
Submitted:
05 July 2025
Posted:
07 July 2025
You are already at the latest version
Abstract
In recent decades, the growing interest in fertility rate improvements in livestock species, considering their high economic value in terms of production, has prompted the devel-opment of new methodological approaches associated with the use of male germline stem cells. The significant potential of spermatogonial stem cells (SSCs) to self-renew and dif-ferentiate into mature spermatozoa makes them potentially useful for transplantation into testicular tissue, as well as for new biotechnological methodologies. More recently, canine SSCs have also been investigated, since dogs are considered an optimal animal model of human reproduction, development, and disease. Characterization studies of canine SSCs (cSSCs) have revealed interesting aspects in relation to their potential clinical application in reproduction. Against this background, this review provides an update on the main aspects of cSSC biology, focusing also on the genetic regulators of self-renewal and differ-entiation processes and different isolation methods. Finally, novel views on the potential clinical transplantation of canine SSCs into recipient testicular tissue are also discussed.
Keywords:
spermatogonial stem cells (SSCs)
; canine testis
; dog
; reproduction
; self-renewal
; differentiation
; cryptorchidism
1. Introduction
In recent decades, the focus of scientific research in the area of fertility rate improvement has been on livestock species, in line with their high economic value in terms of production. In fact, the application of already developed reproductive biotechnologies (artificial insemination, embryo transfer, etc.) to shorten the generation interval and improve reproductive performances has been crucial to dairy cattle breeding. However, these biotechnologies have been shown to have some limitations, including a relatively slow rate of genetic progress. In this context, more recently, the development of new methodological approaches associated with the use of male germline stem cells has been rapid, opening numerous possibilities in this field. In particular, spermatogonial stem cells (SSCs), which are not only adult stem cells that are responsible for the spermatogenesis process but capable of converting into pluripotent stem cells, have been studied extensively for the new opened opportunities that they have created in the field of regenerative medicine. SSCs are a population of resident stem cells located at the basal membrane of seminiferous tubules of the testis and play a role in normal spermatogenesis. In fact, their main activities, including self-renewal, and the balance between differentiated cells and the reserve of stem cells in the tissue are controlled by the testis microenvironment, which produces specific factors and signals [1,2,3].
Compared with livestock species, for whom more innovative aspects of SSCs relating to their prolonged cultivation in vitro and the possibility of their transplantation into testicular tissue for regenerative purposes have been elucidated, the research on companion animals is more scarce. It is worth noting that companion animals, such as canine species, could be considered an optimal animal model for experimental and clinical testing for translation to human models. The very recent literature reports several findings demonstrating the importance of using alternative species to laboratory animals [4]. Moreover, since the sequencing of the canine genome [5], and in light of the evidence that canine genetic disorders are similar to those of humans, the scientific interest in this species and the knowledge of its biological characteristics have increased.
Recently, the exploration of innovative methods for the culturing SSCs, as well as the influence of the testis microenvironment and several other factors influencing spermatogenesis, has emphasized the potential of these cells for therapeutic applications. In particular, SSCs could be a biologic tool (complementing the availability of other advanced assisted reproductive technologies and various genome editing tools) for the therapy of subfertility/infertile diseases or diseases associated with oncology.
The aim of this review is to highlight some recent advances in the characterization of canine SSCs (cSSCs), focusing on their experimental and potential clinical applications in reproduction. The interest in this topic originates from our research focusing on aquaporin characterization at the level of the male reproductive tract of the dog [6,7]. Our results indicating a different mechanism of modulation in aquaporin (AQP) expression according to a physiological/pathophysiological condition (cryptorchidism) suggest the potential value of a better understanding of the canine reproductive tract, specifically the testis tubule microenvironment [6,7]. Such studies on different approaches to the use of SSCs could be important for functional studies and methodologies for restoring fertility. As was also recently reported, the proper function of SSCs and their niche is an important prerequisite for successful spermatogenesis [8].
2. Spermatogonial Stem Cells (SSCs): Characteristics and Main Roles
The process of continuous spermatogenesis is fundamental for male fertility and occurs in the testis. It is dependent on the differentiation of a group of adult stem cells called SSCs, which represent a subgroup of undifferentiated spermatogonia (uSPG). Spermatogenesis is an intricate and cyclic process, controlled by hormones and cell signalling mechanisms. SSCs reside in a niche at the basal membrane of seminiferous tubules of the testis and are found in a limited number in relation to the total cellular population (calculated as 0.01-0.02% in mouse) [9]. SSCs are characterized by an absence of heterochromatin in the nuclei, which allows them to be distinguished from differentiating SPG. Their main role is to maintain the balance between the self-renewal and differentiation processes during spermatogenesis. In fact, a serious consequence of the dysfunction of these mechanisms is the Sertoli cell-only syndrome (SCOS) due to SSC exhaustion [10].
The origin of SSCs has primarily been investigated in mice, leading to different theories. The latest of these, the “Fragmentation model”, demonstrates how undifferentiated spermatogonia continuously interconvert between spermatogonia, As (presenting as a single cell), and short syncytial states via fragmentation called spermatogonia, Apr [11,12].
The characterization of SSCs demonstrated that the functional mechanisms that are regulated in party by these cells are quite complex. In fact, the niche in which SSCs reside constitute the microenvironment (mainly comprising Sertoli cells, Leydig cells, peritubular myoid cells (PMCs), macrophages, and the ECM) [13,14,15] and regulates the plasticity and fate of the cells influencing the functions of SCs, including homing, self-renewal, and differentiation (Figure 1).
The cellular elements of the SSC niche, as well other factors (i.e. ageing, hormones), play a particular role in the maintenance of SSCs. In fact, in mice, a higher capability to generate spermatogenic colonies in pup testes compared with that in adult testes has been demonstrated. This result suggests that the microenvironment of the pup testis is different to that of an adult testis, providing a more hospitable environment for the transplantation of male germ line stem cells [16]. However, it is worth noting the possible influence of SSCs on their niche, as demonstrated by results revealing the ability of niche cells to respond to bovine SSC removal by promoting the upregulation of glial cell line-derived neurotrophic factor (GDNF) and fibroblast growth factor 2 (FGF2) to repopulate the niche with germ cells [17].
In this regard, the role of GDNF and FGF2, which are both produced by Sertoli cells, in promoting the self-renewal and expansion of SSCs (in vitro and in vivo) is well known (Figure 1) [18,19,20,21,22,23].
Although several studies have focused on GDNF’s role by means of investigations in animal models, as well as in different animal species, further insights regarding its spatiotemporal regulation are required. In addition, FGF is recognized for being involved in spermatogonial differentiation, acting in combination with retinoic acid (RA) at the level of the germline niche to favour the differentiation process (Table 1).
This demonstrates a slightly different role to that of GDNF, also suggesting the existence of GDNF- and FGF2-dominant niches [44]. These studies, which are also important for the setting of the optimal conditions for SSC expansion in vitro, demonstrate that GDNF (40 ng/ml) induces an adequate proliferation of ovine SSCs and is able to maintain their stemness for up to 30 days [45]. In addition, GNDF together with basic FGF (bFGF) and GFRα1 is the best combination of factors for preserving the stem potential of SSCs and guaranteeing their durability for a long period, as shown in a culture of pig SSCs (Table 1) [27]. Other studies have demonstrated that the loss of SSCs is a direct consequence of GDNF inhibition, as well as its related signalling mechanisms, indicating the significative role of this growth factor [25]. Other paracrine factors produced by Sertoli cells—including Insulin-like growth factor 1 (IGF1), Insulin growth factor binding protein 7 (IGFBP7), the Na+-K+-Cl transporter isoform 1 (NKCC1), protein tyrosine phosphatase, and non-receptor type 11 (PTPN11, also known as SHP2)—play important roles in the proliferation of human and mouse SSCs (Table 1) [28].
However, studies on SSC characterization also include the analysis of stemness markers, which are similar to those that are found on the surface of stem cells from different tissues (bone marrow, adipose tissue, etc.). It is worth noting that CD9, which is commonly expressed on other stem cells, has been discovered on the surface of rabbit and mouse SSCs, suggesting its potential association with integrins, including b1- and a6-integrin [47]. With the same goal, several other markers (GPR125, GFR1, THY1, ZBTB16, SSEA-4, and PLZF) have been identified on the surface of rodent and human SSCs, albeit some of them (THY1) have also been found on somatic cells [48,49,50,51,52]. In particular, THY1, a glycosylphosphatidylinositol-anchored glycoprotein of the Ig superfamily, is positively expressed in mouse SSCs. The THY1 gene codes thymocyte antigen, which can be used as a marker for a variety of stem cells, and it has been used as an SSC function marker [53,54].
More recently, the CD2 expression in rat SSCs, as well as that previously identified in mouse SSCs [55], led to the hypothesis that this expression may be conserved in the SSCs of other animal species. In addition, a new marker named forkhead box protein C2 (FOXC2), with a role in the maintenance of the quiescent state of primitive SSCs, has also been identified in an SSC subpopulation in adult mice and humans [56].
However, the discovery of stemness markers has been unsuccessful due to several factors (low surface antigen recognition, the inefficiency of spermatogonial transplantation, and a lack of long-term culture systems), which has increased the development of other methodologies requiring SSC enrichment.
Recently, the use of a transcriptomic approach demonstrated how RNA transport and the MAPK and p53 pathways play vital roles in early SSC differentiation, thus shedding light on the importance of these regulatory mechanisms as possible causes of male infertility [57]. In this regard, a recent study by Qingqing Geng (2025) demonstrated the impact of vitamin B6 on the miR-1458-TBX6 regulatory axis for SSC formation in Rugao Yellow Chicken [58].
In terms of the characterization of “stemness” markers for the individuation of SSCs within cultures of primary testicular cells, more attention has been given to other aspects, such as specific behaviours of stem cells. In this regard, the increase in colony numbers can be considered an indication of the presence of proliferative SSCs [59]. However, there is currently a lack of an unambiguously established SSC marker for human culture [60].
3. Spermatogonial Stem Cells from Domestic Animal Species: Isolation and In Vitro Expansion Techniques
In recent decades, isolation and enrichment protocols for isolating SSCs from testicular tissues of several domestic animal species have been explored. In fact, after the initial failure of SSC isolation (i.e., bovine, swine) [61] due to the application of methods like those employed for rodent species, more recently, several aspects have been discovered to be critical for more consistent success in these procedures [61]. In this regard, a fundamental prerequisite seems to be the improvement of isolation techniques facilitating the separation and enrichment of this rare population of cells from a wider group of testicular cells [62]. The best protocol, which is widely adopted for the isolation of SSCs, is the enzymatic digestion of the testicular tissue (the two–three step protocol) (Table 2), collected from domestic animals at a specific age of their development in order to obtain the maximum size of cellular population [63].
For ovine species, it was recently demonstrated that the best SSC isolation was achieved from prepubertal ram testes by using laminin in combination with bovine serum albumin (BSA) [64] (Table 2). This was probably similar to the observation that for goats at the prepubertal stage, the population of cells that can be isolated from a testis contains a high number of undifferentiated spermatogonia and few gonocytes and thus exhibits specific biochemical characteristics of SSCs [65].
Moreover, the choice of an appropriate medium with the right growth factors for the cell culture, as well as the appropriate methods for potentiating SSC renewal and differentiation, has emerged as important for prolonging the actual maximum time of in vitro SSC culture (no longer than 2 months for the majority of animal species) (Table 2) [66,67,68,69,70].
In this regard, it has been widely demonstrated that GDNF is a principal growth factor that can promote the in vitro proliferation of porcine SSCs [71], also suggesting its pivotal role across different species [72,73] (Table 2). Additional factors, including FGF2, IGF1, and LIF, help porcine SSC proliferation, guaranteeing their survival for more than 25 days with a particular cellular morphology and the formation of grape-like colonies (Table 2) [71].
In this regard, experiments on neonatal and adult Swiss albino mice demonstrated a more prolonged duration of culture for SSCs that were isolated from young testes compared with that in adult testes, with a concomitant expression of pluripotency markers (GFRA1, CD9, Nanog, Oct4, and Sox2) [74]. A similar behaviour was previously observed for bovine SSCs [75], suggesting that the animal’s age, and thus the testis development status, is crucial for the success of an SSC culture system. Recently, a hypothesis regarding this aspect was developed by Xiao-Yuan Zhang et al., who demonstrated differences in the transcript profiles of prepubertal buffalo (PUB) and adult buffalo (ADU) seminiferous tubules [76]. The evidence of a particular enrichment in genes relating to the development of SSCs in PUB compared with ADU suggests that the germ cells grow during this age, in parallel with the morphological development of the testes. This is also demonstrated by the simple structure of the seminiferous tubule in PUB, where the stage of the SSC niche is more established than that observed in ADU [76].
The literature on other species, including chicken and cat [70,77], is scarce, while for horse species, an interesting aspect of SSC renewal activity was observed after thawing cryopreserved SSCs [78] (Table 2).
Other factors, including hormones that can mediate the interaction between germ cells and Sertoli cells during spermatogenesis, are involved in the survival of germ cells [79]. Notably, it has been demonstrated that equine chorionic gonadotropin hormone (eCG), which is known for its FSH-similar activity, can influence SSC proliferation by increasing the number of colonies in relation to the number of control SSCs [80] (Table 3). In addition, studies focusing on prolonging SSCs’ survival in culture showed the efficacy of using melatonin supplementation (100 μM) to improve cell viability and colony formation, suggesting its pivotal role for mouse SSC development in vitro [81] (Table 3). The hormone that is added to the freezing medium can also protect frozen–thawed goat SSCs from cellular damage by activating their antioxidant defence system and reducing the freeze-induced excessive autophagy impairment [82].
More recently, it has been demonstrated that low concentrations of testosterone (60 μg/mL) significantly improve the colonization and viability of goat SSCs in a coculture with Sertoli cells, suggesting an important role of this hormone for improving SSC culture conditions and thus achieving future progress in reproductive technologies (Table 3) [83].
With the same objective, a study by A. Jafarnejad et al. (2018) [84] demonstrated the possible beneficial use of antioxidants (vitamin C and α-tocopherol analogue) to counteract the oxidative stress and apoptosis that are known to be the most common injuries to SSCs [85]. In detail, both products, used separately at an optimal dosage (50 µg/mL of vitamin C or 25 µg/mL of Trolox), improved the viability and colony formation of bovine SSCs after seven days of culture and reduced the levels of apoptosis by means of the regulator bax and anti-apoptotic bcl2 (Table 3) [84]. Recently, similar beneficial effects on SSC viability have also been demonstrated for ovine species (Table 3) [86].
4. Canine Spermatogonial Stem Cells: Characteristics and Regulatory Factors
Research on the spermatogenesis process in canine species, as well as the characterization of SSCs (cSSCs), has recently focused more on other animal species (especially rodents) and is continuously developing. It is worth noting that spermatogenesis in dogs begins at 7 months of age, and the complete cycle lasts approximately 60 days [87].
Differences in the process for other species have been demonstrated, showing that canine spermatogonia type A are stem cells (SSCs or As). These cells are able to self-renew and proliferate, as well as to create, by means of mitosis, intermediate cells and later type B cells, which divide to generate primary spermatocytes [87,88,89].
Since their first identification in 2013 [66], cSSCs have been investigated for the individuation of specific markers, as well as genes regulating self-renewal and differentiation processes (Figure 2). These studies have been conducted either in canine testes (at different stages of the spermatogenesis cycle) or in vitro-cultured cSSCs, demonstrating higher expression levels under this last condition [90].
In detail, the results of these studies demonstrated the expression of several markers, including the CXCR4, IGFBP3, LIN28, and SALL4 genes, in different developmental stages of canine testes [91], albeit significant differences were observed for the immunohistochemical distribution of IGFBP3 and LIN28, which exhibited higher expression levels along SCP3-positive differentiated male germ cells.
More recently, additional characterization studies of cSSCs that were cultured in vitro, also in the presence of FSH, confirmed the presence of the early germline marker OCT4 and demonstrated the expression of the late germline markers PLZF, DAZL, C-kit, and GFRA-1 [92], thus demonstrating a morphological profile that is similar to those of mouse SSC cultures [93]. In particular, C-kit, a marker of spermatogonial differentiation which is also responsible for this process when spermatogonia self-renewal is repressed, was detected at a higher percentage (33,3%) than that demonstrated for humans (only 13% of cells) [89]. In addition, FSH supplementation influenced the self-renewal of these cells, as well as their proliferation process, by activating the GDNF-GFRα1 signalling pathway in them [92]. This study on canine species is of particular interest, considering that similar research has not yet carried out for other animal species (rat, mouse, human) [94,95].
Recently, it has been demonstrated that the mRNAs of THY1 and CDH1 (which are cell-specific to spermatogonia) are expressed in greater abundance in immature canine testes compared with mature ones [96] (Figure 2). The importance of THY1, which is recognized as a surface marker of undifferentiated spermatogonia, was recently evidenced in bull testes, demonstrating that THY1+ cells are enriched from the total testis cell population [97].
5. Potential Effects of Xenobiotic and External Factors on the Biology of Spermatogonial Stem Cells
The first research focusing on the possible effect of xenobiotics and substances interfering with the spermatogenesis process began in the 1970s. Since then, more specific research has demonstrated in recent years that different conditions altering the testicular microenvironment, as well as the effect of xenobiotics, can impact the biology of SSCs and, thus, their suitability for therapeutic use. In fact, conditions of moderate hypoxia (2.5% and 5% O2) improve the proliferation of mouse SSCs, while severe hypoxia induces cells to quiescence [99]. Moreover, it has been confirmed that environmental conditions, such as a high temperature inducing heat stress, can cause a reduction in fertility. However, an interesting study on stallion testes under both normal and cryptorchid conditions (used as a model to evaluate the impact of heat stress) demonstrated that undifferentiated SSCs are not any more affected by long-term exposure to heat stress than other germ cells in the spermatogenesis process [100]. The authors ascribed this result to the incomplete development of the testis under cryptorchid conditions. A different result, by using in vitro-cultured SSCs, was reported by the group of Jia Wang and co-authors [101]. The authors showed a negative effect of a high temperature (45 min of 43 °C), which led to the alteration of the SSCs’ self-renewal ability due to SSC cycle arrest. These data demonstrate an inhibition of the JAK/STAT signalling pathway, which is commonly known for its role in mediating cell proliferation, differentiation, and migration.
Apart from environmental and local conditions affecting the normal physiology of the reproductive tract and its components, research has been particularly focused on the possible effects of endocrine disrupting chemicals (EDCs) on reproductive function, considering their widespread use [102,103]. In this regard, genistein (GEN) and Mono(2-ethylhexyl) Phthalate (MEHP) exposure has been demonstrated to alter the eicosanoid pathway that is involved in the differentiation process that is responsible for human SSCs [104]. Similarly, bisphenol and two of the main analogues (bisphenol-F BPF and bisphenol-S BPS) have been demonstrated to have cytotoxic effects on SSCs [105,106,107]. It is worth noting that chemotherapy has a severe impact on spermatogenesis. Particularly of interest is the effect of cisplatin exposure in reducing the total number of germ cells in human foetal testes, with a negative effect on spermatogonia. Similar results have also been demonstrated on the total germ cell count (including spermatogonial stem cells) in prepubertal human testicular tissue [108].
6. Canine Spermatogonial Stem Cells and Pathophysiological Conditions Affecting Fertility
Recently, scientific interest in the field of reproduction has focused on clarifying the possible impact of several pathophysiological conditions (i.e., cryptorchidism and bacterial infections) on the mechanisms of the spermatogenesis process. Cryptorchidism is one of the most frequent congenital anomalies (with an incidence of approximately 6.8%) and characterized by the testes not descending into the scrotum [109]. Research focusing on the mechanisms of diminished fertility in cryptorchid patients revealed impaired spermatogenesis, which was primarily attributed to dysfunction in the self-renewal and differentiation of spermatogonia [110]. The results of research on cryptorchidism showed defects in the canine testicular germ lines as a result of decreased expressions of PGP9.5 and VASA [111,112]. These results agree perfectly with that obtained recently in horses [100]. A decrease in PGP9.5 expression, along with other markers such as DAZL and FOX, has also been demonstrated in the progression of chronic asymptomatic idiopathic orchitis (CAO), which is known to induce non-obstructive azoospermia (NOA) in male dogs, affecting the regular spermatogenesis process [113]. At the cellular level, different DAZL localizations have been shown for human and animal species. In fact, in dogs, a location in the cytoplasm of undifferentiated and differentiating spermatogonia and spermatocytes was observed [89], while a transition from the nucleus to cytoplasm during meiosis was shown for humans and mice.
The mechanisms that are implicated in the failure of spermatogenesis through cryptorchidism still need to be completely explored. As demonstrated by Hirata, J et al., the direct consequence of heat stress on the development of undescended testes in a cryptorchid condition is a decrease in spermatogonial cells due to their death, which is probably related to oxygen stress [114]. Similar results emerged in previous studies using a model of experimentally induced unilateral cryptorchidism in mice, demonstrating a precocial onset of DNA fragmentation, accompanied by germ cell loss [115]. It is worth noting that the incidence of germ cell apoptosis was more severe in adults than in juvenile animals. However, cryptorchidism can impact spermatogonial compartmentalization slightly differently in different species, with a decrease in UTF1 undifferentiated spermatogonia in humans [116] and impaired spermatogenesis differentiation in rodents [117]. This data indicated that the biochemical mechanisms at the testicular level due to cryptorchid conditions are more complex, and that the onset of different protective mechanisms for undifferentiated spermatogonia must also be considered [110].
7. Canine Spermatogonial Stem Cells for Transplantation
As previously reported briefly, the potential goal of SSCs is their use for transplantation. The development of this technology is important in order to evaluate novel insights regarding the entire spermatogenesis process and improve germline repopulation [118,119]. Several aspects, including knowledge of the spermatogenic process, as well as the enrichment of donor SSCs and preparation of germ cells with mitotically active Sertoli cells, need to be taken into account to guarantee successful transplantation.
It is widely known that for this process to succeed, the endogenous donor SSCs that are isolated from the testicular tissue and transplanted into the recipient’s testis have to pass through the vas deferens, rete testis, and seminiferous tubules, and donor-derived spermatogenesis is re-established in the specific microenvironment, namely, the niche [120,121].
In the course of transplantation, it has been demonstrated that transplanted donor SSCs migrate from the lumen to the basement membrane, behaving differently than during the normal spermatogenesis process.
The first experiments by the group of Dobrinski and collaborators, who aimed to investigate the effect of increasing the phylogenetic distance between donor and recipient animals on the outcome of spermatogonial transplantation, demonstrated a minor success for transplantation of dog SSCs compared with rabbit SSCs, when mice were used as recipients [122]. This important result led the authors to hypothesize that the increase in the phylogenetic distance between donor and recipient could decrease the success of xenogenic transplantation [122]. At the same time, this study has helped the scientific community to understand the factors controlling the spermatogonial process. To improve the methodology and obtain a completely sterile recipient testis for better results in SSC transplantation, dedicated studies have been performed to test protocols and evaluate the different methods for the depletion of host germ cells. Success of transplantation through the use of canine seminiferous tubule cells was obtained using recipient canine testes that were previously irradiated to deplete their endogenous male germ cells (Figure 3) [123]. In relation to the irradiation, a different approach was used by using Busulfan treatment (15-17.5 mg/kg), considering its ability to deplete germ cells and disrupt the junctions between Sertoli cells, thus permitting the migration of transplanted spermatogonia (Figure 3) [124].
The results, demonstrating the success of germ cell depletion in canine recipient testes and the possibility to conserve it at least eight weeks after treatment, suggest that this method is useful for preparing the recipient testes for transplantation. However, also considering the possible toxic effects of busulfan, surgically induced cryptorchidism was proposed as an alternative method [112].
Other experimental studies using cSSCs for xenotransplantation have been conducted to further clarify the spermatogenesis process and the factors influencing the success of this method (Figure 3) [92]. The performed experiments showed that supplemented with FSH to cSSCs improved their colonization along mouse seminiferous tubules in vivo after xenotransplantation [92]. This result was supported by the presence of GFP+ cSSCs along the basal layer of the tubules and the improved percentage of seminiferous tubules that were positive for GFP+ cSSCs at 10 weeks (70 days) after transplantation.
Other authors examined the possibility of transplanting vitrified canine testicular cells into nude mice, demonstrating that SSCs can colonize the seminiferous tubules of the recipient, although the spermatogenesis process was not completed (Figure 3) [125]. The authors conclude that the microenvironment of mouse seminiferous tubules is not ideal for domestic animal-derived germ cell transplantation.
8. Conclusions
Although the research on canine spermatogonial stem cells still has a long way to go, new methodological possibilities appear and help improve cell cultivation techniques in vitro, as well as the characterization of the molecular mechanisms underlying spermatogenesis in vivo. Future research on these cells could facilitate increased knowledge of the various physiological events during spermatogenesis, also clarifying the influence of complex molecular mechanisms and other factors (including, for example, endocrine disruptors) that cause fertility disorders.
Author Contributions
conceptualization, A.P.; literature search - original draft preparation C.S., A.P.; writing review, editing A.P., C.S., N.M., S.T. M.I., V.I.; A.P. C.S. supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yoshida, S.; Sukeno, M.; Nabeshima, Y. A vasculature-associated niche for undifferentiated spermatogonia in the mouse testis. Science 2007, 317, 1722–1726. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L. , Wagers, A.J. No place like home: anatomy and function of the stem cell niche. Mol. Cell. Biol. 2008, 9, 11–21. [Google Scholar]
- La, H.M.; Hobbs, R.M. Mechanisms regulating mammalian spermatogenesis and fertility recovery following germ cell depletion. Cell. Mol. Life Sci. 2019, 76, 4071–4102. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Roy, S.; Ghosh, D.; Nandi, S.K. Role of animal models in biomedical research: a review. Lab. Anim. Res. 2022, 38, 18. [Google Scholar] [CrossRef]
- Lindblad-Toh et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog, Nature 2005, 438, 803–819.
- Mirabella, N.; Pelagalli A., Liguori, G.; Rashedul, M.A., Squillacioti, C. Differential abundances of AQP3 and AQP5 in reproductive tissues from dogs with and without cryptorchidism, Anim. Reprod. Sci. 2021, 228 106735.
- Squillacioti, C.; Mirabella, N.; Liguori, G.; Germano, G.; Pelagalli A. Aquaporins are differentially regulated in canine cryptorchid efferent ductules and epididymis. Animals 2021, 11, 1539. [Google Scholar] [CrossRef]
- Sadek, A.; Khramtsova, Y.; Yushkov, B. Mast cells as a component of spermatogonial stem cells' microenvironment. Int. J. Mol. Sci. 2024, 25, 13177. [Google Scholar] [CrossRef]
- Ibtisham, F.; Awang-Junaidi, A.H.; Honaramooz, A. The study and manipulation of spermatogonial stem cells using animal models. Cell Tissue Res. 2020, 380, 393–414. [Google Scholar] [CrossRef]
- Du, G.; Oatley, M.J.; Law, N.C.; Robbins, C.; Wu, X.; Oatley, J.M. Proper timing of a quiescence period in precursor prospermatogonia is required for stem cell pool establishment in the male germline. Development 2021, 148, 194571. [Google Scholar] [CrossRef]
- Nakagawa, T.; Sharma, M.; Nabeshima, Y.; Braun, R.E.; Yoshida, S. Functional hierarchy and reversibility within the murine spermatogenic stem cell compartment. Science 2010, 328, 62–67. [Google Scholar] [CrossRef]
- Hara, K.; Nakagawa, T.; Enomoto, H.; Suzuki, M.; Yamamoto, M.; Simons, B.D.; Yoshida, S. Mouse spermatogenic stem cells continually interconvert between equipotent singly isolated and syncytial states. Cell. Stem Cell 2014, 14, 658–672. [Google Scholar] [CrossRef]
- Potter, S.J.; De Falco, T. Role of the testis interstitial compartment in spermatogonial stem cell function. Reprod. 2017, 153, R151–R162. [Google Scholar] [CrossRef] [PubMed]
- Oatley, J.M.; Brinster, R.L. The germline stem cell niche unit in mammalian testes. Physiol. Rev. 2012, 92, 577–595. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; Kim, R.; Carrell, D.T.; Goriely, A.; Hotaling, J.M.; Cairns, B.R. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, T.; Orwig, K.E.; Avarbock, M.R.; Brinster, R.L. Remodeling of the postnatal mouse testis is accompanied by dramatic changes in stem cell number and niche accessibility. Proc. Natl. Acad. Sci. USA 2001, 98, 6186–6191. [Google Scholar] [CrossRef]
- Vahid, A.; Parviz, T.; Mansoureh, M.; Reza, Y. Effect of removal of spermatogonial stem cells (sscs) from in vitro culture on gene expression of niche factors in bovine. Avicenna J. Med. Biotechnol. 2016, 8, 133–138. [Google Scholar]
- Meng X, Lindahl M, Hyvonen ME, Parvinen M, de Rooij DG, Hess MW, Raatikainen-Ahokas A, Sainio K, Rauvala H, Lakso M, Pichel JG, Westphal H et al. Regulation of cell fate decision of undifferentiated spermatogonia by GDNF. Science 2000; 287(5457):1489–1493. 37.
- Hofmann, M.C.; Braydich-Stolle, L.; Dym, M. Isolation of male germ-line stem cells; influence of GDNF. Dev. Biol. 2005, 279, 114–124. [Google Scholar] [CrossRef]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Growth factors essential for selfrenewal and expansion of mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16489–16494. [Google Scholar] [CrossRef]
- Martin Laura, A.; Assif, N.; Gilbert, M.; Wijewarnasuriya, D.; Seandel, M. Enhanced fitness of adult spermatogonial stem cells bearing a paternal age-associated FGFR2 mutation. Stem Cell. Rep. 2014, 3, 219–226. [Google Scholar] [CrossRef]
- Chen, S.R.; Liu, Y.X. Regulation of spermatogonial stem cell self-renewal and spermatocyte meiosis by Sertoli cell signaling. Reprod. 2015, 149, R159–R167. [Google Scholar] [CrossRef]
- De Falco, T.; Potter, S.J.; Williams, A.V.; Waller, B.; Kan, M.J.; Capel, B. Macrophages contribute to the spermatogonial niche in the adult testis. Cell Rep. 2015, 12, 1107–1119. [Google Scholar] [CrossRef]
- Oatley, J.M.; Avarbock, M.R.; Brinster, R.L. Glial cell line-derived neurotrophic factor regulation of genes essential for self-renewal of mouse spermatogonial stem cells is dependent on Src family kinase signaling. J. Biol. Chem. 2007, 282, 25842–25851. [Google Scholar] [CrossRef] [PubMed]
- Oatley, M.J.; Kaucher, A.V.; Racicot, K.E. , Oatley J.M. Inhibitor of DNA binding 4 is expressed selectively by single spermatogonia in the male germline and regulates the self-renewal of spermatogonial stem cells in mice. Biol. Reprod. 2011, 85, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.E.; Kim, D.; Kaucher, A.; Oatley, M.J.; Oatley, J.M. CXCL12-CXCR4 signaling is required for the maintenance of mouse spermatogonial stem cells. J. Cell. Sci. 2013, 126, 1009–1020. [Google Scholar] [CrossRef]
- Zhao, H.; Li, T.; Yang, H.; Mehmood, M.U.; Lu, Y.; Liang, X.; Yang, X.; Xu, H.; Lu, K.; Lu, S. The effects of growth factors on proliferation of spermatogonial stem cells from Guangxi Bama mini-pig. Reprod. Dom. Anim. 2019, 54, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Wang, Z.; Li, P.; Tang, J.; Jiao, T.; Li, Y.; Ou, J.; Zou, D.; Li, M.; Mang, X.; Liu, J.; Ma, Y.; Wu, X.; Shi, J.; Chen, S.; He, M.; Lu, Y.; Zhang, N.; Miao, S.; Sun, F.; Wang, L.; Li, K.; Yu, J.; Song, W. Decoding the spermatogonial stem cell niche under physiological and recovery conditions in adult mice and humans. Sci Adv. 2023, 9, eabq3173. [Google Scholar] [CrossRef]
- Pellegrini, M.; Filipponi, D.; Gori, M. , Barrios, F.; Lolicato, F.; Grimaldi, P.; Rossi, P.; Jannini, E.A.; Geremia, R.; Dolci, S. ATRA and KL promote differentiation toward the meiotic program of male germ cells. Cell Cycle 2008, 7, 3878–3888. [Google Scholar] [CrossRef]
- Carlomagno, G.; van Bragt, M.P.; Korver, C.M.; Repping, S.; de Rooij, D.G.; van Pelt, A.M. BMP4-induced differentiation of a rat spermatogonial stem cell line causes changes in its cell adhesion properties. Biol. Reprod. 2010, 83, 742–749. [Google Scholar] [CrossRef]
- Barrios, F.; Filipponi, D.; Campolo, F.; Gori, M.; Bramucci, F.; Pellegrini, M.; Ottolenghi, S.; Rossi, P.; Jannini, E.A.; Dolci, S. SOHLH1 and SOHLH2 control kit expression during postnatal male germ cell development. J. Cell. Sci. 2012, 125, 1455–1464. [Google Scholar]
- Yang, Q.E.; Racicot, K.E.; Kaucher, A.V.; Oatley, M.J.; Oatley, J.M. MicroRNAs 221 and 222 regulate the undifferentiated state in mammalian male germ cells. Development 2013, 140, 280–290. [Google Scholar] [CrossRef]
- Hobbs, R.M.; Fagoonee, S.; Papa, A.; Webster, K.; Altruda, F.; Nishinakamura, R. , Chai, L.; Pandolfi, P.P. Functional antagonism between Sall4 and Plzf defines germline progenitors. Cell Stem Cell 2012, 10, 284–298. [Google Scholar] [CrossRef]
- Goertz, M.J.; Wu, Z.; Gallardo, T.D.; Hamra, F.K.; Castrillon, D.H. Foxo1 is required in mouse spermatogonial stem cells for their maintenance and the initiation of spermatogenesis. J. Clin. Invest. 2011, 121, 3456–3466. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gao, C.; Lin, X.; Ning, Y.; He, W.; Zheng, C.; Zhang, D.; Yan, L.; Jiang, B.; Zhao, Y.; Hossen, M.A.; Han, C. The microRNA miR-202 prevents precocious spermatogonial differentiation and meiotic initiation during mouse spermatogenesis. Development 2021, 148, 199799. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Cui, Y.; Liu, B.; Li, C.; Du, L.; Tang, R. Qin, L.; Jiang, Y.; Li, J.; Yu, X., He, Q.; He, Z. Hsa-miR-1908-3p mediates the self-renewal and apoptosis of human spermatogonial stem cells via targeting KLF2. Nucl. Acids 2020, 20, 788–800. [Google Scholar] [CrossRef]
- Zhou, F.; Chen, W.; Cui, Y.; Liu, B.; Yuan, Q.; Li, Z.; He, Z. miRNA-122-5p stimulates the proliferation and DNA synthesis and inhibits the early apoptosis of human spermatogonial stem cells by targeting CBL and competing with lncRNA Casc7. Aging 2020, 12, 25528–25546. [Google Scholar] [CrossRef]
- Fu, H.; Zhou, F.; Yuan, Q.; Zhang, W.; Qiu, Q.; Yu, X.; He, Z. miRNA-31-5p mediates the proliferation and apoptosis of human spermatogonial stem cells via targeting JAZF1 and Cyclin A2. Nucl. Acids 2019, 14, 90–100. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, F.; Ren, Y.; Zhou, Z.; Chen, N.; Huang, Y.; Lei, C.; Chen, H.; Dang, R. MiRNAs expression profiling of bovine (Bos taurus) testes and effect of bta-miR-146b on proliferation and apoptosis in bovine male germline stem cells. Int. J. Mol. Sci. 2020, 21, 3846. [Google Scholar] [CrossRef]
- Yu, M.; Mu, H.; Niu, Z.; Chu, Z.; Zhu, H.; Hua, J. miR-34c enhances mouse spermatogonial stem cells differentiation by targeting Nanos2. J. Cell. Biochem. 2014, 115, 232–242. [Google Scholar] [CrossRef]
- Li, Q.; Li, H.; Liang, J.; Mei, J.; Cao, Z.; Zhang, L.; Luo, J.; Tang, Y. , Huang, R., Xia, H., Zhang, Q.; Xiang, Q.; Yang, Y.; Huang, Y. Sertoli cell-derived exosomal microRNA-486-5p regulates differentiation of spermatogonial stem cell through PTEN in mice. J. Cell. Mol. Med. 2021, 25, 3950–3962. [Google Scholar] [CrossRef]
- Tong, M.H.; Mitchell, D.A.; McGowan, S.D.; Evanoff, R.; Griswold, M.D. Two miRNA clusters, Mir-17-92 (Mirc1) and Mir-106b-25 (Mirc3), are involved in the regulation of spermatogonial differentiation in mice. Biol. Reprod. 2012, 86, 72. [Google Scholar] [CrossRef]
- Xie, R.; Lin, X.; Du, T.; Xu, K.; Shen, H.; Wei, F.; Hao, W.; Lin, T.; Lin, X.; Qin, Y. , Wang, H., Chen, L., Yang, S.; Yang, J.; Rong, X.; Yao, K., Xiao, D., Jia, J.; Sun, Y. Targeted disruption of miR-17-92 impairs mouse spermatogenesis by activating mTOR signaling pathway. Medicine 2016, 95, e2713. [Google Scholar] [CrossRef]
- Masaki, K.; Sakai, M.; Kuroki, S.; Jo, J.I.; Hoshina, K.; Fujimori, Y.; Oka, K.; Amano, T.; Yamanaka, T.; Tachibana, M.; Tabata, Y.; Shiozawa, T.; Ishizuka, O.; Hochi, S.; Takashima, S. FGF2 has distinct molecular functions from GDNF in the mouse germline niche. Stem Cell Rep. 2018, 10, 1782–1792. [Google Scholar] [CrossRef] [PubMed]
- Binsila, B.K. , Selvaraju, S. ; Ghosh, S.K.; Ramya, L.; Arangasamy, A.; Ranjithkumaran, R.; Bhatta, R. EGF, GDNF, and IGF-1 influence the proliferation and stemness of ovine spermatogonial stem cells in vitro. J. Assisted Reprod. Genetics 2020, 37, 2615–2630. [Google Scholar]
- Parker, N.; Laychur, A.; Sukwani, M.; Orwig, K.E.; Oatley, J.M.; Zhang, C.; Rutaganira, F.U.; Shokat, K.; Wright, W.W. Spermatogonial stem cell numbers are reduced by transient inhibition of GDNF signaling but restored by self-renewing replication when signaling resumes. Stem Cell Rep. 2021, 16, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Kanatsu-Shinohara, M.; Toyokuni, S.; Shinohara, T. cd9 is a surface marker on mouse and rat male germline stem cells. Biol. Reprod. 2004, 70, 70–75. [Google Scholar] [CrossRef]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Spermatogonial stem cells share some, but not all, phenotypic and functional characteristics with other stem cells. Proc. Natl. Acad. Sci. USA 2003, 100, 6487–6492. [Google Scholar] [CrossRef]
- Shinohara, T.; Avarbock, M.R.; Brinster, R.L. beta1- and alpha6-integrin are surface markers on mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 1999, 96, 5504–5509. [Google Scholar] [CrossRef]
- Zi, H.; Kokkinaki, M.; Jiang, J.; Dobrinski, I. , Dym, M. Isolation, characterization, and culture of human spermatogonia. Biol. Reprod. 2010, 82, 363–372. [Google Scholar]
- Wu, X.; Schmidt, J.A.; Avarbock, M.R. Tobias, Y.W.; Carlson, C.A.; Kolon, T.F.; Ginsberg, J.P.; Brinster, R.L. Prepubertal human spermatogonia and mouse gonocytes share conserved gene expressionof germline stem cell regulatory molecules. Proc. Natl. Acad. Sci. USA. 2009, 106, 21672–21677.66. [Google Scholar] [CrossRef]
- Dym, M.; Kokkinaki, M. , He, Z. Spermatogonial stem cells: mouse and human comparisons. Birth Defects Res C Embryo Today. 2009, 87, 27–34. [Google Scholar] [CrossRef]
- Oatley, J.M.; Avarbock, M.R.; Telaranta, A.I.; Fearon, D.T.; Brinster, RL. Identifying genes important for spermatogonial stem cell selfrenewal and survival. Proc. Natl. Acad. Sci. USA 2006, 103, 9524–9529. [Google Scholar] [CrossRef]
- Oatley, J.M.; Brinster, R.L. Regulation of spermatogonial stem cell selfrenewal in mammals. Ann. Rev. Cell. Dev. Biol. 2008, 24, 263–286. [Google Scholar] [CrossRef] [PubMed]
- Kanatsu-Shinohara, M.; Chen, G.; Morimoto, H.; Shinohara, T. CD2 is a surface marker for mouse and rat spermatogonial stem cells. J. Reprod. Develop. 2020, 66, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jin, C.; Li, P.; Li, Y.; Tang, J.; Yu, Z.; Jiao, T.; Ou, J.; Wang, H.; Zou, D. , Li, M.; Mang, X.; Liu, J.; Lu,Y.; Li, K., Zhang, N., Miao, S.; Yu, J.; Wang, L.; Song, W. FOXC2 marks and maintains the primitive spermatogonial stem cells subpopulation in the adult testis. eLife 2023, 1–34. [Google Scholar] [CrossRef]
- Lu, X.; Yin, P.; Li, H.; Gao, W.; Jia, H.; Ma, W. Transcriptome analysis of key genes involved in the initiation of spermatogonial stem cell differentiation. Genes 2024, 15, 141. [Google Scholar] [CrossRef]
- Geng, Q.; Hu, C.; Zhao, Z.; Wang, Z.; Cheng, F.; Chen, J.; Zuo, Q.; Zhang, Y. miR-1458 is inhibited by low concentrations of Vitamin B6 and targets TBX6 to promote the formation of spermatogonial stem cells in Rugao Yellow Chicken. Poultry Sci. 2025, 104, 104583. [Google Scholar] [CrossRef]
- Yeh, J.R.; Zhang, X.; Nagano, M.C. Establishment of a short-term in vitro assay for mouse spermatogonial stem cells. Biol. Reprod. 2007, 77, 897–904. [Google Scholar] [CrossRef]
- Di Persio, S.; Neuhaus, N. Human spermatogonial stem cells and their niche in male (in)fertility: novel concepts from single-cell RNA-sequencing. Hum. Reprod. 2023, 38, 1–13. [Google Scholar] [CrossRef]
- Izadyar, F.; Spierenberg, G.T.; Creemers, L.B. , den Ouden, K.; de Rooij, D.G. Isolation and purification of type A spermatogonia from the bovine testis. Reproduction 2002, 124, 85–94. [Google Scholar] [CrossRef]
- Damyanova, K.B.; Nixon, B.; Johnston, S.D.; Gambini, A.; Benitez, P.P.; Lord, T. Spermatogonial stem cell technologies: applications from human medicine to wildlife conservation, Biol. Reprod. 2024, 111, 757–779. [Google Scholar] [CrossRef]
- Xi, H.M.; Ren, Y.J.; Ren, F.; Li, Y.; Feng, T.Y.; Wang, Z.; Du, Y.Q.; Zhang, L.K.; Hu, J.H. Recent advances in isolation, identification, and culture of mammalian spermatogonial stem cells. Asian J. Androl. 2022, 24, 5–14. [Google Scholar] [CrossRef]
- Binsila, K.B.; Selvaraju, S.; Ghosh, S.K. , Parthipan, S. ; Archana, S.S.; Arangasamy, A.; Prasad, J.K.; Bhatta, R.; Ravindra, J.P. Isolation and enrichment of putative spermatogonial stem cells from ram (Ovis aries) testis, Anim. Reprod. Sci. 2018, 196, 9–18. [Google Scholar] [PubMed]
- Nabulindo, N.W.; Nguhiu-Mwangi, J.; Kipyegon, A.N.; Ogugo, M.; Muteti, C.; Tiambo, C.; Oatley, M.J.; Oatley, J.M.; Kemp, S. Culture of kenyan goat (Capra hircus) undifferentiated spermatogonia in feeder-free conditions, Front. Vet. Sci. 2022, 9, 894075. [Google Scholar]
- Harkey, M.A.; Asano, A.; Zoulas, M.E. , Torok-Storb, B.; Nagashima, J., Travis, A. Isolation, genetic manipulation, and transplantation of canine spermatogonial stem cells: progress toward transgenesis through the male germ-line. Reproduction 2013, 146, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Das Dipak Bhuyan, A.; Lalmalsawma, T. , Pratim Das, P., Koushik, S., Chauhan, M.S.; Bhuyan, M. Propagation of porcine spermatogonial stem cells in serum-free culture conditions using knockout serum replacement. Reprod. Dom. Anim. 2023, 58, 219–229. [Google Scholar]
- Binsila, B.K.; Selvaraju, S.; Ghosh, S.K.; Ramya, L.; Arangasamy, A.; Ranjithkumaran, R.; Bhatta, R. EGF, GDNF, and IGF-1 influence the proliferation and stemness of ovine spermatogonial stem cells in vitro. J. Assist. Reprod. Genet. 2020, 37, 2615–2630. [Google Scholar] [CrossRef]
- Sahare, M.; Kim, S.M.; Otomo, A.; Komatsu, K.; Minami, N.; Yamada, M.; Imai, H. Factors supporting long-term culture of bovine male germ cells. Reprod. Fertil. Dev. 2015, 28, 2039–2050. [Google Scholar] [CrossRef]
- Bedford-Guaus, S.J.; Kim, S. , Mulero, L.; Vaquero, J.M., Morera, C., Adan-Milanès, R., Veiga, A.; Raya, A. Molecular markers of putative spermatogonial stem cells in the domestic cat. Reprod. Dom. Anim. 2017, 52, 177–186. [Google Scholar] [CrossRef]
- Zhao, X.; Wan, W. , Li, B., Zhang, X., Zhang, M., Wu, Z.; Yang, H. Isolation and in vitro expansion of porcine spermatogonial stem cells. Reprod. Dom. Anim. 2022, 57, 210–220. [Google Scholar] [CrossRef]
- Shirazi, M.S.; Heidari, B. , Naderi, M.M.; Behzadi, B.; Sarvari, A.; Borjian-Boroujeni, S., Farab, M., Shirazi, A. Transplantation of goat spermatogonial stem cells into the mouse rete testis. Int. J. Anim. Biol. 2015, 1, 61–68. [Google Scholar]
- You, F.W.; Bei, C.S.; Giang, D.D.; You, L.Q. , YanFei, D., XiaoCan, L.; Chan, L.; Ben, H.; DeShun, S. Isolation and identification of prepubertal buffalo (Bubalus bubalis) spermatogonial stem cells. Asian Australas. J. Anim. Sci. 2016, 29, 1407–1415. [Google Scholar]
- Indu, S.; Devi, A.N.; Sahadevan, M.; Sengottaiyan, J.; Basu, A.; Raj K, S.; Kumar, P.G. , Expression profiling of stemness markers in testicular germline stem cells from neonatal and adult Swiss albino mice during their transdifferentiation in vitro. Stem Cell Res. Ther. 2024, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Ikeda, S.; Minami, N.; Yamada, M.; Imai, H. Long-term culture of ndifferentiated spermatogonia isolated from immature and adult bovine testes. Mol. Reprod. Dev. 2018, 85, 236–249. [Google Scholar]
- Zhang, X.Y.; Li, T.T.; Liu, Y.R.; Geng, S.S.; Luo, A.L.; Jiang,M.S.; Liang, X.W.; Shang, J.H.; Lu, K.H.; Yang, X.G. Transcriptome analysis revealed differences in the microenvironment of spermatogonial stem cells in seminiferous tubules between pre-pubertal and adult buffaloes. Reprod. Dom. Anim. 2021, 56, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Ibtisham, F.; Tang, S.; Song, Y.; Wanze, W.; Xiao, M.; Honaramooz, A.; An, L. Optimal isolation, culture, and in vitro propagation of spermatogonial stem cells in Huaixiang chicken. Reprod. Dom. Anim. 2024, 59, e14661. [Google Scholar] [CrossRef]
- Costa, G.M.J.; Avelar, G.F.; Lacerda, S.M.S.N.; Figueiredo, A.F.A.; Tavares, A.O.; Rezende-Neto, J.V.; Martins, F.G.P.; França, L.R. Horse spermatogonial stem cell cryopreservation: feasible protocols and potential biotechnological applications. Cell Tissue Res. 2017, 370, 489–500. [Google Scholar] [CrossRef]
- Baarends, W.; Grootegoed, J. Molecular biology of male gametogenesis. Molecular biology in reproductive medicine New York, USA, 1999, 271-295.
- Kashfi, A.; Sani,R.N.; Ahmadi-hamedani, M. The beneficial effect of equine chorionic gonadotropin hormone (eCG) on the in vitro co-culture of bovine spermatogonial stem cell with Sertoli cells. Comp. Clin. Pathol. 2019, 28, 701–704. [Google Scholar] [CrossRef]
- Navid, S.; Rastegar, T.; Baazm, M.; Alizadeh, R.; Talebi, A.; Gholami, K.; Khosravi-Farsani, S.; Koruji, M.; Abbasi, M. In vitro effects of melatonin on colonization of neonate mouse spermatogonial stem cells. Syst. Biol. Reprod. Med. 2017, 63, 370–381. [Google Scholar] [CrossRef]
- Feng, T.Y.; Li, Q.; Ren, F.; Xi, H.M.; Lv, D.L.; Yu Li, Y.; Hu, J.H. Melatonin protects goat spermatogonial stem cells against oxidative damage during cryopreservation by improving antioxidant capacity and inhibiting mitochondrial apoptosis pathway. Oxid. Med. Cell. Longev. 2020, 31, 5954635. [Google Scholar] [CrossRef]
- Salimi, H.; Feyli, P.R.; Yari, K. , Wong, A.; Moghaddam, A.A. Low testosterone concentration improves colonisation and viability in the co-cultured goat spermatogonial stem cell with Sertoli cells. Reprod. Domest. Anim. 2024, 59, e14729. [Google Scholar] [CrossRef]
- Jafarnejad, A.; Aminafshar, M.; Zandi, M. , Sanjabi, M.R.; Kashan, N.E. Optimization of in vitro culture and transfection condition of bovine primary spermatogonial stem cells. S. Afr. J. Anim. Sci. 2018, 48, 108–116. [Google Scholar] [CrossRef]
- Kushki, D.; Azarnia, M.; Gholami, M.R. Antioxidant effects of selenium on seminiferous tubules of immature mice testis. Zahedan J. Res. Med. Sci. 2015, 17, 29–33. [Google Scholar] [CrossRef]
- Zolfaghari, K.; Zandi, M.; Sanjabi, M.R.; Ghaedrahmati, A. In Vitro culture of ovine spermatogonial stem cells: effects of grape seed extracts and vitamin C. Gene Cell Tissue 2024, 11, e135750. [Google Scholar] [CrossRef]
- Soares, J.M.; Avelar, G.F.; De França, L.R. The seminiferous epithelium cycle and its duration in different breeds of dog (Canis familiaris). J. Anat. 2009, 215, 462–471. [Google Scholar] [CrossRef]
- Fayomi, A.P.; Orwig, K.E. Spermatogonial stem cells and spermatogenesis in mice, monkeys and men. Stem Cell Res. 2018, 29, 207–214. [Google Scholar] [CrossRef]
- Pieri, N.C.G.; De Souza, A.F.; Mançanares, A.C.F.; Roballo, K.; Casals, J.; Martins, D.D.S.; Ambrósio, C.E. Immunolocalization of proteins in the spermatogenesis process of canine. Reprod. Domest. Anim. 2016, 52, 170–176. [Google Scholar] [CrossRef]
- Lee, K.H.; Lee, R.; Lee, W.Y.; Kim, D.H.; Chung, H.J.; Kim, J.H.; Kim, N.H.; Choi, S.H.; Kim, J.H. , Song, H. Identification and in vitro derivation of spermatogonia in beagle testis. PLoS One 2014, 9, e109963. [Google Scholar]
- Lee, W.Y.; Lee, R.; Park, H.J.; Do, J.T.; Park, C.; Kim, J.H.; Jhunc, H.; Leed, J.H.; Hure, T.; Song, H. Analysis of putative biomarkers of undifferentiated spermatogonia in dog testis. Anim. Reprod. Sci. 2017, 185, 174–180. [Google Scholar] [CrossRef]
- Pieri, N.C.G.; Mançanares, A.C.F.; de Souza, A.F.; Fernandes, H.; Diaza, A.M.G.; Bressan, F.F.; Roballo, K.C.S.; Casals, J.B.; Binelli, M.; Ambrósio, C.E.; dos Santos Martins, D. Xenotransplantation of canine spermatogonial stem cells (cSSCs) regulated by FSH promotes spermatogenesis in infertile mice. Stem Cell Res. Ther. 2019, 10, 135. [Google Scholar] [CrossRef]
- Ding, L.; Yan, G.J.; Ge, Q.Y.; Yu, F.; Zhao, X.; Diao, Z.Y.; Wang, Z.Q.; Yang, Z.Z.; Sun, H.X.; Hu, Y.L. FSH acts on the proliferation of type a spermatogonia via Nur77 that increases GDNF expression in the Sertoli cells. FEBS Lett. 2011, 15, 2437–2444. [Google Scholar] [CrossRef]
- Sluka, P.; O’Donnell, L.; Bartles, J.R.; Stanton, P.G. FSH regulates the formation of adherens junctions and ectoplasmic specialisations between rat Sertoli cells in vitro and in vivo. J. Endocrinol. 2006, 189, 381–395. [Google Scholar] [CrossRef]
- Orth, J.M.; McGuinness, M.P.; Qiu, J.; Jester, W.F.; Li, L.H. Use of in vitro systems to study male germ cell development in neonatal rats. Theriogenol. 1998, 49, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, V.R. Kasimanickam, R.K. Sertoli, Leydig, and spermatogonial cells’ specific gene and protein expressions as dog testes evolve from immature into mature states. Animals 2022, 12, 271. [Google Scholar] [CrossRef] [PubMed]
- Reding, S.C.; Stepnoski, A.L.; Cloninger, L.W.; Oatley, J.M. THY1 is a conserved marker of undifferentiated spermatogonia in the pre-pubertal bull testis. Reprod. 2010, 139, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Tarnawa, E.D.; Baker, M.D.; Aloisio, G.M.; Carr, B.R.; Castrillon, D.H. Gonadal expression of foxo1, but not foxo3, is conserved in diverse mammalian species. Biol. Reprod. 2013, 88, 103. [Google Scholar] [CrossRef]
- Wang, J.; Xue, X.; Fan, K.; Liu, Q.; Zhang, S.; Peng, M.; Zhou, J.; Cao, Z. Moderate hypoxia modulates ABCG2 to promote the proliferation of mouse spermatogonial stem cells by maintaining mild ROS levels. Theriogenol. 2020, 145, 149e157. [Google Scholar] [CrossRef]
- Shakeel, M.; Choi, Y.; Yoon, M. Expression pattern of germ cell markers in cryptorchid stallion Testes, Reprod. Dom. Anim. 2024, 59, e14561. [Google Scholar] [CrossRef]
- Wang, J.; Gao, W.J.; Deng, S.L.; Liu, X.; Jia, H.; Ma, W.Z. High temperature suppressed SSC self renewal through S phase cell cycle arrest but not apoptosis. Stem Cell Res. Ther. 2019, 10, 227. [Google Scholar] [CrossRef]
- Lymperi, S.; Giwercman, A. Endocrine disruptors and testicular function. Metabolism 2018, 86, 79–90. [Google Scholar] [CrossRef]
- Sharpe, R.M.; Skakkebaek, N.E. Are oestrogens involved in falling sperm counts and disorders of the male reproductive tract? The Lancet 1993, 341, 1392–1396. [Google Scholar] [CrossRef]
- Tran-Guzman, A.; Moradian, R.; Cui, H.; Culty, M. In vitro impact of genistein and mono(2-ethylhexyl) phthalate (MEHP) on the eicosanoid pathway in spermatogonial stem cells. Reprod. Toxicol. 2022, 107, 150–16. [Google Scholar] [CrossRef]
- Kim, S.H.; Shin, S.H.; Kim, S.M.; Jung, S.E.; Shin, B.J.; Ahn, J.S.; Lim, K.T.; Kim, D.H.; Lee, K.; Ryu, B.J. Bisphenol analogs downregulate the self-renewal potential of spermatogonial stem cells. World J. Mens Health 2025, 43, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Stanojević, M.; Sollner Dolenc, M. Mechanisms of bisphenol A and its analogs as endocrine disruptors via nuclear receptors and related signaling pathways. Arch. Toxicol. 2025, 99, 2397–2417. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.S.; Won, J.H.; Kim, D.Y.; Jung, S.E.; Kim, B.J.; Kim, J.M.; Ryu, B.Y. Transcriptome alterations in spermatogonial stem cells exposed to bisphenol A. Anim. Cells Syst. 2022, 26, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Tharmalingam, M.D.; Matilionyte, G.; Wallace, W.H.B.; Stukenborg, J.B.; Jahnukainen, K.; Oliver, E.; Goriely, A.; Lane, S.; Guo, J.; Cairns, B.; Jorgensen, A.; Allen, C.M.; Lopes, F.; Anderson, R.A.; Spears, N.; Mitchell, R.T. Cisplatin and carboplatin result in similar gonadotoxicity in immature human testis with implications for fertility preservation in childhood cancer. BMC Med. 2020, 18, 374. [Google Scholar] [CrossRef]
- Yates, D.; Hayes, G.; Heffernan, M.; Beynon, R. Incidence of cryptorchidism in dogs and cats. Vet. Rec. 2003, 152, 502–504. [Google Scholar] [CrossRef]
- Liu, X.W.Q.; Zhuang, Z.; Cheng, J.; Zhang, W.; Jiang, Q.; Guo,Y.; Li, R.; Lu, X.; Cui, L.; Weng, J.; Tang, Y.; Yue, J.; Gao, S.; Hong, K.; Qiao, J.; Jiang, H.; Guo, J.; Zhang, Z. Decoding the pathogenesis of spermatogenic failure in cryptorchidism through single-cell transcriptomic profiling, Cell Rep. Med. 2024, 5, 101709.
- Jhun, H.; Lee, W.Y.; Park, J.K.; Hwang, S.G.; Park, H.J. Transcriptomic analysis of testicular gene expression in a dog model of experimentally induced cryptorchidism. Cells 2022, 11, 2476. [Google Scholar] [CrossRef]
- Lee, W.Y.; Lee, R.; Song, H.; Hur, T.Y.; Lee, S.; Ahn, J.; Jhun, H. Establishment of a surgically induced cryptorchidism canine recipient model for spermatogonial stem cell transplantation. Lab. Anim. Res. 2016, 32, 257–266. [Google Scholar] [CrossRef]
- Reifarth, L.; Körber, H.; Packeiser, E.M.; Goericke-Pesch, S. Detection of spermatogonial stem cells in testicular tissue of dogs with chronic asymptomatic orchitis. Front. Vet. Sci. 2023, 10, 1205064. [Google Scholar] [CrossRef]
- Hirata, J.; Ko, J.A.; Mochizuki, H.; Funaishi, K.; Yamane, K.; Sonoda, K.-H.; Kiuchi, Y. Oxidative stress regulates expression of claudin-1 in human RPE cells. Open Life Sci. 2014, 9, 461–468. [Google Scholar] [CrossRef]
- Yin, Y.; Hawkins, K.L.; DeWolf, W.C.; Morgentaler, A. Heat stress causes testicular germ cell apoptosis in adult mice. J. Androl. 1997, 18, 159–165. [Google Scholar] [CrossRef]
- Di Persio, S.; Tekath, T.; Siebert-Kuss, L.M.; Cremers, J.F.; Wistuba, J.; Li, X.; Meyer Zu Horste, G.; Drexler, H.C.A.; Wyrwoll, M.J.; Tuttelmann, F. , Dugas, M.; Kliesch, S.; Schlatt, S.; Laurentino, S.; Neuhaus, N. Single-cell RNA-seq unravels alterations of the human sper matogonial stem cell compartment in patients with impaired spermatogenesis. Cell Rep. Med. 2021, 2, 100395. [Google Scholar] [CrossRef] [PubMed]
- Agoulnik, A.I.; Huang, Z.; Ferguson, L. Spermatogenesis in cryptorchidism. Methods Mol. Biol. 2012, 825, 127–147. [Google Scholar] [PubMed]
- Ogawa, T.; Dobrinski, I.; Avarbock, M.R.; Brinster, R.L. Transplantation of male germ line stem cells restores fertility in infertile mice. Nat Med. 2000, 6, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Arregui, L.; Dobrinski, I. Xenografting of testicular tissue pieces: twelve years of an in vivo spermatogenesis system. Reprod. 2014, 148, R71–R84. [Google Scholar] [CrossRef]
- Ogawa, T.; Aréchaga, J.M.; Avarbock, M.R.; Brinster, R.L. Transplantation of testis germinal cells into mouse seminiferous tubules. Int. J. Dev. Biol. 1997, 41, 111–122. [Google Scholar]
- Nagano, M.; Avarbock, M.R.; Brinster, R.L. Pattern and kinetics of mouse donor spermatogonial stem cell colonization in recipient testes. Biol. Reprod. 1999, 60, 1429–1436. [Google Scholar] [CrossRef]
- Dobrinski, I.; Avarbock, M.R.; Brinster, R.L. Transplantation of Germ Cells from Rabbits and Dogs into Mouse Testes, Biology of Reproduction 1999, 61, 1331–1339.
- Yeunhee, Kim; et al. , Production of donor-derived sperm after spermatogonial stem cell transplantation in the dog. Reprod. 2008, 136, 823–831. [Google Scholar]
- Hur, T.J.; Lee, S.H.; Ock, S.A.; Song, H.; Park, H.J.; Lee, R.; Sung, S.H.; Jhun, H.; Lee, W.Y. Dose-dependent effects of busulfan on dog testes in preparation for spermatogonial stem cell transplantation, Lab. Anim. Res. 2017, 33, 264–269. [Google Scholar]
- Lee, K.H.; Lee, W.Y., Kim, D.H.; Lee, S.H.; Do, J.T.; Park, C.; Kim, J.H., Choi, Y.S.; Song, H. Vitrified canine testicular cells allow the formation of spermatogonial stem cells and seminiferous tubules following their xenotransplantation into nude mice, Sci. Rep. 2016, 6, 21919.
Figure 1.
Organization of the SSC niche and list of main components and secreted factors.

Figure 2.
Main characteristics of canine spermatogonial stem cells.
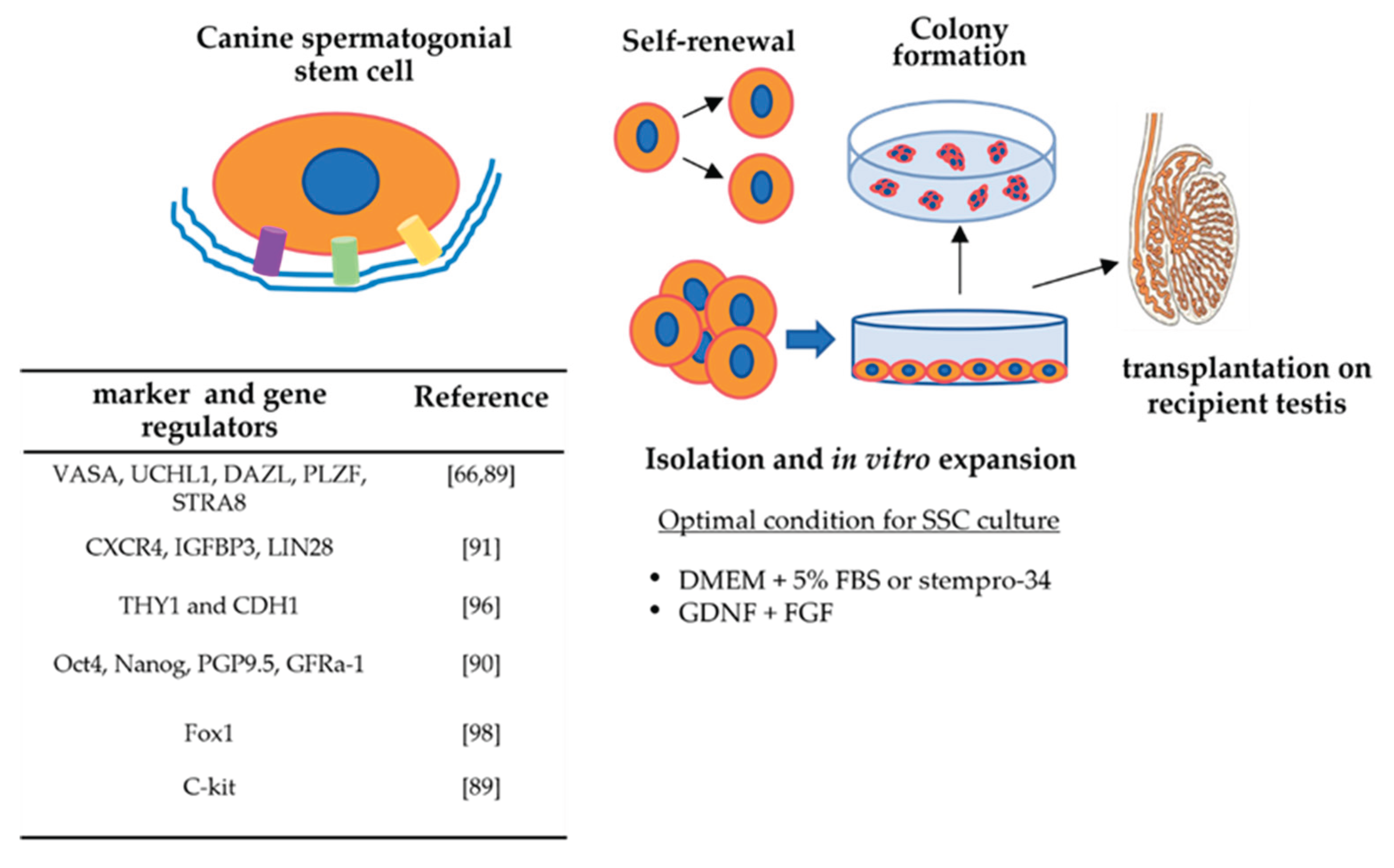
Figure 3.
Transplantation of canine SSCs (SSCT) into recipient testis.
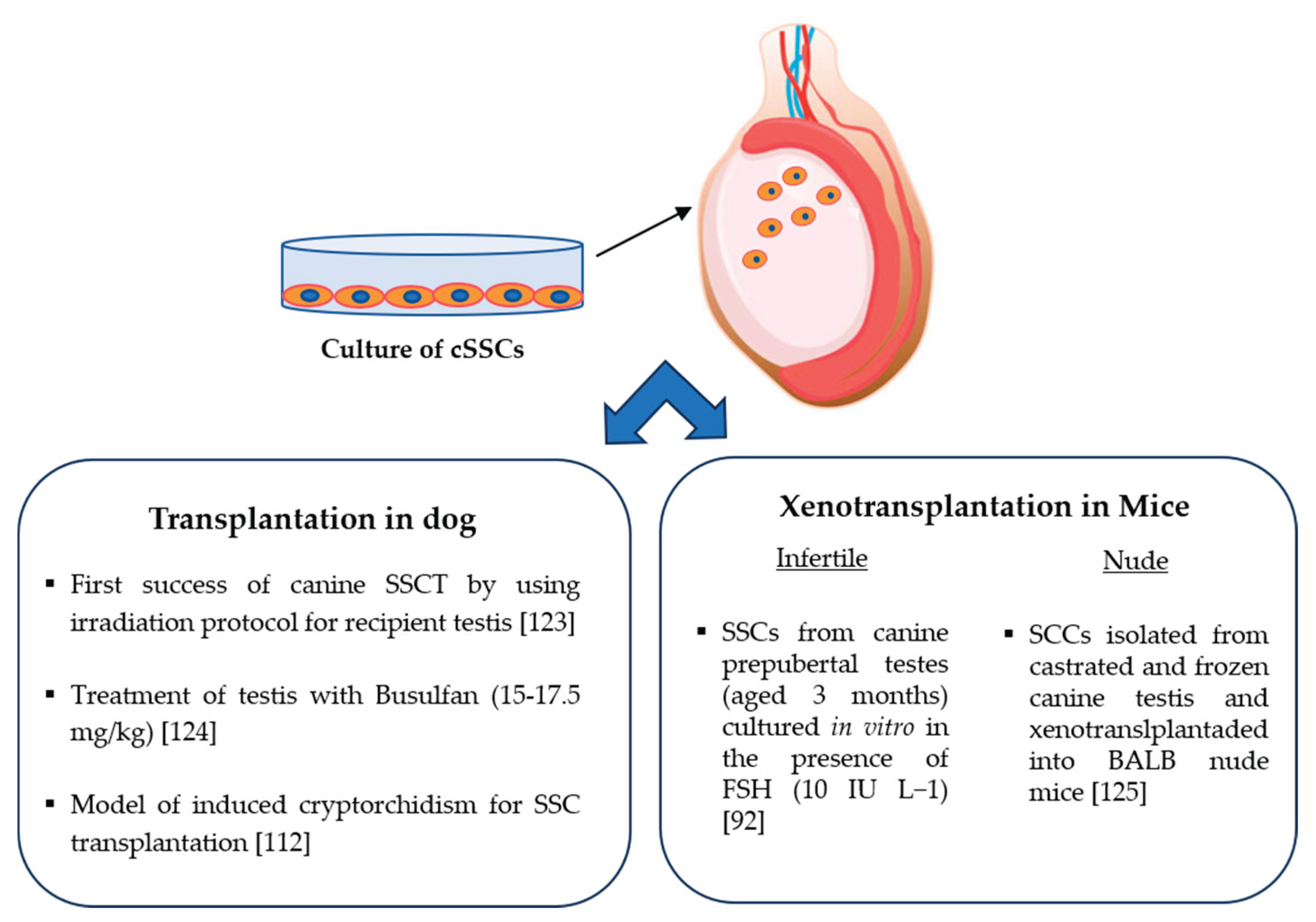

Table 1.
Main factors and gene regulators involved in self-renewal and differentiation of SSCs.
| Factor | Role at level of SSC | Mechanism involved | Species | Reference |
|---|---|---|---|---|
|
GDNF |
self-renewal self-renewal and proliferation |
Nanos2, Etv5, Lhx1, T, Bcl6b, Id1, and Cxcr4 |
mouse swine |
[20,24,25,26] [27] |
| IGF1, IGFBP7, NKCC1, and protein-tyrosine phosphatase | self-renewal and proliferation | CCL24, IGFBP7, and TEK | mouse and human |
[28] |
| retinoic acid | differentiation | Downregulation of GDNF expression activation of differentiation factors (BMP and SCF, SOHL1, SOHL2 |
mouse, rat |
[29,30,31,32] |
| PLZF transcription factor | self-renewal | SALL4 protein | mouse | [33] |
| FOXO1 transcription factor | self-renewal | PI3K-Akt signaling | mouse | [34] |
|
micro-RNAs miR-202 |
self-renewal | Influence of regulators such as STRA8 and DMRT6 | mouse | [35] |
| Hsa-miR-1908-3p | self-renewal |
Kruppel-like factor 2 (KLF2) | human | [36] |
| miRNA-122-5p and miRNA-31-5p | proliferation | transcription factor CBL | human | [37,38] |
| bta-miR-146b | inhibit proliferation and promote apoptosis | n.d. | bovine | [39] |
| miR-34c | differentiation | Inhibition of the function of NANOS2 gene | mouse | [40] |
| miR-486-5p | differentiation | up regulating the expression of STRA8 and SYCP3 | mouse | [41] |
| miR-17-92 and miR-202 | spermatogenesis | Involvement of Bcl2l11, Kit, Socs3, and Stat3 | mouse | [35,42,43] |
Table 2.
Protocols for the isolation and enrichment of SSCs from domestic animals.
| Animal | Optimal age for testis collection | Isolation method | Enrichment method | Factors added to the Culture Medium |
Evaluation of SSC proliferation | Reference |
|---|---|---|---|---|---|---|
| dog | 3–5 month old (pre-pubertal stage) | collagenase-only digestion | SG medium enriched with GDNF, FGF2, EGF, soluble GFRA1, LIF, and a laminin substratum | Note :the enriched cells can survive for several weeks | [66] | |
| pig | 1 month 7-15 days |
two-step enzymatic digestion two-step enzymatic treatment with collagenase,hyaluronidase type II, DNase I and trypsin-EDTA |
gelatin-coated differential plating (laminin and PLL) Sertoli cell feeder layer |
GDNF, FGF2, IGF1 and LIF EGF, epidermal growth factor; FGF, fibroblast growthfactor; GDNF, glial cell line-derived neurotrophic factor; KSR, knockout serum replacement; |
25 days > 30 days |
[71] [67] |
| goat | 4 months | two-step enzymatic digestion |
percoll gradient 32% | LIF (10 ng ml-1), EGF (20 ng ml-1), bFGF (10 ng ml-1), GDNF |
15 days | [72] |
| sheep | two-step enzymatic digestion |
ficoll gradient (12%) and plating (laminin [20 μg/ml in combination with BSA] | GDNF (40ng/ml, EGF (20 ng/ml), and IGF1 (100 ng/ml) | 30 days | [68] | |
| calf | 5-7 months | three-step enzymatic digestion 1°(collagenase Type IV), 2° (collagenase Type IV + hyaluronidase), 3° trypsin and DNase I |
poly-L-lysine-coated method | knockout serum replacement (KSR) (15%) | > 2 months | [69] |
| chicken | 21 days | two-step enzymatic digestion |
differential plating |
2% FBS, GDNF (20 ng/ mL), bFGF (30 ng/mL), or LIF (5 ng/mL) |
7 days | [77] |
| buffalo | two-step enzymatic digestion |
FBS (2.5%) and GDNF (40 ng/mL) | days | [73] | ||
| cat | two-step enzymatic digestion |
gelatin-coated method |
GDNF (15 ng/mL) | 43 days | [70] | |
| horse | two-step enzymatic digestion |
percoll gradient (40%) | FBS (10%) | isolated SSCs cryopreserved after thawed demonstated metabolic activity as the fresh cells | [78] |
Table 3.
Factors improving biological aspects of SSC isolated from animals and cultured in vitro.
| Animals | Factor | Influence on aspect of SSCs biology | Reference |
|---|---|---|---|
| mouse | melatonin (100 µM) |
cell viability improvement | [81] |
| goat | melatonin (1 μM) added to the culture medium | cell viability improvement during cryopreservation | [82] |
| goat | testosterone (60 μg/mL) |
improvement of cell viability and colonization | [83] |
| calf | equine chorionic gonadotropin hormone (eCG) (5 IU/ml) | cell colony formation improvement | [80] |
| calf | vitamin C (50 µg/mL) |
improvement of cell viability and colonization | [84] |
| sheep | vitamin C (50 g/mL) |
cell viability improvement | [86] |
| calf | α-tocopherol analogue (25 µg/mL ) |
improvement of cell viability and colonization | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



