Submitted:
27 June 2025
Posted:
30 June 2025
You are already at the latest version
Abstract
(Background): Developments in biology, genetics, soil science, plant breeding, engineering, and agricultural microbiology are driving advances in soil microbiology and microbial biotechnology. (Material and methods): The literature for this review was collected by searching leading scientific databases such as Embase, Medline/Pubmed, Scopus, and Web of Science. Results: Recent advances in soil microbiology and biotechnology are discussed, emphasizing the role of microorganisms in sustainable agriculture. It has been shown that soil and plant microbiomes significantly contribute to improving soil fertility and plant and soil health. Microbes promote plant growth through various mechanisms, including potassium, phosphorus, and zinc solubilization, biological nitrogen fixation, production of ammonia, HCN, siderophores, and other secondary metabolites with antagonistic effects. The diversity of microbiomes related to crops, plant protection, and the environment is analyzed, as well as their role in improving food quality, especially under stress conditions. Particular attention was paid to the diversity of microbiomes and their mechanisms supporting plant growth and soil fertility. (Conclusion): The key role of soil microorganisms in sustainable agriculture was highlighted. They can support the production of natural substances used as plant protection products, as well as biopesticides, bioregulators or biofertilizers. Microbial biotechnology also offers potential in the production of sustainable chemicals, such as biofuels or biodegradable plastics (PHA) from plant sugars, and in the production of pharmaceuticals, including antibiotics, hormones or enzymes.
Keywords:
bioherbicides
; bioinsecticides
; biopesticides
; biomarkers
; effective microorganisms
; mycorrhizal fungi
; seaweed extracts
1. Introduction
Microbial biotechnology, focusing on microorganisms, plant extracts, and minerals, is crucial for sustainable development by reducing reliance on harmful chemicals and improving environmental health. It offers solutions like green chemistry, biopesticides, bioherbicides, bioinsecticides, and effective microorganisms (EM) for sustainable agriculture. However, ethical concerns exist, including the release of GMOs, increased resistance in weeds, diseases, and pests, highlighting the need for further research into their potential and limitations [1,2].
Sustainable agriculture employs various farming systems to minimize environmental impact, adhering to economic, social, and environmental pillars. Common systems include:
- -
- -
- -
- No-tillage: Improves water retention, reduces soil erosion, and increases soil organic matter.
- -
- Agroforestry: Boosts biodiversity and positively impacts water, climate, and ecological balance [3].
- -
- Precision farming: Optimizes resource use (water, energy, fertilizers, plant protection products) through accurate monitoring [4].
- -
Alongside these, technologies like farming automation and robotics enhance productivity and reduce costs [5,6], and algae to enhance plant productivity and health while protecting the environment. Key applications include:
- -
- -
- -
- -
These systems offer benefits like increased yields, enhanced plant resistance, reduced environmental impact, and lower production costs. However, risks such as potential health impacts on humans and animals, environmental pollution, food contamination, lack of effectiveness, undesirable interactions, and the development of microbial resistance necessitate careful application according to best practices and recommendations [3,6,7,8,9,10,11,12,13,14,15,16,17].
Soil microorganisms are vital for sustainable agriculture, participating in pathogen reduction and nutrient transformation [1,4,7]. They constitute a significant portion of environmental biomass and the carbon cycle [1,6]. The soil is a rich reservoir of diverse microorganisms, leading to efforts in developing microbiological preparations for soil fertility [1,5]. These biopreparations, often containing symbiotic microorganisms, improve plant growth and yield [1,6].
Microorganisms play a pivotal role in agriculture, horticulture, and animal production such as biofertilizers, bioherbicides, bioinsecticides, and growth bioregulators [7,8,9,10,11,12,13,14]. Recent research also explores combining EM applications with herbal extracts [6,7,8,9,10,11,12,13,14]. Continued research in agricultural microbiology is crucial to understanding microbial interactions with soil, plants, and the environment, unlocking their full potential for increased yields, enhanced plant resistance, improved nutritional value, and ultimately, food security and sustainable development [2,3,5,6].
Hence, the aim of this study is to highlight the key role of microorganisms in the natural environment, with particular emphasis on their use in agriculture, horticulture and animal production in the form of biofertilizers, bioherbicides, bioinsecticides, growth bioregulators and in denitrification processes [4,7,8,9,10,11,12,13,14]. This review aims to synthetically present the current achievements of microbiology in the development and use of biopreparations in sustainable agriculture, such as biostimulants, biopesticides, biofungicides, effective microorganisms and other microbiological organisms [1,2,3,4,5,6]. Additionally, the manuscript indicates new directions of research in agricultural microbiology, focusing on increasing plant resistance to diseases, improving the biodiversity of microbial species contributing to crop growth, as well as the impact on plant health, their nutritional and energy value, which is crucial for food security and sustainable development [2,4,5,6].
This paper puts forward an alternative hypothesis to the null hypothesis:
H0:
assumes no significant effect of microorganisms on the efficiency and sustainability of agricultural production.
H1:
alternative hypothesis, according to which the targeted use of specific groups of microorganisms (e.g., bacteria, fungi) in biopreparations (biofertilizers, biopesticides, biostimulants) significantly improves plant health, increases yields, reduces dependence on chemical plant protection products and artificial fertilizers, and thus contributes to the sustainable development of agriculture, minimizing the negative impact on the environment and human health [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16].
2. Materials and Methods
Articles used in this literature review were retrieved from PubMed/Medline, Scopus, Embase, Web of Science, Google Scholar, and ScienceDirect. No restrictions were applied to the year of publication or language in order to cover the widest possible range of literature. A total of 165 publications were included. The search was conducted using relevant keywords such as “fermentation,” “bioactive compounds,” “health benefits,” and the names of specific plants and microorganisms. Articles were selected based on the criteria of (1) relevance to the re-view topic, (2) publication in peer-reviewed journals, and (3) methodological soundness and substantive contribution. Preference was given to newer studies, while older key papers were also included. Titles and abstracts were reviewed independently by the authors; full texts were assessed in detail and any discrepancies were resolved through discussion. The review provides a comprehensive summary of current knowledge on the use of microorganisms and biotechnology in agricultural science and practice.
3. Review
3.1. Agricultural Microbiome
The soil microbiome is a complex community of microorganisms, including bacteria, fungi, viruses, protozoa and other single-celled organisms that inhabit the soil layers. Soil microbiome plays a key role in soil processes such as mineralization, degradation of organic matter, nitration, phosphorylation and others. The microbes in the soil are also important for maintaining plant health as they aid in the process of nitrogen transformation and nutrient absorption. The soil microbiome is very diverse and can vary greatly depending on soil type, climate, geographic location, and other abiotic and biotic factors. The different microbial species in the soil are also linked to different soil processes, meaning that the soil microbiome has a significant impact on the functioning of ecosystems and on agricultural productivity [1].
Changes in soil cultivation methods, related to the introduction of modern agriculture, chemicalization and intensive fertilization, have contributed to significant changes in the soil microbiome. The introduction of modern agricultural technologies, modern, high-performance machines, which, however, destroy the soil structure, mineral fertilizers, pesticides, growth regulators, caused a change in the composition of soil microorganisms. In particular, the use of mineral fertilizers and chemical pesticides can lead to a reduction in the number of soil microorganisms, especially saprotrophic bacteria and fungi, which are responsible for the decomposition processes and processing of organic matter in the soil [2].
Compared to intensively fertilized and chemically treated soils, poorly fertilized or organic soils are usually more diverse in terms of microorganisms. Microorganisms such as nitrogen bacteria and organic matter processing microorganisms predominate in such soils and contribute to soil fertility by converting organic matter into plant nutrients. The introduction of modern agricultural technologies may lead to some negative effects on the soil microbiome; at the same time, they may contribute to increasing the efficiency of agricultural production. It is important to find a balance between increasing the efficiency of agricultural production and protecting the soil microbiome and ensuring sustainable agricultural production in the long term. The introduction of modern, intensive agriculture, chemicalization and intensive fertilization can significantly change the soil microbiome. Although these activities are aimed at increasing plant production, they can lead to disturbances in the microbial balance in the soil [1,2].
The use of chemicals in agriculture, especially the use of pesticides, plant growth regulators and artificial mineral fertilizers, may lead to a decrease in the diversity and number of microorganisms in the soil. For example, pesticides can kill beneficial microorganisms, such as nitrogen bacteria, which take nitrogen from the air and convert it into nutrients. Intensive cultivation can lead to changes in the soil microbiome. Reducing plant diversity in farmland can also lead to bacterial poverty, as different plant species differ in ways that are beneficial to different microorganisms. In addition, changing the cultivation method can also affect the composition of the microorganisms, as different plant species support different populations of microorganisms. Climate change also affects the interactions between microorganisms and between microorganisms and plants in a direct and indirect way. Consequently, they had an impact on soil ecosystem functions such as carbon storage, productivity, fertility and fertility. Climate change may also affect the species composition of the microbiome inhabiting plant roots, as well as on the activity of mycorrhizal fungi and rhizosphere bacteria PGPR (Plant Growth Promoting Rhizobacteria). It comes to numerical and qualitative changes in microbial populations. The processes affecting the biological activity and fertility of soils may be affected intensification or inhibition [2,15,16].
Compared to intensively fertilized and chemise soils, weakly fertilized and organic soils usually have a higher number and greater variety of microorganisms. These soils contain more organic matter, which promotes the development of soil microorganisms. As the content of organic matter in the soil increases, the variety of microorganisms increases, which affects plant efficiency and general soil quality. The introduction of modern agriculture, chemigations and intensive fertilization leads to changes in soil microbiome, which are unfavorable for plant performance and general soil health. Compared to poorly fertilized and organic soils, intensively fertilized and chemise soils usually have a smaller number and variety of microorganisms, which can lead to a reduction in quality and efficiency of crops [3,4].
The variety of soil microbiomes in arable soils around the world is very diverse and depends on many factors, such as climate, soil type, crop plants and cultivation practices, for example: in Europe, soil microbiomes are strongly associated with the history of crops and types of crops. In regions with a long cultural history, soils usually have a lower variety of microorganisms. For example, in the regions of cereal cultivation in Central and Eastern Europe, soils usually have a lower number and variety of microorganisms, compared to soils in vegetable cultivation regions. Asia: Asia has various soil environments, which leads to the differentiation of soil microbiomes in different regions. For example, in Southeast Asia, where rice is grown, soil usually has a high number and variety of nitrification bacteria, which are crucial for the nitrogen cycle in soil. In China, soil in tea growing areas usually have a higher variety of fungi than soil in other regions. Africa: Africa usually has low organic matter and pH soils, which affects the composition of the soil microbiome. In the regions of wheat and corn cultivation, soil usually has a lower variety of nitrification bacteria compared to soils in vegetable cultivation regions. North and South America: in North and South America, soil in vegetable cultivation regions usually have a higher number and variety of nitrification bacteria and other microorganisms compared to soils in cereal cultivation regions [3,4,5,6,7,12,13,14,15,16]. To sum up, in brief, the variety of soil microbiomes in arable soils is very diverse around the world and depends on many factors. Looking at the diversity of microorganisms in arable soils on continents, countries and regions allows for a better understanding of the complexity of interaction between soil, plants and microorganisms, which is crucial to increasing the efficiency of crop and soil protection.
3.2. Sustainable Green Chemistry
The term “Green Chemistry” has been used for the first time first used by P.T. Anastas in called to life in 1991 by the US Environmental Protection Agency “green chemistry program”. It is a discipline that aims to design and develop chemical products and processes that reduce or completely eliminate the use of hazardous substances and the generation of waste. The approach to “Green Chemistry” is also known in the literature under other terms: “Environmentally Benign Chemistry” - chemistry that is gentle to the environment, “Clean Chemistry” - clean chemistry, “Atom Economy” - economy of atoms; “Benign by Design Chemistry” - gentle chemistry by design [6,7]. Green Chemistry is defined as environmentally friendly chemical synthesis, or as alternative methods of chemical synthesis, preventing environmental contamination, and its methodological distinctiveness, unlike traditional modern chemistry - in the search (design), development and implementation of new technologies and chemical materials, with considering the consequences related to use of materials [7,15,16,17,18,19,20,21,22,23,24,25,26].
Microbial biotechnology can be used to produce sustainable chemicals for a variety of purposes such as pest control and fertilization, by utilizing microorganisms that naturally produce these chemicals, or by genetically modifying microorganisms to produce specific chemicals [6].
One example of the use of microbial biotechnology in sustainable green chemistry is the production of biopesticides, which are pest control products made from natural materials such as microorganisms, plant extracts and minerals. They are generally considered safer and more environmentally friendly than synthetic pesticides because they are often biodegradable and have less toxicity to non-target organisms. Microbial biotechnology can be used to produce fertilizers and biopesticides by using microorganisms that produce toxins that are harmful to pests, such as Bacillus thuringiensis (Bt), or by genetically modifying the microorganisms to produce specific toxins [12,16]. Bacillus sp., especially B. thuringiensis, is widely used on the biopesticide market around the world due to its ability to effectively combat pests in economically important crops [16]. Traditional fertilizers are often derived from non-renewable resources such as fossil fuels. Both their production and use may have a negative impact on the natural environment. Microbial biotechnology can be used to produce fertilizers from renewable resources such as plant residues and animal manure by using microorganisms that convert these materials into nutrients that can be taken up by plants. As an example, nitrogen-fixing bacteria, such as rhizobia, can be mentioned, which can change atmospheric nitrogen into a form that is easily absorbed by crop plants, or phosphorus-dissolving bacteria, which increase the availability of insoluble forms of phosphorus for plants [6,7].
Green Technology has a significant, beneficial effect on increasing the efficiency of HEM (High-end Equipment Manufacturing), in medical use. Microbial biotechnology can also be used to produce other sustainable chemicals such as biofuels, biodegradable plastics, polyhydroxy alkanoates (PHAs) by fermenting plant sugars or other organic materials. And finally, microorganisms can be used to produce various pharmaceuticals such as antibiotics, hormones and enzymes through fermentation processes [12,13,14,15].
Microbial biotechnology in sustainable green chemistry can bring numerous benefits, such as reducing our dependence on non-renewable resources, reducing greenhouse gas emissions, and improving the sustainability of the production and use of chemicals. However, there are also various challenges and ethical issues to consider, such as the potential for GMOs to be released into the environment. Therefore, further research is needed to better explain the strengths and weaknesses of this technology and to develop safer ways to use it [6,7].
3.3. Biofertilizers
The problems of hidden hunger in the world can be solved with the help of new farming systems and methods, and one of them is biofortification and the use of biofertilizers in food crops [27,28,29,30,31,32]. The classification of biofertilizers based on their functions and nature is shown in Figure 1.
Preferred mechanisms for improving plant growth include increased nutrient availability (i.e., N, P, K, Mg, Zn and S), phytohormonal modulation, biological management of phytopathogens and stress mitigation (e.g., drought stress, salt stress) [33,34].
Biofertilizers are substances containing a variety of microorganisms that have a high ability to absorb nutrients by plants due to the colonization of the rhizosphere, and thanks to this, it is easily available to plant root hairs. They are a good alternative to synthetic, chemical fertilizers. There are different types of microbial biofertilizers, including symbiotic and free-living nitrogen fixers, solubilizers, and phosphorus mobilizers. Their formulas and applications, the latest approaches to the development of new generation biofertilizers are extremely important for sustainable agriculture [32,33,34]. In the future, it is expected that biofertilizers will contribute to a significant reduction in the application of both chemical fertilizers and pesticides. According to Mathur et al. [34,35] inoculation of crop plant seeds with an effective plant-arbuscular mycorrhizal fungi (AMF) vaccine or spores of obscure mycorrhizal fungi is an effective way to increase the symbiosis of plants with AMF in the soil [35,36]. The AMF symbiosis enhances the transport of water and nutrients from the soil to the host plants, which then allows the transit of carbohydrates from the photosynthesis pathway to an arbuscular mycorrhizal fungus [36,37].
Phosphate biofertilizers are one of the most important groups of beneficial microorganisms, which play an important role in the preparation of nutrients for crops. They are recognized as phosphate suppliers for various cropping systems. They can also provide other cropping systems with macro- and micronutrients. Both fungi and bacteria form the main groups of phosphate biofertilizers. They can live freely, independently or as symbiotic organisms in cultivated soils. Mycorrhiza is the so-called symbiont fungus that increases the uptake of both phosphates, nitrogen and micronutrients by plants. In addition, it improves the soil structure by creating a very extensive, dense network of mycelium connected to the roots of plants. Phosphate-dissolving microorganisms, on the other hand, are usually free-living, capable of dissolving insoluble phosphate compounds in the soil by releasing organic acids as well as chelating metabolites. The effectiveness of these microorganisms is significantly influenced by abiotic factors and farmland management practices. Tillage has a negative impact on the activity of mycorrhizal fungi. Also, the application of synthetic fertilizers also reduces the survival and effectiveness of phosphate biofertilizers [31,37,38].
Plant extracts or metabolites are absorbed by weed seeds, which first initiates cell membrane damage, DNA damage, mitosis, increases amylase activity and other biochemical processes, and delays or inhibits seed germination. The production of reactive oxygen species (ROS) and stress-related hormones increases, including the effects of antioxidants. Enzymes and toxic substances secreted by microbes degrade the seed coat of weeds and use the endosperm for survival, which inhibits seed germination. Microbes grow through the intercellular spaces, reaching the root core, and the toxins deposited in the cells affect cell division and their functions. Metabolites of harmful microorganism’s cause disease, necrosis and chlorosis, which in turn contribute to inhibition of germination and growth of weed seeds by inhibiting photosynthesis and gibberellin activity and enhancing ROS, abscisic acid and ethylene (Figure 2).
Figure 2 shows a comprehensive approach to weed control that combines the use of natural chemicals, microbial bioherbicides and potentially modified chemical derivatives (such as organophosphates). The key mechanism of microbial bioherbicides is the use of microorganisms (or their metabolic products) to directly control weeds, potentially through the production of herbicides such as organophosphates. This points to future strategies that integrate biological control methods (microorganisms) with natural chemicals, offering an alternative to synthetic herbicides and supporting the concept of sustainable agriculture.
3.3. Development of EM and Biofertilizer Formulation
The development of an effective EM (Effective Microorganisms) formulation that ensures good survival for all strains in a consortium is a challenge due to the different optimal conditions for individual species [31,39,40,41,42]. Recent research focuses on microencapsulation and prebiotics to improve the stability and synergism of strains in complex EM formulations, enabling their better survival under variable environmental conditions and ensuring long-term efficacy in the field [43].
Rhizobium, a bacterium widely used in the production of biofertilizers, is an example of the most environmentally friendly options [29]. Biofertilizers, including those based on fungi, contribute to the reduction of plant diseases by inhibiting the development of infections and limiting the biological functions of pathogens [31,39,40,41,42]. These fungi are seen as key contributors to biological life in the soil. Nitrogen fixation strategies, using both bacteria and fungi, focus on increasing plant nutrient utilization and reducing the dependence on synthetic inorganic fertilizers [31,39,40,41,42]. Particular attention has been paid to endophytic bacteria and fungi that fix nitrogen within plant tissues, which may open new possibilities for non-legumes [44].
Biofertilizers increase soil and plant nutrient content through a number of processes, including nitrogen fixation, phosphorus solubilization, and plant growth biostimulation [28,29,30,31]. Recent studies have shown that arbuscular mycorrhizal fungi (AMF) can reduce the structural and functional damage in the PSII and PSI photosystems induced by drought stress. For example, the colonization of D. moldavica roots by AMF promotes the growth of plants of this species by activating the antioxidant system and reducing the production of reactive oxygen species (ROS) [42,45,46,47,48]. Another novelty is the use of mycorrhizal fungi in combination with other biopreparations, which enhances the synergy and complex resistance of plants to many abiotic and biotic stresses [49]. Microbes associated especially with plants from the Fabaceae family (legumes) have great potential in solving environmental problems. Rhizobium, a bacterium used to produce biofertilizers, lives in symbiosis with the roots of legumes in structures called root nodules. These nodules enable the assimilation of atmospheric nitrogen in the form of ammonium, available to plants [30]. Rhizobia are common in soils, and “attracted” by lectins secreted by the roots of legumes, penetrate the hair cells via an infection thread. Infection of the adjacent epidermal cells of the root occurs, growth hormone is secreted by the plant, and finally the formation of a root nodule. During the period of intensive atmospheric nitrogen fixation (the flowering phase of the plant), the nodule takes colors from light pink to red, while white or brown colors indicate the cessation of this process. A protein called leghemoglobin, biochemically similar to animal hemoglobin, is responsible for the pink or red color of the nodules, as well as for maintaining appropriate oxygen conditions inside the nodule. Legume nodules differ anatomically and physiologically, and each plant species is usually capable of symbiosis only with the appropriate strain of bacteria. For example, beans form symbiosis with the bacteria Rhizobium leguminosarum by. phaseoli, while clover with Rhizobium leguminosarum by trifolii, etc. Some wild Rhizobium strains can coexist with many legume species and then have the ability to so-called cross-infect plants. Soybean, on the other hand, has the ability to coexist only with Bradrhizobium japonicum bacteria, which do not naturally occur in Europe. Therefore, it is necessary to inoculate them with a vaccine with these bacteria [31,39,40,41,42].
The source of phosphorus in biofertilizers is often ashes and bones, and microorganisms are used to solubilize it, e.g., Bacillus megaterium [3,40]. The source of phosphorus in biofertilizers is often ash and bones, and microorganisms, e.g., Bacillus megaterium [3,40] are used for its solubilization. For example, the only high-quality bacterial soil preparation on the Polish market, which enriches the soil with easily assimilable phosphorus from the decomposition of complex and insoluble forms, enhances plant growth, is based on a single, carefully selected strain of Bacillus megaterium ssp. phosphaticum bacteria. Fungal biopreparations, e.g., Nod, support plant growth and protect them against pathogens. Thanks to this, it is possible to increase the efficiency of legume cultivation, reduce the intensity of synthetic nitrogen fertilization and protect crops against pathogens in a sustainable way [43]. A novelty in this field is research on the use of endophytic fungi other than mycorrhizal, which can also affect the availability of phosphorus and potassium, as well as plant resistance [44].
Research is being conducted to obtain biopreparations that protect and stimulate plant growth. For example, it has been shown that the active ingredients of biopreparations derived from fungi (i.e., Fusarium spp., Trichoderma spp., Mortierella spp.) and bacteria (e.g., Pseudomonas luteola, P. fluorescens) stimulate crop growth and the absorption of phosphorus and potassium from soil minerals (apatites, phosphates). Bioprotective biopreparations against phytopathogens have been developed, containing non-pathogenic fungi from the genera Fusarium, Trichoderma, Mortierella or inducers of plant resistance pathways. It has been shown that plants treated with these biopreparations are resistant to phytopathogens [40,41,42,43]. Many studies in this area are covered by patent law. For example, plants treated with biopreparations developed by the Polish team of Jaroszuk-Ściseł from Maria Curie-Skłodowska University in Lublin are resistant to phytopathogens, because biopreparations activate systemic resistance pathways that are quickly activated in the event of contact with pathogens. This innovative invention has been submitted for patent protection (patent application P.428832) and uses natural biological methods of plant protection without the need to resort to chemical fungicides. Copper (and its compounds, e.g., copper sulfate, copper oxychloride): These are substances with fungicidal (fight fungi) and bactericidal (fight bacteria) effects. They are widely used in agriculture, horticulture and forestry to combat plant diseases caused by fungi (e.g., apple scab, downy mildew) and bacteria. Currently, there is a growing interest in precise sequencing of microorganism genomes in order to identify and optimize genes responsible for the production of biostimulants and protective compounds, which opens the way to designing “tailor-made” biopreparations [50]. Recently, seaweed extracts (SwE) are used for the strengthening and fertilization of plants under drought stress conditions, mainly from brown macroalgae, due to their composition, rich in polysaccharides, betaines, macro- and microelements and phytohormones, which are considered as beneficial signaling molecules for improving plant response [43]. The application of SwE – under favorable or critical conditions, such as drought stress, is associated with physiological and biochemical processes concerning the stimulation of enzymes involved in carbon and nitrogen metabolic pathways, Krebs cycle and glycolysis, stimulation of phytohormones and enhancement of mineral absorption and accumulation by biostimulated plants due to modification of root morphological structure [43,44]. The use of SwE can be considered as an appropriate and sustainable approach to overcoming agricultural problems caused by anthropogenic climate change. It is worth adding that the development of extraction and fractionation technologies allows for obtaining more concentrated and specific bioactive compounds from seaweed, which increases their efficiency and precision of action [51].
3.4. Biopesticides
The principles of Green Chemistry, related to research and the search for safe chemical products, indicate a growing interest in and introduction of new pesticides to agrochemical practice [44,45,46,47]. New chemical products introduced onto the market, including pesticides, should meet the basic requirements of Green Chemistry, i.e., principal no. 10, according to which after use they should not remain permanently in the environment, but degrade to products harmless to ecosystems. Biopesticides are a special group of active substances used in plant protection. They usually occur naturally or are produced synthetically but are identified with natural substances. These compounds usually decompose quite quickly, and semi-chemical pesticides are used in very low doses [44,45,46,47]. Biopesticides or biological pesticides are methods of pest and disease control that use predatory compounds, parasitism, and their chemical action [45,46,47,48].
Historically, the definition of biopesticides has been related to the biological control and manipulation of living organisms. Public opinion can also influence the perception of this issue and its regulatory significance. Thus, in the European Union, biopesticides are defined as “a form of pesticide based on microorganisms or natural products”, while in the USA, the EPA (Environmental Protection Agency) states that biopesticides include “pesticides of natural origin (biochemical pesticides), microorganisms, bacterial pesticides, and pesticides produced by plants with added genetic material (Protectants or PIPs)” [45,46,47,48]. Typically, such products are produced by culturing and concentrating naturally occurring organisms or their metabolites, including bacteria and other microorganisms, as well as proteins, fungi, nematodes, etc., which are recognized as important components of integrated pest management programs and considered as substitutes for chemical plant protection products [45,46,47,48]. The development of new fermentation technologies, including fermentation in bioreactors, allows for scaling up biopesticide production and reducing their costs, which increases their availability on the market [52]. There are three main groups of biopesticides: microbial pesticides, biochemical pesticides and plant-produced protectants (PIPs) [45,51,52]. Microbial pesticides refer to microorganisms such as bacteria, pathogenic fungi or viruses, and sometimes metabolites produced by bacteria or fungi. Entomicidal nematodes are often classified as microbial pesticides, although they are multicellular organisms. Biochemical (or herbal) pesticides are naturally occurring substances used to control pests and diseases caused by various microorganisms, or for monitoring in the case of pheromones [53]. The novelty in this area is the identification and synthesis of new, highly specific semiochemicals and pheromones that act at very low concentrations, minimizing the impact on the environment [54]. Plant-produced protectants (PIPs), which are produced by plants that have incorporated genes from other species into their own genetic material (GMOs), are problematic due to controversy, especially in European countries [45,51,52]. Despite the controversy, research on gene editing (e.g., CRISPR/Cas9) in plants to increase their natural resistance to pests and diseases, without introducing foreign genetic material, is a promising new avenue in the development of PIPs that may be more socially acceptable [55]. Biopesticides, although their biological activity, especially against fungi, nematodes, insect pests and other organisms, are well documented, have no known function in photosynthesis, growth or other aspects of plant physiology [45,51,52,55]. All vascular plant species have developed a unique, integrated chemical structure that protects them from pests. These biodegradable and renewable alternatives to conventional pesticides have been successfully used, especially in organic farming systems [20]. In order to explain the mechanisms of pesticide action, genetics and molecular physiology are used as tools to overcome plant resistance to new generations of chemicals and antimicrobials [44,48,49]. Human and animal exposure to pesticide residues involves substances originating from various sources, such as the environment or food. The adverse effects of such combinations can occur unexpectedly and manifest themselves in different ways, leading to a change in the assessment of health risks [3,46,50,51,52]. According to the latest research and international and European regulatory changes in this area, the analysis of pesticide residues used in plant protection products (PPPs) aims to determine the risk and precisely establish maximum pesticide residue limits and plan their control [8,44,50,51,52,53,54]. The novelty here is the development of fast and precise analytical methods (e.g., mass spectrometry techniques), which allow monitoring a wide spectrum of pesticide residues in real time, which is crucial for ensuring food safety [56]. Kowalska et al. [45] proved the high potential of 19 isolated strains of yeast-like microorganisms and their antagonism towards the causative agent S. sclerotiorum at different temperatures. They also found a high protective potential of yeast-like organisms Saccharomyces cerevisiae and their suitability for protective treatments against gray mold in strawberries, reduction of symptoms of Phytophthora infestans on potato plants, which proved useful for dressing planting material and increasing the shelf life of vegetables. In addition, these authors proved the differential survival of P. oligandrum and B. subtilis in different temperature ranges and their suitability for controlling the main potato pathogens in field conditions, in strategies using them as an alternative to copper preparations.
Pathogenic soil microorganisms can cause diseases or damage to crops. These microorganisms can also be used to control diseases, weeds and pests. Biopesticides are a group of active substances used in plant protection. They occur naturally or are synthetic products identical to natural substances. Biopesticides are a type of pesticide derived from natural materials such as bacteria, plants, animals and some minerals. They include many living organisms (so-called biocontrol organisms) [45,48,49,50,51,52,53,54,55]. They are usually less toxic than conventional pesticides. Their effects are typically low risk to non-target organisms. They tend to degrade quite quickly, and some semichemicals are used at very low doses. Their mechanisms of action and effects on human and animal health are divided into several groups or classes: biochemical biopesticides, microbial biopesticides, and Plant-Incorporated-Protectants (PIPs), which are pesticidal substances that plants produce from genetic material that has been added to plants [4,5,10,11,12,13,14,15,48,51,55]. Various natural substances, including plant extracts, are classified as biopesticides, including: semiochemicals from insect pheromones; various fermentation products such as Spinosad (a macrocyclic lactone); chitosan; natural products of plant origin (alkaloids, natural phenols, terpenoids and other secondary metabolites); some vegetable oils, e.g., rapeseed oil, as well as products made from plant extracts, such as garlic, onion, nettle, have pesticidal properties. They have been registered in the EU and elsewhere and can be used in plant protection. Pesticide residue limits in food are regulated by the Regulation (EC) on official controls of pesticide residues in food, both for products manufactured in the EU and imported [46]. Microbial biopesticides consist of a microorganism (e.g., bacterium, fungus, virus or protozoan) as an active ingredient and can control many different types of pathogens, but each active ingredient is specific to its target pest. For example, there are species of fungi that control certain species of weeds, or fungi that kill certain species of insects. This allows for the improvement of physicochemical and biological properties of the soil, increases plant resistance to pathogens and allows for the proper selection of biologically active compounds [53,54,55,56]. E.g., Bacillus thuringiensis subsp. tenebrionis destroys Leptinotarsa decemlineata larvae [44,55,56]. It is worth emphasizing that the new generation of microbiological biopesticides is designed considering the specificity of pathogen-host interactions at the molecular level, which allows for precise pest; control with minimal impact on non-target organisms (Figure 3) [16,48].
Figure 3 shows the key role of microorganisms in sustainable agriculture. The main elements are: A healthy plant: Its roots are surrounded by symbolic beneficial microorganisms, such as bacteria and fungi, indicating their role in plant nutrition and health. In the background, crop fields are outlined with visible features of sustainable practices, such as mixed cropping or agroforestry, symbolizing the broader context of application. In the margin, there is a small, schematic diagram of a biopesticide attacking a pest, visualizing the application of biological control.
Entomopathogenic fungi are micropesticides that attack insects and cause diseases inside their bodies, ultimately leading to death [36,37,38,39]. Two species of fungi are known to be used as micropesticides: Beauveria bassiana, which causes white muscardine, and Metarhizium anisopliae, which causes green muscardine [36,37]. B. bassiana, a filamentous fungus from the class Deuteromycetes (imperfect), is used against Colorado potato beetles, apple fruit moths, and American cuticle moths. It attacks the hemocoel of insects via spores. Once the spores attach to the insect cuticle, the fungus germinates and its hyphae penetrate the insect cuticle, forming an appressorium and a penetration peg. At the same time, they secrete chitinases, lipases, and proteases that can dissolve the cuticle. The fungal hyphae enter the hemolymph, multiply, and colonize the insect, releasing blastospores. Death of insects occurs due to depletion of nutrients in the hemolymph or poisoning due to the secretion of toxic metabolites [36,37,38,39]. New approaches also include the selection and engineering of entomopathogenic fungi strains with increased virulence, UV resistance and longer survival time in the environment, which is crucial for their commercialization [33,34,35]. The combined use of biologicals in combination with non-chemical weed control methods significantly improves soil structure, increases earthworm numbers and reduces soil carbon dioxide emissions, but still contributes to a decrease in soil enzymatic activity [42]. The implementation of integrated weed, disease and pest control in commercial agriculture has ultimately led to an increased interest in biopesticides, which in turn will reduce food safety risks [56]. Biochemical biopesticides are naturally occurring substances that control pests through non-toxic mechanisms that include reproductive disruptors such as insect sex pheromones, as well as various fragrant plant extracts that in turn attract insect pests to traps [53].
3.5. Bioherbicides
The priority of sustainable agriculture is food security and environmental security [53,57,58,59]. Already in 2001, Lasiodiplodia theobromae MTCC 3068 was used as a potential bioherbicide in pre- or post-emergence treatments, including Parthenium hysterophorus L. from the Asteraceae family, Datura stramonium from the Solanaceae family, and Sida prickly. The mycoherbicide produced from Lasiodiplodia pseudotheobromae C1136 is suitable for controlling both dicotyledonous and monocotyledonous weeds [23]. Pathogenesis induced by phytotoxins of pathogenic fungi can cause physiological, biochemical and metabolic changes in the host plant, and this leads to abnormalities in the translocation of storage materials, which in turn ends in chlorosis and aging of the leaves at the sites of infection [20,57,58,59]. This process usually leads to the degradation of genetic material in plant cells and growth inducer [24,25,50].
Recently discovered the effect of phytotoxic metabolites produced from bioherbicide isolates may significantly prevent environmental threats in the future [60]. Therefore, further research into bioherbicides and all aspects of the safety of phytotoxic metabolites produced from wild and genetically modified strains of L. pseudotheobromae and their bioherbicide effect on various species of weeds is justified.
Microbial biotechnology can be used to produce bioherbicides through the use of microorganisms that produce toxins that are harmful to weeds, or through the genetic modification of microorganisms to produce specific toxins [23]. One example of the use of microbial biotechnology for the production of bioherbicides is the use of herbicide-tolerant microorganisms. Many plants produce compounds that are toxic to weeds, and these compounds can be extracted and used as bioherbicides. Microbial biotechnology can be used to produce plant extracts with herbicidal properties through the fermentation of plant material or through the genetic modification of microorganisms to produce specific compounds [23,24]. In addition to the use of microorganisms and plant extracts, microbial biotechnology can also be used to produce bioherbicides from minerals. For example, sulfur, a naturally occurring mineral, is commonly used as a bioherbicide to control fungi and bacteria in agriculture, forestry, and horticulture. Sulfur compounds (SO2 and H2SO4) are often used as bioherbicides to reduce fungal and bacterial diseases in orchards (e.g., apple scab, black spot or downy mildew). This is about sulfur compounds as fungicides, not about their use as herbicides in relation to diseases. There is a slightly misleading formulation used, sulfur is not a typical bioherbicide for weeds, but rather a fungicide. Overall, the use of microbial biotechnology for the production of bioherbicides has the potential to provide numerous benefits, such as reducing the reliance on synthetic herbicides, improving the safety of weed control, and reducing the environmental impacts of weed control [23,30,38].
One challenge that needs to be dealt with when applying microbiological biotechnology in the production of bioherbicides is the possibility of developing resistance in various weed populations. Like synthetic herbicides, bioherbicides can select for the survival of weeds that are resistant to the toxins or other compounds used in the bioherbicides. This can lead to the development of weeds that are immune to bio-herbicides, which can reduce their effectiveness over time.
In the system of biodynamic and organic agriculture, there are no effective methods of combating plant diseases and pests [55,57,58,59,60,61]. Vaitkeviciene [56] and Levickienė [61,62] used the biodynamic preparation BD 500, in organic cultivation. It has been proven that some cultivars have different abilities to reduce weed infestation when grown in varietal mixtures compared to varietal monoculture [64,65]. For example, some tomato varieties are more tolerant to Cuscuta spp. [66].
In the case of bioherbicides, in the vast majority of cases the pathogen was a fungus, as preparation of mycelium, conidia or spores. Only a few live microbial bioherbicides have been approved for commercial use and have been placed on the market, but their use has unfortunately been very limited for a number of reasons, mainly economic. Examples of bioherbicides are listed in Table 1.
The advantages and disadvantages of live bioherbicides are presented in Table 2. If the bioherbicide is not genetically modified or contains any synthetic chemicals, it can be accepted by organic farmers and thirdly, such a product must not leave any chemical residues in the crop [53,54,55,56,62,67,68]. Another issue is the safety of the tested strains of bacteria for humans [60,61,67].
Varieties of plants with high tolerance to weeds are important in special situations where chemical control of weed infestation is impossible. Such species or varieties are characterized by strong allelopathic properties [67,68]. Secondary metabolites involved in weed control include phenols, flavonoids and alkaloids [63,69]. Such properties of some species and varieties of crop plants are extremely valuable, especially in integrated farming systems, by inhibiting the development of weeds [70]. In conclusion, microbial biotechnology can play a significant role in the production of bioherbicides through the use of microorganisms, plant extracts and minerals. However, various challenges need to be considered, such as the potential for resistance development in weed populations and the need to better understand the environmental impact of bioherbicides.
3.6. Bioinsecticides
Bioinsecticides are insecticides made from natural materials, such as microorganisms, plant extracts, and minerals. Microbial biotechnology can be used to produce bioinsecticides through the use of microorganisms that produce toxins that are harmful to insects, or through the genetic modification of microorganisms to produce specific toxins [71,72,73,74,75,76,77].
One example of the use of microbial biotechnology for the production of bioinsecticides is the use of Bacillus thuringiensis (Bt). Bt is a soil bacterium that produces toxins that are harmful to certain insects, such as caterpillars and mosquitoes. These toxins are called delta-endotoxins and are toxic to insects when ingested but are generally considered to be safe for humans and other mammals [78,79,80,81,82].
Biopesticides are one way of biological control. The active ingredients of biopesticides include microorganisms, either plant extracts or naturally occurring chemicals (e.g., potassium bicarbonate). As a result, some pest control methods (modes of action or MOAs) differ from traditional, artificial chemical pesticides. Biopesticides with MOA can counteract insect pests (e.g., preparations containing Beauveria bassiana) or against plant diseases (e.g., containing Paraconionthyrium minitans strain CON/M/91-08). Many biopesticide preparations with MOA contain fungal spores. These spores germinate only when they are on an insect or on a pathogen that causes disease (Figure 4) [79,80,81,82].
Biopesticide preparations with MOA contain fungal spores. These spores only germinate when they are on an insect or on a disease-causing pathogen.
A biopesticide containing viable fungal spores (blue) that must be on an insect pest or plant pathogen (yellow rectangle). They then germinate, attack and grow, eventually killing the pest (Figure 3). Another example is the use of plant extracts with insecticidal properties as bioinsecticides. Many plants produce compounds that are toxic to insects, and these compounds can be extracted and used as bioinsecticides. For example, pyrethrum, a compound produced by Chrysanthemum sp. flowers, is used as a bioinsecticide to control a variety of insects, including mosquitoes, flies, and beetles. Microbial biotechnology can be used to produce plant extracts with insecticidal properties through the fermentation of plant material or through the genetic modification of microorganisms to produce specific compounds [77,82,83].
However, there are still various challenges and ethical issues to consider, such as the possibility of GMOs being released into the environment. In addition, an important challenge is the possibility of developing resistance in insect populations. Like synthetic insecticides, bioinsecticides can select for the survival of insects that are resistant to the toxins or other compounds used in the bioinsecticides [9,10,11,12,13,14,15]. This fact may lead to faster development of insect populations resistant to bioinsecticides, which in turn may reduce their effectiveness. To mitigate this risk, it is important to use integrated pest management (IPM) strategies that incorporate multiple control measures, such as the use of diverse bioinsecticides, cultural practices, and natural predators, to reduce the selection pressure for resistance. Kowalska et al. [45] demonstrated the huge role and the scope of usefulness of beneficial microorganisms that occur naturally or were deliberately introduced into the environment to ensure the well-being of plants in the ecological farming system. Among others showed a highly competitive potential of the antagonistic fungus T. asperellum used for spraying potatoes, strawberries, rapeseed and lemon balm and used in storage against Botrytis cinerea and S. sclerotiorum [43,77]. The high protective potential of yeastlike organisms Saccharomyces ceravisiae and the suitability for protective treatments against gray mold in strawberries, reducing the symptoms of Phytophthora infestans on potato plants [45].
Viral insecticides (Baculoviruses) are pathogens that attack insects and arthropods. Viral pesticides are used to control Lepidoptera larvae such as Helicoverpa, Spodoptera sp. on cotton, maize, sorghum and tomatoes. Baculoviruses are a commonly used viral biopesticide. They are microscopic in size and consist of double-strand DNA. The genus Baculoviruses includes 3 subgroups: Nuclear polyhedrosis viruses (NPV), Granuloviruses (GV); non-occluded viruses. The NPV virus enters the insect body through ingestion by insects and infects midgut cells through membrane fusion. NPV develops in the cell nucleus and passes through the intestinal epithelium of the insect, causing a systemic infection of the hemocoel [26,45].
Another challenge is the need for further research to better understand the environmental impacts of bioinsecticides [83,84,85]. Therefore, it is important to carefully assess the environmental impacts of bioinsecticides and to implement appropriate measures to mitigate any potential negative impacts. In conclusion, microbial biotechnology has the potential to play a significant role in the production of bioinsecticides through the use of microorganisms, plant extracts, and minerals. It is important to carefully assess the risks and benefits of using bioinsecticides and to implement appropriate measures to ensure their safe and effective use.
3.7. Effective Microorganisms
In the face of drastic climate changes, the growing world population and growing interest in safe food, there is a growing demand for substances capable of replacing or eliminating conventional pesticides [56,84,86,87,88,89,90,91,92,93,94,95,96,97,98,99]. This is of crucial importance for both consumers and producers striving to reduce the use of chemicals in agriculture, e.g., by introducing safe preparations that are an alternative to synthetic fertilizers and conventional plant protection products. Such an alternative is Effective Microorganisms (EM). EM is a mixture of many microorganisms, such as bacteria, fungi, actinomycetes and yeasts, commonly found in the environment (Figure 5) [57,58,59,100,101,102,103,104,105,106,107,108,109,110,111].
Recent studies focus on precise characterization of the composition of EM consortia using metagenomic techniques and identification of key microbiological interactions that are responsible for their synergistic effects and effectiveness in different soil environments [55].
3.7.1. Microorganisms in the Influence of Plant Resistance
The use of microbiological preparations in plant production raises many controversies in the world. There are reports of both benefits from their use [57,58,59] and their ineffectiveness [112,113]. Modern approaches aim to standardize application methods and environmental conditions in which EM show the highest effectiveness in order to reduce the variability of field results [104].
The main strategy of biological plant protection is induced plant resistance. Induced plant resistance is defined in two ways: (1) as Induced Systemic Resistance (ISR) when induced by rhizobacteria (PGPR) and (2) as Systemic Acquired Resistance (SAR) when induced by other factors (e.g., pathogen infection). SAR is expressed at its highest level when the inducing organism causes necrosis, whereas ISR (induced by PGPR) does not show necrosis symptoms in the host plant. Both ISR and SAR are based on the activation of dormant immune mechanisms [70]. The development of molecular techniques such as transcriptomics and metabolomics allows for a deeper understanding of the specific signaling pathways and molecules involved in ISR and SAR, which enables more precise design of biostimulants [114].
Race-specific avr gene products (elicitors) are active only in varieties with complementary resistance genes (R). They then determine the occurrence of race-specific resistance (full resistance). In the case of acquired systemic resistance, i.e., in order to activate the defense system in uninfected parts of the plant, inhibitors are used – i.e., plant defense proteins exhibiting inhibitory activity, e.g., polygalacturonase (PGIP). A total of fourteen plant proteins have been identified [70,102,103], mainly with bactericidal activity, also involved in apoptosis. These are gluconases, chitinases, osmotins, protease inhibitors, proteinases, lysozymes, peroxidases. Numerous interactions are known between the cell walls of fungi and plants, and the expression of the encoding genes depends on the elicitor, which is captured by specific receptors, signaling their induction via kinases. The typical cell wall of dicotyledons contains cellulose (β-1,3; 1,4-glucans), hemicellulose - 30% (xyloglucans), pectin’s - 35% and proteins 1-5%. The typical cell wall of grasses (monocotyledons) contains cellulose - 25% (β-1,3; 1,4-glucans), hemicellulose 55% (arabino-glucuronoxylans and glucomannans), hemicellulose 30% (xyloglucans), pectin’s - 35% and proteins - 1-5%. In turn, the cell wall of fungi contains: β-1,3-glucans - 30-65%; β-1,6-glucan 1-5%, chitin - 2-40%; CWP proteins 30-50% [102]. Common and race-specific elicitors are distinguished. The former (i.e., β-glucans, chitin, polypeptides, glycolipids) are produced by pathotypes of the pathogen. They are responsible for inducing the so-called race-nonspecific immunity (field, horizontal, partial). Race-specific elicitors (products of a virulent genes avr) are active only in varieties with a complementary resistance gene (R). They are responsible for the development of “gene-to-gene” immunity, i.e., race-specific (vertical, complete) [70,102]. Elicitors trigger a cascade of defense reactions. This leads to an increase in the content of phenols, flavonoids and the formation of secondary metabolites (phytoalexins). They can be toxic to pathogens [102]. The signaling cascades (1) salicylic acid (SA) and (2) jasmonic acid (JA) are shown in Figure 6.
This schematic diagram shows the complex interactions between two major signaling pathways in plant defense: the salicylic acid (SA) pathway and the jasmonic acid (JA)/ethylene (ET) pathway. On the left side, the SA pathway is activated by biotrophic pathogens or elicitors of acquired systemic resistance (SAR). It leads to the activation of pathogenesis-related genes (PR) and consequently to resistance to biotrophic pathogens. On the right side, the JA/ET pathway is induced by necrotrophic pathogens, herbivorous pests or elicitors of inducible systemic resistance (ISR). This results in the activation of defense genes and resistance to necrotrophs and pests. Figure 5 also highlights the antagonistic interactions between these two pathways (marked with dashed lines with minus signs), which is crucial for the fine-tuning of plant defense responses. The central element shows that environmental signals and abiotic stresses also affect both pathways, modulating their activity. General components, such as MAPK cascades and transcription factors, involved in these processes were also identified.
The main consequence of pathogen/elicitor recognition by a plant cell is rapid reactions at the cell periphery that do not require gene expression. They include membrane depolarization, ion flow across the membrane (including K+ and Cl- efflux and H+ and Ca2+ influx), and an oxygen burst. The gene-for-gene theory states that pathogen recognition and activation of defense responses occur only when a plant possessing the appropriate resistance gene R is attacked by microorganisms that contribute to the avirulence gene (avr). A pathogen with the avr gene is pathogenic for plants that do not possess the appropriate R gene, because then there is no defense reaction. Studies on the influence of microorganisms on plant immunity processes were undertaken due to the rapid increase in active oxygen species after contact of plant cell membranes with the pathogen. They are cytotoxic, especially for cells infected with the pathogen. This results in cytoskeleton reorganization and increased thickness of cell walls. According to Kowalska et al. [45] and Barbaś et al. [103], acquired systemic immunity, activating the defense system, is acquired by the plant in its healthy, uninfected tissues. The signal to initiate this response is usually salicylic acid (Figure 5). At least fourteen classes of plant proteins are involved in this process, such as: chitinases, glucanases, protease inhibitors, lysozymes, peroxidases (including β-glucosidase), osmotins, proteinases, antibacterial proteins and proteins involved in apoptosis [102,103]. In the places of action of microbiological preparations, as a result of biochemical changes, targeted resistance of crop plants may occur. Recently, the demand for microbiological preparations in the agricultural sector has been gradually increasing due to the development of ecological and pro-ecological methods of plant cultivation [46,96,104,105,106,107,108].
The demand for microbiological preparations is growing in agriculture due to the desire to reduce chemicalization [45,47,109,110,111]. Preparations with beneficial microorganisms, including EM, are widely used in Europe, supporting the growth and yield of crops, such as bacterial symbioses with legumes or mycorrhizal vaccines in forestry [45,47,109,110,111]. However, the registration procedure for these products for organic farming is often too liberal, and there is often a lack of studies confirming their effectiveness [45,47,109,110,111].
3.7.2. EM in Plant Cultivation
The use of bacterial bioproducts is wide and includes:
Supporting the microbiological balance of the soil thanks to the optimal composition of strains [46,98,99,100,101,102,103].
Accelerating mineralization and increasing the availability of nutrients, which reduces the need for high doses of mineral fertilizers [104,105,106,107,108].
Decomposition of difficult-to-degradable plant parts and activation of the humification process, which improves air-water relations in the soil, increases nutrient absorption, maintains soil moisture, prevents soil compaction and protects plants from biotic and abiotic stresses [109,110,111,112].
Helps maintain the lumpy structure of the soil.
Biopreparations are crucial for soil fertility, and their production in bioreactors ensures high concentration of bacteria and full control over the process [97,113,114,115,116,117,118].
EM are tested in many countries, including Japan, China, Poland, and their aim is to improve the quality of soil, water and agricultural products [111,116,117,118,119,120,121,122,123]. Studies have shown that EM with manure improves root growth and photosynthesis [83], increases the yield and health of potatoes [94,95] and wheat [74]. Other authors also confirm the positive effect of EM on yield and plant health [78,80,81,82,83]. Some studies, e.g., Van Vliet et al. [110], did not show a significant effect. Martyniuk [111] criticizes the liberal registration procedure, which indicates the need for further research on the mechanisms of EM action.
EM in vegetable cultivation increased the yield of onion and pea [104], affecting photosynthesis and water use. Pniewska et al. [136] observed increased density and mass of green beans. EM improved growth and physiological parameters and also increased the yield of beans [136]. Górski and Kleiber [86] showed a positive effect of EM on ornamental plants (roses, gerberas), and Trawczyński [80] on the content of nutrients in potatoes. EM strengthens plant immunity and displaces pathogens [45], inhibiting the development of cereal diseases [79,82,121] and reducing the occurrence of Fusarium fungi [104]. The use of EM together with nettle extract reduced the number of potato beetle larvae [71].
Microbiological preparations fill the gap in biological plant protection and increase soil microbiological activity [71,91,96,110,124,125,126,127]. They have yield-forming potential and the ability to transform harmful compounds [71,91,96,110,124,125,126,127]. They can be used for seed dressing, spraying and soaking seedlings [51,103,118,122,128].
Some authors [109,129,130,131] confirm the positive effect of EM preparations on morphological, physiological and immune features of plants. Knowledge of this subject comes mainly from commercial studies and fragmentary scientific research. In order to support the fertility and quality of crops, close cooperation and exchange of experiences between agricultural practice and the scientific community is necessary [132,133].
Systems and technologies using microorganisms have many advantages and disadvantages that should be considered (Table 2).
Powerful scientific techniques caused a dramatic expansion of genetically modified crops, leading to a change in agricultural practices, which has a direct and indirect impact on the environment (Figure 7).
Despite the increased yield potential, the basic problems to be dealt with are threats and concerns about biological security related to such crops of the commune. Among the scientists and decision-makers, the growing interest in the study of unintentional effects of transgen related to gene flow, the flow of naked DNA, weeds and chemical toxicity can be seen. Current knowledge proves that GMO crops exert a harmful impact on the environment, e.g., by changing the invasiveness of crops, the appearance of herbicides and insecticides, tolerance to salt stress or drought stress, disturbing biological diversity and laying transgentransgene, etc. This impact requires confirmation and Critical research. Currently, there is not enough data on a significant negative impact of GM plants or their nutritional value. Therefore, the consumption of GM products is considered to some extent safe to consume, with a few exceptions. Meanwhile, Tsatsakis et al. [135] They change the existing arrangements and GM crops, and their products are divided into target and underdoer species and shed new light on the challenges and threats associated with this problem. However, basic research also indicates that the influence of GM crops on biological diversity disorders, immunity development and evolution somewhat resemble the effects of genetically unmodified crops (Figure 6). The future prospects are also important.
3.7.3. Microbiological Preparations in Composting
Due to the growing problems of managing natural fertilizers and bio-based municipal waste, especially sewage sludge, the methods of their disposal are diverse [10]. The EU is observing the development of legislation on bio-waste and the implementation of modern technologies for their management, especially recovery. Among the popular technologies for processing organic waste (including natural fertilizers) are energy recovery and aerobic technologies, such as various forms of composting [106,116,126,131,132,133,134].
A potential application of EM is composting. EM preparations can improve the quality of compost by increasing the activity of microorganisms, improving the nutrient balance, reducing unpleasant odors and fly populations [2,93,94,95,96,98,106,126]. Studies by Dacha et al. [106] on composting of sewage sludge with EM addition showed that the thermophilic phase was more intensive without EM addition, and microbiological activity (CO2 emission) was also higher in control compost. Nitrogen losses due to ammonia emission were 0.04% lower in the EM treatments [106]. The lack of EM effect could be due to competition with native microflora or parasitism/antibiosis phenomena [106]. Composting, especially of sewage sludge with a low C:N ratio (<10) [116], can lead to gas emissions, therefore it is important to optimize the composition of the mixture by adding materials rich in carbon (e.g., straw, sawdust) [106]. Despite the promotion of EM in agriculture and municipal economy, their role as a panacea for all problems (fertilization, plant protection, crop quality, feed storage, reduction of ammonia and odor emissions, acceleration of composting) should not be overestimated. Further research is needed to better explain the role of EM [106].
3.7.4. EM in Food Processing
EM technology is one of the biotechnologies used in the food industry [95,96,97,105,107,117,118,121,135,136]. It uses microbiological materials (e.g., “Baikal EM1”), such as photosynthetic bacteria, lactic acid bacteria and nitrogen-fixing bacteria, as well as yeasts and molds [118]. Modern food processing is based on enzymatic and microbiological preparations. More than 100 enzymes are used, and their number is growing [9,10,11,12,13,14,15]. Many microbiological preparations, including EM, are also used [118,135,136].
The benefits of using enzyme and microbiological preparations in the food industry are wide, including accelerating technological processes, the possibility of producing new assortments (including functional foods), increasing the attractiveness and durability of products, increasing the efficiency of raw materials and reducing production costs [101,121,137,138,139]. This allows us to obtain higher quality products at lower costs. Some microbiological preparations, such as yeast for wine, beer and bread production [33,40], or lactic acid bacteria for cheese and dairy products [46,47,135,136,137,138,139,140], are used even in households. Acetic acid bacteria are used for vinegar production [89,101]. EM technology, used e.g., in potato cultivation, can reduce digestive system ailments in humans by reducing the absorption of fats during frying potato products and obtaining better quality of the raw material [98,118,122].
3.7.5. Mycorrhizal Preparations
Mycorrhizal preparations contain mycorrhizal fungi that inoculate plant roots, creating a symbiosis called mycorrhiza. In this interaction, fungi use photosynthetic products, and the plant receives water and minerals (nitrogen, phosphorus, potassium, microelements) [33,34,35,36,37,38,39]. The key functions of mycorrhiza are the protection of plants against fungal pathogens, increased resistance to frost and salinity, more abundant flowering, better rooting, and improved plant growth and appearance [37,38,39]. Mycorrhizal fungi are applied once, and plants can benefit from mycorrhiza throughout their lives. Early application (e.g., in seedling production) improves plant adaptation after transplanting [70,141].
In the context of climate change and long-term droughts, the use of arbuscular fungi (AM) is an effective and sustainable strategy [140]. Correira et al. [141] showed that AM fungi affect osmoregulation in H. serratifolius plants under conditions of moderate water deficit via soluble sugars and amino acids, and under severe water deficit via proline and reducing sugars [141]. AM fungi promote greater absorption of phosphorus and nitrates only under conditions of good water supply, increasing dry mass in both water deficit and water supply conditions [141]. Mycorrhizal colonization is directly related to water regime, with well-watered plants showing higher colonization rates and plants with severe water deficit – lower [141]. Recent studies have focused on identifying plant and fungal genes responsible for the efficiency of mycorrhizal symbiosis under abiotic stress conditions, which will allow for the selection of more resistant genotypes [142].
3.7.6. Microorganisms in Aerobic Nitrification and Denitrification
Both heterotrophic nitrifying and aerobic denitrifying microorganisms (HNADM) can oxidize ammonium nitrogen and then denitrify it in the presence of oxygen [141]. HNADM may offer better utilization options [131,141]. To improve the efficiency of nitrogen cycling in agriculture, EM can be used simultaneously in nitrification and denitrification (SND) and HNADM processes [125,126,142,143].
Climate change and drought stress pose challenges to water management, food security, and water and wastewater treatment technologies [no citation in the source text]. Monitoring nitrogen pollution, mainly from nitrogen fertilizers, remains a challenge in wastewater treatment [144]. Nitrogen compounds still cause many environmental problems, because their excess leads to deterioration of air quality, eutrophication of waters and changes in forest processes, which in turn negatively affects water quality, human and animal health and the environment [143]. In practice, biological methods are more often used. In the technology of biological nitrogen removal, two stages are distinguished: aerobic nitrification and anaerobic denitrification [145].
Denitrification is carried out by heterotrophic microorganisms, and nitrates are first converted to nitrites, and then to NO, N2O and finally to N2 [146,147,148]. The discovery of heterotrophic nitrifying bacteria and aerobic denitrifying bacteria disproved this concept [145,146,147,148]. SND bacteria have the ability to both nitrify and denitrify. SND technology has significantly reduced costs and improved the efficiency of denitrification. Simultaneous heterotrophic nitrification and aerobic denitrification (SND) for nitrogen removal have been gaining increasing public interest due to their high efficiency and low cost [149].
Combined research strategies are proposed for future research directions, including the study of the nitrogen removal mechanism of SND using multi-topic strategies. Understanding this mechanism will allow for more efficient and better utilization of SND for nitrogen removal, e.g., from municipal or pharmaceutical wastewater [126]. Simultaneous nitrification and denitrification reactions can occur spontaneously in practice and are widely distributed in the environment [126,149], therefore simultaneous heterotrophic nitrification and aerobic denitrification (SND) play an important role in wastewater treatment plants and nitrogen removal reactors [150]. For example, Acinetobacter sp. is already used in full-scale A2O operation for nitrogen removal from pig farm wastewater (Table 3, Figure 8) [151,152].
This graph visualizes the relative contribution of different strains of bacteria capable of simultaneous nitrification and denitrification (SND) to nitrogen transformation processes. It shows the contribution of individual strains (A, B, C, D, F) and their overall contribution to denitrification and nitrification, emphasizing the role of these microorganisms in the nitrogen cycle.
Recent research in the area of HNADM/SND focuses on microbial engineering to select and optimize strains capable of more efficient nitrogen removal under different environmental conditions, as well as on integrating these technologies with existing wastewater treatment systems to maximize efficiency and minimize carbon footprint [152].
3.7.6. EM and Germanium (Ge)
Germanium (Ge) is a semimetal with atomic number 32. Its organic form is banned in Europe for therapeutic purposes. Ge is associated with oxygen transport in the blood, acting as an excellent oxidant and antioxidant, protecting against aging and degenerative diseases [161]. Some attribute to it the ability to stimulate immune function, deliver oxygen to cells, detoxify, protect against radiation, and inhibit tumor growth [161].
Soil contamination with metalloids such as Ge is a global problem affecting human, plant, and animal health. Even low concentrations are dangerous. Metalloids can significantly affect the structure and biodiversity of soil microbial communities, which can lead to changes in ecosystem functioning [161].
EM can be used with other techniques, such as Germanium. Ge has been shown to have beneficial effects on plants, increasing their growth and stress tolerance. Combining EM with Ge enhances the uptake and utilization of Ge by plants, leading to increased growth and improved yields [162,163].
Overall, the use of EM has the potential to bring many benefits to sustainable agriculture, such as improving soil health, reducing dependence on synthetic chemicals, and increasing crop yields. However, ethical issues need to be considered, such as the fear of releasing GMOs into the environment, and the need for further research to determine the possibilities and limitations of this technology [162,163].
In summary, microbial biotechnology has the potential to play an important role in the use of EM in various applications in sustainable agriculture, such as biological crop protection, composting, and improving nitrogen cycling [164]. However, there are still important ethical issues, such as the possibility of releasing GMOs into the environment, and the urgent need for continued research to better understand both the potential possibilities and limitations of this technology. In sustainable agriculture, the assessment of both benefits and risks associated with the use of EM preparations is important for food safety, consumer safety and the human environment [no citation in the source text].
3.7.8. Inhibition of the Growth of Various Pathogenic Microorganisms Using EM
EM is increasingly used in various areas of life, e.g., in environmental protection [136]. Biological wastewater treatment processes depend on mixtures of microbial communities, including bacteria, fungi, rotifers and algae. Municipal wastewater contains a wide range of pathogenic and non-pathogenic bacteria, including e.g., Salmonella, which can cause diseases in humans and animals [144,159]. Fungi may also occur in wastewater, the presence of which is highly undesirable. These are multicellular, non-photosynthetic organisms, mainly aerobic, but anaerobic species are also known. Due to their different cellular composition, they tend to dominate over bacteria in various wastes and wastewaters poor in nitrogen or with low pH. They may have a filamentous form, which makes their removal by sedimentation techniques difficult [144,145]. A promising technology in wastewater treatment from undesirable microorganisms is EM technology [159].
Mathews and Gowrilekshmi [126] proved that the application of EM to stagnant rice water changed its physical, chemical and biological properties. Chemical parameters (pH, TDS and BOD) showed a decreasing trend. On the other hand, salinity, conductivity and dissolved oxygen increased. Moreover, it was noticed that under the influence of EM technology there was almost a 2-fold reduction in the population of Lactobacillus, Actinomyces and yeasts in the first 2 weeks after EM application [126].
EM used locally in Iraq [114] proved to be one of the environmental solutions for reducing water salinity and thus improving its quality, both in soil and sewage systems. Mouhamad et al. [114] showed that the effectiveness of EM technology can help farmers use drainage and groundwater for irrigation and make better use of water resources [114].
3.8. Microbiological Organisms in Sustainable Agriculture
Microbiological organisms, including bacteria, fungi, viruses and yeasts play an improvement role in sustainable agriculture. They are involved in various processes, such as nutrient cycling, nitrogen fixation, infestation and pest control, and also in weed management, disease management and fertilization management. They can provide numerous benefits, such as improving soil health, increasing crop yields, and reducing the reliance on synthetic chemicals. For example, of the use of microbiological organisms in sustainable agriculture is the applied of nitrogen-fixing bacteria, such as i.e., rhizobia, to improve the nitrogen content in the soil. These macroelements are an essential nutrient for plant growth and development, and nitrogen-fixing bacteria can convert atmospheric nitrogen into a form that plants can use [137,138,140].
This can reduce the need for synthetic nitrogen fertilizers, which can be costly and have negative environmental impacts. Another example is the use of mycorrhizae, a group of fungi that form symbiotic relationships with plants, to improve nutrient uptake and water retention. Mycorrhizae can improve the efficiency of nutrient uptake by extending the root system of plants and by increasing the surface area for absorption. They can also improve water retention by increasing the water capacity of the soil and reducing water loss due to evaporation. In addition to the use of nitrogen-fixing bacteria and mycorrhizae, microbiological organisms can also be used for pest control. For example, bacteria such as Bacillus thuringiensis (Bt) can be used as bioinsecticides to control insect pests, and fungi such as Beauveria bassiana can be used as biofungicides to control fungal diseases [2]. Overall, the use of microbiological organisms in sustainable agriculture has the potential to provide numerous benefits, such as improving soil health, increasing crop yields, and reducing the reliance on synthetic chemicals [1,3].
EM has a comprehensive effect on the environment of crop plants, protecting them against pests and stimulating their resistance to diseases and unfavorable abiotic and biotic conditions. You can apply before sowing for seed bio conditioning, as well as foliar and soil application [44,87,88,89,91,108,109]. In order to better protect the crop environment against weeds, it is beneficial to propose the use of bioherbicides in weed control, knowing their sources, sources, target weeds and ecosystems (Table 4).
Soil is a natural habitat for numerous organisms coexisting and protecting each other. Rich in organic and nutritional substances, appropriate air-water ratios, having an appropriate reaction, creates an optimal environment for the life and development of microorganisms, which, with the participation of enzymes, stimulate the transformation of organic and mineral components in the soil. Thus, they participate in the creation and shaping its fertility, enrich it with biogenic elements, growth substances, antibiotics and other biologically active substances. This is how they influence the conditions for plant growth and development [1,2,41,54,98]. I’ll-considered management of organic substances with microorganisms present in it leads to destruction, erosion and decrease in soil fertility [98,110,115]. In order to eliminate unfavorable changes in the soil environment, attention is drawn to the need to increase the number of beneficial microorganisms by inoculating the soil with various microbiological preparations, such as UG-Max [108,109] or EM preparations [104,107,125]. Their development leads to the enrichment of microflora characterizing a given soil [54,60,61].
3.9. Soil and Microorganisms: Basics and Impact of EM
Soil is a natural environment for countless organisms, coexisting and protecting each other. Rich in organic and nutritional substances, with appropriate air-water relations and proper pH, it creates optimal conditions for the life and development of microorganisms. These, with the participation of enzymes, stimulate the transformation of organic and mineral components in the soil, participating in the formation of its fertility, enriching it with biogenic elements, growth substances, antibiotics and other biologically active compounds. In this way, they affect the conditions for plant growth and development [1,2,41,54,98]. Improper management of organic substances with microorganisms leads to degradation, erosion and a decrease in soil fertility [98,110,115]. To counteract this, the need to increase the number of beneficial microorganisms by inoculating the soil with various microbiological preparations, such as UG-Max [108,109] or EM preparations [104,107,125] is emphasized, which leads to the enrichment of soil microflora [54,60,61].
Kaczmarek et al. [89] showed that microorganisms in microbiological preparations increased the number of bacteria, fungi, actinomycetes and copiotrophic microorganisms in light soil, while inhibiting oligotrophs. Gałązka [54] showed that EM1 and EM2 preparations only affected the growth of cellulolytic bacteria. Kaczmarek et al. [88] proved that EM preparations stimulated the activity of soil dehydrogenases and supported its respiration. Gałązka et al. [54] found that EM affected the physical and water properties of soil material from arable-humus horizons of mineral soils, causing a decrease in density while increasing porosity [89]. Furthermore, the use of EM on soils with a heavier mechanical composition improved their natural drainage, allowing for faster drainage of excess gravitational water while maintaining high field water capacity [87]. Kaczmarek et al. [88] observed an increase in the maximum and minimum capillary capacity. The addition of EM also increases the rate of water filtration in soils with a heavier granulometric composition and reduces it in light soils. These changes have a positive effect on field conditions. A positive effect of EM on the compressive strength of soil aggregates, an increase in their dynamic and static water resistance, and an increase in the quantity and quality of secondary aggregates were also found, which limits soil surface crusting and erosion [54]. EM consists of two-layer gel capsules with an anaerobic interior and an aerobic external environment, limited by a zone of facultative microorganisms [1,54]. EM is absorbed on the surface of mineral particles, and the attached capsules repel soil particles, affecting the loosening of its structure [1,54]. Shah et al. [2] showed that the use of meat and bone meal with EM addition reduces the caking and spraying index and increases the soil structure index. Quiroz et al. [124] noted a beneficial effect of EM on the chemical properties of the tested soil. Ji et al. [73] claim that EM, by increasing the number of beneficial microorganisms, contributes to faster mineralization of organic carbon.
Kaczmarek et al. [88,89] report that EM applied to lessive soil contributes to the increase in the amount of available potassium, magnesium and mineral nitrogen, but causes a decrease in its buffering capacity, which was confirmed by the studies of Pszczółkowski et al. [94,95]. In chernozem, a decrease in the content of macroelements, mineral nitrogen and hydrolytic acidity was observed, as well as an increase in buffering capacity with the increase in the dose of microbiological preparation. However, Sawicka et al. [97] did not find significant effect of EM on the increase in the amount of organic carbon, humus and pH.
Effective microorganisms fulfill their role especially in heavily degraded, damaged and poor soils, where there is a lack of microorganisms necessary for the proper development and even survival of plants in unfavourable conditions [102,116,141,142,143,144]. According to Sawicka et al. [6], in soils degraded by the mining industry, the use of EM in perennial legume crops brings good results.
EM are easy to produce and use, safe for humans, animals and the environment, are low-cost, effective and can contribute to the sustainable improvement of agriculture and the environment [1,6,114]. Cultivation technology using EM preparations is currently used where it has been found that EM is the best way to replace traditional cultivation and management methods, both in agriculture, forestry, horticulture and in water management, using natural methods [1,2,27,99,114,126,144,166].
3.8.1. Soil Microorganisms Enhancing Plant Health
Many researchers have taken a multi-faceted approach to the role of soil microorganisms, including diatoms [42,43,44]. However, a few scientists have tried to determine the potential and mechanism of action of diatom organisms, Stanek-Tarkowska et al. [63] studied the effect of short-term fertilization with ashes from biomass combustion on microbial communities, mainly diatoms, occurring on the surface of lessive soil. As a result, they identified 23 diatom species, including mainly: Hantzschia amphioxis, Manam atomus, Mayamaea allowis, Nitzschia pusilla, Pinnularia obscura, Pinnularia schoenfelderi, but the most numerous populations were definitely Stauroneis thermicola. They showed that the main factor determining the biodiversity of diatoms was soil moisture.
The role of beneficial microorganisms, occurring naturally or intentionally introduced into the environment to ensure plant well-being, is indisputable. The mechanism of their action is based on increasing plant tolerance to stress conditions by supporting root growth, plant development and inducing their resistance. Microorganisms occurring in the soil or introduced into it in the form of microbiological preparations are an excellent alternative to conventional, chemical methods of protection in plant cultivation. The results of these authors confirm that plants, under the influence of contact with beneficial diatoms, bacteria or fungi, produce secondary metabolites. In this respect, this is very important, because diatoms react relatively quickly to chemical changes in the environment [44,105].
Studies on the diversity of diatom communities have shown that these organisms are sensitive to anthropogenic factors, such as soil pollution, plant protection products, fertilization, plant growth regulators, which disrupt their development. Terrestrial diatoms appear to be sensitive to many environmental factors, such as anthropogenic disturbances, soil pH and moisture, or nitrogen content in the soil. Meanwhile, most aquatic diatom species are well known, and their autecology is well known, qualifying them as biological indicators of water purity. This significant potential has not yet been exploited for terrestrial diatoms [126].
Pioneering discoveries were made on autecological values for soil pH and moisture for common terrestrial diatoms and compared with previous indicator values and tolerances. Data from ecological studies conducted under different climatic conditions were compared and it was found that validation indices improved significantly after removing samples taken from anthropogenic, disturbed habitats. This suggests that anthropogenic disturbance is a major factor determining the occurrence of taxon. International work by scientists in this field has been successful and has now allowed extending the list of terrestrial diatoms with taxa that can be treated as environmental markers in various soil studies. Stanek-Tarkowska et al. [63] discovered a new, undescribed species of the genus Microcostatus in Poland and named it Microcostatus dexteri sp. nov., describing it in the paper “Description of a new diatom species—Microcostatus dexteri sp. nov. from terrestrial habitats in southern Poland”. This species is small (3-9 μm) and rarely encountered. It is the fourth species described by this author recently. Discovering and describing new diatom species is very important for science. Studies on soil diatom communities developing in the surface layer of the soil are niche studies, extremely rare, and therefore very valuable for the development of soil science, enriching new knowledge that allows discovering new relationships between abiotic and biotic components of the environment and discovering new diatom species. This research can contribute to the development of environmental sciences, and at the same time be a source of knowledge for agricultural practice. In summary, systems and technologies using microorganisms have many advantages, such as increasing food production, protecting plants and improving soil quality. At the same time, there are also disadvantages, such as uncontrolled growth of microorganisms and the risk associated with GMOs, which should be considered when assessing their use.
4. Conclusions
Microbial biotechnology is key to sustainable agriculture, offering tools to combat pathogens, increasing crop areas and ensuring global food security. Microbial formulations such as Nod accelerate plant germination and increase the number of root nodules, which translates into better colonization of plants by symbiotic bacteria, improved plant nitrogen supply and increased biomass production. Fungal biopreparations support plant growth and protect them from pathogens. With these solutions, farmers can increase the efficiency of legume cultivation, reduce synthetic nitrogen fertilization and protect crops sustainably and naturally from pathogens. Microbial biotechnology plays an important role in sustainable development through the use of microorganisms, plant extracts and minerals in various applications. However, challenges and ethical issues must be considered, such as the potential release of GMOs into the environment, the development of resistance in weed, disease and pest populations, and the need for further research to better understand the possibilities and limitations of this technology. Careful assessment of the risks and benefits of its use and implementation of appropriate measures to ensure safe and effective use will contribute to the sustainable development of the natural environment.
In the context of an interconnected and interdependent world, sustainable solutions to the most pressing challenges are essential. Whether through green chemistry or other sustainable technologies that reduce dependence on hazardous chemicals, the development of biopesticides to protect crops and forests, or the use of Effective Microorganisms (EM) to improve soil and water quality, microbial biotechnology offers numerous opportunities to advance sustainable development. The potential benefits are too great to ignore. Microbial biotechnology can play a transformative role in achieving the Sustainable Development Goals (SDGs).
To maximize the benefits of this technology, continued investment in R&D is necessary to better understand the possibilities and limitations of different approaches and to identify the most effective and sustainable solutions. At the same time, careful management and minimization of risks and ethical issues related to microbial biotechnology are necessary. This may include developing new regulatory frameworks, implementing best management practices, and promoting transparency and dialogue to ensure that all stakeholders have a say in the development and use of this technology. By adopting a responsible and inclusive approach to the development and implementation of microbial biotechnology, we can harness its potential to build a more sustainable, equitable, and prosperous future for all.
In an era of increasing chemicalization of the agricultural environment, scientists and farmers are increasingly turning to natural ways to increase yield and protect plants. Modern agriculture increasingly uses sustainable farming methods, including activities that reduce the impact of agriculture on the environment and enable more efficient and sustainable use of resources (soil, water, energy, biopesticides, biofertilizers). Such activities allow for maintaining the profitability of agricultural production and are socially acceptable.
Precision agriculture and the microbiome: The development of omics technologies (metagenomics, meta transcriptomics) allows for an increasingly detailed understanding of the soil microbiome and the identification of key microorganisms responsible for specific functions (e.g., nitrogen fixation, phosphorus solubilization, phytohormone production). This opens the way to the design of precise biopreparations, adapted to the specific needs of soil and plants.
Microbial engineering: Advances in genetic engineering and synthetic biology allow for the modification of microorganisms to increase their efficiency in agriculture (e.g., strains that produce more metabolites, with increased resistance to stress, or more effective in bioremediation). However, this is associated with additional ethical and regulatory challenges.
Technology integration: Sustainable agriculture of the future is not just about individual solutions, but their integration. Combining microbial biotechnology with regenerative agriculture, agroecology or new irrigation systems is becoming a priority.
Plant Immunity and Microbiome: A growing body of research indicates that the root and rhizosphere microbiome play a key role in inducing plant systemic resistance to pathogens (ISR). This opens up new perspectives for plant bio protection through manipulation of their microbiome. Circular Economy: Microorganisms are irreplaceable in the processes of converting organic waste into valuable resources (e.g., compost, biogas), which is consistent with the idea of a circular economy in agriculture
Author Contributions
Conceptualization, B.S., DS, VV, PB, PP, PN. and BB.; methodology, BS, VV, BP, DS, PP, PN; software: PN, PB, DS; validation, BB, VV. and PN; formal analysis, PB, BB, DS; writing—original draft preparation, BS, DS, VV, PB, PP, PN, BB. and BB; writing—review and editing DS, PP, PB, DS, BB. PN; supervision: BS, VV, PN; funding acquisition, BS. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
no new data created.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AMF | an effective plant-arbuscular mycorrhizal fungi |
| AOA | ammonia-oxidizing archaea |
| AOB | ammonia-oxidizing bacteria |
| COD | chemical oxygen demand |
| DO | dissolved oxygen |
| DS | Drought Stress |
| EM | effective microorganisms |
| EPA | The Environmental Protection Agency |
| EPS | extracellular polymeric substance |
| FF | French fries |
| GV | Granulosis’s |
| HDL | high-density lipoprotein fraction |
| HN-AD | heterotrophic nitrification and aerobic denitrification |
| HNADM | heterotrophic nitrification with oxygen denitrifying microorganisms |
| HRAC | Herbicide Resistance Action Committee |
| LDL | low-density lipoprotein fraction |
| LEMs | effective local microorganisms |
| MBBR | moving bed biofilm reactor |
| MBR | membrane bioreactor |
| MLR | Maximum Residue Levels |
| MoA in Pesticides | Mode of Action |
| NPV | Nuclear polyhedrosis viruses |
| NOB | nitrite-oxidizing bacteria |
| Nod | biofertilizers containing rhizobia Nod |
| PBS | polybutylene succinatepn |
| PC | Potato Chips |
| PGPR | Plant Growth Promoting Rhizobacteria) |
| PP | pomelo peel |
| PSI | Photosystem I |
| PSII | Photosystem II bs |
| QS | secretion of quorum sensing |
| RSM | response surface methodology |
| SBBR | sequencing batch biofilm reactor |
| SBR | sequencing batch reactor |
| SHARON | Single reactor High activity Ammonium Removal Over Nitrite |
| SND | simultaneous heterotrophic nitrification and aerobic denitrification |
| SwF | seaweed extracts |
| tEM | thermoacids of effective microorganisms |
| tEMA | thermoacids of effective microorganisms with shading |
| tEMB | thermo acids of effective microorganisms without shading |
| TIN | total inorganic nitrogen |
| TN | total nitrogen |
| TP | total phosphorus |
References
- Kucharski J, Jastrzębska E. The role of effective microorganisms in shaping the microbiological properties of soil. Ecological Engineer, 2005, 12: 295-296. (in Polish).
- Shah K.K., Tripathi S., Tiwari I., Shrestha J., Modi B., Paudel N., Das BD., Role of soil microbes in sustainable crop production and soil health: A review. Agricultural Science and Technology, 2021, 13(2), 109-118. [CrossRef]
- Sarveswaran S, Vishal Johar*, Vikram Singh and Ragunanthan C. Agroforestry: A Way Forward for Sustainable Development. Eco. Env. & Cons. 2023, 29 (August Suppl. Issue): S300-S309. ISSN 0971–765X. [CrossRef]
- Li, X, Guo, Q, Wang, Y, Xu, J, Wei, Q, Chen, L, Liao, L. Enhancing Nitrogen and Phosphorus Removal by Applying Effective Microorganisms to Constructed Wetlands. Water, 2020, 12, 2443. [Google Scholar] [CrossRef]
- Coskun M, Coskun M, Cayir A. Frequencies of micronuclei (MNi), nucleoplasm bridges (NPBs), and nuclear buds (NBUDs) in farmers exposed to pesticides in Çanakkale, Turkey. Environ Int, 2011, 1:93-96.
- Wong, R. H, Chang S.Y, Ho S.W. Polymorphisms in metabolic GSTP1 and DNA-repair XRCC1 genes with an increased risk of DNA damage in pesticide-exposed fruit growers. Mutat Res 2008,654:168-75.
- Sawicka B., Vambol V., Krochmal-Marczak B., Messaoudi M, Skiba D., Pszczółkowski, P., Barbaś, P., Farhan A.K. Green Technology as a Way of Cleaning the Environment from Petroleum Substances in South-Eastern Poland. Frontier Biosci. (Elite Ed.) 2022; 14(4): 28. [CrossRef]
- Wang, C. Green Technology Innovation, Energy Consumption Structure and Sustainable Improvement of Enterprise Performance. Sustainability 2022, 14, 10168. [Google Scholar] [CrossRef]
- Commission Implementing Regulation (EU), No. 540/2011, OJ L 153 of 11.6.2011: 1–186.
- European Food Safety Authority. EFSA Journal 2012, 10 (10): sf101, https://www.efsa.europa.eu/en/efsajournal/pub/sf101 (accessed 17/12/2021).
- European Food Safety Authority, Conclusion on the peer review of the pesticide risk assessment of the active substance gibberellins. EFSAJ 2012b, 10(1):2502–2551.
- European Food Safety Authority. Conclusion on the peer review of the pesticide risk assessment of the active substance Bacillus thuringiensis ssp. tenebrionis strain NB-176. EFSA Journal 2013a, 11(1): 3024–3059.
- European Food Safety Authority, Conclusion on the peer review of the pesticide risk assessment of the active substance carbon dioxide. EFSA Journal 2013b, 11(5): 3053–3153.
- European Food Safety Authority, Conclusion on the peer review of the pesticide risk assessment of the active substance orange oil. EFSA Journal 2013c, 11(2): 3090–3144.
- European Union Pesticides Database (2019). (Online). Available: http://ec.europa.eu/food/plant/protection/evaluation/database_act_subs_en.html (accessed 02.10.2021).
- Ortiz, A., Sansinenea, E. Chapter 1. Bacillus thuringiensis based biopesticides for integrated crop management, Editor(s): Amitava Rachit, Vijay Singh Meena, P.C. Abhilash, B.K. Sarma, H.B. Singh, Leonardo Fraceto, Manoj Parihar, Anand Kumar Singh, [In:] Advances in Bio-inoculant Science, Biopesticides, Woodhead Publishing, 2022, Pages 1-6, ISBN 9780128233559. [CrossRef]
- Kapka-Skrzypczak L, Cyranka M, Kruszewski M, Turski W.A. Plant protection products and farmers’ health - biomarkers and the possibility of using them to assess exposure to pesticides. Medycyna Ogólna i Nauki o Zdrowiu, 2011, 17(1): 28-32. (in Polish).
- Skiba D., Sawicka B., Pszczółkowski P., Barbaś P., Krochmal-Marczak. Impact of crop management and weed control systems of very early potatoes on weed infestation, biodiversity, and tuber health safety. Life, 2021, 11(826): 33. [CrossRef]
- Bolognesi, C. Genotoxicity of pesticides: a review of human biomonitoring studies. Mutat Res, 2003, 3:251-72.
- Barbaś P., Sawicka B. Comparison of the profitability of various methods of weed infestation in edible potato cultivation. Problems of Agricultural Engineering, 2017, 2 (96): 5–15. ISSN 1231-0093 (in Polish).
- Nowacka A, Gnusowski B, Dąbrowski J. Remains of protection measures in agricultural crops (2006). The remains. Progress in Plant Protection/Postępy w Ochronie Roślin 47(4):79-90. (in Polish).
- Martínez-Valenzuela C, Gómez-Arroyo S, Villalobos-Pietrini R. Genotoxic biomonitoring of agricultural workers exposed to pesticides in the north of Sinaloa State, Mexico. Environ Int, 8:1155-9.
- Bortoli G.M, Azevedo M.B, Silva L.B. Cytogenetic biomonitoring of Brazilian workers exposed to pesticides. Micronucleus analysis in buccal epithelial cells of soybean growers. Mutat Res, 2009, 1-2: 1-4.
- Crestani M, Menezes Ch, Glusczak L, Santos Miron D. Effects of Clomazone Herbicide on biochemical and histological aspects of silver catfish (Rhamdia quelen) and recovery pattern. Chemosphere, 2007, 67: 2305-2311.
- Santos MD, Crestani M, Shettinger M. R, Morsch V.M. Effects of the herbicides clomazone, quinclorac, and metsulfuron methyl on acetylcholinesterase activity in the silver catfish (Rhamadia quelen). Ecotoxicology and Environmental Safety, 2005, 6: 398-403.
- Anonimous. Assessment of the effectiveness of plant protection products. Resistance risk analysis. Organisation Européenne et Méditerranéenne pour la Protection des Plantes, 2002, PP 1/213(2).
- Alam A, Bibi F, Deshwal K, Sahariya A, Bhardwaj C, Emmanuel I. Biofortification of primary crops to eliminate latent hunger: an overview. Natural resources for human health. 2022, 2(1): 91-99. [CrossRef]
- Li X, Pan Y, Q Xi, Zhang S, Zhi Ch, Meng H, Cheng Z. Effects of exogenous germanium and effective microorganisms on germanium accumulation and nutritional qualities of garlic (Allium sativum L.), Scientia Horticulture, 2021, 283: 110114. [CrossRef]
- Gupta K. Profiles of the bio-fertilizer industry on the market. (In:) Biofertilizers: research and impact. Editors: Inamuddin, Mohd Imran Ahamed, Rajender Boddula, Mashallah Rezakazemi, 2021a. [CrossRef]
- Gupta, P.K, Balyan, H.S, Sharma, S, Kumar, R. Biofortification and bioavailability of Zn, Fe and Se in wheat: present status and future prospects. Theoretical and Applied Genetics, 2021b, 134(1): 1-35. [CrossRef]
- El-Ghwas, DE. , Elkhateeb WA., Daba G, M. Fungi: The Next Generation of Biofertilizers. Environmental Science Archives, 2022, 2(1), 34-41. [CrossRef]
- Singh M, Biswas SK, Nagar D, Lal K, Singh J. Effect of biofertilizer growth parameters and potato yield. inside J. Curr. Microbiol. Regret. Science, 2017, 6(5): 1717-1724.
- De Assis RMA, Carneiro JJ, Medeiros APR, de Carvalho AA, da Cunha Honorato A, Carneiro MAC, Bertolucci, SKV, Pinto JEBP. Arbuscular mycorrhizal fungi and organic manure enhance growth and accumulation of citral, total phenols, and flavonoids in Melissa officinalis L. Ind. Crops Prod. 2020, 158: 112981. [CrossRef]
- Mathur S, Tomar RS, Jajoo A. Arbuscular mycorrhizal fungi (AMF) protect photosynthetic apparatus of wheat under drought stress. Photosynth. Res. 2019, 139:227–238. [CrossRef]
- Mathur S, Jajoo A. Arbuscular mycorrhizal fungi protect maize plants from high temperature stress by regulating photosystem II heterogeneity. Ind. Crops Prod. 2020, 143: 111934. [CrossRef]
- Igiehon NO, Babalola OO, Cheseto X, Torto, B. Effects of rhizobia and Arbuscular mycorrhizal fungi on yield, size distribution and fatty acid of soybean seeds grown under drought stress. Microbiol. Res. 2021, 242: 126640. [CrossRef]
- Paravar A, Farahani SM, Rezazadeh A. Lallemantia response to drought stress and the use of Arbuscular mycorrhizal fungi. Crops and Industrial Products 2021,172: 114002.
- Plouznikoff K, Asins MJ, de Boulois HD, Carbonell EA, Declerck S. Genetic analysis of tomato root colonization by arbuscular mycorrhizal fungi. Ann. Bot. 2019, 124:933–946. [CrossRef]
- Liu R, Abdelraouf E, Bicego B, Joshi V, Garcia, AG. Deficit irrigation: a viable option for sustainable confection sunflower (Helianthus annuus L.) production in the semi-arid US. Irrig. Sci. 36: 319–328. [CrossRef]
- Liu J, Liu X, Zhang Q, Li S, Sun Y, Lu W, Ma C. Response of alfalfa growth to arbuscular mycorrhizal fungi and phosphate-solubilizing bacteria under different phosphorus application levels. AMB Express 2020, 10: 1–13. [CrossRef]
- Ghanbarzadeh Z, Mohsenzadeh S, Rowshan V, Zarei M. Mitigation of water deficit stress in Dracocephalum moldavica L. by symbiotic association with soil microorganisms. Sci. Hortic. 2020, 272: 109549. [CrossRef]
- Ramos-González MI, Matilla MA, Quesada JM, Ramos JL, Espinosa-Urgel M. Using genomics to discover bacterial lifestyle determinants in the rhizosphere (in:) Molecular Microbiological Ecology of the Rhizosphere, Editors: Frans J. de Bruijn. Issue 1, March 18, 2013, ISBN: 9781118296172. [CrossRef]
- Mgomi FC, Zhang B-X, Lu Ch-L, Yang Z-Q, Yuan L. Novel biofilm-inspired encapsulation technology enhances the viability of probiotics during processing, storage, and delivery, Trends in Food Science & Technology, 2025, 160, 105032, ISSN 0924-2244. [CrossRef]
- Ferrando L, Rariz G, Martínez-Pereyra A, Fernández-Scavino A, Endophytic diazotrophic communities from rice roots are diverse and weakly associated with soil diazotrophic community composition and soil properties, Journal of Applied Microbiology, 2024, 135(7),157. [CrossRef]
- Kowalska, J. , Niewiadomska J. Głuchowska K. Kaczmarek D., Impact of fertilizers and soil properties in the case of Solanum tuberosum L. during conversion to organic farming. Applied ecology and Environmental Research, 2017, 15(4),369-839.
- Council Directive 91/414/EEC of 15 July 1991 concerning the placing of plant protection products on the market. OJ L 230, 19.8.1991: p. 1–32.
- Regulation (EC) No 1107/2009 of the European Parliament and of the Council on the placing of plant protection products on the market and repealing Council Directives 79/117/EEC and 91/414/EEC. https://www.fao.org/faolex/results/details/en/c/LEX-FAOC103352 (accessed 14.12.2022.
- Villaverde JJ, Sevilla-Morán B, Sandín-España P, López-Goti C, Alonso-Prados JL. Biopesticides in the framework of the European. Pesticide Regulation (EC) No. 1107/2009. Pest Manag Sci 70: 2-5.
- Lu Ch, Zhang Z, Guo P, Wang R, Liu T, Luo J, Hao B, Wang Y, Guo W. Synergistic mechanisms of bioorganic fertilizer and AMF driving rhizosphere bacterial community to improve phytoremediation efficiency of multiple HMs-contaminated saline soil, Science of The Total Environment, 2023, 883: 163708, ISSN 0048-9697. [CrossRef]
- Yang, Y. The Application of Genomics in Agriculture. Agricultural Science and Food Processing, 2025, 2(1), 26–46. [CrossRef]
- Cheng, Y.; Xue, F.; Yu, S.; Du, S.; Yang, Y. Subcritical Water Extraction of Natural Products. Molecules 2021, 26, 4004. [Google Scholar] [CrossRef] [PubMed]
- Wang Z-Z, Zeng D-W, Zhu Y-F, Zhou M-H, Kondo A, Hasunuma T, Zhao X-Q, Fermentation design and process optimization strategy based on machine learning, BioDesign Research, 2025, 7(1): 100002, ISSN 2693-1257. [CrossRef]
- Sawicka B, Egbuna Ch, Nayak AK, Kala S. Chapter 2. Plant diseases, pathogens, and diagnosis. PART I. Green approach to pest and disease control (in:) Natural Remedies for Pest, Disease and Weed Control. Edited by Chukwuebuka Egbuna, Barbara Sawicka, ELSEVIER, Academic Press, London, 1-16, 2020. ISBN: 978-0-12-819304-4. [CrossRef]
- Crandall, L.; Zaman, R.; Duthie-Holt, M.; Jarvis, W.; Erbilgin, N. Navigating the Semiochemical Landscape: Attraction of Subcortical Beetle Communities to Bark Beetle Pheromones, Fungal and Host Tree Volatiles. Insects 2025, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- Han X, Li S, Zeng Q, Sun P, Wu D, Wu J, Yu X, Lai Z, Milne RJ, Kang Z, Xie K, Li G. Genetic engineering, including genome editing, for enhancing broad-spectrum disease resistance in crops, Plant Communications, 2025, 6(2):101195, ISSN 2590-3462. [CrossRef]
- Vaitkeviciene, N. The Effect of Biodynamic Preparations on the Accumulation of Biologically Active Compounds in the Tubers of Different Genotypes of Ware Potatoes. Ph.D. Thesis, Agricultural Sciences, Agronomy (01A), ASU, Akademija, Kaunas, Lithuania, 2016, pp. 212.
- Vaitkeviciene, N, Jariene, E, Danilcenko, H, Sawicka, B. Effect of biodynamic preparations on the content of some mineral elements and starch in tubers of three colored potato cultivars. J. Elem. 2016, 21, 927–935. [Google Scholar]
- Keidan M. Optimization of winter oilseed rape technological parameters in the organic farming system. PhD thesis, Aleksandras Stulginskis University, Kaunas, pp. 221.
- Adetunji, CO., Oloke, J.K., Phazang P., Sarin N.B. Influence of eco-friendly phytotoxic metabolites from Lasiodiplodia pseudotheobromae C1136 on physiological, biochemical, and ultrastructural changes on tested weeds. Environmental Science and Pollution Research. Environ Sci Pollut Res Int 2020, 27(9): 9919-9934.
- Levickienė D, Jarienė, E, Gajewski, M, Danilčenko, H, Vaitkevičienė N, Przybył JL, Sitarek M. Influence of harvest time on biologically active compounds and antioxidant activity in mulberry leaves in Lithuania. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2017, 45(2): 431-436.
- Levickienė D. The Influence of the Biodynamic Preparations on the Soil Properties and Accumulation of Bioactive Compounds in the Leaves of White Mulberry (Morus alba L.). PhD thesis, Aleksandras Stulginskis University, Kaunas 2018: pp. 212.
- Werle R, Lowell D, Sandeln LD, Buhler DD, Hartzler RG, Lundqvists LJ. Predicting the emergence of 23-year-old annual weed species. Weed Science 2014, 62 (2): 267-279. [CrossRef]
- Stanek-Tarkowska, J.; Szostek, M.; Rybak, M. Effect of Different Doses of Ash from Biomass Combustion on the Development of Diatom Assemblages on Podzolic Soil under Oilseed Rape Cultivation. Agronomy 2021, 11, 2422. [Google Scholar] [CrossRef]
- Ngouajio M, McGiffen ME, Hembree, KJ. Tolerance of tomato cultivars to velvetleaf interference. Weed Sci 2009, 49(379): 91-98, 10.1614/0043-1745(2001)049(0091: TOTCTV)2.0.CO, 2 380.
- Goldwasser Y, Lanini W, Wrobel R. Tolerance of tomato varieties to lespedeza dodder. Weed Sci 2009, 49:520-523. [CrossRef]
- Andrew IKS, Storkey J, Sparke D.L. A review of the potential for competitive cereal cultivars as a tool in integrated weed management. Weed Res 2015, 55, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. , Pan, Z., Bajsa-Hirschel, J., Douglas, B.C. The potential future roles of natural compounds and microbial bioherbicides in weed management in crops. Adv Weed Sci, 40(1), e020210054, Jan. 2022.
- Macías FA, Mejpias FJR, Molinillo JMG. Recent advances in allelopathy for weed control: from knowledge to applications. Pest Manag Sci 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Farooq M, Bajwa AA, Cheema SA, Cheema ZA. Application of allelopathy in crop production Int. J. Agric. Biol., 2013, 15(389): 1367–1378. [CrossRef]
- Portela VO, Moro A, Santana NA, Baldoni DB, de Castro IA, Antoniolli ZI, Dalcol II, Seminoti Jacques RJ. First report on the production of phytotoxic metabolites by Mycoleptodiscus indicus under optimized conditions of submerged fermentation. Environ Technol. 2022 43(10):1458-1470. [CrossRef]
- Boligłowa E. Potato protection against diseases and pests using Effective Microorganisms (EM) with herbs. (In:) Selected ecological issues in modern agriculture. Ed. Z. Zbytek, Publisher: PIMR, Poznań: 165–170, 2005. (in Polish).
- Janas R. Possibilities of using effective microorganisms in ecological systems of crop production. Problems of Agriculure Engineering 2009, 3: 111-119. (in Polish).
- Ji B, Hu H, Zhao Y, Mu X, Liu K, Li C. Effects of deep tillage and straw returning on soil microorganism and enzyme activities. Sci. World J. 2014, 1–12. [Google Scholar] [CrossRef]
- Kołodziejczyk M, Szmigiel A, Ropek, D. Effectiveness of potato protection production using selected insecticides to combat potato beetles (Leptinotarsa decemlineata Say). Acta Scientiarum Polonorum s. Agricultura 2009, 8(4): 5-14.
- Kumar A, Singh VK, Singh P, Mishr K. Purification of environmental pollutants via microbes. 1st edition, Editors: Ajay Kumar Vipin, Kumar Singh, Pardeep Singh, Virendra Kumar Mishra, Woodhead Publishing, eBook ISBN: 9780128232071, Paperback ISBN: 978012821199, 2020: pp. 474.
- Daniel C, Wyssa E. Field applications of Beauveria bassiana to control the Rhizolites cerasi fruit fly. Journal of Applied Entomology 2010, 134 (9-10): 675-681. [CrossRef]
- Macías FA, Rodríguez MJ, Molinillo MG. Recent advances in weed control allopathy. From knowledge to application. Pest Control Science 75 (9). [CrossRef]
- Rudeen ML, Jaronski ST, Petzold-Maxwell JL, Gassmann AJ. Entomopathogenic fungi in cornfields and their potential to fight the larvae of western corn rootworm Diabrotica virgifera. J Invertebr Pathol. 2013 Nov,114(3):329-32. [CrossRef]
- Reddy GVP, Tangtrakulwanich K, Wua S, Miller JH, Ophus VL, Prewett J, Jaronski ST. Evaluation of the effectiveness of entomopaths in the control of wireworms (Coleoptera: Elateridae) on spring wheat. J Invertebr Pathol 2014, 120: 43-49. [CrossRef]
- Trawczyński C., 2012. The influence of irrigation and Effective Microorganisms on quantity and chemical composition of the yields of plants cultivation in organic crop rotation on light soil. J. Res. Appl. Agric. Engineering, 57(4): 148–154.
- Laznik, Ž, Tóth, T, Lakatos, T, Vidrih, M, Trdan, S. Inspection of Colorado potato beetle (L. decemlineata Say) on potato under field conditions: comparison of the effectiveness of the two strains Steinernema feltiae (Filipjev) and spraying with thiamethoxam. J Plant Dis Prot 2010, 117(3): 129-135.
- Lepiarczyk, A, Kulig, B, Stępnik, K. The effect of simplified soil cultivation and forecrops on LAI development of selected winter wheat varieties for crop rotation. Fragmenta Agronomica 2005, 2(86): 98-105.
- Xu, HL. Effects of a microbial inoculant and organic fertilizer on the growth, photosynthesis, and yield of sweet corn. Journal of Crop Production, 2000, 3: 183–214.
- Emitazi G, Nader A, Etemadifar Z. Effect of nitrogen fixing bacteria on growth of potato tubers. Adv. Food Sci. 2004, 26: 56–58.
- Kalitkiewicz A, Kępińska E. The use of Rhizobacteria to stimulate plant growth. Biotechnology, 2008, 2: 102–114.
- Górski R., Kleiber T., 2010. Effect of Effective Microorganisms (EM) on nutrient contents in substrate and development and yielding of rose (Rosa x hybrida) and gerbera (Gerbera jamesonii). Ecol. Chem. Eng., S, 17 (4): 505-513.
- Kaczmarek Z, Owczarzak W, Mrugalska L, Grzelak M. Effect of effective microorganisms on selected physical and water properties of arable-humus levels of mineral soils. J. Res. Appl. Agric. Eng. 2007, 52: 73–77. (In Polish).
- Kaczmarek Z, Jakubas M, Grzelak M, Mrugalska L. Impact of the addition of various doses of Effective Microorganisms to arable-humus horizons of mineral soils on their physical and water properties. J. Res. Appl. Agric. Eng. 2008a, 53: 118–121.
- Kaczmarek Z, Wolna-Murawska A, Jakubas M. Change in the number of selected groups of soil microorganisms and enzymatic activity in soil inoculated with effective microorganisms (EM). J. Res. Appl. Agric. Eng. 2008b, 53: 122–128.
- Szembowski, B. Experiences of a Farm in Trankwice with the EM-FarmingTM Biotechnology. Natural Probiotic Microorganisms, Publisher: Associate Ecosystem, Lichen, Poland, 2009: 56–58. (In Polish).
- Kosicka D, Wolna-Murawka A, Trzeciak M. Influence of microbiological preparations on soil and plant growth and development. Kosmos 2015, 64(2): 327-335. (in Polish).
- Kołodziejczyk M. Effectiveness of nitrogen fertilization and application of microbial preparations in potato cultivation. Turkish Journal of Agriculture and Forestry, 2014a, 38: 299–310.
- Kołodziejczyk, M. Effect of nitrogen fertilization and microbial populations on potato yielding. Plant Soil Environ. 2014, 60(8): 379–386.
- Pszczółkowski P, Sawicka B. The impact of the application of biopreparations and fungicides on the yield and selected parameters of the seed value of seed potatoes. Acta Agroph. 2018a, 25(2): 239–255.
- Pszczółkowski P, Sawicka B. The impact of the use of fungicides, microbiological preparations, and herbal extracts on the shaping of the potato yield. Fragm. Agron, 2018b, 35 (1): 81-93.
- Pszczółkowski P, Krochmal-Marczak B, Sawicka B, Pszczółkowski M. Effect of the use of effective microorganisms on the color of raw potato tuber flesh for food processing. Appl Sci 2021, 11 (19): 8959. [CrossRef]
- Sawicka B, Pszczółkowski P, Kiełtyka-Dadasiewicz A, Ćwintal M, Krochmal-Marczak B. Effect of effective microorganisms on the quality of potatoes in food processing. Appl. Sci. 2021, 11: 1415. [CrossRef]
- Koskey G, Mburu SW, Awino R, Njeru EM, Maingi JM. Potential Use of Beneficial Microorganisms for Soil Amelioration, Phytopathogen Biocontrol, and Sustainable Crop Production in Smallholder Agroecosystems. Frontiers in Sustainable Food Systems, 2021, 5: 130. [CrossRef]
- Galarreta JIR, Ezpelata B, Pascualena J, Ritter E. Combining ability in early generations of potato breeding. Plant Breeding, 2006, 125: 183–186.
- Kolasa-Wiącek, A. Will Effective Microorganisms Revolutionize the World? Post. Technol. Convert Food. 2010, 1: 66–69. (In Polish).
- Kowalska J, Sosnowska D, Remlein-Starosta D, Drożdżyński D, Wojciechowska R, Łopatka W. Effective microorganisms in organic farming. Publisher: IOR-PIB, Poznań, 2021. https://www.ior.poznan.pl/plik,3661,sprawozdanie-mikroorganizmy-w-rol-eko-2011-pdf.pdf.
- Barbaś, P., Skiba, D, Pszczółkowski, P, Sawicka B. Natural Resistance of Plants. Subbject: Biochemistry & Molecular Biology. 2022. E Scholary Community Encyclopedia. https://encyclopedia.pub/entry/34721.
- Martyniuk S. Effective and ineffective microbiological preparations used in the protection and cultivation of plants as well as reliable and unreliable methods of their evaluation. Post. Microbiol., 2011, 50(4): 321-328.
- Okorski M, Majchrzak B. Fungi colonizing pea seeds after applying the EM 1 microbiological preparation. Progress in Plant Protection/Postępy w Ochronie Roślin, 2008, 48(4): 1314–1318.
- Higa T. Effective microorganisms, concept, and the latest technological achievements. Materials from the Effective Microorganisms Conference for Sustainable Agriculture and Environment. 4th Kyusei International Wildlife Agriculture Conference, Bellingham-Washington, 1998, USA: 247-248.
- Dach, J., Wolna-Maruwka B., Zbytek Z. Influence of effective microorganisms’ addition (EM) on composting process and gaseous emission intensity Journal of Research and Applications in Agricultural Engineering, 2009, 54(3), 49-55.
- Higa, T. Effective microorganisms – technology of the 21st century. Conference: “Effective Microorganisms in the World”. London, UK, 2005: 20-24.
- Zarzecka, K, Gugała, M. Effect of UGmax soil fertilizer on the potato yield and its structure. Bulletin IHAR, 2013, 267:107–112. (In Polish).
- Baranowska A, Zarzecka K, Gugała M, Mystkowska I. The effect of fertilizer on UGmax soil on the presence of Streptomyces scabies on edible potato tubers. J. Ecol. Eng. 2018, 3:68-73. [CrossRef]
- Van Vliet PCJ, Bloem J, de Goede RGM. Microbial diversity, nitrogen loss and grass production after the addition of Effective Microorganisms® (EM) to slurry manure. Applied Soil Ecology, 2006, 32: 188–198.
- Martyniuk S. Production of microbiological preparations on the example of symbiotic bacteria in legumes. J. Res. Appl. Agric. Eng. 2010, 55: 20–23. (In Polish).
- Martyniuk S, Księżak J. Evaluation of pseudo-microbial biopreparations used in plant production. Polish Agronomic Journal, 2011, 6: 27-33.
- Paśmionka I, Kotarba K. Possibilities of using effective microorganisms in environmental protection. Cosmos, Problems of Biological Sciences, 2015, 64 (1): 173-184.
- Ding, L.-N.; Li, Y.-T.; Wu, Y.-Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.-L. Plant Disease Resistance-Related Signaling Pathways: Recent Progress and Future Prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef] [PubMed]
- Okumoto, S, Shintani, M, Higa, T. The use of effective microorganisms and biochar inhibits the transfer of radioactive caesium from soil to plants during continuous Komatsu cultivation. [In:] Proceedings of the International Scientific Conference “Radiobiology: Present”, Institute of Radiobiology of the NAS of Belarus, Gomel, Belarus, 26–27 September 2019, 22.
- Nigussie, A., Dume, B., Ahmed, M., Mamuye, M., Ambaw, G., Berhiun, G., Biresaw, A., Aticho, A. Effect of microbial inoculation on nutrient turnover and lignocellulose degradation during composting: A meta-analysis. Waste Management, 2021, 125: 220-234, ISSN 0956-053X. [CrossRef]
- Allahverdiyev SR, Kırdar E, Gunduz G, Kadimaliyev D, Revin V, Filonenko V, Rasulova DA, Abbasova ZI, Gani-Zade SI, Zeynalova EM. Effective Microorganisms (EM). Technology in Plants. Technology, 2011, 14:103–106.
- Pszczółkowski P, Sawicka B, Danilcenko H, Jariene E. The role of microbiological preparations in improving the quality of potato tubers. [In:] Proceedings of the International Scientific Conference: ‘New trends in food safety and quality’, Aleksandras Stulginskis University, Akademija, Lithuania, 5–7 October 2017: 22–23.
- Sawicka B, Egbuna C. Chapter 1. Pest of agricultural crops and control measures. PART I. Green approach to pest and disease control (in:) Natural Remedies for Pest, Disease and Weed Control. Edited by Chukwuebuka Egbuna, Barbara Sawicka, Elsevier, Academic Press, 17-28, 2020, ISBN: 978-0-12-819304-4. [CrossRef]
- Sawicka, B. Rate of spread of fungal diseases on potato plants as affected by application of a biostimulator and foliar fertilizer. Biostimulators in modern agriculture. Solanaceous crops. Ed. Z. Dąbrowski. Published by the Editorial House: Wieś Jutra, Limited. Warsaw. Plantpress, 2008, ISBN 83-89503-55-7: 68-76.
- Boligłowa E, Gleń K. Assessment of effective microorganism activity (EM) in winter wheat protection against fungal diseases. Ecol. Chem. Eng. A 2018, 15:23–27.
- Gacka S. Economic aspects of implementing EM-FarmingTM biotechnology. Natural probiotic microorganisms. Publisher: Ecosystems Association, Licheń, 2009, 28–33. (In Polish).
- Tommonaro G, Abbamondi GR, Mikołaja B, Poli A, D’Angelo C, Iodice C, De Prisco R. Productivity and Nutritional Trait Improvements with Various Tomatoes Grown Effective Microorganisms Technology. Agriculture 2021, 11, 2, (112). [Google Scholar] [CrossRef]
- Quiroz M, Céspedes C. Bokashi as a fix and nitrogen source for sustainable farming systems: an overview. Journal of Soil Science and Plant Nutrition 2019, 237–248. [CrossRef]
- Marczakiewicz J. Another year with EM biotechnology at RZD SGGW Chylice. Natural probiotic microorganisms. Ecosystem Association Publishing House, Licheń, 2009, 65. (in Polish).
- Mathews, S. , Gowrilekshmi R. Solid Waste Management with Effective Microbial (EM) Technology. J. Curr. Microbiol. App. Sci, 2016, 5(7): 804-815. [CrossRef]
- Yan Y., Lu H., Zhang J., Shuguang C., Yangqing C., Lei WY, Zhang R., Song L. Simultaneous heterotrophic nitrification and aerobic denitrification (SND) for nitrogen removal: review and future prospects, Environmental Advances, 2022, 9, 100254. [CrossRef]
- Al-Taweil HI, Bin Osman M, Hamid AA, Yusoff WM. Development of microbial inoculants and the impact of soil application on rice seedlings growth. Am. J. Agric. Biol. Sci. 2009, 4: 79–82.
- Szewczuk Cz, Sugier D, Baran S, Bielińska E.J, Gruszczyk M. The impact of fertilizing agents and different doses of fertilizers on selected soil chemical properties as well as the yield and quality traits of potato tubers). Annales Universitatis Mariae Curie-Skłodowska, Sectio E, Agricultura, 2016, LXXI (2): 65–79.
- Development of ecological hop production technology. [in:] 1st Lublin Scientific and Technical Conference – Microorganisms in Environmental Revitalization - Science and Practice.
- Henry AB, Maung CEH, Kim KY. Metagenomic analysis reveals enhanced biodiversity and composting efficiency of lignocellulosic waste by thermoacidophile effective microorganism (tEM), Journal of Environmental Management, 2020, 276: 111252, ISSN 0301-4797. [CrossRef]
- Ney L, Franklin D, Mahmud K, Cabrera M, Hancock D, Habteselassie M, Newcomer Q, Dahal S. Impact of inoculation with local effective microorganisms on soil nitrogen cycling and legume productivity using composted broiler litter, Applied Soil Ecology, 2020, 154: 103567. [CrossRef]
- Faturrahman L, Meryandini A, Junior M.Z, Rusmana I. The Role of Agarolytic Bacteria in Enhancing Physiological Function for Digestive System of Abalone (Haliotis asinine). J. Appl. Environ. Biol. Sci. 2015, 5(5): 49–56.
- Gacka S, Kolbusz S. Biotechnology EM-FarmingTM – comprehensive, natural, solution in animal production ensuring animal welfare. Natural probiotic microorganisms. Publisher: Ekosystem Association, Licheń, 2009: 102–103. (In Polish).
- Tsatsakis, A.M., Nawaz, M.A., Kouretas D., Balias, G., Savolainen, K., Tutelyan, V.A., Golokhvast, K.S., Lee, J.D., Chung, J.G. 2017. Envirtmental impacts of Genetically Modified Plants: A Review, Environmental Research, 156, 2017, 818-833, ISSN 0013-9351. [CrossRef]
- Pniewska I. Wpływ Efektywnych Mikroorganizmów (EM) i dokarmiania pozakorzeniowego nawozami typu Alkalin na plonowanie i cechy jakościowe fasoli szparagowej. Rozprawa Doktorska, UPH, Siedlce, 2015, 94 ss. UPH241d48bcde014585936d1137b4d854ac [in {Polish).
- Fatunbi O, Ncube L. Activities of Effective Microorganism (EM) on the Nutrient Dynamics of Different Organic Materials Applied to Soil. Am. Eurasian J. Agron. 2009, 2(1): 26–35.
- Safwat M., Safwat, Rozaik, E. Growth Inhibition of Various Pathogenic Microorganisms Using Effective Microorganisms (EM). International Journal of Research and Engineering, 2017, 4 (12), 283-286. [CrossRef]
- Gopalasamy, R. The role of microorganisms in sustainable development Envtl. International Journal of Scientific Research 2019, 6 (7): 413.
- Song T, Zhang X, Li J, Wu X, Feng H, Dong W. A review of research progress of heterotrophic nitrification and aerobic denitrification microorganisms (HNADMs), Science of The Total Environment, 2021, 80: 149319. [CrossRef]
- Correia, T.S.; Lara, T.S.; Santos, J.A.d.; Sousa, L.D.S.; Santana, M.D.F. Arbuscular Mycorrhizal Fungi Promote Physiological and Biochemical Advantages in Handroanthus serratifolius Seedlings. Plants 2022, 11, 2731. [Google Scholar] [CrossRef] [PubMed]
- Martin FM, van der Heijden MAG. The mycorrhizal symbiosis: research frontiers in genomics, ecology, and agricultural application. New Phytologist, 2024, 242(4): 1486-1506. [CrossRef]
- Holmes, D.E. Dang, Y., Smith J.A. Nitrogen cycling during wastewater treatment. Adv. Appl. Microbiol., 2019,106: 113-192.
- Xi, H., Zhou, X., Arslan, M., Luo, Z., Wei, J., Wu, Z., El-Din M.G. Heterotrophic nitrification and aerobic denitrification process: Promising but a long way to go in the wastewater treatment. Sci. Total Environ., 2022, 805, Article 150212.
- Lei, Y., Wang, Y., Liu, H., Xi, C., Song L. A novel heterotrophic nitrifying and aerobic denitrifying bacterium, Jobelihle taiwanensis DN-7, can remove high-strength ammonium. Appl. Microbiol. Biotechnol., 2016, 100 (9), 4219-4229.
- Su, JF., Shi, JX., Huang, T.L., Ma F. Kinetic analysis of simultaneous denitrification and biomineralization of novel Acinetobacter sp. CN86. Mar. Pollut. Bull., 109 (1) (2016), pp. 87-94.
- , Shi, J.X., Ma F. Aerobic denitrification and biomineralization by a novel heterotrophic bacterium, Acinetobacter sp. H36. Mar. Pollut. Bull., 2017, 116 (1–2), 209-215.
- Jetten, M.S. The microbial nitrogen cycle. Environ. Microbiol., 2008, 10(11), 2903-2909.
- Liu, S., Chen, Q., Ma, T., Wang, M., Ni J. Genomic insights into metabolic potentials of two simultaneous aerobic denitrification and phosphorus removal bacteria, Achromobacter sp. GAD3 and Agrobacterium sp. LAD9 FEMS Microbiol. Ecol., 2018, 94 (4), 223.
- Chen, J. , Zhao, B., An, Q., Wang, X., Zhang Y.X. Kinetic characteristics and modelling of growth and substrate removal by Alcaligenes faecalis strain NR. Bioprocess. Biosyst. Eng., 2016, 39 (4), 593-601.
- Xia, L., Li, X., Fan, W., Wang J. Heterotrophic nitrification and aerobic denitrification by a novel Acinetobacter sp. ND7 isolated from municipal activated sludg Bioresour. Technol., 301 (2020), Article 122749.
- Msambwa MM, Daniel K, Lianyu C, Integration of information and communication technology in secondary education for better learning: A systematic literature review. Social Sciences & Humanities Open, 2024, 10: 101203, ISSN 2590-2911. [CrossRef]
- Padhi, S.K., Maiti NK. Molecular insight into the dynamic central metabolic pathways of Achromobacter xylosoxidans CF-S36 during heterotrophic nitrogen removal processes J. Biosci. Bioeng., 2017, 123 (1), 46-55.
- Medhi, K. Singhal, A., Chauhan, D.K. Thakur I.S. Investigating the nitrification and denitrification kinetics under aerobic and anaerobic conditions by Paracoccus denitrificans ISTOD1, Bioresour. Technol., 242 (2017), 334-343.
- Huang, F., Pan, L., Lv, N., Tang, X. Characterization of novel Bacillus strain N31 from mariculture water capable of halophilic heterotrophic nitrification-aerobic denitrification J. Biosci. Bioeng., 124 (5) (2017), pp. 564-571.
- He, T., Xie, D., Li, Z., Ni, J., Sun Q. Ammonium stimulates nitrate reduction during simultaneous nitrification and denitrification process by Arthrobacter arilaitensis Y-10. Bioresour. Technol., 2017, 239, 66-73.
- Zhao, B., He, Y.L., Hughes, J., Zhang X.F. Heterotrophic nitrogen removal by a newly isolated Acinetobacter calcoaceticus HNR. Bioresources Technol., 2010, 101 (14), 5194-5200.
- Yang, L., Ren, Y.X., Liang, X., Zhao, SQ., Wang, JP., Xia, Z.H. Nitrogen removal characteristics of a heterotrophic nitrifier Acinetobacter junii YB and its potential application for the treatment of high-strength nitrogenous wastewater. Bioresour. Technol., 2015, 193, 227-233.
- Wang, H., Zhang, W., Ye, Y., He, Q., Zhang S. Isolation and characterization of Pseudoxanthomonas sp. Strain YP1 capable of denitrifying phosphorus removal (DPR) Geomicrobiology J. 2018, 35 (6), 537-543.
- 1Zhang D, Huang, X., Li, W., Qin, W., Wang P. Characteristics of heterotrophic nitrifying bacterium strain SFA13 isolated from the Songhua River. Ann Microbiol, 2015, 66 (1), 271-278.
- Xu, X.; Yang, K.; Dou, Y. High-end equipment development task decomposition and scheme selection method. J. Syst. Eng. Electron. 2021, 32, 118–135. [Google Scholar]
- Luyckx, M.; Reins, L. The Future of Farming: The (Non)-Sense of Big Data Predictive Tools for Sustainable EU Agriculture. Sustainability 2022, 14, 12968. [Google Scholar] [CrossRef]
- Xu, X.; Yang, K.; Dou, Y. High-end equipment development task decomposition and scheme selection method. J. Syst. Eng. Electron. 2021, 32, 118–135. [Google Scholar]
- Bremmer J, Riemens M, Reinders M. The future of crop protection in Europe. Panel for the Future of Science and Technology. European Parliamentary Research Service Scientific Foresight Unit (STOA) PE 656.330 – February 2021 EN. https://www.europarl.europa.eu/RegData/etudes/STUD/2021/656330/EPRS_STU(2021)656330_EN.pdf.
- Mann, R. S, Kaufman, P.E. Natural product pesticides: their development, delivery and use against insect vectors. Mini-Rev. Org Chem 2012, 9:185–202.
Figure 1.
The classification of biofertilizers is based on their functions and nature. Source: own.

Figure 2.
Mechanism of microbial bioherbicide. Source: own.
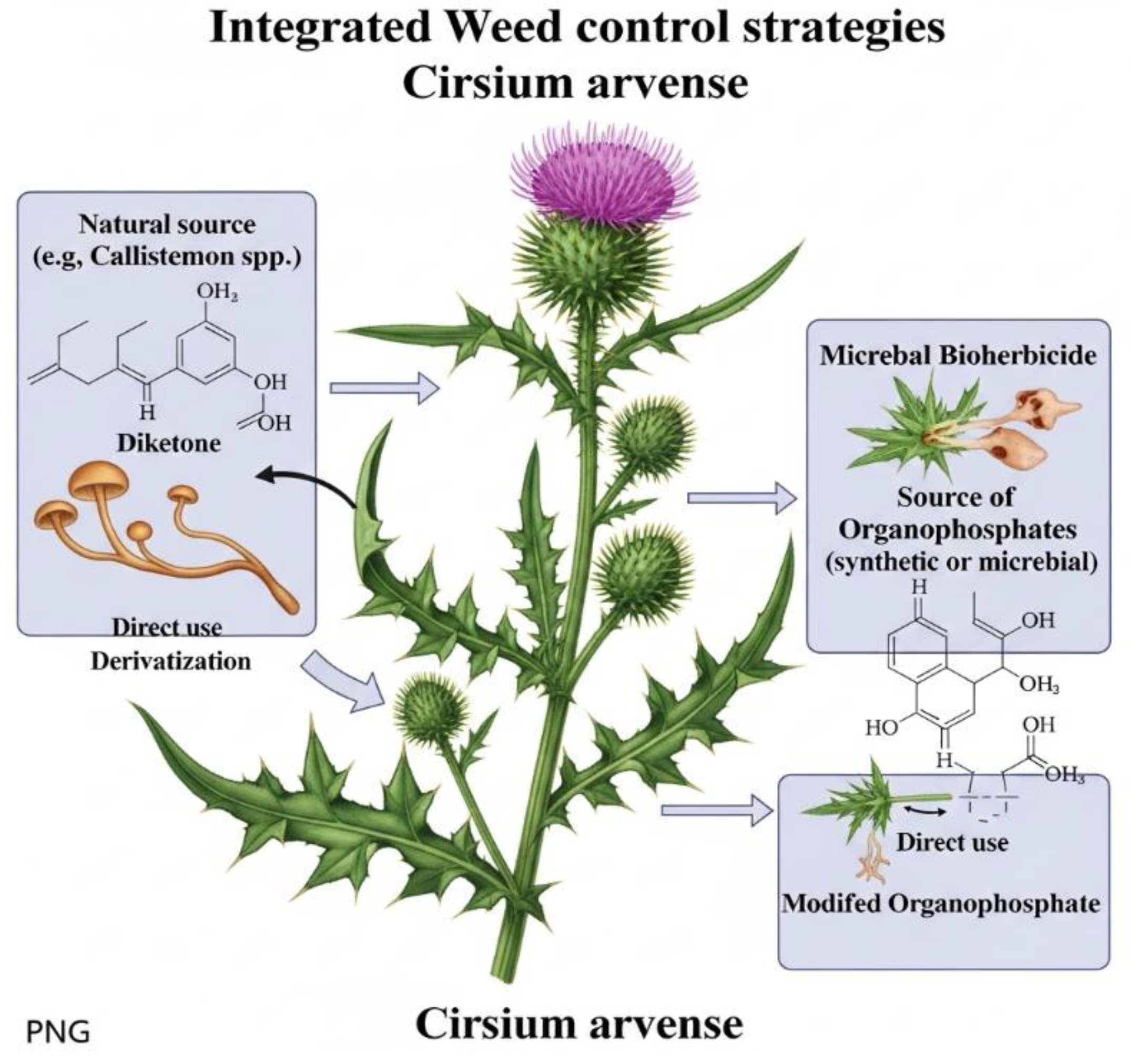
Figure 3.
The key role of microorganisms in sustainable agriculture. Source: own.

Figure 4.
Scheme of action of a biopesticide containing live spores of the fungus. Source: own.
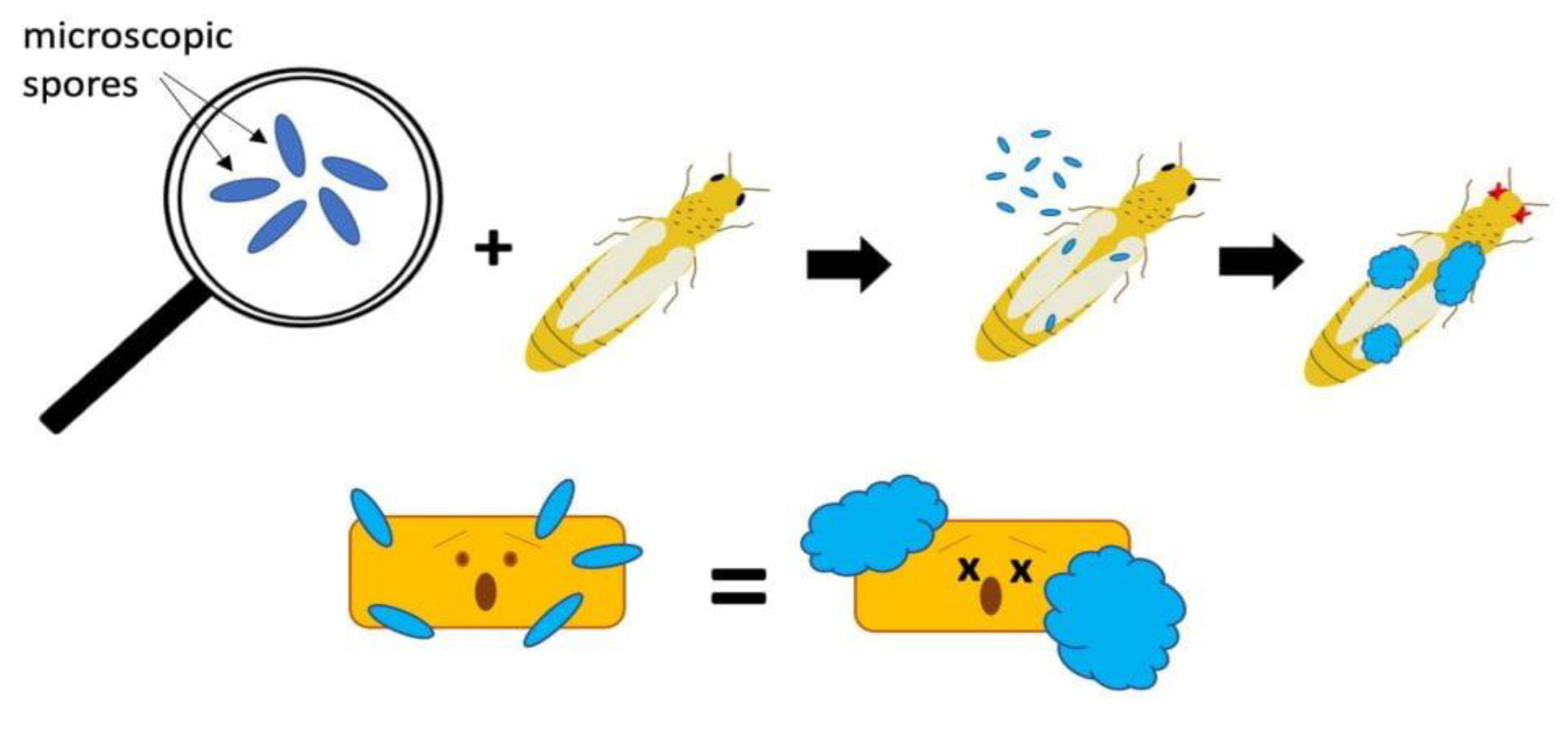
Figure 5.
Effective Microorganisms (EM). Source: own.
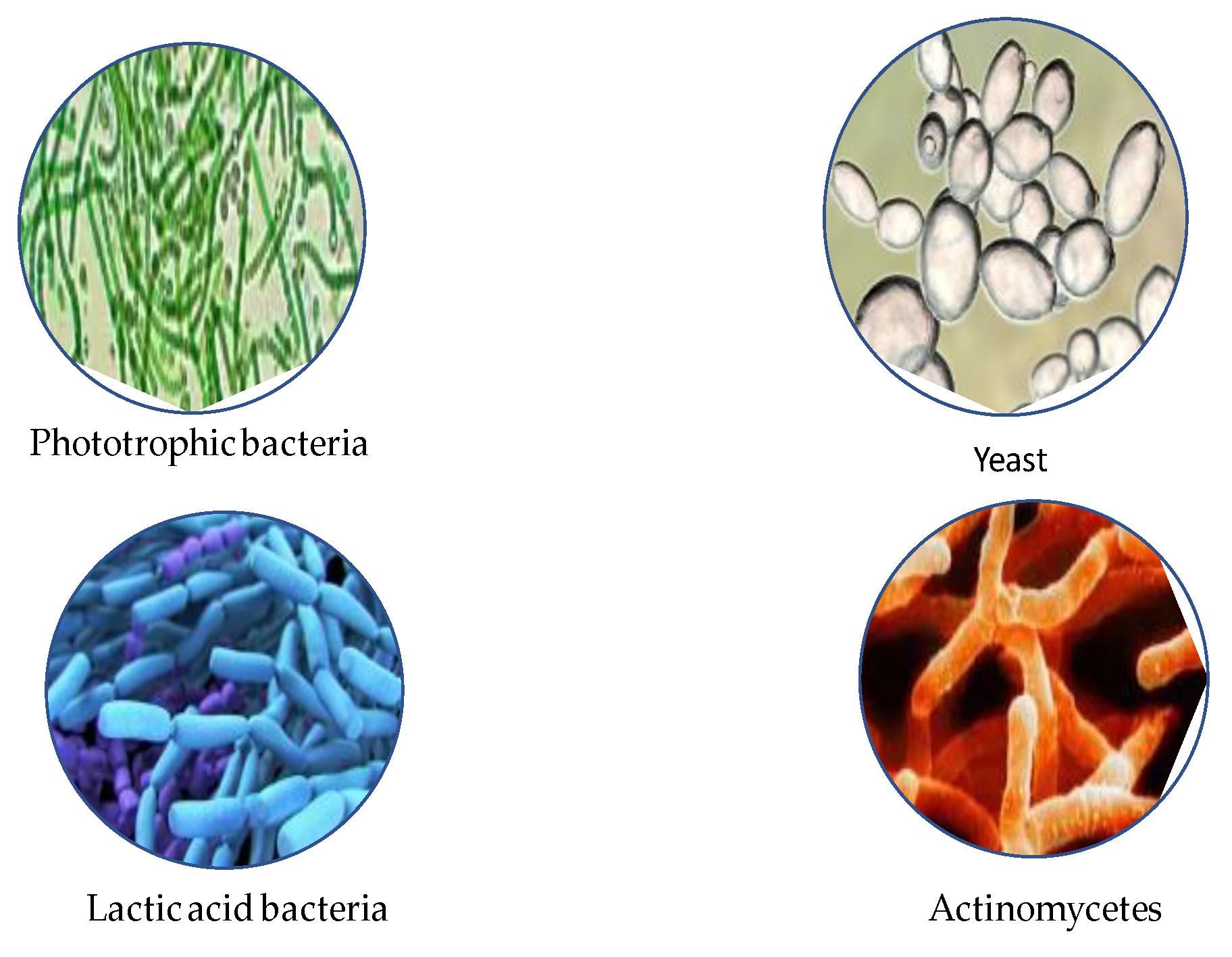
Figure 6.
Interactions between signal cascades (1) Salicylic Acid (SA) and (2) Jasmonic Acid (JA). Source: own.
Figure 6.
Interactions between signal cascades (1) Salicylic Acid (SA) and (2) Jasmonic Acid (JA). Source: own.
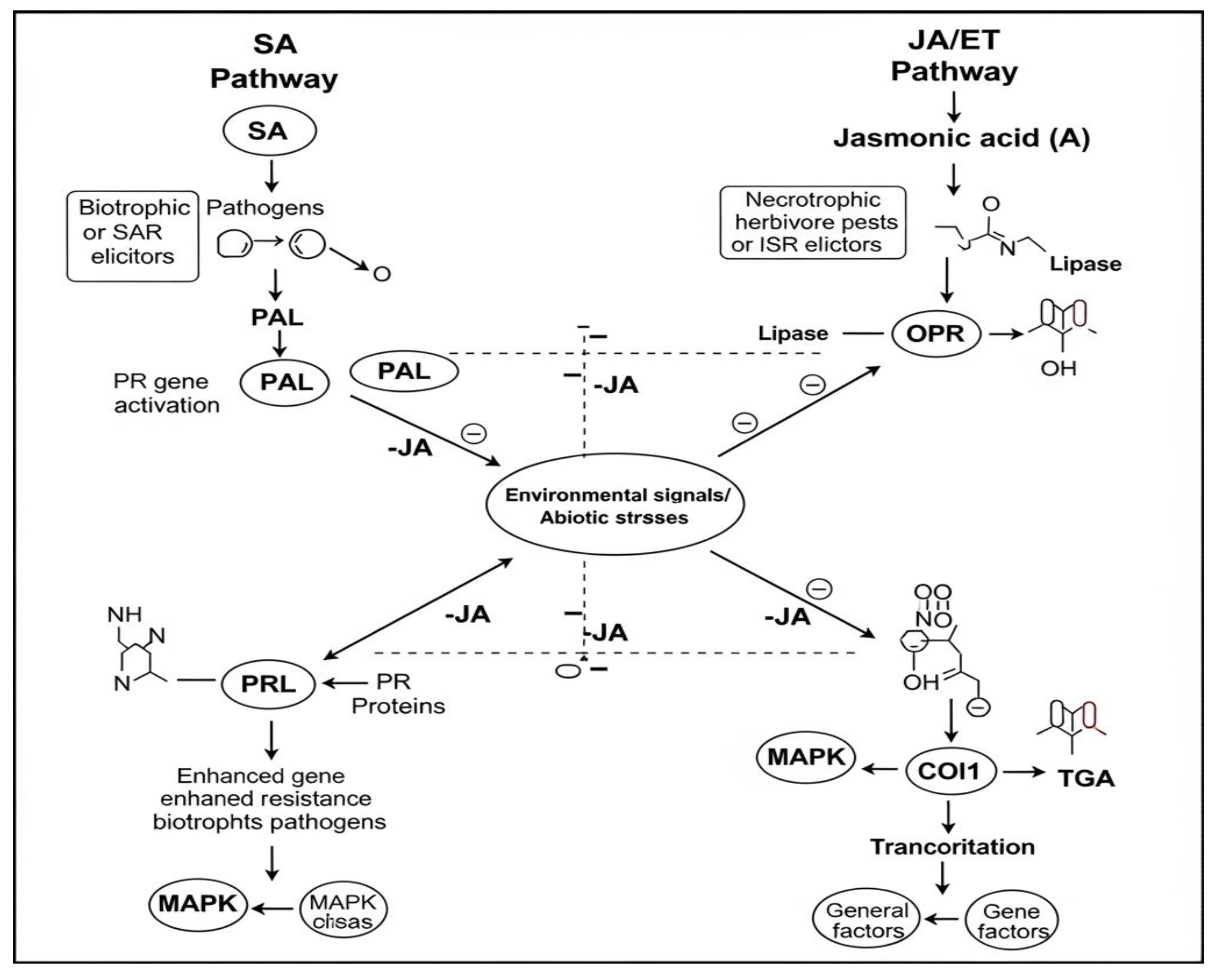
Figure 7.
Environmental implications for genetically modified plants. Source: own.
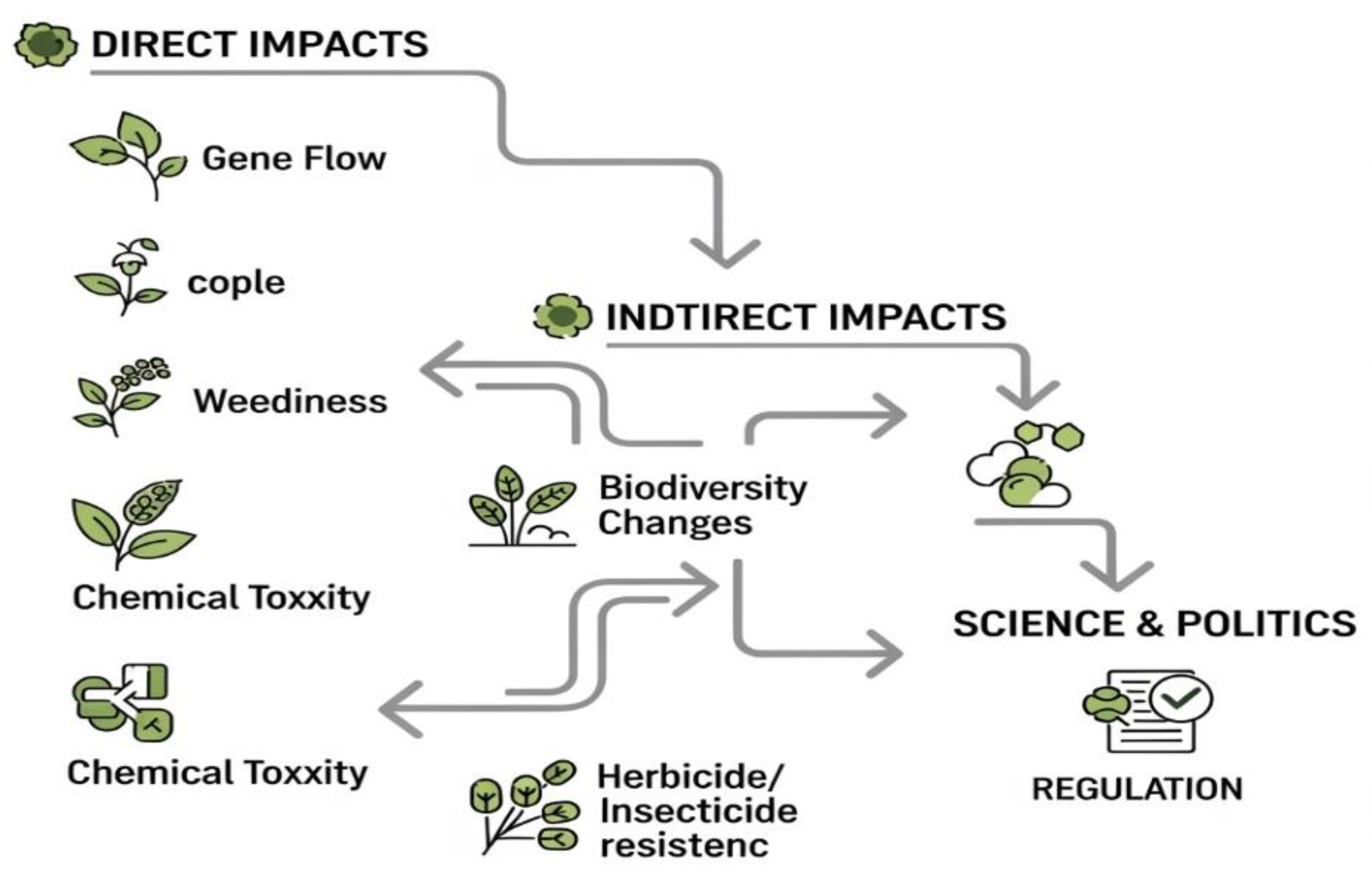
Figure 8.
Participation of individual SND strains in nitrification and denitrification reactions. Source: own.
Figure 8.
Participation of individual SND strains in nitrification and denitrification reactions. Source: own.


Table 1.
Examples of microbial bioherbicides.
| Microbe/ References | Weed target(s) & Status | Trade name | Year of Introduction or registration |
|---|---|---|---|
| Alternaria cassia Bannon,1988 | Cassia obtusifolia C. coccidentalis Crotalaria spectabilis Never commercialized | Casst™ | never |
| Alternaria destruens Bewick et al., 2000 | Cusucta spp. Discontinued | Smolder™ | 2005 |
| Chondrostereum purpureum Hintz, 2007 | Populus and Alnus spp. Unknown | Chontrol™ | 2004 |
| Colletotrichum acutatum Morris, 1989 | Hakea sericea Discontinued | Hakatak | 1990 |
| Colletotrichum gloeosporioides f. sp. aeschynomene Cartwright et al., 2010 | Aeshynomene vigrinica Available on demand | Collego® | 1982 |
| Colletotrichum gloeosporioides f. sp. Malvae Boyetchko et al. 2007 | Acacia mearnsii and A. pycnantha Discontinued | Stumpout™ | 1997 |
| Cylindrobasidium laeve. Morris et al. 1999 | Acacia mearnsii and A. pycnantha Discontinued | Stumpout™ | 1997 |
| Phoma macrostoma Bailey et al. 2011 | many broadleaf weed species Available | Bio-Phoma™ | 2016 |
| Phytophthora palmivora Ridings 1986 | Morrenia odorata Discontinued | DeVine® | 1982 |
| Pseudomonas fluorescens Kennedy et al. 2001 | Bromus tectorum Discontinued | D7® | 2014 |
| Puccinia canaliculata Phatak et al. 1983 | Cyperus esculentus Discontinued | Dr. Biosedge™ | 1987 |
| Puccinia thlaspeos Knopp et al. 2002 | Isatis tinctorial Discontinued | Woad Warrior® | 2002 |
| Sclerotinia minor Watson2018 | Taraxacum officinale Discontinued | Sarritor® | 2009 |
| Several fungi Gale & Goutler, 2013 | Parkinsonia aculeate Available | Di-Bak® | 2019 |
| Streptomyces scabies O’Sullivan et al. 2015 | several grass and broadleaf weeds Never commercialized | Opportune™ | 2012 |
| Tobacco mild green mosaic vírus Charudattan & Hiebert, 2007 | Solanum viarum Available | SolviNix™ | 2014 |
| Xanthomonas campestris pv. poae Imaizaumi et al. 1999 | Poa annua Discontinued | Camperico™ | 1997 |
Source: own adapted on [67].
Table 2.
Advantages and disadvantages in technologies with.
| Advantages | Disadvantages |
|---|---|
| Increasing food production: Some microorganisms are beneficial to plants and help in their growth and production. They can also help produce food by fermenting and processing food. | Uncontrolled growth of microorganisms: In some cases, microorganisms can grow uncontrollably and become harmful to plants, soil and human health. |
| Plant protection: Some microorganisms are able to fight plant diseases and pests, reducing the use of harmful pesticides and other chemicals. | Environmental pollution: Some microorganisms, such as E. coli bacteria, can cause soil and water pollution, which is a public health risk |
| Improving soil quality: Microorganisms can help enrich the soil with nutrients and improve its structure, which positively affects the health of plants. | GMO risks: Some microbial technologies, such as genetic engineering, can lead to genetically modified organisms (GMOs), which raise social and ethical concerns. |
| Sustainable Agriculture: The use of microorganisms can aid sustainable agriculture by reducing the use of harmful chemicals and improving soil quality. | Costs: Some systems and technologies using microorganisms can be expensive, which is a barrier to their widespread use |
Table 3.
Isolated species.
| Origin of species | Specification of Species | References |
|---|---|---|
| Activated sludge |
Acinetobacter sp. ND7 Ochrobactrum anthropic LJ81 Alcaligenes faecalis NR Achromobacter sp. GAD3 Agrobacterium sp. LAD9 Acinetobacter sp. SZ28 Acinetobacter sp. WB-1 Ochrobactrum sp. KSS10 Pseudomonas stutzeri CFY1 Thauera sp. FDN-01 Diaphorobacter sp. PD-7 |
[[152] [146] [151] [150] [150] [126] [126] [28] [151] [[153] [126] |
| Artificial lake |
Acinetobacter sp. H36 Acinetobacter sp. CN86 |
[[147] [147] |
| Domestic wastewater |
Achromobacter xylosoxidans CF-S36 Paracoccus denitrificans ISTOD1 Klebsiella pneumoniae CF-S9 |
[154] [[155] [154] |
| Drinking water reservoir | Zoogloea sp. N299 | [156] |
| Flooded paddy soil |
Arthrobacter arilaitensis Y-10 Pseudomonas tolaasii strain Y-11 |
[157] [157] |
| Laboratory-scale MBR |
Acinetobacter calcoaceticus HNR Bacillus methylotrophicus L7 Serratia sp. LJ-1 |
[158] [[126] [128] |
| Laboratory-scale SBR |
Acinetobacter junii YB Pseudoxanthomonas sp. YP1 |
[[159] [160] |
| Landfill leachate | Zobellella taiwanensis DN- | [146] |
| Seabed sludge | Paracoccus versutus LYM | [161] |
| Songhua River | Microbacterium esteraromaticum SFA13 | [162] |
| Wastewater system |
Acinetobacter sp. YS2 Cupriavidus sp. S1 Pseudomonas sp. yy7 Rhodococcus sp. CPZ24 |
[[143] [160] [151] [151] |
Source: own.
Table 4.
Herbicides and their respective sources, target weeds, ecosystems and registered names.
| Source | Target Weeds | Ecosystem | Registered Name |
References |
|---|---|---|---|---|
| Alternaria cassiae | Cassia obtusifolia L. | Soy | Recipe development - “CASST” | [54] |
| Alternaria destruens | Cuscuta spp. | Cranberry Field | assessment - Smolder | [11] |
| C. purpura | P. Serotina | Forest | Commercoalizedm Biochon TM | [61] |
| C. purpura | Populus euramericana | Guinier Forest | Commercialized - Chontrol® | [57] |
| Cephalospprium diospyri | Diospyras virginiana L. | Pastures, pastures | Oklahoma | [53] |
| Chondrostereum purpureum (Fr.) | Pouz Prunus serotina Ehrh. | Forest, Mountains | Commercialized - Mycotech™ | [54] |
| Citrus lime (L.) Osbeck | D. Sanguinalis | Cultivated areas | Commercial herbicide Avenger® | [51] |
| Citrus sinensis (L.) Osbeck | Solanum nigrum L. | Crop land, roadside | Commercialized - Green Match™ | [62] |
| Colletotrichum gloeosporioides | Hakea sericea Schrad. &J.C. Wendl. | Mountain meadows | Commercialized - Hakak |
[55] |
| Colletotrichum gloeosporioides | Malvae, Malva Pusilla Sm. | Flex, lentils, horticultural crops | Commercialized - BioMal ® [56] |
[56] |
| Colletotrichum gloeosporioidesaeschynomene | Aeschynomene virginica L. | Rice, Soybeans | Commercialized– Colle™ | [54] |
| Cylindrobasidium | leave Acacia spp. | Forest, pasture | Commercialized - Stump-Out™ | [61] |
|
Cymbopogon citratus (DC.) |
Stapf. spp. | Agricultural land | Commercialized - Green Match™ EX | [64] |
| Phoma macrostoma | Reynoutria japonica Houtt. | Golf Courses, Agriculture &Agroforestry | Commercialized - Phoma | [59] |
| Phytophthora palmivora | Morrenia odorata (Hook. &Arn.) Lindl. | Citrus Groves | Commmercialized — Devine™ | [54] |
| Puccinia thlaspeos C. Shub. | Isatis tinctoria L. | Forest, pastures | - Beloukha ® [62] | [63] |
| S. aromaticum | E. crus-galli | Farmland, Rice | Commercialized - Weed Slayer® | [59] |
| Sclerotinia minor Jagger. | Taraxacum sp. | Turf | Commercialized - Sarritor ® [60] | [61] |
| Streptomyces acidi scabies | Taraxacum officinale L. | Turf | Commercialized - Opportune ® |
[59] |
|
Syzygium aromaticum (L.) Merr. & LM Perry & Presl Cinnamomum verum J., |
E. crus-galli | Rice, farmland | Commercialized - WeedZap ® | [53] |
| Xanthomonas campestris | Poa annua L. | Turf, athletic fields | Commercialized - Camperico | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



