
Submitted:
27 September 2024
Posted:
29 September 2024
You are already at the latest version
Abstract
The blood-brain barrier (BBB) strictly regulates the penetration of substances into the brain, which, although important for maintaining brain homeostasis, may delay drug development because of the difficulties in predicting pharmacokinetics/pharmacodynamics (PKPD), toxicokinetics/toxicodynamics (TKTD), toxicity, safety, and efficacy in the central nervous system (CNS). Moreover, BBB functional proteins show species differences; therefore, humanized in vitro BBB models are urgently needed to improve the predictability of preclinical studies. Recently, international trends in the 3Rs in animal experiments and the approval of the FDA Modernization Act 2.0 have accelerated the application of microphysiological systems (MPSs) in preclinical studies, and in vitro BBB models have become synonymous with BBB-MPSs. Recently, we developed an industrialized humanized BBB-MPS, BBB-NET. In our previous report, we reproduced transferrin receptor (TfR)-mediated transcytosis with high efficiency and robustness with hydrogels that include fibrin and collagen I microfibers (CMFs). Herein, we investigated how adding CMFs to the fibrin gel benefits BBB-NETs, showed that CMFs accelerated capillary network formation and maturation by promoting astrocyte (AC) survival, and clarified that integrin 1 is involved in the mechanism of CMFs. Our data suggest that quality control (QC) of CMFs is important for ensuring the stable production of BBB-NETs.
Keywords:
blood-brain barrier (BBB)
; microphysiological system (MPS)
; collagen I microfiber (CMF)
; integrin
; humanized model
1. Introduction
The blood vessels in the central nervous system (CNS) limit the permeability of circulating substances into the brain via the blood-brain barrier (BBB) and protect the CNS from circulating xenobiotic compounds [1]. Essential nutrients (glucose, etc.), on the other hand, are transported by specific transporters on vascular endothelial cells [2,3]. These functions of the BBB cause delays in drug development because it is difficult to predict the pharmacokinetics/pharmacodynamics (PKPD) [4], toxicokinetics/toxicodynamics (TKTD), toxicity, safety, and efficacy in the CNS [5,6]. In addition, BBB functional proteins show species differences [7,8]. Therefore, humanized in vitro BBB models are urgently needed to improve the predictability of preclinical studies [5]. In recent years, international trends in the 3Rs in animal experiments and the approval of the FDA Modernization Act 2.0 have accelerated the application of the microphysiological system (MPS) in preclinical studies. In vitro BBB models have recently become synonymous with BBB-MPS and are being developed in both industry and academia to respond to the BBB context of use (CoU).
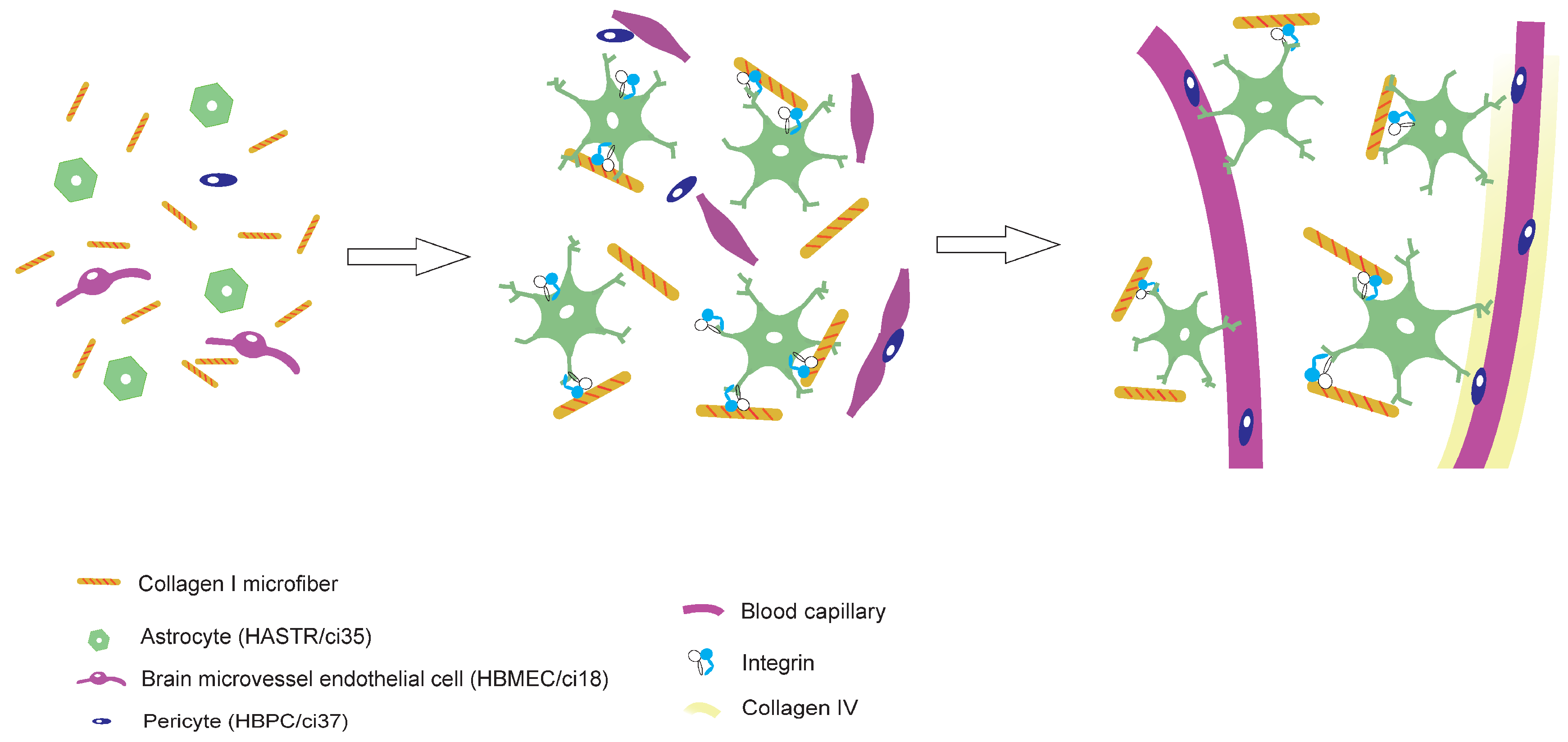
In vivo, the BBB mainly consists of endothelial cells (ECs), pericytes (PCs), basement membrane proteins, and astrocyte (AC) endfeet. Two categories of cell-based BBB-MPSs have been developed thus far: two-dimensional (2D) and three-dimensional (3D). In 2D BBB-MPSs, ECs, PCs, and ACs are arranged in a transwell set in a multiwell plate. These models have been used for drug permeability tests for over 40 years [9,10,11]. Beside organoids, the first report of 3D BBB-MPSs was in 2017. Our group also developed 3D BBB-MPSs, in which immortalized human ECs (HBMECs/ci18), PCs (HBPCs/ci37), and ACs (HASTRs/ci35) self-organize to form a brain capillary network in hydrogels with a BBB [12] (Fig. 1A). The EC monolayers seeded on the bottom side of the hydrogel make opening structures that join the capillaries in the hydrogel. When drugs are placed outside the hydrogel, drug permeability can be assessed by measuring the amount of drug that permeates the BBB and moves into the supernatant above the hydrogel. We have succeeded in reproducing transferrin receptor (TfR)-mediated transcytosis in a efficient and robust manner [13]. We are now working on making this BBB-MPS into an industrial product, denoted BBB-NET.
The BBB-NET hydrogels include fibrin and collagen I microfibers (CMFs). Each component has been reported to play important roles in vasculogenesis [14,15,16,17]. Shang F et al. reported that the size and number of the opening structures on the bottom of BBB-NET depend on the fibrin concentration. In a previous study, hard hydrogels (33 mg/ml fibrinogen) induced more active capillary organization than the soft hydrogels with 7 mg/ml fibrinogen did [18]. Collagen I has been used in tissue engineering and regenerative medicine [9,19] including in vitro BBB models [20] for several decades; however, the detailed effects of CMFs has not been confirmed. In this study, we investigated the advantages of using CMFs in addition to fibrin gel to fabricate BBB-NET. We showed that the CMFs accelerated the formation and maturation of the capillary network by promoting AC survival. We also determined that integrin b1 is involved in the CMF mechanism. Our data suggest that quality control (QC) of CMFs is important for ensuring the stable production of BBB-NET.
2. Materials and Methods
2.1. Materials
VascuLife basal medium (cat. no. LL-0002) and VEGF LifeFactors kit (cat. no. LS-1020) was purchased from Kurabo (Osaka, Japan). Pericyte medium (cat. no. 1201) was purchased from ScienCell Research Laboratories (Carlsbad, CA, USA). Blasticidin S (cat. no. 029-18701) was purchased from Fujifilm Wako (Tokyo, Japan). Astrocyte growth medium (cat. no. A126130), fibronectin (cat. no. 33016015), anti-rabbit IgG conjugated with Alexa Fluor 488 (cat. no. A11031), anti-rabbit IgG conjugated with Alexa Fluor 647 (A21244), anti-rat IgG conjugated with Alexa Fluor 488 (cat. no. A11006), anti-mouse IgG conjugated with Alexa Fluor 488 (cat. no. A11029), anti-rabbit IgG conjugated with Alexa Fluor 647 (cat.no. A31571), and Cell Tracker (cat. no. C7025) were purchased from Thermo Fisher Scientific (Waltham, MA, USA). The anti-CD31 antibody (cat. no.66065-2-1g) was obtained from Proteintech (Rosemont, IL, USA), the anti-CD146 antibody (cat. no. ab75769), anti-integrin b1 antibody (cat. no. ab24693), and mouse IgG, Kappa monoclonal, isotype control (cat. no. ab91353) were purchased from Abcam (Cambridge, UK), and the anti-collagen IV antibody (cat. no. SGE-C-425) was purchased from COSMO BIO (Tokyo, Japan). The anti-aquaporin 4 antibody (cat. no. A5971), fibrinogen (F2006-5MG), and thrombin (T4648-10KU) were purchased from Merk (Darmstadt, Germany). DAPI (cat. no. 342-07431) was purchased from Dojindo (Tokyo Japan). RapiClear 1.47 (cat. no. RC147001) and iSpacer (cat. no. IS002) were obtained from SunJin Laboratory (Hsinchu City, Taiwan, R.O.C.).
2.2. Cell Culture
Human brain microvascular endothelial cells/conditionally immortalized clone 18 (HBMECs/ci18) [21,22], human brain pericytes/conditionally immortalized clone 37 (HBPCs/ci37) [23], and human astrocytes/conditionally immortalized clone 35 (HASTRs/ci35) [24] were established by Prof. Furihata T., et al. For maintenance, HBMECs/ci18 were grown in VascuLife complete medium, while HASTRs/ci35 and HBPCs/ci37 were cultured in astrocyte growth medium and pericyte medium, respectively. The composition of the medium was described in reference 25, and all culture media contained 4 mg/ml blasticidin S. The cells were cultured at 33 °C for growth and at 37 °C for differentiation [24,25].
2.3. CMFs and BBB-NET Preparation
Collagen I microfibers (CMFs) were established by Prof. Matsusaki [26] and supplied by the Toppan Technical Research Institute. Gel preparation was described by Agathe F., et al. [12]. Briefly, 70 μL of gel containing 0.7 mg of CMFs was mixed with 0.4 mg of fibrinogen in 40 μL of DMEM in one tube, while in another tube, 2.0×105 HBMECs/ci18, 4.0×105 HASTRs/ci35, and 1.0×105 HBPCs/ci37 were mixed with 0.3 U of Thrombin in 30 μL of VascuLife complete medium. The solution was mixed just before being added to the top of a culture insert previously plasma treated and coated with fibronectin. After 45 min of gelation, 2 mL of mixed media (1:1:1 complemented VascuLife: Pericyte medium: astrocyte medium) was added. The gels were incubated for 7 days at 37 °C in a humidified atmosphere with 5% CO2, and the media was changed every 3–4 days (Fig.1A).
2.4. Astrocyte (HASTR/ci35) Labelling
HASTRs/ci35 (1.0×106 cells) were centrifuged, and the supernatant was aspirated. The cells were then gently resuspended in prewarmed Cell Tracker working solution (Thermo Fisher Scientific). After 45 min of incubation at 37 °C, the cells were centrifuged, and the working solution was removed.
2.5. Blocking the CMF–Integrin Interaction in Astrocytes
2.6. Immunostaining
After 7 days of culture, the 3D BBB models were with 4% paraformaldehyde (Fujifilm Wako, Osaka, Japan) for 20 min of incubation at room temperature, after which the cells were washed with PBS three times before 2 hr of incubation in blocking solution (10% normal goat serum and 0.2% Triton-X in PBS) at room temperature. The samples were incubated with primary antibody for 2 days at 4 °C in 1% normal serum with 0.2% Triton X-100. The concentrations of the primary antibodies against the human proteins used were as follows: anti-CD31 (200×), anti-CD146 (200×), and anti-collagen IV (50×). The cells were then washed with 0.1% Triton-X in PBS and incubated with PBS containing secondary antibodies (anti-rabbit IgG conjugated with Alexa Fluor 488 or 647 anti-rat IgG conjugated with Alexa Fluor 488, or anti-mouse IgG conjugated with Alexa Fluor 488 or 647; each 500×) for 3 hr at room temperature. The samples were washed and counterstained with DAPI (1000×). After the samples were washed with PBS 3 times, they were cleared with RapiClear according to the manufacturer’s instructions until the samples became transparent. The gels were removed and sandwiched between coverslips using iSpacer (Sunjin Laboratory).
2.7. Analysis of Protein Expression in the 3D BBB Models
Fluorescence images were obtained via a Nikon A1R-A1 confocal microscope system (Nikon, Tokyo, Japan). Z-stack images were taken 30-80 μm from the bottom of the gel at 20× magnification. Four images were taken from each sample. To reconstruct the stained images as 3D images, the Surface tool in IMARIS software v. 9.8.2 (Oxford Instrumentals, Abingdon, UK) was used with the following four steps: Default basic algorithm, Source channel, Threshold, and Classify surface steps. The lowest threshold was set manually so that the target protein signal intensity could be differentiated from the background signal. Nonspecific signals were removed by the Filter program in Classify Surface mode. The 3D rendered images were subsequently constructed, and representative images are shown in Fig. S1. All the images were processed with the same parameters. The vessel volume was calculated and summed according to our previous methods [30,31].
2.8. Statistical Analysis
All data are expressed as the means ± standard deviation unless otherwise noted. Statistical analyses were performed as shown in the figure legends, and p values less than 0.05 were considered to indicate statistical significance. The results were confirmed by 2-3 independent experiments.
3. Results
3.1. The Collagen I Microfibers in BBB-NET Hydrogel Enhanced Autonomous Brain Capillary Network Formation
To clarify the role of the CMFs in the BBB-NET in terms of capillary network formation, we first compared the capillary networks in the hydrogels made from fibrin alone (CMF [-]) with those made from fibrin and CMF (CMF [+]) (Fig. 1A, B). The capillary volume in BBB-NET was quantified with IMARIS analysis of CD31-positive tubes. The CD31-positive capillary volume in the CMF (+) gel was significantly greater than that in the CMF (-) gel (Figure 1B, a-c), reaching 181.93 ± 16.3% that of the CMF (-) gel group. Since CD146 is an immature endothelial cell marker [32], we also compared the CD146-positive capillary volumes between the CMF (-) group and the CMF (+) group (Figure 1B, d-f). The CD146-positive capillary volume in the CMF (+) gel tended to be greater than that in the CMF (-) gel (122.9 ± 19.4% that of the CMF (-) gel), but the difference was not significant. As capillaries mature, the vascular basement membrane develops, which contributes to BBB integrity. The vascular basement membrane forms 3D protein network predominantly composed of collagen IV, laminin, nidogen, and heparan sulfate proteoglycans, which support BBB integrity and interactions among BBB cells [30,33]. We therefore compared the development of basement membrane in the CMF (-) gels and CMF (+) gels in terms of collagen type IV (Col IV) expression levels (Figure 1B, g-h). The volume of Col IV-positive capillaries in the CMF (+) gel was markedly greater than that in the CMF (-) gel (758.8 ± 150.8% of that in the CMF (-) gel group). These results suggest that CMFs enhance autonomous brain capillary network formation in BBB-NET.
3.2. CMFs Supported Astrocyte (HASTRs/ci35) Survival
We found that there were more ACs in the CMF (+) hydrogel than in the CMF (-) hydrogel (Fig. 2A). To examine the effects of the CMFs in more detail, we compared the ACs in the CMF (-) BBB-NET with those in the CMF (+) BBB-NET by aquaporin 4 (AQP4) immunostaining (Fig. 2A, a-c) and cell-tracker staining (Fig. 2A, d, e). AQP4-positive ACs surrounded CD31-positive capillaries in the CMF (+) gel (Fig. 2A-c). We therefore investigated the effect of the CMFs on the number of ACs (Fig. 2B-a). We used Cell Tracker and DAPI for cell counting (Fig. 2B, b-d) because immunostaining for AQP4 or GFAP does not reveal the cell morphology due to the localization of these proteins. The number of astrocytes in the CMF (+) hydrogel group was significantly greater than that in the CMF (-) hydrogel group (Figure 2B-d; 174.1 ± 70.7% that of the CMF [-] group). These results showed that CMFs supported AC survival in BBB-NET.
3.3. Astrocytes Promote the Formation and Maturation of Capillary Networks
On the basis of the results described above, we speculated that an increase in the number of ACs would be advantageous for capillary network formation. We therefore generated BBB-NET with and without ACs (Fig. 3A). After quantifying the CD31-positive capillary volume with IMARIS software (Fig. 3B, c-e), it was revealed that the capillary volume significantly decreased when ACs were removed from the CMF (+) hydrogel (Figure 3B-e, 54.6 ± 16.7% that of the AC (+) group). We also examined the effects of ACs on capillary basement membrane formation by observing the expression of collagen IV, one of the major components of the basal membrane (Fig. 3B, f-h). The volume of collagen IV-positive capillaries was significantly decreased when the ACs were removed (Fig. 3B-h; 37.1 ± 33.9% that of the AC (+) group). In addition, the signal strengths of both CD31 and collagen IV themselves increased. These results indicate that the positive effects of CMFs are mediated by an increase in AC survival, which promotes capillary formation and maturation.
3.4. CMFs Enhanced Astrocyte (HASTR/ci35) Survival by Interacting with Integrin b1
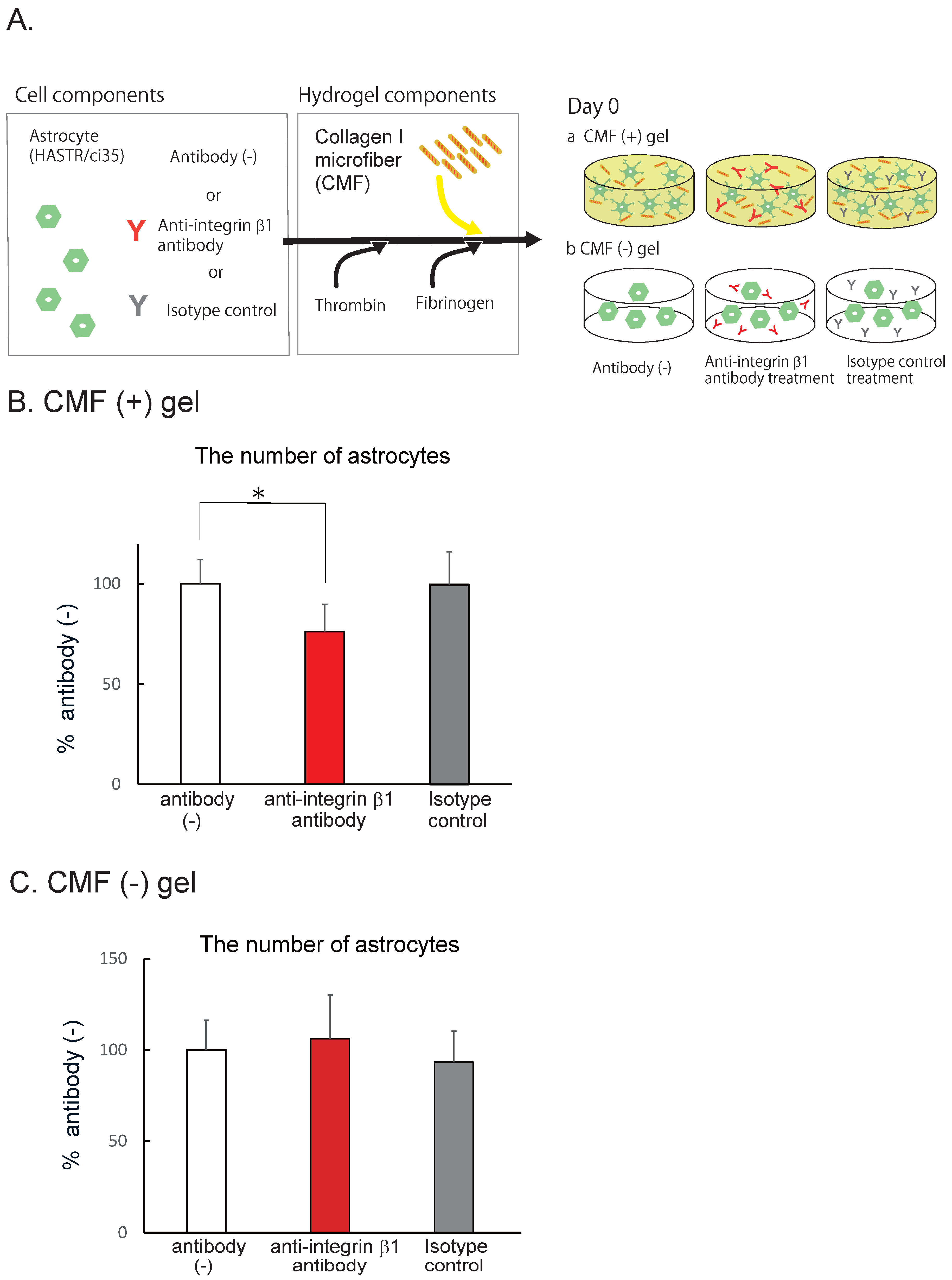
The CMFs used in this study are collagen I fragments with a size of 20 mm that crosslink with each other [26]. Collagen I has been reported to interact with several membrane proteins, such as integrins (α1β1, α2β1, α10β1, and α11β1), discoidin domain receptor (DDR) 1, DDR2, osteoclast-associated receptor (OSCAR), glycoprotein VI (GPVI), G6b-B, leukocyte-associated Ig-like receptor-1 (LAIR-1) of the leukocyte receptor complex (LRC), and the mannose family receptor uPARAP/Endo 180 [34]. Among these membrane proteins, the integrins and DDR1 are expressed in ACs [35], suggesting that integrins and/or DDR1 function as collagen receptors in our experiments. Integrins are active as heterodimers, and there are a wide variety of combinations of subunits [36]. When integrins function as collagen receptors, the b1 subunit is included [34]. We therefore examined the contribution of the CMF–integrin b1 interaction using an anti-integrin β1 antibody [29] or an isotype control (Fig. 4A). After 7 days of AC coculture with the antibody, we counted the number of cells. In the anti-integrin β1 antibody-treated group, the number of ACs was significantly lower than that in the antibody (-) group (Fig. 4B, 76.2 ± 13.8% that of the antibody (-) group), whereas the number of ACs in the isotype control-treated group was almost the same as that in the antibody (-) group (99.6 ± 16.4% that of the antibody (-) group). The BBB-NET hydrogel contains fibrinogen, which contains an Arg-Gly-Asp (RGD) sequence that also binds to the integrin β1 subunit. We therefore confirmed the extent to what the interaction between fibrin and integrin b1 contributed to AC survival using the fibrin hydrogels (Fig. 4C). In the hydrogel without CMFs, the anti-integrin b1 antibody did not affect the number of surviving cells. These data suggest that the CMFs interact with integrin b1 on ACs and promote their survival and that increasing the number of ACs further promotes the formation of capillary networks in BBB-NET.
4. Discussion
Our 3D-type BBB-MPS, BBB-NET uses the hydrogel which includes fibrin and CMFs fabricated from collagen I [26]. This combination has long been used in the field of cell engineering because of its reasonable cost [14,15,16]. In vivo, brain capillaries are characterized by their unique basement membrane (BM) structure, which is composed of extracellular matrix (ECM) components [37], such as laminin, collagen IV, fibronectin, and heparan sulfate proteoglycans [38]. In addition that collagen IV is one of the major components of the BM [33,37], collagen I has also been reported to be involved in the development of the CNS [38].
The CMFs used in this study were designed to be ≈20 mm, which is shorter than the cell body lengths of ECs and ACs [26]. This size enables the CMFs to have a sedimentation velocity similar to that of these cells because of their size ranges, thereby achieving homogeneous cell distribution in BBB-NET. Our study revealed that CMFs promoted AC survival, thereby increasing capillary network formation. The data also suggest that the interaction between ECs and ACs is important for capillary self-organization. Furthermore, the increased number of ACs also upregulated the expression of CD31 and collagen IV. Originally, collagen I and fibrin were used as angiogenic polymers in tissue engineering [39] and CMFs have been reported to enhance the survival of human umbilical vein endothelial cells (HUVEC) [40]. Our data suggest that in the presence of astrocytes, CMFs further enhance capillary maturation and capillary formation through the indirect effects from astrocytes. Astrocytes release many kinds of factors used in the interaction with EC, and the identification is our next step to work on.
Figure 5.
Our hypothesized mechanism by which CMFs promote capillary formation and maturation in BBB-NET. CMFs increased the number of surviving ACs by interacting with integrin b1, and the increased number of ACs supported capillary formation and maturation.
Figure 5.
Our hypothesized mechanism by which CMFs promote capillary formation and maturation in BBB-NET. CMFs increased the number of surviving ACs by interacting with integrin b1, and the increased number of ACs supported capillary formation and maturation.
In mammals, integrin heterodimers are composed of α and β subunits that form a noncovalent complex. To date, 18 α subunits and 8 β subunits have been identified, and 24 functionally distinct heterodimeric transmembrane receptors with different combinations of a and b subunits have been reported [34,36]. Among the integrin receptors, α1β1, α2β1, α10β1, and α11β1 bind to collagen I [36,41], and some amino acid sequences that are necessary for binding have been identified [such as Gly-Phe-Hyp-Gly-Glu-Arg (GFOGER)] [42]. Our data indicated that the effects of the CMFs on astrocytes are partly mediated through interactions with integrin receptors, including integrin b1. The CMFs have protective effects on AC survival through binding integrin receptors, including b1. Integrins transactivate growth factor receptors even in the absence of growth factor ligands and subsequently activate intracellular signal transduction cascades [43]. By binding to collagen or fibronectin, integrin β1 has also been reported to induce tyrosine phosphorylation on growth factor receptors, such as the EGF receptor, PDGFRβ, and vascular endothelial growth factor (VEGF) receptor-3, in the absence of their respective ligands [29,44]. On the other hand, ACs have been reported to support angiogenesis through integrin β1 [45]. These previous studies support our hypothesis that CMFs promote the survival of ACs via transactivation of cell surface growth factor receptors through the interaction with integrin b1, thereby promoting BBB-NET capillary network formation and maturation. Although the effects of the functional blocking antibody were significant, they remained at 76.2 ± 13.8% that of the control, suggesting that other mechanisms also contribute to the effects of CMFs. As described above, the collagen-binding integrins are α1β1, α2β1, α10β1, and α11β1. Further experiments are necessary to identify which α subunits are involved in the effects of the CMFs. In addition to integrins, DDR1 is another CMF candidate receptor. It has been reported that DDR1 interacts with collagen I [28] and that DDR1 is expressed in astrocytes [35,46]; however, little information is available concerning the role of DDR1 in angiogenesis. It is therefore necessary to confirm whether or not the effects of the CMFs are mediated through the mechanisms described above in our next study.
5. Conclusions
In BBB-NET, the CMFs in the hydrogel are advantageous for brain capillary network formation and maturation. The CMFs increased the number of surviving ACs by interacting with integrin b1, and the increased number of ACs supported capillary formation and maturation (Fig. 5). Our data suggest that to produce a stable BBB-NET, quality control (QC) of CMFs is quite important.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Representative Z-stack images of CD31 (a, b) and the Imaris surface (c, d) in hydrogels with (b, d) and without (a, c) CMFs
Funding
This research was supported by AMED (23be1004101j0102, 24be1004101j0103, 23mk0101222j0102, and 24mk0101222j0103).
Acknowledgments
This research was supported by AMED (23be1004101j0102, 24be1004101j0103, 23mk0101222j0102, and 24mk0101222j0103).
Abbreviations
| BBB | blood-brain barrier; |
| MPS | microphysiological system; |
| CNS | central nervous system; |
| PKPD | pharmacokinetics/pharmacodynamics; |
| TKTD | toxicokinetics/toxicodynamics; |
| CoU | context of use; |
| TfR | transferrin receptor; |
| CMF | collagen microfiber; |
| QC | quality control; |
| EC | endothelial cell; |
| AC | astrocyte; |
| PC | pericyte; |
| BM | basement membrane; |
| ECM | extracellular matrix; |
| SOP | standard operating procedure |
References
- Pardridge, W.M. The blood-brain barrier: bottleneck in brain drug development. NeuroRx. 2005, 2, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Xu, C.; Sun, P.; Wu, J.; Yan, C.; Hu, M.; Yan, N. Crystal structure of the human glucose transporter GLUT1. Nature. 2014, 510, 121–125. [Google Scholar] [CrossRef]
- McAllister, M.S.; Krizanac-Bengez, L.; Macchia, F.; Naftalin, R.J.; Pedley, K.C.; Mayberg, M.R.; Marroni, M.; Leaman, S.; A Stanness, K.; Janigro, D. Mechanisms of glucose transport at the blood-brain barrier: an in vitro study. Brain Res. 2001, 904, 20–30. [Google Scholar] [CrossRef]
- Booth, R.; Kim, H. Characterization of a microfluidic in vitro model of the blood-brain barrier (μBBB). Lab. Chip. 2012, 12, 1784–1792. [Google Scholar] [CrossRef]
- Bicker, J.; Alves, G.; Fortuna, A.; Falcão, A. Blood-brain barrier models and their relevance for a successful development of CNS drug delivery systems: a review. Eur. J. Pharm. Biopharm. 2014, 87, 409–432. [Google Scholar] [CrossRef]
- Sato, K. Consideration for future in vitro BBB models - technical development to investigate the drug delivery to the CNS. Nihon Yakurigaku Zasshi. 2018, 152, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Ohtsuki, S.; Katsukura, Y.; Ikeda, C.; Suzuki, T.; Kamiie, J.; Terasaki, T. Quantitative targeted absolute proteomics of human blood-brain barrier transporters and receptors. J. Neurochem. 2011, 117, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Warren, M.S.; Zerangue, N.; Woodford, K.; Roberts, L.M.; Tate, E.H.; Feng, B.; Li, C.; Feuerstein, T.J.; Gibbs, J.; Smith, B.; et al. Comparative gene expression profiles of ABC transporters in brain microvessel endothelial cells and brain in five species including human. Pharmacol. Res. 2009, 59, 404–413. [Google Scholar] [CrossRef]
- Potjewyd, G.; Kellett, K.A.B.; Hooper, N.M. 3D hydrogel models of the neurovascular unit to investigate blood-brain barrier dysfunction. Neuronal Signal 2021, 5, Ns20210027. [Google Scholar] [CrossRef]
- Bhalerao, A.; Sivandzade, F.; Archie, S.R.; Chowdhury, E.A.; Noorani, B.; Cucullo, L. In vitro modeling of the neurovascular unit: advances in the field. Fluids Barriers CNS 2020, 17, 22. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Hui, H.; Cai, T.; Huang, H.; Yang, F.; Feng, J.; Zhang, J.; Yan, X. CD146 coordinates brain endothelial cell-pericyte communication for blood-brain barrier development. Proc. Natl. Acad. Sci. U S A 2017, 114, E7622–e31. [Google Scholar] [CrossRef] [PubMed]
- Agathe, F.; Yasuhiro, N.; Yukari, S.M.; Tomomi, F.; Kaoru, S.; Matsusaki, M. An in vitro self-organized three-dimensional model of the blood-brain barrier microvasculature. Biomed. Mater. 2020, 16, 015006. [Google Scholar] [CrossRef]
- Piantino, M.; Kang, D.H.; Furihata, T.; Nakatani, N.; Kitamura, K.; Shigemoto-Mogami, Y.; Sato, K.; Matsusaki, M. Development of a three-dimensional blood-brain barrier network with opening capillary structures for drug transport screening assays. Mater. Today Bio. 2022, 15, 100324. [Google Scholar] [CrossRef]
- Rao, R.R.; Ceccarelli, J.; Vigen, M.L.; Gudur, M.; Singh, R.; Deng, C.X.; Putnam, A.J.; Stegemann, J.P. Effects of hydroxyapatite on endothelial network formation in collagen/fibrin composite hydrogels in vitro and in vivo. Acta Biomater. 2014, 10, 3091–3097. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.R.; Peterson, A.W.; Ceccarelli, J.; Putnam, A.J.; Stegemann, J.P. Matrix composition regulates three-dimensional network formation by endothelial cells and mesenchymal stem cells in collagen/fibrin materials. Angiogenesis 2012, 15, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Seyedhassantehrani, N.; Li, Y.; Yao, L. Dynamic behaviors of astrocytes in chemically modified fibrin and collagen hydrogels. Integr. Biol. (Camb). 2016, 8, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Critser, P.J.; Kreger, S.T.; Voytik-Harbin, S.L.; Yoder, M.C. Collagen matrix physical properties modulate endothelial colony forming cell-derived vessels in vivo. Microvasc. Res. 2010, 80, 23–30. [Google Scholar] [CrossRef]
- Shang, Y.; Piantino, M.; Zeng, J.; Louis, F.; Xie, Z.; Furihata, T.; Matsusaki, M. Control of blood capillary networks and holes in blood-brain barrier models by regulating elastic modulus of scaffolds. Mater. Today Bio. 2023, 21, 100714. [Google Scholar] [CrossRef]
- Kornev, V.A.; Grebenik, E.A.; Solovieva, A.B.; Dmitriev, R.I.; Timashev, P.S. Hydrogel-assisted neuroregeneration approaches towards brain injury therapy: A state-of-the-art review. Comput. Struct. Biotechnol. J. 2018, 16, 488–502. [Google Scholar] [CrossRef]
- Wevers, N.R.; Kasi, D.G.; Gray, T.; Wilschut, K.J.; Smith, B.; van Vught, R.; Shimizu, F.; Sano, Y.; Kanda, T.; Marsh, G.; et al. A perfused human blood-brain barrier on-a-chip for high-throughput assessment of barrier function and antibody transport. Fluids Barriers CNS 2018, 15, 23. [Google Scholar] [CrossRef]
- Kamiichi, A.; Furihata, T.; Kishida, S.; Ohta, Y.; Saito, K.; Kawamatsu, S.; Chiba, K. Establishment of a new conditionally immortalized cell line from human brain microvascular endothelial cells: a promising tool for human blood-brain barrier studies. Brain Res. 2012, 1488, 113–22. [Google Scholar] [CrossRef] [PubMed]
- Furihata, T.; Kawamatsu, S.; Ito, R.; Saito, K.; Suzuki, S.; Kishida, S.; Saito, Y.; Kamiichi, A.; Chiba, K. Hydrocortisone enhances the barrier properties of HBMEC/ciβ, a brain microvascular endothelial cell line, through mesenchymal-to-endothelial transition-like effects. Fluids Barriers CNS 2015, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Umehara, K.; Sun, Y.; Hiura, S.; Hamada, K.; Itoh, M.; Kitamura, K.; Oshima, M.; Iwama, A.; Saito, K.; Anzai, N.; et al. A New Conditionally Immortalized Human Fetal Brain Pericyte Cell Line: Establishment and Functional Characterization as a Promising Tool for Human Brain Pericyte Studies. Mol. Neurobiol. 2018, 55, 5993–6006. [Google Scholar] [CrossRef] [PubMed]
- Furihata, T.; Ito, R.; Kamiichi, A.; Saito, K.; Chiba, K. Establishment and characterization of a new conditionally immortalized human astrocyte cell line. J. Neurochem. 2016, 136, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Umehara, K.; Suzuki, S.; Kitamura, K.; Nunoya, K.I.; Yamaura, Y.; Imawaka, H.; Izumi, S.; Wakayama, N.; Komori, T.; et al. A Human Immortalized Cell-Based Blood-brain Barrier Triculture Model: Development and Characterization as a Promising Tool for Drug-Brain Permeability Studies. Mol. Pharm. 2019, 16, 4461–4471. [Google Scholar] [CrossRef]
- Naka, Y.; Kitano, S.; Irie, S.; Matsusaki, M. Wholly vascularized millimeter-sized engineered tissues by cell-sized microscaffolds. Mater. Today Bio. 2020, 6, 100054. [Google Scholar] [CrossRef]
- Kanamoto, T.; Hikida, M.; Sato, S.; Oyama, S.; Tachi, Y.; Kuroda, S.; Mazuka, T.; Ebina, K.; Nakai, T.; Nakata, K. Integrin α2β1 plays an important role in the interaction between human articular cartilage-derived chondrocytes and atelocollagen gel. Sci. Rep. 2021, 11, 1757. [Google Scholar] [CrossRef]
- Xu, H.; Bihan, D.; Chang, F.; Huang, P.H.; Farndale, R.W.; Leitinger, B. Discoidin domain receptors promote α1β1- and α2β1-integrin mediated cell adhesion to collagen by enhancing integrin activation. PLoS ONE 2012, 7, e52209. [Google Scholar] [CrossRef]
- Wang, D.; Duan, H.; Feng, J.; Xiang, J.; Feng, L.; Liu, D.; Chen, X.; Jing, L.; Liu, Z.; Zhang, D.; et al. Soluble CD146, a cerebrospinal fluid marker for neuroinflammation, promotes blood-brain barrier dysfunction. Theranostics 2020, 10, 231–246. [Google Scholar] [CrossRef]
- Shigemoto-Mogami, Y.; Nakayama-Kitamura, K.; Sato, K. The arrangements of the microvasculature and surrounding glial cells are linked to blood-brain barrier formation in the cerebral cortex. Front. Neuroanat. 2024, 18, 1438190. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, D. Three-dimensional remodeling of functional cerebrovascular architecture and gliovascular unit in leptin receptor-deficient mice. J. Cereb. Blood Flow. Metab. 2021, 41, 1547–1562. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Luo, Y.; Hui, H.; Cai, T.; Huang, H.; Yang, F.; Feng, J.; Zhang, J.; Yan, X. CD146 coordinates brain endothelial cell-pericyte communication for blood-brain barrier development. Proc. Natl. Acad. Sci. USA 2017, 114, E7622–e31. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.S.; Routhe, L.J.; Moos, T. The vascular basement membrane in the healthy and pathological brain. Journal of cerebral blood flow and metabolism: official journal of the International Society of Cerebral Blood Flow Metab. 2017, 37, 3300–3317. [Google Scholar] [CrossRef]
- Elango, J.; Hou, C.; Bao, B.; Wang, S.; Maté Sánchez de Val, J.E.; Wenhui, W. The Molecular Interaction of Collagen with Cell Receptors for Biological Function. Polymers 2022, 14, 876. [Google Scholar] [CrossRef] [PubMed]
- Vilella, E.; Gas, C.; Garcia-Ruiz, B.; Rivera, F.J. Expression of DDR1 in the CNS and in myelinating oligodendrocytes. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118483. [Google Scholar] [CrossRef]
- Pang, X.; He, X.; Qiu, Z.; Zhang, H.; Xie, R.; Liu, Z.; Gu, Y.; Zhao, N.; Xiang, Q.; Cui, Y. Targeting integrin pathways: mechanisms and advances in therapy. Signal Transduct. Target. Ther. 2023, 8, 1. [Google Scholar] [CrossRef]
- Soles, A.; Selimovic, A.; Sbrocco, K.; Ghannoum, F.; Hamel, K.; Moncada, E.L.; Gilliat, S.; Cvetanovic, M. Extracellular Matrix Regulation in Physiology and in Brain Disease. Int. J. Mol. Sci. 2023, 24, 7049. [Google Scholar] [CrossRef]
- Lauren, K.; Wareham, Robert O. Lauren K. Wareham, Robert O. Baratta, Brian J. Del Buono, Eric Schlumpf & David J. Calkins. Molecular Neurodegeneration 2024, 19, 11. [Google Scholar]
- Twardowski, T.; Fertala, A.; Orgel, J.P.; San Antonio, J.D. Type I collagen and collagen mimetics as angiogenesis promoting superpolymers. Curr. Pharm. Des. 2007, 13, 3608–3621. [Google Scholar] [CrossRef]
- Liu, H.; Kitano, S.; Irie, S.; Levato, R.; Matsusaki, M. Collagen Microfibers Induce Blood Capillary Orientation and Open Vascular Lumen. Adv. Biosyst. 2020, 4, e2000038. [Google Scholar] [CrossRef]
- Mezu-Ndubuisi, O.J.; Maheshwari, A. The role of integrins in inflammation and angiogenesis. Pediatr. Res. 2021, 89, 1619–1626. [Google Scholar] [CrossRef] [PubMed]
- Malcor, J.D.; Mallein-Gerin, F. Biomaterial functionalization with triple-helical peptides for tissue engineering. Acta Biomater. 2022, 148, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Kitazato, K.; Basaki, Y. Integrin-linked kinase regulates vascular morphogenesis induced by vascular endothelial growth factor. J. Cell Sci. 2004, 117 Pt 3, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Moro, L.; Venturino, M.; Bozzo, C.; Silengo, L.; Altruda, F.; Beguinot, L.; Tarone, G.; Defilippi, P. Integrins induce activation of EGF receptor: role in MAP kinase induction and adhesion-dependent cell survival. Embo J. 1998, 17, 6622–6632. [Google Scholar] [CrossRef] [PubMed]
- Tabata, H.; Sasaki, M.; Agetsuma, M.; Sano, H.; Hirota, Y.; Miyajima, M.; Hayashi, K.; Honda, T.; Nishikawa, M.; Inaguma, Y.; et al. Erratic and blood vessel-guided migration of astrocyte progenitors in the cerebral cortex. Nat. Commun. 2022, 13, 6571. [Google Scholar] [CrossRef]
- Itoh, Y. Discoidin domain receptors: Microenvironment sensors that promote cellular migration and invasion. Cell Adh. Migr. 2018, 12, 378–385. [Google Scholar] [CrossRef]
Figure 1.
CMFs are important for the autonomous production of capillary networks. A. Schematic of the BBB-NET components. B. Representative z-stack images of the endothelial cell markers CD31 (a, b) and CD146 (d, e) and the basement membrane protein collagen IV (g, h) in hydrogels with (b, e, h) and without (a, d, g) CMFs. Scale bar: 100 mm. Comparison of the capillary volumes positive for CD31 (c), CD146 (f), and collagen IV (i) among hydrogels with and without CMFs. The percentage of positive staining in the presence of CMFs is shown, and without CMFs was taken as 100%. All data are expressed as the means ± standard errors of the mean (SEMs). Each representation shown is from three independent experiments, each with four images; Student’s t test, *: p < 0.05.
Figure 1.
CMFs are important for the autonomous production of capillary networks. A. Schematic of the BBB-NET components. B. Representative z-stack images of the endothelial cell markers CD31 (a, b) and CD146 (d, e) and the basement membrane protein collagen IV (g, h) in hydrogels with (b, e, h) and without (a, d, g) CMFs. Scale bar: 100 mm. Comparison of the capillary volumes positive for CD31 (c), CD146 (f), and collagen IV (i) among hydrogels with and without CMFs. The percentage of positive staining in the presence of CMFs is shown, and without CMFs was taken as 100%. All data are expressed as the means ± standard errors of the mean (SEMs). Each representation shown is from three independent experiments, each with four images; Student’s t test, *: p < 0.05.
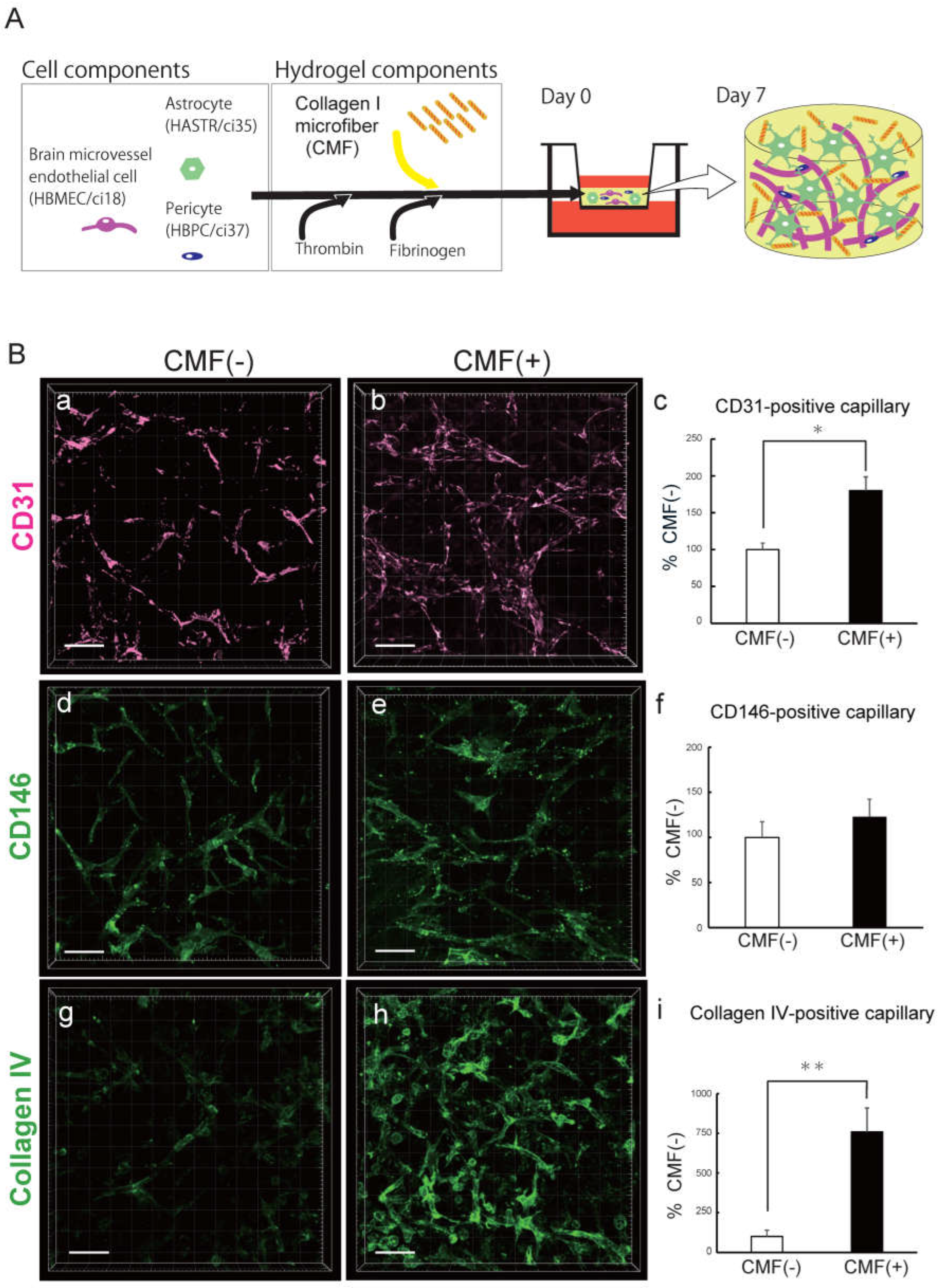
Figure 2.
CMFs support astrocyte survival. A. Representative z-stack images of the astrocyte marker Aquaporin 4 (a, b, c) and cell tracker-labelled astrocytes (d, e) in hydrogels with (b, c, e) and without (a, d) CMFs. Scale bar in a, b, d, e: 100 mm; scale bar in c: 50 mm. B. Schematic of the astrocyte-only gel (a). Representative z-stack images of cell tracker-labelled astrocytes in hydrogels with (c) and without (b) CMFs. Comparison of the number of astrocytes in hydrogels with and without CMFs (d). Each representation shown is from three independent experiments, each with four images; Student’s t test, *: p < 0.05.
Figure 2.
CMFs support astrocyte survival. A. Representative z-stack images of the astrocyte marker Aquaporin 4 (a, b, c) and cell tracker-labelled astrocytes (d, e) in hydrogels with (b, c, e) and without (a, d) CMFs. Scale bar in a, b, d, e: 100 mm; scale bar in c: 50 mm. B. Schematic of the astrocyte-only gel (a). Representative z-stack images of cell tracker-labelled astrocytes in hydrogels with (c) and without (b) CMFs. Comparison of the number of astrocytes in hydrogels with and without CMFs (d). Each representation shown is from three independent experiments, each with four images; Student’s t test, *: p < 0.05.
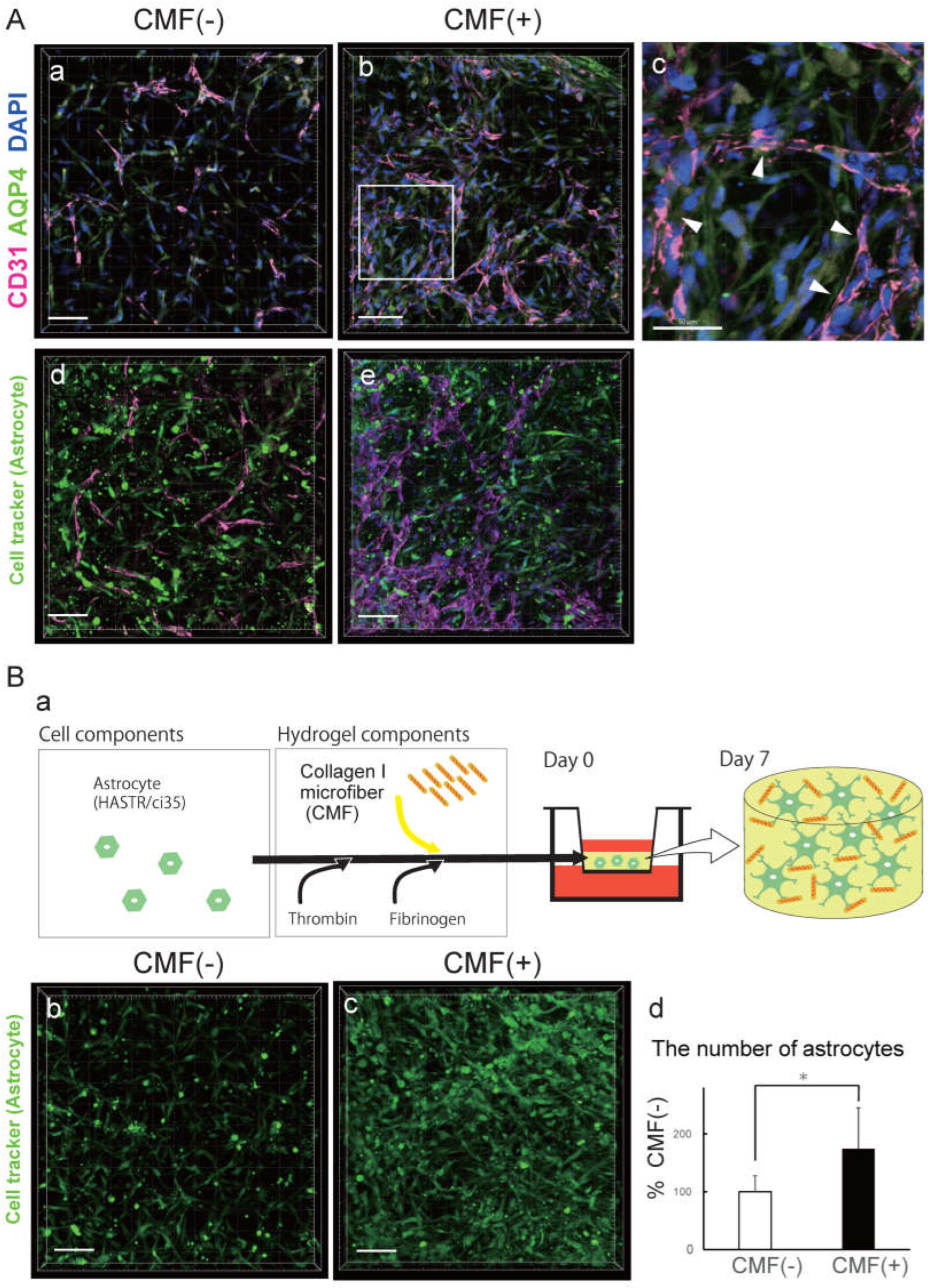
Figure 3.
Astrocytes promote the formation and maturation of capillary networks. A. Schematic of hydrogels with CMFs and with (a) and without (b) ACs. B. Representative z-stack images of CD31 (c, d) and collagen IV (f, g) in the presence (c, f) and absence (d, g) of astrocytes in the hydrogel with CMFs. Scale bar: 100 mm. Comparison of CD31-positive capillary volume (e) and collagen IV-positive blood vessels (h) in the absence and presence of astrocytes in the hydrogel with CMFs. Each representation shown is from three independent experiments, each with four images. Student’s t test, *: p < 0.05 **: p < 0.01.
Figure 3.
Astrocytes promote the formation and maturation of capillary networks. A. Schematic of hydrogels with CMFs and with (a) and without (b) ACs. B. Representative z-stack images of CD31 (c, d) and collagen IV (f, g) in the presence (c, f) and absence (d, g) of astrocytes in the hydrogel with CMFs. Scale bar: 100 mm. Comparison of CD31-positive capillary volume (e) and collagen IV-positive blood vessels (h) in the absence and presence of astrocytes in the hydrogel with CMFs. Each representation shown is from three independent experiments, each with four images. Student’s t test, *: p < 0.05 **: p < 0.01.
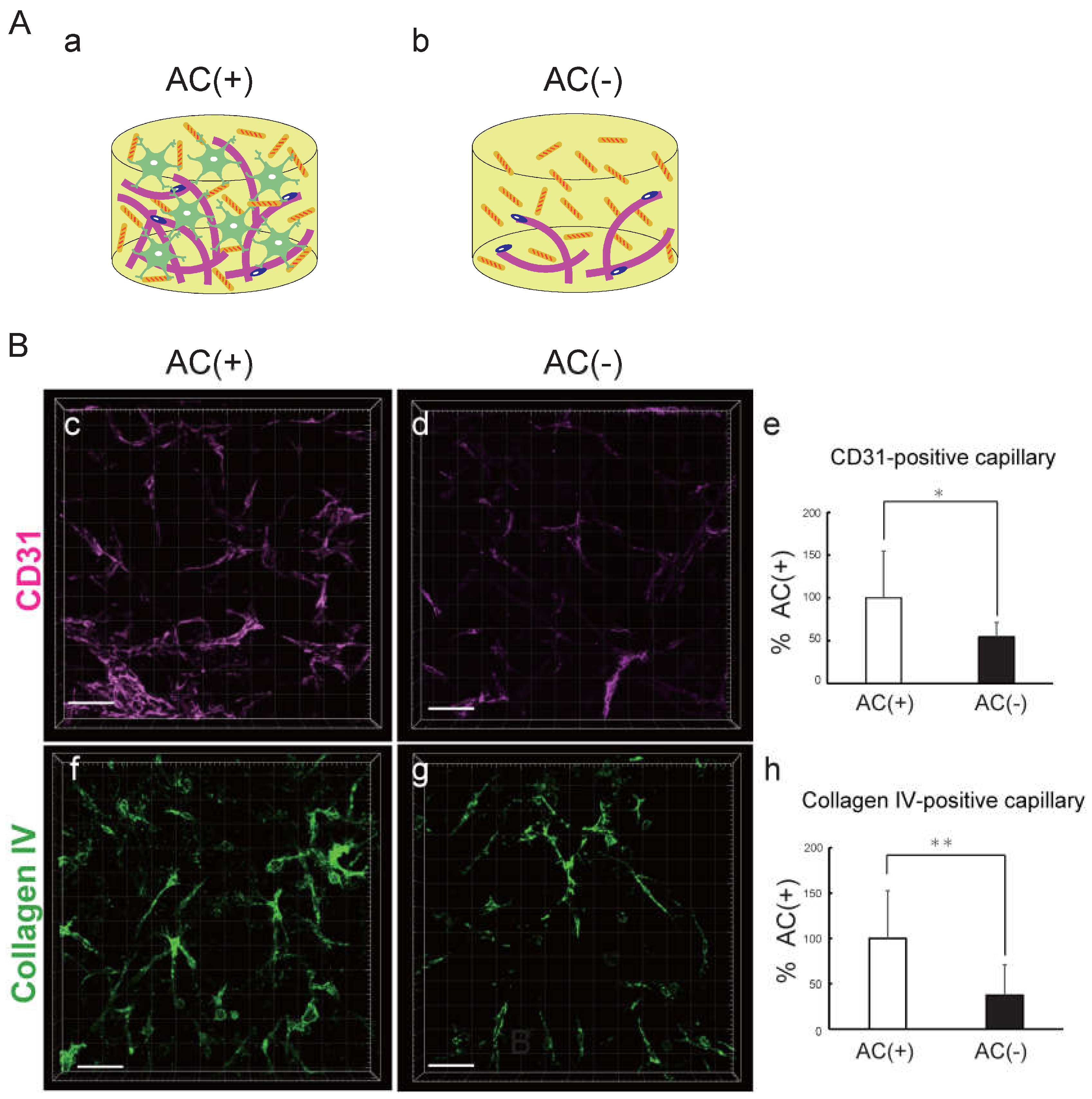
Figure 4.
CMFs interact with astrocytes via the integrin β1 subunit. A Schematic of AC pretreated with and without each antibody in hydrogels with (a) and without CMF(b). B. Comparison of the number of astrocytes after pretreatment with an integrin β1 antibody or an isotype control in hydrogels with CMFs. Each representation shown is from three independent experiments, each with four images. Student’s t test, *: p < 0.05. C. Comparison of the number of astrocytes after pretreatment with an integrin β1 antibody or an isotype control in hydrogels without CMFs. Each representation shown is from two independent experiments, each with four images.
Figure 4.
CMFs interact with astrocytes via the integrin β1 subunit. A Schematic of AC pretreated with and without each antibody in hydrogels with (a) and without CMF(b). B. Comparison of the number of astrocytes after pretreatment with an integrin β1 antibody or an isotype control in hydrogels with CMFs. Each representation shown is from three independent experiments, each with four images. Student’s t test, *: p < 0.05. C. Comparison of the number of astrocytes after pretreatment with an integrin β1 antibody or an isotype control in hydrogels without CMFs. Each representation shown is from two independent experiments, each with four images.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



