
Submitted:
30 June 2025
Posted:
30 June 2025
You are already at the latest version
Abstract
The "dark genome," comprising pseudogenes and various non-coding DNA elements, has historically been overlooked due to the assumption of its non-functionality. Recent advances in genomics and epigenetics have overturned this view, revealing that these sequences play crucial roles in genetic regulation, development, disease, and evolution. Pseudogenes, once dismissed as evolutionary relics, are now recognized for their regulatory potential via RNA interference, decoy functions, and epigenetic modulation. Non-coding regions such as long non-coding RNAs (lncRNAs), enhancer RNAs (eRNAs), and other untranslated elements contribute to transcriptional control and chromatin architecture. This review explores the biological functions of these components, their implications in health and disease, and their growing relevance in biomedical research. Furthermore, we examine how emerging technologies such as single-cell sequencing, CRISPR-based editing, and integrative multi-omics are shedding light on the regulatory functions of the dark genome. Despite significant progress, many challenges persist, including functional validation, annotation inconsistency, and interpretation of non-coding variants. This paper aims to synthesize current findings, highlight biomedical applications, discuss limitations, and propose future research directions, emphasizing the need to embrace the dark genome for a more comprehensive understanding of gene regulation and genome complexity.
Keywords:
dark genome
; pseudogenes
; non‐coding DNA
; Gene regulation
; long non‐coding RNAs
; genome evolution
; transcriptomics
1. Introduction
The human genome comprises approximately 3 billion base pairs, yet only about 1.5% codes for proteins. The remaining 98.5%, often referred to as non-coding DNA, was historically regarded as "junk DNA" due to its apparent lack of function. This view has changed dramatically over the past two decades with the advent of high-throughput genomic technologies, which have uncovered the functional complexity and regulatory significance of these regions [1]. Of particular interest is a subset of the non-coding genome collectively termed the "dark genome," encompassing pseudogenes, regulatory elements, and various classes of non-coding RNAs (ncRNAs).
Pseudogenes are genomic DNA sequences similar to functional genes but typically lack the ability to encode proteins due to disabling mutations or lack of transcriptional machinery. However, recent research has shown that some pseudogenes are transcriptionally active and may function in gene regulation through multiple mechanisms, including acting as competing endogenous RNAs (ceRNAs) [2]. Similarly, long non-coding RNAs (lncRNAs), microRNAs (miRNAs), and enhancer RNAs (eRNAs) have emerged as vital regulators of gene expression, development, and cellular differentiation [3]. The dark genome's relevance extends to numerous fields including developmental biology, oncology, and evolutionary genetics. Misregulation of non-coding regions is increasingly implicated in diseases such as cancer, neurological disorders, and cardiovascular conditions [4]. Moreover, evolutionary conservation and selection patterns in non-coding sequences underscore their functional importance across species.
This review explores the components of the dark genome, their regulatory roles, biomedical applications, current challenges, and future research directions, aiming to provide a comprehensive overview of this burgeoning area in genetic science.
2. Core Concepts or Technologies
The central dogma of molecular biology, DNA to RNA to protein, has long shaped our understanding of gene expression. However, emerging evidence has drastically reshaped this paradigm by emphasizing the significance of the "dark genome," which comprises genomic regions that do not code for proteins yet exert considerable regulatory influence on cellular processes. This section delineates the core components of this cryptic yet functional genomic landscape: pseudogenes, long non-coding RNAs (lncRNAs), microRNAs (miRNAs), and enhancer RNAs (eRNAs).
2.1. Pseudogenes: Evolutionary Relics with Functional Potential
Traditionally considered "junk DNA," pseudogenes are genomic sequences that resemble known genes but harbor mutations or deletions rendering them incapable of producing functional proteins, see Figure 1. Pseudogenes can be categorized into three major types:
- Processed pseudogenes: Resulting from retrotransposition events, lacking introns and often flanked by direct repeats.
- Unprocessed pseudogenes: Arise from gene duplication followed by deleterious mutations.
- Unitary pseudogenes: Formed when a functional gene becomes inactivated without duplication.
2.2. Long Non-Coding RNAs (lncRNAs)
lncRNAs are transcripts longer than 200 nucleotides that do not encode proteins. They function through diverse mechanisms including:
- Chromatin remodeling by interacting with epigenetic modifiers like PRC2 (polycomb repressive complex 2) [3].
- Transcriptional interference by directly binding to transcription factors.
- Post-transcriptional regulation, including splicing and mRNA decay.
Their tissue-specific expression patterns and dysregulation in diseases, particularly cancer, underscore their importance in genetic regulation [4].
2.3. Small Regulatory RNAs: miRNAs and siRNAs
MicroRNAs (miRNAs) are ~22 nucleotide long RNAs that suppress gene expression by targeting mRNAs for degradation or translational repression. They are integral to developmental timing, cell cycle regulation, and immune responses [5].
Small interfering RNAs (siRNAs), typically exogenous or experimentally introduced, share a similar mechanism and are utilized in gene silencing technologies [6].
2.4. Enhancer RNAs (eRNAs) and Chromatin Architecture
Enhancer RNAs are transcribed from active enhancer regions and are thought to facilitate enhancer-promoter looping and chromatin remodeling, allowing efficient transcriptional activation [7]. In addition, 3D genome mapping techniques (Hi-C, ChIA-PET) reveal thattopologically associating domains (TADs) andchromatin looping play crucial roles in orchestrating gene expression by spatial organization of non-coding regions [8].
2.5. Technologies Uncovering the Dark Genome
The functional annotation of non-coding regions has been enabled by high-throughput and integrative approaches, see Table 1:
Together, these tools have revolutionized our ability to decode previously unannotated genomic territories and assign regulatory roles to the so-called “junk DNA.”
3. Applications Across Sectors
Understanding the functional implications of non-coding DNA and pseudogenes has revolutionized diverse fields, from biomedical research to agricultural biotechnology and environmental sciences. This section highlights practical applications and emerging innovations driven by the exploration of the dark genome.
3.1. Healthcare and Precision Medicine
a. Biomarkers for Disease Diagnosis
Non-coding RNAs and pseudogenes have emerged as reliable biomarkers for early detection and prognosis of various diseases, particularly cancer. For instance, the pseudogene PTENP1 acts as a ceRNA to regulate PTEN expression in prostate and breast cancer, serving as a potential tumor suppressor [9]. Similarly, HOTAIR, a lncRNA, is overexpressed in breast, colorectal, and liver cancers and correlates with poor prognosis [10].
b. Therapeutic Targets
RNA-based therapeutics targeting non-coding elements are advancing into clinical pipelines:
c. Neurodegenerative and Rare Genetic Disorders
Misregulated non-coding RNAs have been implicated in Alzheimer’s disease, Huntington’s disease, and amyotrophic lateral sclerosis (ALS). For example, BACE1-AS, an antisense RNA, stabilizes BACE1 mRNA, promoting amyloid-beta plaque formation in Alzheimer’s disease [13]. These insights are fueling RNA-based diagnostics and interventions in neurology.
3.2. Agriculture and Crop Engineering
a. Regulatory Elements for Trait Optimization
Crops engineered with specific non-coding RNAs or modified pseudogenes show enhanced resistance to environmental stresses. For instance:
b. Transgene-Free Editing
CRISPR interference systems (CRISPRi) targeting regulatory lncRNAs allow epigenetic modifications in crops without introducing foreign DNA—offering a non-GMO alternative for agricultural enhancement [16].
3.3. Environmental Science and Microbial Genomics
a. Stress Adaptation in Microbial Communities
Metagenomic studies have revealed that non-coding RNAs and pseudogenes play adaptive roles in microbial communities facing extreme environments. In hydrothermal vents and hypersaline lakes, pseudogenes exhibit regulatory activity under stress [17].
b. Environmental Monitoring Using eDNA and lncRNA Signatures
Environmental DNA (eDNA) profiling increasingly includes analysis of non-coding RNA transcripts to track ecosystem health and biodiversity. Changes in microbial lncRNA profiles can serve as early indicators of environmental perturbations such as pollution or climate shifts [18].
3.4. Industry and Synthetic Biology
a. Designing Regulatory RNA Circuits
Synthetic biology has begun leveraging non-coding RNAs to build programmable RNA-based regulatory circuits. lncRNAs are being engineered to control gene expression in microbial cell factories producing biofuels, bioplastics, and pharmaceuticals [19].
b. Biocomputing and RNA Logic Gates
4. Challenges and Limitations
While the exploration of the dark genome holds transformative potential across biology and biotechnology, significant obstacles impede the full realization of its utility. These challenges span technical, interpretative, and translational domains.
4.1. Annotation and Functional Characterization
Challenge: Many pseudogenes and non-coding RNAs lack functional annotation in existing genome databases.
- The human genome contains over 15,000 pseudogenes and tens of thousands of lncRNAs, yet less than 5% have confirmed biological roles [21].
- Tools like ENCODE and FANTOM5 have improved mapping, but experimental validation remains limited.
Potential Solutions:
- Develop standardized functional assays for non-coding RNA screening.
- Incorporate machine learning to predict function from sequence and structure [22].
4.2. Context-Dependent Activity and Tissue Specificity
Challenge: Non-coding elements often exhibit cell type- and condition-specific expression, complicating generalization.
- For instance, lncRNAs such as NEAT1 and MEG3 may be oncogenic in one tissue and tumor-suppressive in another [23].
Potential Solutions:
- Employ single-cell RNA sequencing to resolve context-specific roles.
- Use conditional knockout models for in vivo validation.
4.3. Genetic Redundancy and Compensation
Challenge: Functional redundancy across pseudogenes and ncRNAs can mask phenotypic consequences in knockout studies.
- Loss of a pseudogene may be buffered by the presence of homologous sequences, making loss-of-function phenotypes difficult to interpret [24].
Potential Solutions:
- Use multiplex CRISPR systems to knock out entire gene families or ncRNA clusters simultaneously.
- Apply synthetic lethality screens to uncover dependencies.
4.4. Translational and Therapeutic Hurdles
Challenge: Delivering ncRNA-based therapeutics remains difficult due to:
- Instability in circulation.
- Off-target effects and poor tissue-specific delivery.
- Potential Solutions:
- Develop RNA stabilization chemistries and ligand-targeted delivery systems (e.g., aptamer-conjugates).
- Apply exosome-based delivery platforms for precision targeting [25].
4.5. Ethical and Regulatory Ambiguities
Challenge: Genome editing targeting non-coding regions poses ethical concerns and lacks clear regulatory pathways.
- Modifying enhancers or pseudogenes may have unintended long-range effects on gene expression [26].
- Potential Solutions:
- Introduce predictive modeling frameworks to simulate genome-wide effects before interventions.
- Promote international bioethical consensus on non-coding genome editing. See Table 3.
5. Future Directions
The burgeoning interest in the non-coding genome promises to reshape the future of biology and medicine. As research tools become more precise and computational models more predictive, novel strategies are emerging to decode and harness the functional roles of pseudogenes and non-coding RNAs.
5.1. AI-Powered Functional Annotation
The integration of artificial intelligence (AI) and deep learning has revolutionized genome interpretation. Predictive models like DeepSEA and Basenji are being employed to:
- Forecast the regulatory impact of non-coding variants.
- Infer enhancer-promoter interactions and ncRNA function from sequence alone [27].
In the future, AI-driven annotation pipelines will accelerate the identification of disease-associated regulatory RNAs and streamline biomarker discovery.
5.2. CRISPR-Based Functional Genomics in Non-Coding Regions
The use of CRISPR tools is evolving beyond protein-coding genes:
- CRISPRi (interference) and CRISPRa (activation) allow targeted regulation of lncRNAs, pseudogenes, and enhancers without altering DNA sequence.
- CRISPR tiling screens offer high-resolution maps of functional non-coding elements in disease loci [28].
These approaches will be pivotal in assigning biological meaning to vast intergenic territories.
5.3. Single-Cell and Spatial Transcriptomics
Recent innovations enable in situ mapping of non-coding RNA expression:
- Single-cell RNA-seq (scRNA-seq) reveals lncRNA heterogeneity across individual cells.
- Spatial transcriptomics captures tissue-specific expression of regulatory elements, vital for developmental biology and cancer studies [29].
These tools will clarify how the dark genome orchestrates spatiotemporal gene regulation.
5.4. Multi-Omics and Systems Biology Approaches
Combining epigenomics, transcriptomics, proteomics, and metabolomics enables a systems-level understanding of ncRNA-mediated control. This integrative view is essential for:
- Dissecting complex regulatory networks.
- Modeling genotype-to-phenotype transitions driven by non-coding elements [30].
Future studies will increasingly adopt multi-omics platforms for comprehensive analysis of regulatory landscapes.
5.5. Synthetic Biology and ncRNA Engineering
Synthetic biology is now applying ncRNAs as programmable tools:
- Engineered lncRNAs can act as scaffolds, decoys, or sponges in synthetic gene circuits.
- Riboregulators—RNA-based switches—are being used to control gene expression in response to environmental cues [31].
Such synthetic constructs will support the development of smart therapeutics and biosensors.
5.6. Clinical Translation and Personalized Medicine
Non-coding variants are gaining attention in genome-wide association studies (GWAS) and personalized risk prediction. The next decade may see:
- Routine inclusion of pseudogene and lncRNA panels in diagnostic assays.
- Personalized therapies targeting individual ncRNA profiles for precision oncology and neurology.
5.7. Ethical Frameworks and Governance
With increasing power to edit and interpret the dark genome, there arises a need for robust ethical frameworks. Future directions include:
6. Conclusions
The historical neglect of non-coding and pseudogenic sequences as mere "junk" or evolutionary debris has yielded to a deeper appreciation of their vast and nuanced regulatory roles. As demonstrated throughout this review, the dark genome—encompassing pseudogenes, long and small non-coding RNAs, and regulatory intergenic elements—plays a central part in orchestrating gene expression, maintaining genomic architecture, and modulating developmental, physiological, and pathological processes.
Breakthroughs in transcriptomics, chromatin profiling, and genome editing have illuminated the functional relevance of these elements, with pseudogenes acting as competitive endogenous RNAs and lncRNAs modulating transcription, splicing, and chromatin state. This non-coding machinery is increasingly implicated in cancer, neurodegeneration, metabolic syndromes, and microbial adaptation.
Beyond human health, the implications of dark genome research span agriculture, synthetic biology, and environmental science—paving the way for transgene-free crop improvements, biocomputational circuits, and novel biosensors.
However, despite the promise, challenges persist. The context-specificity of expression, redundancy, limited annotation, and ethical uncertainties around editing non-coding DNA demand a balanced approach, integrating technical innovation with responsible governance.
Looking forward, the convergence of artificial intelligence, CRISPR technologies, spatial transcriptomics, and multi-omics systems will catalyze a new era of functional genomics. This will not only demystify the regulatory logic encoded in the dark genome but will also translate into clinical, ecological, and industrial innovations with far-reaching impact.
The imperative now is to continue investing in interdisciplinary research, open genomic data repositories, and ethical consensus frameworks to ensure that the dark genome is not only mapped but meaningfully understood and responsibly harnessed.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Data Availability Statement
No new datasets were generated or analyzed during this study. All data supporting this review are derived from previously published sources, which have been appropriately cited.
Acknowledgments
The author gratefully acknowledges the support and resources provided by Prince Sattam Bin Abdulaziz University, particularly the Common First Year and Basic Science Department. Appreciation is extended to colleagues and peers whose insights and feedback enriched the scope of this review. The author also thanks the global scientific community whose innovative research in synthetic biology, nanotechnology, and environmental DNA provided the foundation for this work. Special recognition is given to the open-access platforms and databases that facilitated comprehensive literature retrieval essential for the development of this article.
AI Declaration
No artificial intelligence (AI) tools or automated writing assistants were used in the research, drafting, or editing of this manuscript. The content, including the literature review, analysis, and writing, was entirely produced by the authors. All conclusions and interpretations are based on human expertise, critical evaluation of the literature, and independent scholarly work.
Conflict of Interest
The author declare no conflicts of interest related to this study. No competing financial interests or personal relationships could have influenced the content of this research review.
References
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: what is functional and what is junk? Front Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature. 2010, 465, 1033–1038. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu Rev Biochem.
- Esteller, M. Non-coding RNAs in human disease. Nat Rev Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: target recognition and regulatory functions. Cell. 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Kim, T.K.; Shiekhattar, R. Architectural and functional commonalities between enhancers and promoters. Cell. 2015, 162, 948–959. [Google Scholar] [CrossRef]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 2012, 485, 376–380. [Google Scholar] [CrossRef]
- Poliseno, L.; Pandolfi, P.P. PTENP1 and the ceRNA hypothesis: an intricate balance. Methods.
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Arun, G.; Diermeier, S.; Spector, D.L. Therapeutic targeting of long non-coding RNAs in cancer. Trends Mol Med. 2018, 24, 257–277. [Google Scholar] [CrossRef]
- Liu, S.J.; Horlbeck, M.A.; Cho, S.W.; Birk, H.S.; Malatesta, M.; He, D.; et al. CRISPRi-based genome-scale identification of functional long noncoding RNA loci in human cells. Science. 2017, 355, eaah7111. [Google Scholar] [CrossRef]
- Faghihi, M.A.; Modarresi, F.; Khalil, A.M.; Wood, D.E.; Sahagan, B.G.; Morgan, T.E.; et al. Expression of a noncoding RNA is elevated in Alzheimer’s disease and drives rapid feed-forward regulation of β-secretase. Nat Med. 2008, 14, 723–730. [Google Scholar] [CrossRef]
- Si-Ammour, A.; Windels, D.; Arn-Bouldoires, E.; Kutter, C.; Ailhas, J.; Meins, F.; et al. miR393 and secondary siRNAs regulate expression of the auxin-related gene TIR1 and AFB2 in Arabidopsis. Plant J. 2011, 68, 452–461. [Google Scholar]
- Ariel, F.; Romero-Barrios, N.; Jégu, T.; Benhamed, M.; Crespi, M. Battles and hijacks: noncoding transcription in plants. Trends Plant Sci. 2015, 20, 362–371. [Google Scholar] [CrossRef]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; et al. A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Valera, F.; Martin-Cuadrado, A.B.; López-Pérez, M.; García-Heredia, I. Pseudogenes in microbial genomes: theory and practice. Res Microbiol. 2016, 167, 664–673. [Google Scholar]
- Odah, M.A.A. Temporal dynamics of environmental DNA (eDNA) as early-warning indicators of climate-driven ecosystem shifts in diverse Saudi habitats. Preprints. 2025. [Google Scholar]
- Odah, M.A.A. Ultra-short DNA satellites as environmental sensing elements in soil microbiomes: a frontier review. Preprints. 2025. [Google Scholar]
- Odah, M.A.A. Programmable DNA devices: the next generation of living sensors in agriculture, health, and the environment. Preprints. 2025. [Google Scholar]
- Odah, M.A.A. Photosynthetic reprogramming enhancing carbon fixation in crops through synthetic biology. Preprints. 2025. [Google Scholar]
- Odah, M.A.A. Microbiota-immune-brain crosstalk: synthetic biology solutions for neuroinflammatory disorders. Preprints. 2025. [Google Scholar]
- Pei, B.; Sisu, C.; Frankish, A.; Howald, C.; Habegger, L.; Mu, X.J.; et al. The GENCODE pseudogene resource. Genome Biol. 2012, 13, R51. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Troyanskaya, O.G. Predicting effects of noncoding variants with deep learning–based sequence model. Nat Methods. 2015, 12, 931–934. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Rice, K.; Wang, Y.; Chen, W.; Zhong, Y.; Nakayama, Y.; et al. Maternally expressed gene 3 (MEG3) noncoding RNA: isoform structure, expression, and functions. Nucleic Acids Res. 2010, 38, 4740–4751. [Google Scholar]
- Schmitz, J.F.; Zimmer, F.; Bornberg-Bauer, E. Mechanisms of transcription factor evolution in Metazoa. Nucleic Acids Res. 2016, 44, 6287–6297. [Google Scholar] [CrossRef]
- Li, Z.; Rana, T.M. Therapeutic targeting of microRNAs: current status and future challenges. Nat Rev Drug Discov. 2014, 13, 622–638. [Google Scholar] [CrossRef]
- Cwiklinska, M.; Kolb, G.; Michalakis, S. Non-coding RNAs in inherited retinal diseases. Cell Tissue Res. 2021, 386, 435–450. [Google Scholar]
- Kelley, D.R.; Reshef, Y.A.; Bileschi, M.; Belanger, D.; McLean, C.Y.; Snoek, J. Sequential regulatory activity prediction across chromosomes with convolutional neural networks. Genome Res. 2018, 28, 739–750. [Google Scholar] [CrossRef]
- Sanjana, N.E.; Wright, J.; Zheng, K.; Shalem, O.; Fontanillas, P.; Joung, J.; et al. High-resolution interrogation of functional elements in the noncoding genome. Science. 2016, 353, 1545–1549. [Google Scholar] [CrossRef]
- Stahl, P.L.; Salmen, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science. 2016, 353, 78–82. [Google Scholar] [CrossRef]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Chappell, J.; Takahashi, M.K.; Lucks, J.B. Creating small transcription activating RNAs. Nat Chem Biol. 2015, 11, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Iyer, E.P.; Salvado, O.; Gaikwad, S. A policy framework for genome editing. Nat Biotechnol. 2017, 35, 484–486. [Google Scholar]
Figure 1.
Structural Comparison Between Functional Genes and Pseudogenes: Key Mutational Disruptions.
Figure 1.
Structural Comparison Between Functional Genes and Pseudogenes: Key Mutational Disruptions.
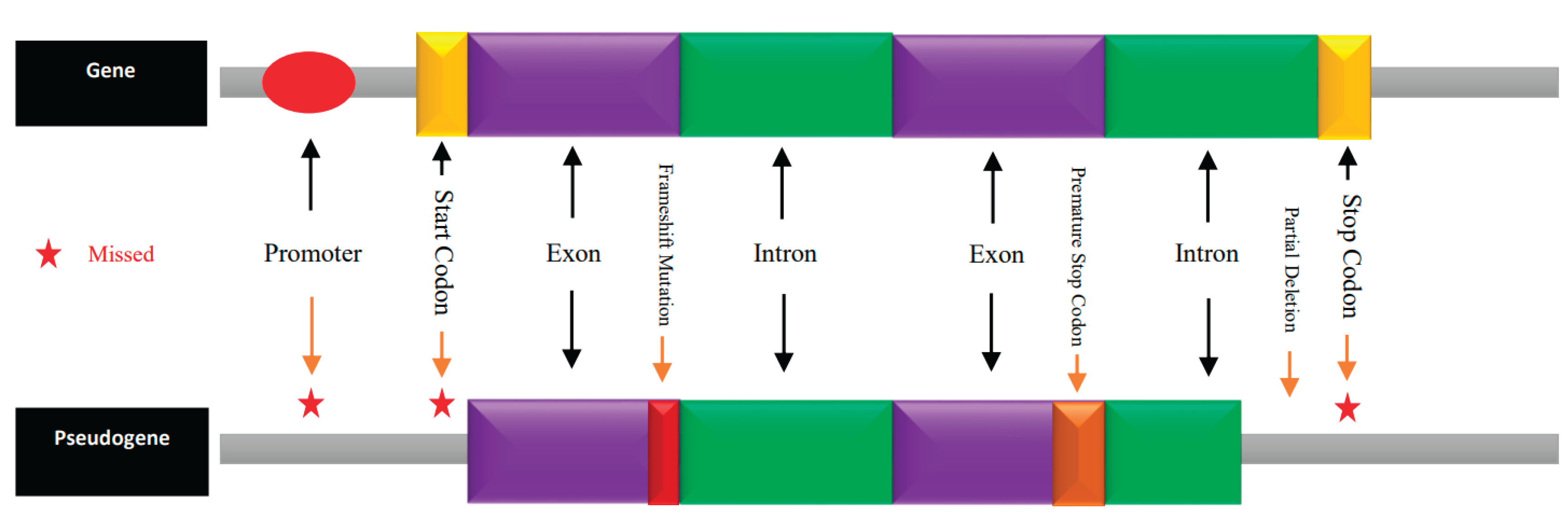

Table 1.
Key Technologies for Functional Annotation of the Dark Genome.
| Technology | Application |
|---|---|
| RNA-Seq | Transcriptome profiling of non-coding RNAs |
| ChIP-Seq | Identifying transcription factor binding sites |
| ATAC-Seq | Mapping chromatin accessibility |
| Hi-C/3C | Studying 3D genome architecture |
| CRISPR interference (CRISPRi) | Functional dissection of non-coding elements |
Table 2.
Applications of the Dark Genome Across Sectors.
| Sector | Application Example | Key Molecule |
|---|---|---|
| Healthcare | PTENP1 in tumor suppression | Pseudogene |
| Agriculture | miR393 enhances drought resistance in rice | miRNA |
| Environment | eDNA-based stress markers in marine microbiomes | lncRNAs |
| Industry | RNA-based logic gates for biosensors | Synthetic lncRNAs |
Table 3.
Key Challenges and Mitigation Strategies.
| Challenge | Description | Proposed Solution |
|---|---|---|
| Poor functional annotation | Limited understanding of roles of ncRNAs/pseudogenes | Functional assays, ML-based prediction |
| Context specificity | Varying function across tissues | Single-cell transcriptomics |
| Redundancy and compensation | Masking of phenotypes by similar elements | Multiplexed CRISPR screens |
| Therapeutic delivery limitations | Instability and off-target effects | RNA modifications, targeted delivery vehicles |
| Ethical and regulatory uncertainties | Editing regulatory DNA with unknown consequences | Simulation models, ethical frameworks |
Table 4.
Emerging Trends in Ethical Governance and Technological Advances in Dark Genome Research.
| Domain | Emerging Direction |
|---|---|
| AI & Bioinformatics | Predictive ncRNA function and variant annotation |
| CRISPR Technology | Targeted manipulation of non-coding elements |
| Single-Cell Biology | Context-specific ncRNA mapping |
| Multi-Omics | Integrated regulatory network modeling |
| Synthetic Biology | Engineered lncRNAs for smart applications |
| Clinical Translation | Diagnostic panels & RNA-targeting therapies |
| Ethics & Governance | Non-coding genome editing regulations |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



