston, J.; Mills, A.; et al. An AAV capsid reprogrammed to bind human transferrin receptor mediates brain-wide gene delivery. Science 2024, 384, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.R.; Lee, A.S.; Kim, S.M.; Heo, H.R.; Kim, C.S. Virus-like nanoparticles as a theranostic platform for cancer. Front. Bioeng. Biotechnol. 2023, 10, 1106767. [Google Scholar] [CrossRef]
- Kumar, M.A.; Baba, S.K.; Sadida, H.Q.; Marzooqi, S.A.; Jerobin, J.; Altemani, F.H.; Algehainy, N.; Alanazi, M.A.; Abou-Samra, A.-B.; Kumar, R.; et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct. Target. Ther. 2024, 9, 1–41. [Google Scholar] [CrossRef]
- Khatami, S.H.; Karami, N.; Taheri-Anganeh, M.; Taghvimi, S.; Tondro, G.; Khorsand, M.; Fard, E.S.; Sedighimehr, N.; Kazemi, M.; Jaberi, K.R.; et al. Exosomes: Promising Delivery Tools for Overcoming Blood-Brain Barrier and Glioblastoma Therapy. Mol. Neurobiol. 2023, 60, 4659–4678. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Iqbal, Z.; Lu, J.; Wang, J.; Zhang, H.; Chen, X.; Duan, L.; Xia, J. Cell-derived nanovesicle-mediated drug delivery to the brain: Principles and strategies for vesicle engineering. Mol. Ther. 2022, 31, 1207–1224. [Google Scholar] [CrossRef]
- Cui G hong, Guo H dong, Li H, Zhai Y, Gong Z bin, Wu J, et al. RVG-modified exosomes derived from mesenchymal stem cells rescue memory deficits by regulating inflammatory responses in a mouse model of Alzheimer’s disease. Immun Ageing. 2019 Dec;16(1):10.
- Chu, L.; Sun, Y.; Zhao, Y.; Wang, A.; Sun, Y.; Duan, X.; Li, N.; Xia, H.; Liu, W.; Sun, K. Exosome-mediated delivery platform of biomacromolecules into the brain: Cetuximab in combination with doxorubicin for glioblastoma therapy. Int. J. Pharm. 2024, 660, 124262. [Google Scholar] [CrossRef]
- Zhao, M.; Li, Q.; Chai, Y.; Rong, R.; He, L.; Zhang, Y.; Cui, H.; Xu, H.; Wang, Z.; Yuan, S.; et al. An anti-CD19-exosome delivery system navigates the blood–brain barrier for targeting of central nervous system lymphoma. J. Nanobiotechnology 2025, 23, 1–21. [Google Scholar] [CrossRef]
- Moos, T.; Morgan, E.H. Restricted transport of anti-transferrin receptor antibody (OX26) through the blood–brain barrier in the rat. J. Neurochem. 2001, 79, 119–129. [Google Scholar] [CrossRef]
- Dixit, S.; Novak, T.; Miller, K.; Zhu, Y.; Kenney, M.E.; Broome, A.-M. Transferrin receptor-targeted theranostic gold nanoparticles for photosensitizer delivery in brain tumors. Nanoscale 2014, 7, 1782–1790. [Google Scholar] [CrossRef]
- Dixit, S.; Novak, T.; Miller, K.; Zhu, Y.; Kenney, M.E.; Broome, A.-M. Transferrin receptor-targeted theranostic gold nanoparticles for photosensitizer delivery in brain tumors. Nanoscale 2014, 7, 1782–1790. [Google Scholar] [CrossRef]
- Kuang, Y.; An, S.; Guo, Y.; Huang, S.; Shao, K.; Liu, Y.; Li, J.; Ma, H.; Jiang, C. T7 peptide-functionalized nanoparticles utilizing RNA interference for glioma dual targeting. Int. J. Pharm. 2013, 454, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, Y.; Jiang, Y.; Lv, W.; Wu, L.; Wang, B.; Lv, L.; Xu, Q.; Xin, H. Enhanced anti-ischemic stroke of ZL006 by T7-conjugated PEGylated liposomes drug delivery system. Sci. Rep. 2015, 5, 12651. [Google Scholar] [CrossRef]
- Giugliani, R.; Giugliani, L.; de Oliveira Poswar, F.; Donis, K.C.; Corte, A.D.; Schmidt, M.; Boado, R.J.; Nestrasil, I.; Nguyen, C.; Chen, S.; et al. Neurocognitive and somatic stabilization in pediatric patients with severe Mucopolysaccharidosis Type I after 52 weeks of intravenous brain-penetrating insulin receptor antibody-iduronidase fusion protein (valanafusp alpha): an open label phase 1-2 trial. Orphanet J. Rare Dis. 2018, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Nyberg, S.; Poma, A.; Cecchin, D.; Jain, S.A.; Kim, K.-A.; Shin, Y.-J.; Kim, E.-H.; Kim, M.; Baek, S.-H.; et al. LRP-1 functionalized polymersomes enhance the efficacy of carnosine in experimental stroke. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Zhu J, Zhang Y, Chen X, Zhang Y, Zhang K, Zheng H, et al. Angiopep-2 modified lipid-coated mesoporous silica nanoparticles for glioma targeting therapy overcoming BBB. Biochemical and Biophysical Research Communications. 2021 Jan;534:902–7.
- Chen, H.; Qin, Y.; Zhang, Q.; Jiang, W.; Tang, L.; Liu, J.; He, Q. Lactoferrin modified doxorubicin-loaded procationic liposomes for the treatment of gliomas. Eur. J. Pharm. Sci. 2011, 44, 164–173. [Google Scholar] [CrossRef]
- Wu, Y.; Qian, Y.; Peng, W.; Qi, X. Functionalized nanoparticles crossing the brain–blood barrier to target glioma cells. PeerJ 2023, 11, e15571. [Google Scholar] [CrossRef]
- Kong, S.D.; Zhang, W.; Lee, J.H.; Brammer, K.; Lal, R.; Karin, M.; Jin, S. Magnetically Vectored Nanocapsules for Tumor Penetration and Remotely Switchable On-Demand Drug Release. Nano Lett. 2010, 10, 5088–5092. [Google Scholar] [CrossRef]
- Kong, S.D.; Lee, J.; Ramachandran, S.; Eliceiri, B.P.; Shubayev, V.I.; Lal, R.; Jin, S. Magnetic targeting of nanoparticles across the intact blood–brain barrier. J. Control. Release 2012, 164, 49–57. [Google Scholar] [CrossRef]
- Pitek AS, Wen AM, Shukla S, Steinmetz NF. The Protein Corona of Plant Virus Nanoparticles Influences their Dispersion Properties, Cellular Interactions, and In Vivo Fates. Small. 2016 Apr;12(13):1758–69.
- Kaiser, C.R.; Flenniken, M.L.; Gillitzer, E.; Harmsen, A.L.; Harmsen, A.G.; A Jutila, M.; Douglas, T.; Young, M.J. Biodistribution studies of protein cage nanoparticles demonstrate broad tissue distribution and rapid clearance in vivo. 2007, 2, 715–733.
- Bai, L.; Yu, L.; Ran, M.; Zhong, X.; Sun, M.; Xu, M.; Wang, Y.; Yan, X.; Lee, R.J.; Tang, Y.; et al. Harnessing the Potential of Exosomes in Therapeutic Interventions for Brain Disorders. Int. J. Mol. Sci. 2025, 26, 2491. [Google Scholar] [CrossRef]
- Meyer, C.; Losacco, J.; Stickney, Z.; Li, L.; Marriott, G.; Lu, B. Pseudotyping exosomes for enhanced protein delivery in mammalian cells. Int. J. Nanomed. 2017, ume 12, 3153–3170. [Google Scholar] [CrossRef]
- Chen, J.; Chopp, M. Exosome Therapy for Stroke. Stroke 2018, 49, 1083–1090. [Google Scholar] [CrossRef]
- Drommelschmidt, K.; Serdar, M.; Bendix, I.; Herz, J.; Bertling, F.; Prager, S.; Keller, M.; Ludwig, A.-K.; Duhan, V.; Radtke, S.; et al. Mesenchymal stem cell-derived extracellular vesicles ameliorate inflammation-induced preterm brain injury. Brain, Behav. Immun. 2017, 60, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Nishida, H.; An, S.Y.; Shetty, A.K.; Bartosh, T.J.; Prockop, D.J. Chromatographically isolated CD63 + CD81 + extracellular vesicles from mesenchymal stromal cells rescue cognitive impairments after TBI. Proc. Natl. Acad. Sci. 2015, 113, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Komatsu, H.; Poku, E.K.; Olafsen, T.; Huang, K.X.; Huang, L.A.; Chea, J.; Bowles, N.; Chang, B.; Rawson, J.; et al. Biodistribution of Intra-Arterial and Intravenous Delivery of Human Umbilical Cord Mesenchymal Stem Cell-Derived Extracellular Vesicles in a Rat Model to Guide Delivery Strategies for Diabetes Therapies. Pharmaceuticals 2022, 15, 595. [Google Scholar] [CrossRef]
- Ophelders, D.R.; Wolfs, T.G.; Jellema, R.K.; Zwanenburg, A.; Andriessen, P.; Delhaas, T.; Ludwig, A.-K.; Radtke, S.; Peters, V.; Janssen, L.; et al. Mesenchymal Stromal Cell-Derived Extracellular Vesicles Protect the Fetal Brain After Hypoxia-Ischemia. STEM CELLS Transl. Med. 2016, 5, 754–763. [Google Scholar] [CrossRef]
- Waseem A, Saudamini, Haque R, Janowski M, Raza SS. Mesenchymal stem cell-derived exosomes: Shaping the next era of stroke treatment. Neuroprotection. 2023 Dec;1(2):99–116.
- Zhang, Y.; Chopp, M.; Meng, Y.; Katakowski, M.; Xin, H.; Mahmood, A.; Xiong, Y. Effect of exosomes derived from multipluripotent mesenchymal stromal cells on functional recovery and neurovascular plasticity in rats after traumatic brain injury. J. Neurosurg. 2015, 122, 856–867. [Google Scholar] [CrossRef]
- Liu, W.; Wang, W.; Dong, X.; Sun, Y. Near-Infrared Light-Powered Janus Nanomotor Significantly Facilitates Inhibition of Amyloid-β Fibrillogenesis. ACS Appl. Mater. Interfaces 2020, 12, 12618–12628. [Google Scholar] [CrossRef]
- Pang, L.; Qin, J.; Han, L.; Zhao, W.; Liang, J.; Xie, Z.; Yang, P.; Wang, J. Exploiting macrophages as targeted carrier to guide nanoparticles into glioma. Oncotarget 2016, 7, 37081–37091. [Google Scholar] [CrossRef]
- Li, J.; Angsantikul, P.; Liu, W.; de Ávila, B.E.; Chang, X.; Sandraz, E.; Liang, Y.; Zhu, S.; Zhang, Y.; Chen, C.; et al. Biomimetic Platelet-Camouflaged Nanorobots for Binding and Isolation of Biological Threats. Adv. Mater. 2017, 30. [Google Scholar] [CrossRef]
- ASTM E2524-08. Standard Test Method for Analysis of Hemolytic Properties of Nanoparticles (ASTM E2524-08). [Internet]. ASTM International; 2008. Available from: https://www.astm.org/e2524-08r13.
- FDA S7A. S7A Safety Pharmacology Studies for Human Pharmaceuticals. U.S. Department of Health and Human Services [Internet]. U.S. Food and Drug Administration.; 2001. Available from: https://www.fda.gov/media/72033/download.
- ICH. ICH Harmonised Tripartite Guideline: Safety Pharmacology Studies for Human Pharmaceuticals S7A. [Internet]. International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use (ICH).; 2000. Available from: https://database.ich.org/sites/default/files/S7A_Guideline.pdf European Medicines Agency (EMA)+5.
- Gao, J.; Song, Q.; Gu, X.; Jiang, G.; Huang, J.; Tang, Y.; Yu, R.; Wang, A.; Huang, Y.; Zheng, G.; et al. Intracerebral fate of organic and inorganic nanoparticles is dependent on microglial extracellular vesicle function. Nat. Nanotechnol. 2023, 19, 376–386. [Google Scholar] [CrossRef] [PubMed]
- FDA. Drug Products, Including Biological Products, that Contain Nanomaterials: Guidance for Industry [Internet]. U.S. Food and Drug Administration; 2022. Available from: https://www.fda.gov/media/157812/download.
- ICHS8. ICH Harmonised Tripartite Guideline: Immunotoxicity Studies for Human Pharmaceuticals S8 [Internet]. 2005. Available from: https://www.ema.europa.eu/en/documents/scientific-guideline/ich-s-8-immunotoxicity-studies-human-pharmaceuticals-step-5_en.pdf U.S. Food and Drug Administration+.
- Liz-Marzán LM, Kagan CR, Millstone JE. Reproducibility in Nanocrystal Synthesis? Watch Out for Impurities! ACS Nano. 2020 Jun 23;14(6):6359–61.
- ASTM F756-17. ASTM F756-17: Standard Practice for Assessment of Hemolytic Properties of Materials. [Internet]. ASTM International.; 2017. Available from: https://www.astm.org/f0756-17.html.
- Braune, S.; Lendlein, A.; Jung, F. Developing standards and test protocols for testing the hemocompatibility of biomaterials. In Hemocompatibility of Biomaterials for Clinical Applications; Siedlecki, C.A., Ed.; Woodhead Publishing: Sawston, UK, 2018; pp. 51–76. [Google Scholar] [CrossRef]
- FDA. Use of International Standard ISO 10993-1, “Biological evaluation of medical devices – Part 1: Evaluation and testing within a risk management process”. [Internet]. U.S. Food and Drug Administration.; 2020. Available from: https://www.fda.gov/media/142959/download U.S. Food and Drug Administration+.
- Giannakou C, Park MVDZ, Bosselaers IEM, de Jong WH, van der Laan JW, van Loveren H, et al. Nonclinical regulatory immunotoxicity testing of nanomedicinal products: Proposed strategy and possible pitfalls. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2020 Sep;12(5):e1633.
- ISO 10993-4:2017. ISO 10993-4:2017 Biological evaluation of medical devices — Part 4: Selection of tests for interactions with blood [Internet]. International Organization for Standardization.; 2017. Available from: https://www.iso.org/standard/63448.html.
- Totea, G.; Ionita, D.; Demetrescu, I.; Mitache, M.M. In vitro hemocompatibility and corrosion behavior of new Zr-binary alloys in whole human blood. Open Chem. 2014, 12, 796–803. [Google Scholar] [CrossRef]
- Weber, M.; Steinle, H.; Golombek, S.; Hann, L.; Schlensak, C.; Wendel, H.P.; Avci-Adali, M. Blood-Contacting Biomaterials: In Vitro Evaluation of the Hemocompatibility. Front. Bioeng. Biotechnol. 2018, 6, 99. [Google Scholar] [CrossRef]
- European Commission. Commission Directive 2009/120/EC of amending Directive 2001/83/EC of the European Parliament and of the Council on the Community code relating to medicinal products for human use as regards advanced therapy medicinal products. [Internet]. 2009. Available from: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32009L0120.
- FDA S7A. Extended release oral dosage forms: Development, evaluation, and application of in vitro/in vivo correlations [Internet]. U.S. Department of Health and Human Services, Center for Drug Evaluation and Research; 1997. Available from: https://www.fda.gov/media/72033/download.
- OECD. Test No. 424: Neurotoxicity Study in Rodents. OECD Guidelines for the Testing of Chemicals, Section 4. [Internet]. Organización para la Cooperación y el Desarrollo Económicos.; 1997. Available from: https://www.oecd.org/en/publications/test-no-424-neurotoxicity-study-in-rodents_9789264071025-en.html.
- Vervaeke, P.; Borgos, S.; Sanders, N.; Combes, F. Regulatory guidelines and preclinical tools to study the biodistribution of RNA therapeutics. Adv. Drug Deliv. Rev. 2022, 184, 114236. [Google Scholar] [CrossRef]
- Gu, X.; Song, Q.; Zhang, Q.; Huang, M.; Zheng, M.; Chen, J.; Wei, D.; Wei, X.; Chen, H.; Zheng, G.; et al. Clearance of two organic nanoparticles from the brain via the paravascular pathway. J. Control. Release 2020, 322, 31–41. [Google Scholar] [CrossRef]
- Liu, R.; Jia, W.; Wang, Y.; Hu, C.; Yu, W.; Huang, Y.; Wang, L.; Gao, H. Glymphatic System and Subsidiary Pathways Drive Nanoparticles Away from the Brain. Research 2022, 2022, 9847612. [Google Scholar] [CrossRef]
- Pietroiusti, A.; Campagnolo, L.; Fadeel, B. Interactions of Engineered Nanoparticles with Organs Protected by Internal Biological Barriers. Small 2012, 9, 1557–1572. [Google Scholar] [CrossRef]
- Le Meur, M.; Pignatelli, J.; Blasi, P.; Palomo, V. Nanoparticles targeting the central circadian clock: Potential applications for neurological disorders. Adv. Drug Deliv. Rev. 2025, 220, 115561. [Google Scholar] [CrossRef]
- Dobrovolskaia, M.A. Lessons learned from immunological characterization of nanomaterials at the Nanotechnology Characterization Laboratory. Front. Immunol. 2022, 13, 984252. [Google Scholar] [CrossRef]
- Pan, J.; Wang, Y.; Chen, Y.; Zhang, C.; Deng, H.; Lu, J.; Chen, W. Emerging strategies against accelerated blood clearance phenomenon of nanocarrier drug delivery systems. J. Nanobiotechnology 2025, 23, 1–22. [Google Scholar] [CrossRef]
- Crist, R.M.; Dasa, S.S.K.; Liu, C.H.; Clogston, J.D.; Dobrovolskaia, M.A.; Stern, S.T. Challenges in the development of nanoparticle-based imaging agents: Characterization and biology. WIREs Nanomed. Nanobiotechnology 2020, 13, e1665. [Google Scholar] [CrossRef]
- FDA. Process validation: General principles and practices. [Internet]. U.S. Food and Drug Administration; 2011. Available from: www.fda.gov/media/71021/download.
- ICHQ13. ICH guideline Q13 on continuous manufacturing of drug substances and drug products. [Internet]. International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use (ICH); 2021. Available from: https://www.ema.europa.eu/en/documents/scientific-guideline/ich-guideline-q13-continuous-manufacturing-drug-substances-drug-products-step-5_en.pdf European Medicines Agency (EMA)+3.
Figure 1.
Structural composition of the BBB. The BBB is formed primarily by a monolayer of endothelial cells joined by tight junctions, which restrict paracellular transport. Pericytes and astrocytic end-feet provide structural and functional support, contributing to barrier integrity and regulation of nutrient exchange. The neurovascular unit also includes neurons and microglia, which interact closely with vascular components to maintain central nervous system (CNS) homeostasis. Various blood cells circulate in the lumen, erythrocytes, monocytes, lymphocytes, and neutrophils, but are largely excluded from the brain parenchyma under physiological conditions due to the barrier's selectivity.
Figure 1.
Structural composition of the BBB. The BBB is formed primarily by a monolayer of endothelial cells joined by tight junctions, which restrict paracellular transport. Pericytes and astrocytic end-feet provide structural and functional support, contributing to barrier integrity and regulation of nutrient exchange. The neurovascular unit also includes neurons and microglia, which interact closely with vascular components to maintain central nervous system (CNS) homeostasis. Various blood cells circulate in the lumen, erythrocytes, monocytes, lymphocytes, and neutrophils, but are largely excluded from the brain parenchyma under physiological conditions due to the barrier's selectivity.
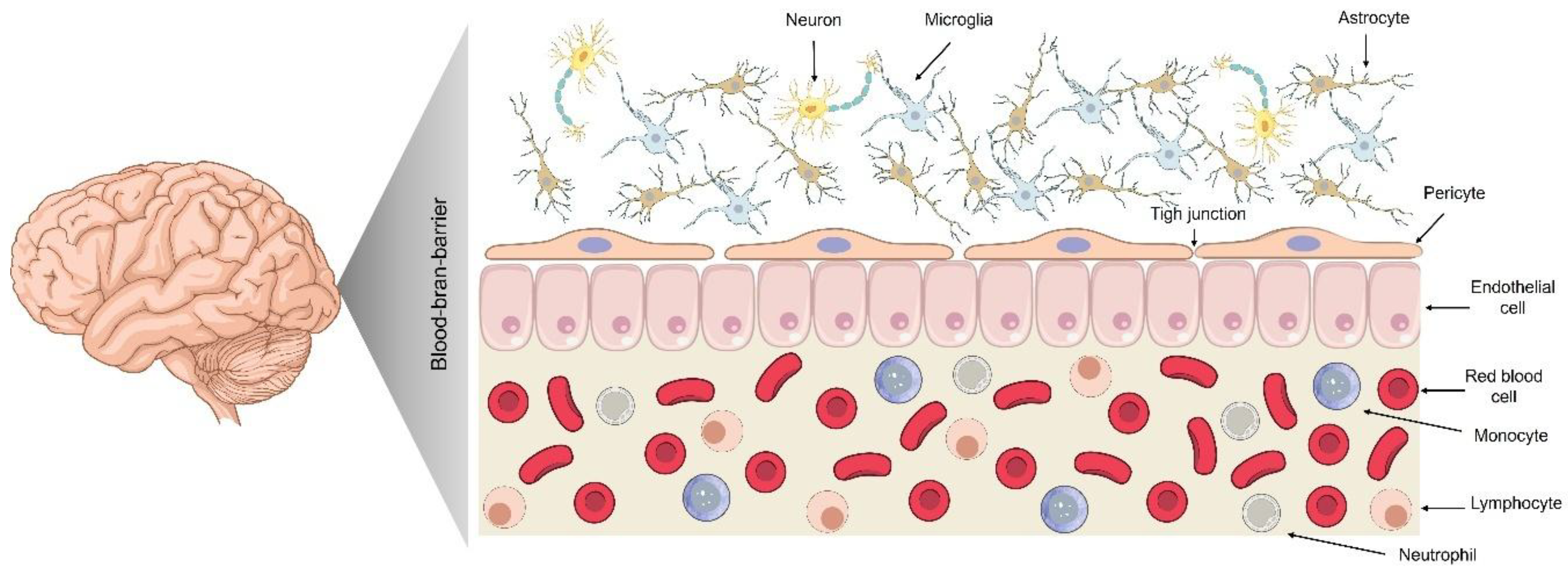
Figure 2.
Key physicochemical parameters influencing nanoparticle transport across the BBB. Effective BBB penetration by nanocarriers depends on the optimization of multiple design parameters. Material class influences biodistribution and immune interactions; VLPs, liposomes, exosomes, and polymeric or metallic nanoparticles exhibit distinct advantages. A size of 10–100 nm is typically optimal for transcytosis. Shape affects cellular uptake and biodistribution, with nanorods and nanotubes (aspect ratio ~2–5) showing enhanced BBB permeability. Surface charge modulates electrostatic interactions with the endothelium; a near-neutral zeta potential (-10 to +10 mV) minimizes opsonization and promotes translocation. Stiffness also plays a critical role; rigid particles (Young’s modulus >3000 MPa) tend to favor endothelial uptake over softer counterparts. Rational tuning of these parameters is essential for the design of CNS-targeting nanomedicines.
Figure 2.
Key physicochemical parameters influencing nanoparticle transport across the BBB. Effective BBB penetration by nanocarriers depends on the optimization of multiple design parameters. Material class influences biodistribution and immune interactions; VLPs, liposomes, exosomes, and polymeric or metallic nanoparticles exhibit distinct advantages. A size of 10–100 nm is typically optimal for transcytosis. Shape affects cellular uptake and biodistribution, with nanorods and nanotubes (aspect ratio ~2–5) showing enhanced BBB permeability. Surface charge modulates electrostatic interactions with the endothelium; a near-neutral zeta potential (-10 to +10 mV) minimizes opsonization and promotes translocation. Stiffness also plays a critical role; rigid particles (Young’s modulus >3000 MPa) tend to favor endothelial uptake over softer counterparts. Rational tuning of these parameters is essential for the design of CNS-targeting nanomedicines.
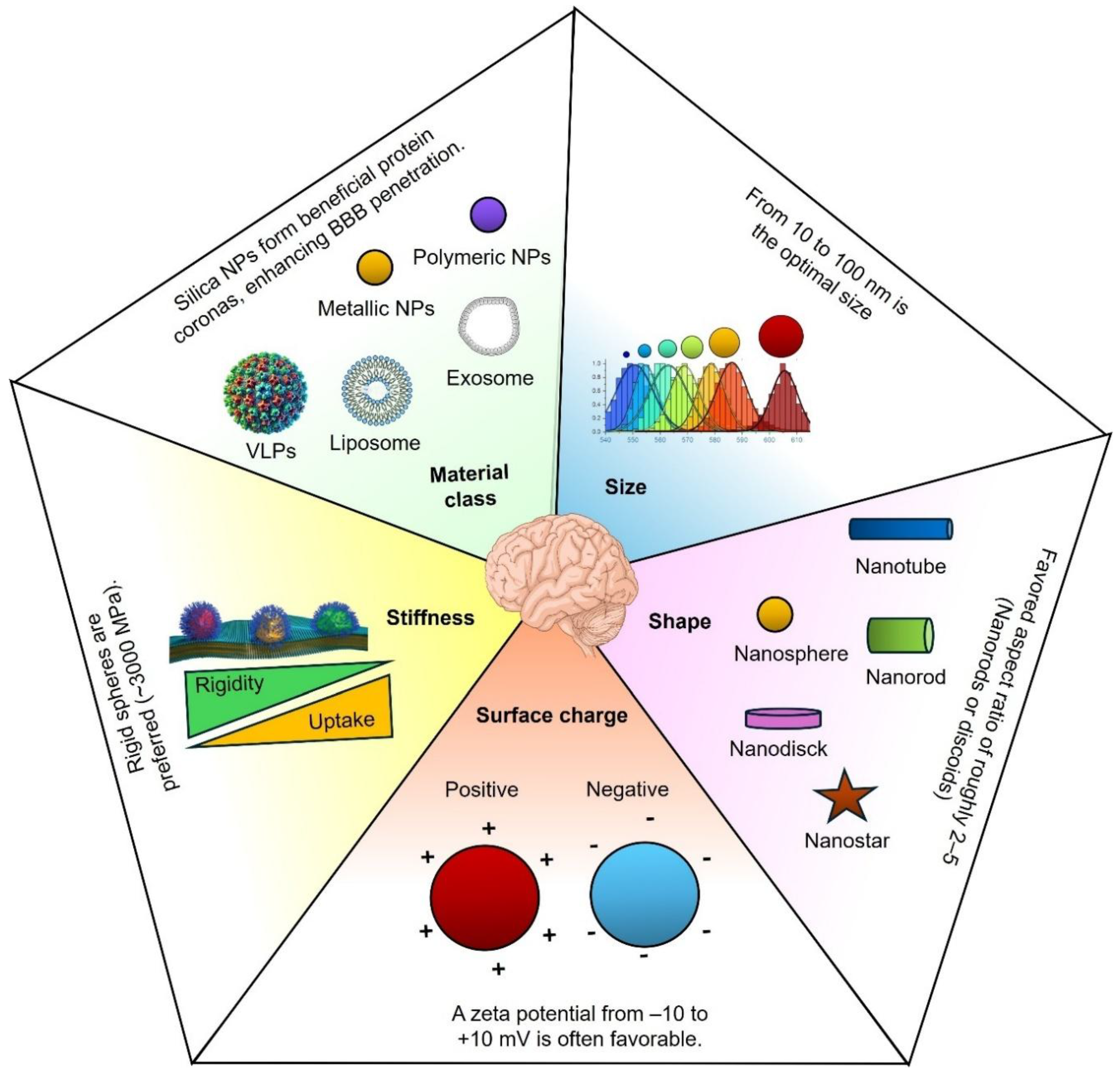
Figure 3.
Mechanisms of nanoparticle transport across the BBB. NPs can traverse the BBB via multiple pathways, each with distinct biophysical and biochemical requirements. These include: (1) Passive transmembrane diffusion, mainly for lipophilic, low-molecular-weight compounds; (2) Paracellular transport, which is highly restricted by tight junctions; (3) Receptor-mediated transcytosis, utilizing ligands that bind to endothelial receptors (e.g., transferrin, insulin); (4) Carrier-mediated transport, which exploits endogenous solute transporters; (5) Magnetic guidance, where externally applied magnetic fields steer magnetic nanoparticles; (6) Adsorptive-mediated transcytosis, driven by electrostatic interactions between cationic nanocarriers and negatively charged endothelial membranes; (7) Virotechnological strategies, leveraging viral vectors or virus-like particles for efficient delivery; and (8) Use of exosomes and extracellular vesicles, which offer endogenous transport capabilities and biocompatibility.
Figure 3.
Mechanisms of nanoparticle transport across the BBB. NPs can traverse the BBB via multiple pathways, each with distinct biophysical and biochemical requirements. These include: (1) Passive transmembrane diffusion, mainly for lipophilic, low-molecular-weight compounds; (2) Paracellular transport, which is highly restricted by tight junctions; (3) Receptor-mediated transcytosis, utilizing ligands that bind to endothelial receptors (e.g., transferrin, insulin); (4) Carrier-mediated transport, which exploits endogenous solute transporters; (5) Magnetic guidance, where externally applied magnetic fields steer magnetic nanoparticles; (6) Adsorptive-mediated transcytosis, driven by electrostatic interactions between cationic nanocarriers and negatively charged endothelial membranes; (7) Virotechnological strategies, leveraging viral vectors or virus-like particles for efficient delivery; and (8) Use of exosomes and extracellular vesicles, which offer endogenous transport capabilities and biocompatibility.
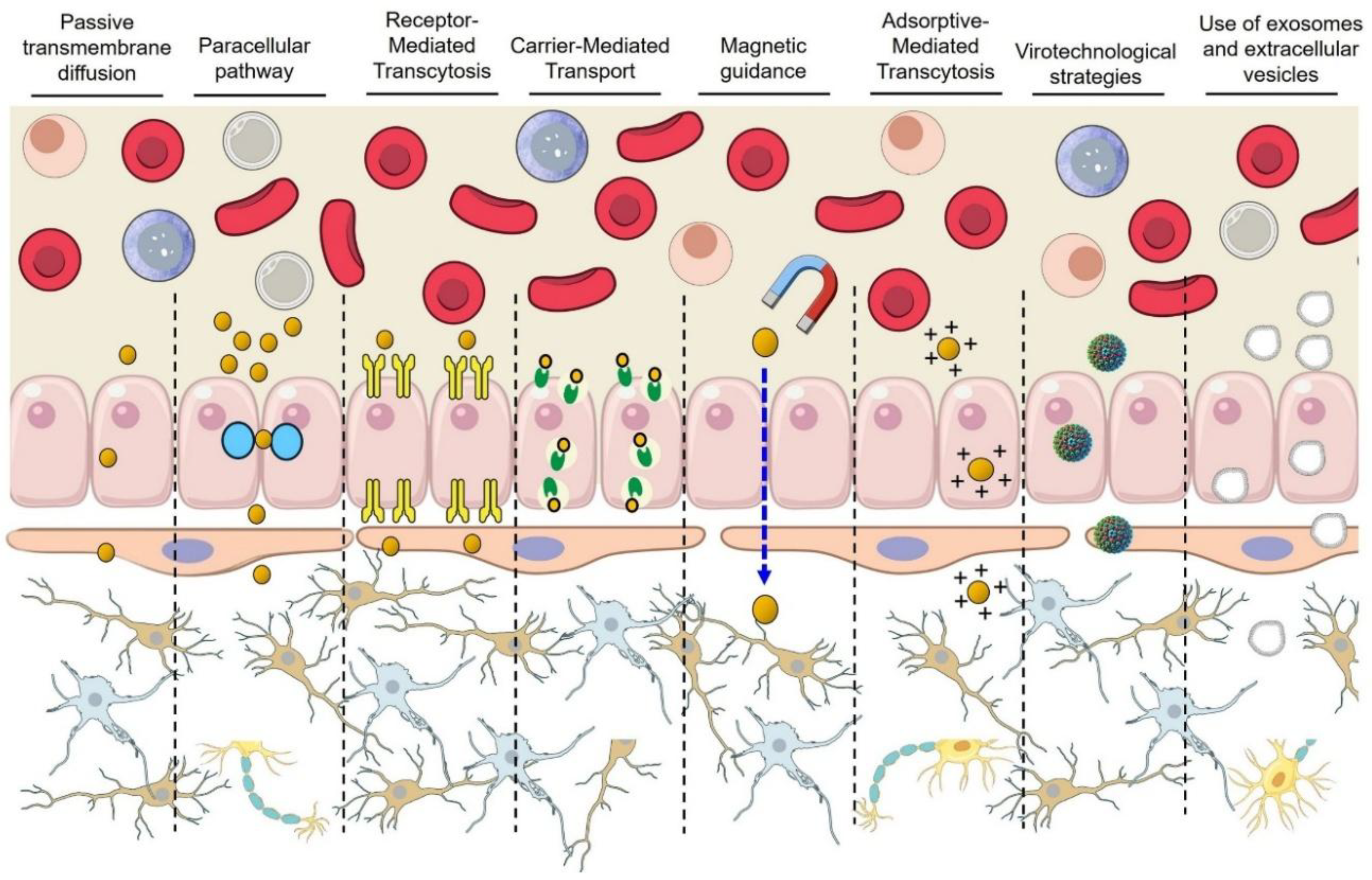

Table 1.
Summary of major transport mechanisms across the BBB.
| Mechanism | Molecule type | Size limit | Transport rate/efficiency | Key examples | References |
|---|---|---|---|---|---|
| Passive diffusion | Small, lipophilic | <400–500 Da | ~2–6% small molecules | O₂, CO₂, ethanol, nicotine | [28,29]. |
| Paracellular transport | Small ions, water | ~1–2 nm pore size | 10⁻⁷–10⁻⁸ cm/s (e.g., sucrose ~340 Da) | Water, sucrose | [6,21,22]. |
| CMT | Polar nutrients | Variable (nutrient-specific) | GLUT1: ~0.5 μmol/g/min (glucose uptake) | Glucose (GLUT1), leucine (LAT1) | [27]. |
| RMT | Specific peptides/proteins | Extensive (kDa range) | ~0.1–2% injected dose (antibody conjugates) | TfR, LDLR, insulin receptor | [30]. |
| AMT | Cationic/amphipathic macromolecules | Large (proteins, NPs) | ~10–20-fold increase (cationic albumin) | Cationic albumin, TAT peptide | [31]. |
CMT: carrier-mediated transport; GLUT1: glucose transporter 1; LAT1: large neutral amino acid transporter 1; RMT: receptor-mediated transcytosis; TfR: transferrin receptor; LDLR: low-density lipoprotein receptor; AMT: adsorptive-mediated transcytosis; TAT: trans-activator of transcription.
Table 2.
Experimental models for NPs delivery across the BBB.
| Model type | Specific description | Preparation/Sourcing | Key characteristics | Advantages | Limitations | Recommended use cases | References |
|---|---|---|---|---|---|---|---|
| In Vitro | Immortalized Cell Monolayer (hCMEC/D3) – human brain endothelial cell line grown as a monolayer on inserts | Human brain microvascular endothelial cells immortalized with hTERT and SV40 large T-antigen. Cells are obtained from repositories and cultured on collagen-coated Transwell inserts (e.g., 12-well) in defined medium. | Retains human BBB markers but forms a leaky barrier (TEER ~30–100 Ω·cm²). Low tight junction proteins cause high permeability. Expresses the efflux of transporters (e.g., P-gp) at reduced activity compared to in vivo. | Simple, robust, high-throughput human model. Ideal for rapid screening, reproducible monolayers, drug uptake, and toxicity assays. Expresses key BBB transporters and enzymes. | Very low barrier tightness (low TEER, high permeability). Incomplete tight junctions, limited BBB transporter expression. Poor predictor of in vivo permeability; lacks astrocytes, pericytes, and flow. |
This is a high-throughput initial screen for NP BBB penetration and cytotoxicity. It helps assess human-specific transport. The precision of the permeability measurement is limited; it should be validated in stricter models. |
[35,36,37,38]. |
| In Vitro | Primary Endothelial Co-culture, e.g., primary rodent or porcine brain endothelial cells with astrocytes/pericytes | Freshly isolated brain endothelial cells seeded on permeable inserts, co-cultured with astrocytes (direct/indirect) to induce BBB phenotype, with optional inclusion of pericytes or primary human cells. | It develops tight junctions and low permeability, like in vivo. Glial co-culture elevates TEER (porcine > rat > mouse). Expresses major BBB transporters (polarized P-gp, BCRP) and influx receptors (transferrin). Mimics in vivo drug exclusion (low paracellular flux). | Physiologically relevant model with high TEER, correct tight junction organization, and native efflux and nutrient transport activity. Supports endothelial–glial interaction studies and BBB regulation. Gold standard for drug permeability assays, closely matching in vivo results. | Labor-intensive, low yield requires fresh tissue and specialized isolation: high batch variability, short cell lifespan. Animal cells differ from human BBB. Primary human cells are scarce and quickly lose BBB properties (TEER drops to ~40–50 Ω·cm²). | Mechanistic studies under near-physiological conditions. Moderate-throughput screening when a tighter barrier is needed than cell lines. Validates findings from simpler models. Species choice tailored to study goals; confirmation in human models recommended. | [39,40,41]. |
| In Vitro | hiPSC-Derived BBB-on-Chip – human iPSC-derived endothelial cells with astrocytes/pericytes in microfluidic device. | hiPSCs differentiated to endothelial-like cells (Wnt/RA), cultured on Transwell or chip with astrocytes/pericytes. Shear flow and stimuli (e.g., hypoxia) improve maturation. | Human-like BBB phenotype: proper tight junction localization (claudin-5, ZO-1, occludin), high TEER (~1000–5000 Ω·cm²), and relevant transporter/receptor expression. Efflux pumps (P-gp, BCRP) are present but sometimes have reduced activity. Supports dynamic modulation. | Entirely human BBB model avoids species differences, maintains high barrier integrity (TEER >20,000 Ω·cm², >2 weeks), supports patient-derived iPSCs, and enables mechanistic and permeability studies. |
Reflects systemic influences (metabolism, protein binding, immune clearance). Measures real brain uptake and therapeutic effects. Genetically tractable. Cost-effective and accessible. | Human BBB model for preclinical NP testing. Ideal for studying transport mechanisms (receptor-mediated, junction modulation) and validating compounds targeting human-specific transporters. Best for focused, low-throughput studies. | [32,33] |
| In Vivo | Mouse Model – rodent in vivo BBB (adult mice, healthy or disease models) | Brain uptake is assessed via post-mortem analysis, imaging (MRI/PET), or genetic models. | Complete neurovascular unit with tight junctions, high TEER (~1000–6000 Ω·cm²), dynamic blood flow, active efflux, and realistic NP biodistribution. Note: Higher P-gp expression in mice than in humans. | Captures systemic factors influencing NP delivery. Directly measures brain uptake and efficacy. Genetically modifiable (e.g., Mdr1a^−/−). Cost-effective, suitable for disease modeling. | Species differences can underestimate human BBB uptake. The small size complicates surgeries and sampling. Limited blood volume and rapid metabolism impact NP circulation. Low throughput requires ethical approval. Results need confirmation in other models. | In vivo proof-of-concept to confirm NP BBB crossing and payload delivery. Mechanistic analyses and preclinical efficacy testing. Intermediate validation step: positive mouse results typically require follow-up in NHPs for human translation. | [34,42,43] |
| In Vivo | Non-Human Primate Model – rhesus or cynomolgus macaque BBB in vivo. | NPs are administered intravenously (often under anesthesia), with brain uptake monitored by MRI, PET, or post-mortem analysis. Allows repeated blood/CSF sampling. Small sample sizes (N≤4–6) due to cost and ethical constraints. | Most similar to human BBB in structure and function. Monkeys share transporter profiles highly similar to humans' (96% amino acid identity in P-gp). Comparable brain anatomy, capillary tight junctions, and pericyte coverage. Enables NP testing in a human-like brain. | Highly predictive of human BBB outcomes. Supports clinical imaging (PET, SPECT) for detailed in vivo NP tracking. Captures physiological BBB modulators, ensuring translational relevance. Essential for safety/toxicology evaluations required by regulatory agencies. | High cost, ethical, and logistical complexity limit throughput and statistical power. Requires specialized facilities and veterinary expertise. Minor physiological differences from humans exist. Handling stress and anesthesia may affect BBB properties. Genetic manipulation is impractical. |
Late-stage validation confirms NP BBB crossing, safety, and pharmacokinetics before human trials. The final translational step uses primate models with human-like BBB and metabolism. This step is not for screening but informs critical go/no-go decisions. | [44,45] |
| In Vivo | Zebrafish Larval Model – zebrafish embryo/larva with developing BBB | Transparent zebrafish embryos (~3 dpf) with functional BBB. NPs are administered by microinjection or water exposure. Fluorescent transgenic lines visualize NP BBB crossing in vivo. | Zebrafish BBB with tight junctions and conserved transporters form by 3–4 dpf, selectively restricting molecules similarly to mammals. Key regulators (e.g., Mfsd2a) share analogous functions. Enables live tracking of NP BBB crossing. | The high-throughput, low-cost in vivo model enables parallel testing, real-time imaging, and genetic manipulation, with fewer ethical constraints. | The non-mammalian model has differences in BBB maturity, immunity, and pharmacokinetics; there is a limited predictive value, requiring mammalian validation and injection variability for some NPs. | Rapid, early-stage in vivo screening of NP brain uptake and toxicity. Ideal for visualizing NP–BBB interactions. Helpful intermediate step before rodent studies; positive hits require mammalian validation. | [46,47,48]. |
Table 3.
Comparative data on NPs crossing the BBB.
| Class | NPs (formulation and surface) | Size | Zeta | Brain uptake | t₁/₂ | Therapeutic efficacy | Model | Reference |
|---|---|---|---|---|---|---|---|---|
| Lipid-based | Liposome – PEGylated (untargeted) | ~90 nm | Negative | 0.023 %ID/g (4 h post-IV) | Short (less than targeted) | N/A (no CNS therapy tested; baseline delivery) | Healthy mice (C57BL/6) | [80]. |
| Lipid-based | Liposome–scFv antibody-targeted (BBB receptor-specific) | ~90 nm | Negative | 0.24 %ID/g (4 h; ~10× over untargeted) | Longer circulation vs. Control | Improved brain drug levels (2-PAM); distribution study (no disease model) | Healthy mice | [80]. |
| Lipid-based | Liposome – TAT peptide-functionalized | ~100 nm (est.) | Cationic (+) | ~0.1 %ID/g (1 h; ~background level) | NA | N/A (no improved uptake; no efficacy) | Healthy mice | [81,82,83] |
| Polymeric | PLGA NP – Poloxamer 188-coated (MTX + PTX combo) | 133 nm and 221 nm | –29 mV / –18 mVnature.com | 17.2 %ID/g (48 h post-IV) | Detected in the brain up to 48 h | ↓ Tumor volume, Ki-67; improved survival vs. control | Rat glioma (C6 orthotopic) | [84] |
| Polymeric | PLGA NP – unmodified (PEG-PLGA) | ~100 nm (typical) | ≈ –15 mV (typical) | < 1 %ID/g (generally low) | Hours (moderate) | N/A (minimal BBB penetration) | Healthy rodents (general) | [85]. |
| Polymeric | PAMAM Dendrimer – G4 (OH-terminated) | 4.3 nm | ~0 mV | 1.9 ± 0.3 μg/g in tumor (24 h) | Rapid renal clearance | N/A (carrier targeted to microglia/Mϕ) | Rat 9L gliosarcoma / GL261 GBM | (86). |
| Polymeric | PAMAM Dendrimer – G6 (OH-terminated) | 6.7 nm | ~0 mV | 17.6 ± 4.5 μg/g in tumor (24 h) | Extended (slower clearance) | N/A (selective TAM uptake; immunotherapy vehicle) | Mouse GL261 GBM | [86]. |
| Protein-based | H-Ferritin nanocage (human heavy-chain) | ~12 nm | – (native) | Effective BBB penetration; slow clearance in brain | Long (persistent in brain) | N/A (proposed CNS drug carrier; no drug loaded) | Healthy mice | [87]. |
| Protein-based | Virus-Like Particle (JC polyomavirus VLP) | ~40 nm | NA | ~0 %ID/g (negligible brain uptake after IV) | NA | N/A (gene vector; no therapeutic cargo in study) | Healthy mice (IV vs. carotid) | [88,89]. |
| Inorganic | Gold NP – PEGylated (no targeting) | ~15 nm (core) | ~0 mV (PEG-coated) | 0.04 %ID/g (baseline) | ~2.3 h | N/A (used as BBB photomodulation agent) | Healthy mice | [90]. |
| Inorganic | Gold NP – anti-JAM-A antibody (BV11) coated | ~15 nm (core) | ~0 mV | 0.13 %ID/g (baseline; ~3× PEG-NP) | ~0.17 h (≈10 min) | N/A (facilitates laser-induced BBB opening) | Healthy mice | [90]. |
NA: Not assessed.Nanotechnological strategies for crossing the BBB.
Table 4.
Comparison of nanotechnological strategies for BBB crossing, highlighting the delivery system, mechanism, NP type, model, brain uptake, and therapeutic effects.
Table 4.
Comparison of nanotechnological strategies for BBB crossing, highlighting the delivery system, mechanism, NP type, model, brain uptake, and therapeutic effects.
| Strategy and system | Target/Mechanism | NP or vector | Model used | Brain uptake metrics | Therapeutic outcome | References | |
|---|---|---|---|---|---|---|---|
| TfR-mediated (RMT), e.g., OX26 antibody NP. | Transferrin receptor on the BBB endothelium. | OX26-conjugated PEGylated liposome or gold NP. | Rat (in vivo) | ~0.3% ID in brain vs 0.03% for IgG (10-fold increase); parenchymal 0.23% ID/g with optimized affinity. | Enhanced brain drug levels; basis for enzyme therapy (ETV:IDS) yielding 50–76% substrate reduction in CNS. | [91,106,126]. | |
|
T7-PLGA NPs |
Transferrin receptor (TfR) targeting, T7 peptide (HAIYPRH) binds TfR on the BBB endothelium, triggering receptor-mediated transcytosis. | PLGA polymer NPs decorated with T7 peptide (often PEGylated; can carry drugs or genes). | Murine brain tumor models (orthotopic glioma) and healthy mice (distribution studies). | T7-functionalized NPs increased brain accumulation by ~6 fold, gene expression by 1.7 fold, and photosensitizer delivery to gliomas by ~6 fold versus untargeted controls. |
T7-targeting improved outcomes: T7-liposomes (ZL006) reduced infarct volume and improved neurological recovery in stroke; T7-NPs enhanced tumor suppression and survival in glioma. | [127,128,129,130]. | |
| INSR-mediated (RMT), e.g., 29B4 antibody. | Insulin receptor (ubiquitous, BBB, and neurons). | Human insulin receptor mAb (29B4) on HSA NP. | Mouse (in vivo) | Qualitative crossing confirmed (therapeutic levels achieved); clinical fusion protein ~2–3% CSF: plasma ratio in patients (phase 1). | CNS enzyme delivery in Hunter syndrome (valanafusp alpha) – reduced CNS pathology; in rodents, INSR-NPs showed functional neuroprotection. | [93,94,131]. | |
| LRP1-mediated (RMT): Angiopep-2 peptide. | LRP1 on the endothelium (also in tumors). | Angiopep-2 decorated polymeric NP. | Mouse (in vivo); in vitro BBB models. | ~2–4× higher brain uptake vs non-targeted NP (biodistribution studies); transcytosis of Ang2-NPs observed in iPSC-derived human BBB model. | In patients, Ang2-NP delivering paclitaxel (ANG1005) showed tumor shrinkage; Ang2-polymersomes with carnosine reduced stroke infarct volume. | [95,96,132]. | |
| Angiopep-2 lipid–silica NPs | Angiopep-2 targets LRP1 receptors on the BBB and glioma cells, mediating transcytosis into the brain. | Lipid-coated mesoporous silica NPs loaded with paclitaxel and functionalized with Angiopep-2. | Rat intracranial glioma model (C6 glioma-bearing rats; IV administration). | Angiopep-2 NPs enhanced paclitaxel brain delivery (~20.6% vs. ~10.6% targeting efficiency), doubling brain drug concentrations compared to untargeted controls. | Angiopep-2 targeting enhanced brain tumor therapy, prolonging survival and increasing tumor apoptosis compared to untargeted NPs. | [133] | |
| Lactoferrin R-mediated (RMT) – Lf-NC. | Lactoferrin receptor (on BBB and glioma cells). | Lactoferrin-coated lipid nanocapsule (Lf-LNC). | Rat (PTZ epilepsy model). | Brain APO concentration ↑ (significant, e.g., 1.5-fold vs uncoated); Lf coating improved BBB permeability. | Suppressed seizures: ~0.67 Racine score with Lf-LNC vs ~3 (uncoated); reduced neuroinflammation. | [100,101,134]. | |
| Folate-mediated (RMT) – FA-NP. | Folate receptor-α (high in glioma, low BBB). | Folic acid-conjugated gold NP (or polymer NP). | Mouse glioma (orthotopic) | Tumor: brain ratio >5:1 uptake in FR-positive tumor; minimal uptake in normal brain. | Enhanced GBM cell kill and imaging contrast; extended survival in folate-R expressing tumor models. | [104,105,135]. | |
| Adsorptive (AMT) – PepH3 peptide NP. | Electrostatic adsorptive uptake. | PepH3 (7-aa cationic) tagged vesicular NP. | Rat and human BBB cell culture; Mouse IV. | Endothelial uptake ↑ (~3–5× vs no peptide); in vivo high brain localization, low off-target (radiotracer). | Delivered anti-Aβ single-domain antibody across the BBB in vitro; potential Alzheimer’s therapy shuttle (in vivo efficacy pending). | [107,108,109]. | |
| Adsorptive (AMT) – Chitosan NP. | Electrostatic (polycationic polymer) | Chitosan DNA NP (~260 nm) | Mouse (in vivo, i.p. injection). | Confirmed BBB crossing: GFP gene expressed in brain cells; brain transfection efficiency ~53% (FACS, vs 27% with control vector). | Successfully expressed therapeutic gene (GFP) in brain parenchyma; proof-of-concept for gene therapy in brain tumors or neurodegeneration. | [111] | |
| Magnetic Targeting – Liposomal SPION (LTF). | External static magnetic field (SMF) guides NP. | Temozolomide + ferucarbotran liposome (LTF). | Mouse glioma (GL261 in the brain). | Tumor NP concentration was ~2 times higher with the magnet (MRI-based); the magnet-guided group showed a p < 0.01 reduction in tumor volume by day 7. | Tumor growth suppressed; Median survival ↑ vs non-magnet (e.g., ~25 days to >31 days with magnet). | [114]. | |
| Magnetic nanocapsules | Magnetic targeting via external field enhances iron oxide nanocapsule transcytosis across the BBB. | ~100 nm silica-coated magnetic NPs (iron oxide core) with RF-triggered drug release capability. |
Healthy mice with intact BBBs were subjected to localized magnetic targeting post-IV injection. | Localized magnetic fields increased brain NP delivery by ~25–26-fold versus controls; ~30% of peak brain signal persisted at 48 h, while non-magnetized delivery remained near background levels. | Magnetic nanocapsules enabled non-invasive BBB crossing without acute toxicity; histology confirmed vessel integrity. Though therapeutic efficacy remains untested, they allow on-demand drug release via radio-frequency heating. | [136,137] | |
| Magnetic Targeting, Tween-SPION. | Magnetic field induces BBB transport. | 20 nm Tween-80 coated SPIONs. | Rat (normal BBB, iv + EMF). | Crossed intact BBB under EMF; SPIONs detected in brain parenchyma (none without EMF). | No therapeutic payload (diagnostic); demonstrates non-invasive BBB crossing by physical force. | [114] | |
| Viral Vector – Engineered AAV (CAP-Mac). | Capsid-mediated transcytosis (evolved tropism). | AAV.CAP-Mac (neurotropic AAV variant). | Non-human primates (marmoset, macaque). | ~1.1–1.3% of all neurons transduced (green monkey) vs <0.5% with AAV9; broad CNS distribution (11 of 11 regions positive). | Enabled IV gene delivery: e.g. widespread GCaMP expression for imagingnature.com; supports CNS gene therapy (potential for autism, Alzheimer’s). | [117]. | |
| Viral Vector – RVG-pseudotyped LV. | Viral glycoprotein-mediated entry. | Lentivirus coated with RVG peptide. | Mouse (in vivo). | Qualitative BBB crossing (RVG-LV detected in brain, unlike unmodified LV); transgene in neurons. | Partial motor function restoration in a neurodegenerative mouse model (using RVG-LV to deliver therapeutic gene). | [116,123]. | |
| TMV-VLPs | Size/shape EPR + ligand (albumin, etc.) | Tobacco Mosaic Virus nanorod + albumin coat | Mouse (brain tumor model) | Accumulated in brain tumor (MRI and NIR imaging); higher tumor: normal brain ratio than spherical NP. | Improved tumor imaging and delivery of photothermal therapy; significant tumor cell apoptosis in combination treatment. | [68,138]. | |
| CCMV VLPs | None (passive) – Natural 28 nm protein cage (plant virus capsid) with no specific targeting; crosses BBB at low levels, possibly via adsorptive transcytosis. | Empty CCMV capsid as a drug nanocarrier. | Healthy mice (IV injection, no disruption of BBB). | Approximately 0.3% ID/g was detected in the brain at one h post-injection, decreasing to <0.01% by 24 h; brain distribution was comparable to other protein-based NPs. |
No therapeutic payload was tested; safety studies showed no overt toxicity or immune response in mice. | [139]. | |
| Exosome – RVG-modified MSC exosomes | Endogenous vesicle uptake + neuron targeting | RVG-peptide engineered exosomes (MSC-derived) | Mouse Alzheimer’s model | Preferential localization to cortex/hippocampus; exosomal cargo (siRNA) in brain increased ~2× vs free siRNA. | Restored memory function (exosome-treated mice performed significantly better in Morris water maze)mdpi.com; reduced brain Aβ and inflammation. | [123,140,141]. | |
| Exosome – Cetuximab-Exo-Dox | Endogenous vesicle + tumor targeting (EGFR) | Exosomes loaded with doxorubicin + Cetuximab | Mouse glioblastoma model | Brain delivery of cetuximab ↑ (~2-fold) with exosomes vs free Ab; doxorubicin brain concentration also higher (HPLC quantification). | Enhanced GBM growth inhibition and prolonged survival vs free drug; exo combo therapy induced greater tumor cell apoptosis (histology). | [124]. | |
| MSC-derived exosomes | MSC-derived exosomes (~50–150 nm) cross the BBB via endocytosis, with enhanced uptake under inflammatory conditions. |
MSC-derived EVs carrying therapeutic cargo (proteins/miRNA or drugs). | Rodent models of CNS injury (stroke, TBI) for therapy; healthy rats for biodistribution. | Baseline brain uptake was low (~0.03–0.04% ID/g) after IV administration; entry increased in neuroinflammatory models with preferential accumulation in injured regions. |
MSC-exosomes reduced infarct volume by ~50%, improved neurological function in stroke, and attenuated neuroinflammation with cognitive recovery in TBI models. | [142,143,144,145,146,147,148]. | |
| Micromotor – NIR Janus nanomotor | Photothermal propulsion (active movement) | Gold-Janus NPs (NIR-responsive) | Mouse (in vivo experiment) | BBB penetration significantly improved under NIR (qualitative: increased dye leakage into brain); no crossing without NIR. | Facilitated brain delivery of a model drug (dye) with spatiotemporal control; concept validated for on-demand BBB opening. | [149]. | |
| Macrophage-mediated “Trojan Horse” delivery | Monocytes/macrophages naturally cross the BBB, delivering internalized drug-loaded NPs to inflamed or tumor sites. | Macrophages loaded ex vivo with drug-encapsulated NPs, then injected intravenously. |
Mouse glioblastoma and neuroinflammation (e.g., Parkinson’s) models using macrophage adoptive transfer. |
NP-loaded macrophages greatly enhanced brain tumor localization versus free NPs; intrathecal macrophage transfer achieved ~8.1% ID/g brain uptake, surpassing standard IV delivery. | Macrophage-mediated delivery improved glioma drug deposition, potentially limiting tumor growth, and increased GDNF levels with functional recovery in Parkinson’s, leveraging immune-cell homing to the CNS. | [150]. | |
| Microrobot – Magnetic spiral (platelet cloaked) | Magnetic rotation (swimming) | Helical nanorobot with Fe coating + platelet membrane | In vitro blood flow; proposed in vivo mouse | Propulsion sustained in blood-mimicking flow; able to navigate and marginate toward vessel walls. (BBB crossing has not yet been directly measured). | Demonstrated long circulation and targeting potential; aims to mechanically traverse the BBB and deliver drugs (studies in progress). | [151]. | |
Quantitative brain uptake is given where available (percentage of injected dose in brain, %ID/g tissue, or fold-change vs controls). ID: injected dose; TfR: transferrin receptor; INSR: insulin receptor; LRP1: low-density lipoprotein receptor-related protein-1; FR: folate receptor; RMT: receptor-mediated transcytosis; AMT: adsorptive-mediated transcytosis; SPION: superparamagnetic iron oxide NP; EMF: electromagnetic field; AAV: adeno-associated virus; LV: lentivirus; VLP: virus-like particle; RVG: rabies virus glycoprotein peptide; MSC: mesenchymal stem cell; EGFR: epidermal growth factor receptor; NIR: near-infrared; GCaMP: calcium indicator gene; siRNA: small interfering RNA: MSC: Mesenchymal stem cell; EVs: extracellular vesicles; TBI: traumatic brain injury; CCMV-VLPs: Cowpea chlorotic mottle virus-like particles; RF: remotely heated; TMV: Tobacco mosaic virus-like particles.
Table 5.
Toxicological parameters, regulatory requirements, and scale-up challenges for BBB-targeted nanomaterials.
Table 5.
Toxicological parameters, regulatory requirements, and scale-up challenges for BBB-targeted nanomaterials.
| Parameter | Recommended studies/assays | Minimal data required (thresholds) | Key considerations | Reference |
|---|---|---|---|---|
| Hemocompatibility (blood compatibility of IV nanomedicine) |
In vitro blood tests: Hemolysis assay (human RBCs); Complement activation (C3a, C5a, SC5b-9); Platelet aggregation/coagulation (platelet markers, thrombin, aPTT, PT). |
Hemolysis: % hemoglobin release; < 5% considered low risk (non-hemolytic). (ISO 10993-4 standard). Ideally, <2% (negligible hemolysis). Complement: No abnormal complement consumption or excessive anaphylatoxin rise (C3a/C5a) compared to negative control (i.e., should not trigger significant CARPA). Platelets/Coagulation: No significant platelet aggregation or >10% change in clotting times relative to baseline. (No official numeric limit; ensure values remain in a normal range of variation.) |
Use fresh human blood to avoid species-specific platelets and complement differences. Account for NP interference with assay readouts via proper controls. Follow ISO 10993-4 and ASTM F756 standards to ensure hemocompatibility (to prevent thrombosis, hemolysis, and infusion reactions). If complement activation occurs in vitro, evaluate the risk of infusion reaction in vivo and consider methods for complement inhibition. |
[152,154,159,160,161,162,163,164,165]. |
| Neurotoxicity (CNS behavioral and histopathological safety) |
Safety pharmacology and neurotoxicity: Perform FOB or modified Irwin test (locomotion, reflexes, coordination, sensorimotor responses, convulsions). Include behavioral assays (open field, rotarod, cognition if needed). Assess CNS histopathology (brain, spinal cord) in subchronic/chronic studies for neuronal/glial damage, inflammation, or vacuolization. |
Neurobehavioral outcomes: No significant adverse effects on motor activity, gait, reflexes, or behavior at therapeutic levels. Minor changes (<20% compared to the control), reversible, and dose-dependent effects are acceptable. Neuropathology: No significant CNS lesions (neuronal degeneration, gliosis, demyelination) exist beyond background; minimal changes are allowed only at doses above therapeutic exposure. Neurofunctional tests: Grip strength, rotarod, and maze performance impairment should remain minimal (<10–15%) at clinical doses, excluding sedation effects. |
FDA S7A Guidance: Evaluate CNS effects (behavior, reflexes, coordination, temperature), especially for CNS-targeted NPs. Assess neuroinflammation (microglia/astrocytes: Iba1, GFAP). Include behavioral tests for chronic CNS exposure. Use recovery groups to check reversibility. |
[154,166,167,168,169]. |
| Long-term Accumulation (Brain retention and clearance) |
Biodistribution studies: Labeled NPs (radioactive/fluorescent) track long-term brain and organ distribution. Multiple time-point assessments (weeks to months) are conducted, and imaging methods (PET/MRI) are preferred for non-invasive monitoring. Chronic toxicity: Extended observation periods post-treatment should be included to evaluate CNS persistence and delayed neurotoxicity. Brain and CSF should be analyzed at intervals to assess clearance. |
Brain retention half-life: Biodegradable NPs should clear significantly (>50% within weeks); non-biodegradable NPs must plateau without progressive accumulation. Residual brain burden: post-treatment brain levels should substantially decrease (<10% peak) within 1–3 months. Clearance pathways: Identify clearance routes (e.g., glymphatic). Rapid clearance (hours–days) is preferred; persistent presence (>6 months) needs justification. |
NP biodegradability: Assess persistence of nonbiodegradable NPs (e.g., gold, silica) and potential chronic neurotoxicity. Biological fate (FDA): Evaluate NP distribution, accumulation, and clearance from the brain and organs. Brain clearance: Examine glymphatic and phagocytic pathways; test in healthy and impaired clearance models. Human translation: Use animal retention data to inform human risk; persistent retention may require clinical imaging or dose adjustments. |
[139,156,167,170,171,172]. |
| Biodistribution and Pharmacokinetics (PK) (Systemic and CNS distribution, drug exposure) |
Animal ADME studies: Radiolabeled or tracer methods track the distribution of NPs and their payloads over time. They measure plasma pharmacokinetics (Cmax, T½, AUC, clearance) and tissue distribution (e.g., percentage of dose in brain vs. organs). Brain penetration metrics: Calculate brain: plasma ratios or brain targeting indices. Include CSF levels if relevant. PK modeling: Apply compartmental or PBPK models using NP properties to predict human PK and dosing. |
Animal ADME studies: Radiolabeled/tracer methods measure the distribution of nanoparticles and their payloads in plasma, brain, and organs over time. Report plasma PK (C_max, T_½, AUC, clearance) and tissue distribution (% dose in brain vs. other organs). Brain penetration: Calculate brain: plasma ratios or targeting index; include CSF levels if relevant. PK modeling: Apply compartmental or PBPK models with NP parameters to predict human PK and dosing. |
Plasma half-life (T<sub>1/2</sub>): Report half-life versus free drug; nanoformulations typically extend circulation time (~10× longer than expected). Brain uptake: Higher brain/plasma ratios (>0.1 generally, >1.0 if targeted) indicate improved CNS targeting compared to a free drug. Bioavailability/distribution: Quantify brain delivery fraction. Minimize systemic exposure intrathecally; characterize off-target accumulation intravenously. PK linearity: Confirm dose-proportional exposure; investigate significant non-linearities (e.g., saturation, aggregation). |
[153,156,157,173]. |
| Immunogenicity and immunotoxicity (Regulatory requirement) |
In vitro assays: Cytokine release (IL-6, TNFα, IFNγ) in human PBMCs. Complement activation (C5a, SC5b-9; CARPA risk). Immune cell function (macrophage uptake/ROS, T-cell activation, dendritic cell maturation). Myelosuppression (bone marrow colony assays). In vivo immunotoxicity: Evaluate immune organ histopathology (spleen, lymph nodes) and blood leukocyte subsets. If immunotoxic signals appear, targeted studies should be conducted per ICH S8. Monitor anti-drug antibodies (ADAs), especially against proteins or PEG. |
Cytokine induction: Minimal cytokine release (e.g., IL-6 <3× baseline). Use controls for comparison. Complement activation: Low complement activation (<50% positive control). High levels signal hypersensitivity risk. Immunogenic antibodies: Monitor anti-nanoparticle antibodies; incidence typically ≤20%. Significant anti-PEG IgM or clearance changes (ABC phenomenon) require attention. Immune cell counts: Maintain WBC subsets within ±30% of control. Investigate consistent suppression or activation (e.g., T-cell drop, eosinophilia). |
Nanomedicine immunogenicity: Assess risks (patient, route, dose). Include NP-specific assays (complement, inflammasome, immune cells). Carrier vs. Payload: Identify carrier (anti-PEG) vs. payload reactions; mitigate significant responses (e.g., ABC phenomenon). Clinical monitoring: Monitor immune reactions (anaphylaxis, complement). Investigate mechanisms; justify and plan mitigation strategies. |
[156,157,162,174,175]. |
| Scale-up and reproducibility | Chemistry, Manufacturing and Controls (CMC): Implement robust cGMP processes. Characterize NP CQAs: particle size (DLS, laser diffraction), ζ-potential, morphology (TEM), drug loading/encapsulation (HPLC, spectroscopy), purity, endotoxin (LAL), and sterility (injectables). Conduct stability studies (size, potency, aggregation over time). Scale-up validation: Using statistical quality control, ensure batch consistency (size, PDI, drug content/release) between pilot and production scales. |
Particle size and PDI: Maintain consistent size (±10% target), low PDI (<0.3; ideally ≤0.2). Avoid aggregates (>1000 nm). Drug content/release: Drug content within 90–110% of label; consistent batch-to-batch release profiles. Other CQAs: Stable zeta potential, impurities within ICH Q3D, endotoxin below USP limits (<5 EU/kg). Reproducibility: Consistent CQAs batch-to-batch (%RSD <5–10%). Scale-up should not affect critical attributes. |
A QbD approach is recommended to control critical parameters, ensuring consistent NP quality at scale. Analytical methods require NP-specific validation, reporting particle size confirmed by orthogonal methods. Adjustments during scale-up (e.g., homogenization parameters) and documented comparability (FDA/EMA guidelines) are essential. Regulatory compliance requires cGMP manufacturing, aseptic processing (especially if >200 nm), comprehensive CQA testing, and detailed CMC documentation before clinical approval. | [156,176,177,178]. |
ADAs: Anti-Drug Antibodies; ADME: Absorption, Distribution, Metabolism, Excretion; AMT: Adsorptive-Mediated Transcytosis; AUC: Area Under Curve; ASTM: American Society for Testing and Materials; BBB: Blood–Brain Barrier; CARPA: Complement Activation-Related Pseudoallergy; CMC: Chemistry, Manufacturing, and Controls; CNS: Central Nervous System; CQAs: Critical Quality Attributes; CSF: Cerebrospinal Fluid; DLS: Dynamic Light Scattering; EMA: European Medicines Agency; FDA: Food and Drug Administration; FOB: Functional Observational Battery; GFAP: Glial Fibrillary Acidic Protein; HPLC: High-Performance Liquid Chromatography; ICH: International Council for Harmonisation; IL-6: Interleukin-6; IV: Intravenous; LAL: Limulus Amebocyte Lysate; MRI: Magnetic Resonance Imaging; NOAEL: No Observed Adverse Effect Level; OECD: Organisation for Economic Co-operation and Development; PBMCs: Peripheral Blood Mononuclear Cells; PBPK: Physiologically Based Pharmacokinetic; PDI: Polydispersity Index; PEG: Polyethylene Glycol; PET: Positron Emission Tomography; PK: Pharmacokinetics; PT: Prothrombin Time; QbD: Quality by Design; RBCs: Red Blood Cells; RES: Reticuloendothelial System; ROS: Reactive Oxygen Species; TEM: Transmission Electron Microscopy; TNFα: Tumor Necrosis Factor-alpha; WBC: White Blood Cell; ζ-potential: Zeta Potential. Notes: When specific quantitative criteria are not established in guidelines, the values above are based on standard industry practices or literature and should be interpreted as general guidance, not fixed regulatory limits.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



