Submitted:
28 June 2025
Posted:
30 June 2025
Read the latest preprint version here
Abstract
Antibiotic resistance (ABR) has become a serious threat to humans. the One Earth-One Health (OE-OH) concept based on the self-regulation of ecosystems was proposed for preventing AMR in 2024. To define and clarify this concept, here the dual mutation pattern of proactive evolution (most) and passive selection (few) for ABR was put forward based on the principle of parsimony (Occam’s Razor), involving the understanding integration of Lamarck theory, Darwinian evolution, and Niche construction theory. Subsequently, the theoretical logic of the OE-OH concept was clarified based on the strong self-regulatory ability and perpetual reconstruction of ecosystems, and a basic mathematical model was first presented for the amount of antibiotic resistance genes (ARGs) renewing with the evolution of the Earthʹs ecosystem. From this concept, it was deduced that ABR and ARGs emerge 3.5 billion years ago after microbial ecosystems formed and exist in every corner on the Earth prior to the production and use of antibiotics by humans. Moreover, the self-regulation of the Earth's ecosystem on the generation, spread, weakening and elimination of ABR and ARGs, according to various abiotic and biotic factors, was sorted out for understanding the OE-OH concept. Regarded as an intervention on ecosystems, the impact of antibiotic use especially overuse and abuse on this regulation have led to a sharp increase in ARGs and a worry beyond the self-regulation of ecosystems, and some important measures from the OE-OH concept are suggested for offsetting the ecosystemic imbalance in ARG generation and elimination, combatting ABR. Among them, the simulation and utilization to the self-regulation mechanisms of ecosystems from the OE-OH concept are emphasized, together with avoiding the excessive aggregation of population, and encouraging antibiotics in combination with plant antimicrobial ingredients.
Keywords:
OE-OH
; Mutation
; ecosystem
; antimicrobial agent
; antibiotic
; microbe
; plant
; climate
; environment
; combination
; parsimony
; Occam
; Lamarck
; Niche
; Darwin
1. Introduction
Antibiotic resistance (ABR) has become a serious threat to global public health and economic development, and the COVID-19 pandemic has further exacerbated this crisis [1,2,3]. It was showed that ABR has rapidly spread from various settings of antibiotic use into ambient environments [4,5], and antibiotic resistance genes (ARGs) can be widely detected in various waterbody and soil environments [4,5,6,7], even in the north slope of Mount Everest [8]. Given these threats, World Health Organization (WHO) predicted that antibiotic resistance would cause 10 million deaths annually if no action is taken until 2050 [9,10]. Subsequently, FAO, UNEP, WHO, and WOAH have developed the One Health Joint Plan of Action (2022–2026) (OH JPA), and appeal working together for the health of humans, animals, plants and the environment [7,11].
As we known, the use of antibiotics, especially their overuse and abuse, has led to the crisis of ABR. However, it is debatable whether these reports can reflect the actual situation of ABR. On the one hand, influenced by the interest and focus of research, many studies pay more attention to the spread and evolution of ABR among microorganisms and/or in the ambient environment, while neglecting or studying less on the weakening and elimination effects on ABR and ARGs from the self-regulatory ability of the Earth's ecosystem [12,13,14,15], including the interactions among human, plants, animals, microorganisms and environment, and the functional redundancy and removal of ARGs [4,16,17,18]. On the other hand, it is also worth considering whether the detection of ARGs means that ABR has been transmitted to the sampling location. These two aspects have prompted a deeper reflection for the strategies and measures to address ABR. Based on the discoveries of various laws on drug combination preventing ABR [19,20] and the effects of many plant metabolites reversing ABR [12,21,22], the One Earth-One Health (OE-OH) concept for preventing ABR was proposed at the 6th International Caparica Conference in Antibiotic Resistance 2024 (IC2AR 2024) [12,23].
The One Health (OH) concept points out that ABR involves multiple aspects such as humans, animals, plants and the environment, and emphasizes the role of humans in addressing ABR. Different from this, the OE-OH concept emphasizes the self-regulation of the Earth's ecosystem and its sub-ecosystems to ABR, especially giving equal consideration to the generation, spread, weakening and elimination of ABR by the ecosystem, and regards the production and use of antibiotics as an intervention to ABR in the ecosystem. Here, the theoretical logic of this concept, together with some amazing and innovative prospective, reasoning, deduction, and induction related to ABR and ARGs from the OE-OH concept, are presented as follows. Based on these, some distinctive suggestions for combatting ABR are subsequently put forward and elaborated.
2. The OE-OH Concept
2.1. Dual Mutation Pattern of Bacterial Resistance
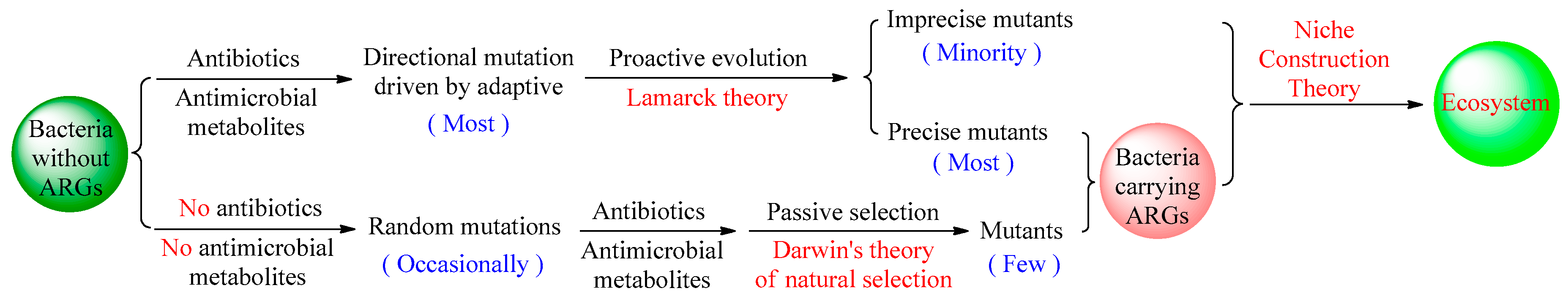
Based on the principle of parsimony (Occam’s Razor) [24,25], the mutation theory of bacterial resistance can be reconsidered, and we believe that bacterial resistance mainly adopts proactive evolution driven by adaptive, which aims to avoid the unnecessary energy and resource consumption associated with random mutations. This is consistent with Lamarck theory which was also supported by recent research [26]. However, there may also be minor imprecise mutants during the process of proactive resistance mutations. The mutants carrying ARGs in the occasional and non-adaptive random mutations of microbes can also be passively screened by natural selection according to Darwinian evolution. From these, the mutants carrying ARGs originate from two types of mutations shown on Figure 1. This dual mutation pattern of proactive evolution (mainly) and passive selection (occasionally) in bacterial resistance is extremely consistent with Niche construction theory being related to ecosystems [27,28].
Figure 1.
Dual mutation pattern of bacterial resistance through proactive evolution and passive selection.
Figure 1.
Dual mutation pattern of bacterial resistance through proactive evolution and passive selection.
2.2. Theoretical Logic of the OE-OH Concept Based on Ecosystems
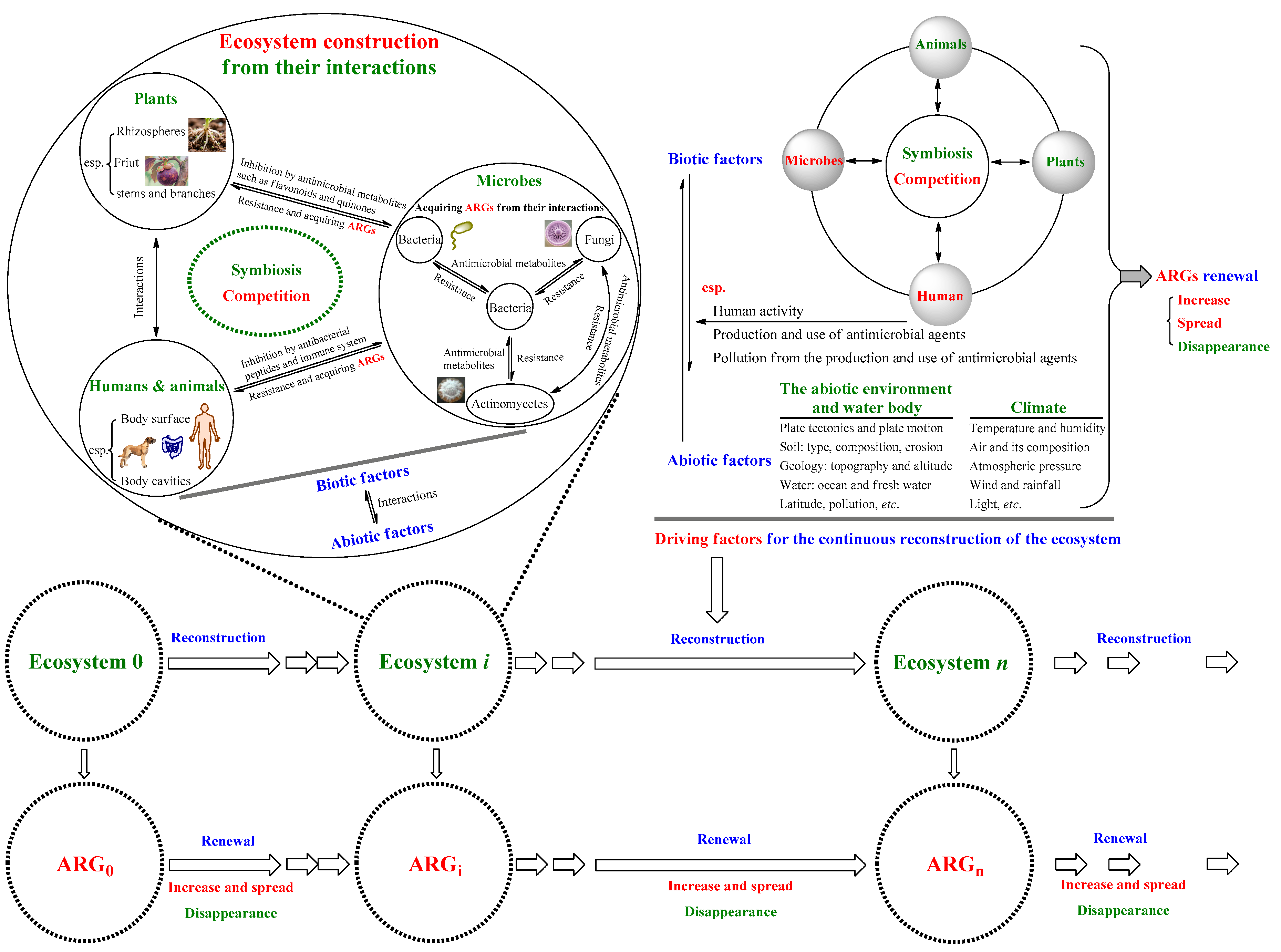
Inspired by the dual mutation pattern on Figure 1, it was deduced that antimicrobial metabolites produced by microbes (some are termed antibiotics now), microbial resistance and ARGs should emerge along with the formation of the microbial ecosystem, since they are generated from the competition among microorganisms in ecosystems [29,30,31], renewing with the reconstructions of ecosystems [32]. Namely, bacteria carrying ARGs likely emerged when some microbial ecosystems were formed 3.5 billion years ago [33,34], and have continuously spread and renewed until now (Figure 2). Simultaneously, the Earth including soil, various environments and all the organisms on it can be regarded as a giant ecosystem which is composed of countless sub-ecosystems sharing similar mechanisms of operation. Therefore, here the strong self-regulatory ability and perpetual reconstruction of ecosystems, especially of the Earth's one [13,14,15], is regarded as the theoretical logic of the OE-OH concept, and various aspects related to microbial resistance can be reexamined from this concept. Moreover, although the proactive mutation is a main pattern of microbial resistance to antibiotics, a large number of antibiotic-resistant microbes can still be discovered before contacting with antibiotics, as there are a large number of ARG-carrying pathogens in the Earth's ecosystem after accumulated over a long period of time.
Figure 2.
Schematic for the theoretical logic of the OE-OH concept based on ecosystems. The ecosystem can be the whole Earthʹs ecosystem or its various sub-ecosystems; ecosystem 0, i, and n indicate the ecosystem is at different time points during its evolution, and their antibiotic resistance genes (ARGs) correspondingly renew with the continuous reconstruction of the ecosystem.
Figure 2.
Schematic for the theoretical logic of the OE-OH concept based on ecosystems. The ecosystem can be the whole Earthʹs ecosystem or its various sub-ecosystems; ecosystem 0, i, and n indicate the ecosystem is at different time points during its evolution, and their antibiotic resistance genes (ARGs) correspondingly renew with the continuous reconstruction of the ecosystem.
2.3. Basic Mathematical Model for the ARGs Renewing with the Ecosystem
The OE-OH concept regards the Earth as a giant ecosystem which is composed of various sub-ecosystems sharing similar mechanisms of operation. The amount of ARGs in an ecosystem can be presented as a basic mathematical model.
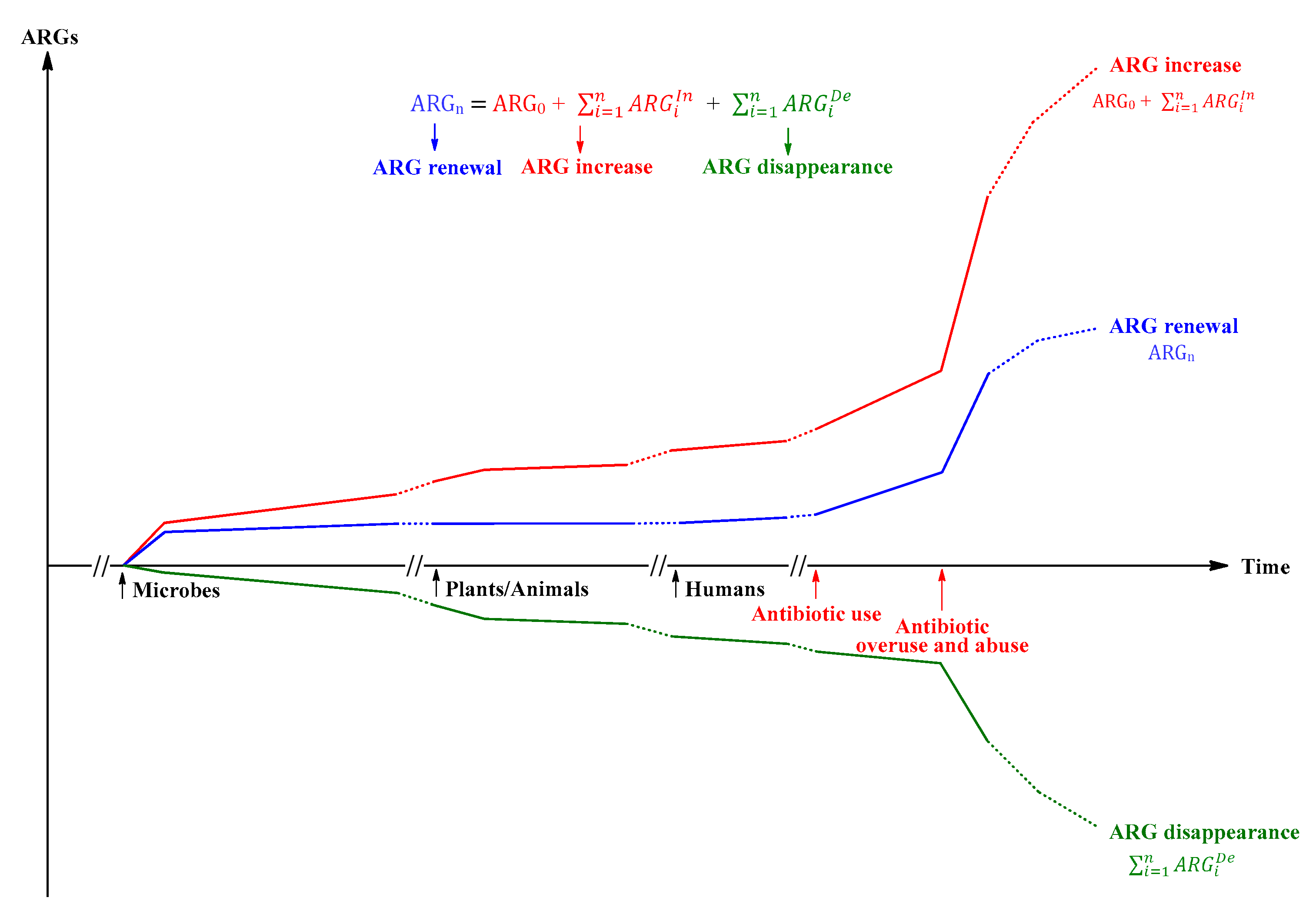
ARGi = ARG0 + (ARG1In + ARG2In +……+ARGiIn +……+ARGnIn) + (ARG1De + ARG2De +……+ ARGiDe+……+ARGnDe)
Where ARGi and ARGn are the amount of ARGs at two time points (Figure 3) of the ecosystem which can be the Earth′s one or its various sub-ecosystem, and time point i can be equal to n; ARG0 can be the amount of ARGs at any time point while time points i and n are more than or equal to time point 0, and especially the amount of ARG0 is zero before the microbial ecosystem emerged on the Earth; ARGIn is the increased amount of ARGs in the ecosystem, for example ARG1In is that from time point 0 to 1; ARGDe (defines as negative value) is the decreased amount of ARGs in the ecosystem, for example ARG1De is that from time point 0 to 1; is the sum of the increased amount of ARGs from time point 1 to n, and is the sum of the decreased amount of ARGs from time point 1 to n.
Figure 3.
Schematic of the amount of ARGs (ARGs, y) in an ecosystem changing with different evolutionary periods (Time, x) according to the basic mathematical model. The ecosystem can be the whole Earthʹs ecosystem or its various sub-ecosystems.
Figure 3.
Schematic of the amount of ARGs (ARGs, y) in an ecosystem changing with different evolutionary periods (Time, x) according to the basic mathematical model. The ecosystem can be the whole Earthʹs ecosystem or its various sub-ecosystems.
3. ARG Analyses from the OE-OH Concept Based on Ecosystems
3.1. ARGs Emerging Before Humans and Existing Everywhere
Antimicrobial metabolites are a class of natural products generated from the competition for ecological niches among microorganisms in ecosystems or facing survival stress [29,30,31]. From the evolutionary history of the Earth, it is known that microbes emerge about 3.5 billion years ago, and various ecosystems containing microbial communities have already formed at that time [33,34]. After a long period of evolution, there are sufficient reasons to infer that the structural skeletons of most clinical antibiotics can be biosynthesized by environmental microorganisms. This can be confirmed by the fact that the structural skeletons of clinical antibiotics were mostly discovered from soil microorganisms [29]. In other words, microorganisms carrying the genetic information biosynthesizing the structural skeletons of most clinical antibiotics, including microbial strains belonging to the same or different genera and species, are widely distributed in various sub-ecosystems of the Earth. Even synthetic quinolone antimicrobial agents have similar structural frameworks in nature, such as plant-derived flavonoids and benzofuranones [12,35]. This is also a reason that natural products with antibacterial activity can be continuously discovered from environmental microorganisms. Likely, the genetic information of microorganisms producing these natural products has existed on the Earth before the emergence of humans [36], while only was discovered and excavated after entering into the 20th century.
It is reported that the pathogenic microorganisms nowadays come from the soil [37,38]. Therefore, clinical pathogens should be widely distributed in various ecosystems of the Earth, also likely emerged prior to the existence of humans (Figure 3). Since antimicrobial metabolites generated before billions of years ago, it is reasonable to deduce that the ARGs of pathogenic microorganisms in response to the survival stress from antimicrobial metabolites (some now as antibiotics) produced by other environmental ones may had been already generated at that time, due to the competition among microorganisms in ecosystems.
Along with the evolution and reconstruction of the Earthʹs ecosystem and its various sub-ones driven by various biotic and abiotic factors (Box 1) [39], new ARGs have been constantly generating, and all ARGs have been constantly spreading and renewing everywhere on the Earth (Figure 3). Therefore, it can be also inferred that same or similar microorganisms that produce the structural skeleton of a certain antibiotic, together with some pathogens carrying corresponding ARGs, are located in certain ecosystems. When some antibiotics were isolated from the soil or ocean microbes, actually they had already been in contact with some pathogens during the evolution of the Earth's ecosystem, and these pathogens have also developed resistance to those antibiotics through proactive adaptive mutation and acquired heritable ARGs. This is likely a reason that sometimes ARG-carrying pathogens can be correspondingly discovered soon after some antibiotics had been launched on the market [40], or before they seemingly had no contact with the corresponding antibiotics, even before these antibiotics approved [41]. Namely, these pathogens in fact had been in contact with corresponding antibiotics long ago. Therefore, it can be foreseen that ARGs can not only be found on Mount Everest and the Mariana Trench, but also in the Antarctica and Arctic of the Earth. These are also supported by recent researches [42,43,44,45,46], and means that ARGs are likely detected in different degrees from every ecological corner of the Earth containing microbes if the detection method is enough sensitive. Moreover, a certain amount of ARGs should contain in various ecosystems and on the Earth prior to the production and use of antibiotics by humans (Figure 3).
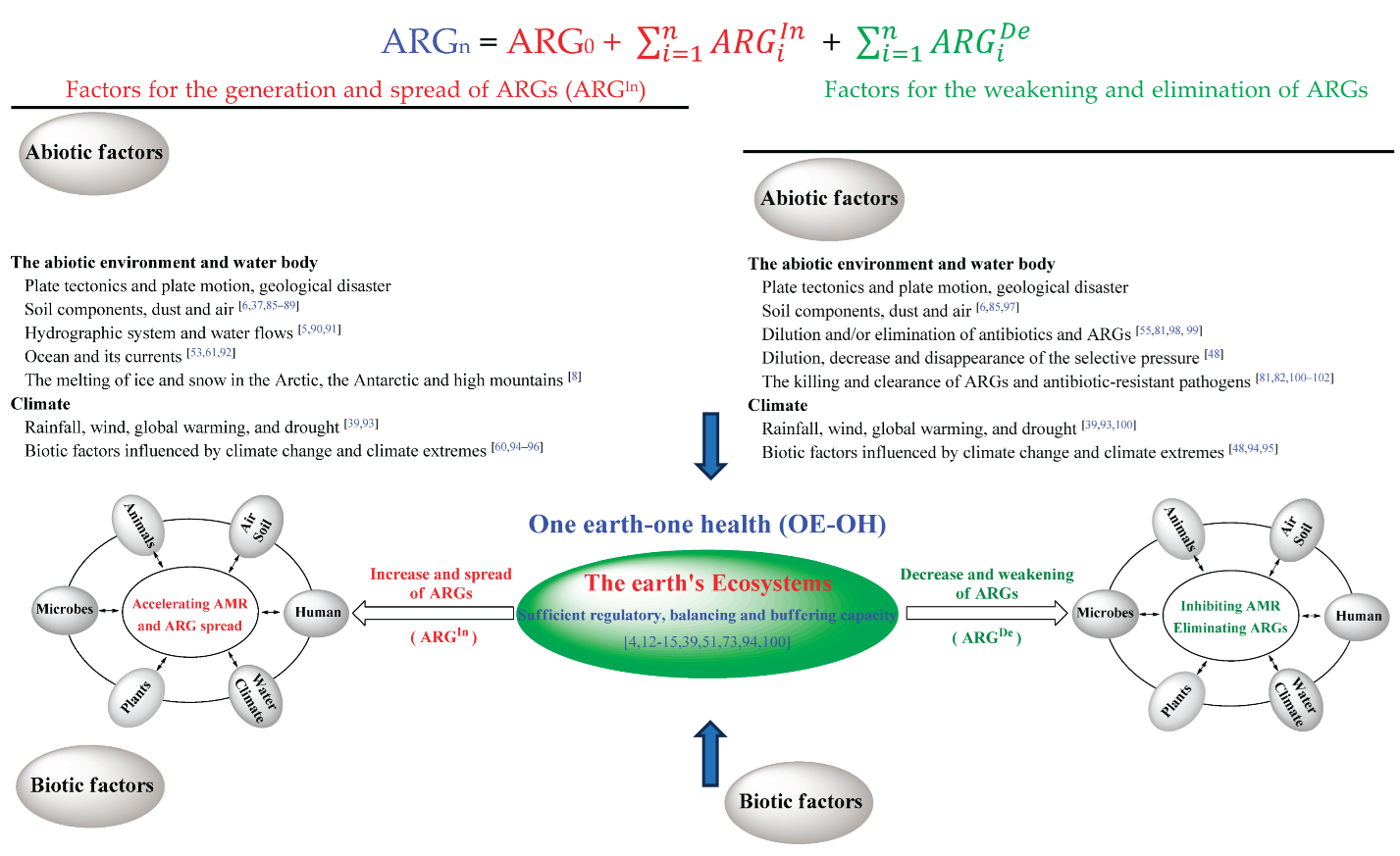
Box 1. The OE-OH concept based on the self-regulation of the Earth's ecosystem for ABR and ARGs.

3.2. ARGs by the Self-Regulation of Ecosystems Before the Industrial Production and Use of Antibiotics
After microbes emerged on the earth, various ecosystems comprising microbial communities have been gradually formed, and continuously evolved along with the incorporation of new organisms and the elimination of existing ones. During the evolution of the Earth, the information of ARGs have continuously renewed with the unceasing reconstruction of ecosystems, including the generation and spread of new ARGs and the weakening and elimination of existing ones, and which was also supported by recent researches [4,6,47,48,49,50,51]. As shown in Box 1, all biotic and abiotic factors, such as abiotic environment, water body, climate, microbes, plants, animals and humans, can impact the renewals of ARGs in the whole Earth′s ecosystem including its various sub-ecosystems. Owing to the sufficient self-regulation, self-balancing and buffering capacity of the Earth′s ecosystem [12,13,14,15], the evolution of ABR and ARGs continuously maintains a balanced and controllable state before the industrial production and use of antibiotics by humans. However, the situation is becoming increasingly worrying due to the extensive use of antibiotics, especially their overuse and abuse in clinical settings, livestock, poultry farming and aquaculture, as well as the resulting widespread dissemination. This has been also reflected in the rapid increase of ABR and the swift spread of ARGs after the industrial production and use of antibiotics.
3.3. Impact of Antibiotic Use on ARGs by the Self-Regulation of Ecosystems
In the 20th century, many secondary metabolites with antimicrobial activities at low concentrations were discovered from environmental microorganisms especially actinomycetes, fungi and bacteria, in various habitats of land, ocean and the body surface/feces of animals and humans [29,31]. Some of these metabolites and their derivatives have been developed as clinical antibiotics. In fact, the genetic information of microorganisms producing these antibiotics, together with the corresponding ARGs carried by pathogens resistant to these antibiotics, is already generated long ago, and can be considered as the products of competition among various microorganisms during the evolution of the Earth. Although most of the clinical antibiotics currently in use are the structural derivatives of these natural antibiotics, they share similar structural skeletons with corresponding naturally sourced antibiotics. Therefore, the information of their corresponding ARGs carried by pathogens has existed in nature long ago. Moreover, pathogens carrying these resistance genes have also been spreading everywhere in the globe, including that some have spread through soil and some have colonized in certain parts of the human body [37].
From the above, the widespread use of antibiotics, such as in medical activities, livestock, poultry farming and aquaculture, can be considered as a human intervention on the evolution of ARGs in the whole Earth's ecosystem [52]. Besides that the general impact of human activities together with the interactions between human body and microbes promote the spread of ARGs (Box 1) before the use of antibiotics, the following main activities related to the industrial production and use of antibiotics by humans have led to a sharp increase in ABR and the rapid spread of ARGs: (1) The use, overuse and abuse of antibiotics in medical activities, livestock, poultry farming, and aquaculture, etc. [4,48,53]; (2) The pollution of the Earth's environment by wastewater discharged from the settings of antibiotic production and use [3,4,7,53,54,55,56,57]; (3) The excessive aggregation of population and the large amounts of domestic sewage under the circumstance in the overuse and abuse of antibiotics [4,58,59]; (4) The impacts of other human activities on all biotic and abiotic factors that generate and spread ARGs in the Earth′s ecosystem [60,61,62,63,64,65].
Theoretically, the wider the application of antibiotics and the greater their usage, the stronger the intervention on ecosystems. When the intervention intensity keeps within a certain and controllable range, the Earth's ecosystem and its various sub-ecosystems have sufficient self-regulation ability to restore or reconstruct a new balance (Box 1). But if the intervention intensity exceeds the self-regulation ability of ecosystems, the balance of ecosystems will inevitably be disrupted, endangering larger ecosystems and even the whole Earth's ecosystem [66]. Therefore, our efforts in discovering new antibiotics seem to have the potential to solve the problem of antibiotic shortage caused by ABR, however they are only a passive defense even if the development of new antibiotics can be accelerated with the assistance of artificial intelligence (AI) [67]. Although the microbial resistance to existing antibiotics can be predicted by AI and a proactive defense is made one day, the struggle between humans and microbes is at most evenly matched. More significantly, the damage to various ecosystems caused by the overuse and abuse of antibiotics, together with the impact and deterioration of new ecosystem reconstruction on human living conditions, should arouse sufficient attention in this struggle.
Therefore, if the overuse and abuse of antibiotics are not controlled, the more the approval and application of new antibiotics, the faster the spread of ABR and ARGs. This will lead to the larger and more ecosystems centered on the application environment are continuously destroyed, and various ones containing more pathogens carried with ARGs will be restructured, ultimately endangering the survival and development of humans. So, scientific and reasonable measures must be taken to control the spread of ABR and ARGs, for maintaining the balance of the Earth′s ecosystem and its various sub-ecosystems.
Figure 4.
Analysis schematic of some actions should be taken for combating the ABR from the OE-OH concept, based on the self-regulation and balancing capacity of ecosystems.
Figure 4.
Analysis schematic of some actions should be taken for combating the ABR from the OE-OH concept, based on the self-regulation and balancing capacity of ecosystems.
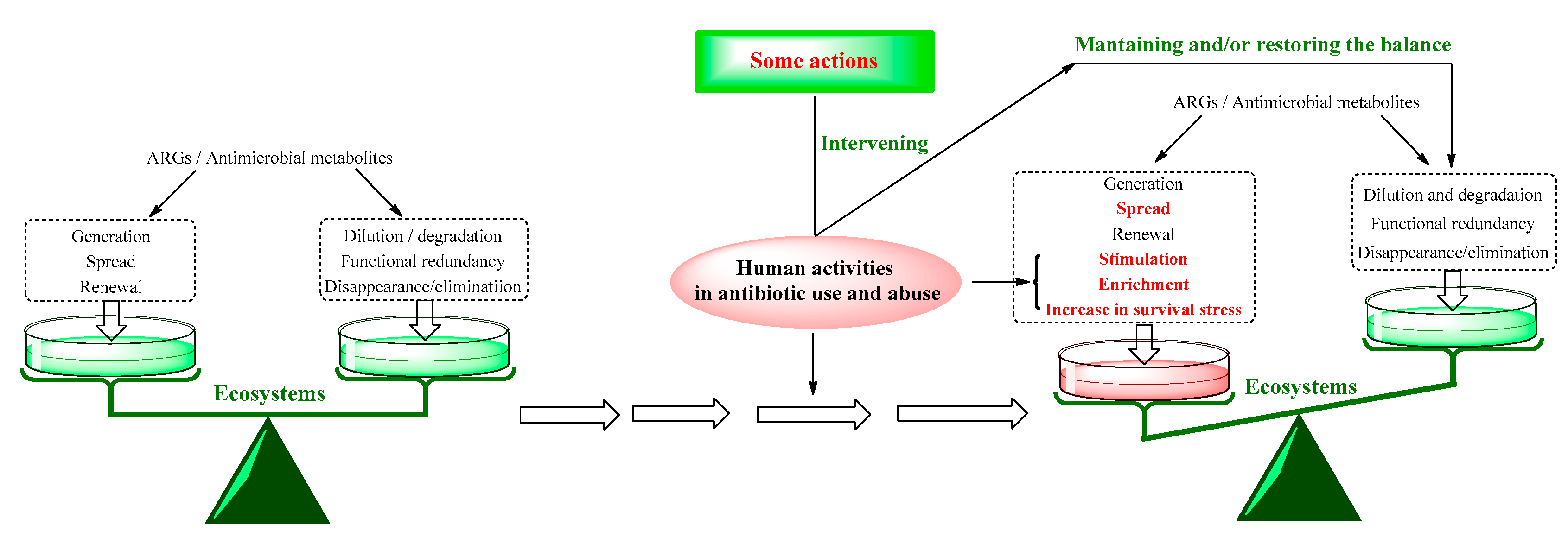
4. Measures Combating ABR from the OE-OH Concept Based on Ecosystems
As above mentioned, the use of antibiotics is considered as an intervention on ecosystems from the OE-OH concept. Thereby, a reverse intervention maintaining the balance of ecosystems should be taken for combating ABR (Figure 4) [68].
Simultaneously, the self-regulation and self-balance capabilities of ecosystems can be fully understood and utilized for the risk evaluation on some measures taken for the research and development, application and management of antibiotics. Combining with the support by literature, some important measures from the OE-OH concept, learning from the approach of problem management [69,70], are suggested as follows:
4.1. Minimizing the Use of Antibiotics, While Fully Utilizing the Regulatory ROLE of Plants on the Body's Ecosystem
From the OE-OH concept, the bodies of humans or animals themselves are ecosystems with sufficient self-regulation capabilities. For example, their immune systems [49], antimicrobial peptides [71], colonized probiotics, and competition among microorganisms in the body's ecosystem can eliminate infected pathogens including antibiotic-resistant ones. Meanwhile, many plants such as traditional Chinese medicine and ethnic medicines can also regulate the balance between the bodies of humans or animals and microbes [12,72,73], which was also proved in China during the COVID-19 pandemic [74]. Therefore, it is entirely possible to reduce the use of antibiotics through utilizing the self-regulation and self-balance ability of both humans and animals, as well as the regulation on this ability by plants. Furthermore, even if the bodies of humans and animals are infected, the balance between the bodies of humans or animals and microbes can be also regulated or restored by plants, reducing the development of microbial infection and avoiding unnecessary use of antibiotics as much as possible.
4.2. Minimizing the Emissions of Antibiotics and ARGs, While Fully Utilizing the Self-Regulation of Ecosystems.
From the OE-OH concept, microbial resistance mainly adopts the proactive pattern of adaptive evolution, occasionally the passive one of random mutation by natural selection. Along with the evolution and development of the Earth, ARG-carrying pathogens distributing everywhere can be activated, screened and enriched under the stresses from antibiotics. Therefore, the emission of antibiotics into the surrounding environment can not only stimulate the overexpression of ARGs in pathogens and enrich the information of ARGs, but also enable sensitive bacteria to proactively evolve into resistant pathogens. So, the emission of antibiotics and ARGs should be minimized.
Alternatively, from the OE-OH perspective, the transmission processes of antibiotic and ARGs are also the ones of their dilution, redundancy, or disappearance by various ecosystems. Therefore, when it is difficult to effectively control and emissions are necessary, the weakening and elimination of ecosystems to antibiotics and ARGs can be fully utilized [73,75,76,77]. At this moment, some scientific and reasonable measures should be taken, based on the risk management [78], for graded emissions to keep the emissions of antibiotics and ARGs within the controllable and balanced range of ecosystems, minimizing the damage caused by excessive accumulation of antibiotics and ARGs to the original ecosystem.
4.3. Avoiding the Excessive Aggregation of Population
From the OE-OH concept, the entire Earth's ecosystem has sufficient self-regulation ability for the generation, dissemination, enrichment, dilution, weakening and elimination of ARGs to maintain its own balance. However, the overuse and abuse of antibiotics by humans will continuously enrich ARGs and stimulate their copies in their living environment, leading to an increasing diversity and abundance of ARGs. If the population is excessively aggregated at this moment, it will greatly exceed the weakening and elimination of ARGs by various biotic and abiotic factors (Box 1) and the self-regulatory ability of ecosystems in the environment, leading to excessive enrichment of ARGs in ecosystems centered around the population agglomeration areas. Therefore, the excessive aggregation of population should be avoided [76]. Specifically, the scale of cities should not be too large, and the urban layout should be appropriately decentralized. This can also be indirectly proved by the transmission situation of the COVID-19 pandemic.
4.4. Accelerating the Development and Reserve of New Antibiotics Based on the Understanding for the Proactive Defense Mechanisms of Microbes
From the OE-OH concept, the proactive defense mechanisms of microbes in ecosystems can be comprehensively understood for better developing new antibiotics that are difficult to be resistance against, and it is also encouraged to develop new antibiotics with strong specificity and minimal disruption to the gut microbes and organismal ecosystems. Meanwhile, it is necessary to fully explore the unknown ARGs in ecosystems for predicting the potential ABR by AI technology. Based on the potential mechanisms of microbial resistance, the development of new antibiotics that are difficult for pathogens to be resistant to should be accelerated to ensure the reserves of new antibiotics. In addition, the protection period of patents for new antibiotics can be extended from policies, for reducing the unnecessary use of new antibiotics.
4.5. Encouraging Them in Combination with Plant Antimicrobial Ingredients
From the OE-OH perspective, the more the application of new antibiotics, the faster the spread and enrichment of ABR and ARGs, if the overuse and abuse of antibiotics are not controlled. So, it is necessary to use antibiotics reasonably. Among various strategies for the rational use of antibiotics, combination therapy is both economical and effective in enhancing the efficacy of antibiotics, reversing microbial resistance, and prolonging the life cycle of antibiotics, buying more time for the development of new antibiotics. This can be proved by the actual clinical effect of combination therapy, such as sulfamethoxazole and trimethoprim, β-lactamase inhibitors and β-lactam antibiotics, and the combination use of multiple anti-tuberculosis drugs. Therefore, combination therapy is worth promoting [12,52]. However, it is noteworthy that an inappropriate antibiotic combination may instead lead to an increase in the risk of ABR since the effect preventing resistance is associate with the fractional inhibitory concentration index, and the proportion and concentration of two antibiotics in a combination [19,20,79].
Antibiotics originate from the competition among microorganisms, and both the bacteria/fungi producing the structural skeleton of antibiotics and the pathogenic bacteria belong to microbes. Therefore, their individual defense mechanisms are familiar to each other, pathogens are prone to develop resistance to antibiotics in the combination. However, the antimicrobial ingredients of plants originate from the interaction between microorganisms and plants in ecosystems, and the defense mechanisms between plants and microbes are less familiar to each other. Simultaneously, the antimicrobial ingredients originated from plants generally present weaker antibacterial activity and smaller stresses on microbial survival than antibiotics. Therefore, it is difficult for microbes to develop resistance to them [20]. Moreover, the combination of plant antibacterial ingredients and antibiotics often has a wide range of synergistic effects [12], and the damage to the gut microbes is also weaker. Therefore, the antibacterial ingredients generated from plants are ideal candidates for combination therapy with antibiotics.
4.6. Simulating the Elimination of Antibiotics and ARGs in Ecosystems
From the OE-OH concept, ecosystems have sufficient ability to regulate ABR, including the dilution, weakening and elimination of ARGs. Therefore, it is encouraged to simulate the elimination of antibiotics and ARGs in ecosystems based on the understanding of self-regulation mechanisms of ecosystems [68,80]. For example, using bacterial ecology to combat ARG dissemination [4]; photocatalysis enhanced constructed wetlands to remove ARGs [81]; simulating sunlight-induced inactivation of tetracycline resistant bacteria [82]; using bacteria-microalgae-fungi symbionts or plants to remove antibiotics [83,84]. Another, these methods and technologies of ecological simulation can also avoid, as much as possible, the probable adverse effects on ecosystems, caused by the measure taken.
5. Methods
5.1. The OE-OH Concept
The OE-OH concept was put forwards from the previous results and the sufficient self-regulation ability of the Earth’s ecosystem in the evolution of ABR [12,23]. Here this concept has been further defined, improved and clarified using logical, reasoning and deductive methods based on the principle of parsimony [24,25], involving the understanding integration of Lamarck theory, Darwinian evolution, and Niche construction theory. It includes dual mutation patterns of bacterial resistance, theoretical logic of the OE-OH concept based on ecosystems, and basic mathematical model for the ARGs renewing with the ecosystem. The literature supporting the reasoning and deduction, together with all other literature, was unsystematically searched from PubMed database and Google academic search engine, using various relevant keywords. Furthermore, some highly persuasive references in the obtained literature were also tracked.
5.2. Analyses of ARG Generation, Spread and Elimination from the OE-OH Concept
From the OE-OH concept, the generation, spread and elimination of ARGs along with different time nodes of the Earthʹs evolution were analyzed using reasoning, deductive and inductive methods based on the self-regulation of ecosystems, together with the supporting of literature also searched from PubMed database and Google academic search engine. These especially include the emergence and distribution of ARGs emerging before humans, the ARG regulation by ecosystems such as its generation, spread, weakening and elimination of ARGs before the industrial production and use of antibiotics, the sharp increase in ARGs after the industrial production and use of antibiotics together with which impact on the self-regulation and self-balance of ecosystems for ARGs.
5.3. Measures Combating ABR from the OE-OH Concept Based on Ecosystems
Regarding the use of antibiotics as an intervention on ecosystems, some important measures for the research and development, application and management of antibiotics are suggested, from the OE-OH prospective, for maintaining the balance of ecosystems regulating on ARGs, learning from the approach of problem management [69,70].
6. Conclusions
The OE-OH concept was defined and clarified, which includes the dual mutation pattern of proactive evolution (most) and passive selection (few) for bacterial resistance mutation, the theoretical logic of the OE-OH concept based on ecosystems, and a basic mathematical model for the ARGs renewing with the ecosystem. From this concept, it deduced that ABR and ARGs emerge 3.5 billion years ago and exist in every corner on the Earth prior to the production and use of antibiotics by humans. Moreover, the self-regulation of the Earth's ecosystem on the generation, spread, weakening and elimination of ABR and ARGs, along with different time nodes of the Earth’s evolution, was sorted out for understanding the OE-OH concept. Regarded as an intervention on ecosystems, the industrial production and use of antibiotics, especially their overuse and abuse, have led to a sharp increase in ARGs and a worry beyond the self-regulation of ecosystems. Based on this, some important measures from the OE-OH concept are suggested for offsetting the imbalance of ecosystems in ARG generation and elimination and combatting ABR, especially emphasizing on the simulation and utilization for the self-regulation mechanisms of ecosystems, together with minimizing the use and emission of antibiotics, avoiding excessive aggregation of population, encouraging them in combination with plant antimicrobial ingredients, and so on.
Author Contributions
Conceptualization, G.Y.; Methodology, G.Y.; Software, G.Y.; Validation, G.Y.; Formal analysis, G.Y.; Investigation, G.Y., J.H., M.Z., X.C. and Y.L.; Resources, G.Y.; Data curation, G.Y.; Writing—original draft preparation, G.Y. J.H., M.Z., X.C. and Y.L.; Writing—review and editing, G.Y. J.H., M.Z., X.C. and Y.L.; Visualization, G.Y.; Supervision, G.Y.; Project administration, G.Y.; Funding acquisition, G.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant Nos. 82073745 and 82360691).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Mantilla-Calderon, D.; Xiong, Y.; Alkahtani, M.; Bashawri, Y.M.; Al Qarni, H.; Hong, P.Y. Investigation of antibiotic resistome in hospital wastewater during the COVID-19 pandemic: is the initial phase of the pandemic contributing to antimicrobial resistance? Environ. Sci. Technol. 2022, 56, 15007–15018. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Yutao, L.; Goh, S.G.; Ng, C.; Luhua, Y.; Tran, N.H.; Gin, K.Y. Quaternary ammonium compounds of emerging concern: classification, occurrence, fate, toxicity and antimicrobial resistance. J. Hazard. Mater. 2023, 445, 130393. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhou, X.; Fu, Q.; Li, Y.; Ni, B.J.; Liu, X. Understanding bacterial ecology to combat antibiotic resistance dissemination. Trends Biotechnol. 2025, 23, S0167–7799. [Google Scholar] [CrossRef]
- Wang, B.; Ma, B.; Zhang, Y.; Stirling, E.; Yan, Q.; He, Z.; Liu, Z.; Yuan, X.; Zhang, H. Global diversity, coexistence and consequences of resistome in inland waters. Water Res. 2024, 253, 121253. [Google Scholar] [CrossRef]
- Hu, X.; Fu, Y.; Shi, H.; Xu, W.; Shen, C.; Hu, B.; Ma, L.; Lou, L. Neglected resistance risks: cooperative resistance of antibiotic resistant bacteria influenced by primary soil components. J. Hazard Mater. 2022, 429, 128229. [Google Scholar] [CrossRef]
- Gao, F.Z.; He, L.Y.; Liu, Y.S.; Zhao, J.L.; Zhang, T.; Ying, G.G. Integrating global microbiome data into antibiotic resistance assessment in large rivers. Water Res. 2024, 250, 121030. [Google Scholar] [CrossRef]
- Fu, C.X.; Chen, C.; Xiang, Q.; Wang, Y.F.; Wang, L.; Qi, F.Y.; Zhu, D.; Li, H.Z.; Cui, L. Hong WL, et al. Antibiotic resistance at environmental multi-media interfaces through integrated genotype and phenotype analysis. J. Hazard Mater. 2024, 480, 136160. [Google Scholar] [CrossRef]
- Naddaf, M. 40 million deaths by 2050: toll of drug-resistant infections to rise by 70. Nature 2024, 633, 747–748. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial antibiotic resistance: the most critical pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- FAO, UNEP, WHO, WOAH. One Health Joint Plan of Action (2022–2026). Working together for the health of humans, animals, plants and the environment. Rome, 2022. [CrossRef]
- Yuan, G.; Lian, F.; Yan, Y.; Wang, Y.; Zhang, L.; Zhu, J.; Fatima, A.; Qian, Y. One Earth-One Health (OE-OH): antibacterial effects of plant flavonoids in combination with clinical antibiotics with various mechanisms. Antibiotics, 2025, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Lenton, T.M. Gaia and natural selection. Nature 1998, 394, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, A.E.; Wilkinson, D.M.; Williams, H.T.P.; Lenton, T.M. Multiple states of environmental regulation in well-mixed model biospheres. J. Theor. Biol. 2017, 414, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Maull, V.; Pla Mauri, J.; Conde Pueyo, N.; Solé, R. A synthetic microbial Daisyworld: planetary regulation in the test tube. J. R. Soc. Interface 2024, 21, 20230585. [Google Scholar] [CrossRef]
- R16. Lynch, M.; O'Hely, M.; Walsh, B.; Force, A. The probability of preservation of a newly arisen gene duplicate. Genetics 2001, 159, 1789–1804. [Google Scholar] [CrossRef]
- Qian, S.H.; Chen, L.; Xiong, Y.L.; Chen, Z.X. Evolution and function of developmentally dynamic pseudogenes in mammals. Genome Biol. 2022, 23, 235. [Google Scholar] [CrossRef]
- Smith, W.P.J.; Wucher, B.R.; Nadell, C.D.; Foster, K.R. Bacterial defences: mechanisms, evolution and antimicrobial resistance. Nat. Rev. Microbiol. 2023, 21, 519–534. [Google Scholar] [CrossRef]
- Xu, X.; Xu, L.; Yuan, G.; Wang, Y.; Qu, Y.; Zhou, M. Synergistic combination of two antimicrobial agents closing each other's mutant selection windows to prevent antimicrobial resistance. Sci Rep. 2018, 8, 7237. [Google Scholar] [CrossRef]
- Yi, H.; Yuan, G.; Li, S.; Xu, X.; Guan, Y.; Zhang, L.; Yan, Y. Drug combinations to prevent antimicrobial resistance: various correlations and laws, and their verifications, thus proposing some principles and a preliminary scheme. Antibiotics 2022, 11, 1279. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: an update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Yi, H.; Zhang, L.; Guan, Y.; Li, S.; Lian, F.; Fatima, A.; Wang, Y. Drug combinations to prevent antimicrobial resistance: theory, scheme and practice. In Book of Abstracts, Proceedings of the 6th International Caparica Conference on Antibiotic Resistance.; Caparica, Portugal, 8–12 September 2024; Capelo-Martinez, J.L., Santos, H.M., Oliveira, E., Fernández, J., Lodeiro, C., Eds.; PROTEOMASS Scientific Society: Caparica, Portugal, 2024. [Google Scholar]
- Van Den Berg, H.A. Occam's razor: from Ockham's via moderna to modern data science. Sci Prog. 2018, 101, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Khandai, K.; Navarro-Martinez, C.; Smith, B.; Buonopane, R.; Byun, S.A.; Patterson, M. Determining significant correlation between pairs of extant characters in a small parsimony framework. J. Comput. Biol. 2022, 29, 1132–1154. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Tang, S.; Liu, H.; Meng, Y.; Luo, H.; Wang, B.; Hou, X.L.; Yan, B.; Yang, C.; Guo, Z.; et al. Inheritance of acquired adaptive cold tolerance in rice through DNA methylation. Cell, 0092. [Google Scholar] [CrossRef]
- Laland, K.; Matthews, B.; Feldman, M.W. An introduction to niche construction theory. Evol. Ecol. 2016, 30, 191–202. [Google Scholar] [CrossRef]
- Constant, A.; Ramstead, M.J.D.; Veissière, S.P.L.; Campbell, J.O.; Friston, K.J. A variational approach to niche construction. J. R. Soc. Interface 2018, 15, 20170685. [Google Scholar] [CrossRef]
- Zhang, C.; Straight, P.D. Antibiotic discovery through microbial interactions. Curr. Opin. Microbiol. 2019, 51, 64–71. [Google Scholar] [CrossRef]
- Booth, S.C.; Smith, W.P.J.; Foster, K.R. The evolution of short- and long-range weapons for bacterial competition. Nat. Ecol. Evol. 2023, 7, 2080–2091. [Google Scholar] [CrossRef]
- Nachmias, N.; Dotan, N.; Rocha, M.C.; Fraenkel, R.; Detert, K.; Kluzek, M.; Shalom, M.; Cheskis, S.; Peedikayil-Kurien, S.; Meitav, G.; et al. Systematic discovery of antibacterial and antifungal bacterial toxins. Nat. Microbiol. 2024, 9, 3041–3058, Erratum in: Nat. Microbiol. 2025, 10, 1024. [Google Scholar] [CrossRef]
- Callahan, B.J.; Fukami, T.; Fisher, D.S. Rapid evolution of adaptive niche construction in experimental microbial populations. Evolution 2014, 68, 3307–3316. [Google Scholar] [CrossRef]
- Sykes, R. The 2009 Garrod lecture: the evolution of antimicrobial resistance: a Darwinian perspective. J. Antimicrob. Chemother. 2010, 65, 1842–1852. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; van Elsas, J.D.; Olff, H.; Salles, J.F. Dispersal-competition tradeoff in microbiomes in the quest for land colonization. Sci. Rep. 2018, 8, 9451. [Google Scholar] [CrossRef] [PubMed]
- Ohemeng, K.A.; Schwender, C.F.; Fu, K.P.; Barrett, J.F. DNA gyrase inhibitory and antibacterial activity of some flavones (l). Bioorg. Med. Chem. Lett. 1993, 3, 225–230. [Google Scholar] [CrossRef]
- Schwartzman, J.A.; Lebreton, F.; Salamzade, R.; Shea, T.; Martin, M.J.; Schaufler, K.; Urhan, A.; Abeel, T.; Camargo, I.L.B. C, Sgardioli, B.F.; et al. Global diversity of enterococci and description of 18 previously unknown species. Proc. Natl. Acad. Sci. U. S. A. 2024, 121, e2310852121. [Google Scholar] [CrossRef] [PubMed]
- Heidrich, V.; Valles-Colomer, M.; Segata, N. Human microbiome acquisition and transmission. Nat. Rev. Microbiol. 2025. [Google Scholar] [CrossRef]
- Munk, P.; Brinch, C.; Møller, F.D.; Petersen, T.N.; Hendriksen, R.S.; Seyfarth, A.M.; Kjeldgaard, J.S.; Svendsen, C.A.; van Bunnik, B.; Berglund, F.; et al. Genomic analysis of sewage from 101 countries reveals global landscape of antimicrobial resistance. Nat. Commun. 2022, 13, 7251, Erratum in: Nat. Commun. 2023, 14, 178. [Google Scholar] [CrossRef]
- Wang, C.; Kuzyakov, Y. Mechanisms and implications of bacterial-fungal competition for soil resources. ISME J. 2024, 18, wrae073. [Google Scholar] [CrossRef]
- Plough, H.H. Penicillin resistance of Staphylococcus aureus and its clinical implications. Am. J. Clin. Pathol. 1945, 15, 446–51. [Google Scholar] [CrossRef]
- Kirby, W.M. Extraction of a highly potent penicillin inactivator from penicillin resistant staphylococci. Science 1944, 99, 452–453. [Google Scholar] [CrossRef]
- Liao, X.; Hou, L.; Zhang, L.; Grossart, H.P.; Liu, K.; Liu, J.; Chen, Y.; Liu, Y.; Hu, A. Distinct influences of altitude on microbiome and antibiotic resistome assembly in a glacial river ecosystem of Mount Everest. J. Hazard Mater. 2024, 479, 135675. [Google Scholar] [CrossRef]
- Arros, P.; Palma, D.; Gálvez-Silva, M.; Gaete, A.; Gonzalez, H.; Carrasco, G.; Coche, J.; Perez, I.; Castro-Nallar, E.; Galbán, C.; et al. Life on the edge: microbial diversity, resistome, and virulome in soils from the union glacier cold desert. Sci. Total Environ. 2024, 957, 177594. [Google Scholar] [CrossRef]
- Roy, S.; Dawson, R.A.; Bradley, J.A.; Hernández, M. Prevalence and dynamics of antimicrobial resistance in pioneer and developing Arctic soils. BMC Microbiol. 2025, 25, 50. [Google Scholar] [CrossRef]
- Bisaccia, M.; Berini, F.; Marinelli, F.; Binda, E. Emerging Trends in antimicrobial resistance in polar aquatic ecosystems. Antibiotics 2025, 14, 394. [Google Scholar] [CrossRef] [PubMed]
- Na, G.; Zhang, W.; Gao, H.; Wang, C.; Li, R.; Zhao, F.; Zhang, K.; Hou, C. Occurrence and antibacterial resistance of culturable antibiotic-resistant bacteria in the Fildes Peninsula, Antarctica. Mar. Pollut. Bull. 2021, 162, 111829. [Google Scholar] [CrossRef] [PubMed]
- Rasouly, A.; Shamovsky, Y.; Epshtein, V.; Tam, K.; Vasilyev, N.; Hao, Z.; Quarta, G.; Pani, B.; Li, L.; Vallin, C.; et al. Analysing the fitness cost of antibiotic resistance to identify targets for combination antimicrobials. Nat. Microbiol. 2021, 6, 1410–1423. [Google Scholar] [CrossRef] [PubMed]
- Maddamsetti, R.; Yao, Y.; Wang, T.; Gao, J.; Huang, V.T.; Hamrick, G.S.; Son, H.I.; You, L. Duplicated antibiotic resistance genes reveal ongoing selection and horizontal gene transfer in bacteria. Nat. Commun. 2024, 15, 1449. [Google Scholar] [CrossRef]
- Leeansyah, E.; Boulouis, C.; Kwa, A.L.H.; Sandberg, J.K. Emerging role for MAIT cells in control of antimicrobial resistance. Trends Microbiol. 2021, 29, 504–516. [Google Scholar] [CrossRef]
- Woelfel, S.; Silva, M.S.; Stecher, B. Intestinal colonization resistance in the context of environmental, host, and microbial determinants. Cell Host Microbe 2024, 32, 820–836. [Google Scholar] [CrossRef]
- Lund, D.; Parras-Moltó, M.; Inda-Díaz, J.S.; Ebmeyer, S.; Larsson, D.G.J.; Johnning, A.; Kristiansson, E. Genetic compatibility and ecological connectivity drive the dissemination of antibiotic resistance genes. Nat. Commun. 2025, 16, 2595. [Google Scholar] [CrossRef]
- Perry, E.K.; Meirelles, L.A.; Newman, D.K. From the soil to the clinic: the impact of microbial secondary metabolites on antibiotic tolerance and resistance. Nat. Rev. Microbiol. 2022, 20, 129–142. [Google Scholar] [CrossRef]
- He, L.X.; He, L.Y.; Tang, Y.J.; Qiao, L.K.; Xu, M.C.; Zhou, Z.Y.; Bai, H.; Zhang, M.; Ying, G.G. Deciphering spread of quinolone resistance in mariculture ponds: cross-species and cross-environment transmission of resistome. J. Hazard Mater. 2025, 487, 137198. [Google Scholar] [CrossRef]
- Wang, X.; Wang, M.; Zhang, W.; Li, H.; Tiedje, J.M.; Zhou, J.; Topp, E.; Luo, Y.; Chen, Z. Treatment of antibiotic-manufacturing wastewater enriches for Aeromonas veronii, a zoonotic antibiotic-resistant emerging pathogen. ISME J. 2025, 19, wraf077. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.E.; Farkas, K.; Kiss, A.; Jones, D.L. National-scale insights into AMR transmission along the wastewater-environment continuum. Water Res. 2025, 282, 123603. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, I.T.; Santos, L. Antibiotics in the aquatic environments: a review of the European scenario. Environ. Int. 2016, 94, 736–757. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Yang, L.; Zhang, L.; Ye, B.; Wang, L. Antibiotics in soil and water in China-a systematic review and source analysis. Environ. Pollut. 2020, 266, 115147. [Google Scholar] [CrossRef]
- Danko, D.; Bezdan, D.; Afshin, E.E.; Ahsanuddin, S.; Bhattacharya, C.; Butler, D.J.; Chng, K.R.; Donnellan, D.; Hecht, J.; Jackson, K.; et al. A global metagenomic map of urban microbiomes and antimicrobial resistance. Cell 2021, 184, 3376–3393. [Google Scholar] [CrossRef]
- Xiang, Q.; Chen, Q.L.; Zhu, D.; An, X.L.; Yang, X.R.; Su, J.Q.; Qiao, M.; Zhu, Y.G. Spatial and temporal distribution of antibiotic resistomes in a peri-urban area is associated significantly with anthropogenic activities. Environ. Pollut. 2018, 235, 525–533. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Jia, S.; Qu, M.; Pei, Y.; Qiu, S.; Zhang, J.; Liu, Y.; Ma, S.; Lyu, N.; et al. A global atlas and drivers of antimicrobial resistance in Salmonella during 1900-2023. Nat. Commun. 2025, 16, 4611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Q.; Wang, T.; Xu, N.; Lu, T.; Hong, W.; Penuelas, J.; Gillings, M.; Wang, M.; Gao, W.; et al. Assessment of global health risk of antibiotic resistance genes. Nat. Commun. 2022, 13, 1553. [Google Scholar] [CrossRef]
- Wang, Y.F.; Liu, Y.J.; Fu, Y.M.; Xu, J.Y.; Zhang, T.L.; Cui, H.L.; Qiao, M.; Rillig, M.C.; Zhu, Y.G.; Zhu, D. Microplastic diversity increases the abundance of antibiotic resistance genes in soil. Nat. Commun. 2024, 15, 9788. [Google Scholar] [CrossRef]
- Tang, X.; Yuan, X.; Lu, W.; Wen, X.; Wu, Y.; Chen, T. ; Microplastics in livestock manure and compost: environmental distribution, degradation behavior, and their impact on antibiotic resistance gene dissemination, Chem. Eng. J. 2025, 513, 162881. [Google Scholar] [CrossRef]
- Li, K.; Rahman, S.U.; Rehman, A.; Li, H.; Hui, N.; Khalid, M. Shaping rhizocompartments and phyllosphere microbiomes and antibiotic resistance genes: The influence of different fertilizer regimes and biochar application. J. Hazard Mater. 2025, 487, 137148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, X.; Sun, H.; Ling, H.; Xie, R.; Fang, L.; Guo, M.; Wu, X. Polyvinyl chloride microplastic triggers bidirectional transmission of antibiotic resistance genes in soil-earthworm systems. Environ. Int. 2025, 198, 109414. [Google Scholar] [CrossRef] [PubMed]
- Dorninger, C.; Menéndez, L.P.; Caniglia, G. Social-ecological niche construction for sustainability: understanding destructive processes and exploring regenerative potentials. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2024, 379, 20220431. [Google Scholar] [CrossRef] [PubMed]
- Wong, F.; Zheng, E.J.; Valeri, J.A.; Donghia, N.M.; Anahtar, M.N.; Omori, S.; Li, A.; Cubillos-Ruiz, A.; Krishnan, A.; Jin, W.; et al. Discovery of a structural class of antibiotics with explainable deep learning. Nature 2024, 626, 177–185. [Google Scholar] [CrossRef]
- Laland, K.; Odling-Smee, J.; Endler, J. Niche construction, sources of selection and trait coevolution. Interface Focus 2017, 7, 20160147. [Google Scholar] [CrossRef]
- Morgan, M.A.; Long, J.S.; Logan, M.W.; Benton, F. Pareto in prison. Behav. Sci. Law 2025, 43, 299–313. [Google Scholar] [CrossRef]
- Olson, B.A.; Mazzuchi, T.A.; Sarkani, S.; Forsberg, K. Problem management process, filling the gap in the systems engineering processes between the risk and opportunity processes. Systems Eng 2012, 15, 275–286. [Google Scholar] [CrossRef]
- Goldberg, K.; Lobov, A.; Antonello, P.; Shmueli, M.D.; Yakir, I.; Weizman, T.; Ulman, A.; Sheban, D.; Laser, E.; Kramer, M.P.; et al. Cell-autonomous innate immunity by proteasome-derived defence peptides. Nature 2025, 639, 1032–1041. [Google Scholar] [CrossRef]
- Takahashi, K.M.; Nakasato, J.A.; de Jesus, G.S.; Micheletti, A.C.; Pott, A.; Yoshida, N.C.; Paulo. P.L. Antimicrobial activity of Pantanal macrophytes against multidrug resistant bacteria shows potential for improving nature-based solutions. Plant Stress 2024, 11, 100304. [Google Scholar] [CrossRef]
- Feigin, S.V.; Wiebers, D.O.; Lueddeke, G.; Morand, S.; Lee, K.; Knight, A.; Brainin, M.; Feigin, V.L.; Whitfort, A.; Marcum, J.; et al. Proposed solutions to anthropogenic climate change: A systematic literature review and a new way forward. Heliyon 2023, 9, e20544. [Google Scholar] [CrossRef]
- Ren, W.; Liang, P.; Ma, Y.; Sun, Q.; Pu, Q.; Dong, L.; Luo, G.; Mazhar, M.; Liu, J.; Wang, R.; Yang, S. Research progress of traditional Chinese medicine against COVID-19. Biomed. Pharmacother. 2021, 137, 111310. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, K.; Gao, Z.; Harbottle, M.; Sapsford, D.; Cleall, P. Life cycle assessment and cost-benefit analysis of nature-based solutions for contaminated land remediation: a mini-review. Heliyon 2023, 9, e20632. [Google Scholar] [CrossRef]
- Mercado, G.; Wild, T.; Hernandez-Garcia, J.; Baptista, M.D.; van Lierop, M.; Bina, O.; Inch, A.; Ode Sang, Å.; Buijs, A.; Dobbs, C.; et al. Supporting nature-based solutions via nature-based thinking across European and Latin American cities. Ambio 2024, 53, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Randrup, T.B.; Buijs, A.; Konijnendijk, C.C.; Wild, T. Moving beyond the nature-based solutions discourse: introducing nature-based thinking. Urban Ecosyst. 2020, 23, 919–926. [Google Scholar] [CrossRef]
- Sparrow, A.; Smith-Torino, M.; Shamamba, S.M.; Chirakarhula, B.; Lwaboshi, M.A.; Benn, C.S.; Chumakov, K. A risk management approach to global pandemics of infectious disease and anti-microbial resistance. Trop. Med. Infect. Dis. 2024, 9, 280. [Google Scholar] [CrossRef]
- Liu, J.; Gefen, O.; Ronin, I.; Bar-Meir, M.; Balaban, N.Q. Effect of tolerance on the evolution of antibiotic resistance under drug combinations. Science 2020, 367, 200–204. [Google Scholar] [CrossRef]
- Odling-Smee, J.; Erwin, D.H.; Palkovacs, E.P.; Feldman, M.W.; Laland, K.N. Niche construction theory: a practical guide for ecologists. Q. Rev. Biol. 2013, 88, 4–28. [Google Scholar] [CrossRef]
- Chen, P.; Yu, X.; Zhang, J. Photocatalysis enhanced constructed wetlands effectively remove antibiotic resistance genes from domestic wastewater. Chemosphere 2023, 325, 138330. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Zhang, T.; Liu, H.; Qu, J.; Li, C.; Chen, J.; Peijnenburg, W.J.G.M. Simulated sunlight-induced inactivation of tetracycline resistant bacteria and effects of dissolved organic matter. Water Res. 2020, 185, 116241. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Z.; Zhao, C.; Lu, B.; Zhao, Y. Phytohormone gibberellins treatment enhances multiple antibiotics removal efficiency of different bacteria-microalgae-fungi symbionts. Bioresoure Technol. 2024, 394, 130182. [Google Scholar] [CrossRef]
- Zhou, T.; An, Q.; Zhang, L.; Wen, C.; Yan, C. Phytoremediation for antibiotics removal from aqueous solutions: a meta-analysis. Environ. Res. 2024, 240, 117516. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.X.; Anupoju, S.M.B.; Nguyen, A.; Zhang, H.; Ponder, M.; Krometis, L.A.; Pruden, A.; Liao, J. Evidence of horizontal gene transfer and environmental selection impacting antibiotic resistance evolution in soil-dwelling Listeria. Nat. Commun. 2024, 15, 10034. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Liang, J.L.; Su, J.Q.; Jia, P.; Lu, J.L.; Zheng, J.; Wang, Z.; Feng, S.W.; Luo, Z.H.; Ai, H.X.; et al. Globally distributed mining-impacted environments are underexplored hotspots of multidrug resistance genes. ISME J. 2022, 16, 2099–2113. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Hu, H.W.; Maestre, F.T.; Guerra, C.A.; Eisenhauer, N.; Eldridge, D.J.; Zhu, Y.G.; Chen, Q.L.; Trivedi, P.; Du, S.; et al. The global distribution and environmental drivers of the soil antibiotic resistome. Microbiome 2022, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.C.; Shuai, X.Y.; Lin, Z.J.; Zheng, J.; Chen, H. Comprehensive profiling and risk assessment of antibiotic resistance genes in a drinking water watershed by integrated analysis of air-water-soil. J. Environ. Manage. 2023, 347, 119092. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, X.; Yang, T.; Su, J.; Qin, Y.; Wang, S.; Gillings, M.; Wang, C.; Ju, F.; Lan, B.; et al. Air pollution could drive global dissemination of antibiotic resistance genes. ISME J. 2021, 15, 270–281. [Google Scholar] [CrossRef]
- Rao, C.; Liu, X.; Xue, L.; Xiang, D.; Xian, B.; Chu, F.; Fang, F.; Tang, W.; Bao, S.; Fang, T. Determining the spatiotemporal variation, sources, and ecological processes of antibiotic resistance genes in a typical lake of the middle reaches of the Yangtze River. Sci. Total Environ. 2024, 907, 167779. [Google Scholar] [CrossRef]
- Mao, X.; Yin, X.; Yang, Y.; Gao, F.; Li, S.; Shi, X.; Deng, Y.; Li, L.; Leung, K.M.Y.; Zhang, T. Longitudinal metagenomic analysis on antibiotic resistome, mobilome, and microbiome of river ecosystems in a sub-tropical metropolitan city. Water Res. 2025, 274, 123102. [Google Scholar] [CrossRef]
- Dželalija, M.; Kvesić-Ivanković, M.; Jozić, S.; Ordulj, M.; Kalinić, H.; Pavlinović, A.; Šamanić, I.; Maravić, A. Marine resistome of a temperate zone: distribution, diversity, and driving factors across the trophic gradient. Water Res. 2023, 246, 120688. [Google Scholar] [CrossRef]
- Yu, W.; Xu, Y.; Wang, Y.; Sui, Q.; Xin, Y.; Wang, H.; Zhang, J.; Zhong, H.; Wei, Y. An extensive assessment of seasonal rainfall on intracellular and extracellular antibiotic resistance genes in Urban River systems. J. Hazard Mater. 2023, 455, 131561. [Google Scholar] [CrossRef]
- Shi, J.; Thakur, M.P. Climate extremes disrupt fungal-bacterial interactions. Nat. Microbiol. 2023, 8, 2226–2229. [Google Scholar] [CrossRef] [PubMed]
- Sokol, N.W.; Slessarev, E.; Marschmann, G.L.; Nicolas, A.; Blazewicz, S.J.; Brodie, E.L.; Firestone, M.K.; Foley, M.M.; Hestrin, R.; Hungate, B.A.; et al. Life and death in the soil microbiome: how ecological processes influence biogeochemistry. Nat. Rev. Microbiol. 2022, 20, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Jansson, J.K.; Hofmockel, K.S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 2020, 18, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wu, Y.; Huang, H.; Li, S.; Zhao, L.; Cao, J.; Wang, C. Revealing the critical role of rare bacterial communities in shaping antibiotic resistance genes in saline soils through metagenomic analysis. J. Hazard Mater. 2025, 491, 137848. [Google Scholar] [CrossRef]
- Zhao, M.; Zhou, X.; Li, Z.; Xu, G.; Li, S.; Feng, R.; Xia, D. The dynamics and removal efficiency of antibiotic resistance genes by UV-LED treatment: an integrated research on single- or dual-wavelength irradiation. Ecotox. Environ. Safe 2023, 263, 115212. [Google Scholar] [CrossRef]
- Huang, S.; Wang, Q.; Fan, Z.; Xu, M.; Ji, R.; Jin, X.; Gu, C. Dry-to-wet fluctuation of moisture contents enhanced the mineralization of chloramphenicol antibiotic. Water Res. 2023, 240, 120103. [Google Scholar] [CrossRef]
- Simonsen, A.K. Environmental stress leads to genome streamlining in a widely distributed species of soil bacteria. ISME J. 2022, 16, 423–434. [Google Scholar] [CrossRef]
- Liu, J.; Xue, S.; Jiang, C.; Zhang, Z.; Lin, Y. Effect of dissolved organic matter on sulfachloropyridazine photolysis in liquid water and ice. Water Res. 2023, 246, 120714. [Google Scholar] [CrossRef]
- Qian, G.; Shao, J.; Hu, P.; Tang, W.; Xiao, Y.; Hao, T. From micro to macro: the role of seawater in maintaining structural integrity and bioactivity of granules in treating antibiotic-laden mariculture wastewater. Water Res. 2023, 246, 120702. [Google Scholar] [CrossRef]
- Ruan, C.; Ramoneda, J.; Kan, A.; Rudge, T.J.; Wang, G.; Johnson, D.R. Phage predation accelerates the spread of plasmid-encoded antibiotic resistance. Nat. Commun. 2024, 15, 5397. [Google Scholar] [CrossRef]
- Xia, R.; Yin, X.; Balcazar, J.L.; Huang, D.; Liao, J.; Wang, D.; Alvarez, P.J.J.; Yu, P. Bacterium-phage symbiosis facilitates the enrichment of bacterial pathogens and antibiotic-resistant bacteria in the plastisphere. Environ. Sci. Technol. 2025, 59, 2948–2960. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Liang, J.L.; Wen, P.; Jia, P.; Feng, S.W.; Liu, S.Y.; Zhuang, Y.Y.; Guo, Y.Q.; Lu, J.L.; Zhong, S.J.; et al. Giant viruses as reservoirs of antibiotic resistance genes. Nat. Commun. 2024, 15, 7536. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Weiss, A.; Ma, H.R.; Son, H.I.; Zhou, Z.; You, L. Antibiotic-mediated microbial community restructuring is dictated by variability in antibiotic-induced lysis rates and population interactions. Nat. Commun. 2025, 16, 2299. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.L.; Correia-Melo, C.; Zorrilla, F.; Herrera-Dominguez, L.; Wu, M.Y.; Hartl, J.; Campbell, K.; Blasche, S.; Kreidl, M.; Egger, A.S.; et al. Microbial communities form rich extracellular metabolomes that foster metabolic interactions and promote drug tolerance. Nat. Microbiol. 2022, 7, 542–555. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Q.; Zhang, Z.; Zhou, S.; Jin, M.; Zhu, D.; Yang, X.; Qian, H.; Lu, T. Plants select antibiotic resistome in rhizosphere in early stage. Sci. Total Environ. 2023, 858, 159847. [Google Scholar] [CrossRef]
- Forster, S.C.; Liu, J.; Kumar, N.; Gulliver, E.L.; Gould, J.A.; Escobar-Zepeda, A.; Mkandawire, T.; Pike, L.J.; Shao, Y.; Stares, M.D.; et al. Strain-level characterization of broad host range mobile genetic elements transferring antibiotic resistance from the human microbiome. Nat. Commun. 2022, 13, 1445. [Google Scholar] [CrossRef]
- Lamberte, L.E.; van Schaik, W. Antibiotic resistance in the commensal human gut microbiota. Curr. Opin. Microbiol. 2022, 68, 102150. [Google Scholar] [CrossRef]
- Wang, F.; Sun, R.; Hu, H.; Duan, G.; Meng, L.; Qiao, M. The overlap of soil and vegetable microbes drives the transfer of antibiotic resistance genes from manure-amended soil to vegetables. Sci. Total Environ. 2022, 828, 154463. [Google Scholar] [CrossRef]
- McInnes, R.S.; McCallum, G.E.; Lamberte, L.E.; van Schaik, W. Horizontal transfer of antibiotic resistance genes in the human gut microbiome. Curr. Opin. Microbiol. 2020, 53, 35–43. [Google Scholar] [CrossRef]
- Yaffe, E.; Dethlefsen, L.; Patankar, A.V.; Gui, C.; Holmes, S.; Relman, D.A. Brief antibiotic use drives human gut bacteria towards low-cost resistance. Nature 2025, 641, 182–191. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, Y.; Mulchandani, R.; Van Boeckel, T.P. Global surveillance of antimicrobial resistance in food animals using priority drugs maps. Nat. Commun. 2024, 15, 763. [Google Scholar] [CrossRef] [PubMed]
- Proctor, D.M.; Sansom, S.E.; Deming, C.; Conlan, S.; Blaustein, R.A.; Atkins, T.K.; NISC Comparative Sequencing Program; Dangana, T. ; Fukuda, C.; Thotapalli, L.; et al. Clonal Candida auris and ESKAPE pathogens on the skin of residents of nursing homes. Nature 2025, 639, 1016–1023, Erratum in: Nat. Commun. 2024, 15, 933. [Google Scholar] [CrossRef] [PubMed]
- Häsler, R.; Kautz, C.; Rehman, A.; Podschun, R.; Gassling, V.; Brzoska, P.; Sherlock, J.; Gräsner, J.T.; Hoppenstedt, G.; Schubert, S.; et al. The antibiotic resistome and microbiota landscape of refugees from Syria, Iraq and Afghanistan in Germany. Microbiome 2018, 6, 37. [Google Scholar] [CrossRef]
- Ren, L.; Yang, J.; Xiao, Y.; Guo, L.; Rao, J.; Wu, C.; Wang, X.; Wang, Y.; Zhang, L.; Zhang, L.; et al. Transmission of the human respiratory microbiome and antibiotic resistance genes in healthy populations. Microbiome 2025, 13, 115. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Huang, Z.; Malakar, P.K.; Pan, Y.; Zhao, Y.; Zhang, Z. Antimicrobial resistomes in food chain microbiomes. Crit. Rev. Food Sci. Nutr. 2024, 64, 6953–6974. [Google Scholar] [CrossRef]
- Lu, J.; Guo, J. Disinfection spreads antimicrobial resistance. Science 2021, 371, 474. [Google Scholar] [CrossRef]
- Wang, Y.; Dagan, T. The evolution of antibiotic resistance islands occurs within the framework of plasmid lineages. Nat. Commun. 2024, 15, 4555. [Google Scholar] [CrossRef]
- Jiang, M.; Zheng, J.; Tang, Y.; Liu, H.; Yao, Y.; Zhou, J.; Lin, W.; Ma, Y.; Liu, J.; Zhou, J. Retrievable hydrogel networks with confined microalgae for efficient antibiotic degradation and enhanced stress tolerance. Nat. Commun. 2025, 16, 3160. [Google Scholar] [CrossRef]
- Yang, L.; Vadiveloo, A.; Chen, A.J.; Liu, W.Z.; Chen, D.Z.; Gao, F. Supplementation of exogenous phytohormones for enhancing the removal of sulfamethoxazole and the simultaneous accumulation of lipid by Chlorella vulgaris. Bioresoure Technol. 2023, 378, 129002. [Google Scholar] [CrossRef]
- Jacoby, R.P.; Koprivova, A.; Kopriva, S. Pinpointing secondary metabolites that shape the composition and function of the plant microbiome. J. Exp. Bot. 2021, 72, 57–69. [Google Scholar] [CrossRef]
- Zhao, Z.; Gao, B.; Henawy, A.R.; Rehman, K.U.; Ren, Z.; Jiménez, N.; Zheng, L.; Huang, F.; Yu, Z.; Yu, C.; et al. Mitigating the transfer risk of antibiotic resistance genes from fertilized soil to cherry radish during the application of insect fertilizer. Environ. Int. 2025, 199, 109510. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, K.; Jia, Y.; Shi, J.; Tong, Z.; Fang, D.; Yang, B.; Su, C.; Li, R.; Xiao, X.; Wang, Z. Gut microbiome alterations in high-fat-diet-fed mice are associated with antibiotic tolerance. Nat. Microbiol. 2021, 6, 874–884. [Google Scholar] [CrossRef] [PubMed]
- de Dios, R.; Proctor, C.R.; Maslova, E.; Dzalbe, S.; Rudolph, C.J.; McCarthy, R.R. Artificial sweeteners inhibit multidrug-resistant pathogen growth and potentiate antibiotic activity. EMBO Mol. Med. 2023, 15, e16397. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Yang, P.; Wang, B.; Xu, Q.; Song, F.; Wang, Y.; Zhao, Y.; Guo, L.; Wang, X.; Xu, M. Divergent mitigation mechanisms of soil antibiotic resistance genes by biochar from different agricultural wastes. Environ. Pollut. 2025, 374, 126247. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



